






γλώσσες
Σελίδες
Νομικός


1
CURS BIOLOGIE MOLECULARĂ
Reglarea Iniţierii Transcripţiei
Prof. Dr. Marieta Costache
2
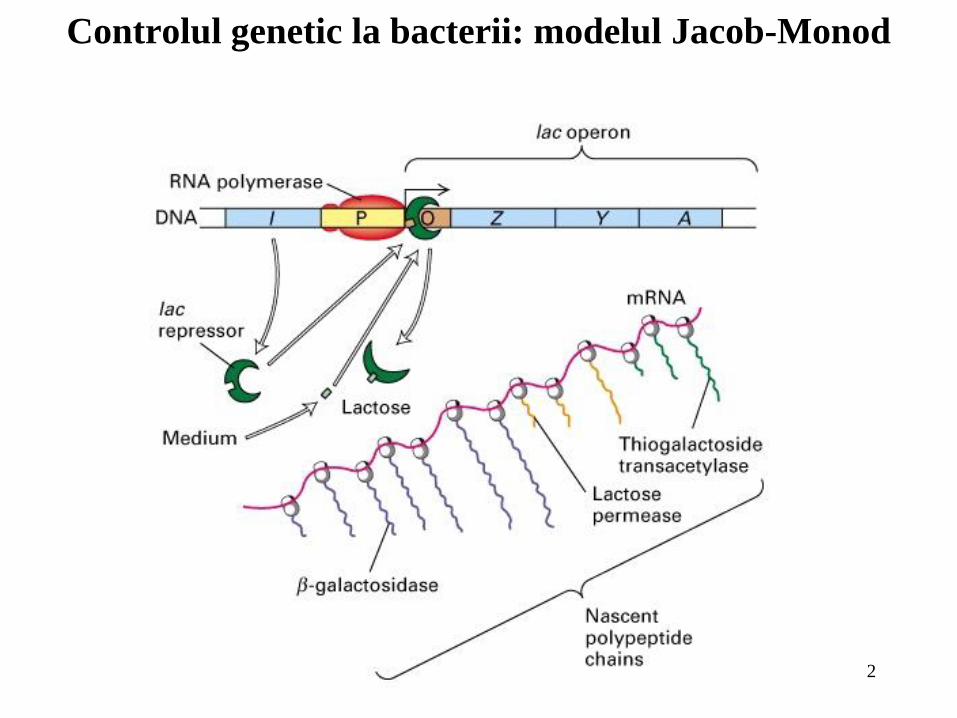
Controlul genetic la bacterii: modelul Jacob-Monod
3
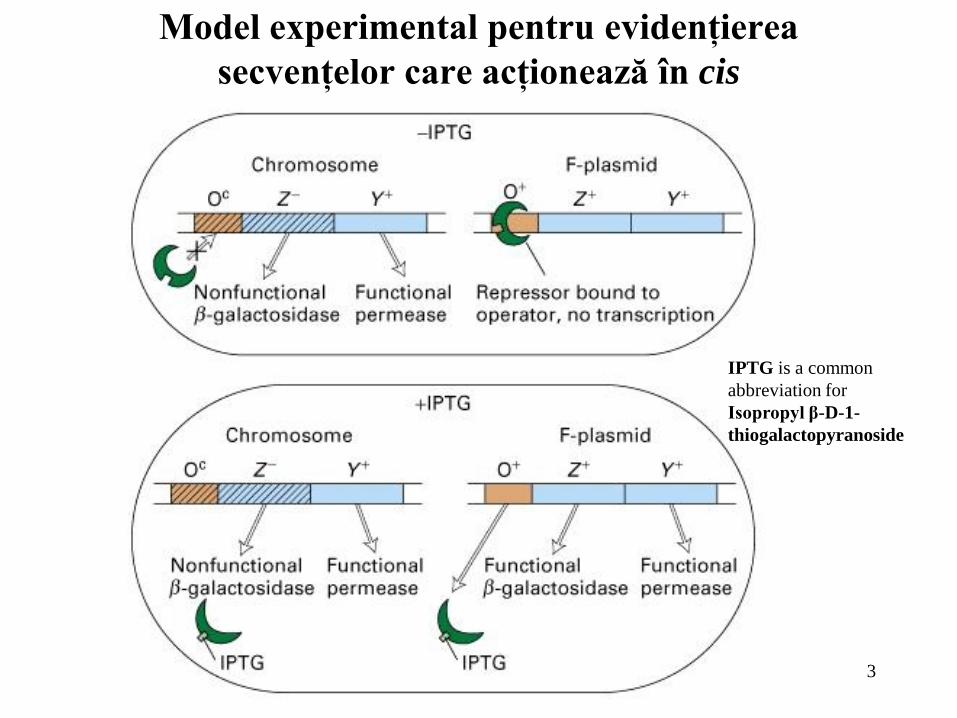
Model experimental pentru evidenţierea
secvenţelor care acţionează în cis
IPTG is a common
abbreviation for
Isopropyl β-D-1-
thiogalactopyranoside
4
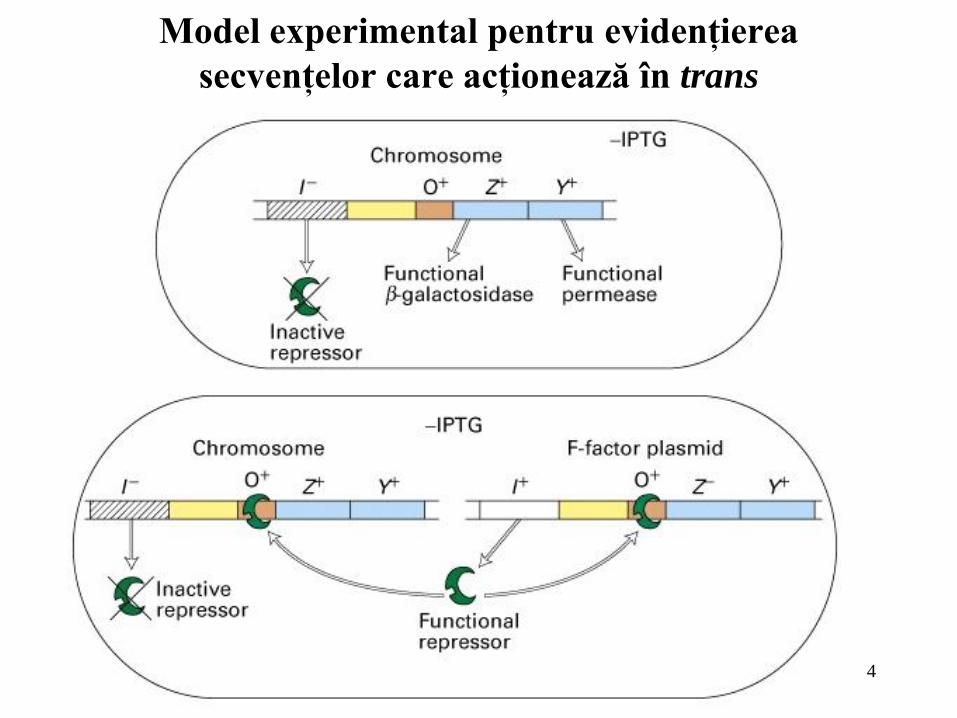
Model experimental pentru evidenţierea
secvenţelor care acţionează în trans
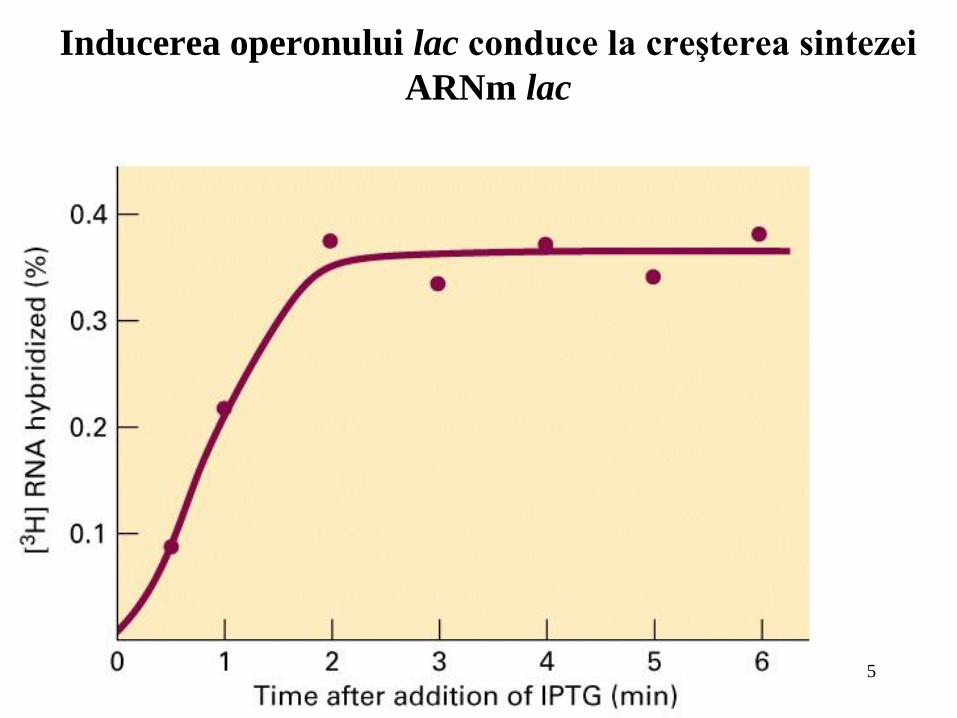
5
Inducerea operonului lac conduce la creşterea sintezei
ARNm lac

6
Concluzii
Multe proteine bacteriene sunt inductibile, sinteza lor fiind dependentă de
statusul nutriţional al celulelor. Expresia diferenţială a genelor care codifică astfel
de proteine apare cel mai adesea la nivelul iniţierii transcripţiei;
În concordanţă cu modelul Jacob şi Monod privind controlul
transcripţiei, transcripţia operonului lac, care codifică trei proteine inductibile este
represată de legarea proteinei represoare lac la secvenţa operator. În absenţa
lactozei sau a altor inductori represia este înlăturată şi operonul lac este transcris;
Mutaţiile la nivelul promotorului, la care se leagă ARN polimeraza, sau la
nivelul operatorului acţionează în cis; aceasta înseamnă că ele afectează numai
expresia genelor de pe aceeaşi moleculă de ADN în care apare mutaţia;
Mutaţiile în secvenţa unui operator care scad legarea unui represor au
drept rezultat o transcripţie constitutivă. Mutaţiile în secvenţa promotorului, care
afectează afinitatea de legarea a ARN polimerazei, pot fie să scadă (mutaţie
“down”) fie să crească (mutaţie “up”) transcripţia;
Represorii şi activatorii acţionează în trans; ei afectează expresia genelor
reglate de ei indiferent de poziţionarea moleculei de ADN în celulă.

7
Inţierea transcripţiei la bacterii
ARN polimeraza iniţiază transcripţia majorităţii genelor la
nivelul unei poziţii unice situată în amonte de secvenţa codantă;
Perechea de baze la nivelul căreia se iniţiază transcripţia este
denumită “situs de iniţiere a transcripţiei” sau Punct START;
Prin convenţie, situl de iniţiere a transcripţiei de pe secvenţa
ADN este desemnată cu poziţia +1, bazele situate în sensul transcripţiei
(downstream) sunt desemnate cu numere pozitive iar cele poziţionate în
sens opus (upstream) sunt asimilate cu numere negative;
Diferitele proteine (ARN polimeraze, activatori, represori)
interacţionează cu ADN la nivelul promotorului sau în vecinătatea
acestuia pentru a regla iniţierea transcripţiei

8
Interacţiunile proteine-ADN identificate prin
tehnica “footprinting”
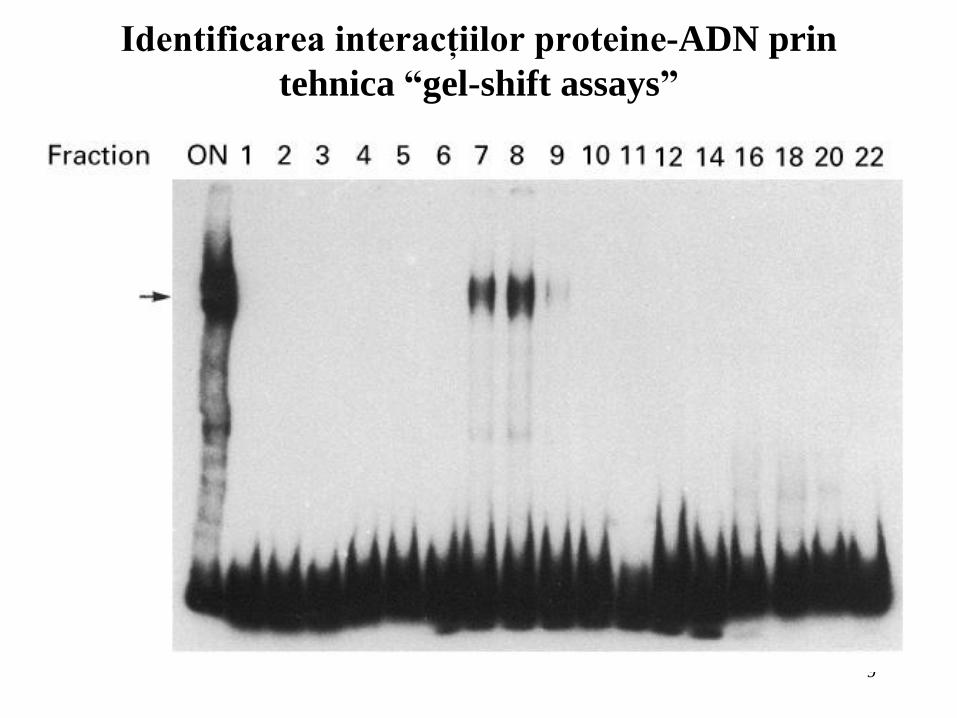
9
Identificarea interacţiilor proteine-ADN prin
tehnica “gel-shift assays”
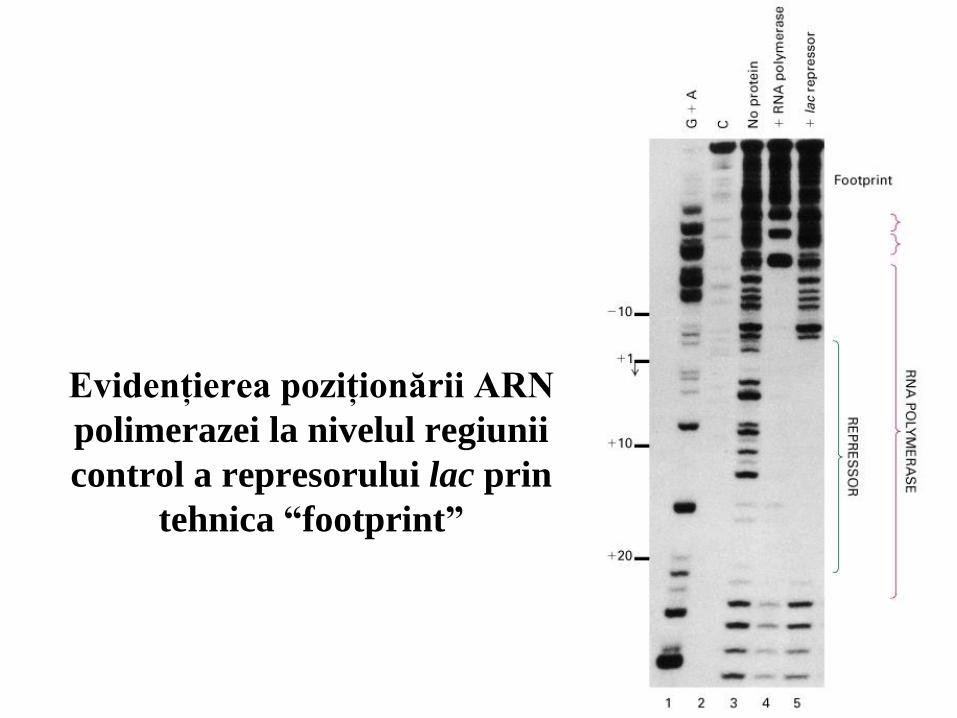
10
Evidenţierea poziţionării ARN
polimerazei la nivelul regiunii
control a represorului lac prin
tehnica “footprint”

11
Regiunea control a operonului lac conţine 3 situri
critice “cis-acting”
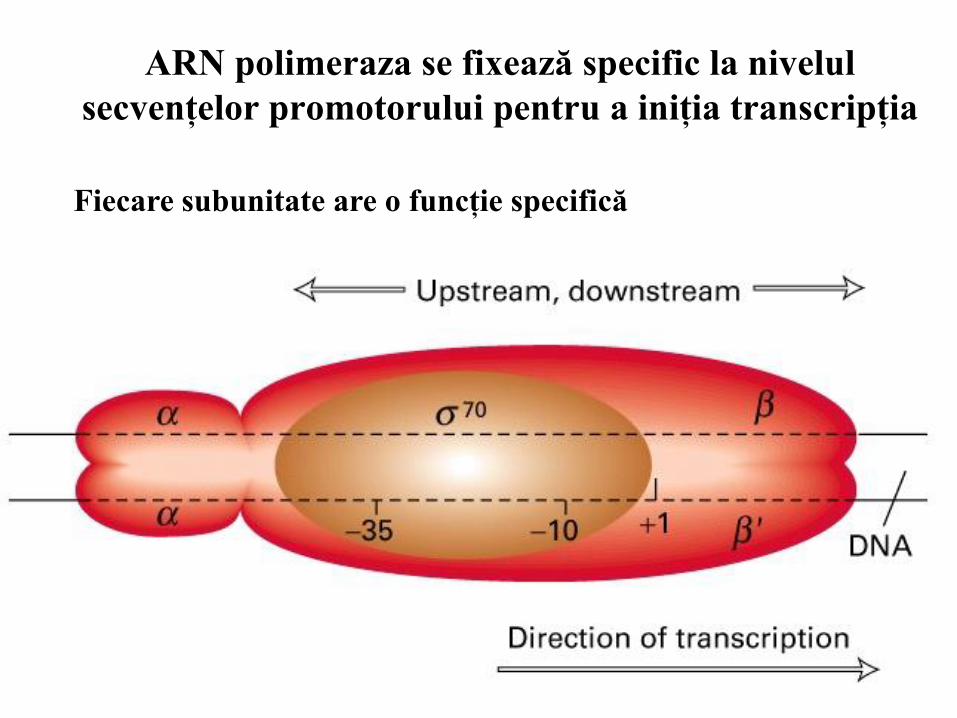
12
ARN polimeraza se fixează specific la nivelul
secvenţelor promotorului pentru a iniţia transcripţia
Fiecare subunitate are o funcţie specifică
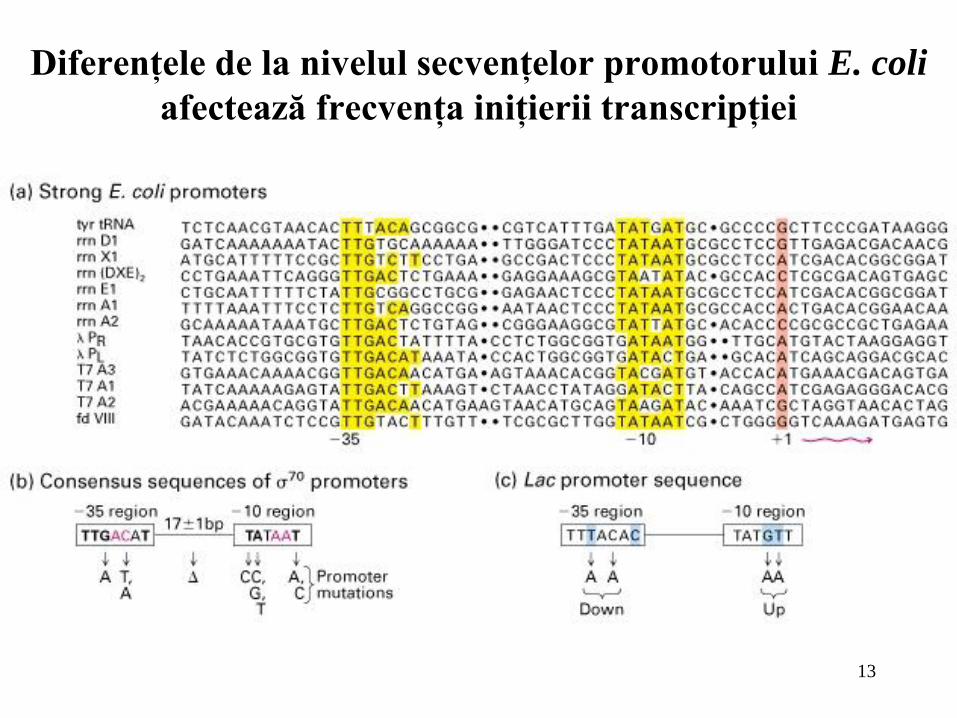
13
Diferenţele de la nivelul secvenţelor promotorului E. coli
afectează frecvenţa iniţierii transcripţiei
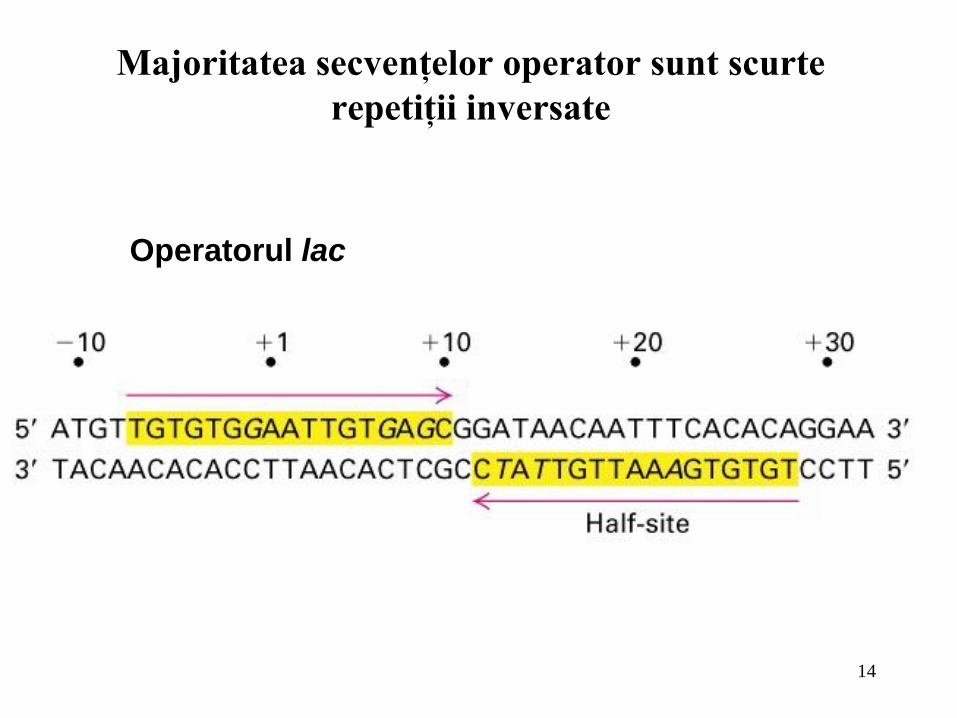
14
Majoritatea secvenţelor operator sunt scurte
repetiţii inversate
Operatorul lac
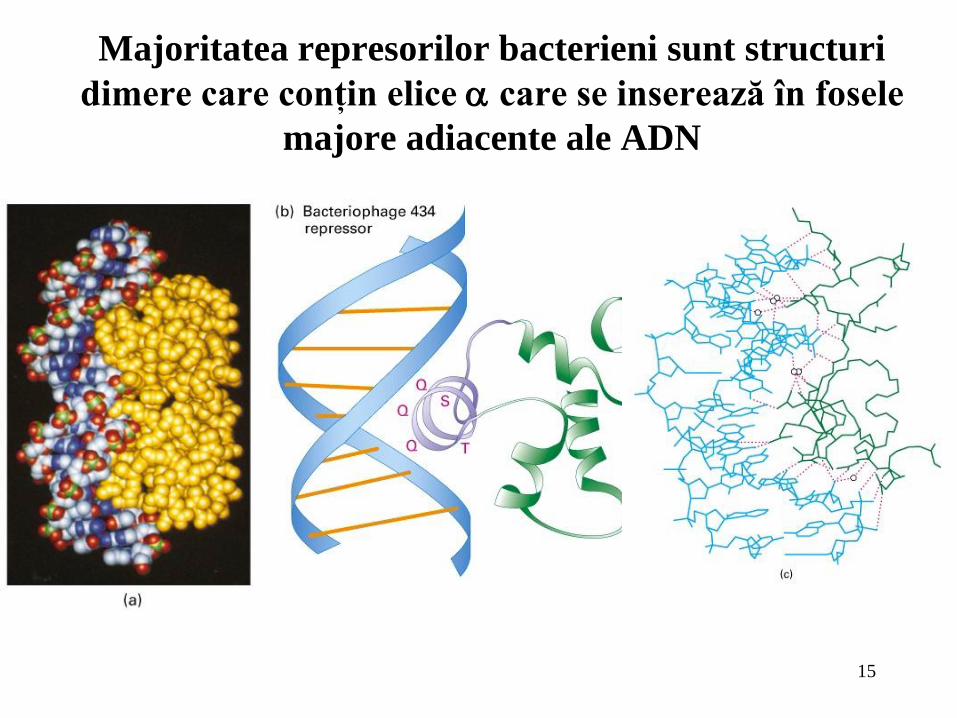
15
Majoritatea represorilor bacterieni sunt structuri
dimere care conţin elice care se inserează în fosele
majore adiacente ale ADN
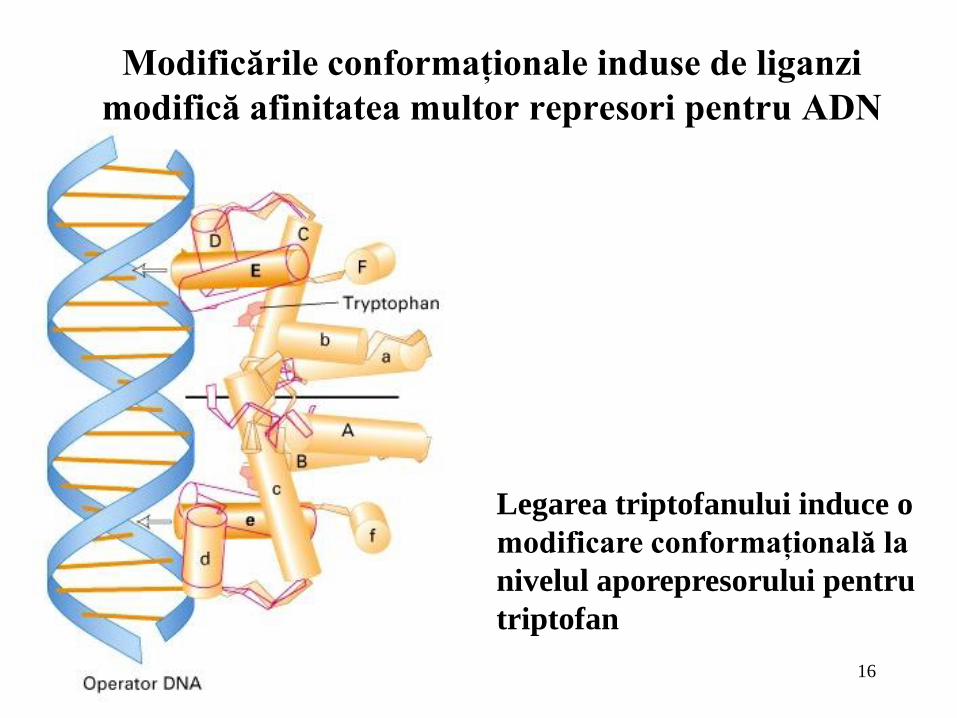
16
Modificările conformaţionale induse de liganzi
modifică afinitatea multor represori pentru ADN
Legarea triptofanului induce o
modificare conformaţională la
nivelul aporepresorului pentru
triptofan
17
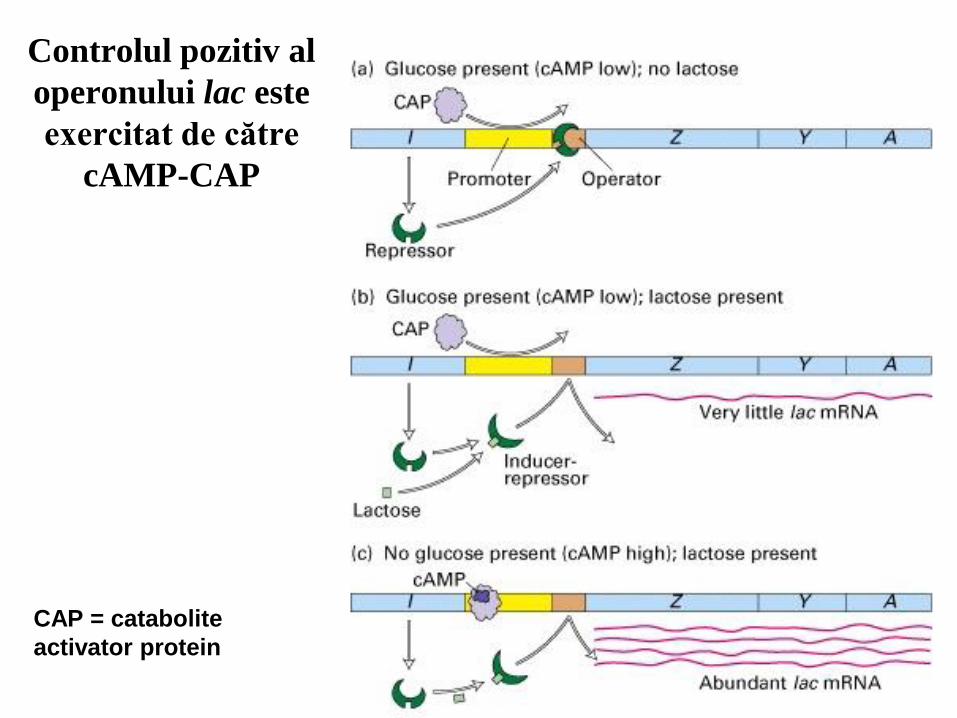
Controlul pozitiv al
operonului lac este
exercitat de către
cAMP-CAP
CAP = catabolite
activator protein
18
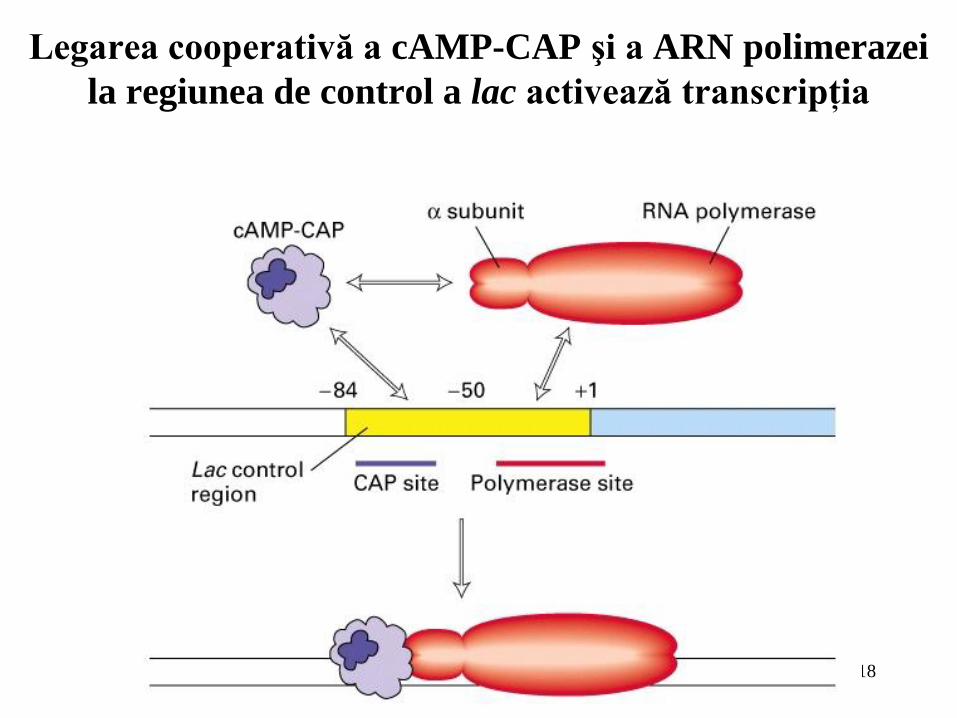
Legarea cooperativă a cAMP-CAP şi a ARN polimerazei
la regiunea de control a lac activează transcripţia
19
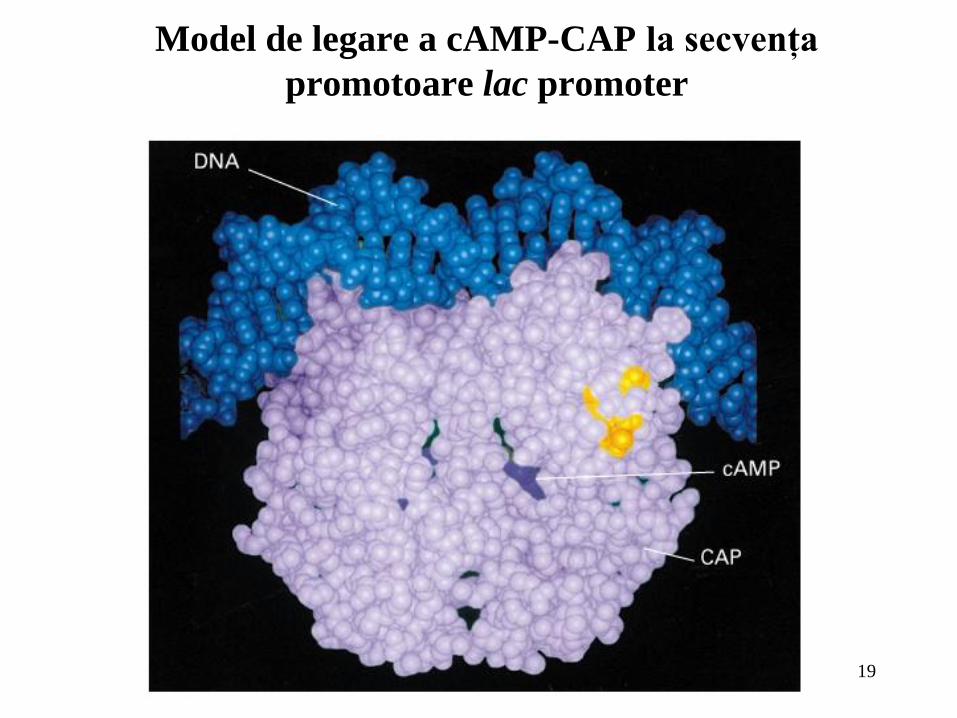
Model de legare a cAMP-CAP la secvenţa
promotoare lac promoter

20
Transcripţia de la nivelul unor promotori este
iniţiată de factorii sigma () alternativi
21
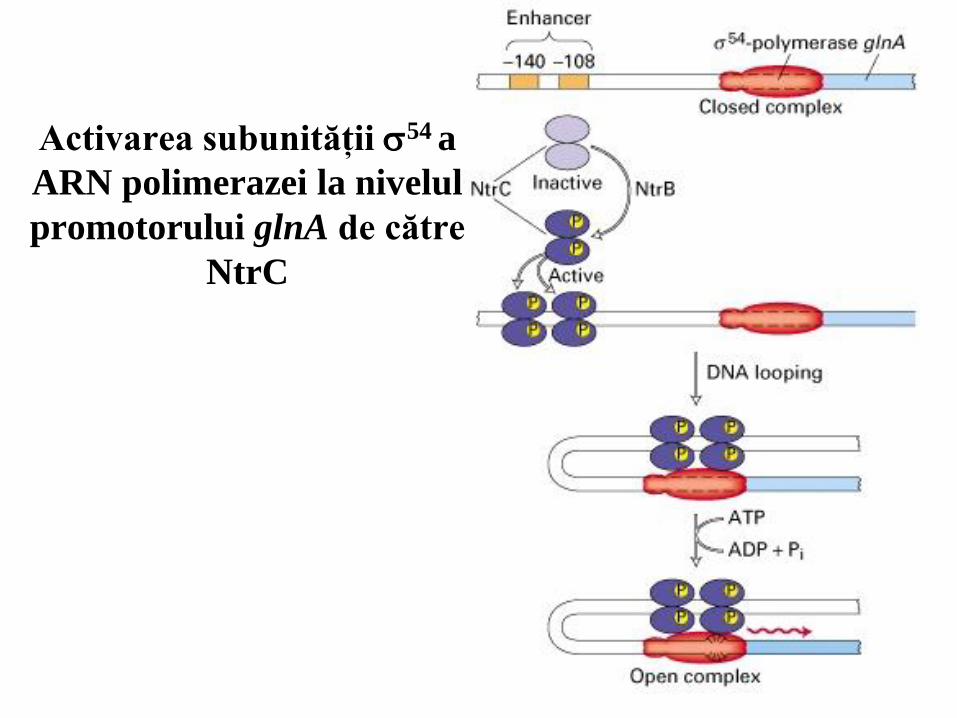
Activarea subunităţii 54 a
ARN polimerazei la nivelul
promotorului glnA de către
NtrC
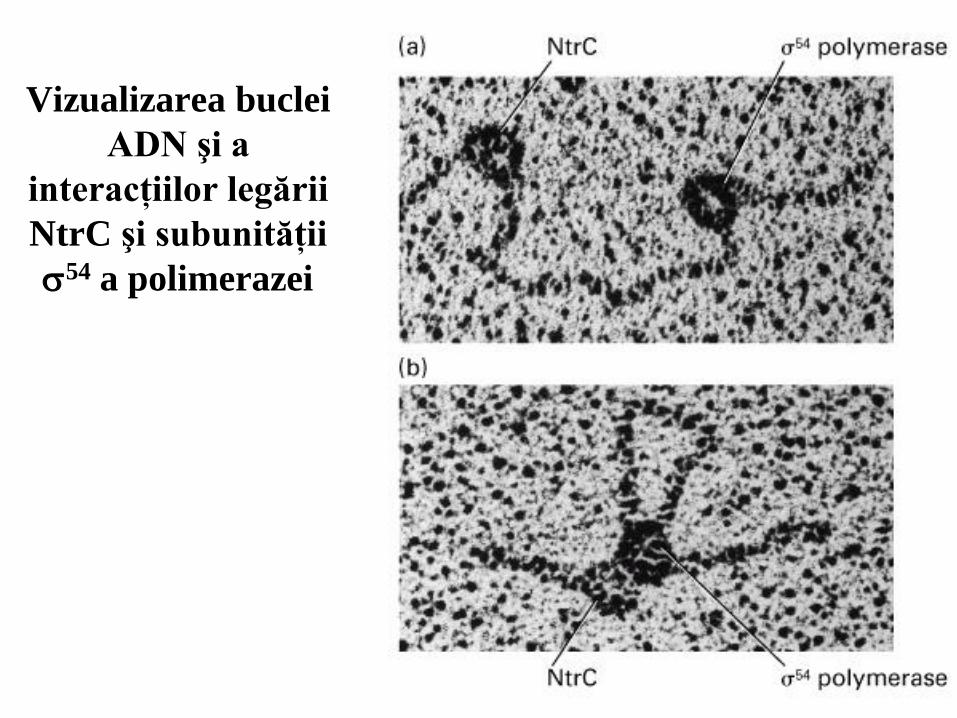
22
Vizualizarea buclei
ADN şi a
interacţiilor legării
NtrC şi subunităţii
54 a polimerazei
23
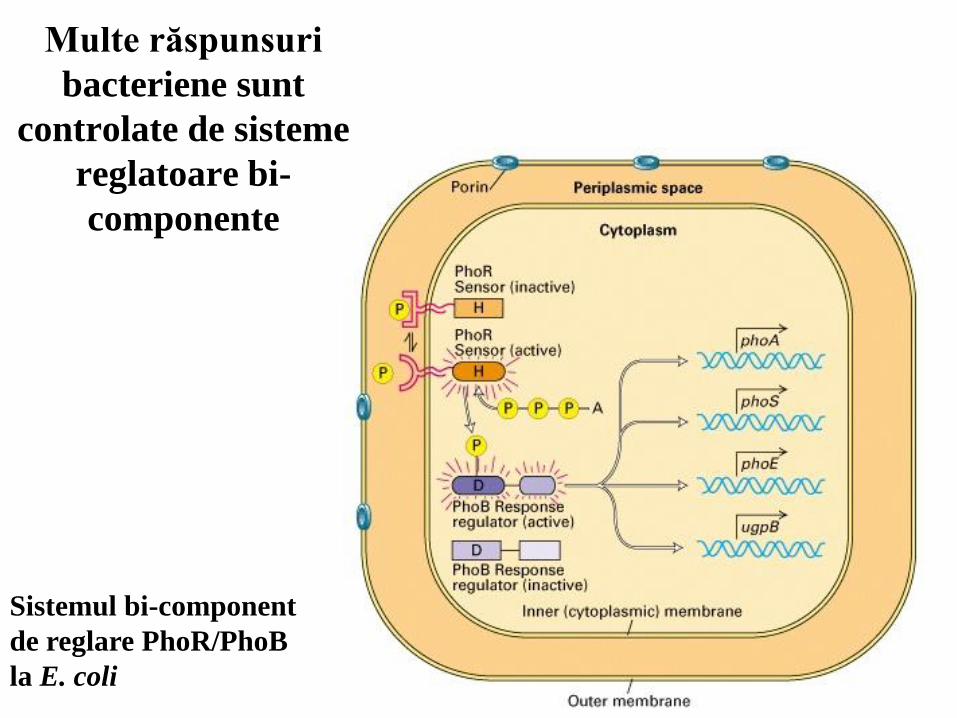
Multe răspunsuri
bacteriene sunt
controlate de sisteme
reglatoare bi-
componente
Sistemul bi-component
de reglare PhoR/PhoB
la E. coli

24
Concluzii (I)
ARN polimerazele sunt proteine mari compuse din subunităţi beta, beta’ şi două subunităţi alfa care formează “core” polimeraza şi o subunitate sigma, din câteva alternative, care funcţionează ca factor de iniţiere; Iniţierea începe când subunitatea sigma a moleculei de polimerază se leagă la secvenţa promotor, formând un complex unit. Polimeraza separă apoi catenele pe o distanţă de 12-13 pb la nivelul situsului de start a transcripţiei, formând un complex deschis. După ce aproximativ 10 ribonucleotide au fost polimerizate, subunitatea sigma este eliberată şi “core” polimeraza continuă transcripţia matriţei; “Puterea” unui promotor se referă la cât de frecvent ARN polimeraza iniţiază transcripţia pornind de la acesta. Subunitatea sigma70, factorul major în iniţiere la E. Coli, interacţionează cu secvenţele promotor situate în regiunea –10 la –35 faţă de situsul de start; Represorii se leagă la secvenţele de ADN numite operatori, care se suprapun parţial cu regiunea promotoare la nivelul căreia se leagă ARN polimeraza. Legarea represorului interferă cu legarea ARN polimerazei şi cu iniţierea transcripţiei; Activatorii subunităţii sigma70 a ARN polimerazei se leagă, în general, la ADN pe partea opusă a helixului faţă de polimerază, în regiunea –20 la –50, sau chiar în amonte de polimerază, în apropierea –60;

25
Concluzii (II)
Activatorul cAMP-CAP stimulează transcripţia prin formarea unui complex cu ARN polimeraza care prezintă o mai mare afinitate pentru situsuri ADN specifice decât proteinele individuale. În plus faţă de stimularea legării polimerazei, activatorii pot stimula şi formarea complexului deschis şi începerea transcripţiei; Mulţi represori bacterieni sunt dimeri. Fiecare monomer conţine un alfa-helix care se inserează în fosa majoră a operatorului ADN, astfel încât dimerul se leagă la fose majore succesive. Afinitatea mare de legare este rezultatul formării multor legături de hidrogen, ionice şi van der Waals dintre proteine şi secvenţe ADN specifice; Secvenţele de ADN care leagă proteinele reglatoare dimere sunt secvenţe repetitive inverse, care reprezintă fiecare jumătaţi de situsuri care leagă un monomer; Operonii transcrişi de către subunitatea sigma54 sunt reglati prin activatori care se leagă la secvenţe “enhancer” situate la aproximativ 100 pb în amonte de situsul de iniţiere. Secvenţele “enhancer” care leagă activatori interacţionează tranzitoriu cu polimeraza echilibrată legată la nivelul promotorului, stimulând formarea unui complex deschis şi iniţierea transcripţiei; În sistemele bi-componente reglatoare, o proteină acţionează ca un senzor, monitorizând nivelul de nutrienţi din mediu. În condiţii adecvate, proteina senzor activează o a doua proteină, reglatorul răspunsului, care se leagă apoi la secvenţele reglatoare, stimulând sau represând astfel transcripţia genelor specifice.

26
Controlul genelor eucariote: scop şi principii
generale
Spre deosebire de celulele bacteriene şi eucariotele
unicelulare, celulele organismelor pluricelulare au relativ
puţine gene reglate reversibil de condiţiile de mediu;
Controlul genetic al organismelor pluricelulare este
important pentru dezvoltare şi diferenţiere şi,
În general, nu este reversibil.
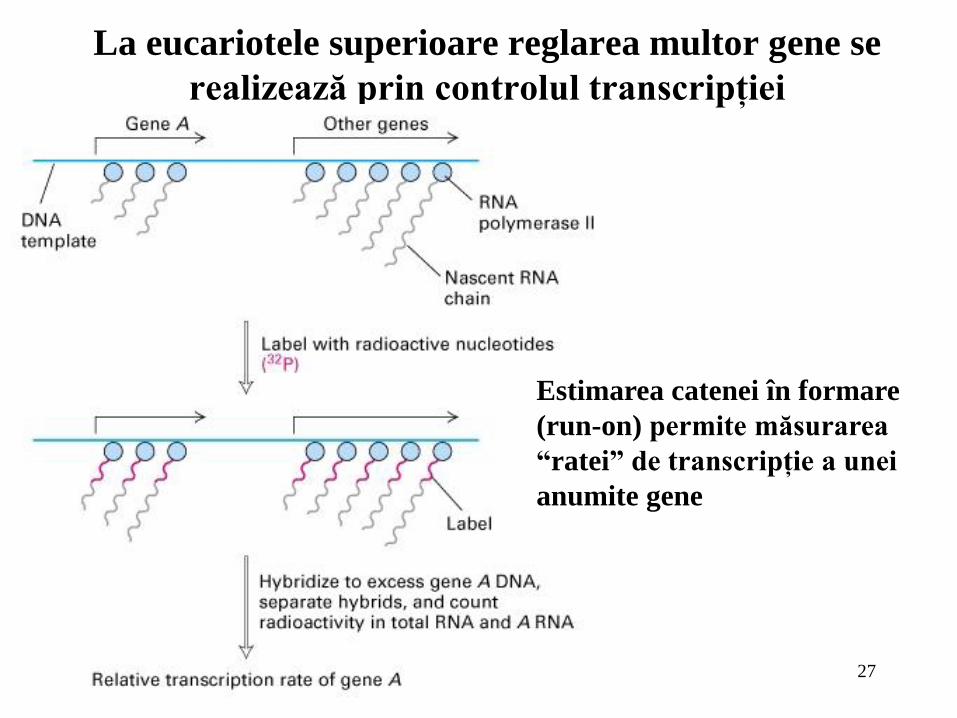
27
La eucariotele superioare reglarea multor gene se
realizează prin controlul transcripţiei
Estimarea catenei în formare
(run-on) permite măsurarea
“ratei” de transcripţie a unei
anumite gene
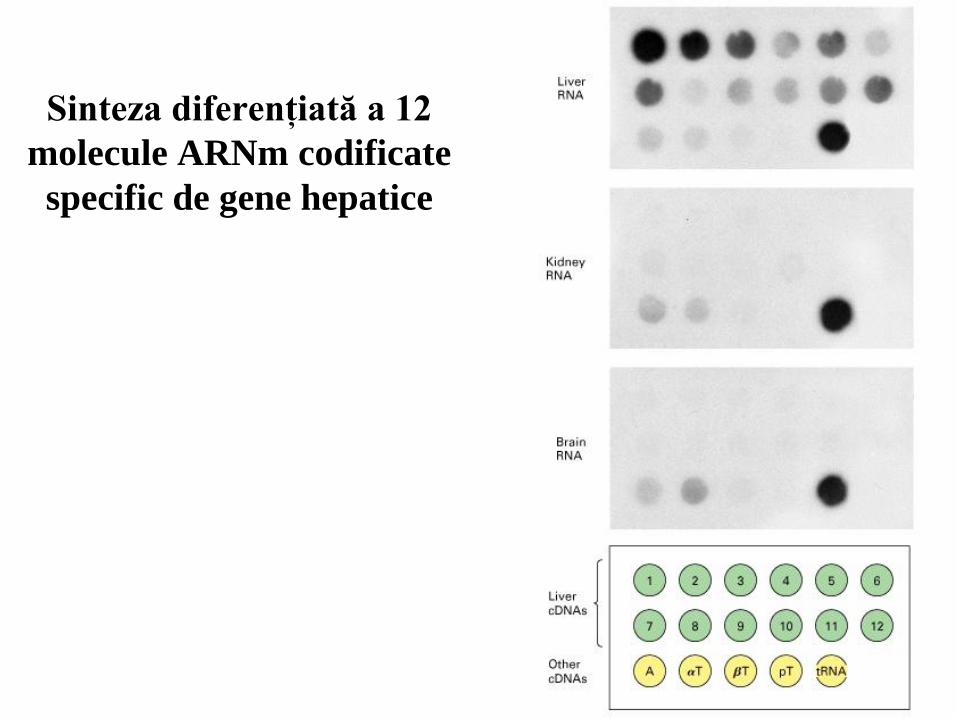
28
Sinteza diferenţiată a 12
molecule ARNm codificate
specific de gene hepatice

29
Analiza ADN
microarray
Analizele ADN microarray ofera o vedere globala asupra
modificarilor in procesul de transcriptie, de exemplu, ca
urmare a aduagarii de SFV (ser fetal) la celulrele umane in
cultura.
Serul contine factori de crestere care stimuleaza celulele in faza
stationara catre crestere si diviziune. Prin analiza ADN
microarray se pot detecta factorii de trasncriptie relative ai
genelor in doua populatii celulare in cultura. (figura). Aceasta
analiza consta in spot-uri foarte fine de ADN atasate pe o lama
de microscop sau pe alt support. Fiecare spot consta in mai
multe copii de secvente ADN dintr-o singura gena umana. Se
realizeaza doua preparate ARN: i) un preparat de ARN, care
contine toate tipurile de ARN provenind de la celulele
stationare care au fost cultivate in fara ser care este marcat cu
molecule fluorescente verzi; ii) un preparat de ARN care
contine toate tipurile de ARN de la celulele in crestere si
diviziune cultivate pe mediu cu SFV si marcate fluorescent in
rosu. Cele doua preparate sunt amestecate si hibridizate pe
lama, unde acestea se vor imperechea cu secventele genelor lor
corespunzatoare. Spoturile verzi (ex, spot 3) indica genele care
sunt transcrise in celulele nedivizate crescute in lipsa de ser.
Spoturile rosii (ex. spot 4) indica genele care sunt transcrise in
celulele in diviziune, iar spoturile galbene (ex. spot 1 si 2)
indica genele care sunt transcrise in cele doua tipuri celulare
(stationare si in diviziune) [From V. R. Iyer et al., 1999, Science
283:83.

30
Elementele reglatoare la eucariote
Reprezintă adesea mii de kilobaze în amonte sau în aval de
START;
Principiile de bază care controlează transcripţia la bacterii există şi
la eucariote: transcripţia este iniţiată la nivelul unei regiuni specifice şi este
controlată de legarea proteinelor “trans-activatoare” (factori de
transcripţie) la secvenţele “cis-activatoare” din structura ADN;
Elementele “cis-activatoare” sunt adesea mult mai departe de
promotorul pe care îl reglează, transcripţia la nivelul unui singur promotor
poate fi reglată prin legarea mai multor factori de transcripţie la
elementele de control alternative;
Secvenţele care controlează transcripţia pot fi identificate prin
analize de deleţie în serie la capătul 5’.
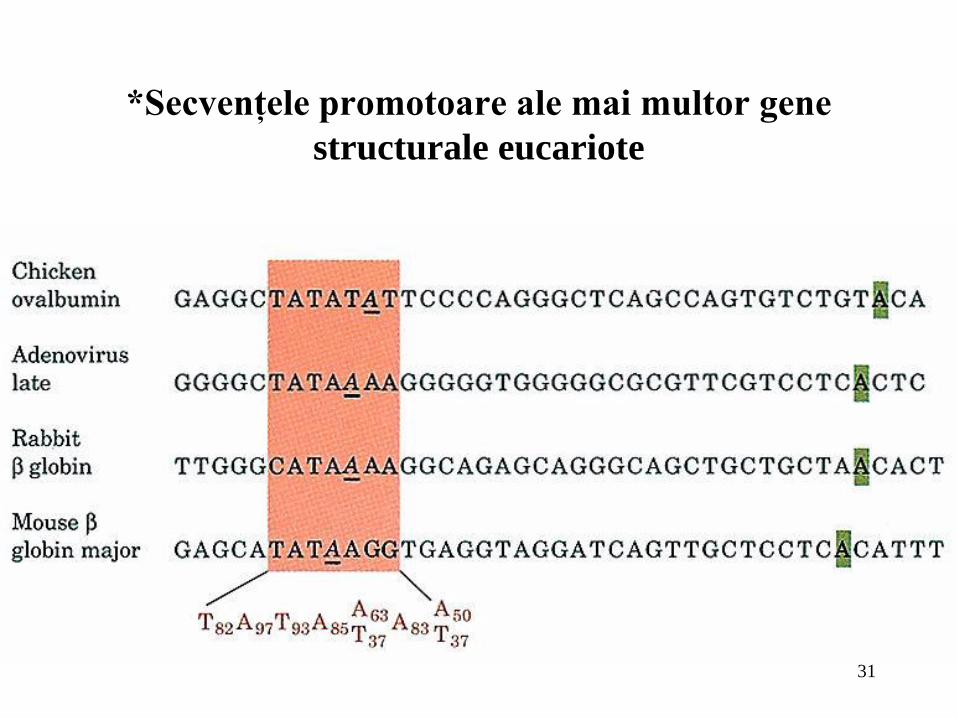
31
*Secvenţele promotoare ale mai multor gene
structurale eucariote
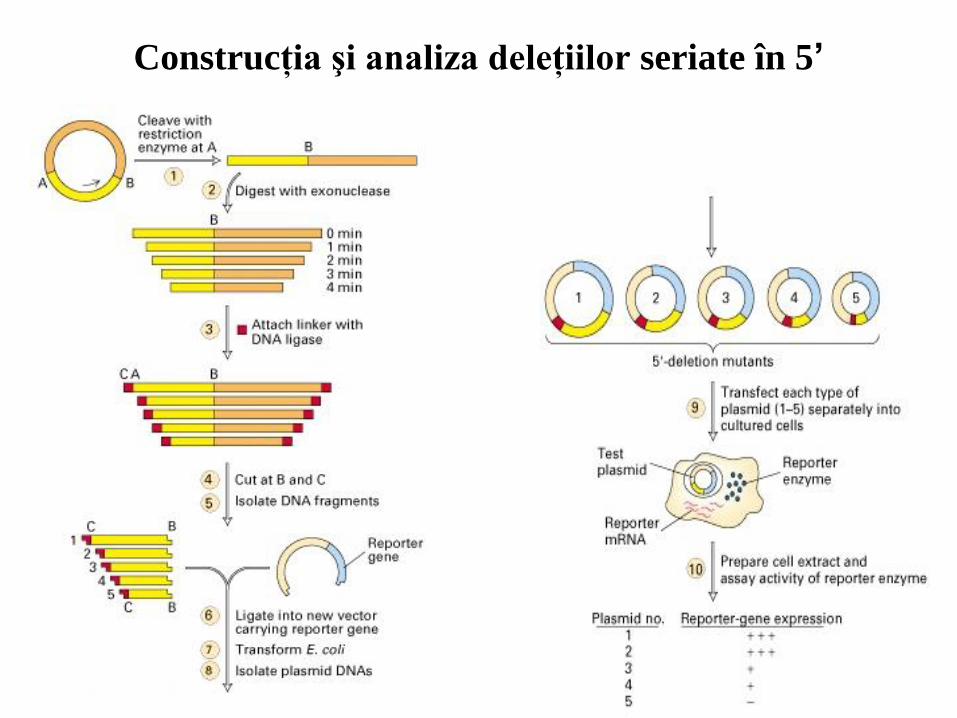
32
Construcţia şi analiza deleţiilor seriate în 5’
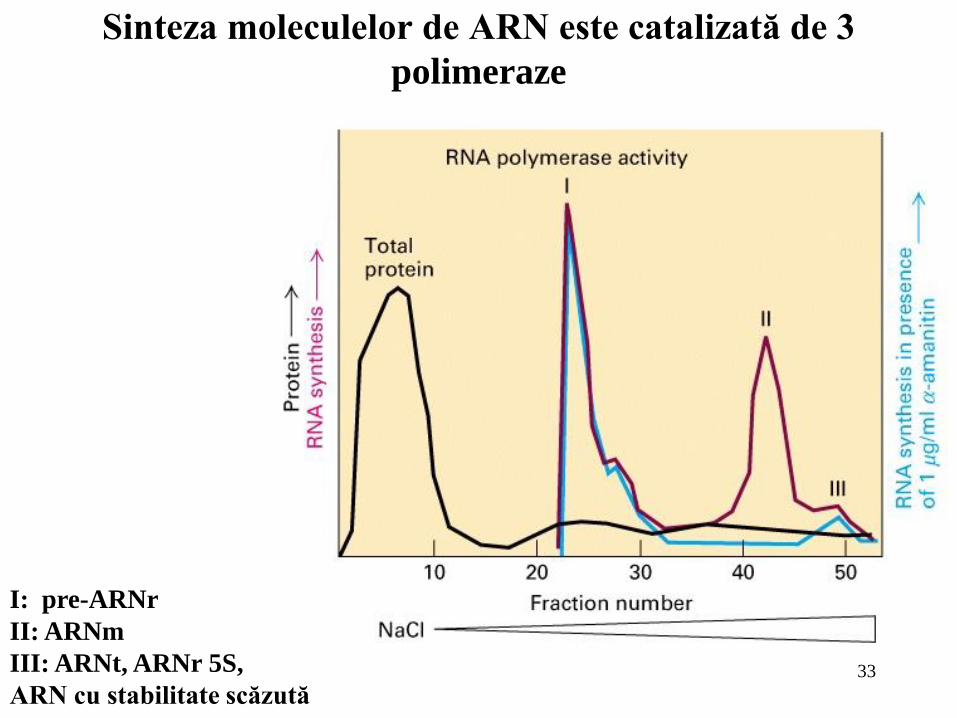
33
Sinteza moleculelor de ARN este catalizată de 3
polimeraze
I: pre-ARNr
II: ARNm
III: ARNt, ARNr 5S,
ARN cu stabilitate scăzută
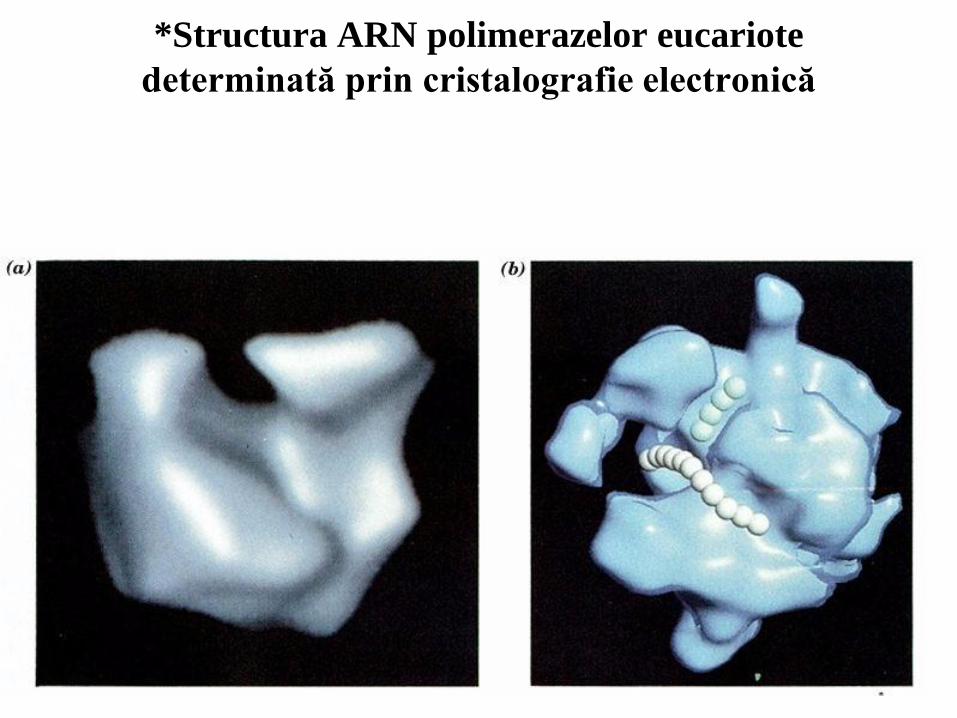
34
*Structura ARN polimerazelor eucariote
determinată prin cristalografie electronică
35
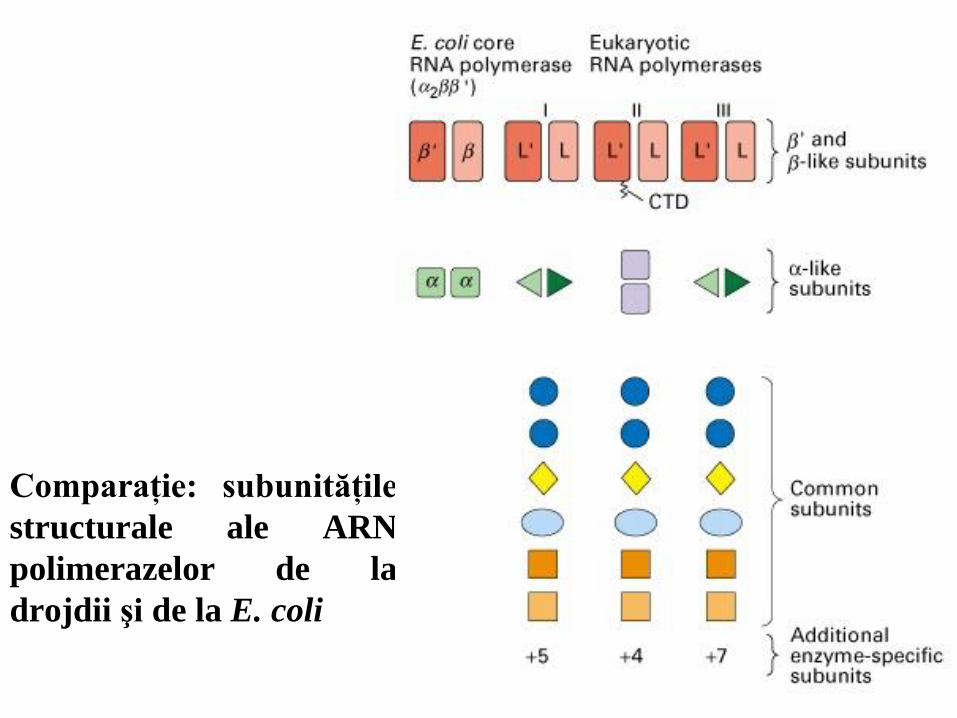
Comparaţie: subunităţile
structurale ale ARN
polimerazelor de la
drojdii şi de la E. coli
36
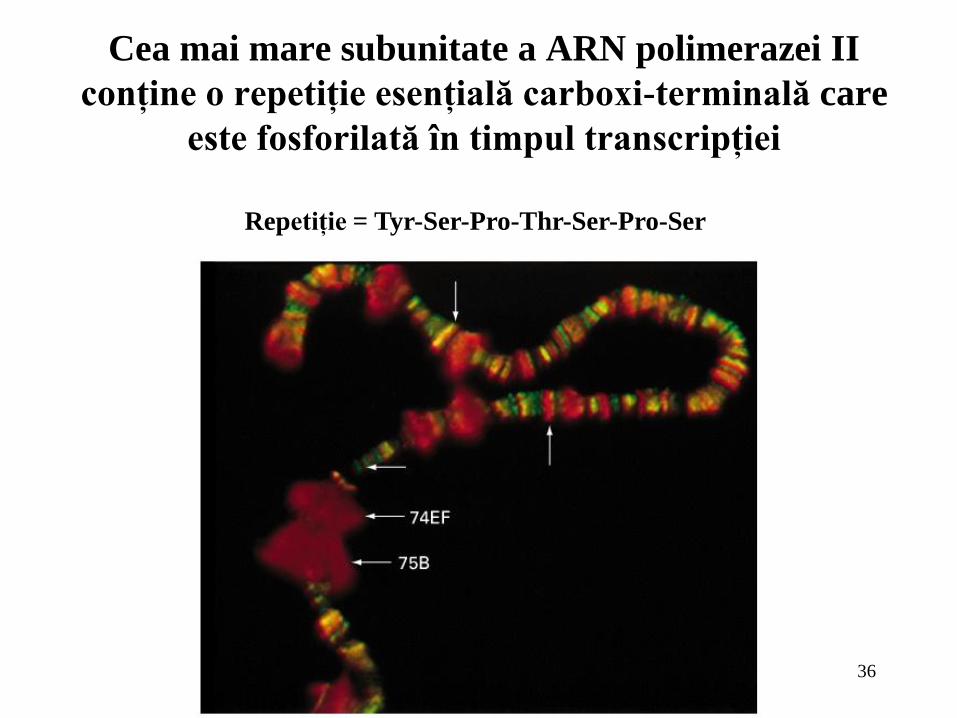
Cea mai mare subunitate a ARN polimerazei II
conţine o repetiţie esenţială carboxi-terminală care
este fosforilată în timpul transcripţiei
Repetiţie = Tyr-Ser-Pro-Thr-Ser-Pro-Ser
37
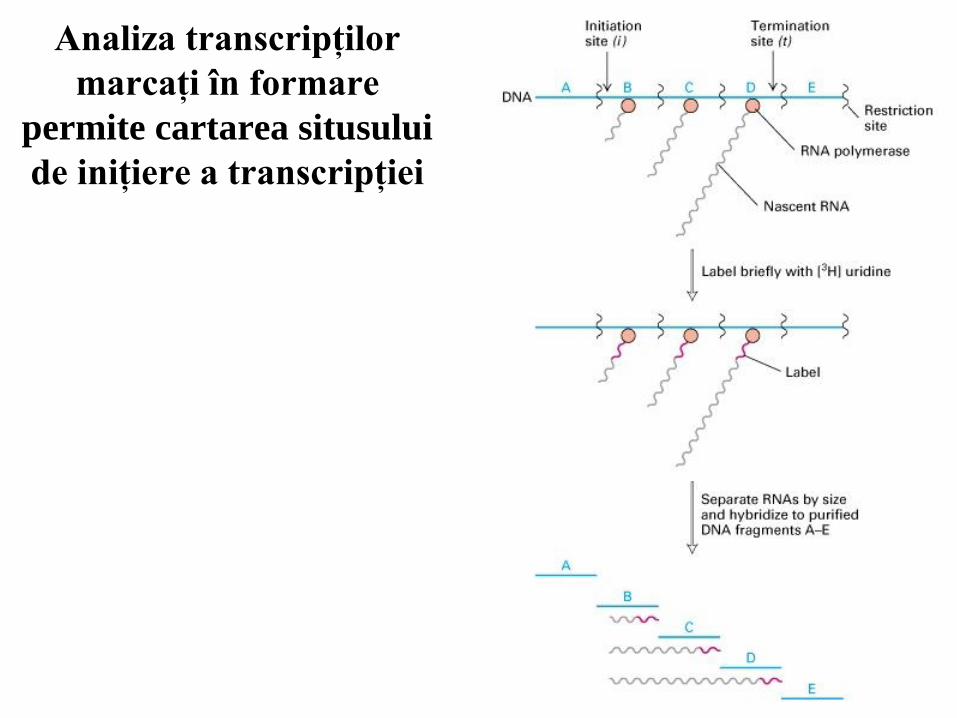
Analiza transcripţilor
marcaţi în formare
permite cartarea situsului
de iniţiere a transcripţiei
38
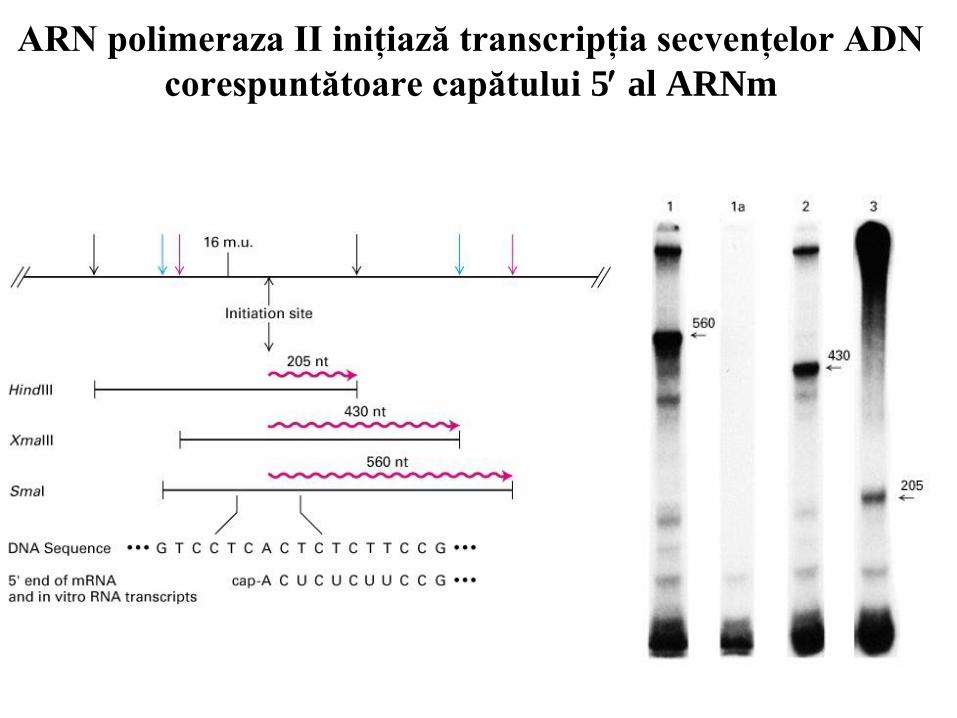
ARN polimeraza II iniţiază transcripţia secvenţelor ADN
corespuntătoare capătului 5 al ARNm

39
Concluzii
Principalul scop al controlului genetic în organismele multicelulare este realizarea deciziilor precise de dezvoltare astfel încât gene particulare sunt exprimate în celule particulare în timpul dezvoltării şi diferenţierii celulare; Controlul transcripţional este principalul sens al reglării expresiei genice atât la eucariote cât şi la procariote; În genoamele eucariote, elementele de control care acţionează în cis şi care reglează expresia de la nivelul promotorului sunt adesea localizate la multe kb depărtare de situsul de start. În contrast, elementele de control bacterian sunt dispuse, în general, pe o distanţă de 60 pb faţă de promotorul pe care aceştia îl reglează; Eucariotele conţin trei tipuri de ARN polimeraze nucleare. Toate trei conţin două subunităţi mari şi două subunităţi mici care constituie structura centrală omologă cu subunităţile , ’ şi ale ARN polimerazei de la E. Coli, cât şi numeroase subunităţi mai mici adiţionale. Unele dintre aceste subunităţi mici sunt comune iar altele sunt unice pentru fiecare polimerază; ARN polimeraza I sintetizează numai pre-ARNr, ARN polimeraza II sintetizează ARNm şi unele molecule mici de ARN nuclear care participă la splicingul ARNm iar ARN polimeraza III sintetizează ARNt, ARNr 5S şi multe alte molecule relativ scurte şi stabile de ARN. O secvenţă heptapeptidică, domeniul carboxi-terminal (CTD), din subunitatea cea mai mare a ARN polimerazei II este fosforilată în timpul iniţierii şi rămâne fosforilată cât timp enzima transcrie matriţa; Similară cu ARN polimerazele bacteriene, ARN polimeraza II iniţiază, de obicei, transcripţia genelor la nivelul unor perechi de baze specifice sau baze vecine alternative din ADN matriţă. Nucleotidul din 5’ căruia i se adaugă capişonul la nivelul ARNm corespunde nucleotidului de pe catena matriţă la care transcripţia este iniţiată.
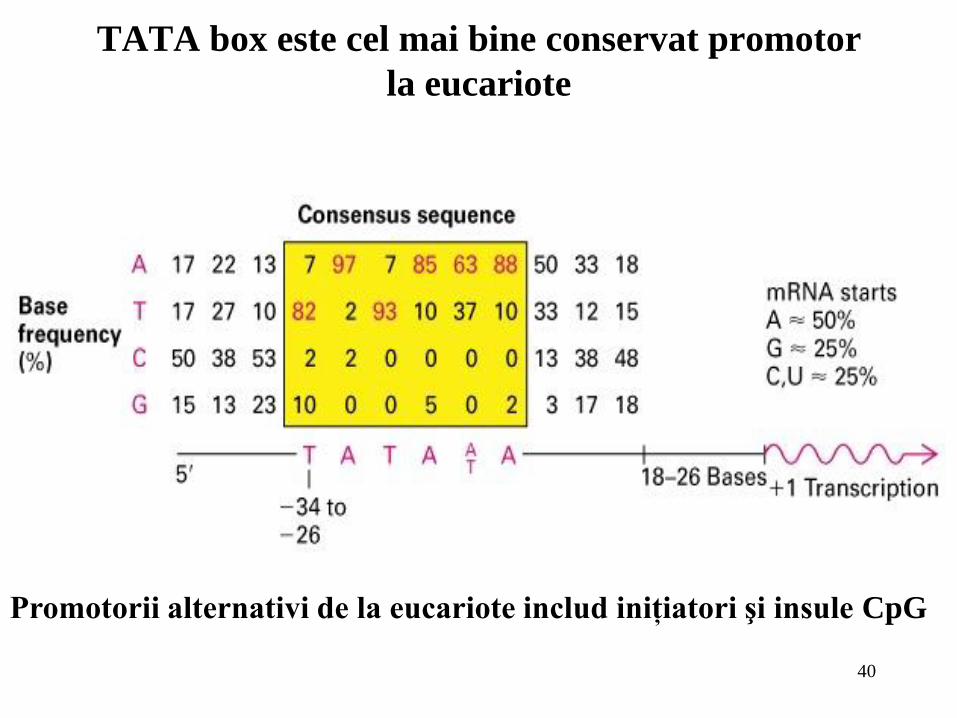
40
TATA box este cel mai bine conservat promotor
la eucariote
Promotorii alternativi de la eucariote includ iniţiatori şi insule CpG
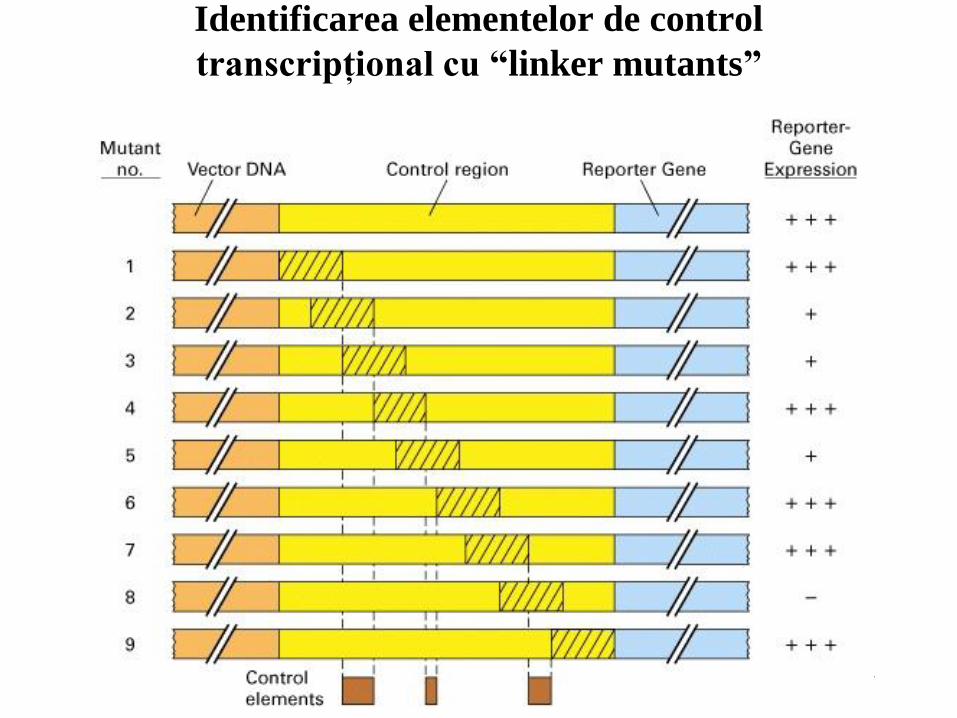
41
Identificarea elementelor de control
transcripţional cu “linker mutants”
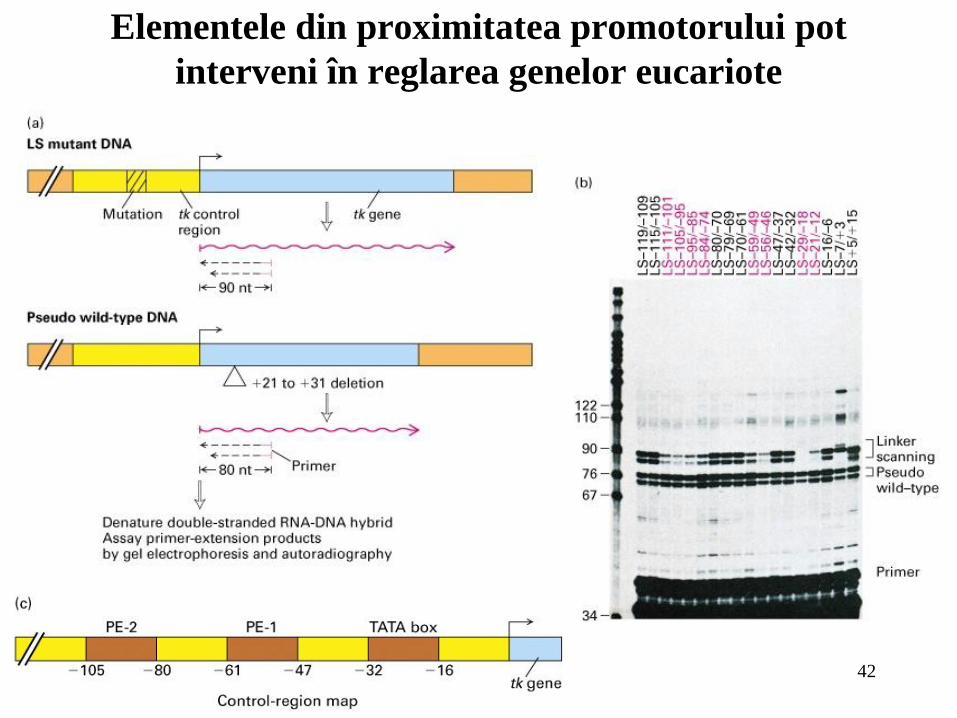
42
Elementele din proximitatea promotorului pot
interveni în reglarea genelor eucariote
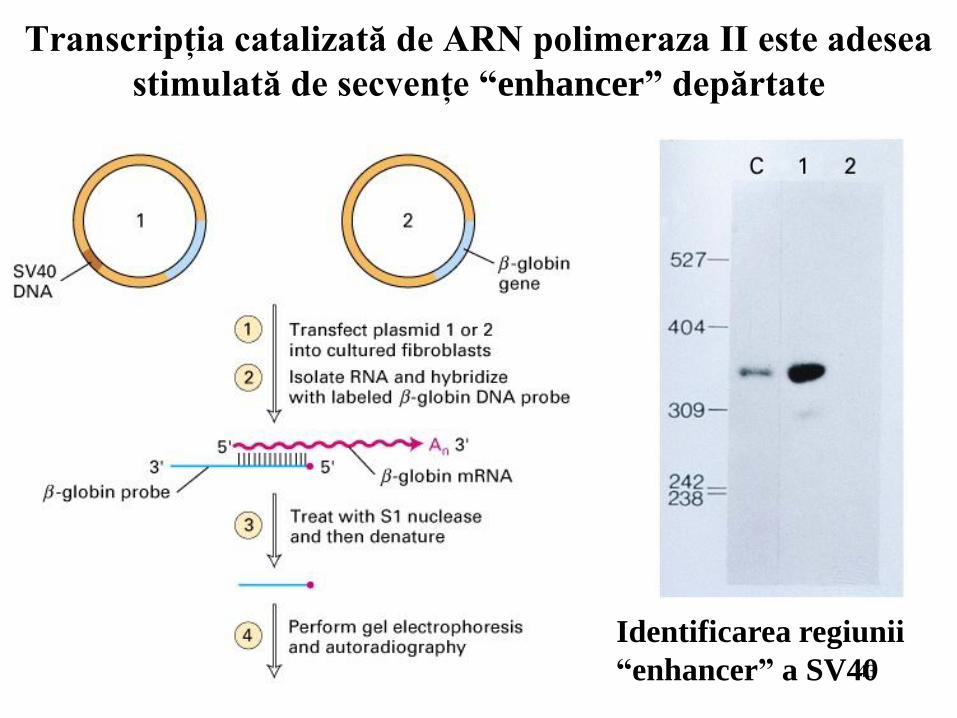
43
Transcripţia catalizată de ARN polimeraza II este adesea
stimulată de secvenţe “enhancer” depărtate
Identificarea regiunii
“enhancer” a SV40
44
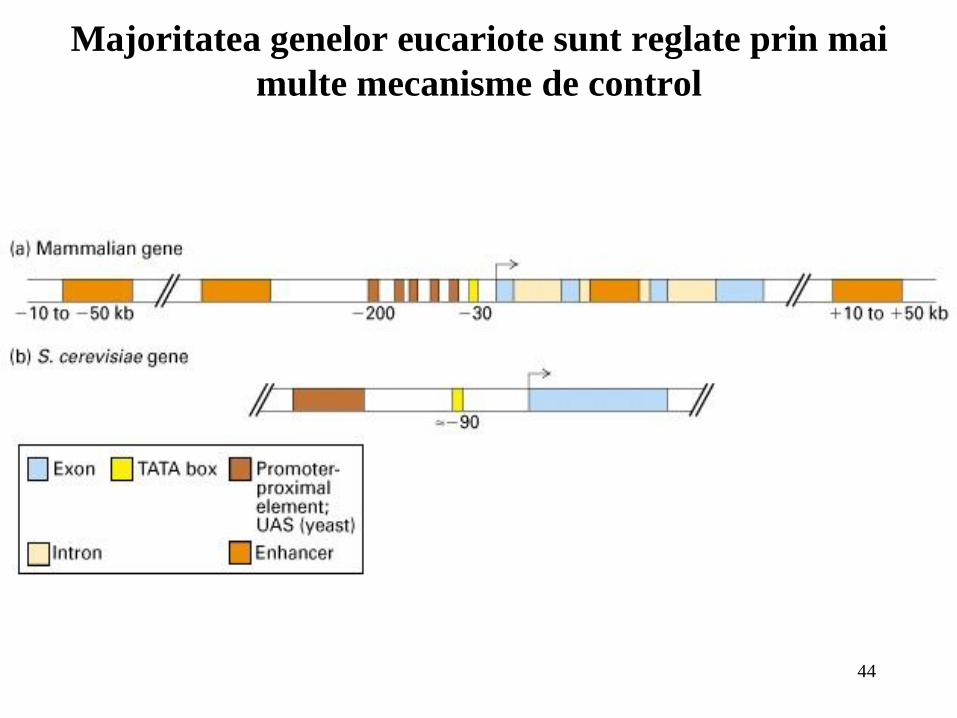
Majoritatea genelor eucariote sunt reglate prin mai
multe mecanisme de control

45
Concluzii
Expresia genelor eucariote care codifică pentru proteine este reglată prin
intervenţia unor elemente multiple care acţionează sub forma unor regiuni de
control în cis. Unele elemente de control sunt localizate în apropierea situsului de
start (elemente din proximitatea promotorului), în timp ce altele sunt localizate la
distanţă (activatori sau “enhanceri”);
Promotorii determină situsul de iniţiere a transcripţiei şi legarea directă a
ARN polimerazei II. Au fost identificate trei tipuri de secvenţe promotor pentru
ADN de la eucariote. TATA box, tipul cel mai comun este dominant pentru genele
transcrise rapid. Promotorii de tip iniţiator sunt reprezentaţi cu o frecvenţă
scăzută în anumite gene în timp ce insulele CpG sunt caracteristice genelor
transcrise;
Elementele din proximitatea promotorului sunt localizate într-un interval
de 200 pb faţă de situsul de start. Multe astfel de elemente, conţinând mai puţin de
20 pb, pot fi utile în reglarea unei gene particulare;
Activatorii, care au de obicei o lungime de aproximativ 100-200 pb, conţin
mai multe elemente control de 8 la 20 pb. Acestea pot fi localizate de la 200 pb la
10 kb în amonte sau în aval de promoter, în interiorul unui intron, sau în aval de
exonul final al genei;
Elementele din proximitatea promotorului şi activatorii sunt adesea
specifici unui tip celular, funcţionând numai tipuri celulare diferenţiate specific.
46
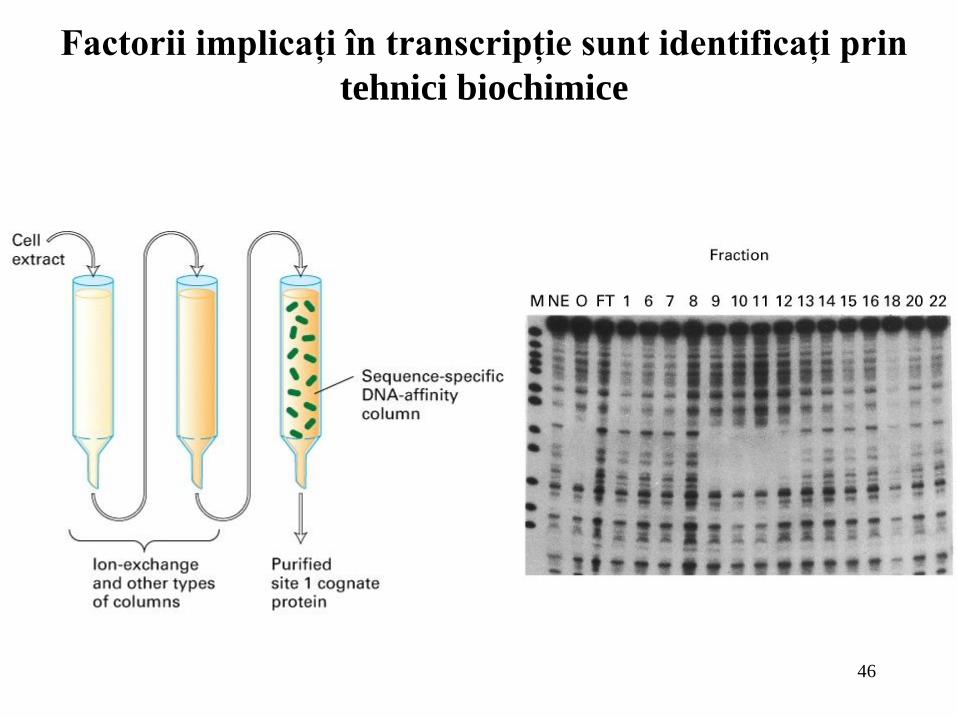
Factorii implicaţi în transcripţie sunt identificaţi prin
tehnici biochimice
47
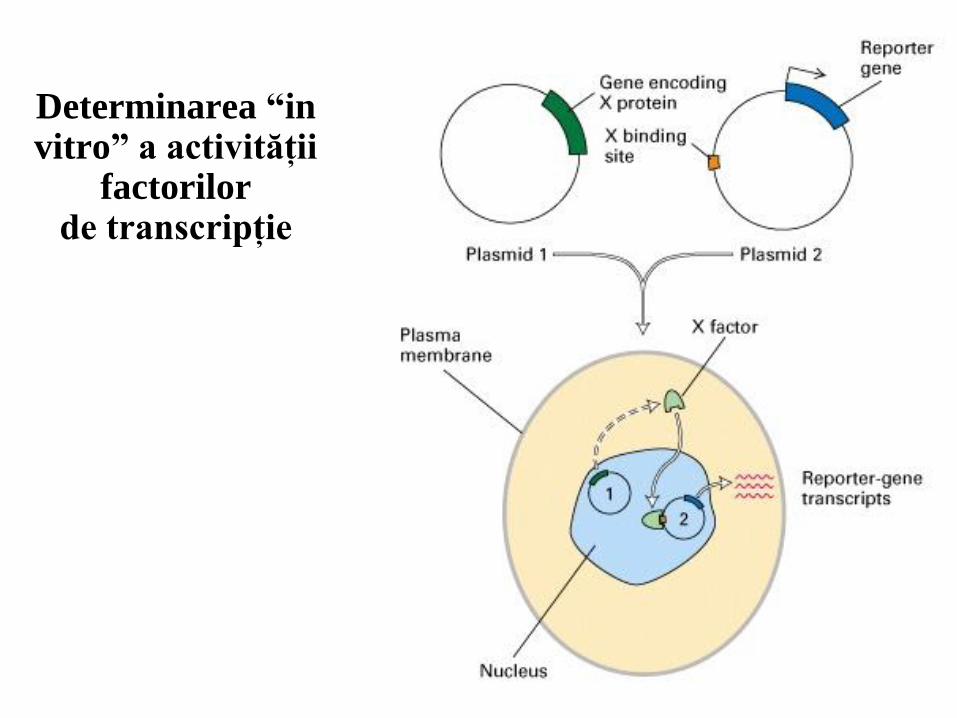
Determinarea “in vitro” a activităţii
factorilor de transcripţie
48
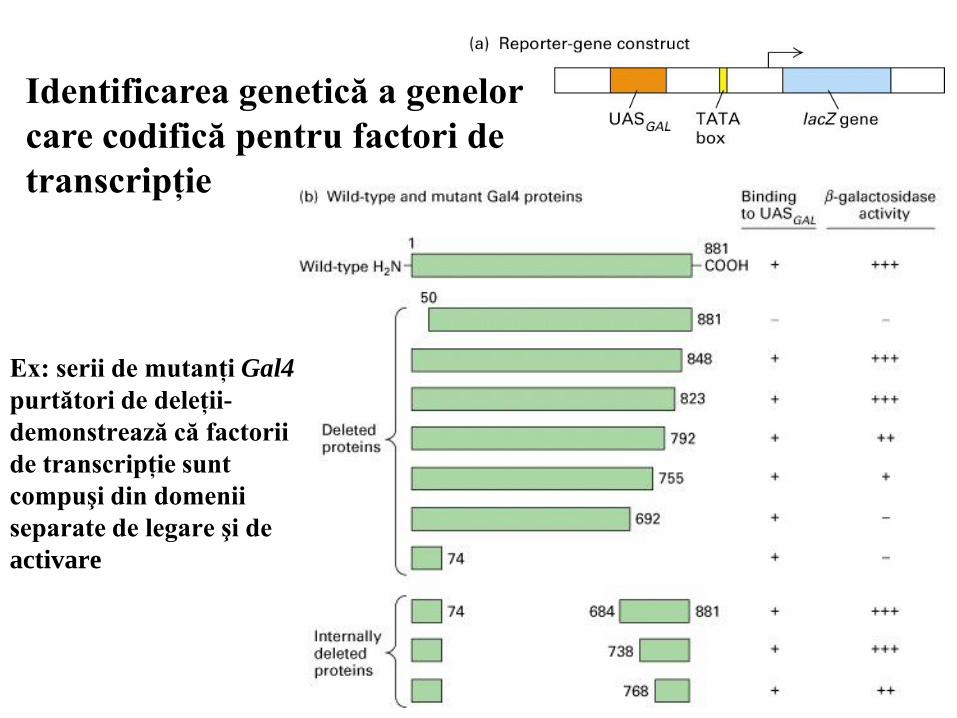
Identificarea genetică a genelor
care codifică pentru factori de
transcripţie
Ex: serii de mutanţi Gal4
purtători de deleţii-
demonstrează că factorii
de transcripţie sunt
compuşi din domenii
separate de legare şi de
activare
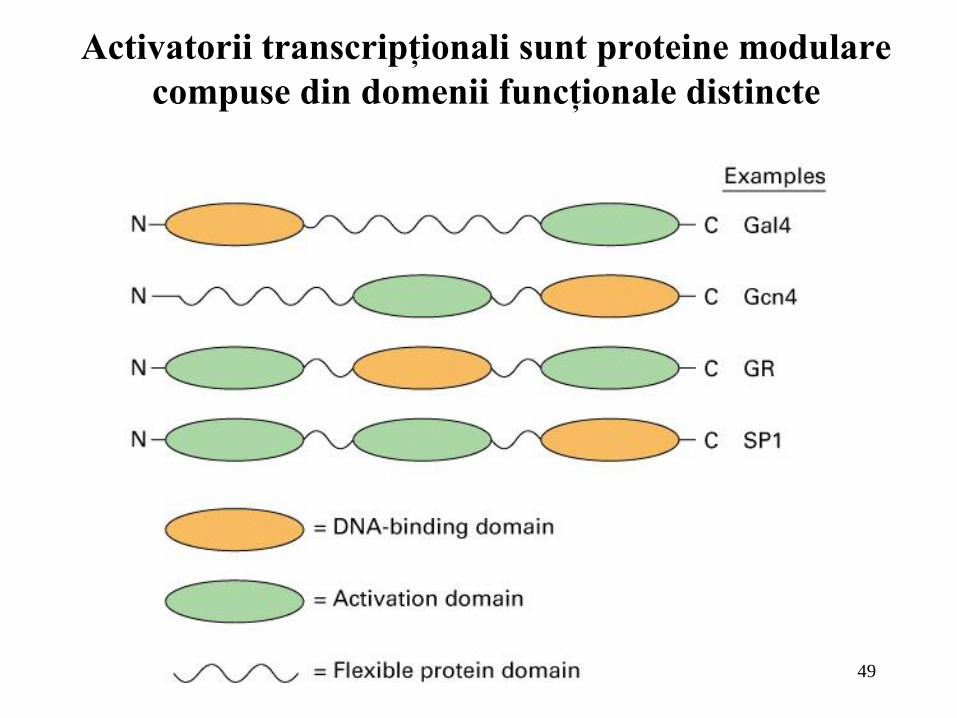
49
Activatorii transcripţionali sunt proteine modulare
compuse din domenii funcţionale distincte

50
Domeniile de legare la ADN pot fi clasificate în
mai multe tipuri structurale
Proteine care conţin homeodomenii
Proteine “Zinc-finger”
Proteine “Winged-helix (forkhead)”
Proteine “Leucine-zipper”
Proteine “Helix-loop-helix”
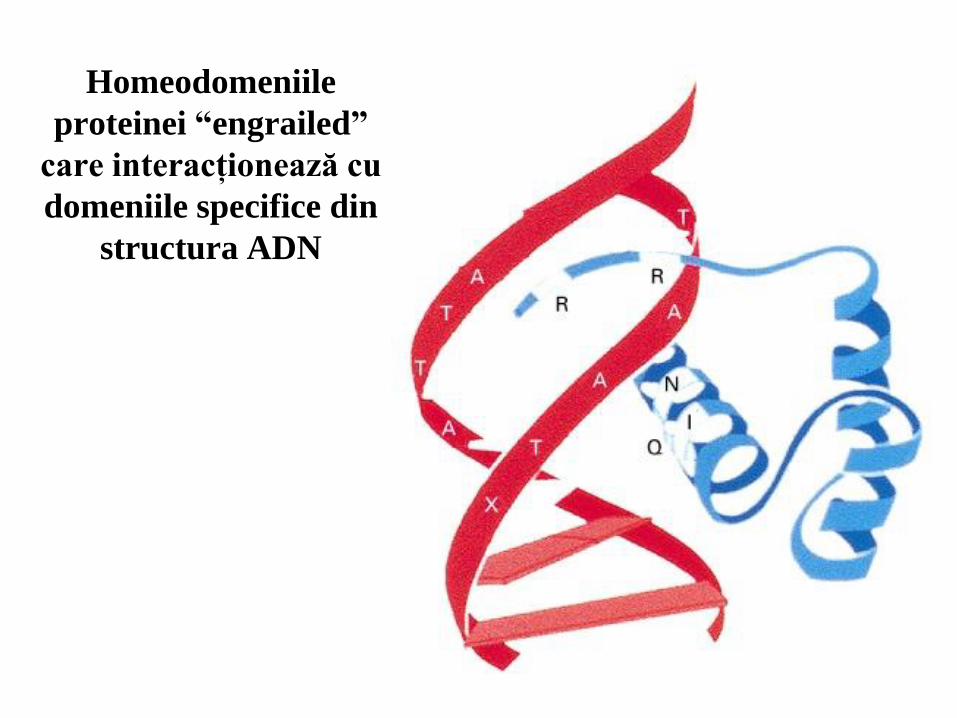
51
Homeodomeniile
proteinei “engrailed”
care interacţionează cu
domeniile specifice din
structura ADN
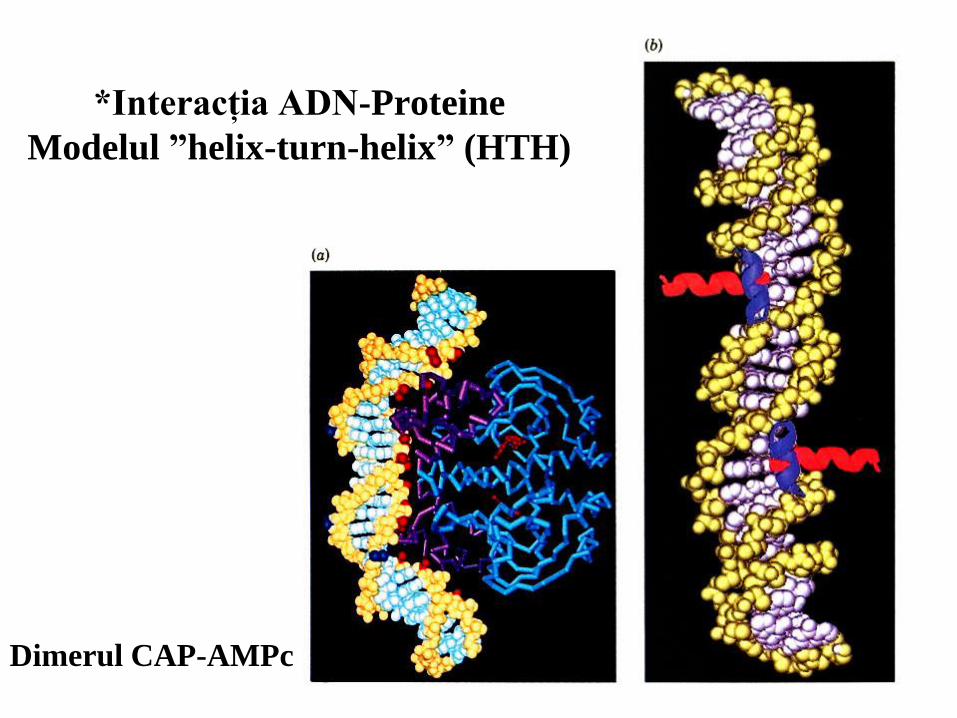
52
*Interacţia ADN-Proteine
Modelul ”helix-turn-helix” (HTH)
Dimerul CAP-AMPc
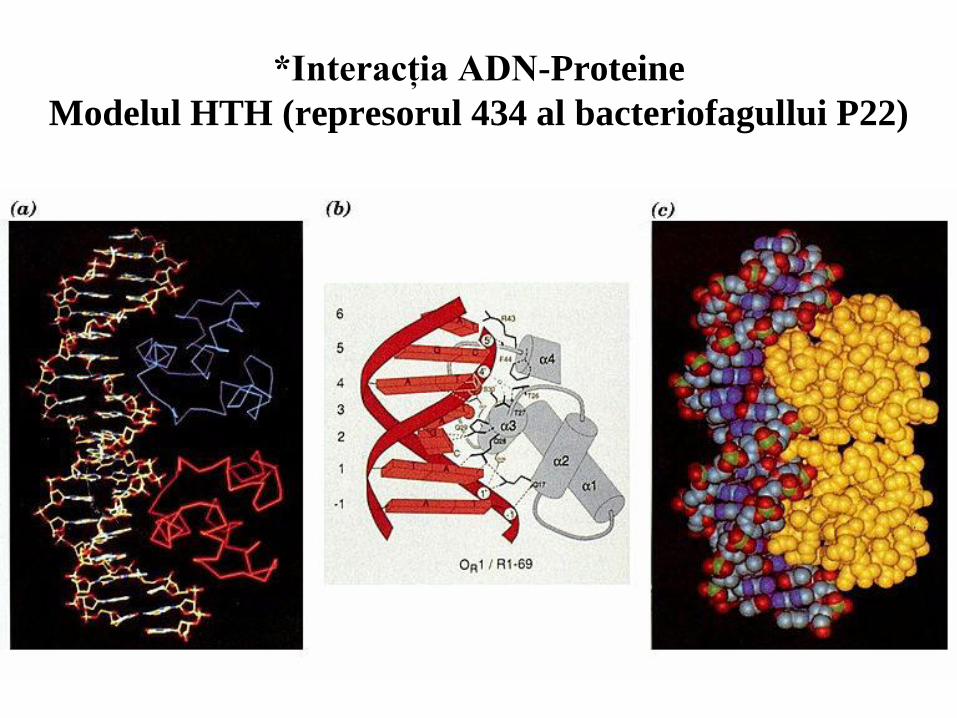
53
*Interacţia ADN-Proteine
Modelul HTH (represorul 434 al bacteriofagullui P22)
54
*Interacţia ADN -
Proteine
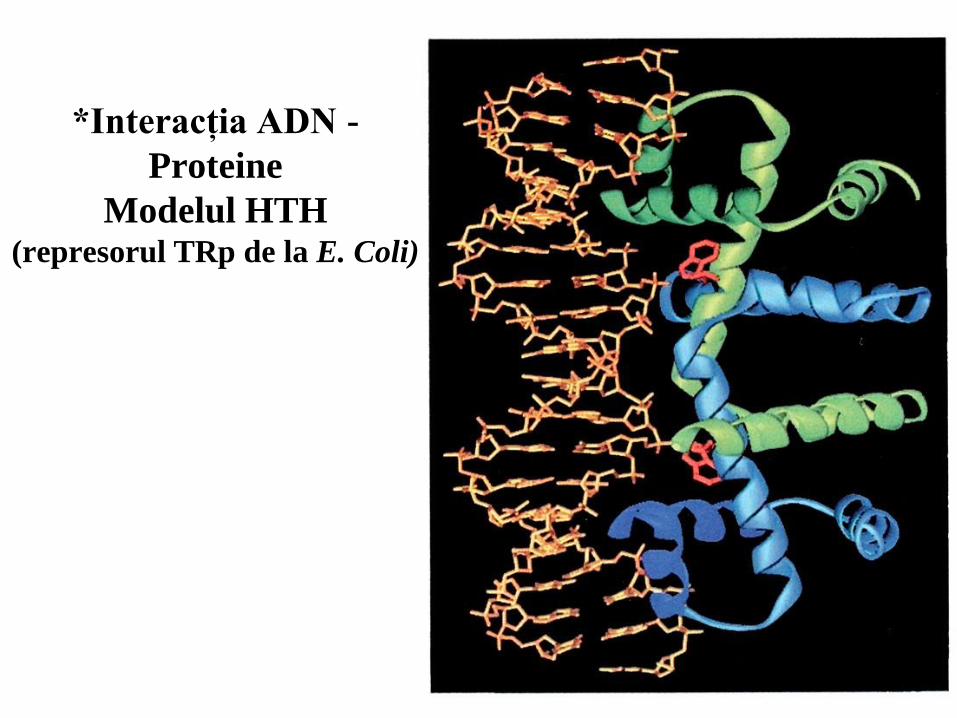
Modelul HTH (represorul TRp de la E. Coli)
55
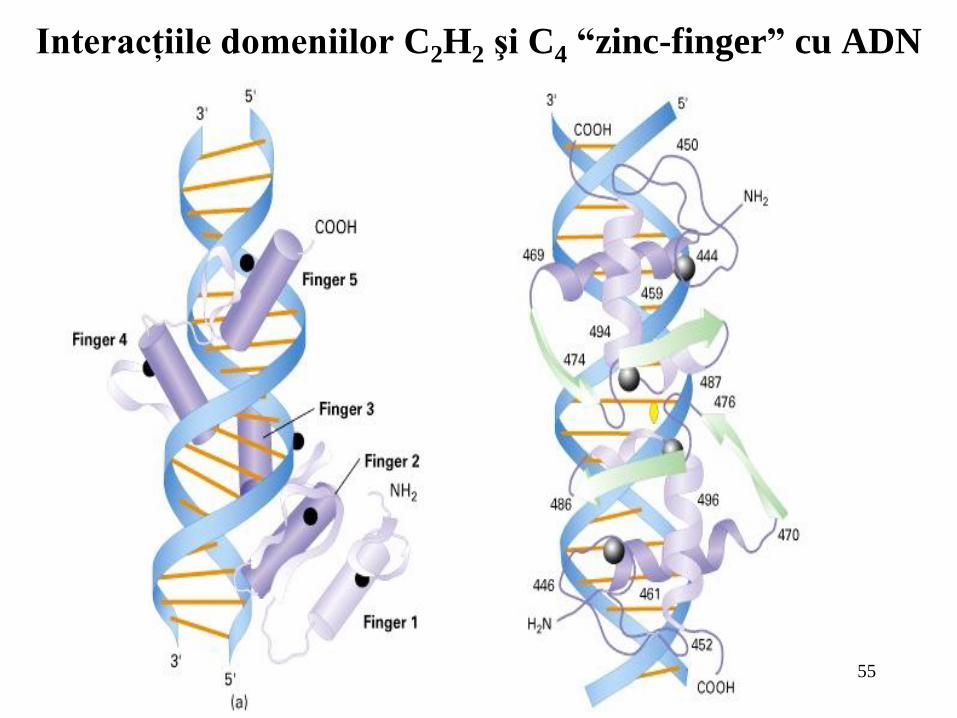
Interacţiile domeniilor C2H2 şi C4 “zinc-finger” cu ADN
56
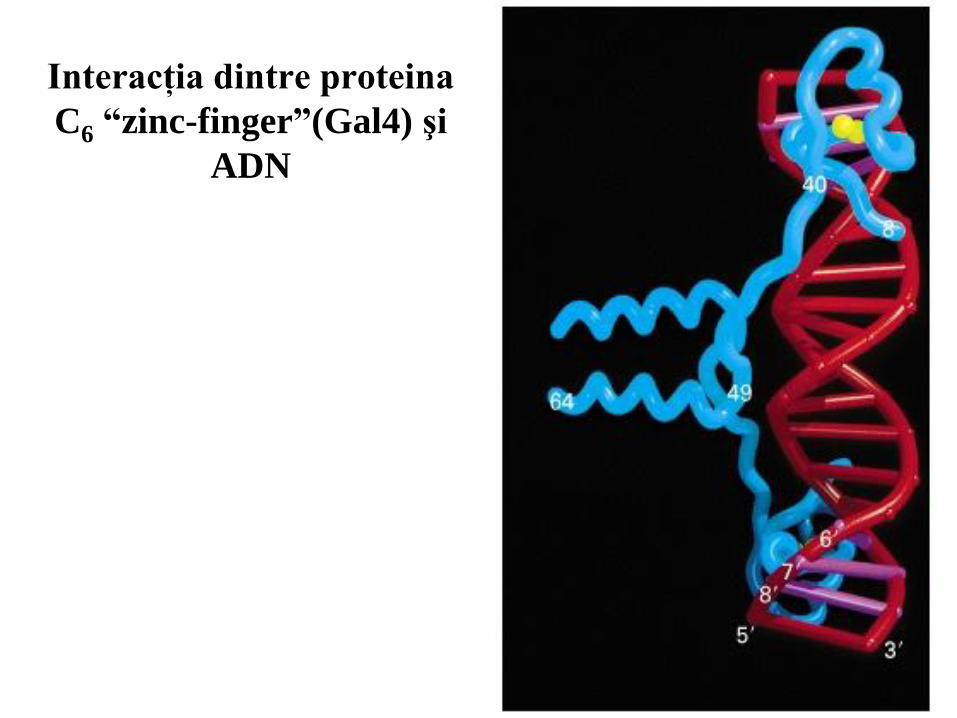
Interacţia dintre proteina
C6 “zinc-finger”(Gal4) şi
ADN
57
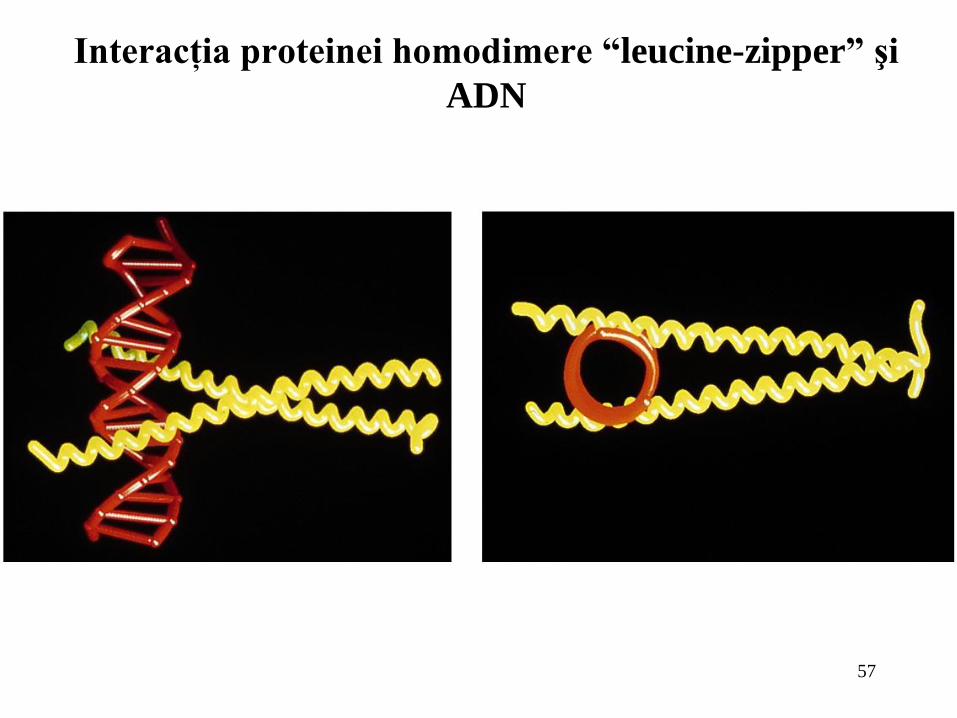
Interacţia proteinei homodimere “leucine-zipper” şi
ADN
58
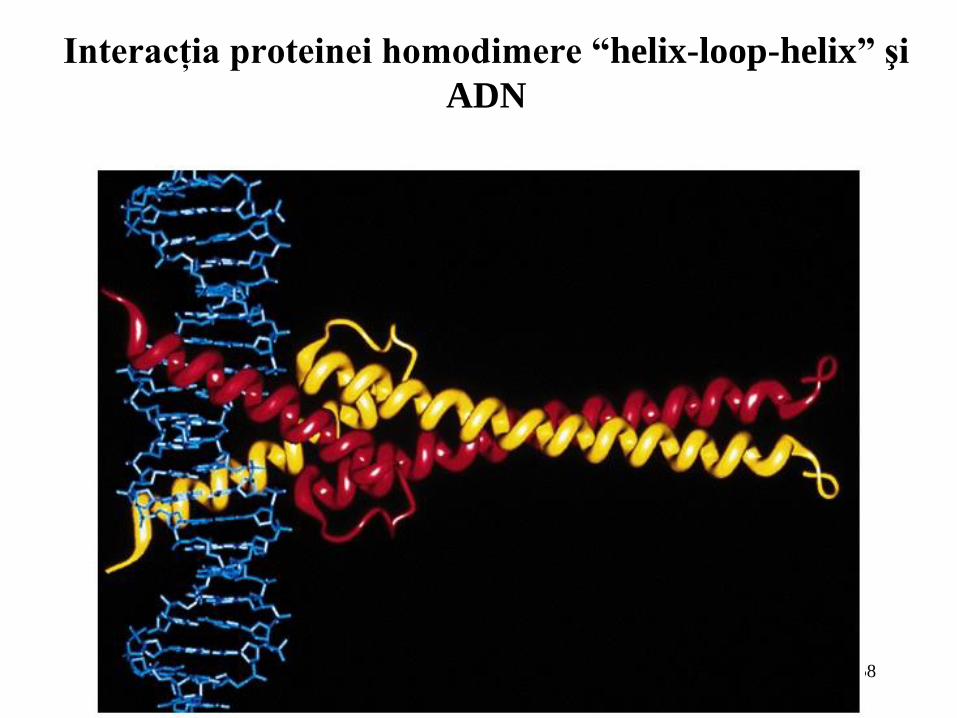
Interacţia proteinei homodimere “helix-loop-helix” şi
ADN
59
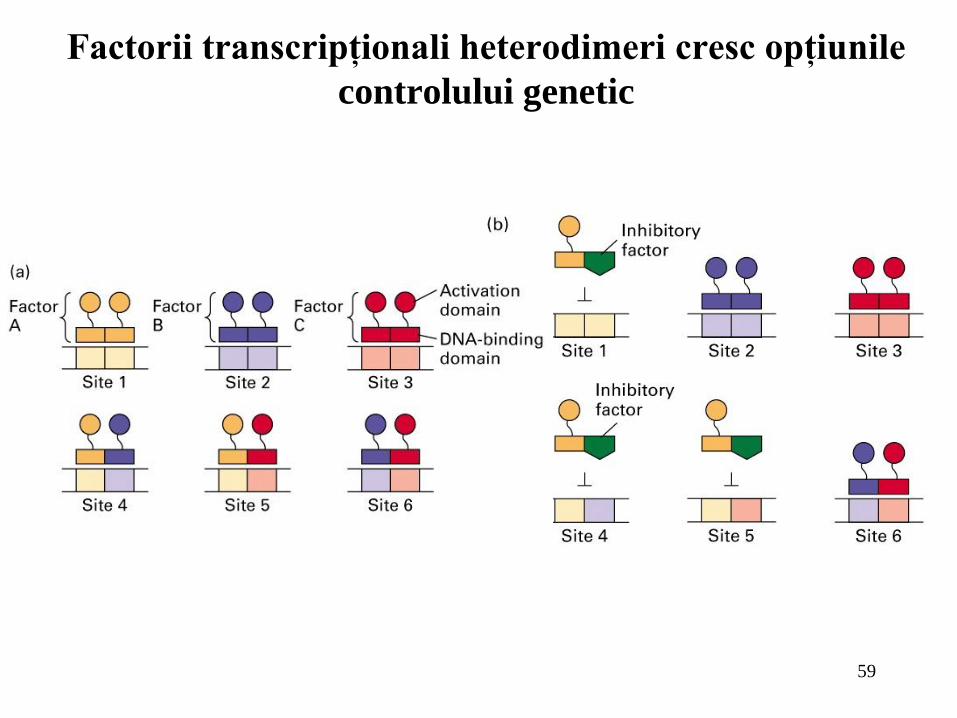
Factorii transcripţionali heterodimeri cresc opţiunile
controlului genetic
60
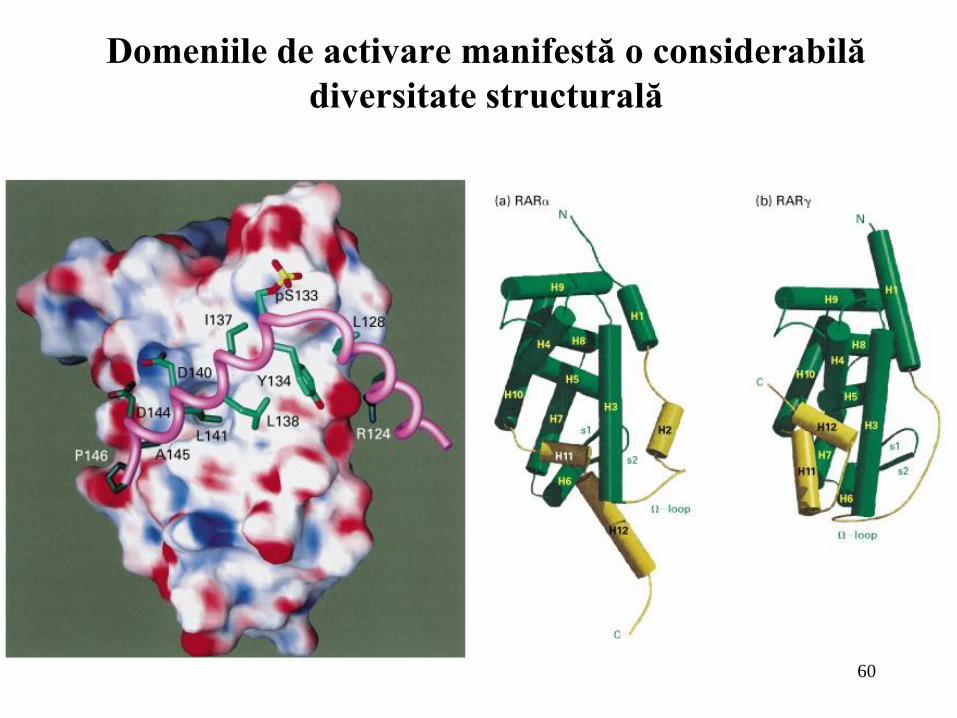
Domeniile de activare manifestă o considerabilă
diversitate structurală
61
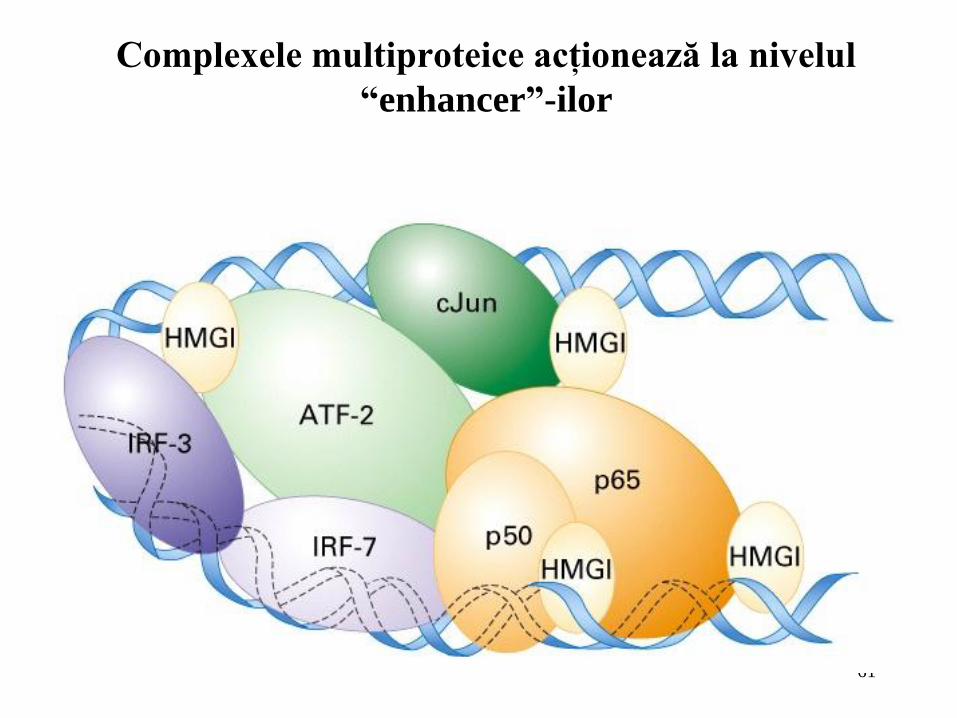
Complexele multiproteice acţionează la nivelul
“enhancer”-ilor

62
Mulţi represori interacţionează cu activatorii
Transcripţia de la eucariote este reglată de represori
cât şi de activatori ;
Situsurile de legare a represorilor pot fi
identificate şi represorii purificaţi prin aceleaşi tehnici folosite
pentru activatori;
Mulţi represori din sistemele eucariote posedă două
domenii: un domeniu de legare a ADN şi un domeniu represor.

63
Concluzii Factorii de transcripţie, care stimulează sau reprimă transcripţia, se leagă la elemente situate în proximitatea promotorului şi la “enhancers”(activatori) din structura ADN la eucariote;
Activatorii sunt în general proteine modulare, care conţin un singur domeniu de legare şi unul sau mai multe domenii activatoare; diferitele domenii sunt adesea legate prin regiuni polipeptidice flexibile. Acestea permit domeniilor diferiţilor activatori să interacţioneze chiar şi când domeniile de legare la ADN recunosc situsuri separate de zeci de perechi de baze;
Activatorii conţin, în general, mai multe situsuri de legare pentru factorii de transcripţie
grupate sub formă de clustere. Legarea cooperativă a mai multor activatori la situsuri învecinate din structura unui element activator formează un complex multi-proteic numit “complex activator”. Asamblarea acestuia necesită adesea proteine mici care se lreagă la fosa minoră a ADN şi determină curbarea rapidă a secventei permitând proteinelor de pe fiecare parte a curburii să interacţioneze mult mai bine;
Majoritatea represorilor de la eucariote sunt proteine modulare. Similar cu activatorii, aceştia conţin de obicei un singur domeniu de legare şi unul sau mai multe domenii represoare, şi pot controla transcripţia când sunt legaţi la situsuri situate la sute sau mii de baze faţă de situsul de start;
Domeniile de legare la ADN ale factorilor de transcripţie de la eucariote exprimă o varietate de structuri. Printre cele mai comune motive structurale sunt homeodomeniile, domeniile bazice “fermoar de leucină” (leucine zipper), HLH şi diferite tipuri de degete zinc (zinc finger). În general una sau mai multe alfa-elice din domeniul de legare la ADN interacţionează cu fosa majoră la nivelul unor situsuri de recunoaştere;
Capacitatea anumitor factori de transcripţie de a forma heterodimeri creşte numărul de situsuri ADN pe care aceşti factori le pot controla şi modalităţile prin care le pot controla;
Deşi anumite domenii de activare şi de represare sunt bogate în aminoacizi particulari, aceste domenii funcţionale manifestă o varietate de secvenţe de aminoacizi şi structuri proteice caracteristice diferiţilor factori transcripţie.

64
Complexul ARN polimerazic II de iniţiere a
transcripţiei
Iniţierea de către Pol II necesită factori generali de
transcripţie, care poziţionează Pol II la nivelul situsului de
iniţiere şi sunt necesari pentru transcrierea marii majorităţi
a genelor transcrise de către această polimerază;
Factorii generali de transcripţie sunt structuri
multimere şi înalt conservate;
Proteinele cuprinse în complexul Pol II de iniţiere a
transcripţiei se asamblează într-o ordine specifică “in vitro”
dar majoritatea proteinelor se pot combina pentru a forma un
complex holoenzimatic “in vivo”.
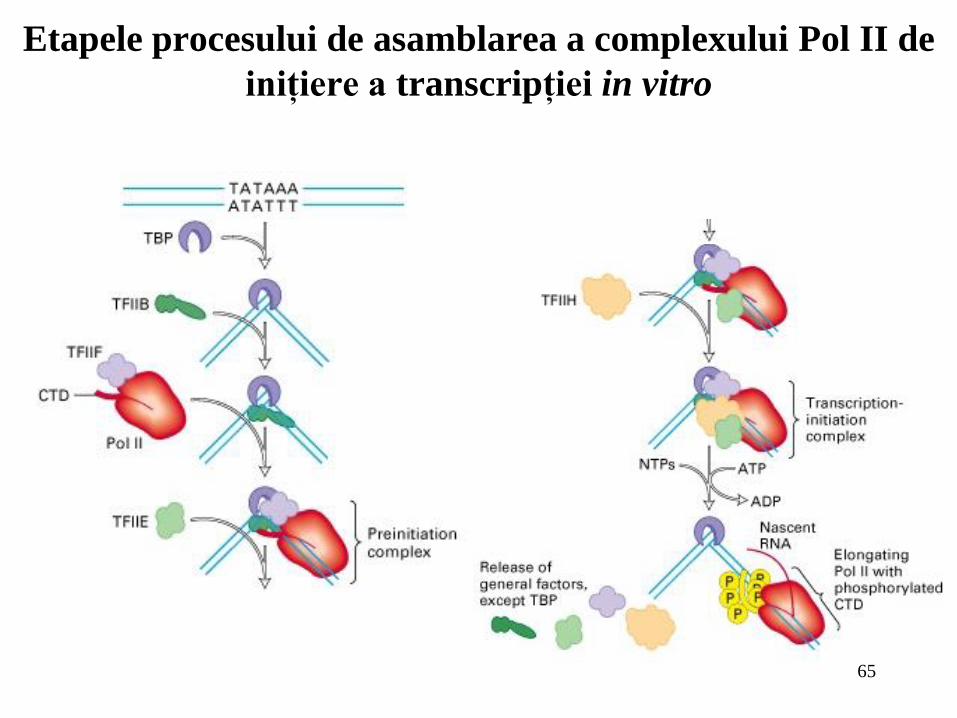
65
Etapele procesului de asamblarea a complexului Pol II de
iniţiere a transcripţiei in vitro

66
In vitro assembly of RNA
polymerase II preinitiation
complex
Factorii de transcriptie generali se leaga secvential la
ARN polimeraza II purificata (Pol II), la nivelul
secventei ADN TATAbox pentru a forma complexul de
preinitiatiere. Hidroliza ATP furnizeaza energia
pentru desfacerea moleculelor de ADN la situsul de
start prin legarea subunitatii TFIIH subunit. Initierea
transcriptiei de catre Pol II conduce la formarea
complexului deschis, iar polimeraza se deplaseaza de
la nivelul promotorului iar domeniul CTD este
fosforilat. In vitro, factorii generali de transcriptie (cu
exceptia TBP) disociaza de complexul TBP si
promotor, dar nu se stie inca care dintre factori
raman asociati cu regiunea promotoare pentru fiecare
runda de initiere a transcriptiei, in vivo.
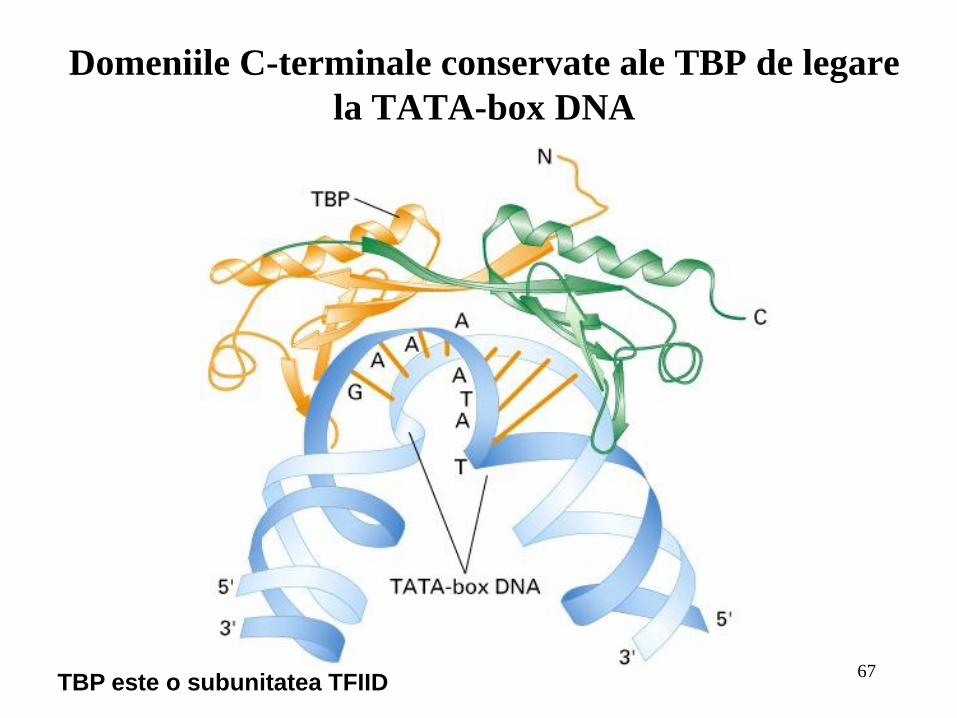
67
Domeniile C-terminale conservate ale TBP de legare
la TATA-box DNA
TBP este o subunitatea TFIID
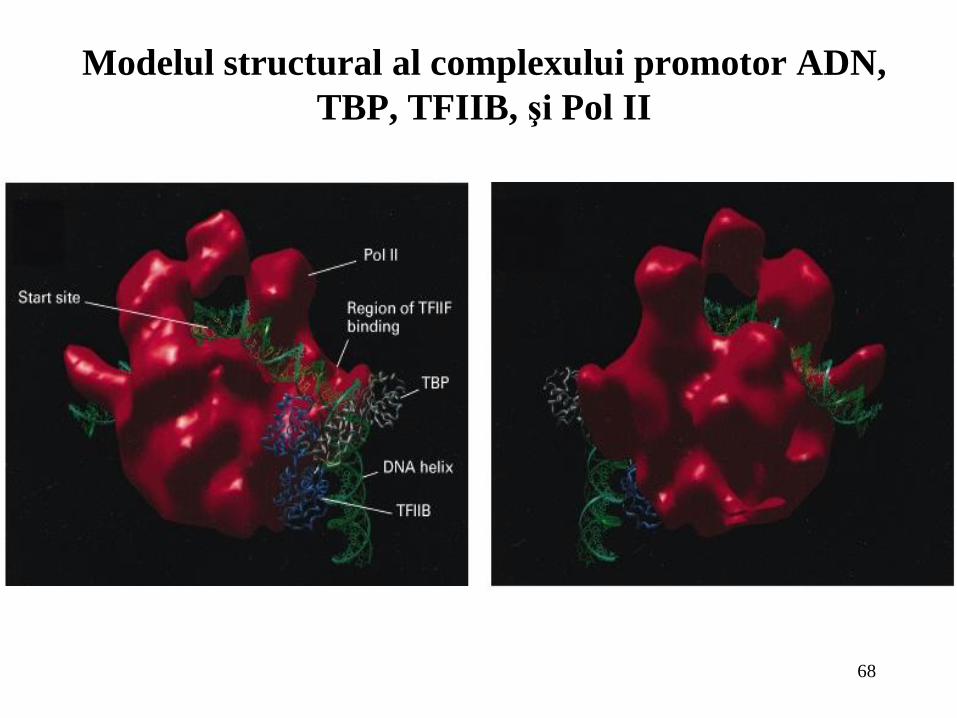
68
Modelul structural al complexului promotor ADN,
TBP, TFIIB, şi Pol II

69
Concluzii
ARN polimeraza II necesită mai mulţi factori generali de transcripţie pentru a
localiza clar punctul de start la nivelul matriţei ADN şi a iniţia transcripţia. Acestea includ
TFIID care se leagă la TATA-box prin intermediul TBP;
Transcripţia genelor care codifică pentru proteine este controlată de către ARN
polimeraza II care poate fi iniţiată in vitro prin legarea secvenţială a factorilor: TBP, care se
leagă la TATA-box; TFIIB; un complex Pol II şi TFIIF; TFIIE; şi final TFIIH;
Activităţile helicazice ale celor două subunităţi TFIIH separă catenele la nivelul
situsului de start la majoritatea promotorilor, un proces care necesită hidroliza ATP. După
ce pol II începe transcrierea dincolo de situsul de start, elementele CTD sunt fosforilate de
către o altă subunitate a TFIIH;
Iniţierea de către Pol II in vivo necesită un complex multimediator multiproteic,
care se asociază cu elementele CTD nefosforilate ale Pol II, formând un mare complex
holoenzimatic care include şi majoritatea factorilor de transcripţie. Se crede că această
holoenzimă preasamblată se leagă la ADN promotor într-o singură etapă in vivo;
Complexul de iniţiere a transcripţiei care se asamblează la nivelul promotorilor in
vivo poate cuprinde 60-70 polipeptide cu o masă totală similară cu cea a unui ribozom.

70
Formarea heterocromatinei încetineşte expresia la nivelul
telomerilor şi altor regiuni
Mecanismele moleculare ale controlului transcripţional
la eucariote
Concentraţia şi activitatea activatorilor şi represorilor reglează structura
cromatinei şi acetilarea-deacetilarea histonelor, cât şi asamblarea complexelor de
iniţiere a transcripţiei şi viteza cu care transcripţia este iniţiată
Genele silenţioase în controlul tip mating la drojdii
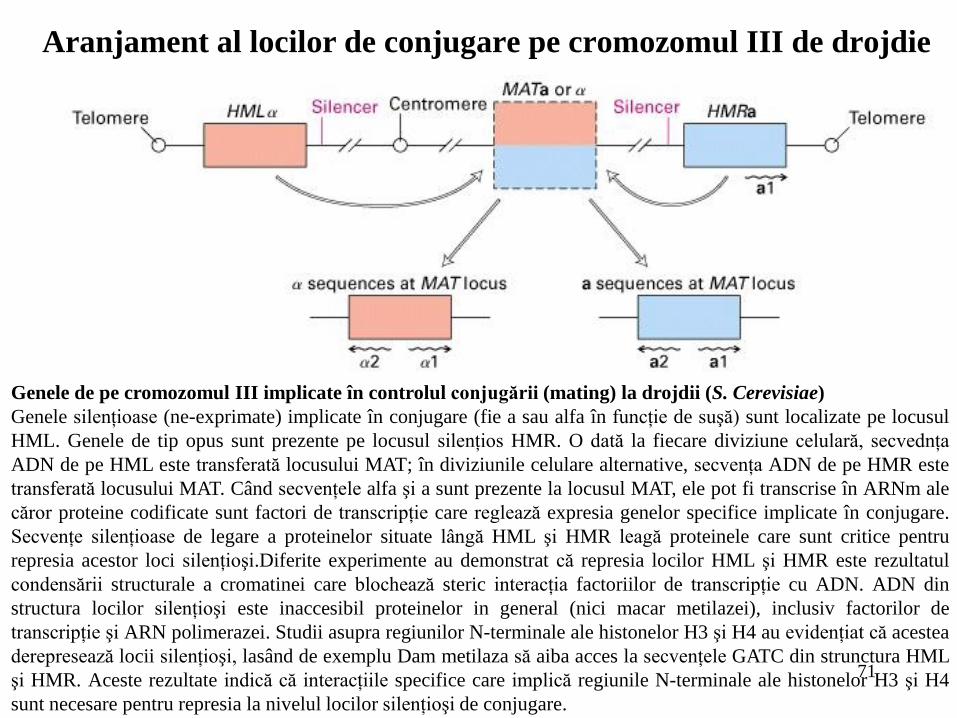
71
Genele de pe cromozomul III implicate în controlul conjugării (mating) la drojdii (S. Cerevisiae)
Genele silenţioase (ne-exprimate) implicate în conjugare (fie a sau alfa în funcţie de suşă) sunt localizate pe locusul
HML. Genele de tip opus sunt prezente pe locusul silenţios HMR. O dată la fiecare diviziune celulară, secvednţa
ADN de pe HML este transferată locusului MAT; în diviziunile celulare alternative, secvenţa ADN de pe HMR este
transferată locusului MAT. Când secvenţele alfa şi a sunt prezente la locusul MAT, ele pot fi transcrise în ARNm ale
căror proteine codificate sunt factori de transcripţie care reglează expresia genelor specifice implicate în conjugare.
Secvenţe silenţioase de legare a proteinelor situate lângă HML şi HMR leagă proteinele care sunt critice pentru
represia acestor loci silenţioşi.Diferite experimente au demonstrat că represia locilor HML şi HMR este rezultatul
condensării structurale a cromatinei care blochează steric interacţia factoriilor de transcripţie cu ADN. ADN din
structura locilor silenţioşi este inaccesibil proteinelor in general (nici macar metilazei), inclusiv factorilor de
transcripţie şi ARN polimerazei. Studii asupra regiunilor N-terminale ale histonelor H3 şi H4 au evidenţiat că acestea
derepresează locii silenţioşi, lasând de exemplu Dam metilaza să aiba acces la secvenţele GATC din strunctura HML
şi HMR. Aceste rezultate indică că interacţiile specifice care implică regiunile N-terminale ale histonelor H3 şi H4
sunt necesare pentru represia la nivelul locilor silenţioşi de conjugare.
Aranjament al locilor de conjugare pe cromozomul III de drojdie
72
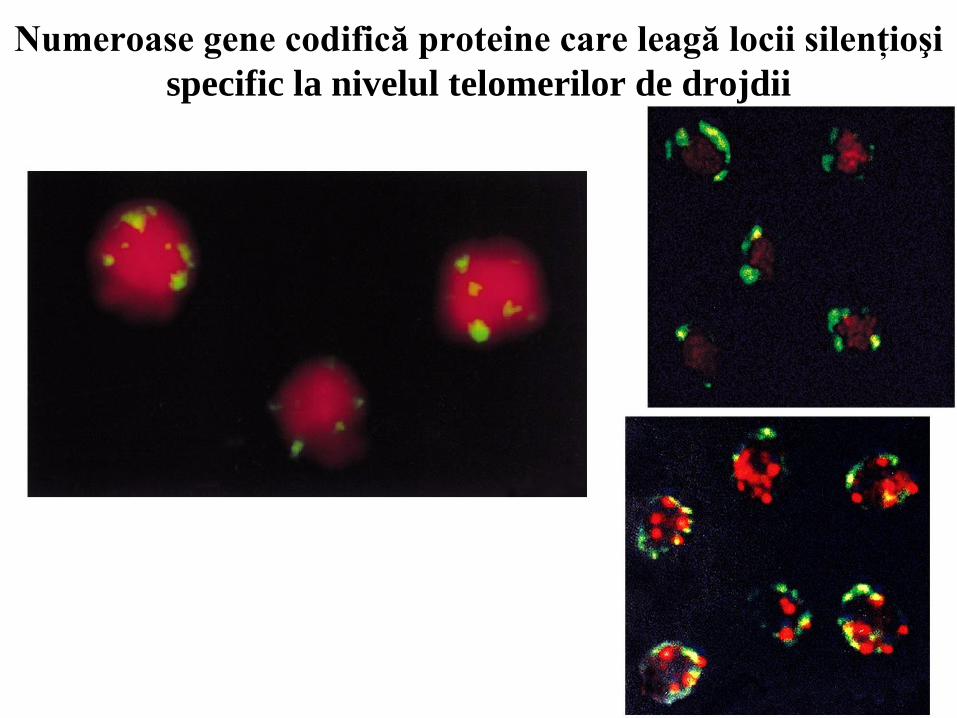
Numeroase gene codifică proteine care leagă locii silenţioşi
specific la nivelul telomerilor de drojdii
73
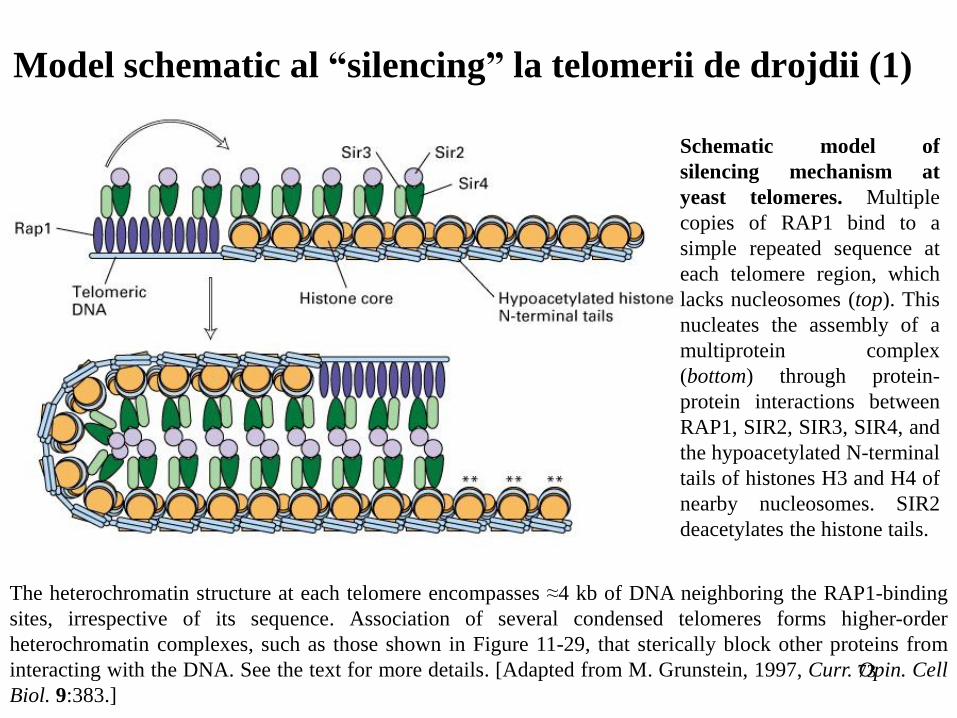
Model schematic al “silencing” la telomerii de drojdii (1)
The heterochromatin structure at each telomere encompasses ≈4 kb of DNA neighboring the RAP1-binding
sites, irrespective of its sequence. Association of several condensed telomeres forms higher-order
heterochromatin complexes, such as those shown in Figure 11-29, that sterically block other proteins from
interacting with the DNA. See the text for more details. [Adapted from M. Grunstein, 1997, Curr. Opin. Cell
Biol. 9:383.]
Schematic model of
silencing mechanism at
yeast telomeres. Multiple
copies of RAP1 bind to a
simple repeated sequence at
each telomere region, which
lacks nucleosomes (top). This
nucleates the assembly of a
multiprotein complex
(bottom) through protein-
protein interactions between
RAP1, SIR2, SIR3, SIR4, and
the hypoacetylated N-terminal
tails of histones H3 and H4 of
nearby nucleosomes. SIR2
deacetylates the histone tails.
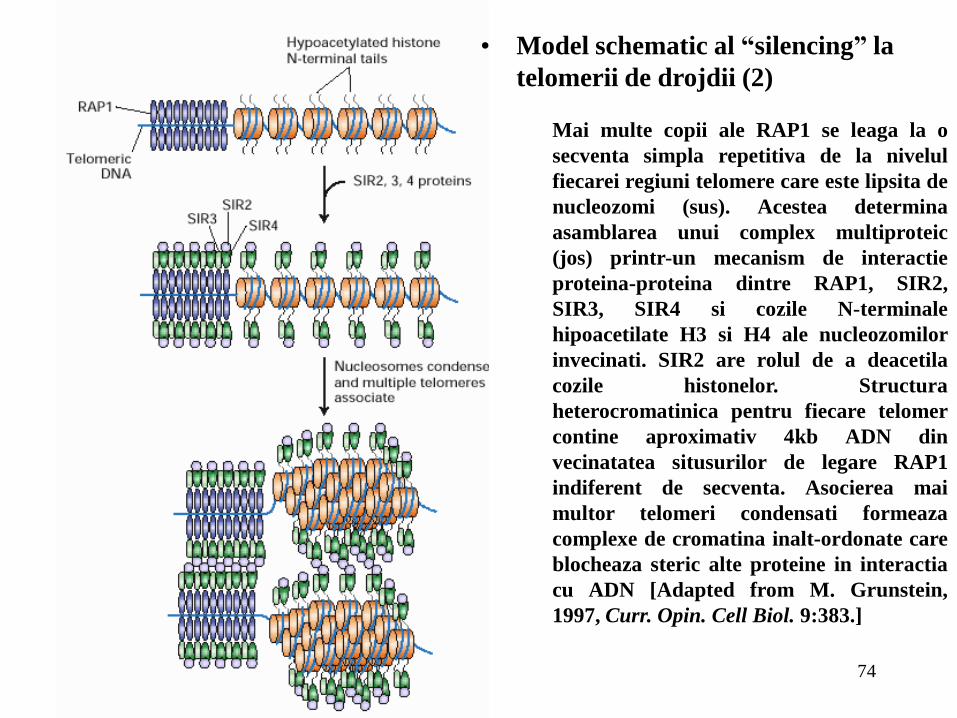
74
Mai multe copii ale RAP1 se leaga la o
secventa simpla repetitiva de la nivelul
fiecarei regiuni telomere care este lipsita de
nucleozomi (sus). Acestea determina
asamblarea unui complex multiproteic
(jos) printr-un mecanism de interactie
proteina-proteina dintre RAP1, SIR2,
SIR3, SIR4 si cozile N-terminale
hipoacetilate H3 si H4 ale nucleozomilor
invecinati. SIR2 are rolul de a deacetila
cozile histonelor. Structura
heterocromatinica pentru fiecare telomer
contine aproximativ 4kb ADN din
vecinatatea situsurilor de legare RAP1
indiferent de secventa. Asocierea mai
multor telomeri condensati formeaza
complexe de cromatina inalt-ordonate care
blocheaza steric alte proteine in interactia
cu ADN [Adapted from M. Grunstein,
1997, Curr. Opin. Cell Biol. 9:383.]
• Model schematic al “silencing” la
telomerii de drojdii (2)
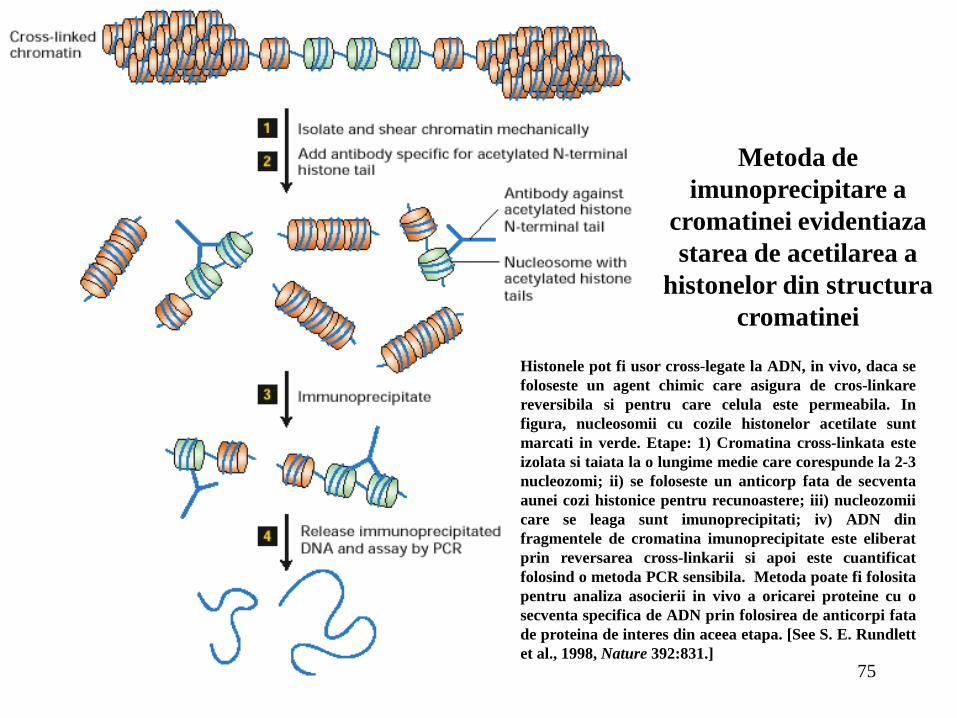
75
Histonele pot fi usor cross-legate la ADN, in vivo, daca se
foloseste un agent chimic care asigura de cros-linkare
reversibila si pentru care celula este permeabila. In
figura, nucleosomii cu cozile histonelor acetilate sunt
marcati in verde. Etape: 1) Cromatina cross-linkata este
izolata si taiata la o lungime medie care corespunde la 2-3
nucleozomi; ii) se foloseste un anticorp fata de secventa
aunei cozi histonice pentru recunoastere; iii) nucleozomii
care se leaga sunt imunoprecipitati; iv) ADN din
fragmentele de cromatina imunoprecipitate este eliberat
prin reversarea cross-linkarii si apoi este cuantificat
folosind o metoda PCR sensibila. Metoda poate fi folosita
pentru analiza asocierii in vivo a oricarei proteine cu o
secventa specifica de ADN prin folosirea de anticorpi fata
de proteina de interes din aceea etapa. [See S. E. Rundlett
et al., 1998, Nature 392:831.]
Metoda de
imunoprecipitare a
cromatinei evidentiaza
starea de acetilarea a
histonelor din structura
cromatinei
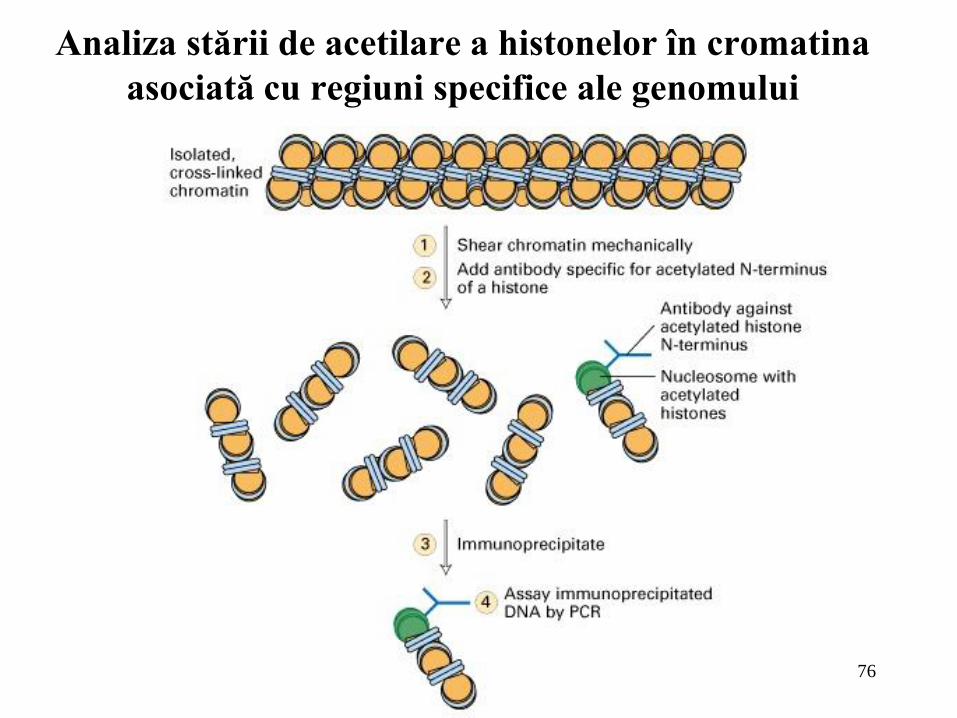
76
Analiza stării de acetilare a histonelor în cromatina
asociată cu regiuni specifice ale genomului
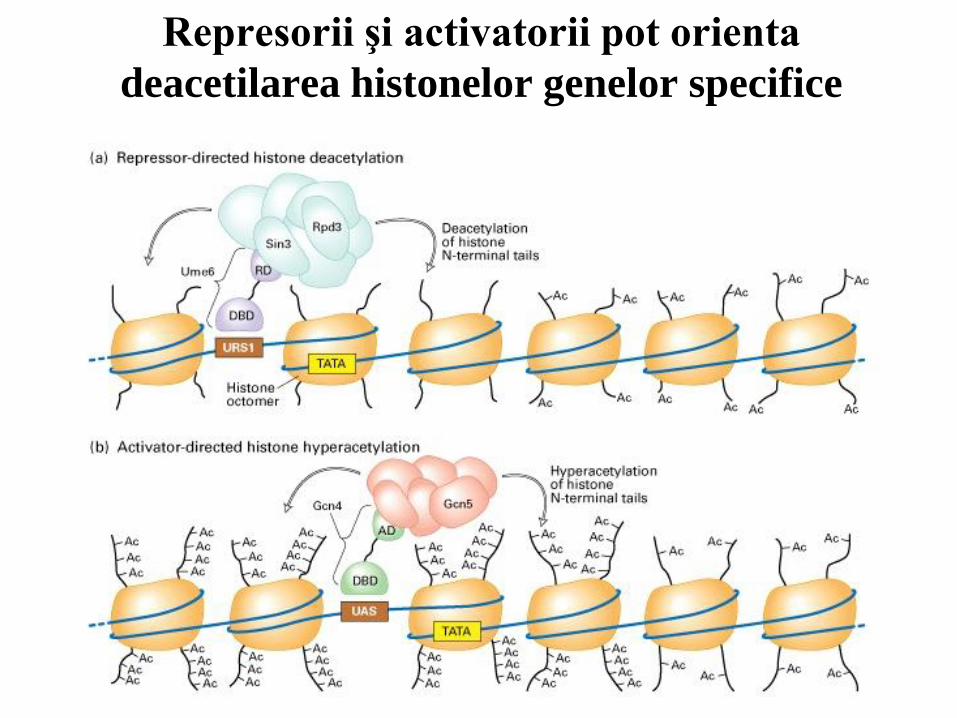
77
Represorii şi activatorii pot orienta
deacetilarea histonelor genelor specifice
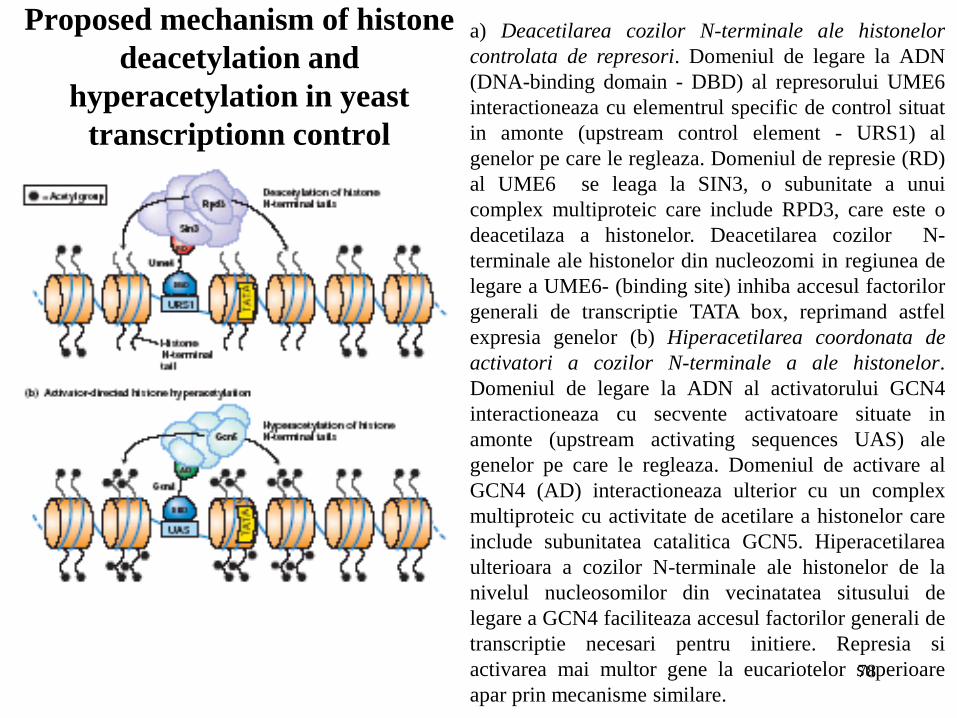
78
Proposed mechanism of histone
deacetylation and
hyperacetylation in yeast
transcriptionn control
a) Deacetilarea cozilor N-terminale ale histonelor
controlata de represori. Domeniul de legare la ADN
(DNA-binding domain - DBD) al represorului UME6
interactioneaza cu elementrul specific de control situat
in amonte (upstream control element - URS1) al
genelor pe care le regleaza. Domeniul de represie (RD)
al UME6 se leaga la SIN3, o subunitate a unui
complex multiproteic care include RPD3, care este o
deacetilaza a histonelor. Deacetilarea cozilor N-
terminale ale histonelor din nucleozomi in regiunea de
legare a UME6- (binding site) inhiba accesul factorilor
generali de transcriptie TATA box, reprimand astfel
expresia genelor (b) Hiperacetilarea coordonata de
activatori a cozilor N-terminale a ale histonelor.
Domeniul de legare la ADN al activatorului GCN4
interactioneaza cu secvente activatoare situate in
amonte (upstream activating sequences UAS) ale
genelor pe care le regleaza. Domeniul de activare al
GCN4 (AD) interactioneaza ulterior cu un complex
multiproteic cu activitate de acetilare a histonelor care
include subunitatea catalitica GCN5. Hiperacetilarea
ulterioara a cozilor N-terminale ale histonelor de la
nivelul nucleosomilor din vecinatatea situsului de
legare a GCN4 faciliteaza accesul factorilor generali de
transcriptie necesari pentru initiere. Represia si
activarea mai multor gene la eucariotelor superioare
apar prin mecanisme similare.
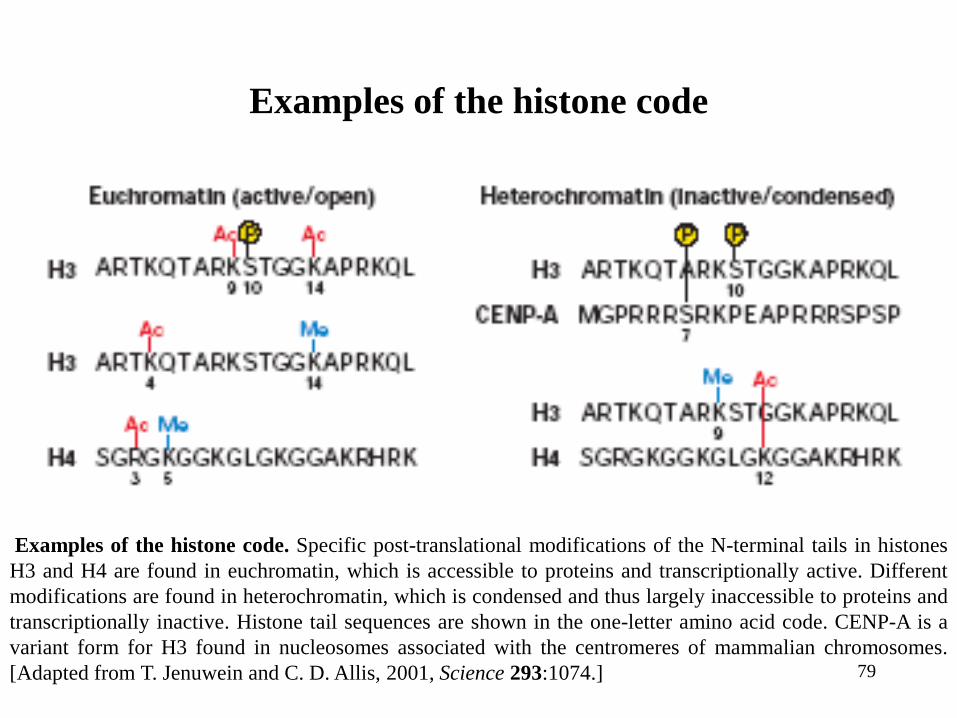
79
Examples of the histone code
Examples of the histone code. Specific post-translational modifications of the N-terminal tails in histones
H3 and H4 are found in euchromatin, which is accessible to proteins and transcriptionally active. Different
modifications are found in heterochromatin, which is condensed and thus largely inaccessible to proteins and
transcriptionally inactive. Histone tail sequences are shown in the one-letter amino acid code. CENP-A is a
variant form for H3 found in nucleosomes associated with the centromeres of mammalian chromosomes.
[Adapted from T. Jenuwein and C. D. Allis, 2001, Science 293:1074.]
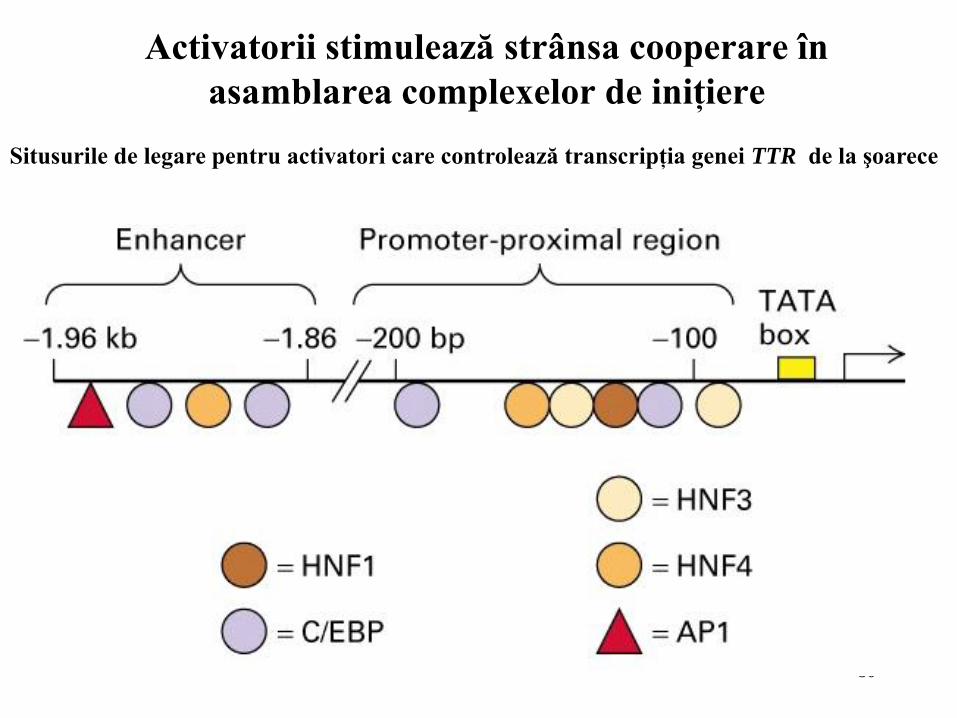
80
Activatorii stimulează strânsa cooperare în
asamblarea complexelor de iniţiere
Situsurile de legare pentru activatori care controlează transcripţia genei TTR de la şoarece

81
Structure of yeast and human mediator
complexes. (a) Reconstructed image of
mediator from S. cerevisiae bound to Pol
II. Multiple electron microscopy images
were aligned and computer-processed to
produce this average image in which the
three-dimensional Pol II structure (light
blue) is shown associated with the yeast
mediator complex (dark blue).
(b) Diagrammatic representation of
mediator subunits from human cells.
Subunits shown in the same color are
thought to form a module. Subunits in
orange, yellow, and green are homologous
with subunits in the yeast mediator
complex. Genetic studies in yeast show
that mutations in one of the subunits in a
module inhibit the association of other
subunits in the same module with the rest
of the complex. [Part (a) courtesy of
Francisco J. Asturias, 2002, Mol. Cell
10:409. Part (b) adapted from S. Malik
and R. G. Roeder, 2000, Trends Biochem.
Sci. 25:277.]
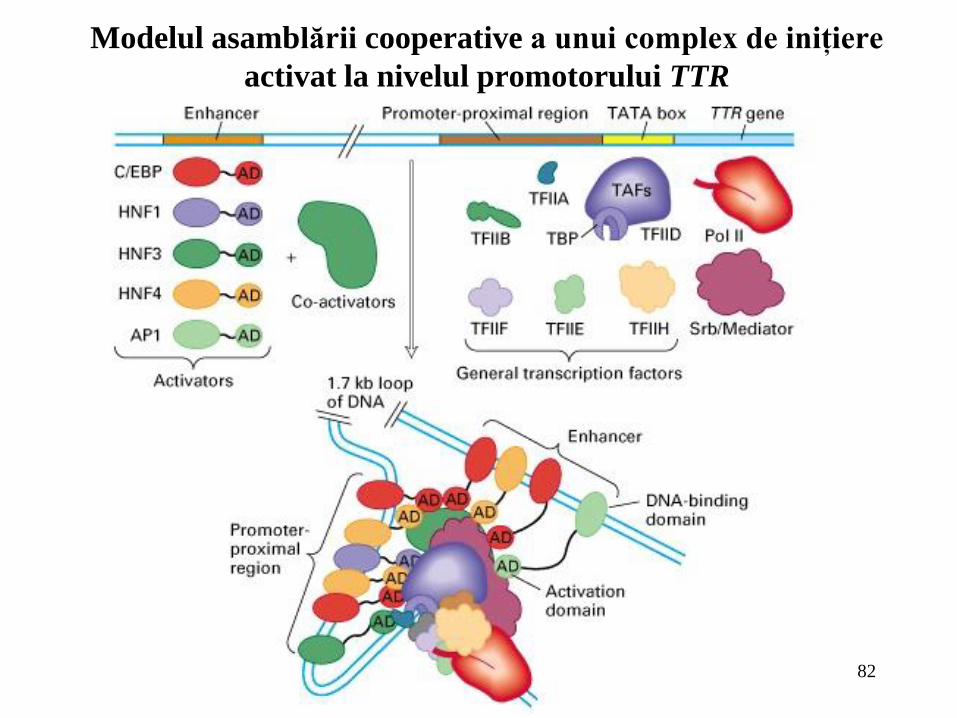
82
Modelul asamblării cooperative a unui complex de iniţiere
activat la nivelul promotorului TTR
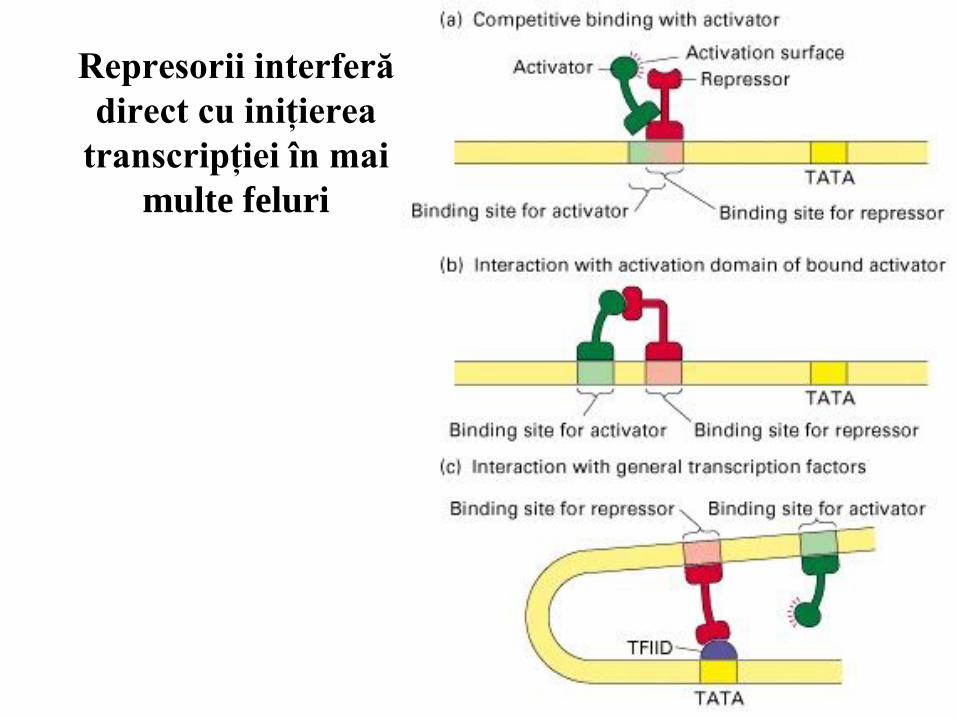
83
Represorii interferă
direct cu iniţierea
transcripţiei în mai
multe feluri
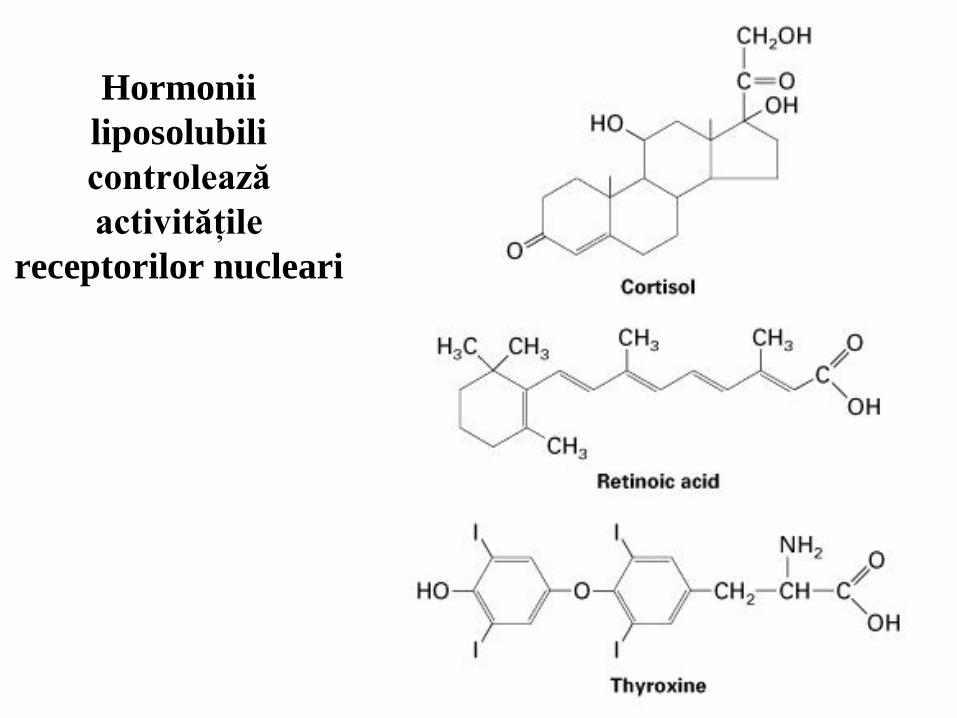
84
Hormonii
liposolubili
controlează
activităţile
receptorilor nucleari
85
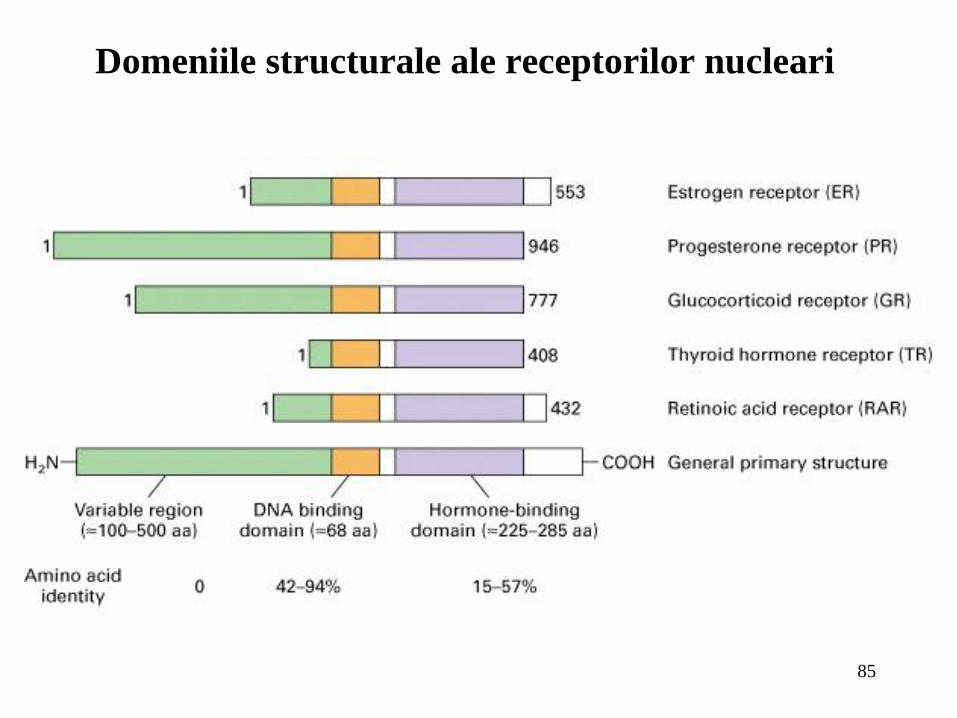
Domeniile structurale ale receptorilor nucleari

86
“Response
elements” sunt
secvenţe de ADN
care leagă mai
mulţi receptori
nucleari majori
87
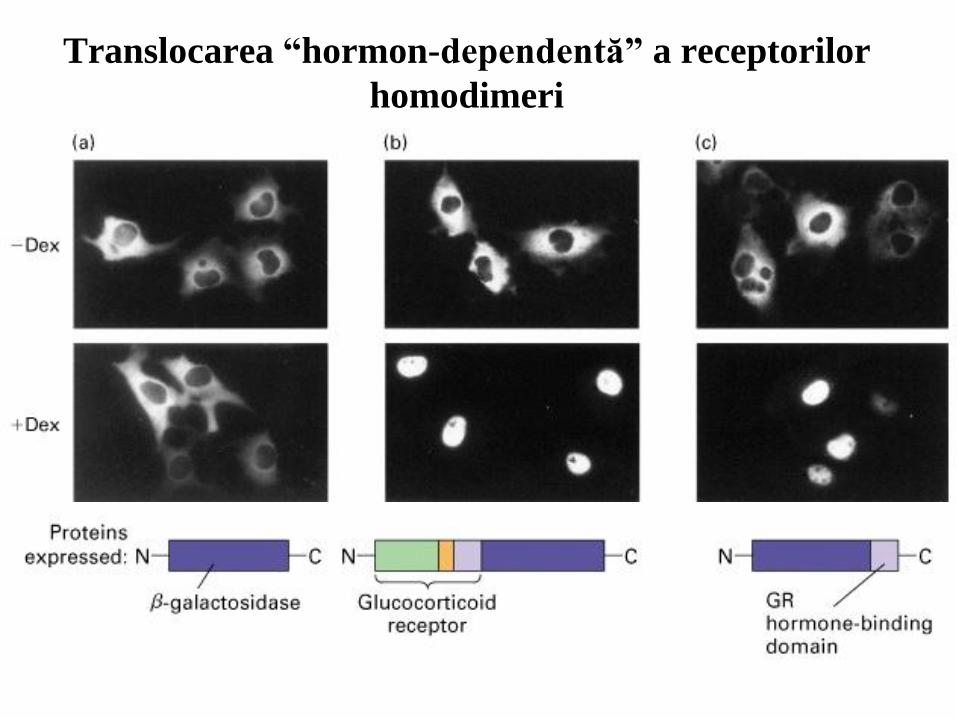
Translocarea “hormon-dependentă” a receptorilor
homodimeri
88
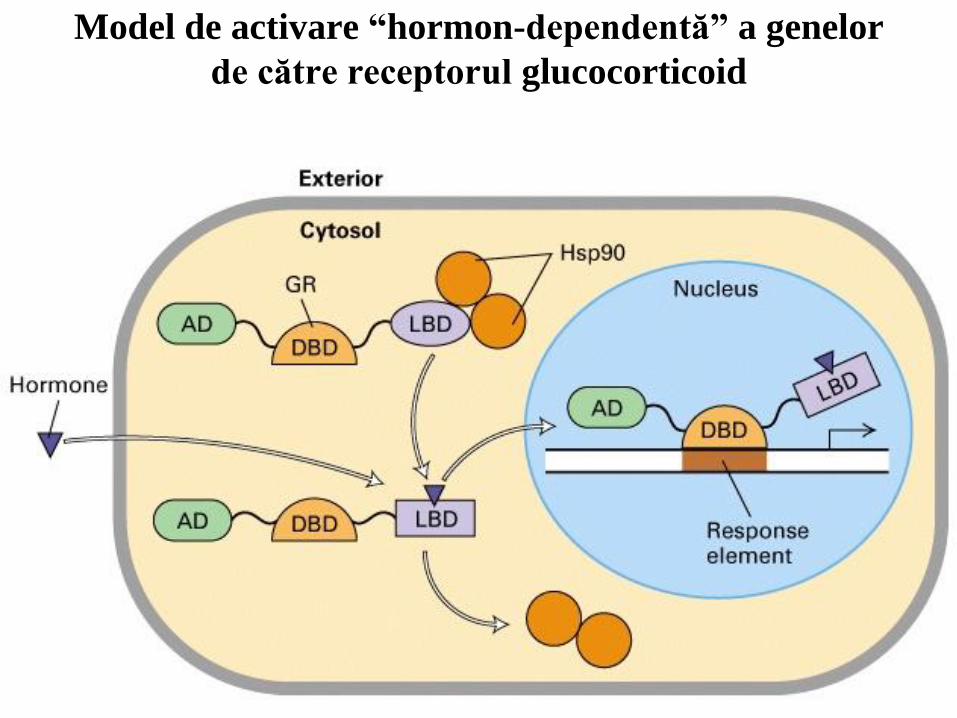
Model de activare “hormon-dependentă” a genelor
de către receptorul glucocorticoid
89
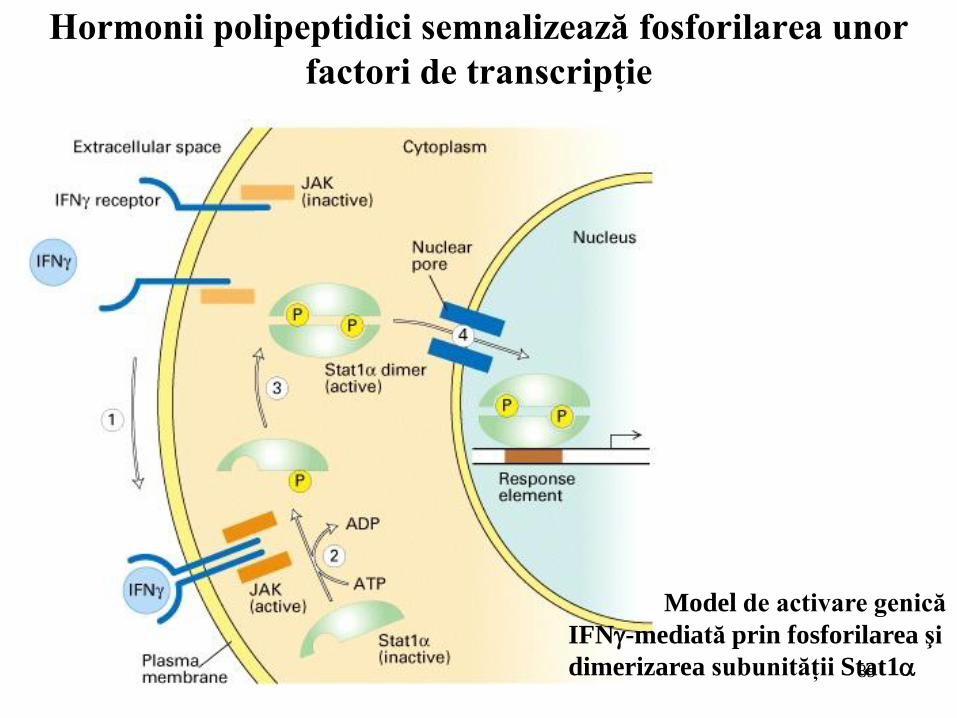
Hormonii polipeptidici semnalizează fosforilarea unor
factori de transcripţie
Model de activare genică
IFN-mediată prin fosforilarea şi
dimerizarea subunităţii Stat1

90
Concluzii
Controlul transcripţional la eucariote operează pe trei nivele: i) modularea
nivelelor şi/sau activităţilor activatorilor şi represorilor; ii) modificări în structura
cromatinei determinate de activatori şi represori; iii) influenţa directă a activatorilor şi
represorilor în asamblarea complexelor de iniţiere;
Heterocromatina se referă la regiunile de cromatină condensată în care ADN este
relativ inaccesibil factorilor de transcripţie şi altor proteine, astfel încât expresia genei este
represată;
Represarea mediată a heterocromatinei apare la nivelul telomerilor şi a locilor
implicaţi în conjugare la S. Cerevisiae. Interacţiile diferitelor proteine şi hipoacetilarea
regiunilor N-terminale ale histonelor H3 şi H4 sunt responsabile pentru represarea
structurii cromatinei în aceste regiuni;
Unii represori funcţionează parţial prin interacţia cu complexele de deacetilare a
histonelor, având drept rezultat deacetilarea histonelor din structura nucleosomilor situaţi
în vecinătate. Aceasta inhibă interacţia dintre promotorul ADN şi factorii generali de
transcripţie, represând astfel iniţierea transcripţiei;

91
Concluzii
Factorii care remodelează cromatina determină disocierea tranzitorie a ADN de
“miezul” histonic printr-o reacţie ATP-dependentă şi promovează astfel legarea altor
proteine necesare procesului de iniţiere care se desfăşoară la nivelul anumitor promotori
ADN;
In vitro, combinarea activatorilor poate stimula asamblarea complexelor de
iniţiere din vecinatatea promotorului. Se crede că acest efect direct al activatorilor apare
in vivo în urma acetilării histonelor;
In vivo, asamblarea “înalt” cooperativă a complexului de iniţiere necesită mai
mulţi activatori. O celulă trebuie să producă un set specific de activatori necesari pentru
transcripţia unei anumite gene în scopul reglării expresiei acesteia;
Unii represori inhibă competitiv legarea activatorilor sau factorilor generali de
transcripţie. Alţii interacţionează direct cu factorii generali sau cu activatorii;
Activităţile super-familiei recptorilor nucleari sunt reglate de hormonii lipo-
solubili. Legarea hormonilor la aceşti factori de transcripţie induce modificări
conformaţionale care modifică interacţiile lor cu alte proteine;
Activităţile unor factori de transcripţie sunt reglate prin fosforilare indusă de
legarea hormonilor polipeptidici la receptorii lor situaţi la suprafaţa celulelor.
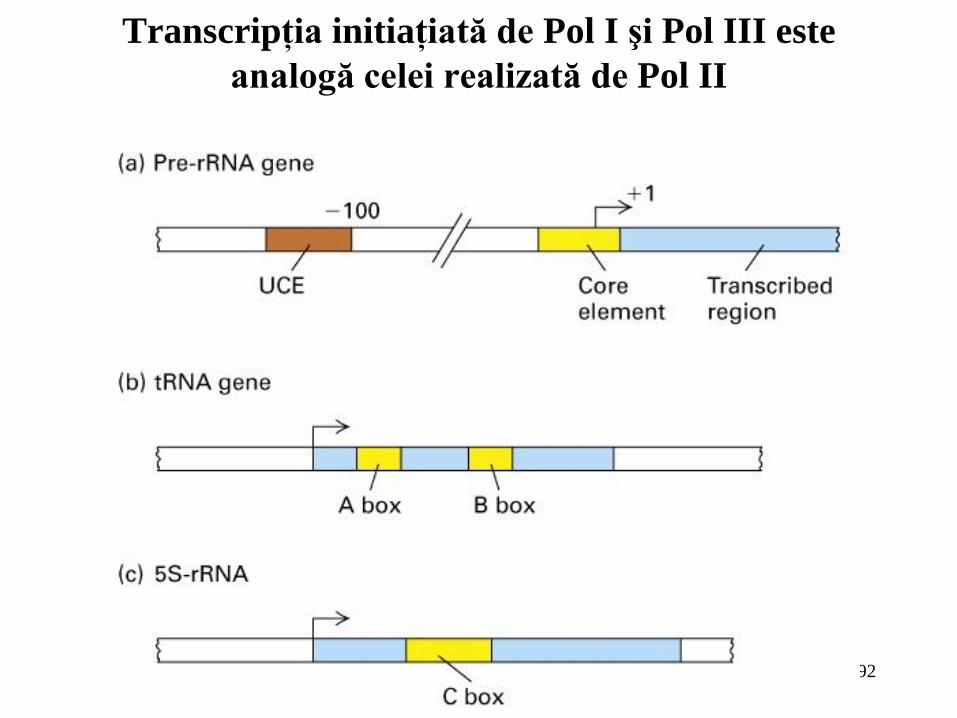
92
Transcripţia initiaţiată de Pol I şi Pol III este
analogă celei realizată de Pol II

93
Alte sisteme de transcripţie
T7 şi alţi bacteriofagi înrudiţi exprimă ARN
polimeraze monomere
ADN mitocondrial este transcris de către ARN
polimeraze care prezintă similarităţi cu enzimele bacteriofagilor
sau bacteriene
Transcripţia ADN din cloroplaste se aseamănă cu
transcripţia bacteriană
Transcripţia de la archaea este mai apropiată de
transcripţia de la eucariote decât de cea de la bacterii
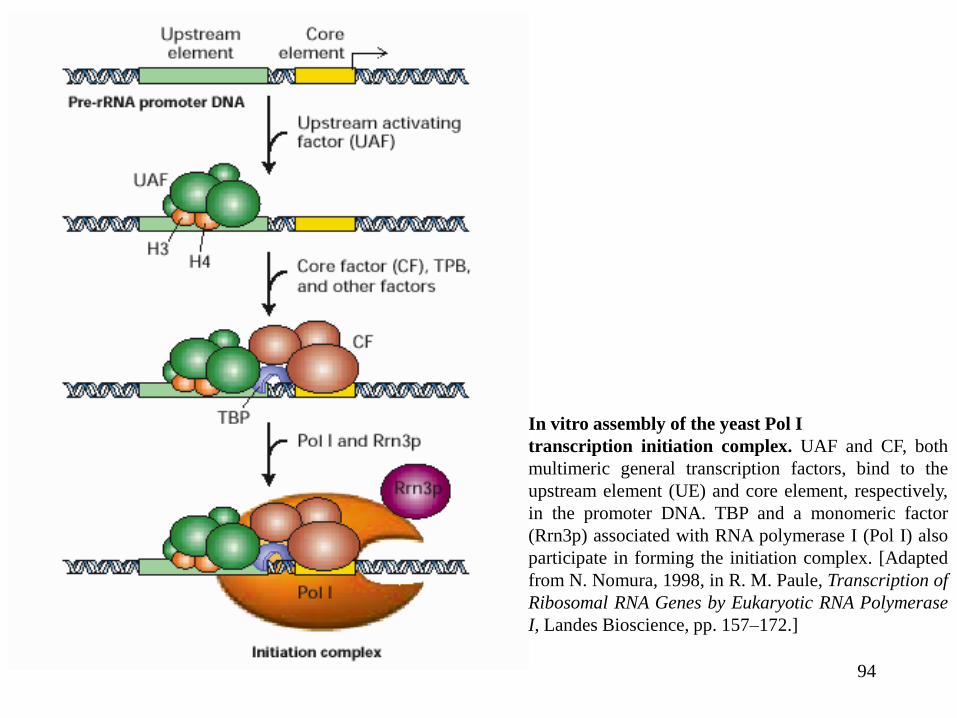
94
In vitro assembly of the yeast Pol I
transcription initiation complex. UAF and CF, both
multimeric general transcription factors, bind to the
upstream element (UE) and core element, respectively,
in the promoter DNA. TBP and a monomeric factor
(Rrn3p) associated with RNA polymerase I (Pol I) also
participate in forming the initiation complex. [Adapted
from N. Nomura, 1998, in R. M. Paule, Transcription of
Ribosomal RNA Genes by Eukaryotic RNA Polymerase
I, Landes Bioscience, pp. 157–172.]
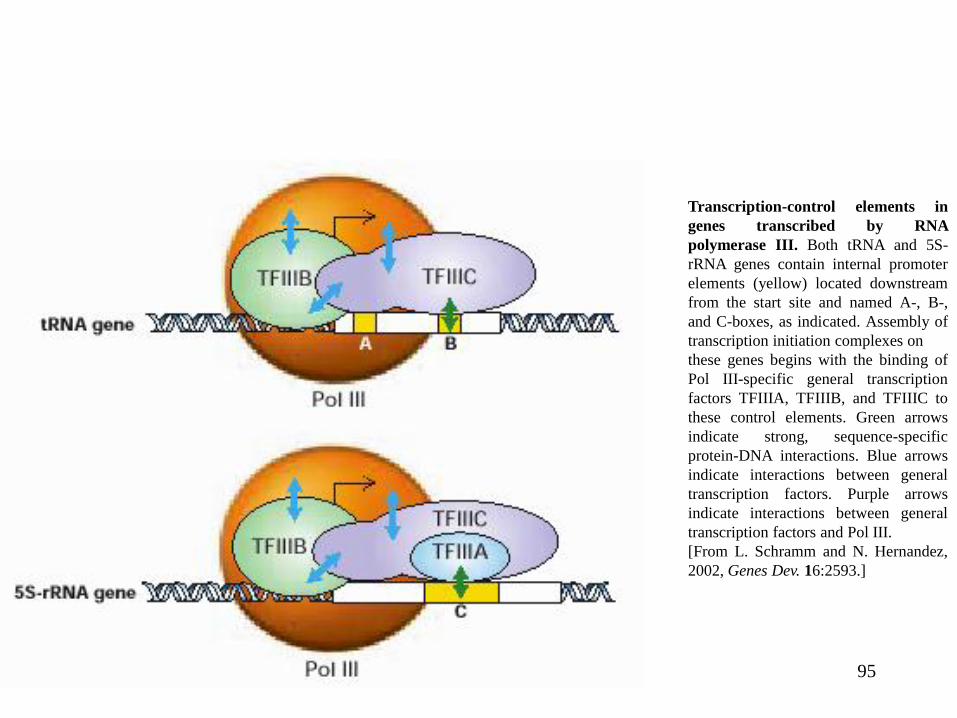
95
Transcription-control elements in
genes transcribed by RNA
polymerase III. Both tRNA and 5S-
rRNA genes contain internal promoter
elements (yellow) located downstream
from the start site and named A-, B-,
and C-boxes, as indicated. Assembly of
transcription initiation complexes on
these genes begins with the binding of
Pol III-specific general transcription
factors TFIIIA, TFIIIB, and TFIIIC to
these control elements. Green arrows
indicate strong, sequence-specific
protein-DNA interactions. Blue arrows
indicate interactions between general
transcription factors. Purple arrows
indicate interactions between general
transcription factors and Pol III.
[From L. Schramm and N. Hernandez,
2002, Genes Dev. 16:2593.]

96
Concluzii
Iniţierea transcripţiei de către Pol I este dirijată de către un element promotor “core”, care
se suprapune cu situsul de start, şi o regiune de control situată în amonte (UCE). Aceata necesită doi
factori de transcripţie generali, SL1 şi UBF;
Iniţierea transcripţiei de către ARN polimeraza III este cel mai adesea dirijată de către
elemente promotoare interne. Doi factori generali de transcripţie sunt necesari pentru iniţierea
transcripţiei ARNt (TFIIIC şi TFIIIB); un factor suplimentar (TFIIIA) este necesar pentru iniţierea
transcripţiei genelor ARNr 5S;
Iniţierea transcripţiei de către toate cele trei ARN polimeraze nucleare eucariote necesită un
factor de transcripţie specific general care conţine TBP ca subunitate;
Iniţierea de către Pol I şi Pol III nu necesită ATP spre deosebire de iniţierea realizată de Pol
II care depinde de hidroliza ATP;
Bacteriofagul T7 şi bacteriofagii înrudiţi exprimă o ARN polimerază monomeră simplă.
Aceste polimeraze recunosc o regiune promotoare de 23 pb care include situsul de start;
Mitocondria conţine molecule circulare de ADN transcrise de către o ARN polimerază
codificată nuclear compusă din două subunităţi. O subunitate este omologă cu ARN polimeraza simplă
din bacteriofagul T7; cealaltă se aseamănă cu factorii bacterieni sigma;
Cloroplastele conţin ADN care este transcris de către o ARN polimerază codificată de
cloroplast omologă cu ARN polimerazele bacteriane, cu excepţia că aceasta este lipsită de factorul
sigma;
Archeobacteriile utilizează o ARN polimerază formată din mai multe subunităţi care se
aseamănă cu ARN polimerazele eucariote nucleare. Iniţierea transcripţiei în celulele de archeobacterii
necesită o proteină, omologă cu factorul eucariotic TBP, care se leagă la un element promotor bogat în
A/T situat imediat în amonte faţă de situsul de start şi o proteină omologă cu TFIIB. Aceste rezultate
susţin ipoteza că eucariotele şi acheobacteriile actuale au evoluat dintr-un strămoş comun.