UNIVERSIDADE ESTADUAL DE CAMPINAS Instituto de Biologia
87
UNIVERSIDADE ESTADUAL DE CAMPINAS Instituto de Biologia FERNANDA GARCIA FÓSSA INTER-RELAÇÃO ENTRE NANOPARTÍCULAS LIPÍDICAS SÓLIDAS, A VIA DO TGF-β E A TRANSIÇÃO EPITÉLIO-MESENQUIMAL EM CÉLULAS DE PRÓSTATA TUMORAIS E NÃO TUMORAIS CAMPINAS 2020
Transcript of UNIVERSIDADE ESTADUAL DE CAMPINAS Instituto de Biologia

UNIVERSIDADE ESTADUAL DE CAMPINAS
Instituto de Biologia
FERNANDA GARCIA FÓSSA
INTER-RELAÇÃO ENTRE NANOPARTÍCULAS LIPÍDICAS SÓLIDAS, A VIA DO
TGF-β E A TRANSIÇÃO EPITÉLIO-MESENQUIMAL EM CÉLULAS DE PRÓSTATA
TUMORAIS E NÃO TUMORAIS
CAMPINAS
2020

FERNANDA GARCIA FÓSSA
INTER-RELAÇÃO ENTRE NANOPARTÍCULAS LIPÍDICAS SÓLIDAS, A VIA DO
TGF-β E A TRANSIÇÃO EPITÉLIO-MESENQUIMAL EM CÉLULAS DE PRÓSTATA
TUMORAIS E NÃO TUMORAIS
Dissertação apresentada ao Instituto de
Biologia da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do Título de Mestra em Biologia Funcional e Molecular, na área de Bioquímica.
Orientador: PROF. DR. MARCELO BISPO DE JESUS
CAMPINAS
2020
ESTE ARQUIVO DIGITAL CORRESPONDE À
VERSÃO FINAL DA DISSERTAÇÃO
DEFENDIDA PELA ALUNA FERNANDA GARCIA
FÓSSA E ORIENTADA PELO PROF. DR.
MARCELO BISPO DE JESUS.


Campinas, 19/02/2020.
COMISSÃO EXAMINADORA
Prof. Dr. Marcelo Bispo de Jesus
Prof.(a). Dr.(a) Carmen Veríssima Ferreira Halder
Prof.(a) Dr(a). Ana Carolina Santos de Souza Galvão Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no processo de vida acadêmica do aluno.
A Ata da defesa com as respectivas assinaturas dos membros encontra-se no
SIGA/Sistema de Fluxo de Dissertação/Tese e na Secretaria do Programa (Biologia
Funcional e Molecular) da Unidade (Instituto de Biologia).

AGRADECIMENTOS
Aos meus pais, Beatriz e Nelson, que me deram toda a estrutura e apoio
emocional para chegar até aqui; obrigada por tudo. Aos meus irmãos, Renata, Lígia,
Flávia e Júnior, por serem bons exemplos de humanos bons, e nos quais eu sempre
me baseio para ser alguém melhor. Em especial à Lígia, minha irmã-mãe que apoia
e torce por mim como ninguém. Vocês são a minha maior torcida. Às minhas avós
Albertina (in memoriam) e Othilia, exemplos de mulheres fortes e guerreiras.
Ao Vlad, meu companheiro nessa jornada do mestrado e da vida, obrigada
pelos conselhos, pelos insights, por me incentivar a crescer, pelo carinho e paciência
que faz meus dias mais felizes. Você me mostra que o caminho pode ser leve,
mesmo quando árduo.
Ao meu orientador Marcelo Bispo, que desde os primórdios da minha
iniciação científica, me mostrou o que é ser uma cientista, me incentivou a pensar
por mim mesma e acreditar; não tenha dúvidas de que me espelho em você.
Obrigada pelos conhecimentos e por me guiar nessa escuridão (que agora vejo com
mais clareza) da vida acadêmica.
Aos queridos professores da minha banca de qualificação, Profa. Dra.
Carmen Veríssima Ferreira, Prof. Dr. Murilo Vieira Geraldo e Prof. Dr. Hernandes
Faustino de Carvalho, sou muito grata pelos conselhos sobre a aula e meu projeto.
Ao Prof. Bonafé pelas conversas esclarecedoras e histórias sempre boas; à
Profa. Eneida por me ensinar um pouquinho perto da sua vasta experiência como
professora, e em quem eu me espelho para quando for ensinar algum dia.
Ao laboratório NanoCell das antigas: Allan, Luiz, Guilherme, Ju Mattoso, em
especial a Carol Cassago que me guiou como ninguém, obrigada por todos os
ensinamentos e conversas, vocês fazem parte da minha formação. À Bruna, pois
compartilhamos muitos dos medos do início, além da grande amiga que se tornou.
Ao NanoCell não tão distante do presente: Monique, por seus cafés
acompanhados de muitos conselhos e suas ideias ótimas, obrigada; Aline, por sua
ajuda, conversas e cafés, obrigada; à Tuanny, agradeço as conversas, conselhos
sobre a vida, experimentos e desabafos, e por sempre estar presente nessa jornada.
Tamo junto!
À Samara, que além de me suportar desde o início me mostrando que a vida
é mais simples do que parece, me confortou quando eu precisei, me socorreu em

todas as crises, e não saiu do meu lado. Sá, obrigada por toda sua luz e sua
amizade.
Aos que encontrei durante o mestrado e que são especiais: à Maria
Fernanda, sempre presente, obrigada pela força e amizade; agradeço à Jéssica, por
me ensinar a fazer scratch, e nossa colaboração que rendeu um trabalho bonito e
uma amizade mais bonita ainda; Thays, que conheci há pouco, mas que trouxe uma
luz gigante para minha vida, obrigada.
Aos amigos de hoje e sempre, Letícia, Fer, Catu, Carol, Bru, por estarem
comigo e saber que mesmo longe, podemos contar. À Pierina, a melhor amiga dos
melhores projetos, e que tenho a sorte de encontrá-la no corredor logo à frente –
obrigado por tudo, quero ser você quando crescer.
Aos que sem eles nosso trabalho não seria possível: Ju Mattoso, Érika, Elzira,
Kiko por todo o suporte, e Vânia, sempre com um bom dia caloroso logo cedo. À
Unicamp, que tem sido minha morada por seis anos de muito aprendizado.
Agradeço à Fundação de Amparo à Pesquisa do Estado de São Paulo
(FAPESP) pelo equipamento multiusuário Leitor Multidetecção Híbrido Cytation 5
(BioTek Instruments, Inc., Winooski, VT, USA), FAPESP #15/06134.
O presente trabalho foi realizado com apoio da Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Código de
Financiamento 001.

“Don't let anyone rob you of your imagination, your creativity, or your curiosity. It's
your place in the world; it's your life. Go on and do all you can with it, and make it the
life you want to live.” – Mae Jemison

RESUMO
O estudo da nanotecnologia, e mais especificamente de nanopartículas têm
crescido, principalmente de suas propriedades e efeitos colaterais. Dentre as
nanopartículas, se destacam as nanopartículas lipídicas sólidas (SLN), produzidas
com componentes biocompatíveis e biodegradáveis. Muito se estuda sobre os
efeitos benéficos de nanopartículas na entrega específica de fármacos e genes às
células, porém pouco se sabe sobre os efeitos em vias de sinalização intracelular.
Nesse trabalho estudamos os efeitos de SLN catiônica em células não tumorais e
tumorais do epitélio de próstata humano, relacionadas à via do TGF-β e transição
epitélio-mesenquimal (EMT). A via do TGF-β está relacionada com o aumento da
invasão e migração celular no câncer, estimulando a metástase através da ativação
da EMT. Nossos resultados mostram que as SLN aumentam a migração de células
de câncer de próstata PC-3, efeito não observado em células não tumorais de
próstata PNT1A – um resultado inesperado e inédito. Observamos que o aumento
da migração depende da nanopartícula e não dos seus componentes, e é um efeito
concentração-dependente. Ainda, proteínas relacionadas à EMT, como Zeb1,
mostraram-se aumentadas após tratamento com a SLN, demonstrando a
importância dessa via no efeito biológico. Além disso, o papel biológico, i.e., a
transfecção, mediado por SLN é significativamente reduzido quando inibimos os
receptores do TGF-β ou a Smad3. Concluímos que a SLN é capaz de interferir na
motilidade de células PC-3 através de sua ativação da via do TGF-β e a EMT. Tal
efeito, ao extrapolar para a clínica, poderia agravar o quadro de um paciente caso
esses resultados reproduzam em ensaios in vivo. A compreensão dos mecanismos
moleculares disparado por nanomateriais, bem como suas consequências para o
funcionamento celular, é importante para o desenvolvimento de novos nanomateriais
e para delimitar seu uso seguro e eficaz.

ABSTRACT
The study of nanotechnology, and more specifically of nanoparticles, has grown,
mainly due to its properties and side effects. Among the nanoparticles, we highlight
the Solid Lipid Nanoparticles (SLN), produced with biocompatible and biodegradable
components. Much has been researched about the beneficial effects of nanoparticles
on the specific delivery of drugs and genes to cells, but little is known about the
effects on intracellular signaling pathways. In this work we studied the effects of
cationic SLN in non-tumor and tumor cells of the human prostate epithelium, related
to the TGF-β pathway and epithelial-mesenchymal transition (EMT). The TGF-β
pathway is related to increased cell invasion and migration in cancer, stimulating
metastasis through activation of EMT. Our results show that SLNs increase the
migration of PC-3 prostate cancer cells, an effect not seen in non-tumor prostate
cells PNT1A - an unexpected and unprecedented result. We observed that the
increase in migration depends on the nanoparticle and not on its components, and is
a concentration-dependent effect. Also, EMT-related proteins, such as Zeb1, were
shown to be increased after treatment with SLN, demonstrating the importance of
this pathway in the biological effect. In addition, the biological effect, i.e., transfection,
mediated by SLN is significantly reduced when we inhibit TGF-β receptors or Smad3.
We conclude that SLN is capable of interfering in the motility of PC-3 cells through its
activation of the TGF-β pathway and EMT. Such an effect, when extrapolating to the
clinic, could worsen a patient's condition if these results reproduce in in vivo trials.
Understanding the molecular mechanisms triggered by nanomaterials, as well as
their consequences for cellular functioning, is important for the development of new
nanomaterials and to delimit their safe and effective use.

LISTA DE ILUSTRAÇÕES
Figura 1 - Principais vias de endocitose em células. ........................................ 19
Figura 2 - Modelo simplificado da via de sinalização de TGF-β.. ..................... 24
Figura 3 – Estrutura química de inibidores dos receptores TβRI e TβRII (LY2109761,
LY364947 e SB431542) e Smad (SIS3 HCl) da via do TGF-β. ................. 27
Figura 4 - Transfecção mediada por SLN em células de próstata. ................... 45
Figura 5 - Velocidade de migração de células de próstata após exposição à SLN
................................................................................................................... 48
Figura 6 – Velocidade de migração de células de próstata na presença de SLN e de
cada um de seus componentes. ................................................................ 50
Figura 7 - SLN induz a migração de células PC-3 de forma dose-dependente.52
Figura 8 – Proliferação de células PNT1A após tratamento com SLN. ............ 54
Figura 9 – Proliferação de células PC-3 após tratamento com SLN.. .............. 56
Figura 10 - Determinação da concentração ótima dos inibidores SIS3 e LY2109761
em células de próstata PNT1A e PC-3.. .................................................... 58
Figura 11 – Transfecção por SLN:GFP em células PNT1A e PC-3 em presença dos
inibidores LY2109761 e SIS3.. .................................................................. 60
Figura 12 – Internalização de SLN-Rodamina em células PNT1A e PC-3 após
tratamento com inibidores LY2109761 e SIS3. .......................................... 61
Figura 13 – Translocação nuclear de Smad2 em células PNT1A em presença ou
ausência de inibidores e do TGF-β.. .......................................................... 63
Figura 14 - Translocação nuclear de Smad2 em células PC-3 após tratamento com
inibidores. .................................................................................................. 65
Figura 15 – Translocação de Smad2 em células PNT1A.. ............................... 67
Figura 16 – Translocação de Smad2 em células PC-3.. .................................. 69
Figura 17 - Proteínas Zeb1 e Snail em células PNT1A após 24h de tratamento.71
Figura 18 – Proteínas Zeb1 e Snail em células PC-3 após 24h de tratamento. 73
Figura 19 – Vimentina em células do fronte migratório após 12h.. ................... 75

LISTA DE TABELAS
Tabela 1 – Composição das formulações produzidas. ..................................... 33
Tabela 2 - Anticorpos primários e suas respectivas diluições. ......................... 40
Tabela 3 – Caracterização das nanopartículas.. .............................................. 43

LISTA DE ABREVIATURAS E SIGLAS
ANOVA Analysis Of Variance (Análise de Variância)
BMP Bone Morphogenetic Protein (Proteína Morfogenética Óssea)
CBS Cascading Bistable Switches
cPML cytoplasmatic ProMyelocytic Leukaemia (Leucemia
Promielocítica citoplasmática)
DAPI 4,6–diamidino-2-fenilindol
DLS Dynamic Light Scattering (dispersão de luz dinâmica)
DMSO Dimethyl Sulfoxide (Dimetilsulfóxido)
DOTAP 1,2 dioleoil propano 3-trimetilamônio
EdU 5-ethinil-2‟deoxiuridina
EEA1 Early Endossome Antigen 1 (Antígeno do endossomo precoce 1)
EMT Epithelial-Mesenchymal Transition (Transição Epitélio-
Mesenquimal)
ERK Extracellular-Signal-Regulated Kinase (quinase regulada por sinal
extracelular)
EthD-1 homodímero de etídio
GDF Growth Differentiation Factor (Fator de diferenciação do
crescimento)
hPLCS High-performance liquid chromatography (Cromatografia Líquida
De Alta Eficiência)
IL-12 Interleucina-12
MH1/2 MAD homology 1/2
p38 Mitogen-activated protein kinase 38
PBS Phosphate-buffered saline (solução salina tamponada de fosfato)
PCTA Prostate Carcinoma Tumor Antigen-1 (sinônimo para Galectina-8)
PDI Índice de polidispersão
PFA Paraformaldehyde (Paraformaldeído)
PLGA-
PEG
Poli-ácidos Lático e Glicólico- Poli-Etileno Glicol
RPMI
1640
Roswell Park Memorial Institute 1640
SARA Smad Anchor for Receptor Activation

SFB Soro Fetal Bovino
SIS3 Specific Inhibitor of Smad3 (Inibidor específico de Smad3)
SLN Solid Lipid Nanoparticles (Nanopartículas lipídicas sólidas)
Smad Small mothers against decapentaplegic
Snail Zinc finger protein SNAI1
TCS Ternary Chimera Switch
TGF-β Transforming Growth Factor (Fator de transformação do
crescimento)
TGIF Transforming Growth-Interacting Factor
TβRI/II/III Transforming Beta Receptor type I/II/III (Receptor do TGF-β do
tipo I/II/III)
Zeb1 Zinc finger E-box-binding homeobox 1

SUMÁRIO
1. Introdução ........................................................................................................... 16
a. Nanomedicina e o câncer de próstata ............................................................. 16
b. Nanopartículas lipídicas sólidas (SLN) ............................................................ 17
c. Endocitose ....................................................................................................... 17
d. Internalização e tráfego de nanopartículas e sinalização celular .................... 20
e. A via do TGF-β ................................................................................................ 21
f. Inibidores da via do TGF-β .............................................................................. 25
g. Efeitos biológicos da via do TGF-β: Migração e EMT...................................... 27
h. Metástase ........................................................................................................ 29
i. PyScratch ........................................................................................................ 30
j. Resultados preliminares .................................................................................. 30
2. Objetivo ............................................................................................................... 32
3. Materiais e métodos............................................................................................ 32
a. Cultura de células ........................................................................................ 32
b. Produção e caracterização de nanopartículas lipídicas sólidas (SLN) ......... 33
c. Transfecção por SLN ................................................................................... 33
d. Ensaio de migração celular .......................................................................... 34
e. Proliferação celular....................................................................................... 36
f. Avaliação da citotoxicidade dos inibidores ................................................... 37
g. Determinação da transfecção na presença dos inibidores ........................... 38
h. Determinação da Internalização dos complexos em presença de inibidores
38
i. Avaliação de proteínas da via do TGF-β e EMT .......................................... 39
j. Imunofluorescência e anticorpos .................................................................. 40
k. Análise de imagens ...................................................................................... 41
l. Análise estatística dos resultados ................................................................ 42
4. Resultados e discussão ...................................................................................... 42

a. Caracterização das nanopartículas lipídicas sólidas (SLN) ...................... 42
b. Eficiência de transfecção .......................................................................... 43
c. Migração na presença da SLN .................................................................. 45
d. SLN e proliferação celular ......................................................................... 52
e. SLN e a via do TGF-β ............................................................................... 57
f. SLN e a transição epitélio-mesenquimal (EMT) ........................................ 70
5. Conclusão ........................................................................................................... 76
6. Referências bibliográficas ................................................................................... 77
7. Anexos ................................................................................................................ 86

16
1. Introdução
a. Nanomedicina e o câncer de próstata
É previsto que em 2030 a população de homens acima de 65 anos
chegue a 72 milhões e dessa forma, a taxa de câncer de próstata aumente
proporcionalmente (Cao e Kyprianou 2015). Dos casos de câncer em homens, o
câncer de próstata correspondeu a 1 em cada 5 novos casos em 2018 (Siegel,
Miller, e Jemal 2019). Desde 1979, o PSA (do inglês prostate-specific antigen) é
utilizado como marcador para o adenocarcinoma de próstata e representa uma
triagem inicial e caso haja suspeita de câncer, o próximo passo é a realização da
biópsia transretal guiada por ultrassom (Lomas e Ahmed 2020). O tratamento para o
câncer de próstata pode ser realizado através de cirurgia laparóspica ou robótica, e
também através da radioterapia associada à terapia de supressão androgênica
(Sathianathen et al. 2018). A castração cirúrgica está sendo substituída por métodos
farmacológicos, principalmente de fármacos anti-receptor de andrógeno, prevenindo
que os receptores estimulem o crescimento do câncer (Sathianathen et al. 2018).
Apesar do grande avanço, existe a necessidade de se desenvolver novas terapias
que tragam menores efeitos adversos, sendo mais eficientes e mais seguras para o
tratamento do câncer de próstata.
O câncer de próstata pode ter várias causas, dentre elas vias de sinalização
desreguladas. Uma das vias desreguladas é a do TGF-β, que em células epiteliais
não tumorais de próstata inibe a proliferação e induz a apoptose através da cascata
de sinalização de Smad (Cao e Kyprianou 2015). Em estágios avançados da
doença, o TGF-β promove a migração e invasão tumoral através de efeitos no
citoesqueleto de actina, aumentando a mobilidade de células de câncer de próstata
(Dicken, Hensley, e Kyprianou 2019). Por isso, a via do TGF-β no câncer de próstata
se tornou alvo de medicamentos, como por exemplo, o inibidor dos receptores do
TGF-β, LY2157299, que já está em fase clínica de testes (Cao e Kyprianou 2015).
O uso da nanotecnologia através de nanopartículas se tornou uma opção
para o tratamento do câncer de próstata, um exemplo é o uso de nanopartículas de
polietilenoglicol (PLGA-PEG) conjugadas a anti-CD24 para entrega de fármacos
especificamente no tumor (Bharali et al. 2017; Sanna e Sechi 2012). Conforme
cresce a aplicação de nanopartículas para o tratamento de câncer de próstata, é

17
importante entender quais os mecanismos utilizados pelas nanopartículas ao
interagirem com essas células. Pergunta-se: há algum tipo de interação entre as
nanopartículas e receptores de membrana celular? Seria uma nanopartícula é capaz
de disparar vias de sinalização intracelulares e qual o efeito causado por tal disparo?
b. Nanopartículas lipídicas sólidas (SLN)
Nanopartículas lipídicas sólidas (SLN, do inglês Solid Lipid Nanoparticles) têm
sido utilizadas para entrega de fármacos e material genético em células de câncer
de próstata (Akanda et al. 2015; Badawi et al. 2018; de Jesus et al. 2010), pois
possuem características desejadas, como por exemplo, o tamanho em escala
nanométrica, grande área de superfície e potencial para aumentar a performance de
outros compostos, como fármacos (Bhatt et al. 2018).
Usualmente, as SLN são produzidas por homogeneização em alta pressão
(HPH) e sonicação (Nasirizadeh e Malaekeh-Nikouei 2020). Nosso grupo de
pesquisa desenvolveu uma técnica de extrusão de microemulsão laboratorial, que
garante a estabilidade das SLN, sendo que sua produção é manual, de baixo custo e
eficiente. Com isso, estabelecemos um sólido conhecimento para desenvolvimento,
caracterização e aplicação de formulações de SLN para o carreamento de genes e
em muitos deles utilizamos células de câncer de próstata (de Jesus et al. 2010; de
Jesus e Zuhorn 2015; Radaic e Jesus 2018; Radaic, de Paula, e de Jesus 2015).
Tais nanopartículas são formadas por lipídios sólidos a temperatura ambiente,
surfactantes que previnem agregação, e agentes hidrossolúveis e lipossolúveis
podem ser incorporados a ela. Devido a sua composição, e por apresentar baixa
toxicidade, as SLN possuem excelente biocompatibilidade e são consideradas
seguras para o uso (Bhatt et al. 2018; Scheideler, Vidakovic, e Prassl 2020).
Entretanto, nosso grupo mostrou que as SLN induzem inflamação de baixa
intensidade em ratos, sendo capazes de ultrapassar a barreira hematoencefálica
sem comprometer sua integridade (Mendonça et al. 2019). Dessa forma, existem
lacunas no entendimento da interação SLN e sistemas biológicos, já que sua
composição segura não garante a falta de efeitos colaterais.
c. Endocitose
O transporte de moléculas da membrana plasmática em direção ao
citoplasma é chamado de endocitose, processo em que pequenas vesículas

18
transportam nutrientes, receptores de membrana e outras moléculas para o interior
da célula (Melkikh e Sutormina 2020). Com o auxílio de proteínas específicas,
vesículas são formadas a partir da invaginação da membrana plasmática; tais
vesículas receberão a carga a ser endocitada (Kaksonen e Roux 2018).
A endocitose é dividida em fagocitose e pinocitose: a fagocitose é realizada
por células do sistema imunológico, enquanto que a pinocitose existe virtualmente
em todas as células. A pinocitose é dividida em endocitose mediada por clatrina
(CME, do inglês clathrin-mediated endocytosis), endocitose mediada por caveolina,
macropinocitose e endocitose independentes de clatrina-caveolina (Figura 1). De
modo geral, a endocitose inicia-se através da invaginação da membrana contendo a
carga a ser internalizada, formando uma vesícula que irá se desligar da membrana e
serão entregues para o endossomo precoce. O endossomo precoce contendo a
carga passa por processos de fusão e fissão, diminuição do pH e acontece a fusão
com endossomos tardios, até chegar ao lisossomo, onde o material será degradado
(Melkikh e Sutormina 2020; Sahay, Alakhova, e Kabanov 2010).
Vários receptores importantes, como o receptor de transferrina, dependem da
CME para serem internalizados pela célula. A clatrina, com auxílio de proteínas
adaptadoras, polimeriza e forma um revestimento em volta da vesícula que está em
formação na membrana plasmática (Hinze e Boucrot 2018). Já a endocitose
mediada por caveolina é feita através de cavéolas, que são vesículas contendo
domínios ricos em colesterol e caracterizados pela presença das proteínas caveolina
e cavina, que também é importante para a endocitose de receptores, como por
exemplo o GLUT4 (Moore et al. 2018). Ressaltamos a importância de considerar o
tipo celular no que concerne a endocitose, pois o tipo de endocitose vai depender da
célula, da carga endocitada e do contexto em que a célula está inserida.

19
Figura 1 - Principais vias de endocitose em células (retirado de Ferreira et al. 2014).
Além do papel de nutrição da célula, a endocitose pode ser definida como um
sistema intimamente integrado às funções e sinalização celular, que constitui a
maior infraestrutura de comunicação da célula - integra a informação, transporte e
regulação das respostas celulares (Barbieri, Di Fiore, e Sigismund 2016). A
endocitose de receptores de superfície define o destino dos receptores, seja
marcando-os para degradação, diminuindo o sinal, ou reciclando-os de volta à
membrana plasmática e mantendo a sinalização ativa (Schmid 2017). Estímulos
contínuos aos receptores podem diminuir sua concentração, evitando um efeito
exacerbado daquele sinal, o que torna necessário um maior número de ligantes para
acionar a resposta celular (Moore et al. 2018). Já para outras vias, os receptores

20
ficam concentrados em endossomos, porque alguns deles necessitam desse
mecanismo para entrarem em contato com proteínas de uma cascata de sinalização
(Moore et al. 2018).
A cascata de sinalização pode ser determinada segundo a rota endocítica de
um receptor. Por exemplo, os receptores do TGF-β podem ser internalizados através
de duas vias: endocitose mediada por clatrina ou caveolina. A entrada via clatrina
resulta na entrega dessa vesícula em endossomo precoce positivo para EEA-1
(EEA-1, do inglês Early Endosome Antigen 1), sendo que EEA-1 se associa a
Rab5GTP presente em endossomos precoces. Além do EEA-1, a proteína SARA (do
inglês Smad Anchor for Receptor Activation) necessita estar presente nos
endossomos para que a sinalização aconteça. Assim, a presença dos receptores na
CME promove a via canônica do TGF-β, aumentando a translocação de Smad2 e
ativando a sinalização intracelular. Após a ativação, os receptores podem ser
degradados ou reciclados de volta à membrana celular. Caso haja uma alta
concentração de receptores associados ao ligante TGF-β (concentrações >10
ng/mL), a internalização ocorre via caveolina. Nessa situação, o receptor ativado
interage com proteína adaptadora Smad7 e SMURF7, induzindo a ubiquitinação e
subsequente degradação do receptor, que controla a resposta da célula à grande
concentração de ligante (Chen 2009; He et al. 2015; Vander Ark, Cao, e Li 2018).
Dessa forma, a endocitose oferece ferramentas para que a célula regule a
propagação, duração e atenuação do sinal (He et al. 2015).
Assim como a endocitose de receptores por vias específicas determina o
efeito causado na célula, a endocitose de nanopartículas por diferentes vias também
determina qual será o efeito na célula, pois tem conexão com a liberação do material
genético/fármaco carreado pela nanopartícula (de Jesus e Kapila 2014). O material
da nanopartícula, tamanho e carga influenciam na via de entrada na célula: segundo
K. Chen et al. (2018), o mecanismo de entrada de nanopartículas está em fase
inicial de estudos e seu entendimento é um desafio para o desenvolvimento da
nanotecnologia.
d. Internalização e tráfego de nanopartículas e sinalização celular
Nanopartículas lipídicas complexadas ao material genético (chamadas de
lipoplexos) são internalizadas por diversas vias de endocitose, dependendo da carga
e do material carreado (Lonez et al. 2009; Rehman, Zuhorn, e Hoekstra 2013).

21
Apesar da versatilidade dos lipoplexos, a liberação do material genético nem sempre
ocorre de forma eficiente, pois depende da via de entrada na célula. Dessa forma,
estudar como a liberação ocorre e aperfeiçoá-la é essencial para o desenvolvimento
de nanopartículas (Chen et al. 2018; Voltan et al. 2017).
Durante a internalização, nanopartículas podem trafegar em direção aos
lisossomos, uma vesícula ácida que pode degradar tanto o carreador quanto o
material genético, impedindo sua efetividade (Foroozandeh e Aziz 2018; de Jesus e
Kapila 2014; Rehman et al. 2013). Uma forma de controlar a entrada das
nanopartículas é funcionalizando sua superfície com moléculas, que interagem com
receptores e aumentam a endocitose por aquela via específica (Ohya, Takahashi, e
Kuzuya 2018).
Ao induzir a interação de nanopartículas com receptores, deve-se considerar
que essa interação pode gerar respostas celulares inesperadas. Por exemplo,
lipídios catiônicos podem ativar cascatas de sinalização em células ao interagir
diretamente com componentes da membrana plasmática. Sabe-se que cargas
positivas podem contribuir para a invaginação da membrana plasmática, auxiliando
no processo de endocitose desses materiais (Lonez, Vandenbranden, e Ruysschaert
2012). Outro exemplo são os lipossomas de Lipofectamina e DOTAP que induzem a
liberação de cálcio dos estoques intracelulares (Lonez et al. 2009). DOTAP pode
ainda induzir a secreção de interleucina 12 (IL-12) em células dendríticas, além de
estimular a formação de espécies reativas de oxigênio (Lonez et al. 2012). Entender
como os lipídios são capazes de ativar essas vias é importante para prever e regular
o uso dessas nanopartículas.
e. A via do TGF-β
As células se comunicam através de ligantes como citocinas, fatores de
crescimento ou hormônios. Dentre as citocinas, a família do TGF-β (do inglês
Transforming Growth Factor-β) é uma das mais difundidas nos animais e possui
efeitos diversos, dependendo do tipo celular. A saber, essa diversidade de efeitos se
mostra na indução da diferenciação de células tronco embrionárias, inibição do
crescimento de progenitores epiteliais, apoptose em células pré-tumorais e invasão
metastática em carcinomas (David e Massagué 2018).
Cerca de 30 ligantes fazem parte da família do TGF-β. Dentre eles, existem
as subfamílias TGF-β e BMP; na subfamília do TGF-β os ligantes são conhecidos

22
como TGF-β1, TGF-β2 e TGF-β3, activinas, Nodal e GDFs (os dois últimos ativos na
embriogênese) (David e Massagué 2018). Nesse projeto, quando nos referirmos ao
TGF-β, estaremos nos restringindo ao TGF-β1, 2 e 3. Todos os ligantes da família
do TGF-β são expressos como precursores, contendo um peptídeo sinalizador na
porção N-terminal, um pró-domínio que é necessário para dobra e secreção do
ligante, seguido por um polipeptídeo maduro na porção C-terminal. O TGF-β fica em
uma forma latente que previne sua ligação aos receptores; essa forma latente é
composta pela ligação do TGF-β a dois pró-domínios, chamados de LAPs (do inglês
latency-associated polypeptides) que configuram um pequeno complexo latente
chamado de SLC (do inglês small latente complex). O SLC pode ainda se associar a
outra proteína chamada de LTBP (do inglês latente TGF-β binding protein) formando
o LLC (do inglês large latente complex), que fica depositado na matriz extracelular.
Para a ativação do TGF-β e sua liberação do LLC, proteínas como integrinas (por
exemplo, β6) e proteases (por exemplo metaloproteinases, MMP) e irá depender do
contexto fisiológico em que as células se encontram (Budi, Duan, e Derynck 2017).
As proteínas da família do TGF-β agem como dímeros e se ligam na
superfície celular aos dois receptores (proteínas quinases transmembrana
serina/treonina) do tipo I (TβRI) e do tipo II (TβRII), o último sendo restrito à ligação
ao TGF-β. Existe ainda o receptor do tipo III (TβRIII), que facilita a ligação do TGF-β
ao TβRII (Vander Ark et al. 2018). Com a ligação do TGF-β ao TβRII, ocorre a
associação deste ao TβRI, que possibilita a fosforilação do domínio glicina-serina
pertencente ao TβRI pelo TβRII. Com isso, o TβRI fica na sua forma ativa (Budi et al.
2017). É necessário ressaltar que apenas uma fração dos receptores do TGF-β
ficam disponíveis na membrana celular, e a maioria fica retida dentro das células.
Uma forma de regulação da quantidade de receptores é através da Akt: a ativação
de Akt pela insulina induz a fosforilação de Rab5, e promove a ida dos receptores do
TGF-β para a membrana celular (Budi, Muthusamy, e Derynck 2015).
Como mencionado, os receptores da via do TGF-β precisam ser endocitados
para que a via dê início. A internalização pode ocorrer por endocitose mediada por
clatrina ou caveolina. Quando endocitados via clatrina, os receptores ficam
localizados em um endossomo precoce (EEA-1) que promove a sinalização do TGF-
β (Vander Ark et al. 2018). Tais endossomos contêm altos níveis de SARA, que
facilita o recrutamento da Smad2/3 (Rozés-Salvador et al. 2018). A fosforilação de
Smad2/3 pelo TβRI é mediada por SARA, que recruta a Smad2 até os receptores de

23
TGF-β. Para que a sinalização do TGF-β seja eficiente, há a necessidade da
colocalização de SARA, Smad2/3, TβRI e TβRII para o endossomo precoce
(Chaikuad e Bullock 2016; Rozés-Salvador et al. 2018). A cPML (do inglês
cytoplasmatic promyelocytic leukaemia) auxilia na aproximação dessas proteínas e
receptores no endossomo precoce. A inibição da via do TGF-β acontece através da
translocação de cPML para o núcleo, que se associa com TGIF. Para ativar
novamente a via, galectina-8 (ou PCTA) liga-se ao TGIF, liberando o cPML que volta
ao citoplasma para cumprir sua função (Faresse et al. 2008; Liu 2008).
Para que os ligantes promovam a sinalização, existem fatores de transcrição
para prolongar a resposta intracelular. No caso do TGF-β, a ativação da via pode ser
canônica ou dependente de Smads (do inglês Small mothers against
decapentaplegic homologues), e não-canônica ou independente de Smads. No caso
da via canônica, as Smads são fosforiladas pelo receptor ativado, oligomerizam e
translocam para o núcleo, onde se ligarão ao genoma e irão ativar ou reprimir genes
relacionados à via. Ainda, o complexo Smad3-Smad4 ativa diretamente a expressão
de fatores de transcrição relacionados à transição epitélio-mesenquimal (EMT, do
inglês epitelial-mesenchymal transition), como Snail1, Snail2/Slug, ZEB1/2,
cooperando com esses fatores para ativar uma expressão de genes relacionados ao
fenótipo mesenquimal (Budi et al. 2017; David e Massagué 2018; Gu e Feng 2018).
O controle fino da resposta celular do TGF-β envolve diversos tipos de
Smads: são oito tipos, sendo que Smad1-Smad5 e Smad8 têm função efetora, e
Smad6/7 possuem função inibitória. Todas contêm os domínios N-terminal e C-
terminal, conhecidos como MH1 e MH2. A região MH1 interage com o DNA e MH2
interage com as demais Smads, coativadores e corepressores, como o TGIF (do
inglês Transforming Growth-Interacting Factor) (David e Massagué 2018).
O TβRI fosforila as Smad2 e Smad3 nos dois resíduos de serina na porção C-
terminal, formando uma cauda ácida que media a interação entre a Smad2/3 e
Smad4. A formação do complexo Smad2/3 e Smad4 são necessárias para disparar
a maioria das respostas ao TGF-β (David e Massagué 2018). A minoria dos genes
pode responder apenas à Smad2, sem participação da Smad3; ou mesmo Smad3
pode se combinar à Smad4 e regular alguns outros genes (Budi et al. 2017).
Em resumo, quando TGF-β 1, 2 ou 3 se liga ao TβRII, o TβRI é recrutado e
fosforilado pelo TβRII. O TβRI então fosforila Smad2/3, que recruta Smad4 e formam
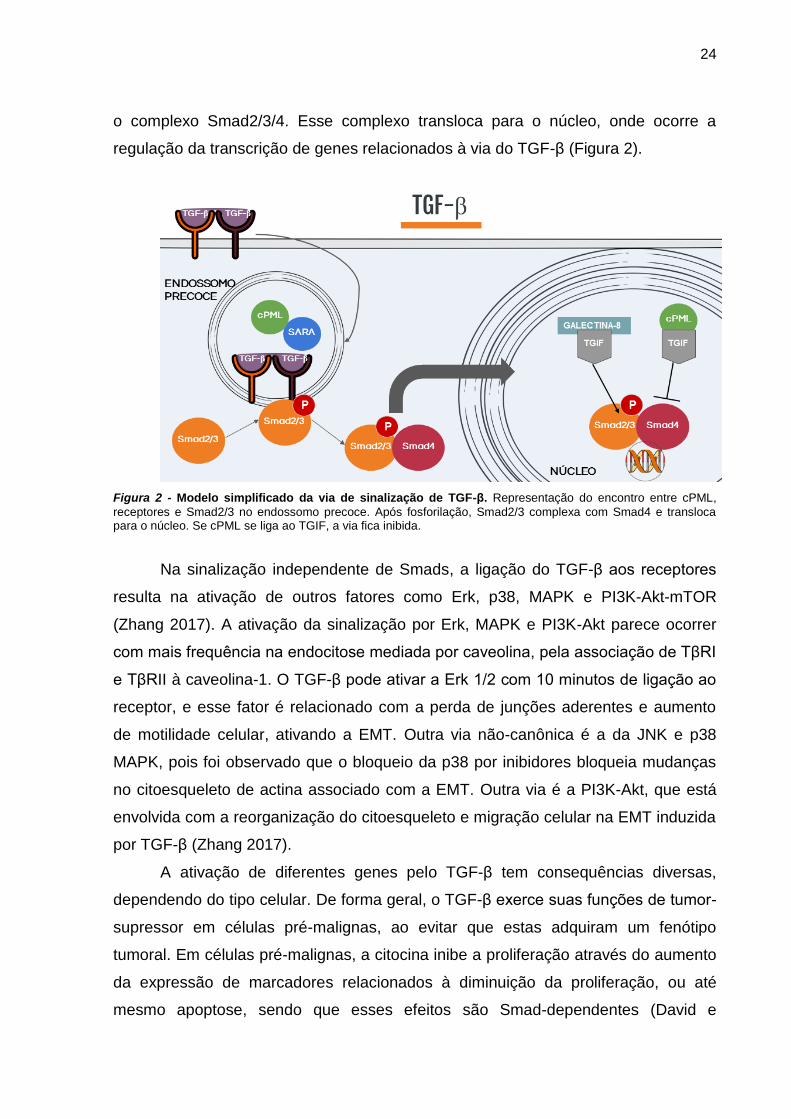
24
o complexo Smad2/3/4. Esse complexo transloca para o núcleo, onde ocorre a
regulação da transcrição de genes relacionados à via do TGF-β (Figura 2).
Figura 2 - Modelo simplificado da via de sinalização de TGF-β. Representação do encontro entre cPML,
receptores e Smad2/3 no endossomo precoce. Após fosforilação, Smad2/3 complexa com Smad4 e transloca para o núcleo. Se cPML se liga ao TGIF, a via fica inibida.
Na sinalização independente de Smads, a ligação do TGF-β aos receptores
resulta na ativação de outros fatores como Erk, p38, MAPK e PI3K-Akt-mTOR
(Zhang 2017). A ativação da sinalização por Erk, MAPK e PI3K-Akt parece ocorrer
com mais frequência na endocitose mediada por caveolina, pela associação de TβRI
e TβRII à caveolina-1. O TGF-β pode ativar a Erk 1/2 com 10 minutos de ligação ao
receptor, e esse fator é relacionado com a perda de junções aderentes e aumento
de motilidade celular, ativando a EMT. Outra via não-canônica é a da JNK e p38
MAPK, pois foi observado que o bloqueio da p38 por inibidores bloqueia mudanças
no citoesqueleto de actina associado com a EMT. Outra via é a PI3K-Akt, que está
envolvida com a reorganização do citoesqueleto e migração celular na EMT induzida
por TGF-β (Zhang 2017).
A ativação de diferentes genes pelo TGF-β tem consequências diversas,
dependendo do tipo celular. De forma geral, o TGF-β exerce suas funções de tumor-
supressor em células pré-malignas, ao evitar que estas adquiram um fenótipo
tumoral. Em células pré-malignas, a citocina inibe a proliferação através do aumento
da expressão de marcadores relacionados à diminuição da proliferação, ou até
mesmo apoptose, sendo que esses efeitos são Smad-dependentes (David e

25
Massagué 2018). Em diferentes tipos celulares, o TGF-β induz a supressão do ciclo
celular (fase G1), por indução de inibidores da CDK (do inglês cyclin-dependent
kinase). Ainda, a indução da apoptose pelo TGF-β depende do contexto celular e
pode ser induzida de forma Smad dependente e independente (Hao, Baker, e ten
Dijke 2019).
No câncer, a história é outra, pois em células malignas o TGF-β age como um
ativador. Conforme o TGF-β age suprimindo populações pré-malignas, são
selecionados clones com Smads modificadas, que irão aumentar os efeitos da via de
invasividade e motilidade (David e Massagué 2018). Considerando que
nanopartículas podem interagir com receptores e a importância da via do TGF-β no
controle de células tumorais, nos perguntamos se existe alguma forma de
nanopartículas afetarem essa via de sinalização em células.
f. Inibidores da via do TGF-β
Estudar a interação entre nanopartículas e células envolve o uso de diversas
técnicas como citometria de fluxo, microscopia confocal e inibidores capazes de
interferir em processos intracelulares (Radaic et al. 2016). O uso de inibidores é uma
ferramenta interessante que permite determinar qual é a via de internalização de
uma nanopartícula. Por exemplo, nosso grupo (A. Radaic e de Jesus (2018))
demonstrou, utilizando inibidores de endocitose, que as SLN são internalizadas
principalmente via clatrina em células embrionárias de rim humano HEK293T.
Os inibidores podem ser classificados como não-competitivos, incompetitivos
ou competitivos pelo sítio ativo (Berg et al. 2002). Para inibidores competitivos, como
aqueles usados nessa dissertação, a constante de dissociação, ou Ki, será menor
quanto mais potente a inibição. A potência de um inibidor de quinase pelo seu alvo
pode ser definida pelo IC50, que representa a concentração do inibidor em que 50%
da atividade da quinase está inibida. No caso de inibidores ATP competitivos essa
relação vai depender também da afinidade do inibidor e ainda da competição pelo
ATP em condições específicas (chamado de Km,ATP). Quando o ATP (substrato) está
em baixas concentrações não há competição pelo sítio ativo e o IC50 será similar ao
Ki, porém conforme a concentração de ATP aumenta e ultrapassa o Km,ATP, são
necessárias maiores concentrações de inibidor para competir, e o IC50 também
aumenta. Dessa forma, a velocidade máxima de reação não se altera, mas sim o Km,

26
que é igual a concentração de substrato em que a velocidade de reação é metade
de seu valor máximo (Berg et al. 2002). Inibidores que se ligam à conformação
inativa da enzima vão competir menos com o ATP, já que sua ação ocorre na
enzima inativa e a previne da ativação, ao invés de inibir a atividade diretamente,
tendo maior eficiência na inibição in vivo (Knight e Shokat 2005).
Através do uso de inibidores, se tornou possível avaliar como as células
respondem à SLN frente à inibição da via do TGF-β, visto que diversos inibidores
têm sido desenvolvidos tendo como alvo enzimas da via do TGF-β. Alguns desses
compostos inibem a sinalização intracelular, agindo em receptores da via ou
bloqueando a ação de Smads (Gu e Feng 2018). Um exemplo é o inibidor da
atividade quinase de TβRI, o Galunisertib (LY2157299), que já passou por testes
clínicos de fase I e fase II e mostrou bons resultados em pacientes de câncer de
pâncreas (Melisi et al. 2018).
Outro exemplo é o LY2109761, uma molécula pequena inibidora do TGF-β,
que foi desenvolvida tendo como alvo tratar angiogênese e metástase, processos
em que o TGF-β tem papel importante (Figura 3). Foi demonstrado que esse inibidor
tem uma seletividade pelos receptores TβRI e TβRII (Luangmonkong et al. 2018).
Em células hepáticas hPLCS tratadas com LY2109761, a fosforilação da Smad2 foi
diminuída em 84% (Luangmonkong et al. 2018). O inibidor diminuiu a migração em
células de carcinoma hepático (He et al. 2017) e suprimiu a invasividade em células
de câncer de próstata C4-2 (Jin et al. 2017).
Outro inibidor é SIS3 HCl, potente inibidor da Smad3 (Figura 3). Estudos
iniciais em fibroblastos humanos demonstraram que SIS3 é capaz de suprimir a
fosforilação de Smad3 na presença do TGF-β em 50% (IC50 = 3 µM) e diminuiu
também em 50% a ligação de Smad3 ao DNA. O SIS3 HCl não alterou a fosforilação
de Smad2 nem a expressão de Smad4, além de não alterar mecanismos em outras
vias como p38, p85 e ERK, se mostrando específico à família do TGF-β (Jinnin, Ihn,
e Tamaki 2006). Chihara et al. (2017) mostrou que SIS3 (IC50 = 10 µM) inibe a
translocação de Smad3 para o núcleo mas não inibe a fosforilação de Smad3, em
células de câncer de mama tratadas previamente com TGF-β durante 14 dias,
sugerindo que SIS3 está envolvido na retenção da Smad3 no citoplasma,
possivelmente através de alguma proteína chave como a SARA. Apesar do
mecanismo de ação de SIS3 ainda não ser completamente elucidado, é importante

27
destacar como cada célula responde de forma diferente ao inibidor, sendo essa
interação célula-inibidor contexto dependente.
Há ainda o SB431542, que inibe especificamente o TβRI, competindo com o
ATP pelo sítio de ligação do TβRI, sendo um inibidor de quinase do tipo I. O TβRI
contém um pequeno resíduo de aminoácido no sítio de ligação ao ATP, o que libera
um espaço que acomoda facilmente o SB431542 (Figura 3) (Ogunjimi et al. 2012).
Os inibidores chamados de pequenas moléculas inibidoras de quinase são divididos
em três classes: tipo I, II e III. Os inibidores do tipo I são pequenas moléculas que
ligam na conformação ativa da quinase no sítio de ligação ao ATP; tipo II se ligam na
conformação inativa da quinase e do tipo III são os inibidores alostéricos. Inibidores
do tipo I competem com o ATP pela ligação ao sítio ativo e mimetizam o anel de
purina da porção adenina do ATP, pois muitas vezes contém um anel heterocíclico
similar à purina. Assim, esses inibidores se ligam no sítio ativo e diminuem a
atividade de fosforilação da proteína (Bhullar et al. 2018).
Figura 3 – Estrutura química de inibidores dos receptores TβRI e TβRII (LY2109761, LY364947 e SB431542)
e Smad (SIS3 HCl) da via do TGF-β.
g. Efeitos biológicos da via do TGF-β: Migração e EMT
A migração celular faz parte da homeostase de um organismo, pois é um
processo essencial na regeneração dos tecidos e órgãos frente a um dano. Em
humanos adultos, a cicatrização de feridas é uma cascata de eventos que inclui
quimiotaxia, divisão celular, neovascularização e deposição de nova matriz

28
extracelular (Kaplani et al. 2018). Apesar de essencial em situações fisiológicas, a
migração é um processo indesejado em células tumorais, pois aumenta a motilidade
dessas células. O aumento da motilidade pode levar ao espalhamento dessas
células pelo corpo, que aumenta as chances de metástase.
O descontrole das células cancerígenas está relacionado a diversas vias de
sinalização, incluindo a via do TGF-β (Lichtman, Otero‐Vinas, e Falanga 2016). No
processo normal de regeneração de feridas, o TGF-β tem papel essencial; o TGF-β3
é a isoforma que prevalece durante as fases iniciais da regeneração, responsável
por aumentar a migração de fibroblastos. Ainda, plaquetas liberam TGF-β1 após o
dano e recrutam outras células para a ferida (Lichtman et al. 2016). Já em cânceres
como gliomas, mama e próstata ocorre uma subversão da via do TGF-β, utilizando-a
para indução de EMT, invasão tumoral e metástase (Gu e Feng 2018).
A EMT é um processo comum na embriogênese, em que células epiteliais
(que possuem adesão coesiva entre as células, baixa motilidade celular) sofrem a
transição para o fenótipo mesenquimal (interação célula-célula perdida, alta
motilidade celular). No câncer, a EMT adquire um novo propósito, permitindo
invasão e migração de células, através da perda de expressão de E-Caderina,
rearranjo do citoesqueleto e aumento da expressão de N-Caderina, Snail e
vimentina (David e Massagué 2018; Gu e Feng 2018; Katsuno et al. 2019).
De modo geral, a EMT induzida pelo TGF-β estimula células tumorais e
contribui para a metástase. No câncer de próstata, TGF-β estimula a EMT
reprimindo a expressão de E-caderina e promovendo a expressão de N-caderina
(Gu e Feng 2018). Considerando o papel da EMT (orquestrada pelo TGF-β)
podemos imaginar quais mecanismos utilizados por nanopartículas para induzir a
migração de células, e que essa indução pode envolver uma ativação direta da via
do TGF-β e EMT.
A EMT pode ser descrita em diversos estágios, possuindo estados
intermediários entre células epiteliais e mesenquimais, chamado de estado parcial
(P-EMT). A ativação da transição envolve diversos fatores, demonstrando a sua
complexidade: em células da pele e de mama, foram encontrados sete estágios
diferentes de EMT. De modo geral, o TGF-β pode ativar a EMT de forma Smad-
dependente ou independente; Smad3 e Smad4 parecem estar mais envolvidas nas
vias Smad-dependente, enquanto que na via Smad-independente, a Erk está

29
envolvida pois tem relação com a remodelação do citoesqueleto, além de outros
fatores como RHOA, RAC e CDC42 (Hao et al. 2019).
h. Metástase
A metástase tumoral é responsável por 90% das mortes por câncer e envolve
diversos processos: células tumorais se distanciam do tumor principal, invadem
através dos tecidos, entram na circulação (sanguínea/linfática), sobrevivem e
extravasam através das veias, onde se estabelecem em novos sítios e proliferam
formando um tumor secundário (Chaffer e Weinberg 2011; Hao et al. 2019). Esses
processos podem ser resumidos em duas fases: (i) a translocação de um câncer de
um tumor primário para um tecido distante e (ii) colonização do novo ambiente.
Para que a fase (i) ocorra, as células do tumor primário devem adquirir a
capacidade de invasão e migração. A invasão permite que as células degradem a
matriz extracelular, abrindo caminhos para as células chegarem ao sistema
circulatório, nessa fase há uma importante reorganização do microambiente tumoral.
Por exemplo, os carcinomas são compostos por células epiteliais e possuem
junções fortes entre elas que as imobilizam no tecido; porém a perda dessas junções
e a capacidade de invasão dão maior motilidade às células. Além disso, essa célula
deve ter a capacidade de auto-renovação e deixar um número ilimitado de
descendentes para que um novo tumor se inicie. O microambiente tumoral possui
células de diversos tipos: fibroblastos, células mesenquimais, granulócitos,
macrófagos, linfócitos, que juntos criam um ambiente de inflamação, resultando na
liberação de sinais como o TGF-β, Wnt e certas interleucinas, que induzem a EMT.
Assim, as células do carcinoma podem responder aos sinais e ativar fatores de
transcrição relacionados à EMT (Chaffer e Weinberg 2011; Lambert, Pattabiraman, e
Weinberg 2017).
Na fase de colonização (ii), as células devem passar por modificações para
serem capazes de se estabelecer em um novo tecido; alguns carcinomas
preferencialmente se estabelecem em certos tecidos, como carcinomas de próstata
que normalmente colonizam o osso, ou metástase do pulmão que é capaz de
colonizar diferentes sítios. O extravasamento da circulação para o novo tecido exige
que fatores sejam liberados para aumentar a permeabilidade da vasculatura: VEGF
(do inglês Vascular Endothelial Growth Factor), MMP e outras moléculas são
importantes para aumentar o extravasamento. A colonização é um sucesso quando

30
as células são capazes de se adaptarem ao novo ambiente, proliferando e gerando
descendentes, além de recrutar células do estroma e suprimento de sangue
(Lambert et al. 2017).
i. PyScratch
Durante o desenvolvimento dos experimentos de migração, foi gerada uma
grande quantidade de imagens para análise da área de migração das células.
Apesar de já existirem alguns softwares disponíveis que realizam essa medição,
como o ImageJ/FIJI (Schindelin et al. 2012), essa análise era feita manualmente,
uma imagem por vez. No nosso caso, cada experimento (placa de 12 poços,
adquirindo imagens a cada 15 minutos durante 48 horas) gera cerca de 1000 fotos.
Dessa forma, em colaboração com Vladimir Gaal (IFGW – UNICAMP),
desenvolvemos o PyScratch, uma nova ferramenta para processar dados de
migração celular (Garcia-Fossa et. al, 2020).
O PyScratch foi desenvolvido e implementado na linguagem de programação
Python, e é um software open-source que analisa as imagens de migração de uma
forma automatizada, sem interferência humana, eliminando influências do usuário no
momento da análise, possibilitando uma análise quantitativa e reprodutível dos
dados. Com o intuito de promover seu uso por toda a comunidade, desenvolvemos
uma interface-usuário que permite a análise por qualquer pesquisador, seja ele
programador ou não. Para utilizar o software acesse:
https://bitbucket.org/vladgaal/pyscratch_public/src/master/. E para maiores
informações acesse www.nanocell.com.br/pyscratch.
j. Resultados preliminares
Resultados preliminares do nosso laboratório indicavam que haveria uma
correlação entre as SLN e via do TGF-β. Ao incubar células da linhagem de câncer
de próstata PC-3 com SLNplex durante 2 horas, encontramos uma intensa
translocação de galectina-8 (ou PCTA) para o núcleo da célula. Interessante
observar que após 4 horas de incubação, a proteína galectina-8 retorna ao
citoplasma (Figura 4). Esse resultado prévio motivou a investigação da inter-relação
da via do TGF-β com a transfecção por SLN, já que a galectina-8 está envolvida com
a ativação da via do TGF-β.
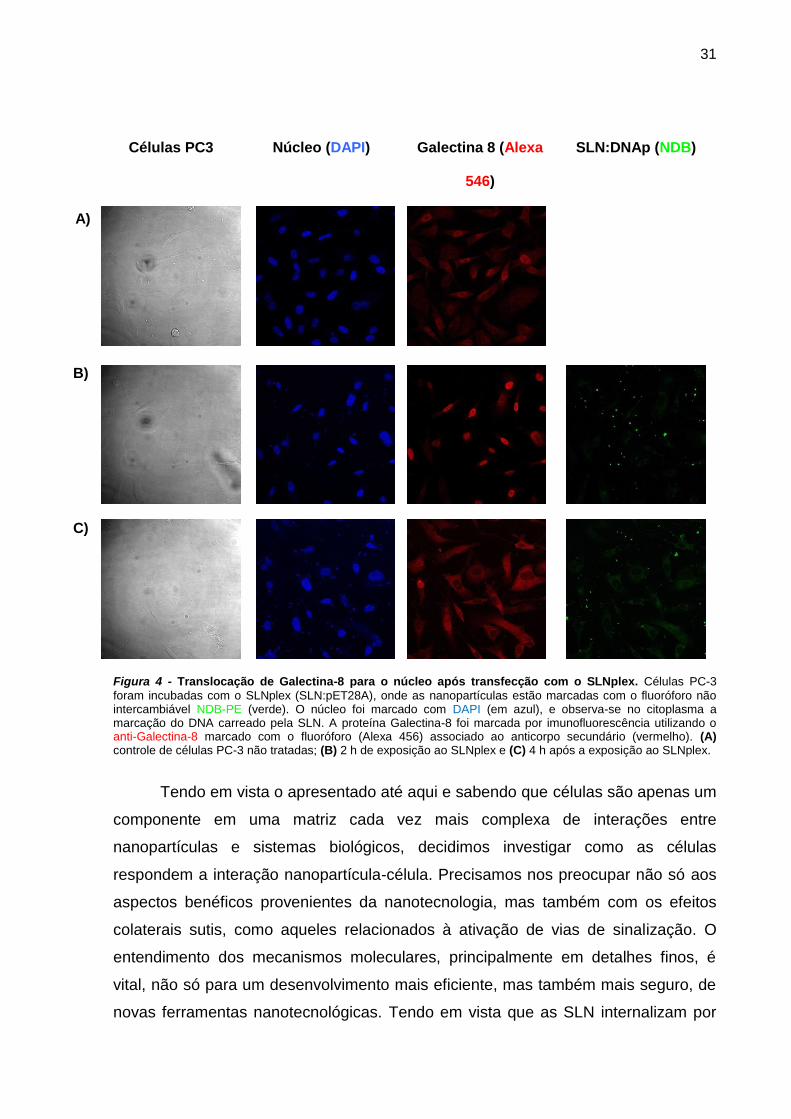
31
Células PC3 Núcleo (DAPI) Galectina 8 (Alexa
546)
SLN:DNAp (NDB)
A)
B)
C)
Figura 4 - Translocação de Galectina-8 para o núcleo após transfecção com o SLNplex. Células PC-3
foram incubadas com o SLNplex (SLN:pET28A), onde as nanopartículas estão marcadas com o fluoróforo não intercambiável NDB-PE (verde). O núcleo foi marcado com DAPI (em azul), e observa-se no citoplasma a marcação do DNA carreado pela SLN. A proteína Galectina-8 foi marcada por imunofluorescência utilizando o anti-Galectina-8 marcado com o fluoróforo (Alexa 456) associado ao anticorpo secundário (vermelho). (A) controle de células PC-3 não tratadas; (B) 2 h de exposição ao SLNplex e (C) 4 h após a exposição ao SLNplex.
Tendo em vista o apresentado até aqui e sabendo que células são apenas um
componente em uma matriz cada vez mais complexa de interações entre
nanopartículas e sistemas biológicos, decidimos investigar como as células
respondem a interação nanopartícula-célula. Precisamos nos preocupar não só aos
aspectos benéficos provenientes da nanotecnologia, mas também com os efeitos
colaterais sutis, como aqueles relacionados à ativação de vias de sinalização. O
entendimento dos mecanismos moleculares, principalmente em detalhes finos, é
vital, não só para um desenvolvimento mais eficiente, mas também mais seguro, de
novas ferramentas nanotecnológicas. Tendo em vista que as SLN internalizam por

32
vias endocíticas que são vias chave para a sinalização do TGF-β, e sua possível
relação, resolvemos estudar os efeitos da SLN na via do TGF-β em células de
próstata.
2. Objetivo
O objetivo desse trabalho foi avaliar os mecanismos moleculares na inter-
relação entre nanopartículas lipídicas sólidas (SLN) e células epiteliais não
tumorais PNT1A e tumorais PC-3 da próstata humana, estabelecendo relação
entre a SLN e seus impactos na via do TGF-β e EMT, considerando o efeito
biológico (i.e. transfecção, migração) nessas células.
Para alcançar o objetivo geral, foram realizados os seguintes objetivos
específicos:
a. Determinar o papel da SLN em diferentes concentrações e seus componentes
na migração celular;
b. Quantificar o papel na proliferação celular ao longo do tempo em presença da
SLN;
c. Determinar a eficiência de transfecção e internalização de SLN em presença
dos inibidores dos receptores do TGF-β;
d. Avaliar a relação entre a via do TGF-β, relacionado à translocação nuclear de
Smad2, e a transfecção por SLN;
e. Avaliar a interação da SLN com as vias do TGF-β e EMT através de proteínas
relacionadas: Smad2 para a via do TGF-β e Zeb1 e Snail para a EMT.
3. Materiais e métodos
a. Cultura de células
Células epiteliais normais de próstata (PNT1A, ECACC 95012614) e células
de câncer de próstata (PC-3, ATCC® CRL-1435™) foram cultivadas em frascos de
cultura de 25 cm2 contendo 5 mL de meio de cultura RPMI 1640 (Invitrogen, Grand
Island, New York), suplementado com 10 % de soro fetal bovino inativado (Gibco,
Brazil), 1% de penicilina/estreptomicina (Gibco, Canadá), chamado de meio RPMI
completo. As células foram mantidas a 37 °C, com atmosfera de 5% de CO2 em
incubadora Panasonic (modelo MCO-170AICUVL-PA).

33
b. Produção e caracterização de nanopartículas lipídicas sólidas (SLN)
Produção: A produção das nanopartículas lipídicas sólidas e seus
componentes (Tabela 1) foram realizados por uma técnica de micro extrusão
desenvolvida por nosso grupo de pesquisa (de Jesus et al. 2013). Esta técnica
consiste na preparação de uma micro emulsão catiônica em água bidestilada
contendo ácido esteárico (Sigma-Aldrich), DOTAP (1,2 Dioleoil-3-propionato de
trimetilamônio, Avanti Polar Lipids Inc., Alabaster, USA), e Pluronic F68 (Sigma-
Aldrich, Alemanha), resultando na concentração final de lipídeos (DOTAP e ácido
esteárico) de 3.8 µg/mL. A micro emulsão foi aquecida 5 °C acima da temperatura
de fusão do ácido esteárico (69 °C). Em seguida o tamanho das partículas foi
uniformizado utilizando uma mini-extrusora (Avanti Polar Lipids Inc., Alabaster, USA)
através da sucessiva passagem (30 vezes) por uma membrana de policarbonato
com poros de 100 nm (IsoporeTM, Millipore). Após 30 passagens, a solução
contendo as nanopartículas lipídicas sólidas foi transferida para um banho de gelo,
para solidificar as SLN e prevenir a agregação das partículas e, em seguida,
estocada a 4 °C para uso futuro (de Jesus e Zuhorn 2015; Radaic et al. 2015). Além
da nanopartícula, também produzimos o ácido esteárico, pluronic F68 e o lipossoma
de DOTAP separadamente, que foram utilizados como controle no experimento de
ensaio de migração celular (Tabela 1).
Caracterização: Após duas semanas em repouso, foi realizada a
caracterização físico-química das nanopartículas, medido o diâmetro médio,
polidispersão (PDI) e potencial Zeta, utilizando o Zetasizer Nanoseries (Malvern
Instruments, Reino Unido). As amostras foram diluídas em água na diluição 1:30 e
submetidas às medidas.
Tabela 1 – Composição das formulações produzidas.
Ácido esteárico (mmol.L-1
) DOTAP (mmol.L-1
) Pluronic F68 (mmol.L-1
)
SLN 7,0 2,5 1,0
Ácido esteárico 7,0 - -
DOTAP - 2,5 -
Pluronic F68 - - 1,0
c. Transfecção por SLN
Células PNT1A e PC-3 foram plaqueadas na densidade de 1x105 células/poço
em placa de 12 poços. No dia seguinte, foi realizada a complexação da SLN (4 μL

34
por poço) com o plasmídeo pEGFP-N1 (Clontech, California) (1 μg de plasmídeo por
poço). O complexo de SLN foi preparado 20 minutos antes da transfecção,
totalizando 100 μL de solução final para cada poço (complexo + meio de cultura
RPMI sem SFB (Soro Fetal Bovino). A solução de SLN foi colocada sobre a solução
de pEGFP-N1 e após 20 minutos, 100 μL foi gotejado sobre cada poço. Após 4h de
experimento, o meio foi substituído por meio RPMI completo. No dia seguinte, as
células foram levadas ao Leitor Multidetecção Híbrido Cytation 5 (BioTek
Instruments, Inc., Winooski, VT, USA) para aquisição de imagens (contraste de fase
e GFP). Logo após, os poços foram lavados com PBS e 300 μL de tripsina 0.1% foi
adicionada. Após cerca de 10 minutos, as células soltaram totalmente. Com mais
100 μL de PBS, as células foram ressuspendidas e colocadas em tubos para
citometria de fluxo. O citômetro de fluxo utilizado foi o FACSCalibur. Os dados foram
analisados no software FlowJo™ (BD, EUA). As células foram selecionadas como
viáveis através de parâmetros de tamanho e granulosidade, ignorando debris
celulares; dentro do gate de células viáveis, selecionamos as células positivas para
GFP com base no controle (nenhuma célula positiva para GFP). Para a
porcentagem de transfecção, foram exportados os dados referentes à Frequency of
Parent, definido como a porcentagem de eventos naquele gate. Para a intensidade
de fluorescência, foram exportados o número total de células e a média da
fluorescência de GFP por célula; esses dois valores foram multiplicados, gerando o
valor de intensidade de fluorescência da pEGFP-N1 na população de células.
d. Ensaio de migração celular
Células PNT1A e PC-3 foram plaqueadas na densidade de 5x104 células/mL
em placa de 24 poços. Densidade de células: O valor de confluência ideal para o
ensaio de migração celular foi determinado segundo estudos prévios (Garcia-Fossa
et al. 2020). Para esse experimento, PNT1A e PC-3 foram plaqueadas na densidade
de 2,5x104, 5x104 e 10x104 células/mL em placa de 12 poços utilizando meio RPMI
completo. As células ficaram em privação de SFB por 24h, para tentar diminuir a
proliferação celular, e no dia seguinte foi feito o risco com o auxílio de uma ponteira
p200. As imagens foram adquiridas no Cytation 5, utilizando objetiva 4x no contraste
de fase, a cada 15 min durante 60h. analisadas com o auxílio do PyScratch e a
velocidade de migração das células foi calculada. Nós observamos uma correlação
linear positiva entre a densidade de células e a velocidade, mostrando que a

35
densidade influencia no aumento da velocidade de migração. Assim, a densidade de
5x104 células/mL foi determinada ideal para os ensaios dessa dissertação.
Para esse ensaio foi utilizado o plasmídeo pET28A que possui expressão
gênica somente em bactérias, portanto seu uso é apenas para garantir as
características físico-químicas do complexo SLN:plasmídeo, que irá interagir com as
células. Para todos os ensaios de migração, as células foram mantidas em privação
de SFB por 24h previamente ao experimento.
Migração celular na presença de SLN: as células foram transfectadas com
SLN:pET28A (4 µL) e TGF-β (10 ng/mL) durante 4 h. A concentração de TGF-β a 10
ng/mL foi escolhida com base na literatura, já que essa concentração ativa vias
dependente de Smad e PI3K em células PC-3 (Barrett, Millena, e Khan 2017;
Chaudhry et al. 2014; Vo et al. 2013). Migração celular na presença de SLN e
seus componentes: as células foram transfectadas durante 4 h com TGF-β 10
ng/mL, SLN:pET28A (4 µL), e os componentes em concentração equivalentes
àquelas da transfecção com SLN: Ácido esteárico (4 µL), Pluronic F68 (4 µL) e
lipossoma de DOTAP (4 µL). Migração celular na presença de SLN em diferentes
concentrações: O complexo de SLN: pET28A foi formado na concentração de 7.6
µg/mL, o dobro da concentração normalmente utilizada de SLN. Após a formação do
complexo, a SLN:pET28A foi diluída em meio RPMI sem SFB para as concentrações
de 3.8 µg/mL (1x), 0.76 µg/mL (0.2x), 0.38 µg/mL (0.1x) e 0.076 µg/mL (0.02x).
Células PC-3 foram tratadas durante 4 h.
Após 4 h, utilizando uma pipeta p200 foi realizado um arranhado em linha
reta em cada poço, segurando a pipeta em um ângulo de 30°. As células foram
lavadas com PBS (do inglês Phosphate-Buffered Saline) 1x. O acompanhamento da
migração foi feito através de aquisição de imagens em contraste de fase, objetiva de
4x, pelo Leitor Multidetecção Híbrido Cytation 5 (BioTek Instruments, Inc., Winooski,
VT, USA), e as imagens foram adquiridas a cada quinze minutos. Ao final do
experimento, as imagens foram analisadas através do programa PyScratch,
desenvolvido pelo nosso grupo em colaboração com Vladimir Gaal (IFGW -
UNICAMP) com o intuito de facilitar a análise da migração no tempo, medindo a área
de migração em pixels de forma automatizada e garantindo a imparcialidade na
análise (Garcia-Fossa et al. 2020). Utilizando os valores da área (eixo y) versus o
tempo em minutos (eixo x), a velocidade de migração foi calculada: inclinação da
curva / 2 x comprimento da imagem em pixels. O valor de velocidade, em pixels/min,

36
foi convertido para µm/min através do valor de conversão da objetiva utilizada para
adquirir as imagens (1,6 µm/pixel).
Para o cálculo do tempo de fechamento do risco, foi utilizado como threshold
o tempo que as células demoram a atingir 5% do valor da área (0.05) total do risco.
Fizemos uma rotina em Python 3.7.3 utilizando o valor normalizado da área do risco,
que começa em um e termina próximo de zero. É criado um loop (for x in
range(len(df['Area normalizada'])) que compara os valores de x ao longo da área
normalizada; quando x é menor do que 0.05, o loop para e retorna o tempo referente
a x < 0.05 (if df[''Area normalizada'][x] < 0.05: break). Assim, obtivemos em minutos
o tempo que as células demoram para chegar em 5% da área, que depois foi
convertido em h.
e. Proliferação celular
Click-iT™ Plus EdU Alexa Fluor™ 647 Imaging Kit: Células PNT1A e PC-3 foram
plaqueadas em lamínulas 13 mm pré-tratadas com L-polilisina (Sigma-Aldrich),
densidade de 5x104 células/poço, e após adesão, o meio de cultura RPMI 1640
completo foi trocado por meio de cultura RPMI 1640 sem SFB, e mantidas em
privação de soro (starvation) durante 24 h em incubadora Panasonic, a 37 °C e 5%
de CO2. Após 24 h, o complexo entre SLN (4 µL, 3.8 µg/mL) e pET28A (1 µg) foi
formado. Os poços foram lavados com PBS e adicionou-se 200 μL de meio de
cultura RPMI 1640 e tratou-se as células com 50 μL de solução SLN:pET28A. Após
4 h, o meio de transfecção foi retirado e lavou-se o poço uma vez com PBS. Por fim,
adicionou-se meio de cultura RPMI 1640 sem SFB contendo EdU (5-ethinil-
2‟deoxiuridina) a 10 μM. Após 5h, 24h e 48h de incubação, os poços
correspondentes ao tempo foram fixados com PFA 4%, e as células fixadas foram
lavados com solução bloqueadora SFB/PBS 10% por duas vezes. Adicionou-se
0.5% de Triton X-100 (PBS) durante 20 minutos. Enquanto isso foi preparado a
solução responsável pela reação Click it: Reaction Buffer 1x, Copper Protectant,
Alexa Fluor® picolyl azide e Buffer additive 1x. As células foram lavadas novamente
com SFB 10%. Foi adicionado 30 μL da solução por poço e a incubou-se as
lamínulas por 30 minutos, a temperatura ambiente. Após 30 minutos, foram lavadas
com SFB 10%, PBS e adicionou-se solução de Hoechst 33342 (Sigma-Aldrich)
1:2000. Incubou-se por 30 minutos a temperatura ambiente, depois as células foram
lavadas duas vezes com PBS e as lâminas foram montadas utilizando meio de

37
montagem para fluorescência Dako (Agilent Dako, California). As fotos foram
capturadas no Leitor Multidetecção Híbrido Cytation 5 (BioTek Instruments, Inc.,
Winooski, VT, USA) na objetiva de 10x e 18 sítios foram adquiridos por tratamento.
Para contagem de núcleos (Hoechst) e núcleos positivos para proliferação (Alexa
Fluor 647) foi utilizado o software aberto CellProfiler 3.1.8 (Carpenter et al. 2006)
através de uma rotina desenvolvida, que identifica os objetos primários, tanto núcleo
total (Hoechst) quanto núcleos positivos para proliferação (Alexa Fluor 647). Após, o
dado é exportado em formato csv (do inglês comma-separated values) contendo
todos os objetos (núcleos) identificados nas imagens e a razão entre núcleos
positivos para proliferação e células totais (EdU/Hoechst * 100), que fornece a Taxa
de Proliferação. Os gráficos foram plotados utilizando a linguagem em Python 3.7
Software Foundation (Python Language Reference, version 3.7. Disponível em
http://www.python.org) com o auxílio da biblioteca de visualização de dados Seaborn
(Disponível em https://seaborn.pydata.org).
f. Avaliação da citotoxicidade dos inibidores
Células PNT1A e PC-3 foram plaqueadas na densidade de 1x104 células/poço
em placa de 96 poços. No dia seguinte, as diluições foram preparadas para os
inibidores: LY2109761, solução estoque a 5 mM em DMSO, foi diluído nas seguintes
concentrações em meio RPMI: 0,625 µM, 1,25 µM, 2,5 µM, 5 µM, 10 µM, 20 µM, 40
µM e 80 µM. O SIS3 HCl, solução estoque a 5 mM em DMSO, foi diluído nas
concentrações também em meio RPMI: 0,375 µM, 0,75 µM, 1,5 µM, 3 µM, 6 µM, 12
µM, 24 µM e 48 µM. Após tratar as células em triplicata, elas foram deixadas em
contato com o inibidor durante 24 h. No dia seguinte, o controle positivo (Mortas) foi
tratado durante 15 minutos com 50 µL de DMSO, para matar as células totalmente.
Logo após a solução de homodímero de etídio (EthD-1) (Santa Cruz Biotechnology,
Dallas) foi diluído na concentração final de 1 µM em Fluorobrite-DMEM, um meio de
cultura incolor (Gibco, Canadá) e Hoechst 33342 (Sigma-Aldrich) foi adicionado na
concentração de 8 µM, para marcar todos os núcleos. Essa solução contendo EthD-
1 e Hoechst foi colocada nas células durante meia h. Ao final, as imagens foram
adquiridas no Leitor Multidetecção Híbrido Cytation 5 (BioTek Instruments, Inc.,
Winooski, VT, USA) com objetiva de 4x em montagem de 4x4 para adquirir imagens
do poço todo. Cerca de 30.000 células foram contabilizadas por poço utilizando o
CellProfiler 3.8.1 (Carpenter et al. 2006). Através da identificação do núcleo

38
(marcação com Hoechst), foram contadas todas as células do poço, valor que foi
definido como 100% das células. Para contabilizar as células mortas, utilizamos o
número de células marcado com EthD-1, utilizado para determinar a porcentagem
de células mortas.
g. Determinação da transfecção na presença dos inibidores
Células PNT1A e PC-3 foram plaqueadas na densidade de 1x105 células/poço
em placa de 12 poços. No dia seguinte, as células foram incubadas com LY2109761
(5 µM) ou SIS3 HCl (1.5 µM) durante 1 h. Após 1 h de incubação, os inibidores foram
removidos e as células foram tratadas com o complexo da SLN (4 μL) com o
plasmídeo pEGFP-N1 (1 μg de plasmídeo por poço). O complexo de SLN foi
preparado 20 minutos antes da transfecção, totalizando 100 μL de solução final para
cada poço (complexo + meio de cultura RPMI sem SFB). A solução de SLN foi
colocada sobre a solução de pEGFP-N1 e após 20 minutos, 100 μL dessa solução
foi pingada sobre cada poço. Após 4h de experimento, o meio foi substituído por
meio RPMI completo. No dia seguinte, os poços foram lavados com PBS e 300 μL
de tripsina 0.1% foram adicionadas. Após cerca de 10 minutos, as células soltaram
totalmente. Com mais 100 μL de PBS, as células foram ressuspendidas e colocadas
em tubos para citometria de fluxo. O citômetro de fluxo utilizado foi o FACSCalibur.
Os dados foram analisados no software FlowJo™ (BD, EUA). As células foram
selecionadas como viáveis através de parâmetros de tamanho e granulosidade,
ignorando debris celulares; dentro do gate de células viáveis, selecionamos as
células positivas para GFP com base no controle (nenhuma célula positiva para
GFP). Para a porcentagem de transfecção, foram exportados os dados referentes à
Frequency of Parent, definido como a porcentagem de eventos naquele gate. Para a
intensidade de fluorescência, foram exportados o número total de células e a média
da fluorescência de GFP por célula; esses dois valores foram multiplicados, gerando
o valor de intensidade de fluorescência da pEGFP-N1 na população de células.
h. Determinação da Internalização dos complexos em presença de
inibidores
Células PNT1A e PC-3 foram plaqueadas na densidade de 1x105 células/poço
em placa de 12 poços. No dia seguinte, as células foram tratadas com LY2109761 (5
µM) ou SIS3 HCl (1,5 µM). Após 1 h de incubação, os inibidores foram removidos e

39
as células foram tratadas com o complexo da SLN-Rodamina (4 μL) com o
plasmídeo pET28A (1 μg de plasmídeo por poço). O complexo de SLN foi preparado
20 minutos antes da transfecção, totalizando 100 μL de solução final para cada poço
(complexo + meio de cultura RPMI sem SFB). A solução de SLN foi colocada sobre
a solução de pET28A e após 20 minutos, 100 μL foi gotejado sobre cada poço. Após
4h de experimento, as células foram preparadas para citometria de fluxo. Os poços
foram lavados com PBS e 300 μL de tripsina 0.1% foram adicionadas. Após cerca de
10 minutos, as células soltaram totalmente. Com mais 100 μL de PBS, as células
foram ressuspendidas e colocadas em tubos para citometria de fluxo. O citômetro de
fluxo utilizado foi o FACSCalibur. Os dados foram analisados no software FlowJo™
(BD, EUA). As células foram selecionadas como viáveis através de parâmetros de
tamanho e granulosidade, ignorando debris celulares; dentro do gate de células
viáveis, selecionamos as células positivas para rodamina (FL-2) com base no
controle. Para a porcentagem de transfecção, foram exportados os dados referentes
à Frequency of Parent, definido como a porcentagem de eventos naquele gate. Para
a intensidade de fluorescência, foram exportados o número total de células e a
média da fluorescência de rodamina por célula; esses dois valores foram
multiplicados, gerando o valor de intensidade de fluorescência da SLN-Rodamina na
população de células.
i. Avaliação de proteínas da via do TGF-β e EMT
Smad2: Células PNT1A e PC-3 foram plaqueadas em lamínulas 13 mm pré-
tratadas com L-polilisina (Sigma-Aldrich) para melhor adesão das células, na
densidade de 5x104 células/poço em placa de 24 poços. No dia seguinte, as células
foram tratadas com TGF-β 10 ng/mL em meio RPMI sem SFB, ou mantidas apenas
em meio RPMI sem SFB (Controle). Após 45 minutos, 1 h, 2 h e 4 h de tratamento,
as células foram fixadas com PFA 4% e preparadas para imunofluorescência de
Smad2.
EMT – Snail e Zeb1: Células PNT1A e PC-3 foram plaqueadas em lamínulas
13 mm pré-tratadas com L-polilisina (Sigma-Aldrich) para melhor adesão das células,
na densidade de 5x104 células/poço em placa de 24 poços. No dia seguinte, 2 µL de
SLN foi adicionada a 0.5 µg do plasmídeo pET28A, formando o complexo de
SLN:pET28A. O complexo de SLN foi preparado 20 minutos antes da transfecção,
totalizando 50 μL de solução final para cada poço (complexo + meio de cultura RPMI

40
sem SFB). As células também foram tratadas com TGF-β 10 ng/mL em meio RPMI
sem SFB, ou mantidas apenas em meio RPMI sem SFB (Controle). Após 4 h de
tratamento, o meio de tratamento foi substituído por meio RPMI sem SFB e as
células mantidas na incubadora por 24h. Após 24h, as células foram fixadas com
PFA 4% e preparadas para imunofluorescência das proteínas Snail e Zeb1.
Vimentina: Células PNT1A e PC-3 foram plaqueadas em placa de 24 poços
na densidade de 5x104 células/poço. Após 24h em privação de SFB, as células
foram transfectadas durante 4h com SLN:pET28A e TGF-β 10 ng/mL. Após 4h, o
poço foi riscado com o auxílio de uma ponteira p200, as células foram lavadas com
PBS 1x e o meio substituído por RPMI. Ao final de 12h de migração, as células
foram fixadas com PFA 4% por 20 minutos, e preparadas para imunofluorescência
da proteína Vimentina.
j. Imunofluorescência e anticorpos
Para a marcação de imunofluoescência, as células submetidas aos
tratamentos, como exemplificado em Material e Métodos seção i, as lamínulas
contendo as células foram incubadas com glicina 0.1 M durante 20 minutos e
permeabilizadas em solução de Triton X-100 (0.2%) durante 2 minutos. Após a
incubação em solução bloqueadora (soro fetal bovino 10%) durante 10 minutos, as
lamínulas foram incubadas com os anticorpos indicados na Tabela 2 durante 1 h a
37 °C. As lamínulas foram lavadas em PBS 1x por 5 minutos, por três vezes e
incubadas com o anticorpo secundário Alexa Fluor®488 produzido em cabra anti-
coelho (Invitrogen, Grand Island, New York) e DAPI (núcleo) durante meia h
protegidas da luz. As lâminas foram montadas em meio de montagem para
fluorescência Dako (Agilent Dako, California) e as imagens foram adquiridas no
aparelho Leitor Multidetecção Híbrido Cytation 5 (BioTek Instruments, Inc., Winooski,
VT, USA) utilizando objetiva de 20x.
Tabela 2 - Anticorpos primários e suas respectivas diluições.
Anticorpo Marca Diluição
Smad2 (D43B4) XP Rabbit mAb Cell Signaling Technology, EUA 1:200
Snail (C15D3) Rabbit mAb Cell Signaling Technology, EUA 1:200
TCF8/ZEB1 (D80D3) Rabbit mAb Cell Signaling Technology, EUA 1:300
Vimentin (V6389) Mouse mAb Sigma-Aldrich 1:100

41
k. Análise de imagens
Todas as imagens foram submetidas ao controle de qualidade, em uma rotina
desenvolvido no CellProfiler 3.8.1 (Bray e Carpenter 2018; Carpenter et al. 2006)
que avalia diversos parâmetros para cada imagem do experimento (módulo
MeasureImageQuality: intensidade, textura, foco, saturação de pixels) e exporta os
dados em formato .db, gerando uma base de dados em SQLite. Esse arquivo é
aberto no software CellProfiler Analyst 2.2.1 (Dao et al. 2016), onde é possível
classificar as imagens em positivas (ou seja, se a imagem está em foco, inexistência
de artefatos) e em negativas (fora de foco, artefato presente) utilizando o
classificador Fast Gentle Boosting, que retorna as 10 características mais
importantes utilizadas pelo classificador para separar as imagens em positivas e
negativas. Um arquivo de texto é gerado contendo tais características, que depois é
adicionado em uma rotina de análise das imagens; caso a imagem contenha as
características negativas, a imagem não é utilizada no processo de análise,
garantindo que analisamos apenas as imagens de qualidade.
Smad2: Utilizando uma rotina no CellProfiler 3.8.1 (Carpenter et al. 2006), as
células foram identificadas através do núcleo (marcação com DAPI), criando um
objeto primário. A partir da imagem da Smad2, o programa identifica o objeto
secundário, que é a célula inteira. O objeto terciário (citoplasma) é formado através
da subtração do objeto secundário do objeto primário. Outros módulos são
adicionados para medida da intensidade integrada (ou seja, a soma) dos pixels de
todos os objetos, medida de tamanho e forma da célula, textura das imagens,
gerando uma tabela que contém mais de 400 colunas. Os gráficos foram plotados
utilizando a linguagem em Python 3.7 Software Foundation (Python Language
Reference, version 3.7. Disponível em http://www.python.org) com o auxílio da
biblioteca de visualização de dados Seaborn (Disponível em
https://seaborn.pydata.org).
EMT – Snail e Zeb1: Utilizando uma rotina no CellProfiler 3.8.1 (Carpenter et
al. 2006), as células são identificadas através do núcleo (marcação com DAPI),
criando um objeto primário. A partir da imagem da Snail ou Zeb1, dependendo da
proteína marcada, o programa identifica o objeto secundário, que é a célula inteira.
O objeto terciário (citoplasma) é formado através da subtração do objeto secundário
do objeto primário. Outros módulos são adicionados para medida da intensidade

42
integrada (ou seja, a soma) dos pixels de todos os objetos, medida de tamanho e
forma da célula, textura das imagens, gerando uma tabela que contém mais de 400
colunas. Os gráficos foram plotados utilizando a linguagem em Python 3.7 Software
Foundation (Python Language Reference, version 3.7. Disponível em
http://www.python.org) com o auxílio da biblioteca de visualização de dados Seaborn
(Disponível em https://seaborn.pydata.org).
l. Análise estatística dos resultados
Os dados de Transfecção, Internalização e Migração foram analisados através
do software GraphPad Prism 6, diferença significativa por Two-way ANOVA seguido
de teste de Tukey (comparação das médias com todas as demais médias)
considerando p < 0,05.
Os dados de Proliferação, Translocação de Smad2, Validação dos Inibidores,
Proteínas da EMT foram analisadas utilizando a linguagem em Python 3.7 Software
Foundation (Python Language Reference, version 3.7. Disponível em
http://www.python.org) com o auxílio da biblioteca scipy.stats para o teste t de
Student que compara as médias de duas amostras independentes, assumindo que
as duas populações tem variâncias idênticas (hipótese nula), considerando p < 0,05
para descartar a hipótese nula.
4. Resultados e discussão
a. Caracterização das nanopartículas lipídicas sólidas (SLN)
As nanopartículas lipídicas sólidas (SLN) foram produzidas como descrito nos
Materiais e Métodos. Medimos seu diâmetro médio, índice de polidispersidade (PDI,
do inglês Polidispersity Index) e potencial Zeta, através de espalhamento de luz
dinâmico (DLS, do inglês Dynamic Light Scattering) (Tabela 3). As SLN são diluídas
em água; por serem submetidas ao meio polar, contendo íons, as nanopartículas
atraem moléculas de carga oposta, o que forma uma dupla camada elétrica.
Determina-se o plano de ruptura entre a carga adsorvida na nanopartícula e o meio
em volta, chamado de potencial Zeta.
A caracterização das nanopartículas foi realizada previamente à sua utilização
na cultura celular. O tamanho e potencial Zeta das nanopartículas ficaram dentro
dos valores esperados, conforme trabalhos anteriores do grupo (de Jesus et al.

43
2014; Radaic e Jesus 2018), possuindo tamanho de cerca de 100 nm e carga
positiva, apontado pelo potencial Zeta (Tabela 3). O valor de PDI revela quão
homogêneo é a amostra; valores abaixo de 0,05 indicam soluções monodispersas,
enquanto que valores maiores que 0,7 indicam soluções heterogêneas, que não
podem ser avaliadas através de DLS (Danaei et al. 2018; Stetefeld, McKenna, e
Patel 2016). No caso das nossas nanopartículas, o valor de PDI mostra que as
soluções possuem nanopartículas de tamanho homogêneo (Tabela 3), sendo assim
adequadas para a utilização em cultura de células.
Tabela 3 – Caracterização das nanopartículas. Duas semanas após a produção, as nanopartículas foram levadas ao ZetaSizer Nano series e foi avaliado o diâmetro, PDI e potencial Zeta. Os valores médios são apresentados ± o desvio padrão (DP).
Nanopartícula Diâmetro (nm) ± DP PDI Potencial Zeta (mV) ± DP
SLN 113,03 ± 40,65 0,08 34,36 ± 19,43 SLN-Rodamina 88,54 ± 36,05 0,17 19,96 ± 34,85
Lipossoma DOTAP 119,20 ± 27,57 0,02 37,60 ± 10,20
b. Eficiência de transfecção
Para determinar a eficiência de transfecção pelas nanopartículas SLN, SLN-
Rodamina e DOTAP, realizamos a complexação das nanopartículas ao plasmídeo
pEGFP-N1. Quando o plasmídeo é entregue às células pelas nanopartículas, ele é
codificado e transcrito, levando à expressão da proteína fluorescente verde (GFP, do
inglês Green Fluorescent Protein). O GFP é utilizado como um sistema repórter, que
possibilita quantificar a porcentagem de células que expressam GFP e
consequentemente, as células que foram transfectadas pela nanopartícula.
Como controle positivo foi utilizado Lipofectamina ™ 2000 (LF2K 2000), um
agente comercial formado por lipossoma catiônico, considerado padrão-ouro por
entregar diversos tipos de material genético para diversos tipos celulares (Cardarelli
et al. 2016). Como podemos observar na Figura 5 A, LF2K 2000 é capaz de
transfectar ambas as linhagens PNT1A e PC-3 com a mesma eficiência (ca. 80%),
sem diferença significativa.
No caso da SLN, SLN-Rodamina e DOTAP, a eficiência de transfecção foi a
mesma dentro de cada linhagem celular (Figura 5 A). Porém quando comparamos a
eficiência de transfecção de SLN, SLN-Rodamina e DOTAP entre as linhagens,
observamos 60% de transfecção na PC-3, enquanto que na PNT1A a transfecção

44
cai para cerca de 20%. Os valores de transfecção em PC-3 condizem com as taxas
de transfecção por SLN publicadas por nosso grupo de pesquisa, na faixa de 60%
de transfecção (de Jesus et al. 2014; Radaic et al. 2015). Além disso, a intensidade
de fluorescência do GFP também foi quantificada (Figura 5 B). A intensidade é
calculada através da média de fluorescência do GFP multiplicado pelo o número de
células transfectadas. Esse valor diz sobre os níveis totais de expressão das
proteínas GFP nas células. Nesse caso, o perfil de intensidade foi parecido com
aquele encontrado para a eficiência transfecção (para todos os grupos).
A diferença de transfecção por nanopartículas em células normais (PNT1A) e
tumorais (PC-3) é interessante, pois mostra uma preferência da ação dessas
nanopartículas na célula tumoral. A busca por nanopartículas que tem como alvo
especificamente células tumorais é constante na terapia gênica, pois possuem
características que afeta apenas células tumorais e causa um efeito direto e
específico (Roacho-Perez et al. 2018). Após a caracterização das formulações
usadas nesse estudo e de que seu efeito biológico era similar aquele encontrado em
trabalhos anteriores do grupo, passamos para a avaliação na nossa hipótese de que
nanopartículas lipídicas sólidas têm efeito na migração de células de próstata.
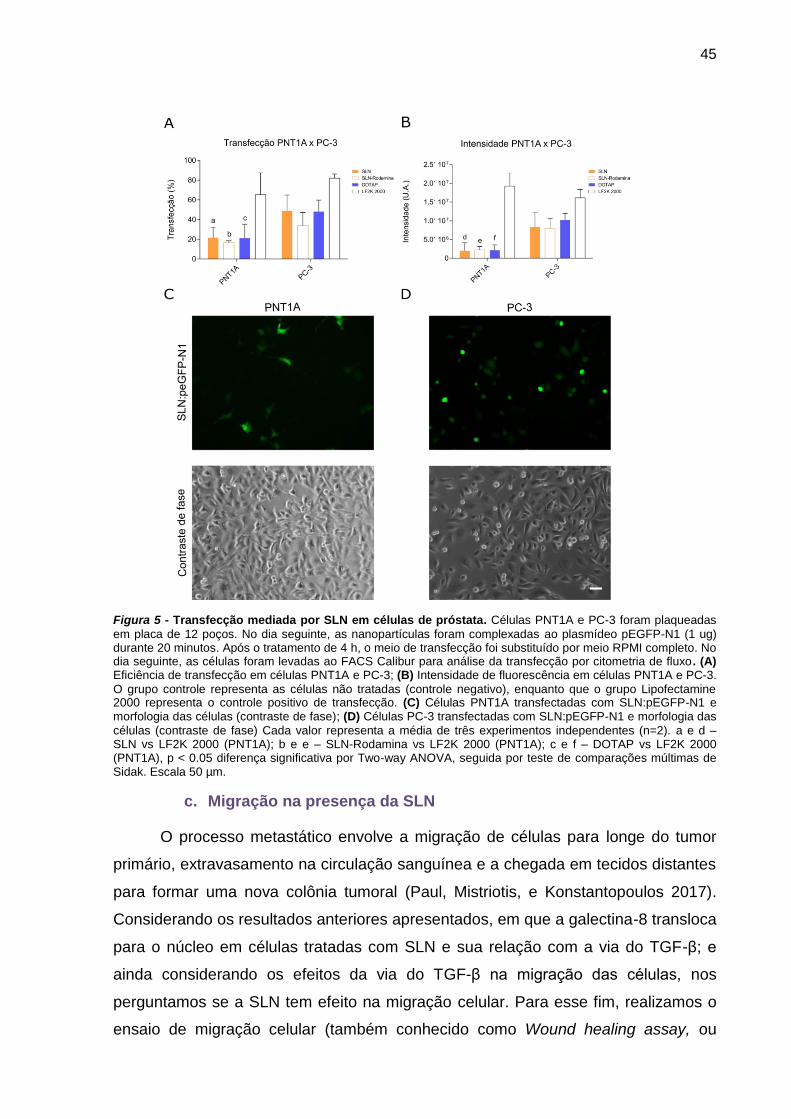
45
Figura 5 - Transfecção mediada por SLN em células de próstata. Células PNT1A e PC-3 foram plaqueadas
em placa de 12 poços. No dia seguinte, as nanopartículas foram complexadas ao plasmídeo pEGFP-N1 (1 ug) durante 20 minutos. Após o tratamento de 4 h, o meio de transfecção foi substituído por meio RPMI completo. No dia seguinte, as células foram levadas ao FACS Calibur para análise da transfecção por citometria de fluxo. (A) Eficiência de transfecção em células PNT1A e PC-3; (B) Intensidade de fluorescência em células PNT1A e PC-3.
O grupo controle representa as células não tratadas (controle negativo), enquanto que o grupo Lipofectamine 2000 representa o controle positivo de transfecção. (C) Células PNT1A transfectadas com SLN:pEGFP-N1 e morfologia das células (contraste de fase); (D) Células PC-3 transfectadas com SLN:pEGFP-N1 e morfologia das
células (contraste de fase) Cada valor representa a média de três experimentos independentes (n=2). a e d – SLN vs LF2K 2000 (PNT1A); b e e – SLN-Rodamina vs LF2K 2000 (PNT1A); c e f – DOTAP vs LF2K 2000 (PNT1A), p < 0.05 diferença significativa por Two-way ANOVA, seguida por teste de comparações múltimas de Sidak. Escala 50 µm.
c. Migração na presença da SLN
O processo metastático envolve a migração de células para longe do tumor
primário, extravasamento na circulação sanguínea e a chegada em tecidos distantes
para formar uma nova colônia tumoral (Paul, Mistriotis, e Konstantopoulos 2017).
Considerando os resultados anteriores apresentados, em que a galectina-8 transloca
para o núcleo em células tratadas com SLN e sua relação com a via do TGF-β; e
ainda considerando os efeitos da via do TGF-β na migração das células, nos
perguntamos se a SLN tem efeito na migração celular. Para esse fim, realizamos o
ensaio de migração celular (também conhecido como Wound healing assay, ou

46
ainda Scratch assay), em que um risco é feito no centro do poço e acompanha-se a
migração das células em direção ao preenchimento da área do risco. Para diminuir a
contribuição da proliferação no processo de fechamento do risco, utilizamos privação
de SFB por 24 h previamente ao tratamento (Grada et al. 2017). Para efeitos de
comparação foi utilizado o TGF-β a 10 ng/mL, já que essa citocina é conhecida por
induzir transformações pró-invasiva em células tumorais de próstata, aumentando a
motilidade celular (Kimbrough-Allah, Millena, e Khan 2018; Vo et al. 2013).
Em células normais PNT1A não houve aumento significativo da migração,
seja na presença da SLN ou TGF-β (Figura 6 A, mediana para controle é de 0.19
µm/min, SLN 0.20 µm/min e TGF-β 0.15 µm/min). O tratamento de 4 h com TGF-β
não foram suficientes para observarmos aumento na migração das células PNT1A.
Entretanto, para incubações mais longas, Freire-de-Lima et al. 2011 demonstrou
aumento no número de células PNT1A na área do risco, após 72 h de tratamento
com TGF-β. Em nossos tratamentos, a similaridade entre os tratamentos reflete no
tempo de fechamento do risco: o controle leva em média 40h, SLN 36h e TGF-β 35h
(Figura 6 B).
No caso das células PC-3, a SLN duplicou a velocidade de migração em
relação ao controle (Figura 6 A, mediana para Controle: 0.31 µm/min, SLN: 0.59
µm/min e TGF-β 0.42 µm/min, i p<0.05). Essa diferença reflete principalmente no
tempo de fechamento do risco: a SLN estimula o fechamento completo em 12h,
enquanto que o controle leva em média 30h (Figura 6 B, ii p<0.05). O TGF-β
estimula a migração, mas o efeito é sutil e o risco fecha em 23h (Figura 6 B).
Podemos acompanhar no tempo o fechamento dos riscos através dos vídeos
representativos disponíveis em https://youtu.be/oJjWehtLyC8 para PNT1A e em
https://youtu.be/fJSd_mUtNBY para PC-3.
O TGF-β é conhecido por aumentar a migração em células PC-3 em tempos
prolongados de exposição ao TGF-β; Walker et al. 2013 mostrou que os tratamentos
com TGF-β1 e TGF-β3 foram capazes de aumentar a migração de células PC-3,
mas tal efeito foi observado na exposição das células ao TGF-β por 48h. No caso
desse ensaio, a citocina foi utilizada para fins de comparação com a SLN, portanto
as células foram expostas apenas por 4h aos tratamentos; sendo assim, o tempo de
exposição à citocina poderia explicar a razão de não observarmos aumento da
migração de PC-3 na presença do TGF-β.

47
Na literatura encontramos trabalhos que relatam nanopartículas que causam
tanto a inibição da migração de células tumorais, quanto aumento da migração em
células importantes para a regeneração tecidual. Guo et al. (2015) mostrou que
nanopartículas de zinco diminuíram a migração de células tumorais de retina de
forma concentração-dependente, ou nanopartículas de grafeno que inibem a
migração de células de glioblastoma pela diminuição de moléculas de adesão
(Wierzbicki et al. 2017). O aumento da migração é relatado para nanopartículas de
quitosana-cobre na regeneração de células da córnea (Tellios et al. 2018), ou
nanopartículas de prata-tanino que aumentam a migração de queratinócitos
(Orlowski et al. 2018). Tais estudos são essenciais para descoberta de novas
terapias, porém pouco é descrito sobre nanopartículas que demonstram efeitos
tóxicos – no nosso caso, a SLN possui alta eficiência de transfecção em PC-3 e em
contrapartida aumenta a migração dessas células significativamente. Assim,
destacamos a importância em entender os mecanismos de aumento de migração de
forma aprofundada.
Ressaltamos ainda que os efeitos causados por nanopartículas são célula-
dependente, como observado nas diferenças entre PNT1A e PC-3. Sabe-se que
uma linhagem celular pode conter subpopulações que respondem de forma
diferenciada, como no caso das células PC-3 que apresenta tanto fenótipos epiteliais
quanto mesenquimais na mesma população. Dessa forma, é necessário aprofundar
o estudo sobre qual mecanismo está sendo utilizado pelas SLN para aumentar a
migração dessas células; nossa hipótese é que esse aumento tem relação com a
ativação da via do TGF-β, visto que na próxima seção, veremos que inibidores da
via do TGF-β alteram a transfecção por SLN apenas em células PC-3. Porém antes
de prosseguir, decidimos investigar em detalhe para entender o papel de cada um
dos componentes da SLN têm efeito no perfil de migração das células de próstata.
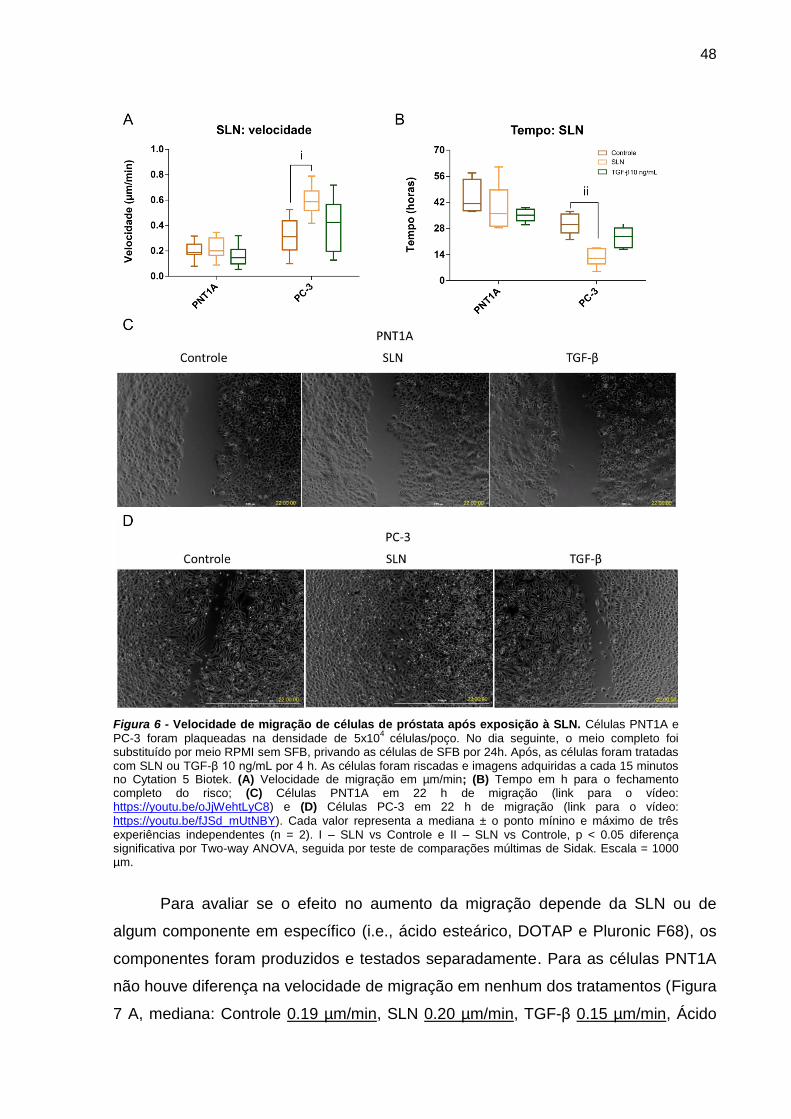
48
Figura 6 - Velocidade de migração de células de próstata após exposição à SLN. Células PNT1A e
PC-3 foram plaqueadas na densidade de 5x104
células/poço. No dia seguinte, o meio completo foi substituído por meio RPMI sem SFB, privando as células de SFB por 24h. Após, as células foram tratadas com SLN ou TGF-β 10 ng/mL por 4 h. As células foram riscadas e imagens adquiridas a cada 15 minutos no Cytation 5 Biotek. (A) Velocidade de migração em µm/min; (B) Tempo em h para o fechamento completo do risco; (C) Células PNT1A em 22 h de migração (link para o vídeo: https://youtu.be/oJjWehtLyC8) e (D) Células PC-3 em 22 h de migração (link para o vídeo:
https://youtu.be/fJSd_mUtNBY). Cada valor representa a mediana ± o ponto mínino e máximo de três experiências independentes (n = 2). I – SLN vs Controle e II – SLN vs Controle, p < 0.05 diferença significativa por Two-way ANOVA, seguida por teste de comparações múltimas de Sidak. Escala = 1000 µm.
Para avaliar se o efeito no aumento da migração depende da SLN ou de
algum componente em específico (i.e., ácido esteárico, DOTAP e Pluronic F68), os
componentes foram produzidos e testados separadamente. Para as células PNT1A
não houve diferença na velocidade de migração em nenhum dos tratamentos (Figura
7 A, mediana: Controle 0.19 µm/min, SLN 0.20 µm/min, TGF-β 0.15 µm/min, Ácido

49
esteárico 0.26 µm/min, DOTAP 0.22 µm/min e Pluronic F68 0.23 µm/min), resultado
que era esperado visto que a nanopartícula em si não foi capaz de induzir a
migração nessas células. O tempo que as células PNT1A levaram para migrar foi
similar entre todos os tratamentos, resultado congruente com a velocidade
encontrada (Figura 7 B).
Em células PC-3 somente a SLN foi capaz de induzir a migração enquanto
que os componentes da SLN em separado não aumentaram a migração das células
(Figura 7 A, mediana para Controle: 0.31 µm/min, SLN: 0.59 µm/min, TGF-β 0.42
µm/min, Ácido esteárico 0.26 µm/min, DOTAP 0.30 µm/min e Pluronic F68 0.40
µm/min). A SLN continuou com o tempo de fechamento do risco diminuído em
relação ao controle, e nos demais tratamentos não houve alteração significativa
(Figura 7 B). O interessante desse resultado é mostrar que a formação da
nanopartícula, uma estrutura organizada, é necessária para que o efeito aconteça, e
não dos componentes isolados – mostrando que a interação da SLN com a célula é
o que causa o efeito. Levando em consideração que a forma com que as
nanopartículas interagem com as células in vitro pode ser influenciada por diversos
fatores, até mesmo se a nanopartícula é adicionada concentrada ou previamente
diluída (Moore et al. 2019), esses resultados nos fazem pensar que diversos fatores
influenciam nessa interação célula-nanopartícula.
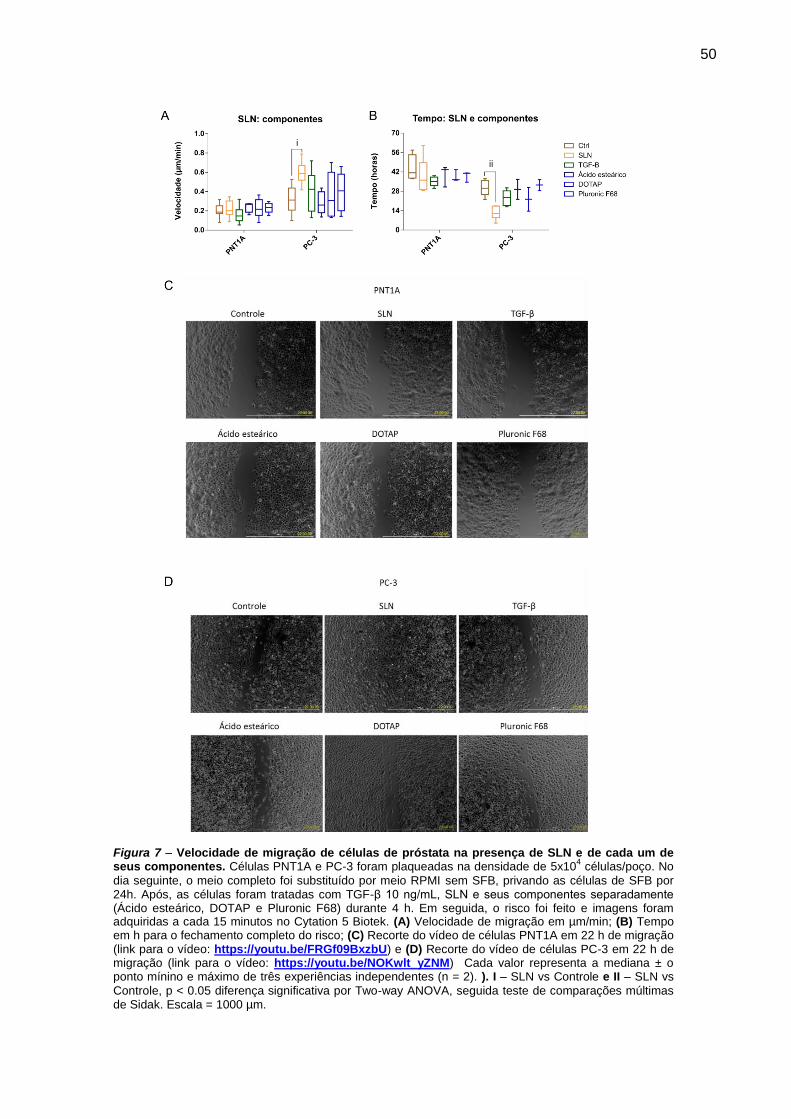
50
Figura 7 – Velocidade de migração de células de próstata na presença de SLN e de cada um de seus componentes. Células PNT1A e PC-3 foram plaqueadas na densidade de 5x10
4 células/poço. No
dia seguinte, o meio completo foi substituído por meio RPMI sem SFB, privando as células de SFB por 24h. Após, as células foram tratadas com TGF-β 10 ng/mL, SLN e seus componentes separadamente (Ácido esteárico, DOTAP e Pluronic F68) durante 4 h. Em seguida, o risco foi feito e imagens foram adquiridas a cada 15 minutos no Cytation 5 Biotek. (A) Velocidade de migração em µm/min; (B) Tempo em h para o fechamento completo do risco; (C) Recorte do vídeo de células PNT1A em 22 h de migração (link para o vídeo: https://youtu.be/FRGf09BxzbU) e (D) Recorte do vídeo de células PC-3 em 22 h de migração (link para o vídeo: https://youtu.be/NOKwlt_yZNM) Cada valor representa a mediana ± o ponto mínino e máximo de três experiências independentes (n = 2). ). I – SLN vs Controle e II – SLN vs
Controle, p < 0.05 diferença significativa por Two-way ANOVA, seguida teste de comparações múltimas de Sidak. Escala = 1000 µm.

51
O mecanismo que envolve o aumento da motilidade de células causada pela
SLN em diferentes concentrações é desconhecido. Moléculas bem descritas como o
TGF-β1 diminuem a porcentagem de células que migraram em direção ao risco: um
maior número de células da córnea migram conforme a concentração de TGF-β1
aumenta, um efeito dose-dependente (Wang et al. 2011). Sabendo que a SLN por si
só induz a migração de células PC-3, nos perguntamos se a concentração de SLN
utilizada até o momento (3.8 µg/mL, considerada SLN 1x) seria a única responsável
pelo aumento da velocidade de migração das células; para isso, fizemos uma curva
dose-resposta variando de 0.076 a 7.6 µg/mL (0.02x a 2x).
As concentrações iniciais de 0.02x (0.076 µg/mL) e 0.1x (0.38 µg/mL) não
aumentam a velocidade e seu valor assemelha-se ao controle não tratado (Figura 8
A). A partir da concentração 0.2x (0.76 µg/mL) a migração é induzida de forma
similar à SLN 1x. Quando as células são expostas à concentração de 2x, há uma
saturação na velocidade e esta não aumenta em relação à SLN 1x (Figura 8 A). A
curva dose-resposta (Figura 8 B) utilizando os valores de velocidade mostra que a
concentração de SLN necessária para induzir a migração das células (ponto em que
a curva começa a subir, IC50) é de 0.79 µg/mL, semelhante à concentração testada
de 0.2x.
O tempo de fechamento do poço se mostra condizente com os valores de
velocidade, já que concentrações menores (0x, 0.02x e 0.1x) levam mais tempo para
fechar o risco do que a SLN 2x (Figura 8 C, iv, v e vi p < 0.05). No nosso caso a SLN
aumenta a migração de células PC-3 mesmo diluída (0.2x), resultado que deve
aumentar o cuidado na administração in vivo, pois mesmo concentrações baixas de
nanopartícula podem ter efeitos inesperados.
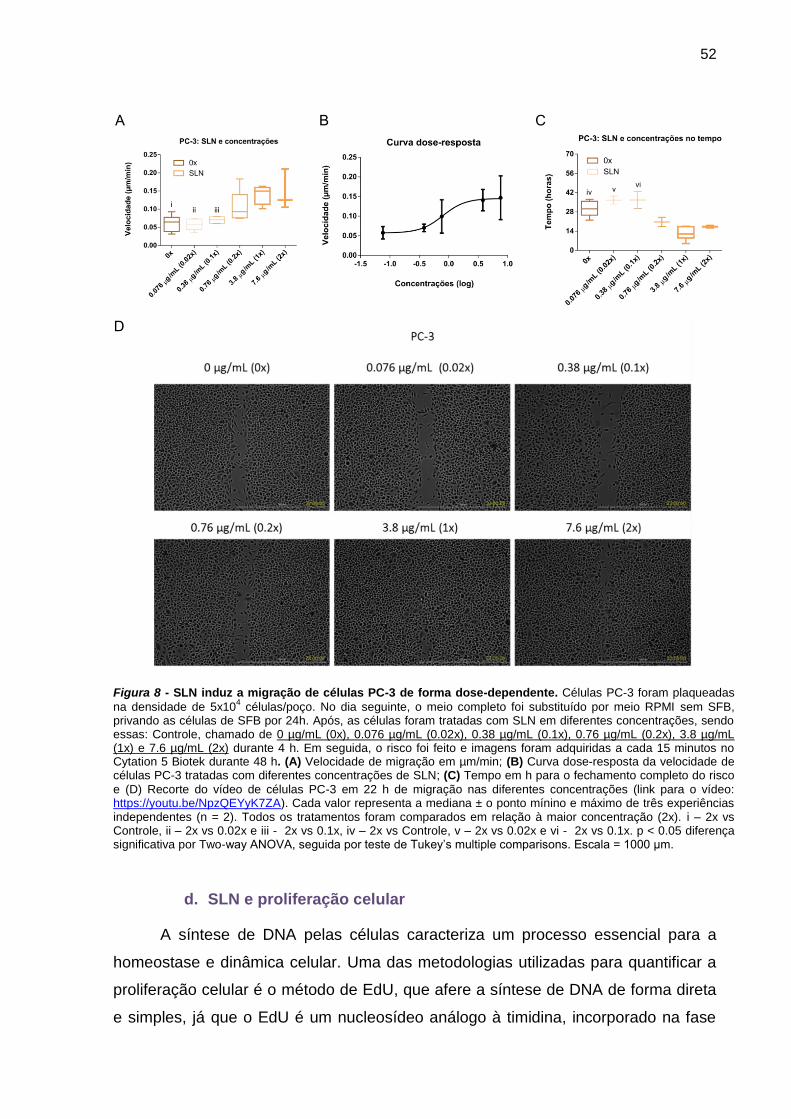
52
Figura 8 - SLN induz a migração de células PC-3 de forma dose-dependente. Células PC-3 foram plaqueadas
na densidade de 5x104 células/poço. No dia seguinte, o meio completo foi substituído por meio RPMI sem SFB,
privando as células de SFB por 24h. Após, as células foram tratadas com SLN em diferentes concentrações, sendo essas: Controle, chamado de 0 µg/mL (0x), 0.076 µg/mL (0.02x), 0.38 µg/mL (0.1x), 0.76 µg/mL (0.2x), 3.8 µg/mL (1x) e 7.6 µg/mL (2x) durante 4 h. Em seguida, o risco foi feito e imagens foram adquiridas a cada 15 minutos no Cytation 5 Biotek durante 48 h. (A) Velocidade de migração em µm/min; (B) Curva dose-resposta da velocidade de células PC-3 tratadas com diferentes concentrações de SLN; (C) Tempo em h para o fechamento completo do risco
e (D) Recorte do vídeo de células PC-3 em 22 h de migração nas diferentes concentrações (link para o vídeo: https://youtu.be/NpzQEYyK7ZA). Cada valor representa a mediana ± o ponto mínino e máximo de três experiências independentes (n = 2). Todos os tratamentos foram comparados em relação à maior concentração (2x). i – 2x vs Controle, ii – 2x vs 0.02x e iii - 2x vs 0.1x, iv – 2x vs Controle, v – 2x vs 0.02x e vi - 2x vs 0.1x. p < 0.05 diferença significativa por Two-way ANOVA, seguida por teste de Tukey‟s multiple comparisons. Escala = 1000 µm.
d. SLN e proliferação celular
A síntese de DNA pelas células caracteriza um processo essencial para a
homeostase e dinâmica celular. Uma das metodologias utilizadas para quantificar a
proliferação celular é o método de EdU, que afere a síntese de DNA de forma direta
e simples, já que o EdU é um nucleosídeo análogo à timidina, incorporado na fase

53
de síntese do ciclo celular, e é detectado por uma reação click it entre uma azida
(contida no fluorófuro) e um alquino (contido no EdU) (Flomerfelt e Gress 2016).
Para avaliar se o aumento da velocidade de migração em células PC3 causado
pelas SLNs estava relacionado com um aumento nas taxas de proliferação,
realizamos o teste de proliferação celular com EdU. Além disso, checar os níveis de
proliferação auxilia a determinar qual a contribuição da proliferação na migração das
células, pois é esperado que mesmo em privação de soro fetal bovino, as células
mantenham níveis basais de proliferação.
Realizamos o teste de proliferação em células PNT1A e PC-3 privadas de
SFB, nos tempos de 5h, 24h e 48h após o tratamento com SLN. As células foram
marcadas com DAPI, corante que tem atração por DNA e por isso cora todas as
células, e as células que proliferaram foram marcadas com EdU. De um modo geral,
notamos que células PNT1A não sofreram alteração significativa na taxa de
proliferação quando tratadas com SLN, em todos os tempos (Figura 9 A, medianas
em 5h: Controle 34.3% e SLN 32.9%, 24h: Controle 62.6% e SLN 59.7%, 48h:
Controle 76.8% e SLN 60.2%, respectivamente). Porém, ao longo do tempo, a
privação de SFB não é capaz de conter a proliferação, que chega a 76.8% em 48h.
Esses resultados vão de acordo com a literatura, pois apesar de células PNT1A
possuírem origem no tecido epitelial de próstata, estas têm alta taxa de proliferação,
cerca de 100% após 24h de privação de SFB (Domińska et al. 2016). Através das
imagens representativas (Figura 9 B) também é possível perceber um aumento na
quantidade de núcleos em vermelho conforme o aumento do tempo, em todos os
tratamentos. Um dos fatores que poderia explicar a alta taxa de proliferação em
células PNT1A pode ser explicado pelo processo de imortalização dessa linhagem
celular, já que a imortalização por si só aumenta o potencial proliferativo dessas
células (Mondello e Chiodi 2013). Outro ponto a se considerar é a forma de
imortalização, através de transformação com o vírus SV40, que pode alterar genes
envolvidos em senescência e apoptose celular (Liu et al. 2009).
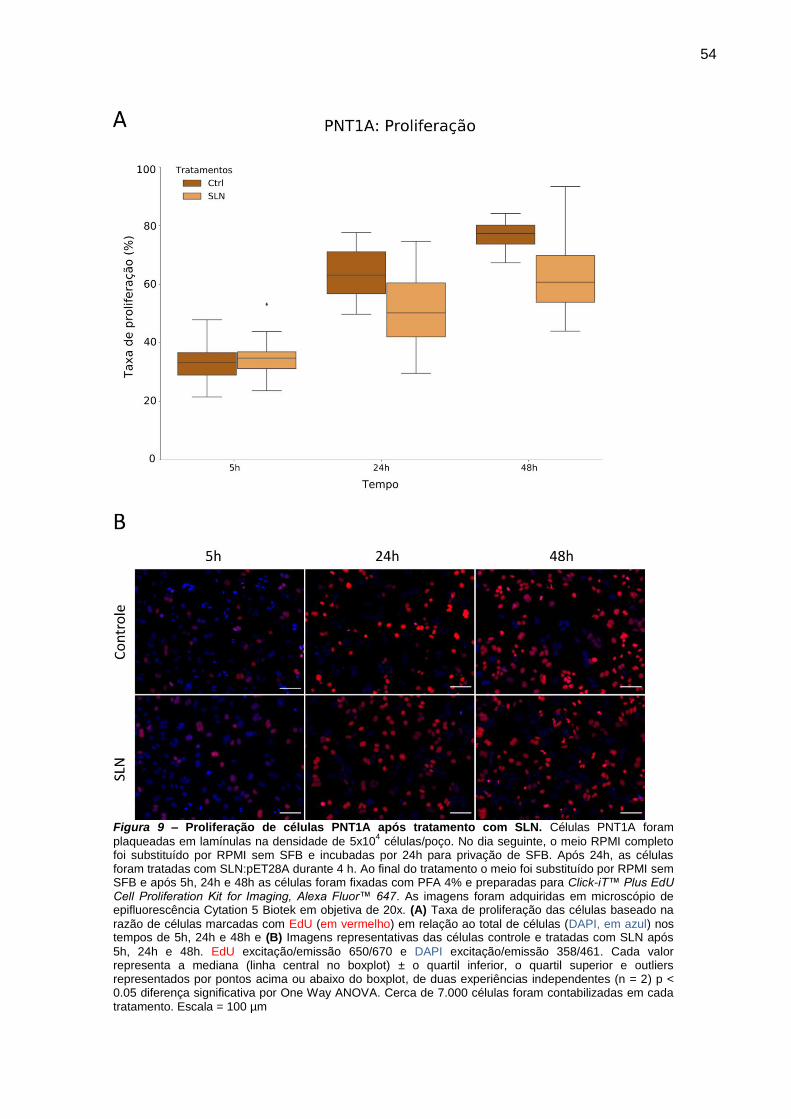
54
Figura 9 – Proliferação de células PNT1A após tratamento com SLN. Células PNT1A foram
plaqueadas em lamínulas na densidade de 5x104 células/poço. No dia seguinte, o meio RPMI completo
foi substituído por RPMI sem SFB e incubadas por 24h para privação de SFB. Após 24h, as células foram tratadas com SLN:pET28A durante 4 h. Ao final do tratamento o meio foi substituído por RPMI sem SFB e após 5h, 24h e 48h as células foram fixadas com PFA 4% e preparadas para Click-iT™ Plus EdU Cell Proliferation Kit for Imaging, Alexa Fluor™ 647. As imagens foram adquiridas em microscópio de epifluorescência Cytation 5 Biotek em objetiva de 20x. (A) Taxa de proliferação das células baseado na
razão de células marcadas com EdU (em vermelho) em relação ao total de células (DAPI, em azul) nos tempos de 5h, 24h e 48h e (B) Imagens representativas das células controle e tratadas com SLN após
5h, 24h e 48h. EdU excitação/emissão 650/670 e DAPI excitação/emissão 358/461. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos acima ou abaixo do boxplot, de duas experiências independentes (n = 2) p < 0.05 diferença significativa por One Way ANOVA. Cerca de 7.000 células foram contabilizadas em cada tratamento. Escala = 100 µm

55
No caso das células PC-3, percebemos que a taxa de proliferação se mantém
estável ao longo dos tempos e que não há diferença significativa entre o controle e
tratado com SLN (Figura 10, mediana em 5h: Controle 13.7% e SLN 15.4%, 24h:
Controle 25.4% e SLN 21.1%, 48h: Controle 24.1% e SLN 19.4%, respectivamente).
Mesmo após 48h, a taxa de proliferação é mantida a níveis basais, indicando que a
privação de SFB auxilia na diminuição da proliferação em células PC-3, pois mesmo
na presença de SFB essas células proliferaram 30% em 24h (Lovitt et al. 2016;
Wang et al. 2019). Através de imagens representativas (Figura 10 B) percebemos
um aumento pequeno nas células marcadas em vermelho, que demonstra não ser
significativo. Em tumores, células da borda costumam migrar mais e proliferar menos
(Gallaher, Brown, e Anderson 2019), informação que condiz com nossos resultados
de aumento de migração em células PC-3.
Esses resultados mostram que a privação de SFB é um método robusto de
manter a proliferação de células PC-3 em níveis basais até 48 h de experimento,
enquanto que para células PNT1A, 24 h é o tempo limite para o controle da
proliferação, que apesar de estar em 30%, representa ainda uma taxa aceitável de
proliferação. Como o aumento da proliferação por SLN foi descartado, para explicar
o aumento na taxa de migração, seguimos os experimentos no sentido de investigar
os mecanismos por trás do aumento da migração das células e sua relação com a
via do TGF-β.
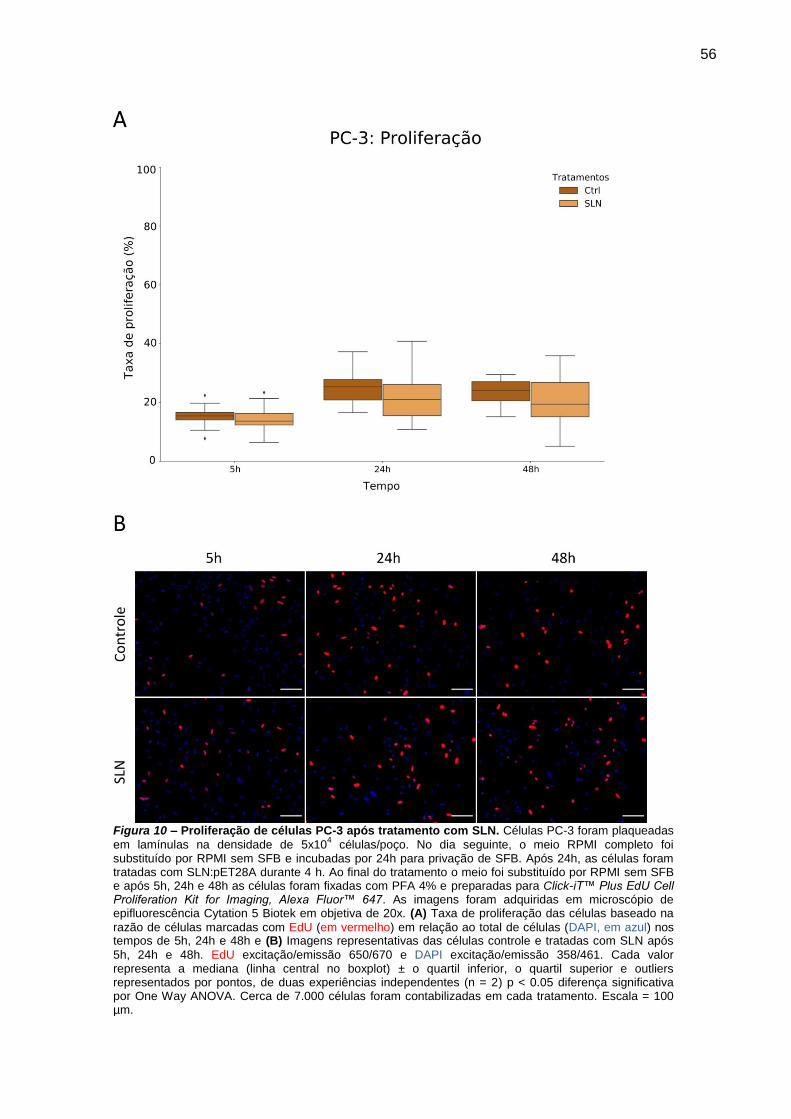
56
Figura 10 – Proliferação de células PC-3 após tratamento com SLN. Células PC-3 foram plaqueadas
em lamínulas na densidade de 5x104 células/poço. No dia seguinte, o meio RPMI completo foi
substituído por RPMI sem SFB e incubadas por 24h para privação de SFB. Após 24h, as células foram tratadas com SLN:pET28A durante 4 h. Ao final do tratamento o meio foi substituído por RPMI sem SFB e após 5h, 24h e 48h as células foram fixadas com PFA 4% e preparadas para Click-iT™ Plus EdU Cell Proliferation Kit for Imaging, Alexa Fluor™ 647. As imagens foram adquiridas em microscópio de epifluorescência Cytation 5 Biotek em objetiva de 20x. (A) Taxa de proliferação das células baseado na
razão de células marcadas com EdU (em vermelho) em relação ao total de células (DAPI, em azul) nos tempos de 5h, 24h e 48h e (B) Imagens representativas das células controle e tratadas com SLN após
5h, 24h e 48h. EdU excitação/emissão 650/670 e DAPI excitação/emissão 358/461. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de duas experiências independentes (n = 2) p < 0.05 diferença significativa por One Way ANOVA. Cerca de 7.000 células foram contabilizadas em cada tratamento. Escala = 100 µm.

57
e. SLN e a via do TGF-β
Para avaliar a influência da via do TGF-β na interação das células com a SLN,
utilizamos inicialmente dois inibidores da via, LY2109761 e SIS3 HCl. O LY2109761
é responsável por inibir os receptores da via (TβRI e TβRII) enquanto que o SIS3
HCl inibe a fosforilação e translocação da Smad3. Determinamos a melhor
concentração de cada inibidor, que mantivesse a viabilidade das células em pelo
menos 80% (Figura 11).
Utilizamos o homodímero de etídio (EthD-1), que é permeável à membrana
celular danificada e é excluído por membranas plasmáticas de células viáveis. Ao
entrar na célula danificada, sua fluorescência aumenta numa escala de 40x na
ligação ao DNA, dessa forma emitindo fluorescência vermelha apenas nas células
mortas, Excitação/Emissão: 495/635 nm (Li et al. 2013). Portanto, células que
apresentaram marcação para EthD-1 foram consideradas não viáveis; para
determinar o número total de células, foi utilizado o marcador de núcleo Hoechst
33342 (Figura 11).
O inibidor LY210976 apresentou o mesmo comportamento nas duas
linhagens celulares ao não apresentar toxicidade nas concentrações testadas
(Figura 11, A e B); todas as concentrações apresentaram diferença em relação ao
controle positivo de morte (Mortas). Com base nas concentrações comumente
utilizadas na literatura, entre 0,1 até 20 µM (Wan et al. 2012), foi escolhida a
concentração de 5 µM para prosseguir com os ensaios celulares.
O inibidior de Smad3, SIS3 HCl foi mais tóxico do que LY2109761 em ambas
as linhagens (Figura 11, C e D). Nas células PNT1A, de 0.375 µM a 12 µM houve
diferença em relação ao controle, enquanto que nas células PC-3 apenas até 1.5 µM
houve diferença em relação ao controle de morte; dessa forma, a concentração de
1.5 µM foi escolhida para continuar com os demais experimentos.
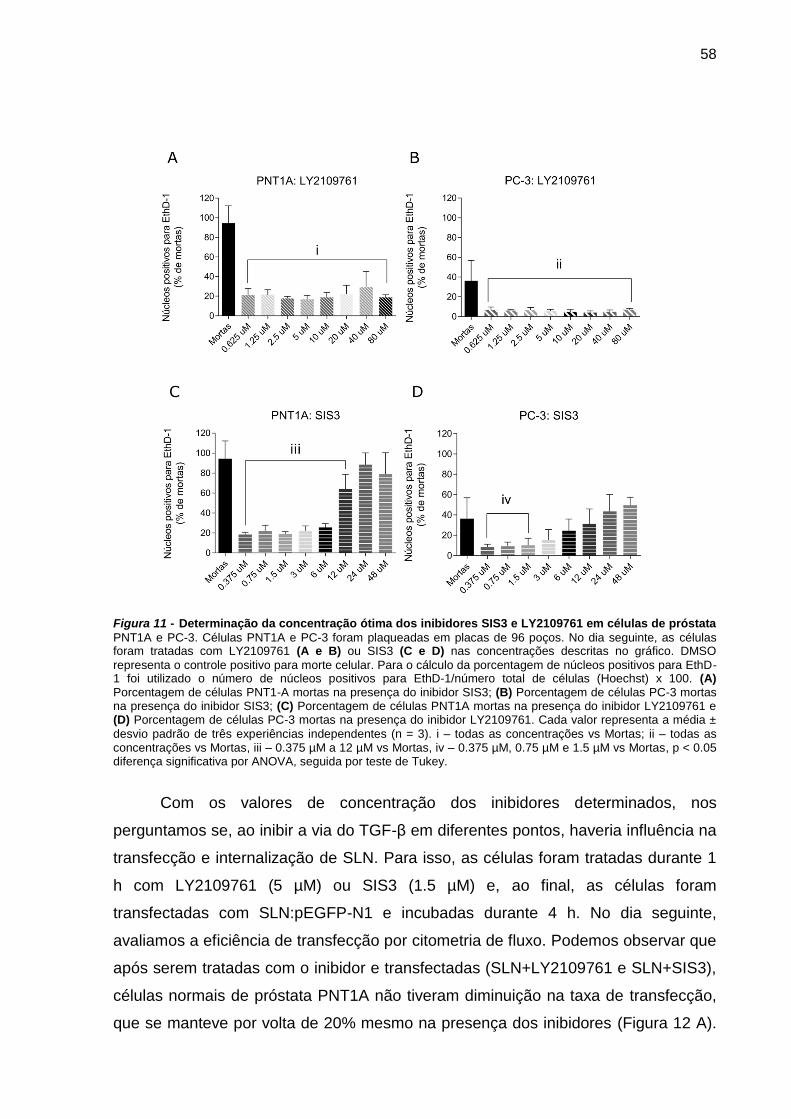
58
Figura 11 - Determinação da concentração ótima dos inibidores SIS3 e LY2109761 em células de próstata
PNT1A e PC-3. Células PNT1A e PC-3 foram plaqueadas em placas de 96 poços. No dia seguinte, as células foram tratadas com LY2109761 (A e B) ou SIS3 (C e D) nas concentrações descritas no gráfico. DMSO
representa o controle positivo para morte celular. Para o cálculo da porcentagem de núcleos positivos para EthD-1 foi utilizado o número de núcleos positivos para EthD-1/número total de células (Hoechst) x 100. (A) Porcentagem de células PNT1-A mortas na presença do inibidor SIS3; (B) Porcentagem de células PC-3 mortas na presença do inibidor SIS3; (C) Porcentagem de células PNT1A mortas na presença do inibidor LY2109761 e (D) Porcentagem de células PC-3 mortas na presença do inibidor LY2109761. Cada valor representa a média ±
desvio padrão de três experiências independentes (n = 3). i – todas as concentrações vs Mortas; ii – todas as concentrações vs Mortas, iii – 0.375 µM a 12 µM vs Mortas, iv – 0.375 µM, 0.75 µM e 1.5 µM vs Mortas, p < 0.05 diferença significativa por ANOVA, seguida por teste de Tukey.
Com os valores de concentração dos inibidores determinados, nos
perguntamos se, ao inibir a via do TGF-β em diferentes pontos, haveria influência na
transfecção e internalização de SLN. Para isso, as células foram tratadas durante 1
h com LY2109761 (5 µM) ou SIS3 (1.5 µM) e, ao final, as células foram
transfectadas com SLN:pEGFP-N1 e incubadas durante 4 h. No dia seguinte,
avaliamos a eficiência de transfecção por citometria de fluxo. Podemos observar que
após serem tratadas com o inibidor e transfectadas (SLN+LY2109761 e SLN+SIS3),
células normais de próstata PNT1A não tiveram diminuição na taxa de transfecção,
que se manteve por volta de 20% mesmo na presença dos inibidores (Figura 12 A).

59
A intensidade de fluorescência não sofreu alterações e as células expressaram GFP
em intensidade similar à SLN isolada (Figura 12 B).
Entretanto, para as células tumorais PC-3, a expressão de GFP nos grupos
SLN+LY2109761 e SLN+SIS3 foi menor do que as células que foram somente
tratadas com SLN:pEGFP, diminuindo cerca de 3x (Figura 12, i p<0.05). Os
inibidores diminuíram a taxa de transfecção em PC-3 a níveis de transfecção
similares ao da célula PNT1A. Interessante notar que mesmo com a diminuição nos
níveis de transfecção, não foram observadas diferenças na intensidade de
fluorescência (Figura 12 B), mostrando inclusive tendência ao aumento na
intensidade em presença dos inibidores. Apesar de uma porcentagem menor de
células transfectarem na presença dos inibidores, tais células expressam GFP em
níveis maiores do que das células transfectadas apenas com SLN:pEGFP. A
primeira hipótese é de que a inibição da via do TGF-β esteja interferindo na via de
internalização da nanopartícula, que na situação de inibição, pode entrar por uma via
mais produtiva e que leva à maior expressão de GFP, ou seja, um efeito upstream,
no início da via de entrada. A segunda hipótese é que inibir a via do TGF-β leva a
nanopartícula a seguir por outro tráfego intracelular; considerando que os receptores
do TGF-β são fosforilados em endossomos precoces, inibir a sua fosforilação pode
interferir no tráfego intracelular da nanopartícula; que seria um efeito downstream,
ou seja, após a entrada da nanopartícula na célula.
A diminuição da expressão de GFP após o tratamento com o inibidor é uma
indicação de que a via do TGF-β é importante para a entrega e expressão do
plasmídeo nas células, mas ainda não temos detalhes precisos de como isso ocorre.
Outros trabalhos mostram a relação entre a inibição por pequenas moléculas e a
diminuição da expressão de GFP (Liu, Krishnan, e Phua 2017), inclusive essa
diminuição de expressão pode ser utilizada para determinar a via de endocitose
responsável pela entrada da nanopartícula na célula (Radaic e Jesus 2018; Ruiz de
Garibay et al. 2013). Rehman, Hoekstra, e Zuhorn (2011) mostraram que ao inibir a
proteína quinase A (PKA), a transfecção por PEI em células HeLa aumentou em 2,5
vezes, sem afetar a internalização da nanopartícula. Esse aumento da transfecção
foi devido à perturbação do trafégo intracelular na endocitose mediada por clatrina, o
que levou ao impedimento da chegada do PEI ao endossomo tardio/lisossomo.
Dessa forma, o estudo de liberação do material genético pela nanopartícula e quais
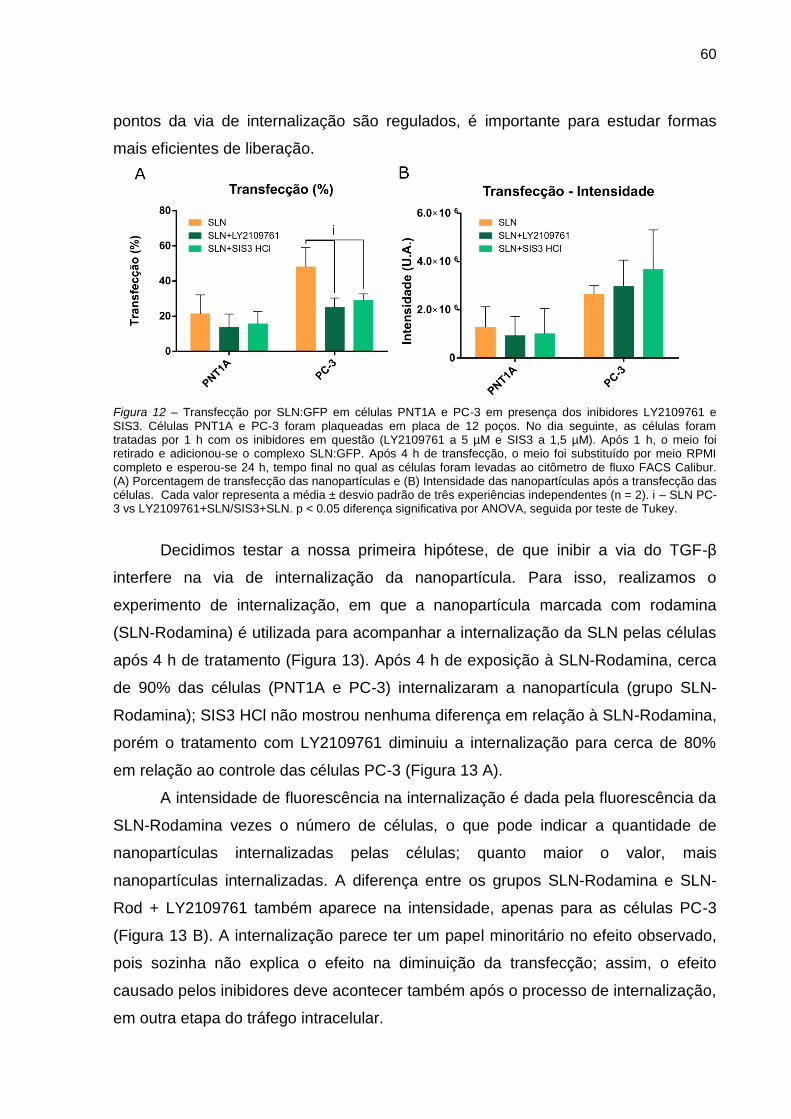
60
pontos da via de internalização são regulados, é importante para estudar formas
mais eficientes de liberação.
Figura 12 – Transfecção por SLN:GFP em células PNT1A e PC-3 em presença dos inibidores LY2109761 e SIS3. Células PNT1A e PC-3 foram plaqueadas em placa de 12 poços. No dia seguinte, as células foram tratadas por 1 h com os inibidores em questão (LY2109761 a 5 µM e SIS3 a 1,5 µM). Após 1 h, o meio foi retirado e adicionou-se o complexo SLN:GFP. Após 4 h de transfecção, o meio foi substituído por meio RPMI completo e esperou-se 24 h, tempo final no qual as células foram levadas ao citômetro de fluxo FACS Calibur. (A) Porcentagem de transfecção das nanopartículas e (B) Intensidade das nanopartículas após a transfecção das células. Cada valor representa a média ± desvio padrão de três experiências independentes (n = 2). i – SLN PC-3 vs LY2109761+SLN/SIS3+SLN. p < 0.05 diferença significativa por ANOVA, seguida por teste de Tukey.
Decidimos testar a nossa primeira hipótese, de que inibir a via do TGF-β
interfere na via de internalização da nanopartícula. Para isso, realizamos o
experimento de internalização, em que a nanopartícula marcada com rodamina
(SLN-Rodamina) é utilizada para acompanhar a internalização da SLN pelas células
após 4 h de tratamento (Figura 13). Após 4 h de exposição à SLN-Rodamina, cerca
de 90% das células (PNT1A e PC-3) internalizaram a nanopartícula (grupo SLN-
Rodamina); SIS3 HCl não mostrou nenhuma diferença em relação à SLN-Rodamina,
porém o tratamento com LY2109761 diminuiu a internalização para cerca de 80%
em relação ao controle das células PC-3 (Figura 13 A).
A intensidade de fluorescência na internalização é dada pela fluorescência da
SLN-Rodamina vezes o número de células, o que pode indicar a quantidade de
nanopartículas internalizadas pelas células; quanto maior o valor, mais
nanopartículas internalizadas. A diferença entre os grupos SLN-Rodamina e SLN-
Rod + LY2109761 também aparece na intensidade, apenas para as células PC-3
(Figura 13 B). A internalização parece ter um papel minoritário no efeito observado,
pois sozinha não explica o efeito na diminuição da transfecção; assim, o efeito
causado pelos inibidores deve acontecer também após o processo de internalização,
em outra etapa do tráfego intracelular.
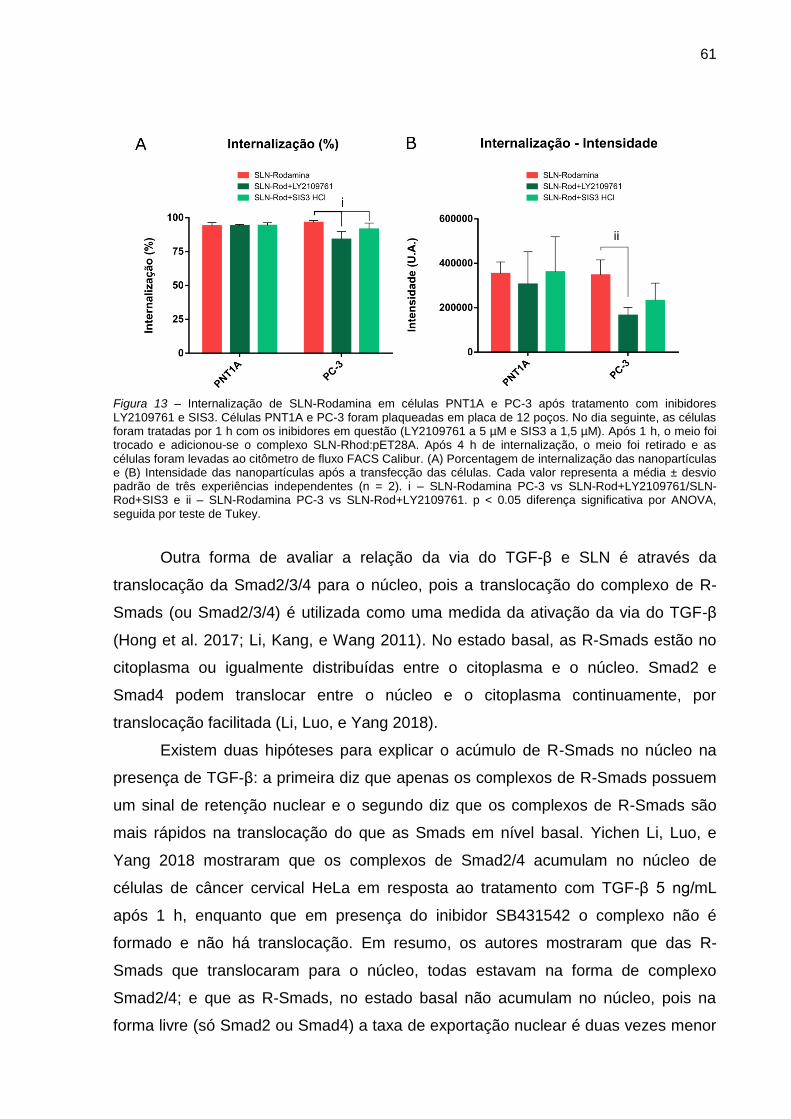
61
Figura 13 – Internalização de SLN-Rodamina em células PNT1A e PC-3 após tratamento com inibidores LY2109761 e SIS3. Células PNT1A e PC-3 foram plaqueadas em placa de 12 poços. No dia seguinte, as células foram tratadas por 1 h com os inibidores em questão (LY2109761 a 5 µM e SIS3 a 1,5 µM). Após 1 h, o meio foi trocado e adicionou-se o complexo SLN-Rhod:pET28A. Após 4 h de internalização, o meio foi retirado e as células foram levadas ao citômetro de fluxo FACS Calibur. (A) Porcentagem de internalização das nanopartículas e (B) Intensidade das nanopartículas após a transfecção das células. Cada valor representa a média ± desvio padrão de três experiências independentes (n = 2). i – SLN-Rodamina PC-3 vs SLN-Rod+LY2109761/SLN-Rod+SIS3 e ii – SLN-Rodamina PC-3 vs SLN-Rod+LY2109761. p < 0.05 diferença significativa por ANOVA, seguida por teste de Tukey.
Outra forma de avaliar a relação da via do TGF-β e SLN é através da
translocação da Smad2/3/4 para o núcleo, pois a translocação do complexo de R-
Smads (ou Smad2/3/4) é utilizada como uma medida da ativação da via do TGF-β
(Hong et al. 2017; Li, Kang, e Wang 2011). No estado basal, as R-Smads estão no
citoplasma ou igualmente distribuídas entre o citoplasma e o núcleo. Smad2 e
Smad4 podem translocar entre o núcleo e o citoplasma continuamente, por
translocação facilitada (Li, Luo, e Yang 2018).
Existem duas hipóteses para explicar o acúmulo de R-Smads no núcleo na
presença de TGF-β: a primeira diz que apenas os complexos de R-Smads possuem
um sinal de retenção nuclear e o segundo diz que os complexos de R-Smads são
mais rápidos na translocação do que as Smads em nível basal. Yichen Li, Luo, e
Yang 2018 mostraram que os complexos de Smad2/4 acumulam no núcleo de
células de câncer cervical HeLa em resposta ao tratamento com TGF-β 5 ng/mL
após 1 h, enquanto que em presença do inibidor SB431542 o complexo não é
formado e não há translocação. Em resumo, os autores mostraram que das R-
Smads que translocaram para o núcleo, todas estavam na forma de complexo
Smad2/4; e que as R-Smads, no estado basal não acumulam no núcleo, pois na
forma livre (só Smad2 ou Smad4) a taxa de exportação nuclear é duas vezes menor

62
do que na presença do TGF-β (Li et al. 2018). Como as proteínas regulam genes
relacionados à via do TGF-β, sua translocação para o núcleo indica que houve
sinalização para que Smad2/3 se concentre no núcleo da célula e realize sua função
como fator de transcrição.
Para avaliar se os inibidores interferem na translocação de R-Smads e validá-
los quanto à capacidade de inibição, avaliamos a translocação de Smad2 após
tratamento com os inibidores LY2109761 e SIS3 HCl individualmente, e em um
segundo momento, como as células pré-tratadas com os inibidores respondem à
exposição ao TGF-β (Figura 14 e Figura 15), que a 10 ng/mL foi utilizado como
controle positivo para translocação nuclear de Smad2.
As células PNT1A respondem ao estímulo por TGF-β após 1h de exposição,
pois observamos a translocação de Smad2 para o núcleo de forma intensa em
praticamente todas as células da população (Figura 14 A e B, 1h TGF-β). Após 4h, a
Smad2 começa a retornar para o citoplasma, mas grande parte ainda é encontrada
no núcleo. O inibidor SIS3 HCl parece não ativar a translocação no mesmo nível do
TGF-β ao observar as imagens (Figura 14 B) apesar do valor da intensidade de
Smad2 no núcleo similar ao TGF-β em ambos os tempos (Figura 14 A). Ao adicionar
o TGF-β nas células pré-tratadas com SIS3 HCl, a translocação de Smad2 ocorre
normalmente em ambos os tempos (Figura 14 B). O SIS3 HCl é um inibidor
específico para Smad3 e podemos ver que nessas células, este não tem efeito na
translocação de Smad2.
O tratamento com o inibidor LY2109761 manteve os níveis de Smad2 nuclear
baixos em todos os tempos, e mesmo a exposição ao TGF-β não ativou a
translocação de Smad2 para o núcleo. Concluimos que em células PNT1A o
tratamento com LY2109761 é suficiente para inibir a translocação de Smad2,
mesmo este sendo um inibidor dos receptores do TGF-β tipo I e tipo II (Figura 14 B).
Sabe-se que o inibidor LY2109761 inibe em 92% a fosforilação de Smad2 em
células hepáticas (Luangmonkong et al. 2018), porém geralmente não é reportado o
nível de translocação desse fator. Já que a Smad2 fica ativa quando é fosforilada, é
esperado que ao inibir sua fosforilação, haja diminuição nos níveis de translocação.
Para a interação e mecanismos de inibição de ambos os inibidores em células
PNT1A não há relatos na literatura atualmente.
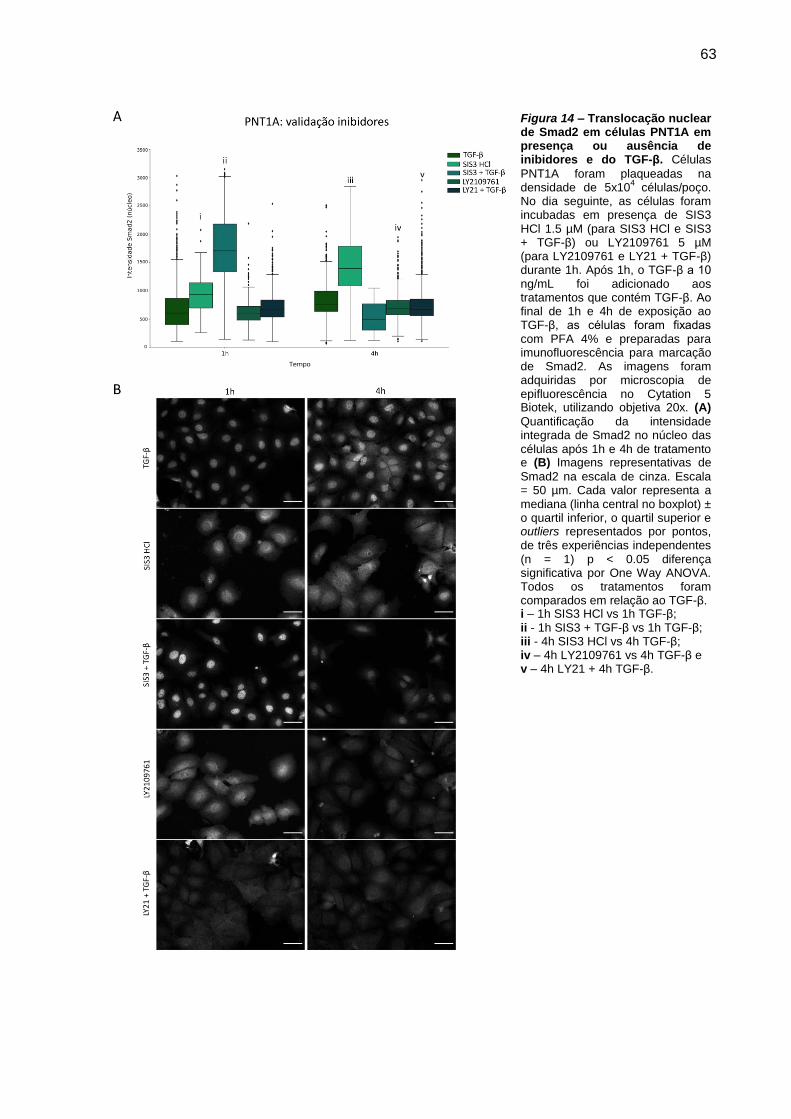
63
Figura 14 – Translocação nuclear de Smad2 em células PNT1A em presença ou ausência de inibidores e do TGF-β. Células
PNT1A foram plaqueadas na densidade de 5x10
4 células/poço.
No dia seguinte, as células foram incubadas em presença de SIS3 HCl 1.5 µM (para SIS3 HCl e SIS3 + TGF-β) ou LY2109761 5 µM (para LY2109761 e LY21 + TGF-β) durante 1h. Após 1h, o TGF-β a 10 ng/mL foi adicionado aos tratamentos que contém TGF-β. Ao final de 1h e 4h de exposição ao TGF-β, as células foram fixadas com PFA 4% e preparadas para imunofluorescência para marcação de Smad2. As imagens foram adquiridas por microscopia de epifluorescência no Cytation 5 Biotek, utilizando objetiva 20x. (A)
Quantificação da intensidade integrada de Smad2 no núcleo das células após 1h e 4h de tratamento e (B) Imagens representativas de
Smad2 na escala de cinza. Escala = 50 µm. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 1) p < 0.05 diferença significativa por One Way ANOVA. Todos os tratamentos foram comparados em relação ao TGF-β. i – 1h SIS3 HCl vs 1h TGF-β; ii - 1h SIS3 + TGF-β vs 1h TGF-β; iii - 4h SIS3 HCl vs 4h TGF-β; iv – 4h LY2109761 vs 4h TGF-β e v – 4h LY21 + 4h TGF-β.

64
O tratamento com TGF-β em células PC-3 leva à translocação de Smad2 em
1h de tratamento, sendo que em 4h ocorre o retorno da Smad2 para o citoplasma
(Figura 17 A e B). Assim como nas células PNT1A, o tratamento com SIS3 HCl
previamente à adição de TGF-β não inibe totalmente a translocação de Smad2 para
o núcleo e após 1h, é possível observá-la no núcleo de algumas células da
população (Figura 17 B), sendo que após 4h há o retorno da Smad2 para o
citoplasma. O inibidor LY2109761 inibiu a translocação de Smad2 na presença do
TGF-β em ambos os tempos. Assim como para células PNT1A, a interação e
mecanismos de inibição de ambos os inibidores em células PC-3 não é descrito na
literatura.
Portanto, concluímos que o inibidor de TβRI e TβRII, LY2109761, mantém a
inibição da via do TGF-β mesmo na presença deste fator de crescimento, em ambas
as linhagens celulares. Por outro lado, o inibidor SIS3 HCl não foi capaz de inibir a
translocação de Smad2 em ambas as células, o que já seria esperado pois o inibidor
tem ação específica na Smad3.
Para avaliar se nanopartículas lipídicas sólidas SLN têm efeito na
translocação de Smad2, utilizamos como controle positivo de translocação o
tratamento com TGF-β 10 ng/mL. Utilizamos então o anticorpo para Smad2 total,
que representa o controle positivo de ativação da via do TGF-β como demonstrado
na seção anterior, em que os inibidores foram validados para translocação de
Smad2.
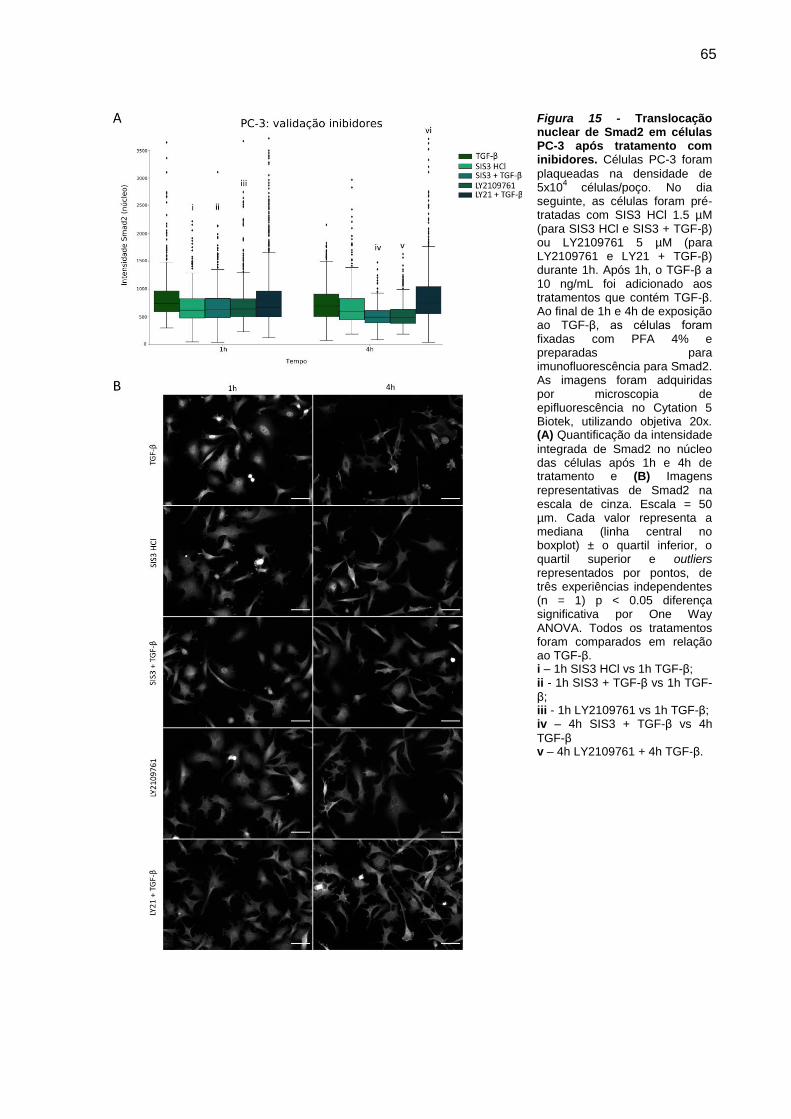
65
Figura 15 - Translocação nuclear de Smad2 em células PC-3 após tratamento com inibidores. Células PC-3 foram
plaqueadas na densidade de 5x10
4 células/poço. No dia
seguinte, as células foram pré-tratadas com SIS3 HCl 1.5 µM (para SIS3 HCl e SIS3 + TGF-β) ou LY2109761 5 µM (para LY2109761 e LY21 + TGF-β) durante 1h. Após 1h, o TGF-β a 10 ng/mL foi adicionado aos tratamentos que contém TGF-β. Ao final de 1h e 4h de exposição ao TGF-β, as células foram fixadas com PFA 4% e preparadas para imunofluorescência para Smad2. As imagens foram adquiridas por microscopia de epifluorescência no Cytation 5 Biotek, utilizando objetiva 20x. (A) Quantificação da intensidade
integrada de Smad2 no núcleo das células após 1h e 4h de tratamento e (B) Imagens
representativas de Smad2 na escala de cinza. Escala = 50 µm. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 1) p < 0.05 diferença significativa por One Way ANOVA. Todos os tratamentos foram comparados em relação ao TGF-β. i – 1h SIS3 HCl vs 1h TGF-β; ii - 1h SIS3 + TGF-β vs 1h TGF-
β; iii - 1h LY2109761 vs 1h TGF-β; iv – 4h SIS3 + TGF-β vs 4h
TGF-β v – 4h LY2109761 + 4h TGF-β.

66
As imagens foram adquiridas em microscópio de fluorescência (Figura 16 e
Figura 17) e além da Smad2 marcada em verde, o núcleo foi marcado com DAPI,
passo necessário para a formação da máscara utilizada na análise das imagens. Na
análise feita através do CellProfiler (Carpenter et al. 2006), o núcleo é utilizado para
formar a primeira máscara, que identifica todas as células; a partir da imagem da
Smad2, o programa encontra as bordas da célula e delimita o perímetro celular. A
medida de intensidade integrada da Smad2 no objeto primário (núcleo) foi utilizada
para avaliar a translocação de Smad2.
As imagens representativas da Smad2 nos diferentes tempos mostram a
distribuição da Smad2 na presença da SLN, e na ausência (controle) ou presença do
TGF-β (Figura 16 e Figura 17). Podemos observar que nos controles de ambas as
linhagens, a Smad2 está igualmente distribuída entre o citoplasma e o núcleo, um
padrão uniforme na população de células.
No caso da célula PNT1A (Figura 16), o tratamento com TGF-β induziu a
translocação de Smad2 para o núcleo e essa translocação perdurou até 4 h de
tratamento; sendo que no tempo de 4 h, é possível observar que o TGF-β ainda se
concentra no núcleo, porém o sinal da proteína no citoplasma começa a aumentar,
indicando a volta da Smad2 para o citoplasma (Figura 16 A, diferenças significativas
i, ii e iii, p < 0.05 e imagens em B). Não houve diferença entre os controles e o
tratamento com SLN, indicando que nos tempos avaliados a SLN não foi capaz de
induzir a translocação de Smad2 em células PNT1A.
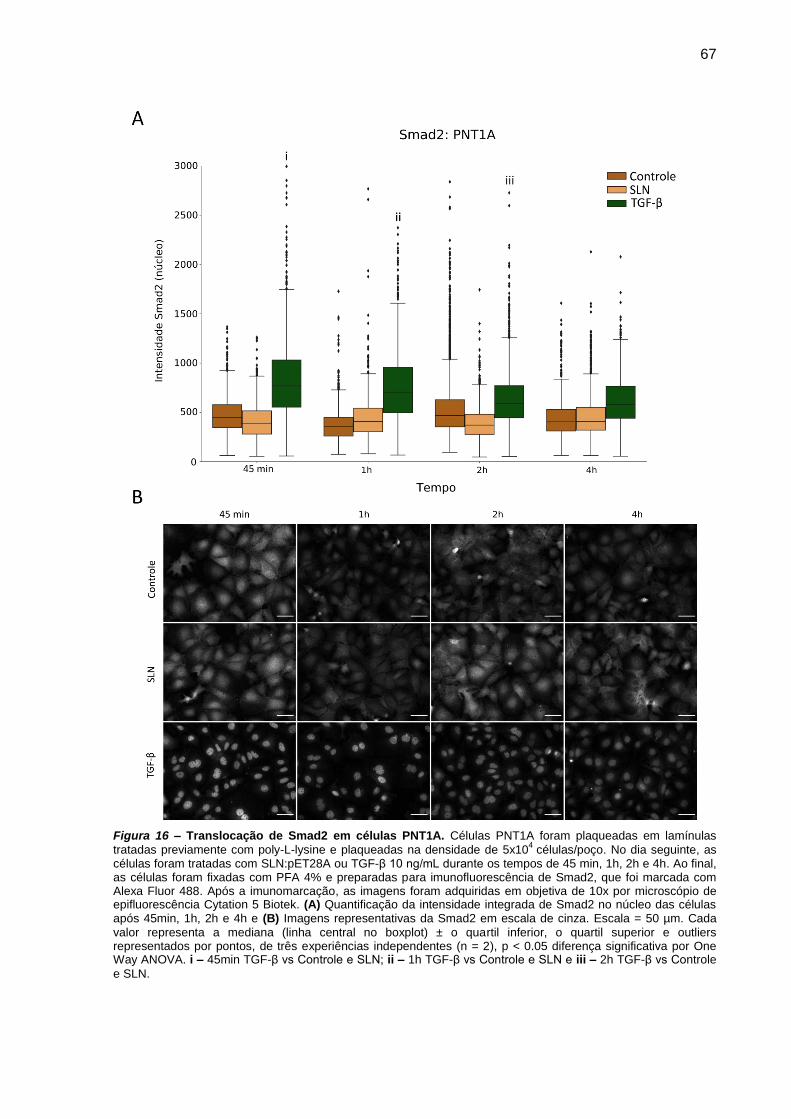
67
Figura 16 – Translocação de Smad2 em células PNT1A. Células PNT1A foram plaqueadas em lamínulas
tratadas previamente com poly-L-lysine e plaqueadas na densidade de 5x104
células/poço. No dia seguinte, as células foram tratadas com SLN:pET28A ou TGF-β 10 ng/mL durante os tempos de 45 min, 1h, 2h e 4h. Ao final, as células foram fixadas com PFA 4% e preparadas para imunofluorescência de Smad2, que foi marcada com Alexa Fluor 488. Após a imunomarcação, as imagens foram adquiridas em objetiva de 10x por microscópio de epifluorescência Cytation 5 Biotek. (A) Quantificação da intensidade integrada de Smad2 no núcleo das células após 45min, 1h, 2h e 4h e (B) Imagens representativas da Smad2 em escala de cinza. Escala = 50 µm. Cada
valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 2), p < 0.05 diferença significativa por One Way ANOVA. i – 45min TGF-β vs Controle e SLN; ii – 1h TGF-β vs Controle e SLN e iii – 2h TGF-β vs Controle
e SLN.

68
No caso das células PC-3, o tratamento com TGF-β também induziu a
translocação da proteína Smad2 para o núcleo, que foi mais evidente na primeira h
de tratamento (Figura 17, TGF-β 45 min e 1h); nos tempos de 2h e 4h, a proteína
transloca de volta para o citoplasma e começa a se distribuir igualmente pela célula.
Nos tempos avaliados, a SLN não induziu a translocação de Smad2 para o núcleo
(Figura 17 B). A ativação da via do TGF-β tem como efeito a translocação de Smad2
na via canônica, enquanto que outros marcadores estão relacionados com a via não-
canônica, que também pode levar a efeitos em células de câncer (Vander Ark et al.
2018). Assim, a SLN pode estar ativando a migração através da regulação de outras
proteínas relacionadas. O TGF-β, através do seu receptor TβRI, poderia ativar
TRAF6 e MAP kinase, levando à ativação de p38 e JNK, induzindo invasão em
células tumorais e a transição epitélio-mesenquimal (Vander Ark et al. 2018). Esses
resultados criaram novas hipóteses, de que a SLN poderia induzir a migração
através da transição epitélio-mesenquimal (EMT) – considerando que células PC-3,
por se encontrarem em um estágio parcial de transição e por isso parte da
população das células ainda é suscetível à indução de EMT – nos fez investigar
algumas proteínas importantes dessa via.
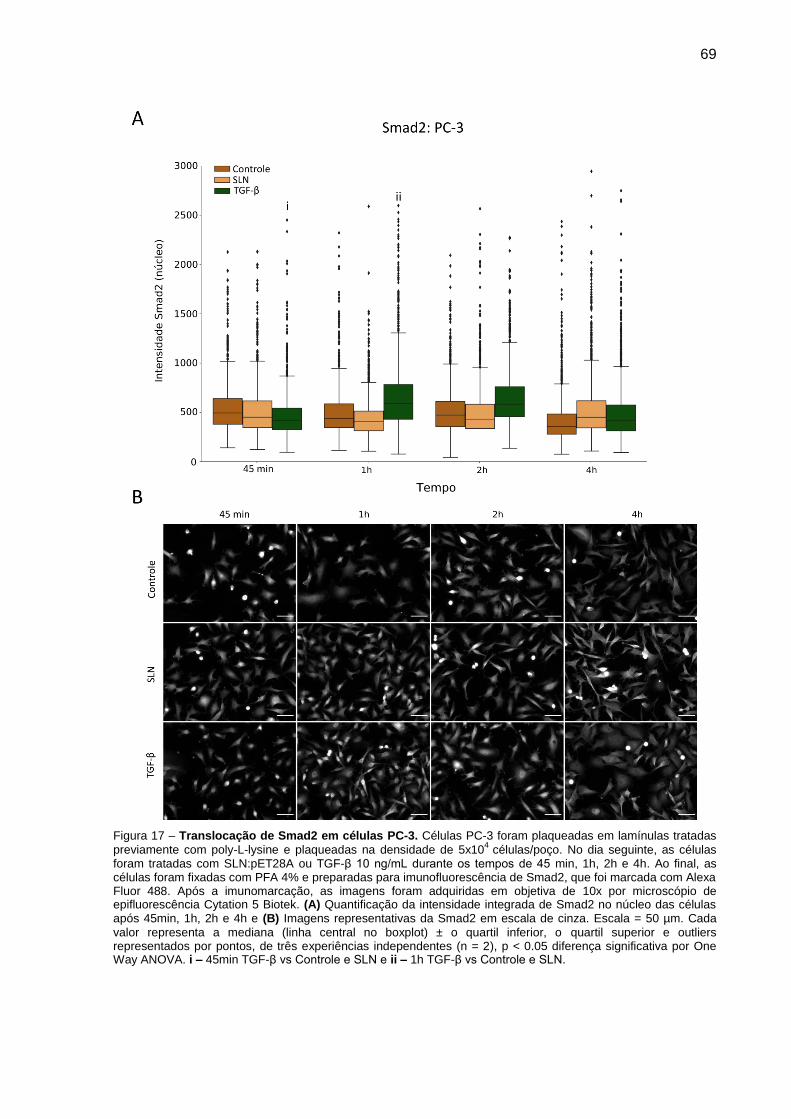
69
Figura 17 – Translocação de Smad2 em células PC-3. Células PC-3 foram plaqueadas em lamínulas tratadas
previamente com poly-L-lysine e plaqueadas na densidade de 5x104
células/poço. No dia seguinte, as células foram tratadas com SLN:pET28A ou TGF-β 10 ng/mL durante os tempos de 45 min, 1h, 2h e 4h. Ao final, as células foram fixadas com PFA 4% e preparadas para imunofluorescência de Smad2, que foi marcada com Alexa Fluor 488. Após a imunomarcação, as imagens foram adquiridas em objetiva de 10x por microscópio de epifluorescência Cytation 5 Biotek. (A) Quantificação da intensidade integrada de Smad2 no núcleo das células após 45min, 1h, 2h e 4h e (B) Imagens representativas da Smad2 em escala de cinza. Escala = 50 µm. Cada
valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 2), p < 0.05 diferença significativa por One Way ANOVA. i – 45min TGF-β vs Controle e SLN e ii – 1h TGF-β vs Controle e SLN.

70
f. SLN e a transição epitélio-mesenquimal (EMT)
Sabe-se que o TGF-β tem capacidade de induzir a EMT e dois modelos se
propõem a explicar o mecanismo de indução. Ambos os modelos, CBS (do inglês
Cascading Bistable Switches) e TCS (do inglês Ternary Chimera Switch), tem como
base loops de Snail1/miR-34 e Zeb1/miR-200, que funcionam como interruptores na
regulação. O TGF-β (exógeno) induz a expressão de Snail, e Snail por sua vez induz
a expressão de Zeb1 e inibe miR-200. Zeb1 é responsável por manter o fenótipo
mesenquimal ativo, inibindo E-caderina, marcador epitelial, e promovendo N-
caderina e vimentina, marcadores mesenquimais (Zhang et al. 2014).
Células PC-3 expressam tanto E-caderina quanto N-caderina, portanto essas
células estão em um estágio intermediário da transição, chamado de estado EMT
parcial. Células tumorais são heterogêneas e podem estar em um estado parcial de
EMT expressando E-caderina, e ao mesmo tempo apresentar invasão e migração
aumentadas (Fontana et al. 2019).
Para investigar os reguladores essenciais da transição Zeb1 e Snail, células
PNT1A e PC-3 foram transfectadas com SLN, o meio foi trocado e após 24 h
observamos os marcadores através de imunofluorescência (Figura 18 e Figura 19).
A intensidade de fluorescência dos marcadores no núcleo foi mensurada (Figura 19
A) para mais de 5x104 células por tratamento. Para células PNT1A, tanto Snail e
Zeb1 não foram afetadas pela transfecção por SLN; apenas as células tratadas com
TGF-β tiveram intensidade de Zeb1 diminuída no núcleo (Figura 18 A).
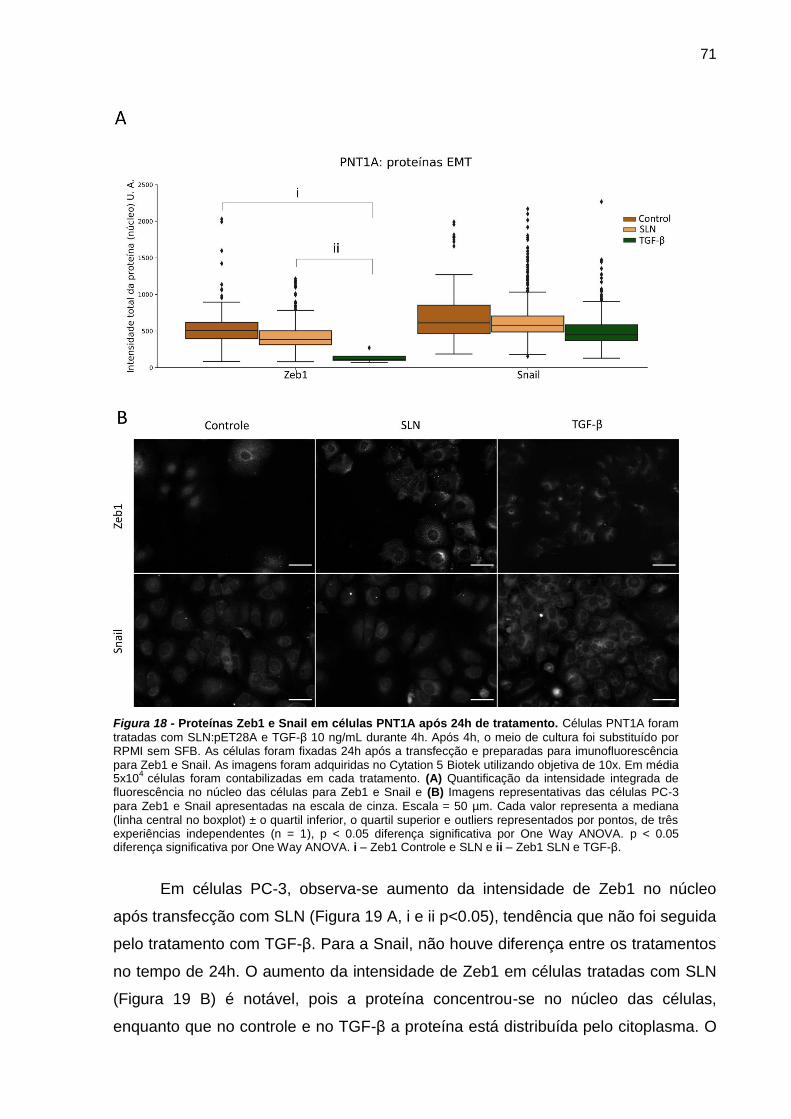
71
Figura 18 - Proteínas Zeb1 e Snail em células PNT1A após 24h de tratamento. Células PNT1A foram
tratadas com SLN:pET28A e TGF-β 10 ng/mL durante 4h. Após 4h, o meio de cultura foi substituído por RPMI sem SFB. As células foram fixadas 24h após a transfecção e preparadas para imunofluorescência para Zeb1 e Snail. As imagens foram adquiridas no Cytation 5 Biotek utilizando objetiva de 10x. Em média 5x10
4 células foram contabilizadas em cada tratamento. (A) Quantificação da intensidade integrada de
fluorescência no núcleo das células para Zeb1 e Snail e (B) Imagens representativas das células PC-3
para Zeb1 e Snail apresentadas na escala de cinza. Escala = 50 µm. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 1), p < 0.05 diferença significativa por One Way ANOVA. p < 0.05 diferença significativa por One Way ANOVA. i – Zeb1 Controle e SLN e ii – Zeb1 SLN e TGF-β.
Em células PC-3, observa-se aumento da intensidade de Zeb1 no núcleo
após transfecção com SLN (Figura 19 A, i e ii p<0.05), tendência que não foi seguida
pelo tratamento com TGF-β. Para a Snail, não houve diferença entre os tratamentos
no tempo de 24h. O aumento da intensidade de Zeb1 em células tratadas com SLN
(Figura 19 B) é notável, pois a proteína concentrou-se no núcleo das células,
enquanto que no controle e no TGF-β a proteína está distribuída pelo citoplasma. O

72
fenótipo parcial (subpopulações epiteliais e mesenquimais) dessas células vai de
encontro a novos conceitos sobre EMT, que antes visto como um processo binário,
hoje é visto como um processo gradual em células tumorais, que exibem fenótipo
intermediário (Pastushenko e Blanpain 2019). Níveis aumentados de Zeb1 são
descritos para células PC-3, e sugere-se que no câncer de próstata Zeb1 tem papel
essencial no extravasamento de células e metástase: células que têm níveis
aumentados de Zeb1 e E-caderina diminuída adquirem características de migração
trans-endotelial in vitro, enquanto que a diminuição de Zeb1 restaura parcialmente o
fenótipo epitelial (Lo et al. 2017).
Estudos em células de mama mostram que Snail é induzido no início da
transição de um estado epitelial para um estado parcial ou transitório; enquanto que
Zeb1 é induzido em um segundo momento, na transição entre o estado parcial e o
estado mesenquimal, sendo que o knockdown de Zeb1 restaura características
epiteliais em células, mostrando que a translocação de Snail é necessária em um
primeiro momento, enquanto que Zeb1 é responsável por manter um estado
mesenquimal (Zhang et al. 2014). Em nosso ensaio nas células PC-3, podemos
hipotetizar que Snail foi expressa em um momento anterior e somente após 24h
vimos o aumento de Zeb1, em concordância com a literatura. Porém é necessário
investigar mais tempos, já que células PC-3 estão no estado parcial/transitório, já
que a transição total para mesenquimal é observada após uma semana de
tratamento (Zhang et al. 2014).
Ainda, tais resultados vão de encontro com o aumento da migração causado
por SLN em células PC-3, pois células que estão localizadas no fronte de migração
passam por EMT parcial e ganham o fenótipo mesenquimal, no sentido de direcionar
a migração, enquanto que retém características epiteliais para se manterem ligadas
às suas células vizinhas durante o avanço do fronte migratório (Lu e Kang 2019).
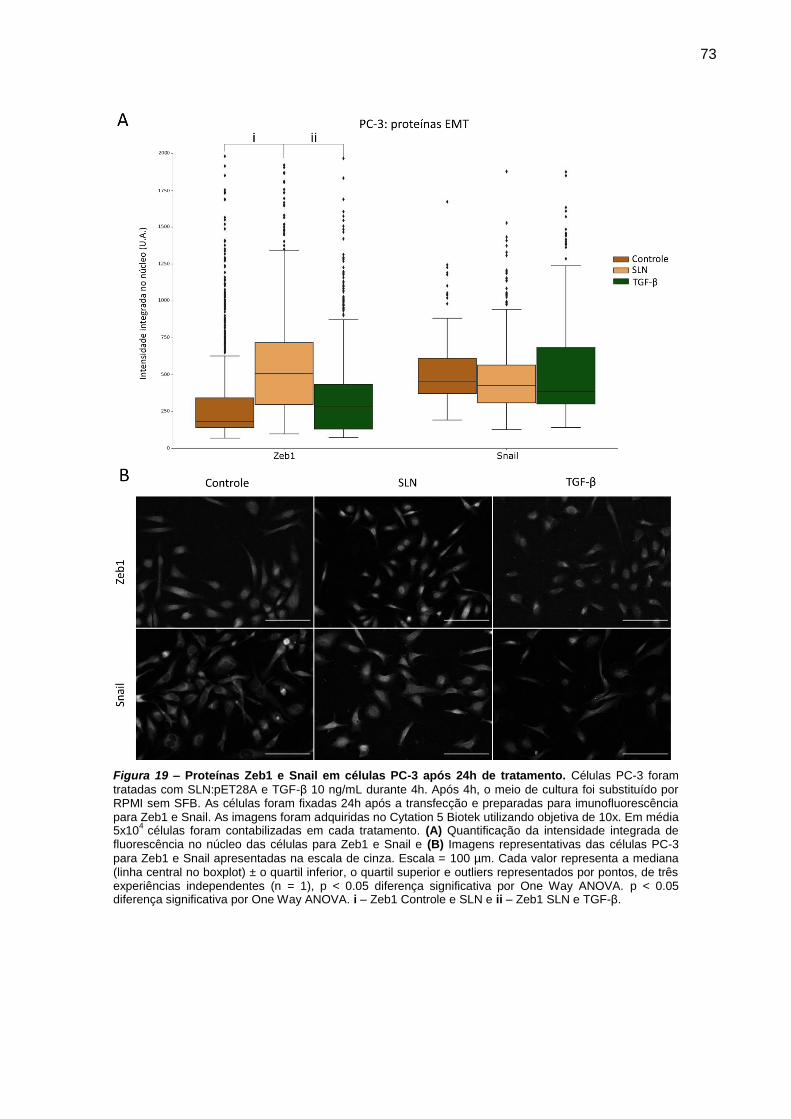
73
Figura 19 – Proteínas Zeb1 e Snail em células PC-3 após 24h de tratamento. Células PC-3 foram
tratadas com SLN:pET28A e TGF-β 10 ng/mL durante 4h. Após 4h, o meio de cultura foi substituído por RPMI sem SFB. As células foram fixadas 24h após a transfecção e preparadas para imunofluorescência para Zeb1 e Snail. As imagens foram adquiridas no Cytation 5 Biotek utilizando objetiva de 10x. Em média 5x10
4 células foram contabilizadas em cada tratamento. (A) Quantificação da intensidade integrada de
fluorescência no núcleo das células para Zeb1 e Snail e (B) Imagens representativas das células PC-3
para Zeb1 e Snail apresentadas na escala de cinza. Escala = 100 µm. Cada valor representa a mediana (linha central no boxplot) ± o quartil inferior, o quartil superior e outliers representados por pontos, de três experiências independentes (n = 1), p < 0.05 diferença significativa por One Way ANOVA. p < 0.05 diferença significativa por One Way ANOVA. i – Zeb1 Controle e SLN e ii – Zeb1 SLN e TGF-β.

74
Um dos marcadores mesenquimais críticos é a vimentina, uma proteína de
filamento intermediário que aparece em altos níveis durante a EMT (Lo et al. 2017),
pois têm papel na adesão e migração em vários tipos celulares (Ridge et al. 2018).
Para observar a distribuição dessa proteína nas células durante a migração,
realizamos a transfecção com SLN e o poço foi riscado; após 12h, as células foram
fixadas e marcadas para vimentina (Figura 20). Esse teste preliminar foi essencial
para mostrar a distribuição da vimentina em células PNT1A, já que observamos que
nem todas as células da população expressam o marcador, sendo que quando
foram tratadas com TGF-β, há uma tendência no aumento das células positivas para
vimentina (células com maior intensidade do marcador). A expressão de vimentina já
foi reportada para PNT1A, e sua presença em células epiteliais pode estar
relacionada às interações com actina e outros filamentos intermediários para uma
efetiva mudança no fenótipo celular (Lang et al. 2002). Para as células PC-3, a
marcação de vimentina é bem distribuída entre a população de células (Figura 20),
sendo que as células no fronte migratório são mais alongadas e apresentam fenótipo
mesenquimal, e podemos observá-las no centro do risco (células apontadas em
amarelo). Uma análise visual sugere um aumento da expressão da proteína nas
células tratadas com SLN. É sabido que esse marcador está presente em células
PC-3 (Steinmetz et al. 2011), e que pacientes com câncer de próstata com
metástase têm células positivas tanto para E-caderina quanto vimentina, sugerindo
um fenótipo in vivo intermediário (Osorio et al. 2016).
Esses resultados preliminares mostram que a vimentina parece ser
importante para a migração de células PC-3, e que o entendimento aprofundado da
expressão de vimentina ao longo do tempo, após a exposição por SLN, poderá
trazer novas hipóteses e resoluções na investigação da ativação da EMT por SLN
em células de próstata.
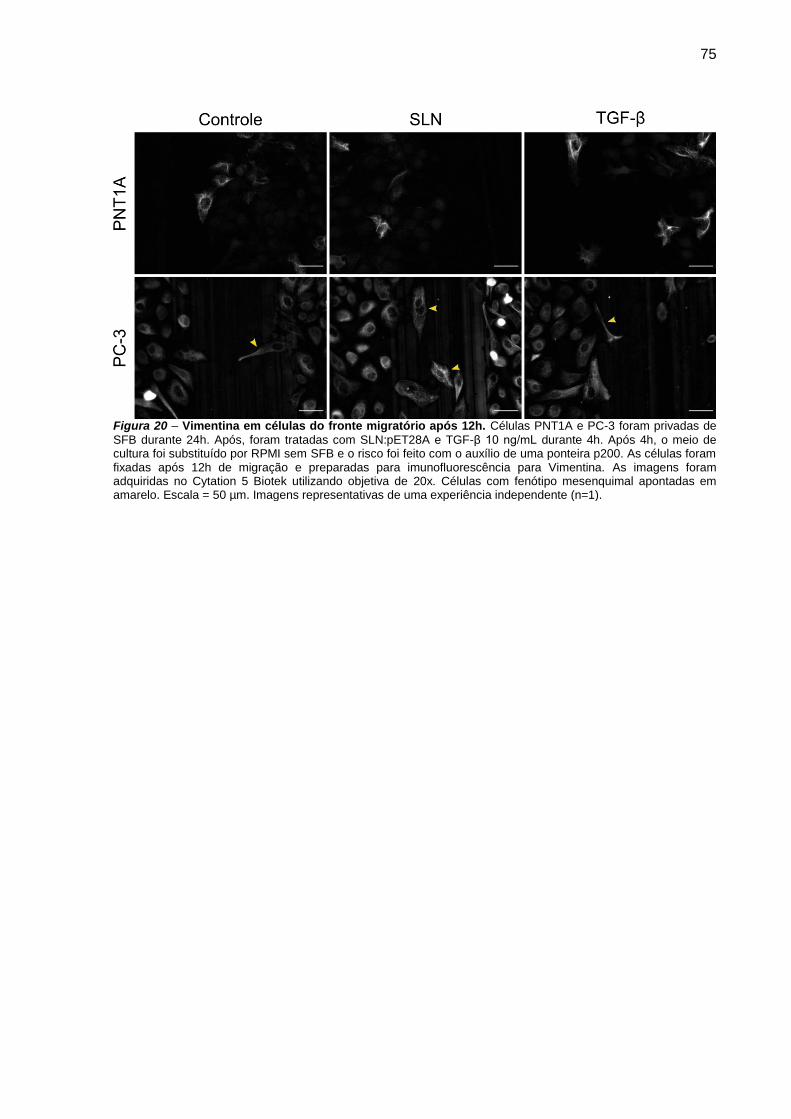
75
Figura 20 – Vimentina em células do fronte migratório após 12h. Células PNT1A e PC-3 foram privadas de
SFB durante 24h. Após, foram tratadas com SLN:pET28A e TGF-β 10 ng/mL durante 4h. Após 4h, o meio de cultura foi substituído por RPMI sem SFB e o risco foi feito com o auxílio de uma ponteira p200. As células foram fixadas após 12h de migração e preparadas para imunofluorescência para Vimentina. As imagens foram adquiridas no Cytation 5 Biotek utilizando objetiva de 20x. Células com fenótipo mesenquimal apontadas em amarelo. Escala = 50 µm. Imagens representativas de uma experiência independente (n=1).

76
5. Conclusão
Nesse projeto observamos que nanopartículas lipídicas sólidas (SLN) aumentam
a migração de células de câncer de próstata (PC-3) e que tal efeito se deve à SLN e não
aos componentes da nanopartícula em separadamente. Vimos também que a SLN
aumenta a migração de forma concentração-dependente nas células PC-3, e que há
uma saturação do aumento da velocidade em concentrações maiores que 3.8 µg/mL.
Mostramos que a migração é um efeito isolado, já que a proliferação celular de ambas
as células não são alteradas pelo tratamento com SLN. Outro ponto importante é que a
via do TGF-β tem relação com o efeito biológico de transfecção pela SLN, pois ao inibir
os receptores da via do TGF-β (LY2109761) e a fosforilação da Smad3 (SIS3 HCl), há
diminuição na transfecção por SLN em células PC-3. Além disso, a internalização da
SLN na presença desses inibidores não foi alterada, o que deixa uma hipótese em
aberto, de que o efeito da SLN na via do TGF-β ocorre no tráfego intracelular, processo
que deverá ser investigado em projetos futuros.
Sabendo que a via do TGF-β é influenciada pela SLN em células PC-3,
investigamos também a translocação de Smad2 para o núcleo, o que responderia
muitas questões. Porém, nos tempos observados, não houve translocação de Smad2 na
presença da SLN, o que gerou novas hipóteses, de que a ativação seria da via do TGF-
β não-canônica. Isso nos fez imaginar se a transição epitélio-mesenquimal (EMT), um
processo que aumenta a motilidade celular através do fenótipo da célula, poderia estar
relacionada com o aumento da migração. Vimos então que em células PC-3, há a
translocação de Zeb1, um importante fator de transcrição relacionado com o fenótipo
mesenquimal na EMT.
O uso de nanopartículas para entrega de genes e fármacos continua sendo
estudado com foco em aplicações terapêuticas, e o entendimento dos efeitos adversos
causados pela mesma se torna necessário, pois limita seu uso e segurança. Dessa
forma, esse trabalho trás a tona questões importantes sobre nanotoxicidade –
nanopartículas que parecem inofensivas à primeira vista, em razão da sua composição
biocompatível, podem não atender às expectativas e apresentarem efeitos sutis, porém
malignos, em células de câncer. Trabalhos in vitro são essenciais para identificar essas
sutilezas e revelar efeitos que poupam ensaios in vivo ou mesmo clínicos, que tem a
intenção de tratar doenças como a próstata, no nosso caso. No futuro, esperamos
desvendar outros mecanismos moleculares envolvendo nanopartículas, relevantes para
o avanço da área de intersecção entre biologia e nanotecnologia.

77
6. Referências bibliográficas
Akanda, Mushfiq H., Rajeev Rai, Ian J. Slipper, Babur Z. Chowdhry, Dimitrios Lamprou, Giulia Getti, e Dennis Douroumis. 2015. “Delivery of Retinoic Acid to LNCap Human Prostate Cancer Cells Using Solid Lipid Nanoparticles”. International Journal of Pharmaceutics 493:161–171.
Badawi, Noha M., Mahmoud H. Teaima, Khalid M. El-Say, Dalia A. Attia, Mohamed A. El-Nabarawi, e Mohey M. Elmazar. 2018. “Pomegranate Extract-Loaded Solid Lipid Nanoparticles: Design, Optimization, and in Vitro Cytotoxicity Study”. International Journal of Nanomedicine 13:1313–26.
Barbieri, Elisa, Pier Paolo Di Fiore, e Sara Sigismund. 2016. “Endocytic control of signaling at the plasma membrane”. Current Opinion in Cell Biology 39:21–27.
Barrett, Cachétne S. X., Ana C. Millena, e Shafiq A. Khan. 2017. “TGF-β Effects on Prostate Cancer Cell Migration and Invasion Require FosB”. The Prostate 77(1):72–81.
Berg, Jeremy M., John L. Tymoczko, Lubert Stryer, Jeremy M. Berg, John L. Tymoczko, e Lubert Stryer. 2002. Biochemistry. 5th ed. W H Freeman.
Bharali, Dhruba J., Thangirala Sudha, Huadong Cui, Badar M. Mian, e Shaker A. Mousa. 2017. “Anti-CD24 Nano-Targeted Delivery of Docetaxel for the Treatment of Prostate Cancer”. Nanomedicine: Nanotechnology, Biology, and Medicine 13(1):263–73.
Bhatt, Shailendra, Jaibharti Sharma, Manishkumar Singh, e Vipin Saini. 2018. “Solid Lipid Nanoparticles: A Promising Technology for Delivery of Poorly Water-Soluble Drugs”. ACTA Pharmaceutica Sciencia 56(3):27.
Bhullar, Khushwant S., Naiara Orrego Lagarón, Eileen M. McGowan, Indu Parmar, Amitabh Jha, Basil P. Hubbard, e H. P. Vasantha Rupasinghe. 2018. “Kinase-Targeted Cancer Therapies: Progress, Challenges and Future Directions”. Molecular Cancer 17(1):48.
Bray, Mark-Anthony, e Anne E. Carpenter. 2018. “Quality control for high-throughput imaging experiments using machine learning in CellProfiler”. Methods in molecular biology (Clifton, N.J.) 1683:89–112.
Budi, Erine H., Dana Duan, e Rik Derynck. 2017. “Transforming Growth Factor-β Receptors and Smads: Regulatory Complexity and Functional Versatility”. Trends in Cell Biology 27(9):658–72.
Budi, Erine H., Baby-Periyanayaki Muthusamy, e Rik Derynck. 2015. “The Insulin Response Integrates Increased TGF-β Signaling through Akt-Induced Enhancement of Cell Surface Delivery of TGF-β Receptors”. Science Signaling 8(396):ra96–ra96.
Cao, Zheng, e Natasha Kyprianou. 2015. “Mechanisms Navigating the TGF-β Pathway in Prostate Cancer”. Asian Journal of Urology 2(1):11–18.
Cardarelli, Francesco, Luca Digiacomo, Cristina Marchini, Augusto Amici, Fabrizio Salomone, Giuseppe Fiume, Alessandro Rossetta, Enrico Gratton, Daniela Pozzi, e Giulio Caracciolo. 2016. “The Intracellular Trafficking Mechanism of Lipofectamine-Based Transfection Reagents and Its Implication for Gene Delivery”. Scientific Reports 6:25879.

78
Carpenter, Anne E., Thouis R. Jones, Michael R. Lamprecht, Colin Clarke, In Han Kang, Ola Friman, David A. Guertin, Joo Han Chang, Robert A. Lindquist, Jason Moffat, Polina Golland, e David M. Sabatini. 2006. “CellProfiler: image analysis software for identifying and quantifying cell phenotypes”. Genome Biology 7(10):R100.
Chaffer, Christine L., e Robert A. Weinberg. 2011. “A Perspective on Cancer Cell Metastasis”. Science 331(6024):1559–64.
Chaikuad, Apirat, e Alex N. Bullock. 2016. “Structural Basis of Intracellular TGF-β Signaling: Receptors and Smads”. Cold Spring Harbor Perspectives in Biology 8(11).
Chaudhry, P., F. Fabi, M. Singh, S. Parent, V. Leblanc, e E. Asselin. 2014. “Prostate Apoptosis Response-4 Mediates TGF- β -Induced Epithelial-to-Mesenchymal Transition”. Cell Death & Disease 5(2):e1044–e1044.
Chen, Kai, Xue Li, Hongyan Zhu, Qiyong Gong, e Kui Luo. 2018. “Endocytosis of Nanoscale Systems for Cancer Treatments”. Current Medicinal Chemistry 25(25):3017–35.
Chen, Ye-Guang. 2009. “Endocytic Regulation of TGF-Beta Signaling”. Cell Research 19(1):58–70.
Chihara, Yoko, Masafumi Shimoda, Ami Hori, Ako Ohara, Yasuto Naoi, Jun-ichiro Ikeda, Naofumi Kagara, Tomonori Tanei, Atsushi Shimomura, Kenzo Shimazu, Seung Jin Kim, e Shinzaburo Noguchi. 2017. “A Small-Molecule Inhibitor of SMAD3 Attenuates Resistance to Anti-HER2 Drugs in HER2-Positive Breast Cancer Cells”. Breast Cancer Research and Treatment 166(1):55–68.
Danaei, M., M. Dehghankhold, S. Ataei, F. Hasanzadeh Davarani, R. Javanmard, A. Dokhani, S. Khorasani, e M. R. Mozafari. 2018. “Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems”. Pharmaceutics 10(2).
Dao, David, Adam N. Fraser, Jane Hung, Vebjorn Ljosa, Shantanu Singh, e Anne E. Carpenter. 2016. “CellProfiler Analyst: Interactive Data Exploration, Analysis and Classification of Large Biological Image Sets”. Bioinformatics 32(20):3210–12.
David, Charles J., e Joan Massagué. 2018. “Contextual Determinants of TGFβ Action in Development, Immunity and Cancer”. Nature Reviews. Molecular Cell Biology 19(7):419–35.
Dicken, Haley, Patrick J. Hensley, e Natasha Kyprianou. 2019. “Prostate tumor neuroendocrine differentiation via EMT: The road less traveled”. Asian Journal of Urology 6(1):82–90.
Domińska, Kamila, Tomasz Ochędalski, Karolina Kowalska, Zuzanna E. Matysiak-Burzyńska, Elżbieta Płuciennik, e Agnieszka W. Piastowska-Ciesielska. 2016. “A Common Effect of Angiotensin II and Relaxin 2 on the PNT1A Normal Prostate Epithelial Cell Line”. Journal of Physiology and Biochemistry 72(3):381–92.
Faresse, Nourdine, Frédéric Colland, Nathalie Ferrand, Céline Prunier, Marie-Francoise Bourgeade, e Azeddine Atfi. 2008. “Identification of PCTA, a TGIF Antagonist That Promotes PML Function in TGF-Beta Signalling”. The EMBO Journal 27(13):1804–15.
Ferreira, L. A. B., A. Radaic, G. O. Pugliese, M. B. Valentini, M. R. Oliveira, e M. B. de Jesus. 2014. “Endocitose e tráfego intracelular de nanomateriais”.

79
Flomerfelt, Francis A., e Ronald E. Gress. 2016. “Analysis of Cell Proliferation and Homeostasis Using EdU Labeling”. P. 211–20 in T-Cell Development: Methods and Protocols, Methods in Molecular Biology, organizado por R. Bosselut e M. S. Vacchio. New York, NY: Springer New York.
Fontana, Fabrizio, Michela Raimondi, Monica Marzagalli, Michele Sommariva, Patrizia Limonta, e Nicoletta Gagliano. 2019. “Epithelial-To-Mesenchymal Transition Markers and CD44 Isoforms Are Differently Expressed in 2D and 3D Cell Cultures of Prostate Cancer Cells”. Cells 8(2).
Foroozandeh, Parisa, e Azlan Abdul Aziz. 2018. “Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles”. Nanoscale Research Letters 13(1):339.
Freire-de-Lima, Leonardo, Kirill Gelfenbeyn, Yao Ding, Ulla Mandel, Henrik Clausen, Kazuko Handa, e Sen-itiroh Hakomori. 2011. “Involvement of O-Glycosylation Defining Oncofetal Fibronectin in Epithelial-Mesenchymal Transition Process”. Proceedings of the National Academy of Sciences 108(43):17690–95.
Gallaher, Jill A., Joel S. Brown, e Alexander R. A. Anderson. 2019. “The Impact of Proliferation-Migration Tradeoffs on Phenotypic Evolution in Cancer”. Scientific Reports 9(1):1–10.
Garcia-Fossa, Fernanda, Vladimir Gaal, e Marcelo B. de Jesus. 2020. “PyScratch: an ease of use tool for analysis of Scratch assays.” in press.
Grada, Ayman, Marta Otero-Vinas, Francisco Prieto-Castrillo, Zaidal Obagi, e Vincent Falanga. 2017. “Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay”. Journal of Investigative Dermatology 137(2):e11–16.
Gu, Shuchen, e Xin-Hua Feng. 2018. “TGF-β Signaling in Cancer”. Acta Biochimica et Biophysica Sinica 50(10):941–49.
Guo, Da Dong, Qing Ning Li, Chun Min Li, e Hong Sheng Bi. 2015. “Zinc oxide nanoparticles inhibit murine photoreceptor-derived cell proliferation and migration via reducing TGF-β and MMP-9 expression in vitro”. Cell Proliferation 48(2):198–208.
Hao, Yang, David Baker, e Peter ten Dijke. 2019. “TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis”. International Journal of Molecular Sciences 20(11):2767.
He, Kangmin, Xiaohua Yan, Nan Li, Song Dang, Li Xu, Bing Zhao, Zijian Li, Zhizhen Lv, Xiaohong Fang, Youyi Zhang, e Ye-Guang Chen. 2015. “Internalization of the TGF-β Type I Receptor into Caveolin-1 and EEA1 Double-Positive Early Endosomes”. Cell Research 25(6):738–52.
He, Xiaojun, Xiaopeng Guo, Hongsen Zhang, Xiangchuang Kong, Fan Yang, e Chuansheng Zheng. 2017. “Mechanism of action and efficacy of LY2109761, a TGF-β receptor inhibitor, targeting tumor microenvironment in liver cancer after TACE”. Oncotarget 9(1):1130–42.
Hinze, Claudia, e Emmanuel Boucrot. 2018. “Endocytosis in Proliferating, Quiescent and Terminally Differentiated Cells”. Journal of Cell Science 131(23).
Hong, Seul-Ki, Eun-Ho Choo, Sang-Hyun Ihm, Kiyuk Chang, e Ki-Bae Seung. 2017. “Dipeptidyl peptidase 4 inhibitor attenuates obesity-induced myocardial fibrosis by

80
inhibiting transforming growth factor-βl and Smad2/3 pathways in high-fat diet-induced obesity rat model”. Metabolism 76:42–55.
de Jesus, Marcelo B., Allan Radaic, Wouter L. J. Hinrichs, Carmen V. Ferreira, Eneida de Paula, Dick Hoekstra, e Inge S. Zuhorn. 2014. “Inclusion of the Helper Lipid Dioleoyl-Phosphatidylethanolamine in Solid Lipid Nanoparticles Inhibits Their Transfection Efficiency”. Journal of Biomedical Nanotechnology 10(2):355–65.
de Jesus, Marcelo B., e Inge S. Zuhorn. 2015. “Solid lipid nanoparticles as nucleic acid delivery system: Properties and molecular mechanisms”. Journal of Controlled Release 201:1–13.
de Jesus, Marcelo Bispo, Carmen Veríssima Ferreira, Eneida de Paula, Dick Hoekstra, e Inge S. Zuhorn. 2010. “Design of Solid Lipid Nanoparticles for Gene Delivery into Prostate Cancer”. Journal of Controlled Release: Official Journal of the Controlled Release Society 148(1):e89-90.
de Jesus, Marcelo Bispo, e Yvonne L. Kapila. 2014. “Cellular Mechanisms in Nanomaterial Internalization, Intracellular Trafficking, and Toxicity”. Nanomedicine and Nanotoxicology 201–27.
de Jesus, Marcelo Bispo, Allan Radaic, Inge S. Zuhorn, e Eneida de Paula. 2013. “Microemulsion Extrusion Technique: A New Method to Produce Lipid Nanoparticles”. Journal of Nanoparticle Research 15(10):1960.
Jin, Yang, Zilian Cui, Xudong Li, Xunbo Jin, e Jian Peng. 2017. “Upregulation of long non-coding RNA PlncRNA-1 promotes proliferation and induces epithelial-mesenchymal transition in prostate cancer”. Oncotarget 8(16):26090–99.
Jinnin, Masatoshi, Hironobu Ihn, e Kunihiko Tamaki. 2006. “Characterization of SIS3, a Novel Specific Inhibitor of Smad3, and Its Effect on Transforming Growth Factor-Β1-Induced Extracellular Matrix Expression”. Molecular Pharmacology 69(2):597–607.
Kaksonen, Marko, e Aurélien Roux. 2018. “Mechanisms of Clathrin-Mediated Endocytosis”. Nature Reviews Molecular Cell Biology 19(5):313–26.
Kaplani, Konstantina, Stamatina Koutsi, Vasileios Armenis, Foteini G. Skondra, Nickolas Karantzelis, Spyridon Champeris Tsaniras, e Stavros Taraviras. 2018. “Wound healing related agents: Ongoing research and perspectives”. Advanced Drug Delivery Reviews 129:242–53.
Katsuno, Yoko, Dominique Stephan Meyer, Ziyang Zhang, Kevan M. Shokat, Rosemary J. Akhurst, Kohei Miyazono, e Rik Derynck. 2019. “Chronic TGF-β Exposure Drives Stabilized EMT, Tumor Stemness, and Cancer Drug Resistance with Vulnerability to Bitopic MTOR Inhibition”. Sci. Signal. 12(570):eaau8544.
Kimbrough-Allah, Mawiyah N., Ana C. Millena, e Shafiq A. Khan. 2018. “Differential Role of PTEN in Transforming Growth Factor β (TGF-β) Effects on Proliferation and Migration in Prostate Cancer Cells”. The Prostate 78(5):377–89.
Knight, Zachary A., e Kevan M. Shokat. 2005. “Features of Selective Kinase Inhibitors”. Chemistry & Biology 12(6):621–37.
Lambert, Arthur W., Diwakar R. Pattabiraman, e Robert A. Weinberg. 2017. “Emerging Biological Principles of Metastasis”. Cell 168(4):670–91.

81
Lang, Shona H., Catherine Hyde, Ian N. Reid, Ian S. Hitchcock, Claire A. Hart, A. A. Gordon Bryden, Jean-Marie Villette, Michael J. Stower, e Norman J. Maitland. 2002. “Enhanced Expression of Vimentin in Motile Prostate Cell Lines and in Poorly Differentiated and Metastatic Prostate Carcinoma”. The Prostate 52(4):253–63.
Li, Wei, Samuel Lee, Minglin Ma, Soo Min Kim, Patrick Guye, James R. Pancoast, Daniel G. Anderson, Ron Weiss, Richard T. Lee, e Paula T. Hammond. 2013. “Microbead-Based Biomimetic Synthetic Neighbors Enhance Survival and Function of Rat Pancreatic β-Cells”. Scientific Reports 3(1).
Li, Yichen, Wangxi Luo, e Weidong Yang. 2018. “Nuclear Transport and Accumulation of Smad Proteins Studied by Single-Molecule Microscopy”. Biophysical Journal 114(9):2243–51.
Li, Yihao, Xianjiang Kang, e Qiang Wang. 2011. “HSP70 decreases receptor-dependent phosphorylation of Smad2 and blocks TGF-β-induced epithelial-mesenchymal transition”. Journal of Genetics and Genomics 38(3):111–16.
Lichtman, Michael K., Marta Otero‐Vinas, e Vincent Falanga. 2016. “Transforming Growth Factor Beta (TGF-β) Isoforms in Wound Healing and Fibrosis”. Wound Repair and Regeneration 24(2):215–22.
Liu, Fang. 2008. “PCTA: A New Player in TGF-Beta Signaling”. Science Signaling 1(46):pe49.
Liu, Jinglan, Gurpreet Kaur, Vikramjit K. Zhawar, Drazen B. Zimonjic, Nicholas C. Popescu, Raj P. Kandpal, e Raghbir S. Athwal. 2009. “Role of SV40 Integration Site at Chromosomal Interval 1q21.1 in Immortalized CRL2504 Cells”. Cancer research 69(19):7819–25.
Liu, Yang, Manoj N. Krishnan, e Kyle K. L. Phua. 2017. “Suppression of mRNA Nanoparticle Transfection in Human Fibroblasts by Selected Interferon Inhibiting Small Molecule Compounds”. Biomolecules 7(3).
Lo, U. Ging, Cheng-Fan Lee, Ming-Shyue Lee, e Jer-Tsong Hsieh. 2017. “The Role and Mechanism of Epithelial-to-Mesenchymal Transition in Prostate Cancer Progression”. International Journal of Molecular Sciences 18(10).
Lomas, Derek J., e Hashim U. Ahmed. 2020. “All Change in the Prostate Cancer Diagnostic Pathway”. Nature Reviews Clinical Oncology 1–10.
Lonez, Caroline, Marc F. Lensink, Michel Vandenbranden, e Jean-Marie Ruysschaert. 2009. “Cationic Lipids Activate Cellular Cascades. Which Receptors Are Involved?” Biochimica Et Biophysica Acta 1790(6):425–30.
Lonez, Caroline, Michel Vandenbranden, e Jean-Marie Ruysschaert. 2012. “Cationic Lipids Activate Intracellular Signaling Pathways”. Advanced Drug Delivery Reviews 64(15):1749–58.
Lovitt, Carrie J., David H. Hilko, Vicky M. Avery, e Sally-Ann Poulsen. 2016. “Development of Ethynyl-2′-Deoxyuridine Chemical Probes for Cell Proliferation”. Bioorganic & Medicinal Chemistry 24(18):4272–80.
Lu, Wei, e Yibin Kang. 2019. “Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis”. Developmental Cell 49(3):361–74.

82
Luangmonkong, Theerut, Su Suriguga, Adhyatmika Adhyatmika, Amirah Adlia, Dorenda Oosterhuis, Chuthamanee Suthisisang, Koert P. de Jong, Henricus A. M. Mutsaers, e Peter Olinga. 2018. “In vitro and ex vivo anti-fibrotic effects of LY2109761, a small molecule inhibitor against TGF-β”. Toxicology and Applied Pharmacology 355:127–37.
Melisi, Davide, Rocio Garcia-Carbonero, Teresa Macarulla, Denis Pezet, Gael Deplanque, Martin Fuchs, Jorg Trojan, Helmut Oettle, Mark Kozloff, Ann Cleverly, Claire Smith, Shawn T. Estrem, Ivelina Gueorguieva, Michael M. F. Lahn, Al Blunt, Karim A. Benhadji, e Josep Tabernero. 2018. “Galunisertib plus Gemcitabine vs. Gemcitabine for First-Line Treatment of Patients with Unresectable Pancreatic Cancer”. British Journal of Cancer 119(10):1208.
Melkikh, Alexey V., e Maria Sutormina. 2020. “Intra- and Intercellular Transport of Substances: Models and Mechanisms”. Progress in Biophysics and Molecular Biology 150:184–202.
Mendonça, Monique Culturato Padilha, Allan Radaic, Fernanda Garcia-Fossa, Maria Alice da Cruz-Höfling, Marco Aurélio Ramirez Vinolo, e Marcelo Bispo de Jesus. 2019. “The in Vivo Toxicological Profile of Cationic Solid Lipid Nanoparticles”. Drug Delivery and Translational Research.
Mondello, Chiara, e Ilaria Chiodi. 2013. “Cellular immortalization and neoplastic transformation”. Cell Cycle 12(11):1804–5.
Moore, Rachel, Marta Giralt Pujol, Zhou Zhu, e Elizabeth Smythe. 2018. “Interplay of Endocytosis and Growth Factor Receptor Signalling”. Progress in Molecular and Subcellular Biology 57:181–202.
Moore, Thomas L., Dominic A. Urban, Laura Rodriguez-Lorenzo, Ana Milosevic, Federica Crippa, Miguel Spuch-Calvar, Sandor Balog, Barbara Rothen-Rutishauser, Marco Lattuada, e Alke Petri-Fink. 2019. “Nanoparticle Administration Method in Cell Culture Alters Particle-Cell Interaction”. Scientific Reports 9(1):1–9.
Nasirizadeh, Samira, e Bizhan Malaekeh-Nikouei. 2020. “Solid Lipid Nanoparticles and Nanostructured Lipid Carriers in Oral Cancer Drug Delivery”. Journal of Drug Delivery Science and Technology 55:101458.
Ogunjimi, Abiodun A., Elton Zeqiraj, Derek F. Ceccarelli, Frank Sicheri, Jeffrey L. Wrana, e Laurent David. 2012. “Structural Basis for Specificity of TGFβ Family Receptor Small Molecule Inhibitors”. Cellular Signalling 24(2):476–83.
Ohya, Yuichi, Akihiro Takahashi, e Akinori Kuzuya. 2018. “Preparation of Biodegradable Oligo(lactide)s-Grafted Dextran Nanogels for Efficient Drug Delivery by Controlling Intracellular Traffic”. International Journal of Molecular Sciences 19(6).
Orlowski, Piotr, Magdalena Zmigrodzka, Emilia Tomaszewska, Katarzyna Ranoszek-Soliwoda, Monika Czupryn, Malgorzata Antos-Bielska, Janusz Szemraj, Grzegorz Celichowski, Jaroslaw Grobelny, e Malgorzata Krzyzowska. 2018. “Tannic acid-modified silver nanoparticles for wound healing: the importance of size”. International Journal of Nanomedicine 13:991–1007.
Osorio, Luis A., Nancy M. Farfán, Enrique A. Castellón, e Héctor R. Contreras. 2016. “SNAIL transcription factor increases the motility and invasive capacity of prostate cancer cells”. Molecular Medicine Reports 13(1):778–86.

83
Pastushenko, Ievgenia, e Cédric Blanpain. 2019. “EMT Transition States during Tumor Progression and Metastasis”. Trends in Cell Biology 29(3):212–26.
Paul, Colin D., Panagiotis Mistriotis, e Konstantinos Konstantopoulos. 2017. “Cancer Cell Motility: Lessons from Migration in Confined Spaces”. Nature Reviews Cancer 17(2):131–40.
Radaic, A., e M. B. de Jesus. 2018. “Solid Lipid Nanoparticles Release DNA upon Endosomal Acidification in Human Embryonic Kidney Cells”. Nanotechnology 29(31):315102.
Radaic, Allan, Eneida de Paula, e Marcelo Bispo de Jesus. 2015. “Factorial Design and Development of Solid Lipid Nanoparticles (SLN) for Gene Delivery”. Journal of Nanoscience and Nanotechnology 15(2):1793–1800.
Radaic, Allan, Gustavo Oliveira Pugliese, Guilherme Capiraço Campese, Francisco Benedito Teixeira Pessine, Marcelo Bispo de Jesus, Allan Radaic, Gustavo Oliveira Pugliese, Guilherme Capiraço Campese, Francisco Benedito Teixeira Pessine, e Marcelo Bispo de Jesus. 2016. “STUDYING THE INTERACTIONS BETWEEN NANOPARTICLES AND BIOLOGICAL SYSTEMS”. Química Nova 39(10):1236–44.
Rehman, Zia ur, Dick Hoekstra, e Inge S. Zuhorn. 2011. “Protein kinase A inhibition modulates the intracellular routing of gene delivery vehicles in HeLa cells, leading to productive transfection”. Journal of Controlled Release 156(1):76–84.
Rehman, Zia ur, Inge S. Zuhorn, e Dick Hoekstra. 2013. “How cationic lipids transfer nucleic acids into cells and across cellular membranes: Recent advances”. Journal of Controlled Release 166(1):46–56.
Ridge, Sarah M., Dibyangana Bhattacharyya, Eoin Dervan, Serika D. Naicker, Amy J. Burke, J. M. Murphy, Karen O‟leary, John Greene, Aideen E. Ryan, Francis J. Sullivan, e Sharon A. Glynn. 2018. “Secreted Factors from Metastatic Prostate Cancer Cells Stimulate Mesenchymal Stem Cell Transition to a Pro-Tumourigenic „Activated‟ State That Enhances Prostate Cancer Cell Migration”. International Journal of Cancer 142(10):2056–67.
Roacho‐Perez, Jorge A., Hugo L. Gallardo‐Blanco, Margarita Sanchez‐Dominguez, Perla E.
Garcia‐Casillas, Christian Chapa‐Gonzalez, e Celia N. Sanchez‐Dominguez. 2018. “Nanoparticles for death‐induced gene therapy in cancer (Review)”. Molecular Medicine Reports 17(1):1413–20.
Rozés-Salvador, Victoria, Sebastian O. Siri, Melina M. Musri, e Cecilia Conde. 2018. “New Player in Endosomal Trafficking: Differential Roles of Smad Anchor for Receptor Activation (SARA) Protein”. Molecular and Cellular Biology 38(24).
Ruiz de Garibay, Aritz Perez, María Ángeles Solinís Aspiazu, Alicia Rodríguez Gascón, Haleh Ganjian, e Renate Fuchs. 2013. “Role of endocytic uptake in transfection efficiency of solid lipid nanoparticles-based nonviral vectors”. The Journal of Gene Medicine 15(11–12):427–40.
Sahay, Gaurav, Daria Y. Alakhova, e Alexander V. Kabanov. 2010. “Endocytosis of Nanomedicines”. Journal of Controlled Release 145(3):182–95.

84
Sanna, Vanna, e Mario Sechi. 2012. “Nanoparticle Therapeutics for Prostate Cancer Treatment”. Nanomedicine: Nanotechnology, Biology, and Medicine 8 Suppl 1:S31-36.
Sathianathen, Niranjan J., Badrinath R. Konety, Juanita Crook, Fred Saad, e Nathan Lawrentschuk. 2018. “Landmarks in Prostate Cancer”. Nature Reviews Urology 15(10):627–42.
Scheideler, Marcel, Ivan Vidakovic, e Ruth Prassl. 2020. “Lipid Nanocarriers for MicroRNA Delivery”. Chemistry and Physics of Lipids 226:104837.
Schindelin, Johannes, Ignacio Arganda-Carreras, Erwin Frise, Verena Kaynig, Mark Longair, Tobias Pietzsch, Stephan Preibisch, Curtis Rueden, Stephan Saalfeld, Benjamin Schmid, Jean-Yves Tinevez, Daniel James White, Volker Hartenstein, Kevin Eliceiri, Pavel Tomancak, e Albert Cardona. 2012. “Fiji: An Open-Source Platform for Biological-Image Analysis”. Nature Methods 9(7):676–82.
Schmid, Sandra L. 2017. “Reciprocal Regulation of Signaling and Endocytosis: Implications for the Evolving Cancer Cell”. J Cell Biol 216(9):2623–32.
Siegel, Rebecca L., Kimberly D. Miller, e Ahmedin Jemal. 2019. “Cancer Statistics, 2019”. CA: A Cancer Journal for Clinicians 69(1):7–34.
Steinmetz, Nicole F., Jochen Maurer, Huiming Sheng, Armand Bensussan, Igor Maricic, Vipin Kumar, e Todd A. Braciak. 2011. “Two Domains of Vimentin Are Expressed on the Surface of Lymph Node, Bone and Brain Metastatic Prostate Cancer Lines along with the Putative Stem Cell Marker Proteins CD44 and CD133”. Cancers 3(3):2870–85.
Stetefeld, Jörg, Sean A. McKenna, e Trushar R. Patel. 2016. “Dynamic light scattering: a practical guide and applications in biomedical sciences”. Biophysical Reviews 8(4):409–27.
Tellios, Vasiliki, Hong Liu, Nikoleta Tellios, Xiangji Li, e Cindy M. L. Hutnik. 2018. “Administration of Nitric Oxide Through a Novel Copper-Chitosan Delivery System in Human Corneal and Limbal Epithelial Cell Injury”. Investigative Ophthalmology & Visual Science 59(2):967–77.
Vander Ark, Alexandra, Jingchen Cao, e Xiaohong Li. 2018. “TGF-β receptors: In and beyond TGF-β signaling”. Cellular Signalling 52:112–20.
Vo, BaoHan T., Derrick Morton, Shravan Komaragiri, Ana C. Millena, Chelesie Leath, e Shafiq A. Khan. 2013. “TGF-β Effects on Prostate Cancer Cell Migration and Invasion Are Mediated by PGE2 through Activation of PI3K/AKT/mTOR Pathway”. Endocrinology 154(5):1768–79.
Voltan, Aline R., Kaila M. Alarcon, Ana M. Fusco-Almeida, Christiane P. Soares, Maria J. S. Mendes-Giannini, e Marlus Chorilli. 2017. “Highlights in Endocytosis of Nanostructured Systems”. Current Medicinal Chemistry 24(18):1909–29.
Walker, Lindsey, Ana C. Millena, Nicole Strong, e Shafiq A. Khan. 2013. “Expression of TGFβ3 and Its Effects on Migratory and Invasive Behavior of Prostate Cancer Cells: Involvement of PI3-Kinase/AKT Signaling Pathway”. Clinical & Experimental Metastasis 30(1):13–23.

85
Wan, Xinhai, Zhi-Gang Li, Jonathan M. Yingling, Jun Yang, Michael W. Starbuck, Murali K. Ravoori, Vikas Kundra, Elba Vazquez, e Nora M. Navone. 2012. “Effect of Transforming Growth Factor Beta (TGF-β) Receptor I Kinase Inhibitor on Prostate Cancer Bone Growth”. Bone 50(3):695–703.
Wang, Jun, Xiaojuan Li, Zhaoming Xiao, Yu Wang, Yuefu Han, Jun Li, Weian Zhu, Qu Leng,
Yuehui Wen, e Xinqiao Wen. 2019. “MicroRNA‐488 inhibits proliferation and glycolysis in human prostate cancer cells by regulating PFKFB3”. FEBS Open Bio 9(10):1798–1807.
Wang, Lingyan, Chun-Ying Ko, Erin E. Meyers, Benjamin S. Pedroja, Nadia Pelaez, e Audrey M. Bernstein. 2011. “Concentration-dependent effects of transforming growth factor β1 on corneal wound healing”. Molecular Vision 17:2835–46.
Wierzbicki, Mateusz, Sławomir Jaworski, Marta Kutwin, Marta Grodzik, Barbara Strojny, Natalia Kurantowicz, Krzysztof Zdunek, Rafał Chodun, André Chwalibog, e Ewa Sawosz. 2017. “Diamond, graphite, and graphene oxide nanoparticles decrease migration and invasiveness in glioblastoma cell lines by impairing extracellular adhesion”. International Journal of Nanomedicine 12:7241–54.
Zhang, Jingyu, Xiao-Jun Tian, Hang Zhang, Yue Teng, Ruoyan Li, Fan Bai, Subbiah Elankumaran, e Jianhua Xing. 2014. “TGF-β–Induced Epithelial-to-Mesenchymal Transition Proceeds through Stepwise Activation of Multiple Feedback Loops”. Science Signaling 7(345):ra91–ra91.
Zhang, Ying E. 2017. “Non-Smad Signaling Pathways of the TGF-β Family”. Cold Spring Harbor Perspectives in Biology 9(2):a022129.

86
7. Anexos
Documento Bioética e Biossegurança

87
Declaração autoria