
POLYMER PHYSICS & DNA
90
POLYMER PHYSICS & DNA Bertrand Duplantier Institut de Physique Théorique, Saclay, France Troisième Cycle de la physique en Suisse romande École Polytechnique Fédérale de Lausanne February-March 2009
Transcript of POLYMER PHYSICS & DNA

POLYMER PHYSICS & DNA
Bertrand Duplantier
Institut de Physique Théorique, Saclay, France
Troisième Cycle de la physique en Suisse romande
École Polytechnique Fédérale de Lausanne
February-March 2009

I. FREELY-JOINTED CHAIN
&
CONSTRAINTS
POLYMER PHYSICS & DNA
Troisième Cycle de la physique en Suisse romande
École Polytechnique Fédérale de Lausanne
12 March 2009
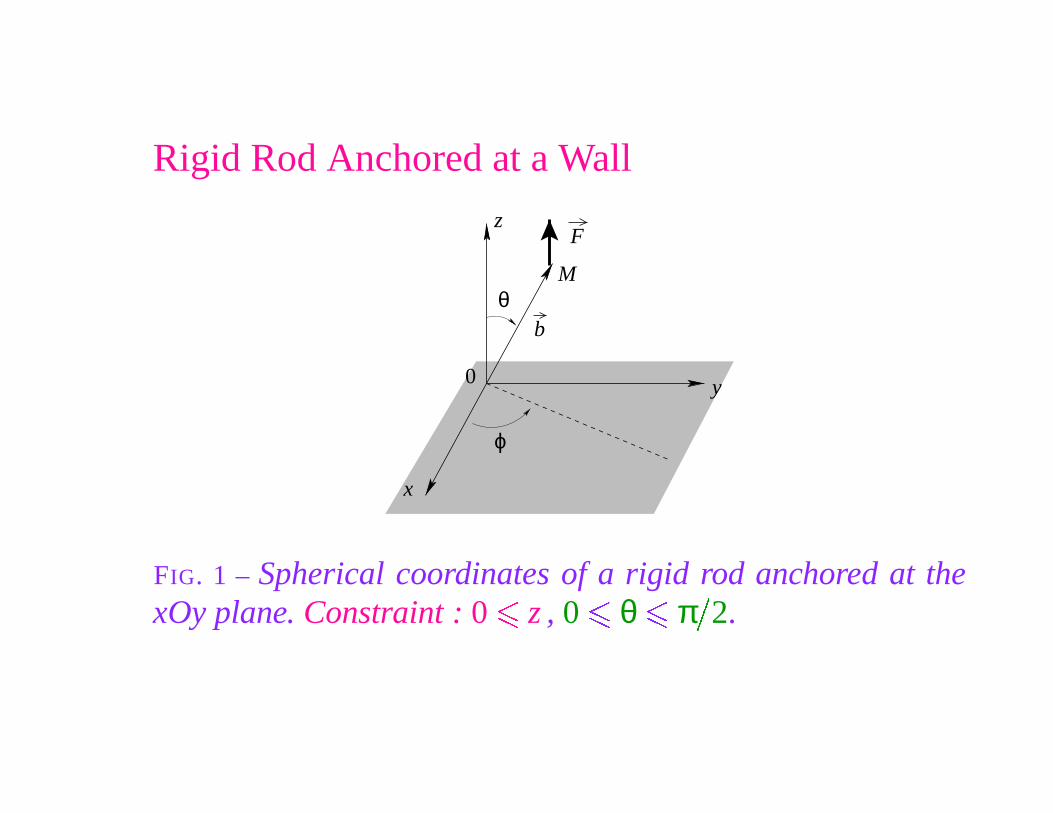
Rigid Rod Anchored at a Wall
F
θ
ϕ
0
b
y
z
x
M
FIG. 1 – Spherical coordinates of a rigid rod anchored at thexOy plane. Constraint : 0
�
z , 0
� θ � π
�
2.

Protein & Rigid Rod Anchored at a Wall
F
R0
θ
ϕ
0
b
y
z
x
M
FIG. 2 – Rigid rod anchored above the xOy plane wall, with abound protein of radius R0. Constraint : R0
�
z , 0
� θ � θ0 ,cosθ0
� R0
�
b.

Partition Functions
Z α
�
u
�
�
θ0
0sinθdθexp
�
βFbcosθ�
� expu � expαuu
��
α � cosθ0
� R0
�
b
�
� expu � 1u
�
�α � 0 � θ0
� π
�
2
�
� 2sinhu
u�
�
α � � 1 � θ0
� π
�
as a function of the dimensionless variable u � βFb.
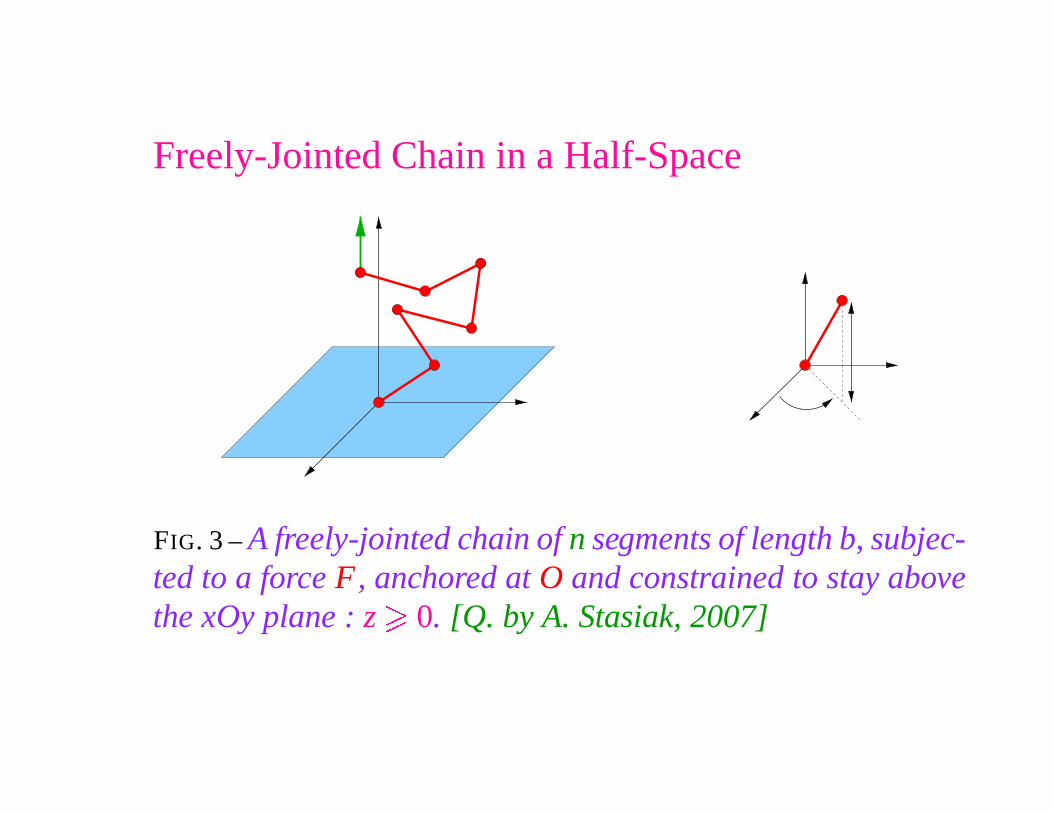
Freely-Jointed Chain in a Half-Space
FIG. 3 – A freely-jointed chain of n segments of length b, subjec-ted to a force F, anchored at O and constrained to stay abovethe xOy plane : z
�0. [Q. by A. Stasiak, 2007]
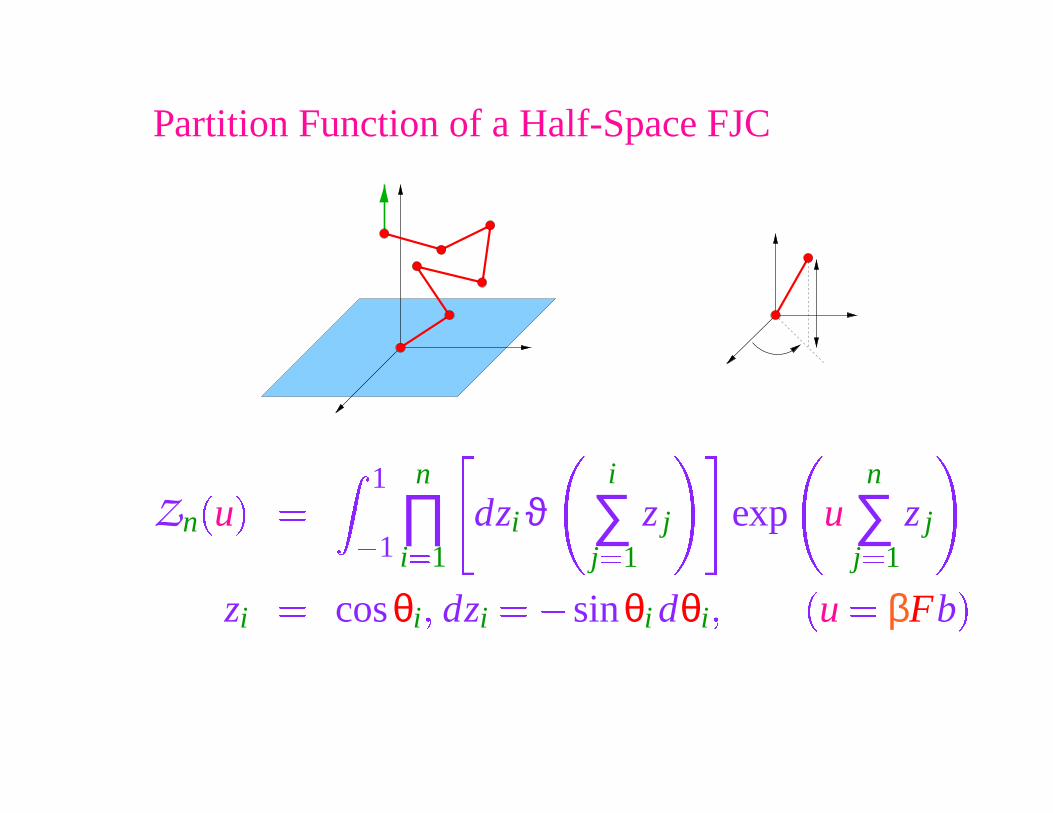
Partition Function of a Half-Space FJC
Zn
�
u
�
�
1
� 1
n
∏i �1
dzi ϑi
∑j �1
z j exp un
∑j �1
z j
zi
� cosθi � dzi
� � sinθi dθi �
�
u � βFb
�
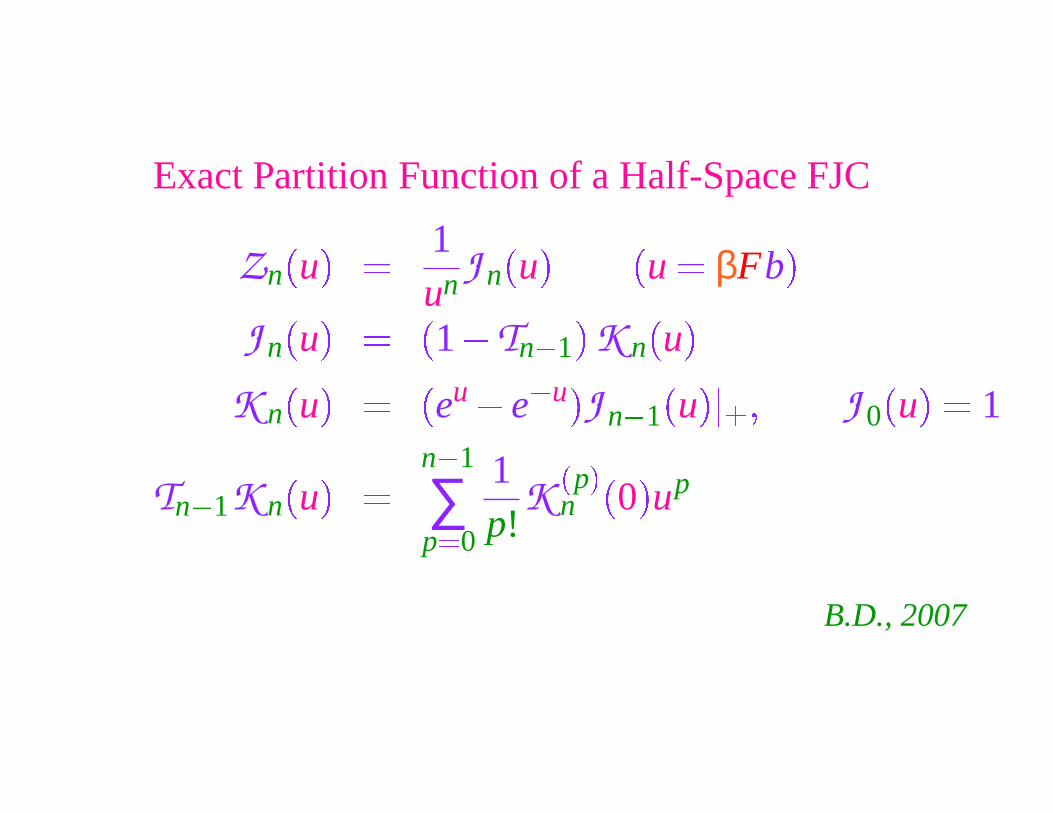
Exact Partition Function of a Half-Space FJC
Zn
�
u
�
� 1un J n
�
u
� �
u � βFb�
J n
�
u
�
�
�
1 � Tn � 1
�
K n�
u�
K n
�
u
�
�
�
eu � e
� u �
J n � 1
�
u
��
� � J 0
�
u
�
� 1
Tn � 1K n
�
u
�
�
n � 1
∑p �0
1p!
K�
p�
n
�
0
�
up
B.D., 2007
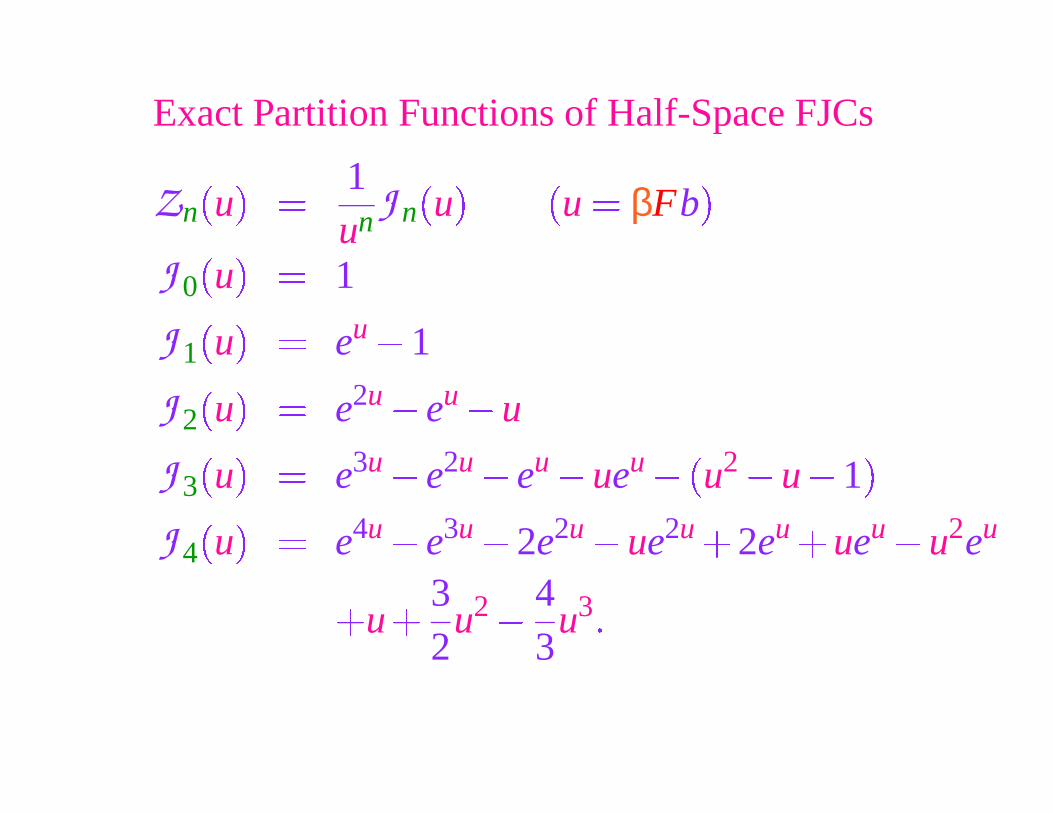
Exact Partition Functions of Half-Space FJCs
Zn
�
u
�
� 1un J n
�
u
� �
u � βFb
�
J 0
�
u
�
� 1
J 1
�
u
�
� eu � 1
J 2
�
u
�
� e2u � eu � u
J 3
�
u
�
� e3u � e2u � eu � ueu ��
u2 � u � 1
�
J 4
�
u
�
� e4u � e3u � 2e2u � ue2u 2eu ueu � u2eu
u32
u2 �
43
u3�
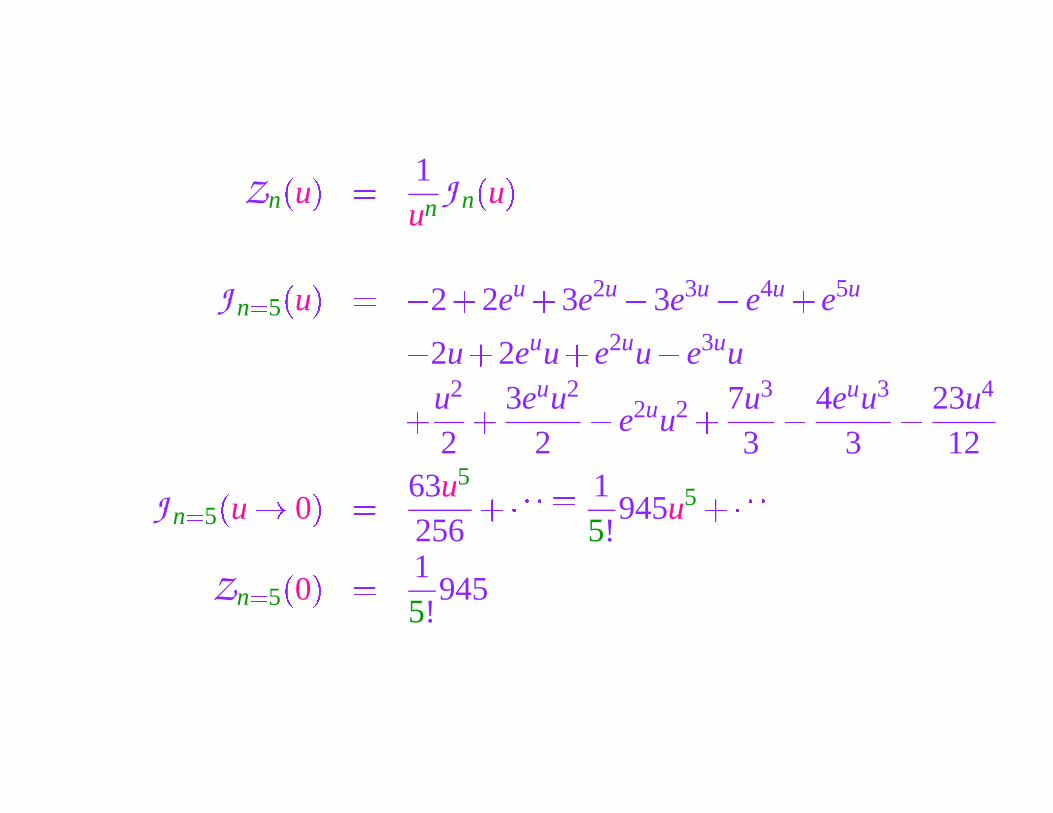
Zn
�
u
�
� 1un J n
�
u
�
J n �5
�
u
�
� � 2
�
2eu �
3e2u � 3e3u � e4u �
e5u
� 2u
�
2euu
�
e2uu � e3uu
� u2
2
� 3euu2
2� e2uu2 � 7u3
3
�
4euu3
3
�
23u4
12
J n �5
�
u � 0
�
� 63u5
256��� � � � 1
5!945u5 ��� � �
Zn �5
�
0
�
� 15!
945

J n �10
�
u
�
� � 42eu �
42e2u �
48e3u � 48e4u � 27e5u �
27e6u �
8e7u � 8e8u � e9u �
e10u
� 14u � 28euu
�
28e2uu
�
20e3uu � 20e4uu � 7e5uu�
7e6uu
�
e7uu � e8uu
� 21u2 �
7euu2 �
21e2uu2 � 11e3uu2 � 9e4uu2 � 112
e5uu2 � 32
e6uu2 � e7uu2
�
23u3
6
� 121euu3
6
�
376
e2uu3 �
232
e3uu3 � 112
e4uu3 � 73
e5uu3 �
43
e6uu3
� 65u4
4
� euu4
24
�
1158
e2uu4 � 214
e3uu4 � 154
e4uu4 �
2312
e5uu4
� 317u5
40
�
253euu5
15
� 24760
e2uu5 � 37160
e3uu5 �
4415
e4uu5
�
1013u6
60
� 22euu6
45� 937
90e2uu6 �
841180
e3uu6
�
799u7
90
� 7489euu7
420
�
2416315
e2uu7 � 208139u8
6720
�
259723euu8
20160
�
62476u9
2835

Zn
�
u
�
� 1un J n
�
u
�
J n �10
�
u � 0
�
� 46189u10
262144
��� � � � 110!
654729075u10 ��� � �
Zn �10
�
u � 0
�
� 110!
654729075
Zn
�
u � 0
�
� 1n!
�
2n � 1
� �
2n � 3
���� �
3� 1
(O. Bernardi, B.D., P. Nadeau, 2009)

II. DNA TOPOLOGY
POLYMER PHYSICS & DNA
Troisième Cycle de la physique en Suisse romande
École Polytechnique Fédérale de Lausanne
12 March 2009
FIG. 4 – Idealized structure of the double helix.
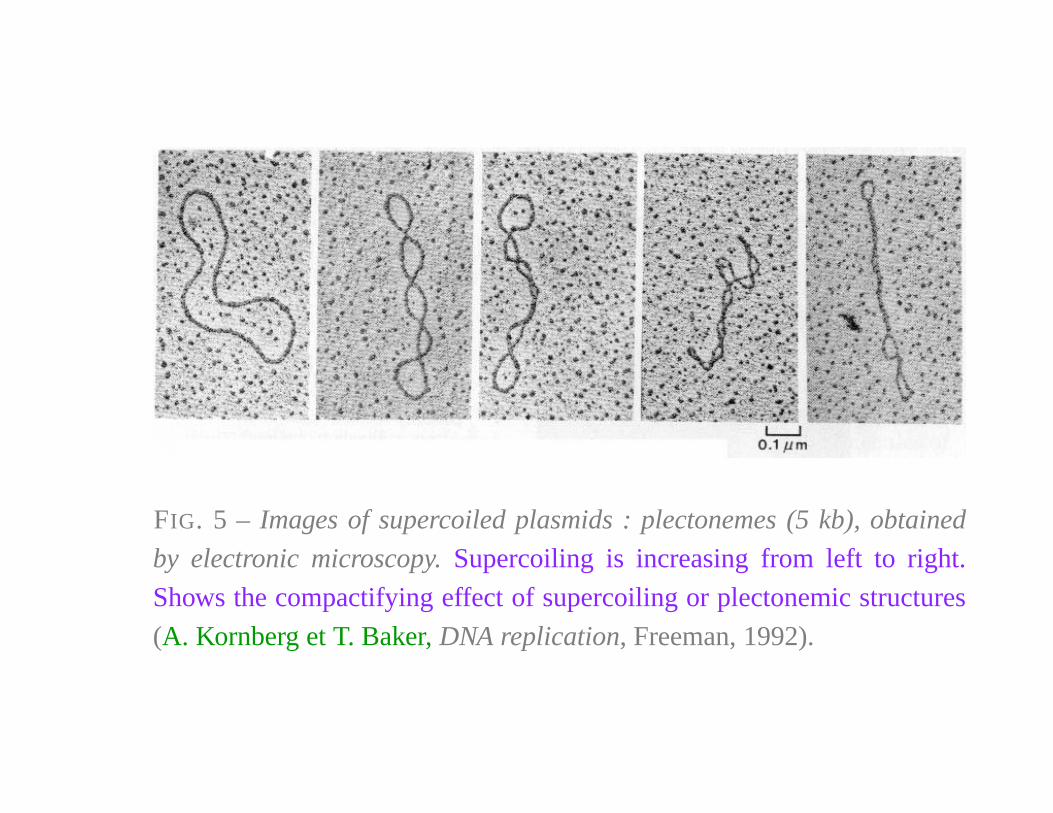
FIG. 5 – Images of supercoiled plasmids : plectonemes (5 kb), obtained
by electronic microscopy. Supercoiling is increasing from left to right.
Shows the compactifying effect of supercoiling or plectonemic structures
(A. Kornberg et T. Baker, DNA replication, Freeman, 1992).

FIG. 6 – Images of plectonemes of supercoiled plasmids (pUC19, 2686 pb
= 913 nm in B form), obtained by atomic force microscopy. [Courtesy by
G. Dietler and G. Witz, EPFL (January 2008).]
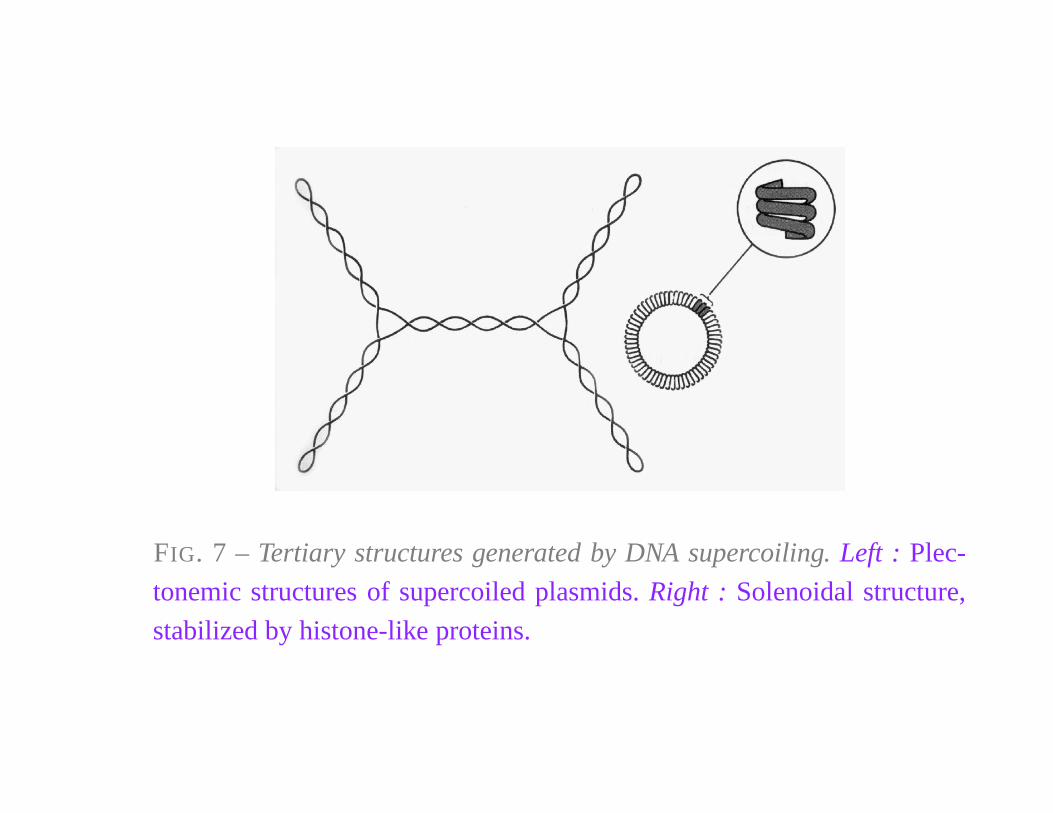
FIG. 7 – Tertiary structures generated by DNA supercoiling. Left : Plec-
tonemic structures of supercoiled plasmids. Right : Solenoidal structure,
stabilized by histone-like proteins.

TOPOLOGICAL FORMALISM
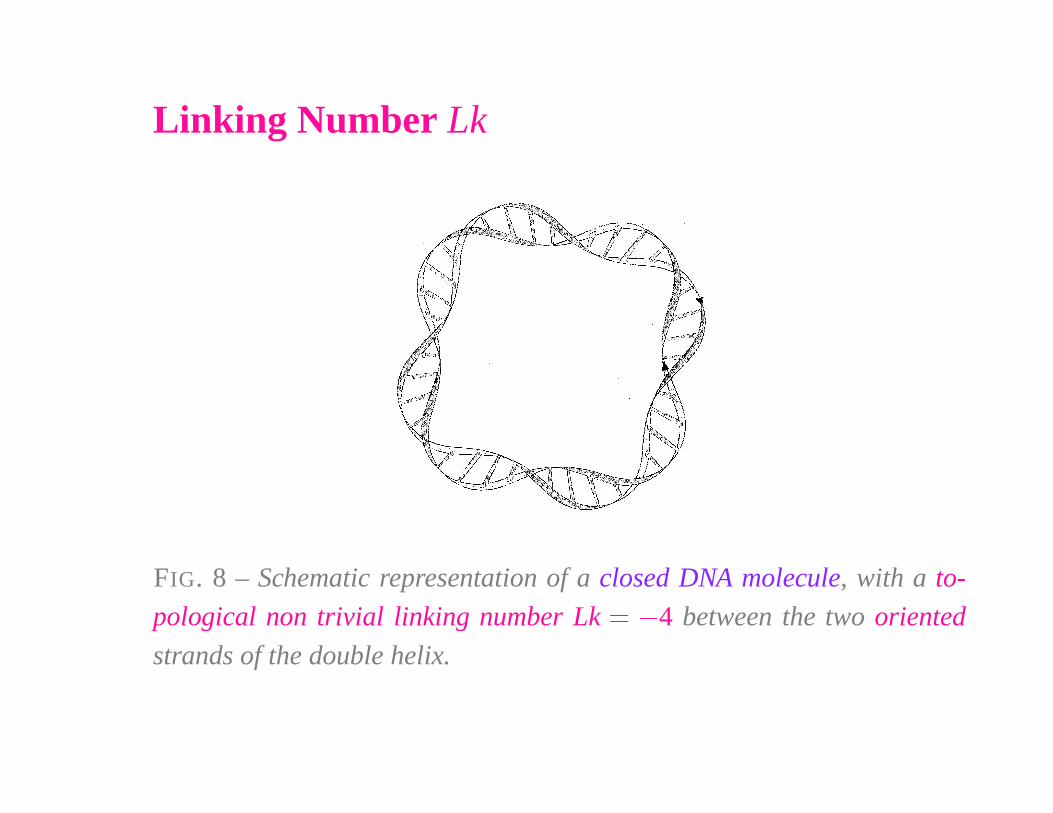
Linking Number Lk
FIG. 8 – Schematic representation of a closed DNA molecule, with a to-
pological non trivial linking number Lk � � 4 between the two oriented
strands of the double helix.
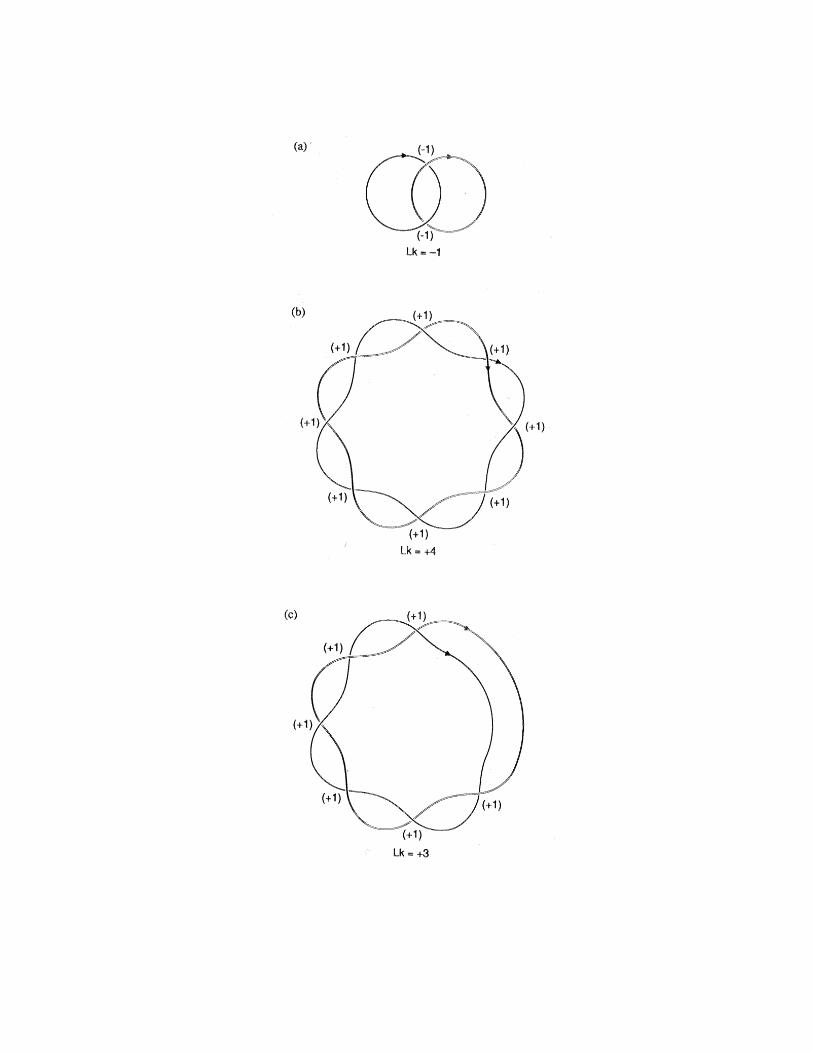
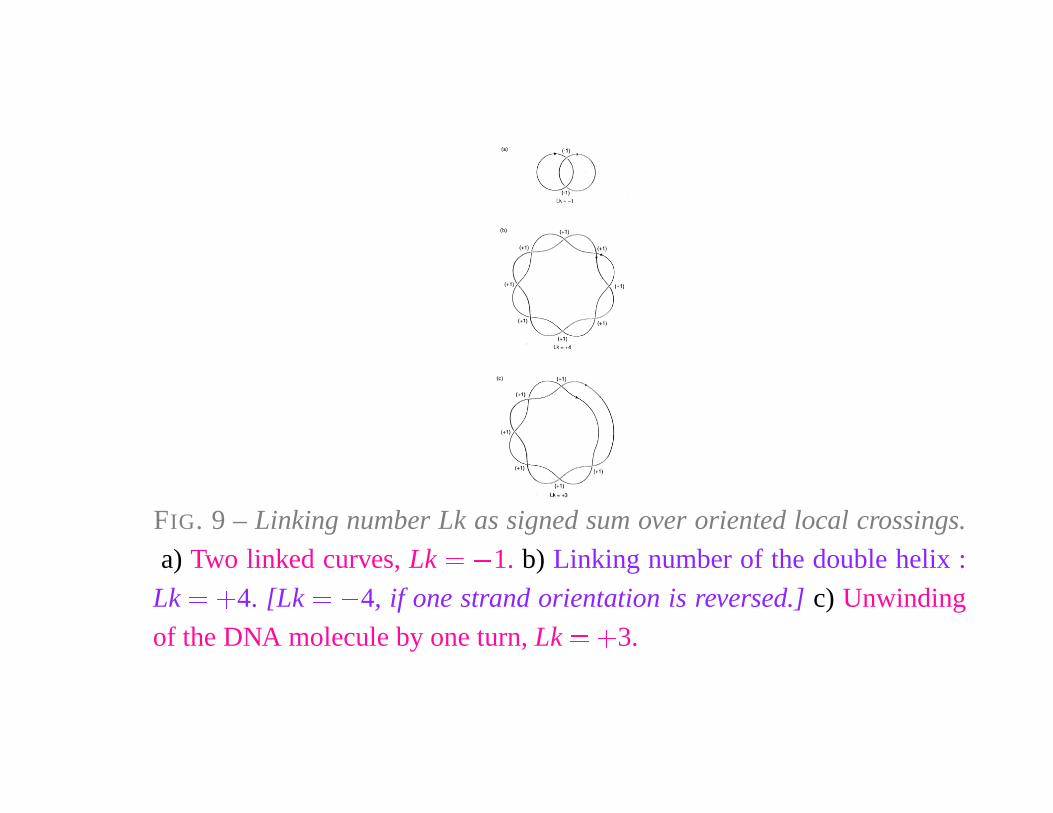
FIG. 9 – Linking number Lk as signed sum over oriented local crossings.
a) Two linked curves, Lk � � 1. b) Linking number of the double helix :
Lk � �
4. [Lk � � 4, if one strand orientation is reversed.] c) Unwinding
of the DNA molecule by one turn, Lk � �
3.

Linking Numbers & Local Crossings
The two DNA strands have a natural linking number Lk0, equal to the
number of base pairs N, divided by the double helix pitch h [h � 10 � 4
base pairs for B-DNA (standard form)] :
Lk0
� Nh
Plane projection of both oriented strands, and signed sum of their
apparent crossings :
Lk � 12 ∑
mutual crossings i
εi �
where εi
� �
1, depending on the right or left orientation of the i crossing.
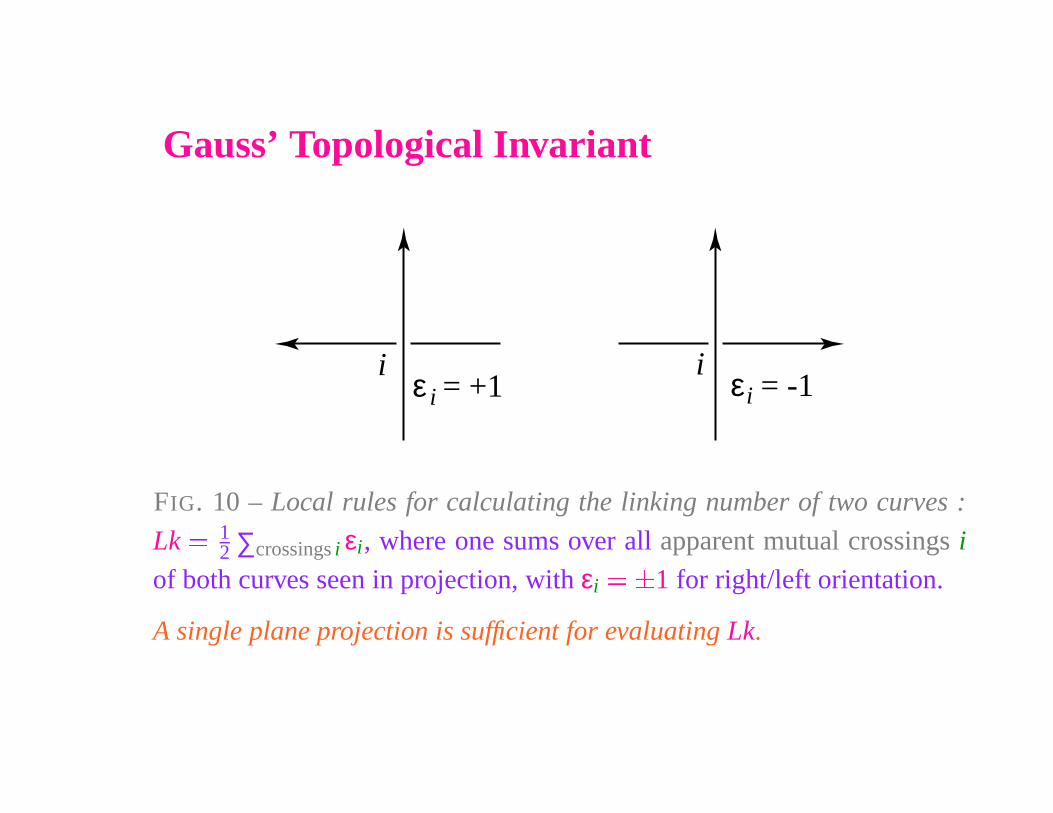
Gauss’ Topological Invariant
iε = +1i
iε = -1i
FIG. 10 – Local rules for calculating the linking number of two curves :
Lk � 12 ∑crossings i εi, where one sums over all apparent mutual crossings i
of both curves seen in projection, with εi
� �
1 for right/left orientation.
A single plane projection is sufficient for evaluating Lk.

The Gauss linking number is an algebraic topologicalinvariant.
It stays invariant as long as no DNA strand break appears. Upon
replication, or during reading of the genetic code by RNA-polymerases,
this topological invariant stays unchanged. When the extremities of a
linear double helix are fixed, as in a chromosome, a Gauss linking number
can also be defined.
DNA can also be supercoiled. One shall characterize itsdegree of supercoiling, together with torsion.

Twist Tw
Description of the geometry (and no longer the topology) of a DNA
double helix by “twist” and “writhe”.
� “Twisting number” Tw : counts the number of turns of the helix in
space. For an unconstrained linear B-DNA, the natural twist Tw0 is equal
to the base pair number N, divided by the helical pitch h (h � 10 � 4) :
Tw0� N
h
�
Identity of linking number and twist for a natural torsion-free DNA :
Lk0
� Tw0 �
Different in presence of supercoiling.

Writhe Wr
FIG. 11 – Example of writhing number�
Wr�
� 3. Counts theapparent self-crossings of the DNA helix backbone.
Definition : DNA arbitrarily oriented, followed by projectionon a random plane with a signed
�
1 counting for eachcrossing. The result is averaged over all planar projections in3-space :
Wr ��
∑self-crossings i
εi
�
plane �

Writhe and Projected Crossings
iε = +1i
iε = -1i
FIG. 12 – To calculate the “writhe” of a single curve, one has to sum over
apparent self-crossings, as seen in a planar projection, and to average over
the projection plane : W r ��
∑i εi
�
. The real number W � �
is the mean
“self-linking number” of the curve, and depends on its geometrical shape.

The Calugareanu-Fuller-White Theorem
� Unconstrained linear DNA : Lk and Tw are both equal tothe number of turns of the double helix : Lk0
� Tw0, withWr0
� 0.
� For a double stranded DNA, the sum of the Twisting andWrithing numbers is a topological invariant, the Linking
number of the two DNA helix strands :
Lk � Tw
�
Wr �
Topological conservation law for DNA functions, relating the intrinsic
(secondary) topological double-stranded structure of the helix to the
tertiary geometrical structure (twist & writhe) ; (G. Calugareanu, 1959, J.
Vinograd, 1965, F.B. Fuller, 1971).

The Calugareanu-Fuller-White Theorem
Historical Note : J. Vinograd and collaborators guessed in 1965 that there
existed a decomposition of the integer number Lk into the sum of two real
numbers, that describe the DNA geometry, and no longer only its
topology. The mathematician F. B. Fuller (1971) clarified Vinograd’s
empirical formula, while the differential geometer W. F. Pohl, contacted
by F. H. C. Crick, remarked that the formula Lk � Tw
�
Wr also resulted
from the PhD thesis of his former student J. H. White (1969), who
considered it in any space dimension. Actually, a definition of the writhe
W r and a proof in three dimensions appeared in earlier work by the
Romanian mathematician G. Calugareanu (1959-61).

Supercoiling
Double stranded DNA is supercoiled when Lk�
� Lk0, i.e.,when the molecule has a linking number excess or deficit,with respect to its torsion-free state
�
Wr0� 0
�
.
∆Lk � Lk � Lk0
� Tw � Tw0�
Wr � ∆Tw
�
Wr
σ � Lk � Lk0
Lk0
� ∆Tw
�
WrLk0
�
σ � 0 � supercoiled DNA ; σ � 0, undercoiled or negativelysupercoiled.
Bacterial plasmids have a natural negative supercoiling σ � � 0 � 06. For a
50 kpb DNA, this represents a deficit of 300 helical turns.

FIG. 13 – Representation of a supercoiled circular DNA, for different par-
titions of the linking number ∆Lk between supplementary twist ∆Tw, as
measured from the natural coiling or torsion Lk0
� Tw0, and writhe W r.
In the figure, the plasmid is modelled as a tube with a square section and
one black face, and its extra linking number ∆Lk and twist ∆Tw are
simply denoted here by Lk and Tw.

FIG. 14 – AFM images of DNA of adsorbed pUC19 plasmids. pUC19 with
5 mM MgCl2 buffer. a) Absence of Dau anthracycline, W r � 0. b) 3.6 µM
Dau : W r � 0. c) 37 µM Dau :W r � 0. Scale : 500 nm. [V. Viglasky et al.,
Electrophoresis 24, 1703-1711 (2003).] Courtesy of G. Dietler, EPFL.

TWISTING DNA :EXPERIMENTS
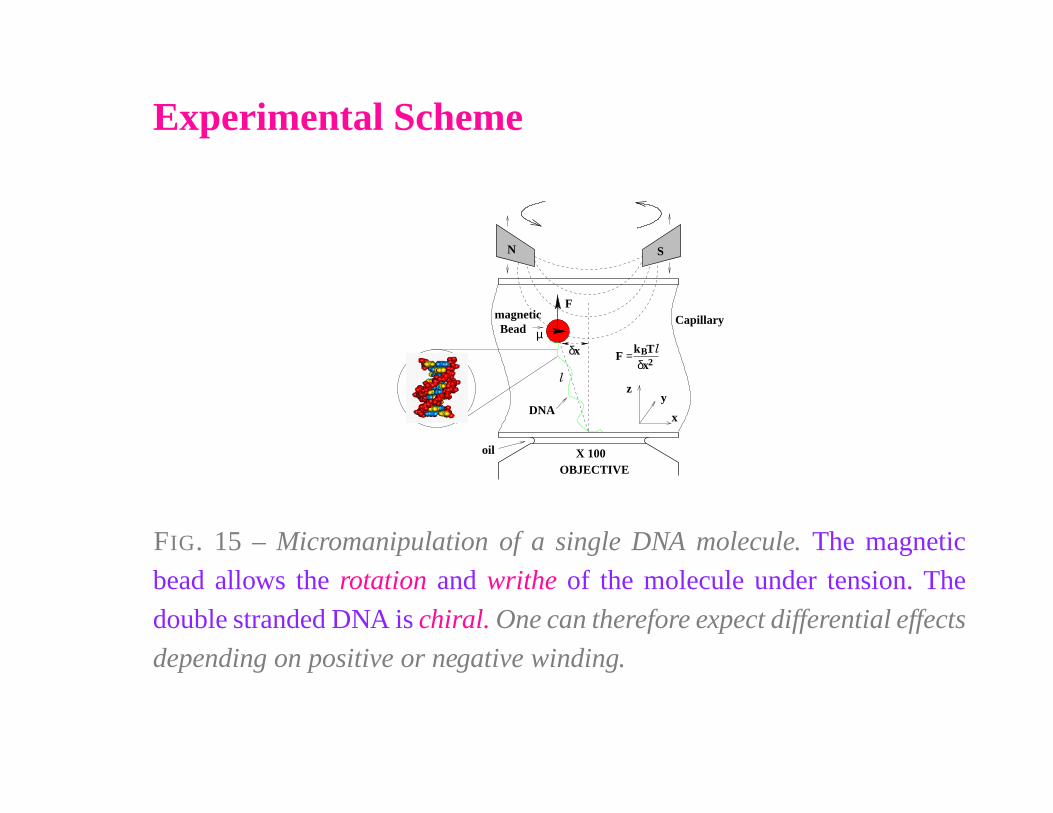
Experimental Scheme
F = k TB lδx2
N S
δx
l
µ
DNA
magneticBead
F
x
zy
oil
Capillary
X 100OBJECTIVE
FIG. 15 – Micromanipulation of a single DNA molecule. The magnetic
bead allows the rotation and writhe of the molecule under tension. The
double stranded DNA is chiral. One can therefore expect differential effects
depending on positive or negative winding.
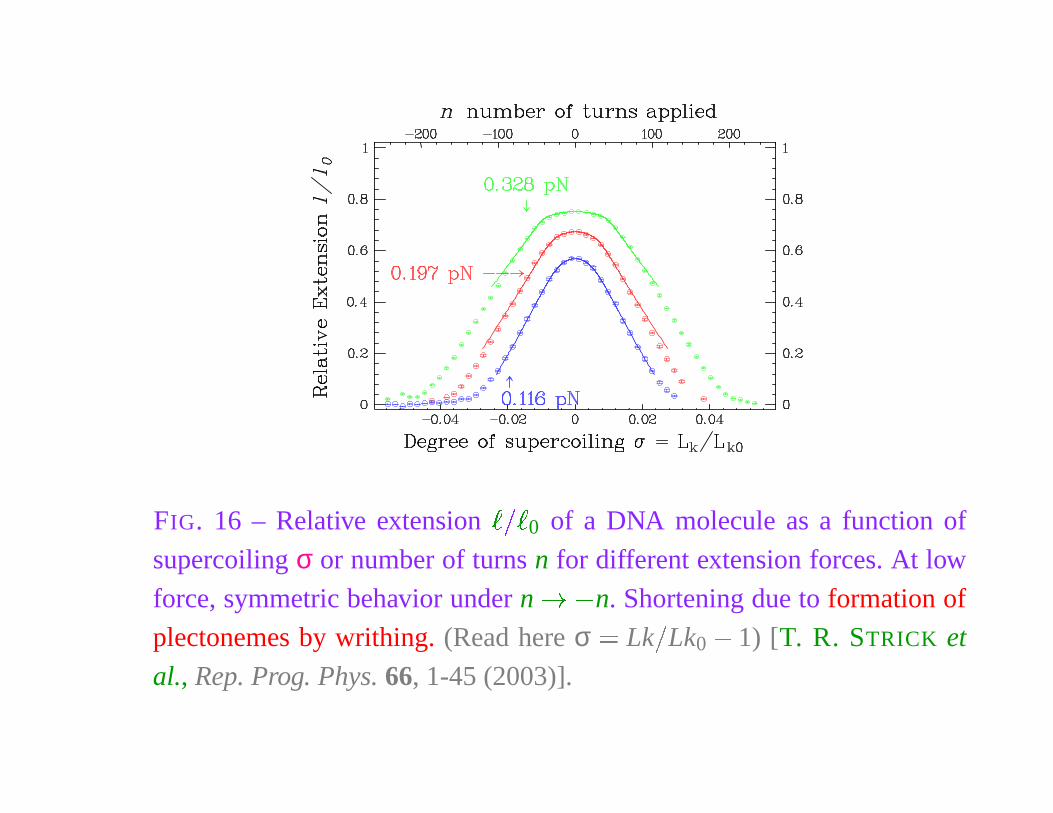
FIG. 16 – Relative extension� � �
0 of a DNA molecule as a function of
supercoiling σ or number of turns n for different extension forces. At low
force, symmetric behavior under n � � n. Shortening due to formation of
plectonemes by writhing. (Read here σ � Lk
�
Lk0
� 1) [T. R. STRICK et
al., Rep. Prog. Phys. 66, 1-45 (2003)].

DNA TWIST
& THERMAL FLUCTUATIONS

Twist Energy
Tw number of turns about tangent vector
�
t
�
s
�
, or rotationangle
Θ � 2πTw
Relaxed DNA Values :
Lk0
� Tw0
��
0
h; Θ0
� 2πTw0
� 2π
�
0
h
�
h helical pitch (h � 3 � 6 nm, 10 � 4 bp for standard DNA).
Twist Elastic Energy :
βHTw
� 12
C�
0
�Θ � Θ0
� 2 �
C torsional modulus
�

Twist Elastic Energy
Lk � Tw
�
Wr � Θ � 2πTw
σ � ∆LkLk0
� Lk � Lk0
Lk0
� Lk0� Tw0 � Wr0
� 0
Θ � Θ0
� 2π
�
Lk0σ � Wr
�
� 2π
�
0
hσ � Wr
βHTw
� 12
C
�
0
�
Θ � Θ0� 2
� βE�
σ�
� b
�
σ
�
2πWr
�
c0
�
2πW r
� 2 �
βE
�
σ
�
� 12
�2π
� 2C
�
0
h2 σ2 � b
�
σ
�
� 2πCh
σ � c0
� 12
C
�
0
�

Writhe (F.B. Fuller, 1978)
t(s)zuC
A
FIG. 17 – Trajectory C of
�
t
�
s
�
on the unit sphere for periodic boundary
conditions :
�
t
�
0
�
��
t
� �
0
�� Area A : solid angle as seen from O.
Fuller’s formula :2πWr � A�

Auxiliary vector potential at
�
r � 3
�
A
� �
r
�
�
�
uz
� �
rr
�
r
� �
uz
� �
r
� � ∇ �
r
�
�
A
� �
r
�
�
�rr3
�
2πW r � A �
S
�
rr3
� d�
S �
C
�
A
� �
t
� � d
�
t
�
�
0
0ds
�uz
� �
t�
s
�
1� �
uz
��
t
�
s
� �
d
�
t
�
s
�
ds
�
[N. B. : Vector potential
�
A� �
r�
defined for r
� �
uz
��
r � 0, undefined for
z � �� ∞ � 0
�
, where r
� �
uz�
�r � 0 ; corresponds to a Dirac magnetic
monopole. ]
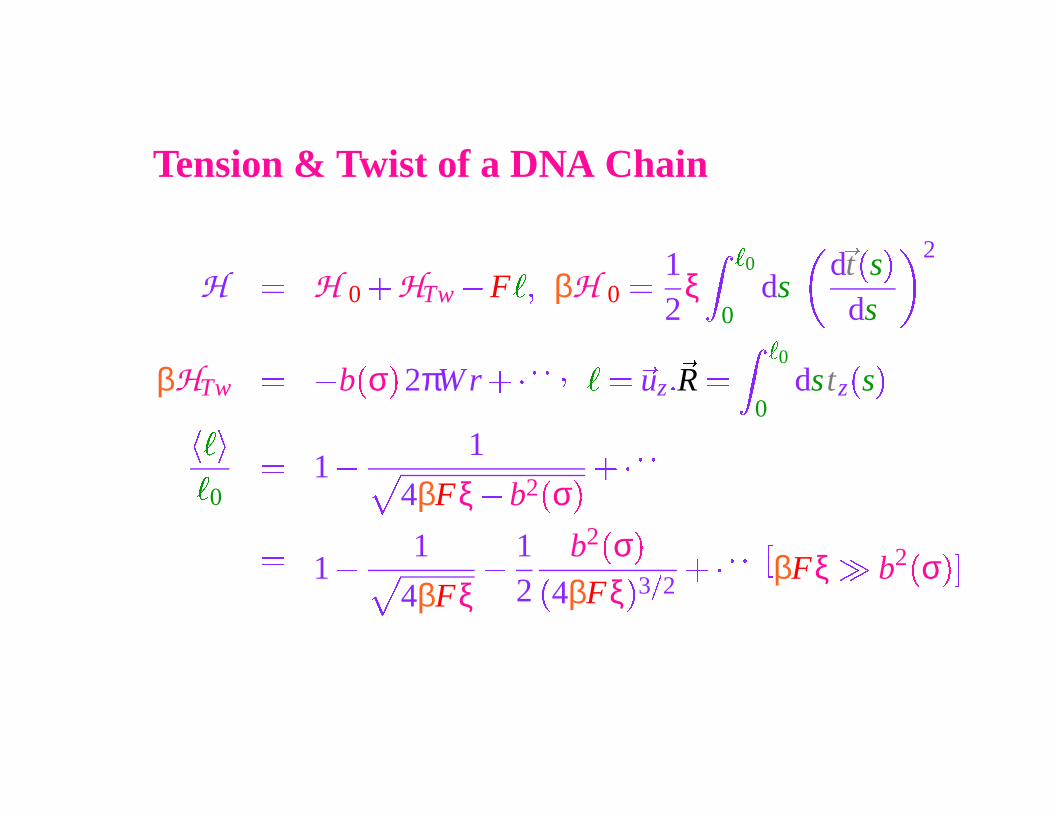
Tension & Twist of a DNA Chain
H � H 0
� HTw
� F
�� βH 0
� 12
ξ�
0
0ds
d
�
t
�
s
�
ds
2
βHTw
� � b
�
σ
�
2πW r
��� � � �
� ��
uz ��
R �
�
0
0dstz
�
s
�
� � ��
0
� 1 �
1
4βFξ � b2�
σ�
��� � �
�
1 �
1
4βFξ
�12
b2 �
σ
�
�
4βFξ
�
3
�
2
��� � �
�
βFξ b2 �
σ
� �
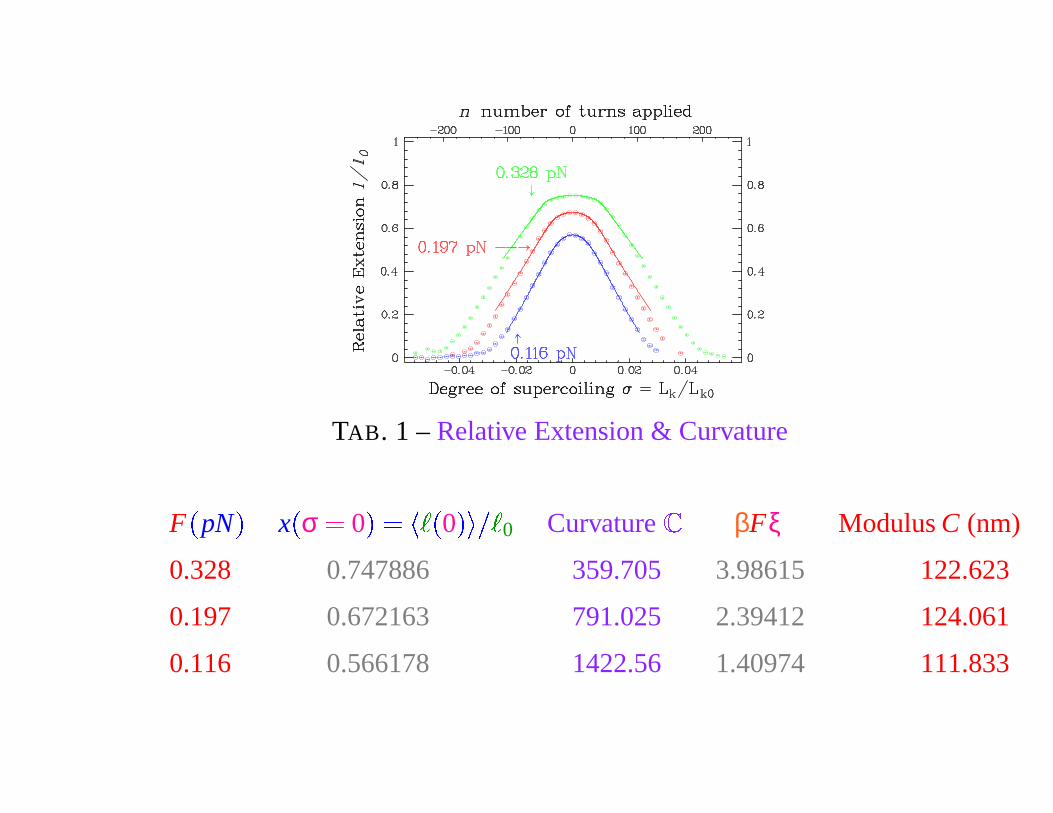
TAB. 1 – Relative Extension & Curvature
F
�
pN
�
x
�
σ � 0
�
�� � �
0
� � � �0 Curvature
�
βFξ Modulus C (nm)
0.328 0.747886 359.705 3.98615 122.623
0.197 0.672163 791.025 2.39412 124.061
0.116 0.566178 1422.56 1.40974 111.833

Curvature & Torsional Modulus
� � � � �
0
� x
�
σ
�
� x
�
0
�
�
�
σ2 ��� � �
� � 116
2πCh
2 1
�
βFξ
�
3
�
2
C � h2π
�
16
� �
βFξ� 3
�
2� 1
�2
�
One first finds ξ � 50 nm. The torsional modulus, which is also the
persistence length for the thermal fluctuations of the torsional angle Θ, is
C � 100 nm. New experiments by the team of D. BENSIMON, V.
CROQUETTE & J.-F. ALLEMAND, (Physique des biomolécules,
LPS-ENS), give the value C � 94
�
7 nm. [F. Mosconi et al., Torque
Determination on DNA with Magnetic Tweezers, arXiv : 0809.0621, Phys.
Rev. Lett., to appear (2009).]

Average Writhe
2π
�
W r
�
�
�
0
2ξb
�
σ
�
21
4βFξ � b2�
σ�
��� � �
�
�
0
2ξb
�
σ
�
4βFξ1
� 12
b2 �
σ
�
4βFξ
��� � � �
βFξ b2 �
σ
� �

Average Torque
β
�
Γ
�
� b
�
σ
�
�
C
�
02π
�
Wr
�
� b
�
σ
�
�
C2ξ
b
�
σ
�
4βFξ1
� 12
b2 �
σ�
4βFξ
� � � � �
To leading order :
β
�
Γ
�
� b
�
σ
��
1 �
C4ξ
1
βFξ�
As b
�
σ
�
� 2πCh σ, one finds a renormalization of the torsional modulus C
into Ceff [J.D. Moroz & P. Nelson, Proc. Nat. Acad. Sci. USA 94, 14418
(1997)]
Ceff� C
�
1 �
C4ξ
1
βFξ
�
�
The torsion rigidity diminishes for weaker force, with plectonemes
induced by thermal fluctuations.
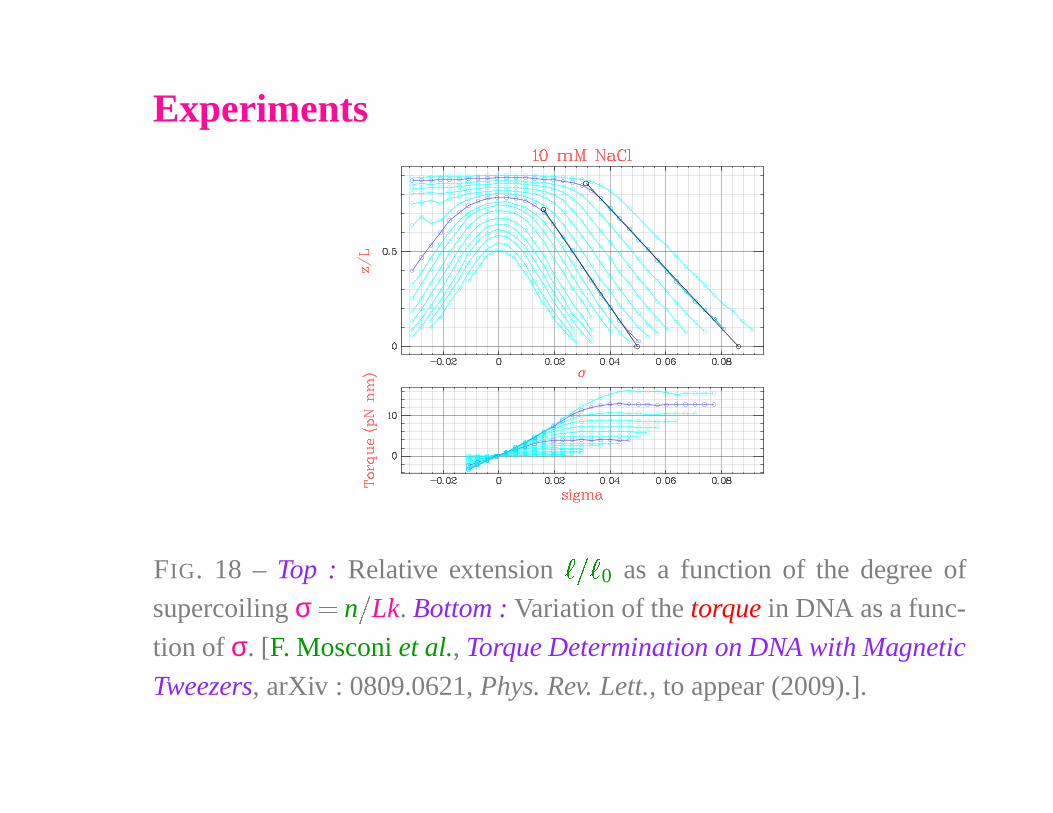
Experiments
FIG. 18 – Top : Relative extension
� � �
0 as a function of the degree of
supercoiling σ � n
�
Lk. Bottom : Variation of the torque in DNA as a func-
tion of σ. [F. Mosconi et al., Torque Determination on DNA with Magnetic
Tweezers, arXiv : 0809.0621, Phys. Rev. Lett., to appear (2009).].
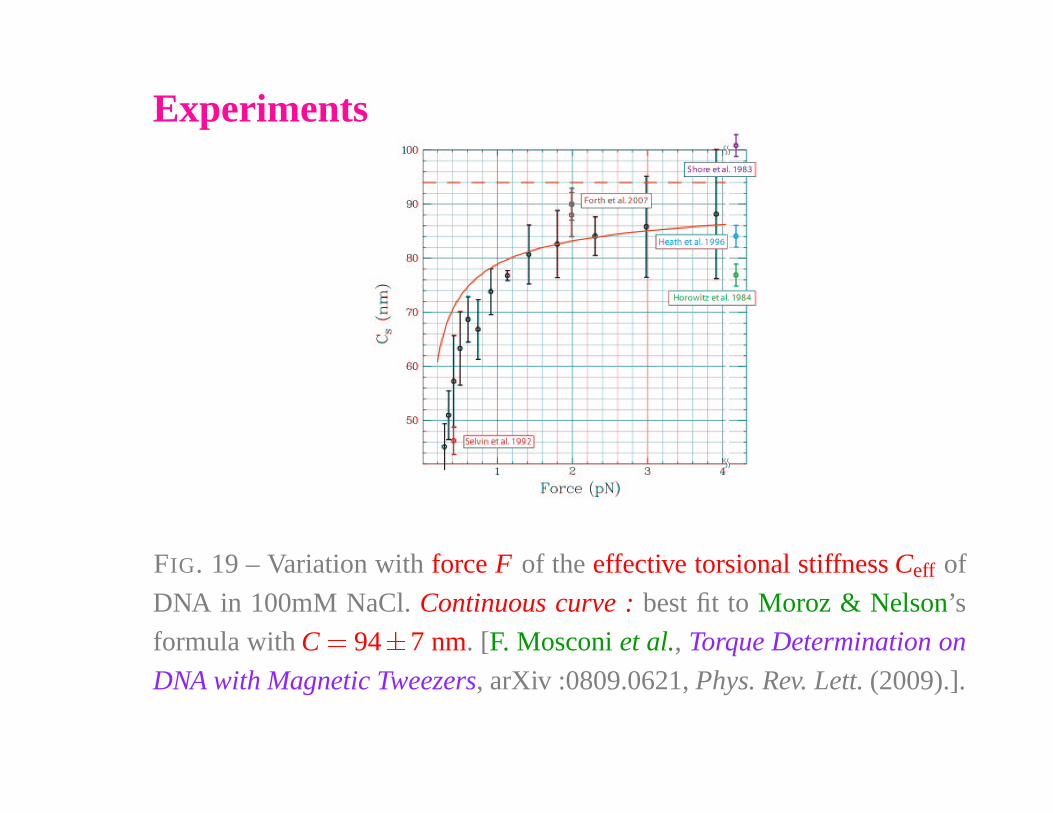
Experiments
FIG. 19 – Variation with force F of the effective torsional stiffness Ceff of
DNA in 100mM NaCl. Continuous curve : best fit to Moroz & Nelson’s
formula with C � 94�
7 nm. [F. Mosconi et al., Torque Determination on
DNA with Magnetic Tweezers, arXiv :0809.0621, Phys. Rev. Lett. (2009).].
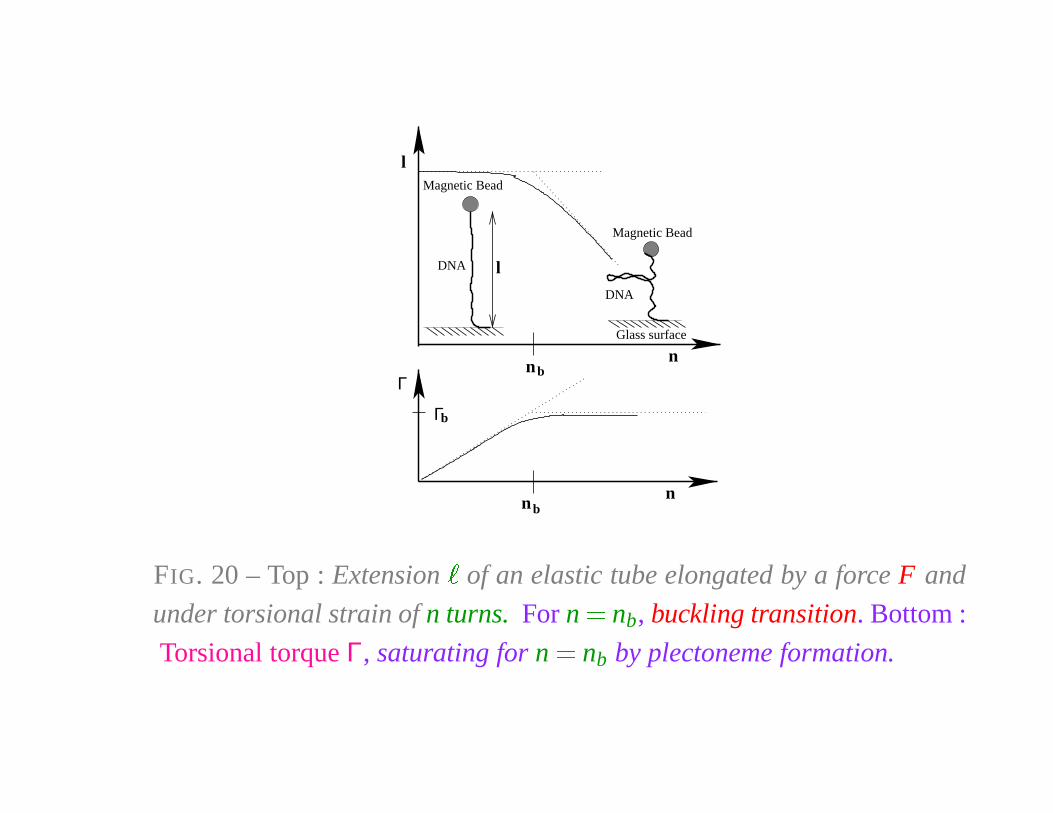
Magnetic Bead
DNA l
DNA
Magnetic Bead
Glass surface
Γ
nnb
Γb
bn
l
n
FIG. 20 – Top : Extension�
of an elastic tube elongated by a force F and
under torsional strain of n turns. For n � nb, buckling transition. Bottom :
Torsional torque Γ, saturating for n � nb by plectoneme formation.
Buckling Transition
�
�
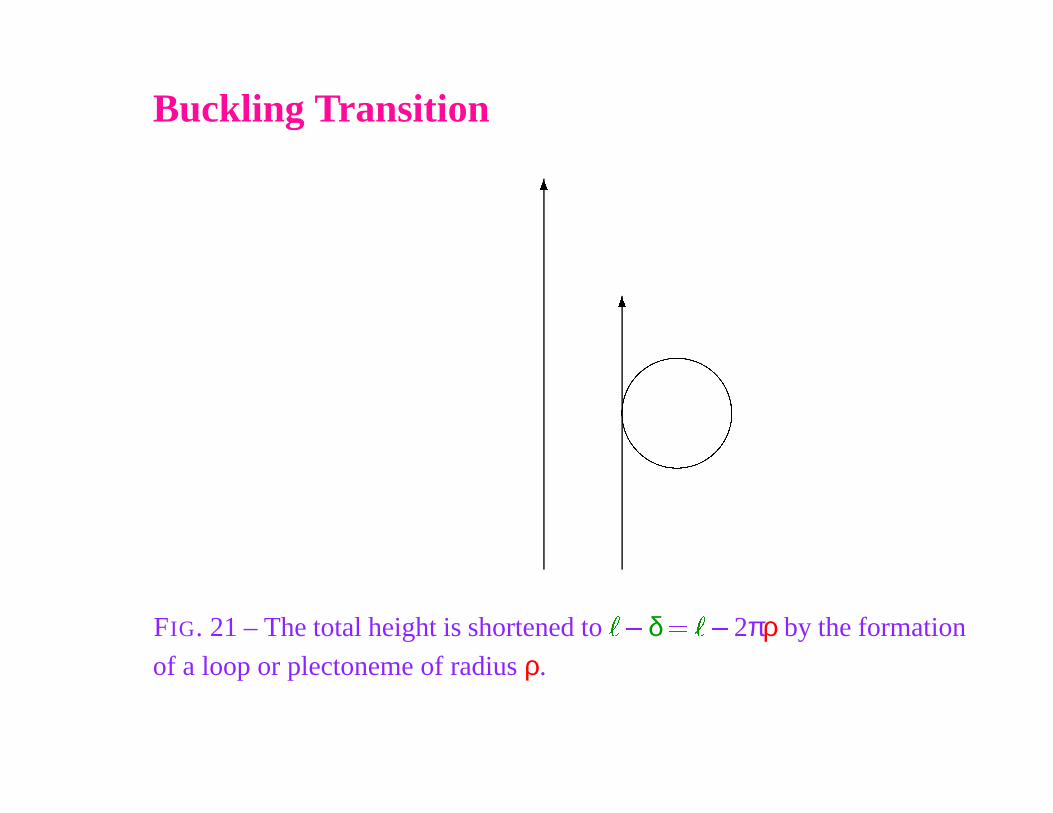
FIG. 21 – The total height is shortened to
�� δ �
�� 2πρ by the formation
of a loop or plectoneme of radius ρ.

Semi-Flexible ChainCurvature Hamiltonian in presence of a pulling force F in the Oz direction
1kBT
H � βH � 12
ξ
�
0
0ds
d
�
t
�
s
�
ds
2
� βF�
�
ξ persistence length and
�
chain elongation along Oz.
Kinematics :d
�
t
�
s
�
ds�
�
n�
s�
ρ�
�
n
�
s
�
unit normal vector, ρ curvature radius. For a circular plectoneme of
radius ρ, tangent to the DNA stretched part, a loop contribution to the
curvature energy
δH0
�
ρ
�
� ξ2β
2πρ
�

Contribution
�
Fδ of shortening δ � 2πρ in the force field :
δH
�
ρ
�
� ξπ
�
βρ �
2πρF �
Equilibrium value ρc of ρ such that δH
�
ρ
�
is minimal :
ρc
��
ξ
�
2βF
� 1
�
2 �
Bead displacement under plectoneme formation :
δc
� 2πρc� 2πξ
12βFξ
1
�
2
�
Numerical data. For F � 1 pN, and a typical DNA persistence length
ξADN
� 53 nm � δc
� 65 nm.
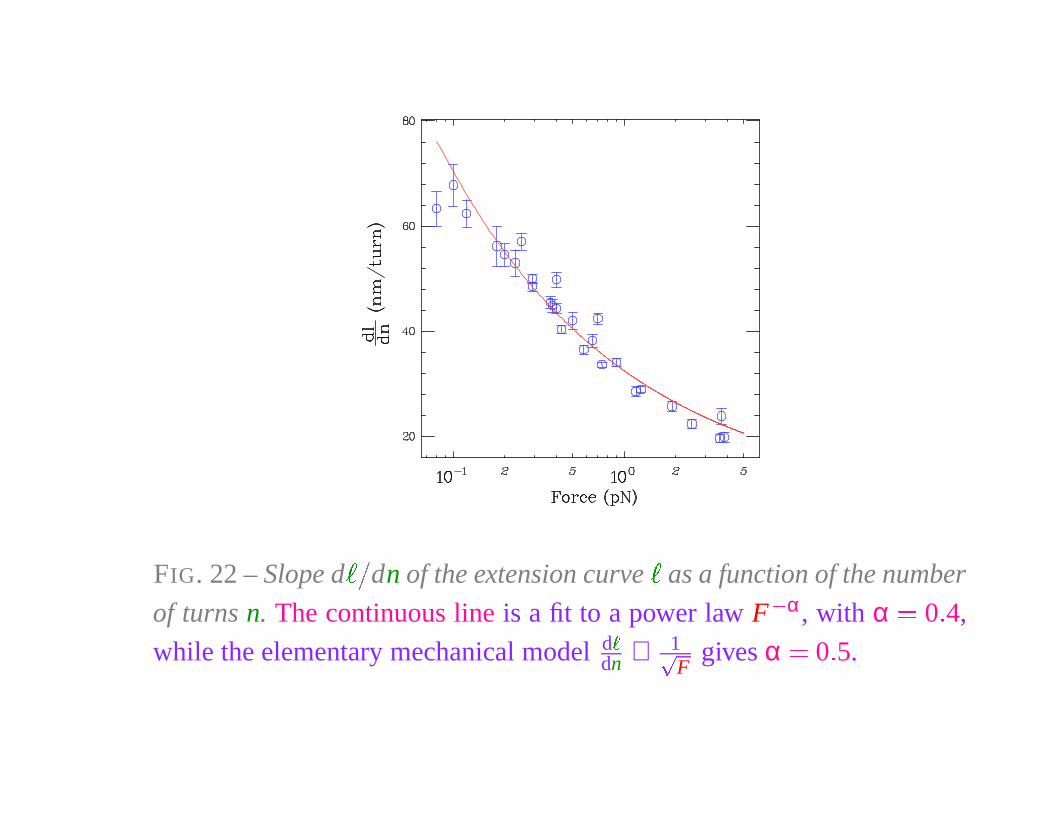
FIG. 22 – Slope d
� �
dn of the extension curve
�
as a function of the number
of turns n. The continuous line is a fit to a power law F
� α, with α � 0 � 4,
while the elementary mechanical model d
�
dn ∝ 1�
Fgives α � 0 � 5.
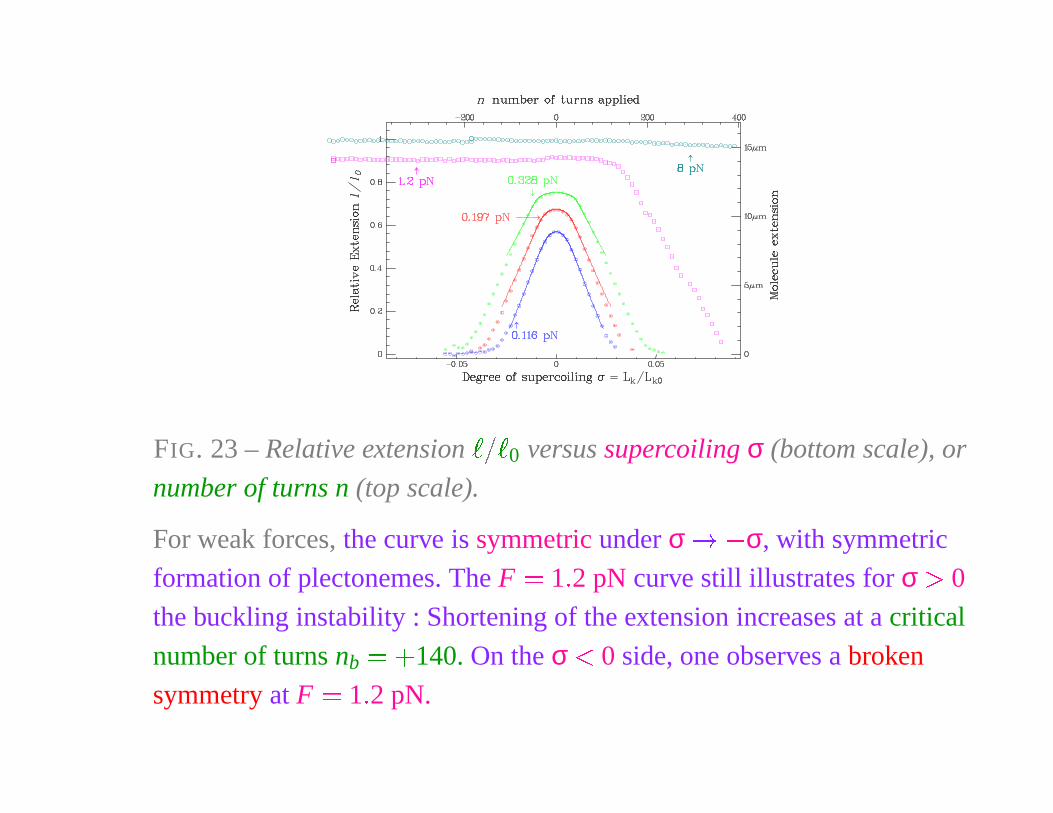
FIG. 23 – Relative extension
� � �
0 versus supercoiling σ (bottom scale), or
number of turns n (top scale).
For weak forces, the curve is symmetric under σ � � σ, with symmetric
formation of plectonemes. The F � 1 � 2 pN curve still illustrates for σ � 0
the buckling instability : Shortening of the extension increases at a critical
number of turns nb� �
140. On the σ � 0 side, one observes a broken
symmetry at F � 1 � 2 pN.
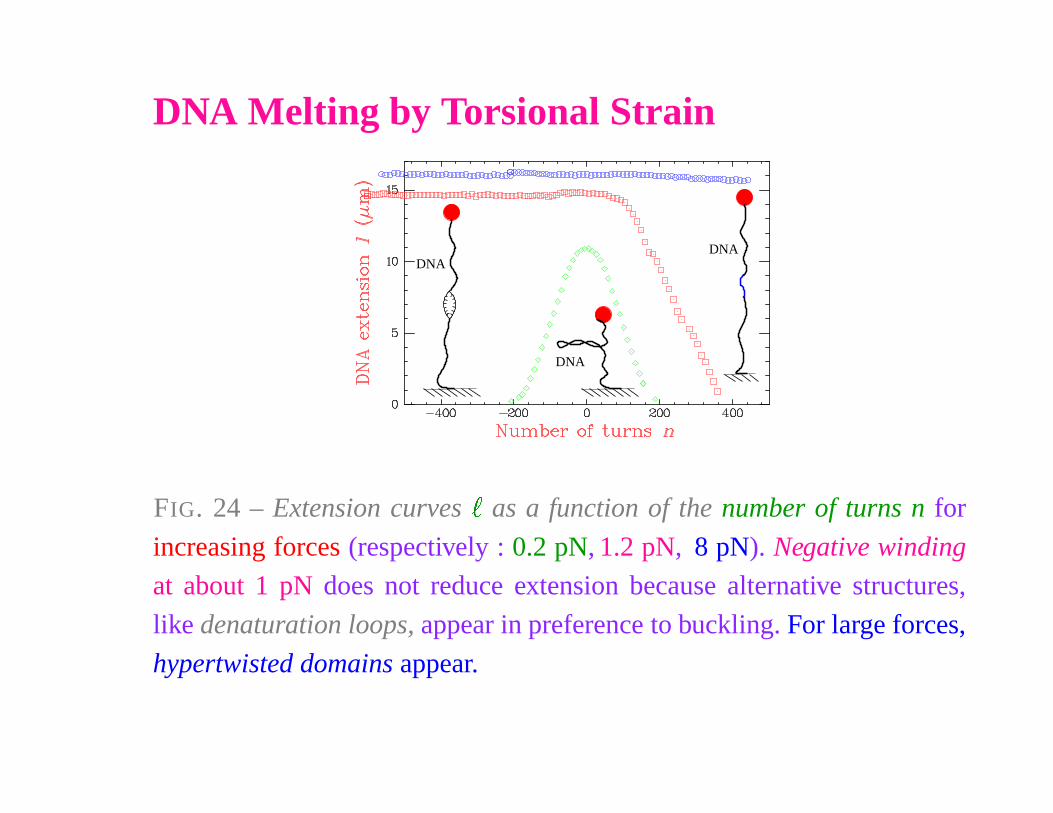
DNA Melting by Torsional Strain
DNA
DNA
DNA
FIG. 24 – Extension curves
�
as a function of the number of turns n for
increasing forces (respectively : 0.2 pN, 1.2 pN, 8 pN). Negative winding
at about 1 pN does not reduce extension because alternative structures,
like denaturation loops, appear in preference to buckling. For large forces,
hypertwisted domains appear.
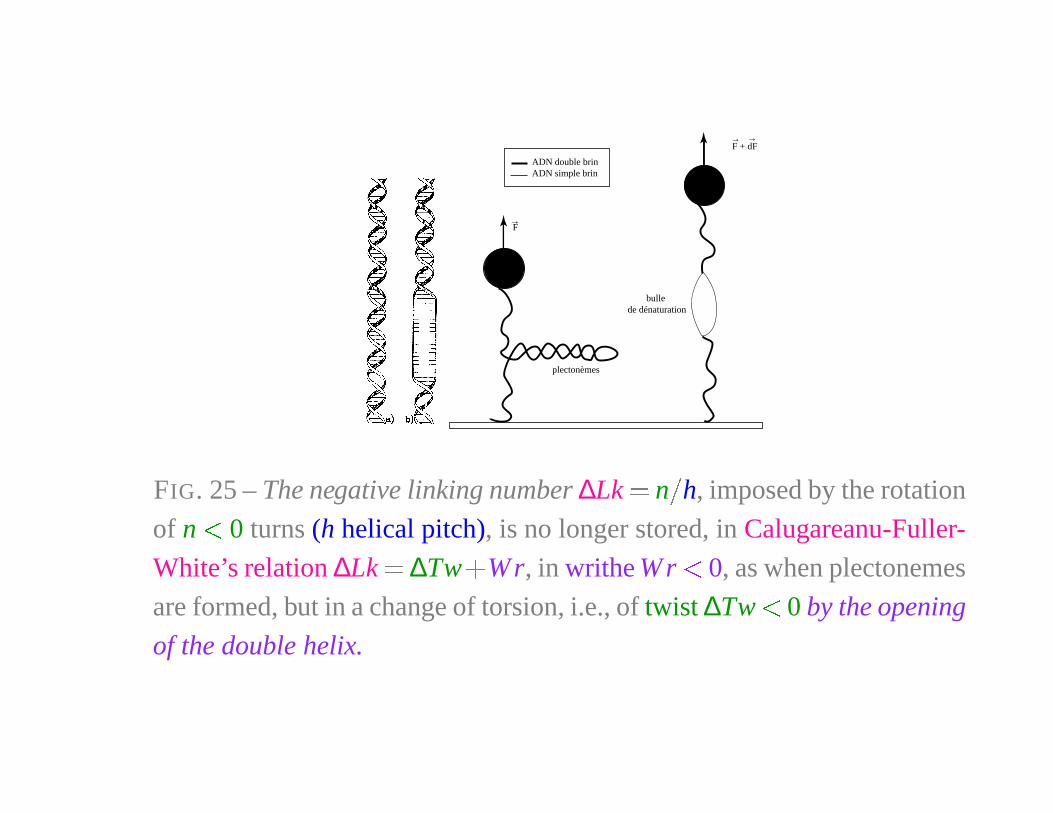
ADN double brinADN simple brin
F + dF→ →
bulle de dénaturation
plectonèmes
F→
FIG. 25 – The negative linking number ∆Lk � n
�
h, imposed by the rotation
of n � 0 turns (h helical pitch), is no longer stored, in Calugareanu-Fuller-
White’s relation ∆Lk � ∆Tw�
Wr, in writhe W r � 0, as when plectonemes
are formed, but in a change of torsion, i.e., of twist ∆Tw � 0 by the opening
of the double helix.
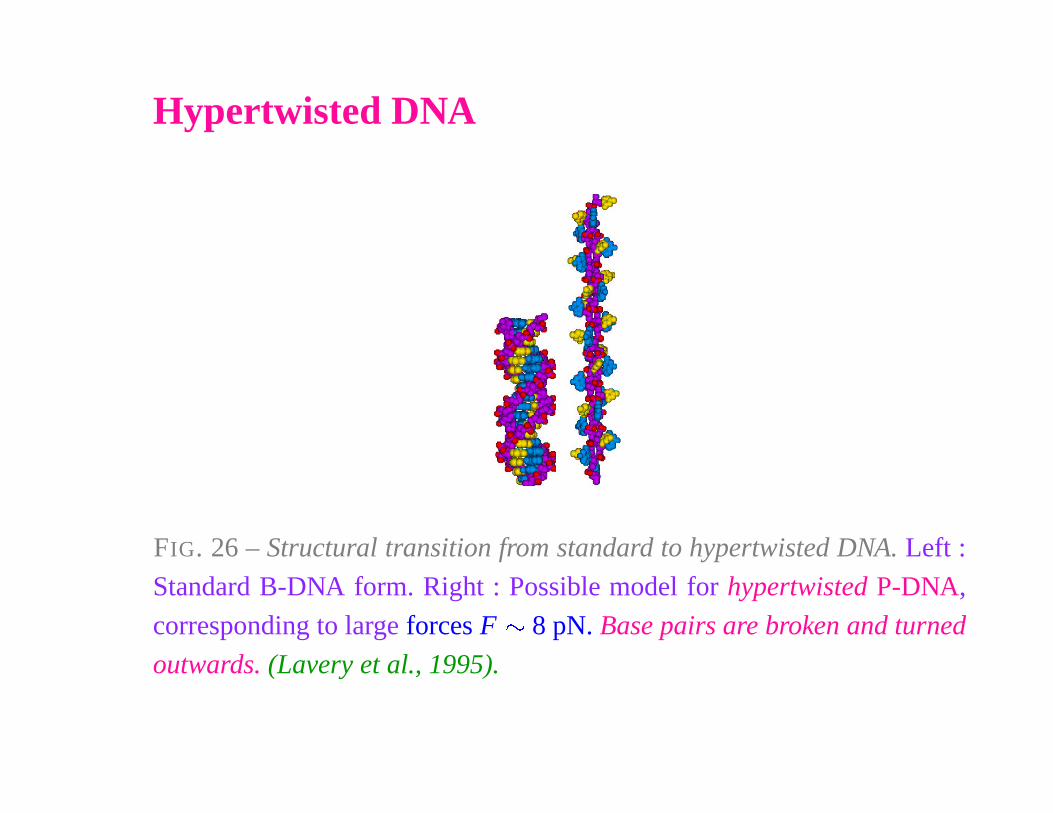
Hypertwisted DNA
FIG. 26 – Structural transition from standard to hypertwisted DNA. Left :
Standard B-DNA form. Right : Possible model for hypertwisted P-DNA,
corresponding to large forces F � 8 pN. Base pairs are broken and turned
outwards. (Lavery et al., 1995).

DNA & TOPOISOMERASES

Knots in DNA
How can two replicated DNAs be separated ? Their mutual linking
number Lk is conserved in replication, thus equal to the linking number of
the two duplex strands before replication, Lk0
� N�
h, where N is the
number of base pairs and h the helical pitch.
Discovery by J.C. Wang a of a family of enzymes, the topoisomerases,
able to modify the topology of DNA molecules by cleaving and resealing
one or two DNA strands (Topo I or Topo II). Topoisomerases are able to
alter DNA supercoiling, the linking number of two molecules, or to
(un)knot a circular DNA. After replication, the daughter DNA chains are
highly linked together. Hence the simultaneous action of multiple
topoisomerases is required during mitosis.
aJ.C. Wang, J. Mol. Biol. 55, 523-533 (1971).
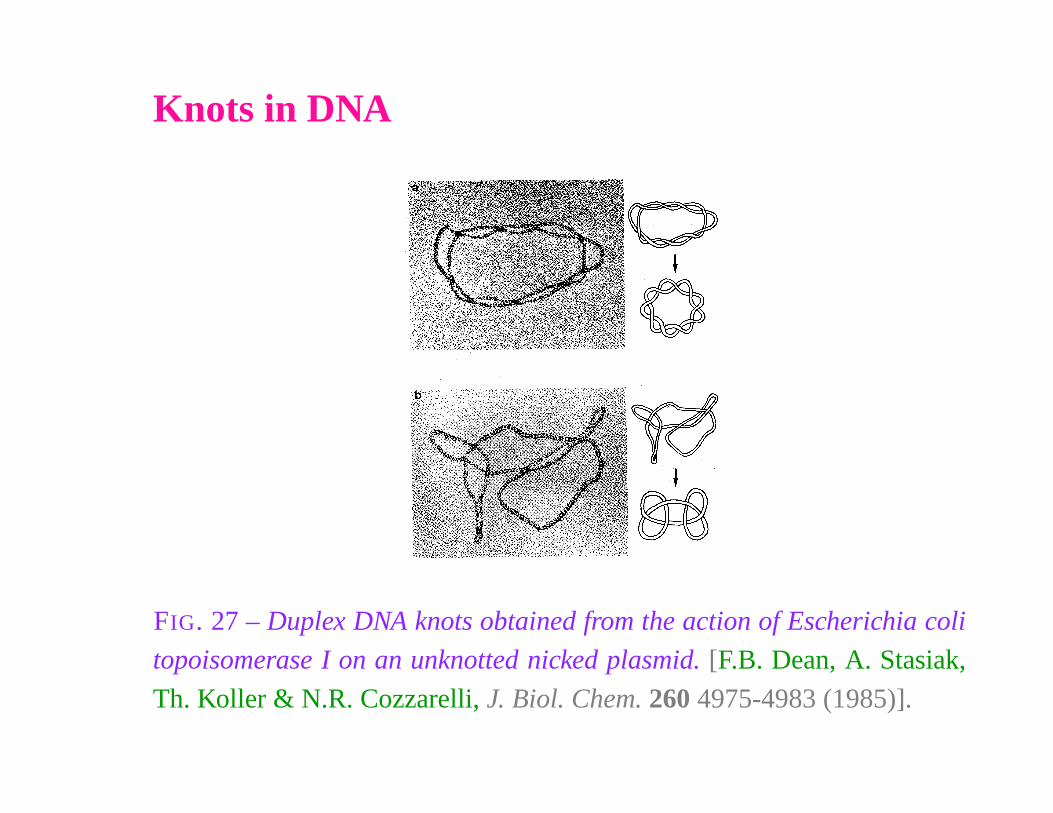
Knots in DNA
FIG. 27 – Duplex DNA knots obtained from the action of Escherichia coli
topoisomerase I on an unknotted nicked plasmid. [F.B. Dean, A. Stasiak,
Th. Koller & N.R. Cozzarelli, J. Biol. Chem. 260 4975-4983 (1985)].

FIG. 28 – AFM images of circular DNA of P4 phage (10346 base pairs,
�
0
� 3 � 5µm). Left column : Strong adsorption, 3D configurations ; Right
column : Weak adsorption on mica, 2D configurations. Top : Simple knots
with essential crossing numbers < 6. Bottom : Complex knots with essen-
tial crossing numbers between 30 and 40. [E. Ercolini et al., Phys. Rev.
Lett. 98, 058102 (2007).] (Courtesy of G. Dietler, EPFL.)

FIG. 29 – AFM images of circular DNA of P4 phage, weakly adsorbed in
two dimensions on mica. Top left : Unknotted configuration. In all other
images the circular DNAs are knotted. For entropic reasons, the knots ap-
pear as localized along the chains. Images from E. Ercolini et al., Phys.
Rev. Lett. 98, 058102 (2007). (Courtesy by G. Dietler, EPFL.)
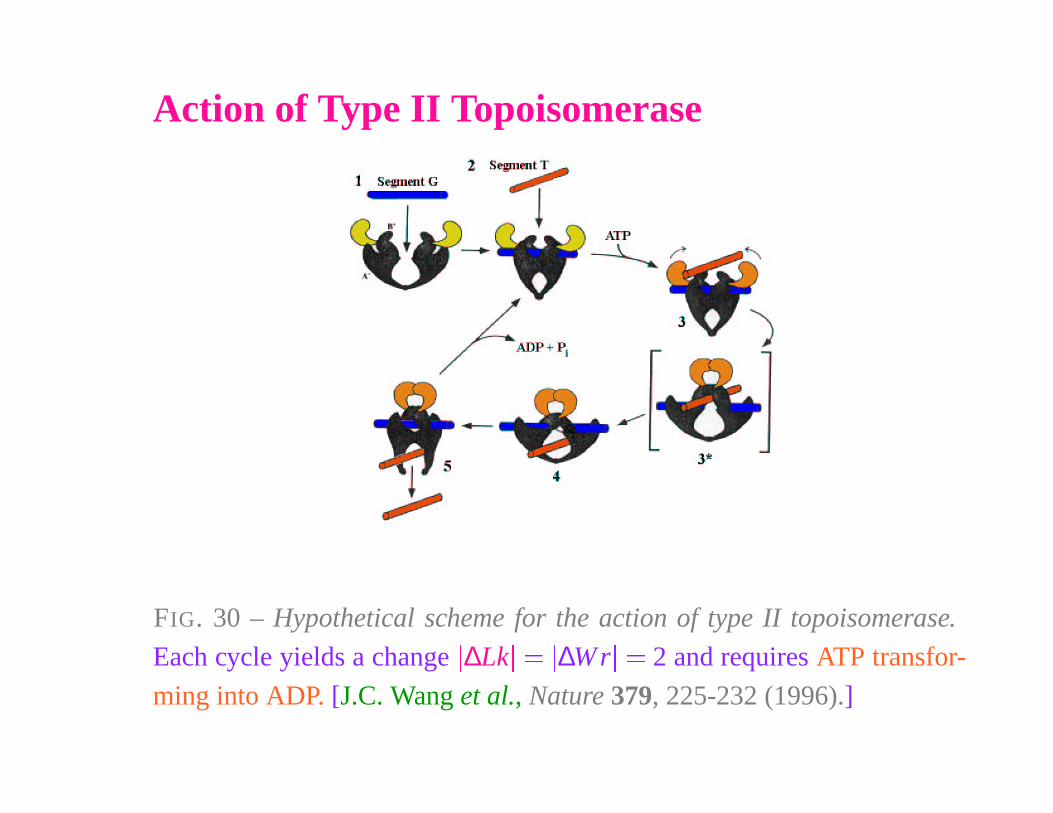
Action of Type II Topoisomerase
FIG. 30 – Hypothetical scheme for the action of type II topoisomerase.
Each cycle yields a change
�
∆Lk
�
��
∆W r
�
� 2 and requires ATP transfor-
ming into ADP. [J.C. Wang et al., Nature 379, 225-232 (1996).]
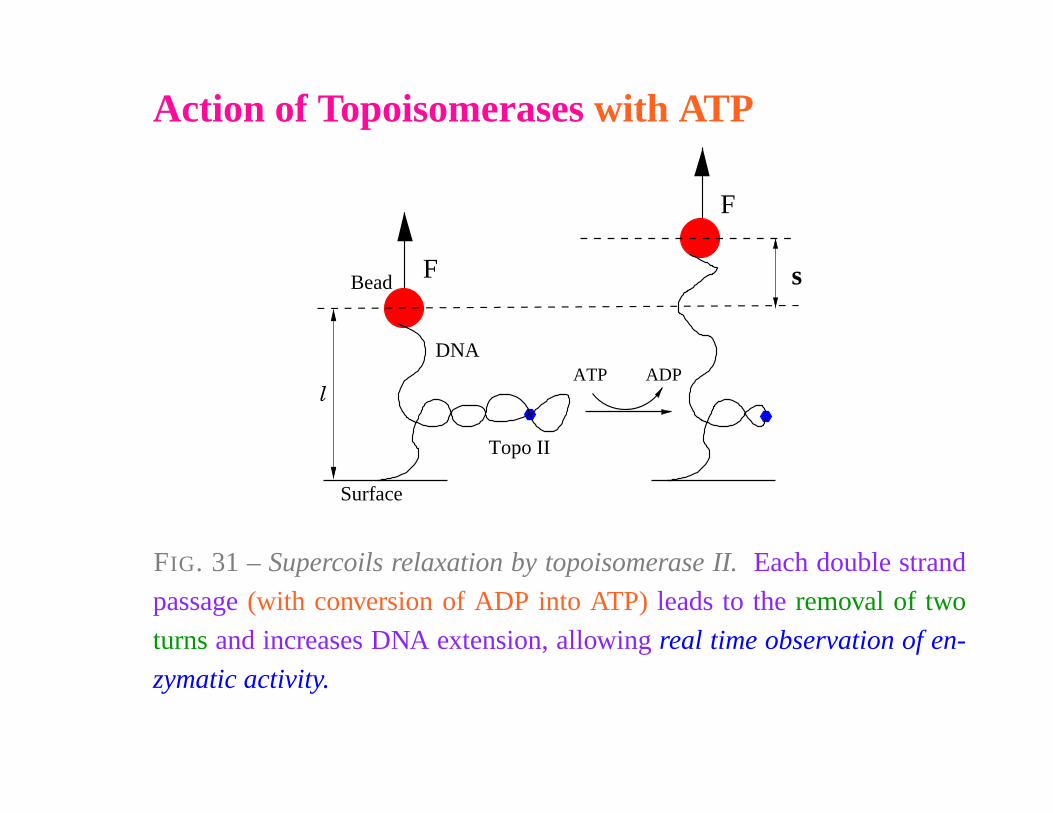
Action of Topoisomerases with ATP
F
ATP ADP
F
Surface
Topo II
l
Bead
DNA
s
FIG. 31 – Supercoils relaxation by topoisomerase II. Each double strand
passage (with conversion of ADP into ATP) leads to the removal of two
turns and increases DNA extension, allowing real time observation of en-
zymatic activity.
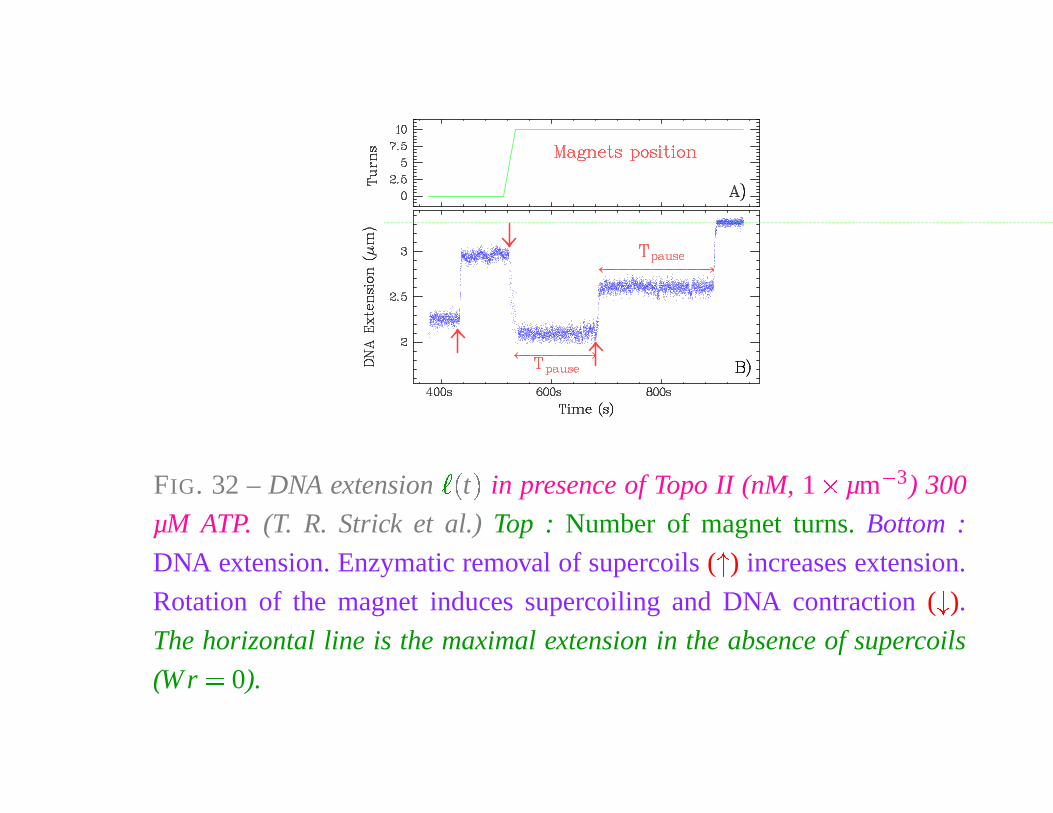
FIG. 32 – DNA extension
� �
t
�
in presence of Topo II (nM, 1 � µm
� 3) 300
µM ATP. (T. R. Strick et al.) Top : Number of magnet turns. Bottom :
DNA extension. Enzymatic removal of supercoils (
�
) increases extension.
Rotation of the magnet induces supercoiling and DNA contraction (
�
).
The horizontal line is the maximal extension in the absence of supercoils
(W r � 0).
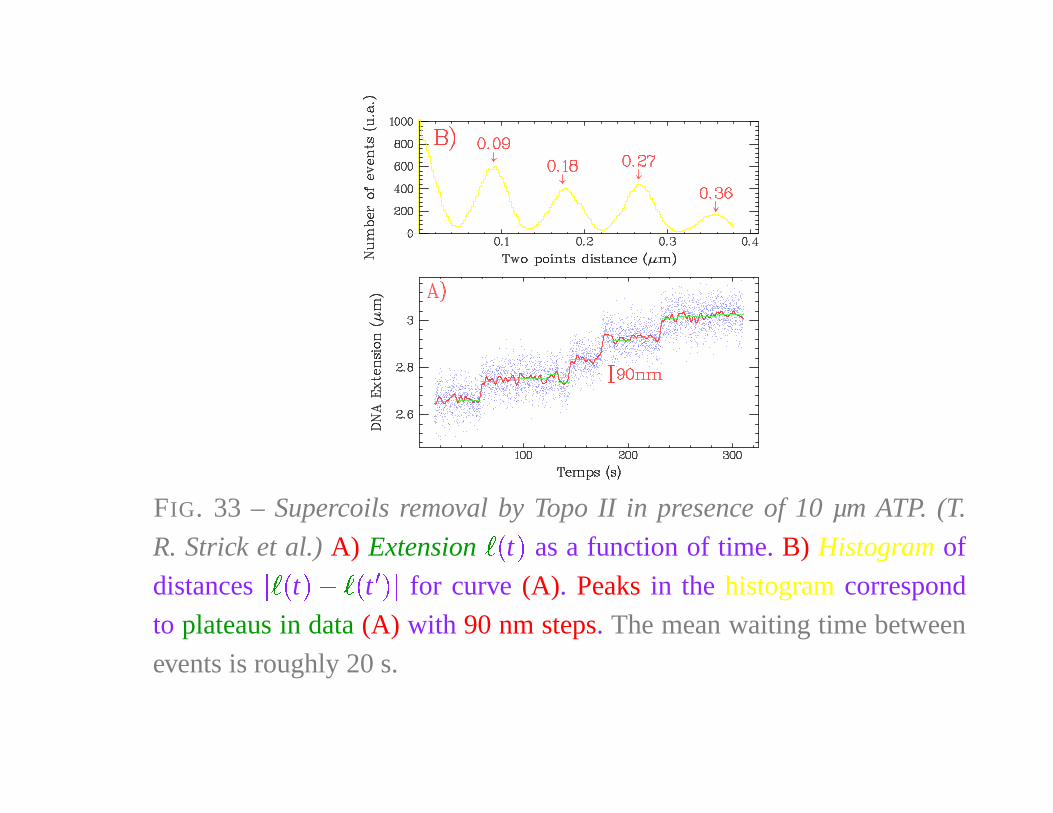
FIG. 33 – Supercoils removal by Topo II in presence of 10 µm ATP. (T.
R. Strick et al.) A) Extension� �
t�
as a function of time. B) Histogram of
distances
� � �
t
�
�� �
t
� � �
for curve (A). Peaks in the histogram correspond
to plateaus in data (A) with 90 nm steps. The mean waiting time between
events is roughly 20 s.
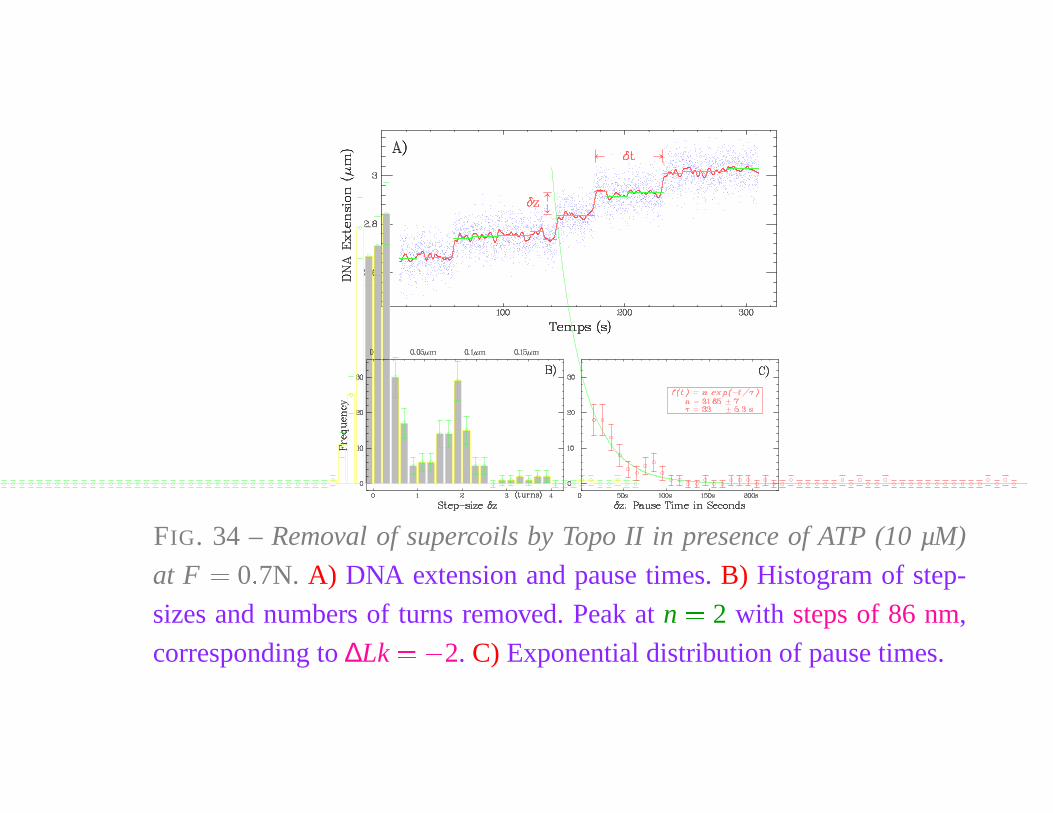
FIG. 34 – Removal of supercoils by Topo II in presence of ATP (10 µM)
at F � 0 � 7N. A) DNA extension and pause times. B) Histogram of step-
sizes and numbers of turns removed. Peak at n � 2 with steps of 86 nm,
corresponding to ∆Lk � � 2. C) Exponential distribution of pause times.

Action of topoisomerases without ATP
FIG. 35 – Traction on plectonemes without Topo II (left) or with Topo II
(right). DNA overwound by n � 60 turns and submitted to a crenel force
from 1pN to 5 pN. In the absence of enzymes, the force removes plecto-
nemes and the extension oscillates between levels (1) (weakly extended)
and (3) (extended). In the presence of enzymes, intermediate extensions
appear (2), due to the protection of plectonemes clamped by Topo II.

Bursting Tractions on Supercoils
l
F = 1 pN
ADN
Bille
F = 5 pN
Surface
Topo II
1. 3.2.
FIG. 36 – Interpretation : When a Topo II clamps a local dsDNA cros-
sing [1.], it momentarily protects a number of supercoils during the next
force increase from 1 to 5 pN [2.]. When the enzymatic clamp relaxes, the
remaining supercoils vanish [3.]
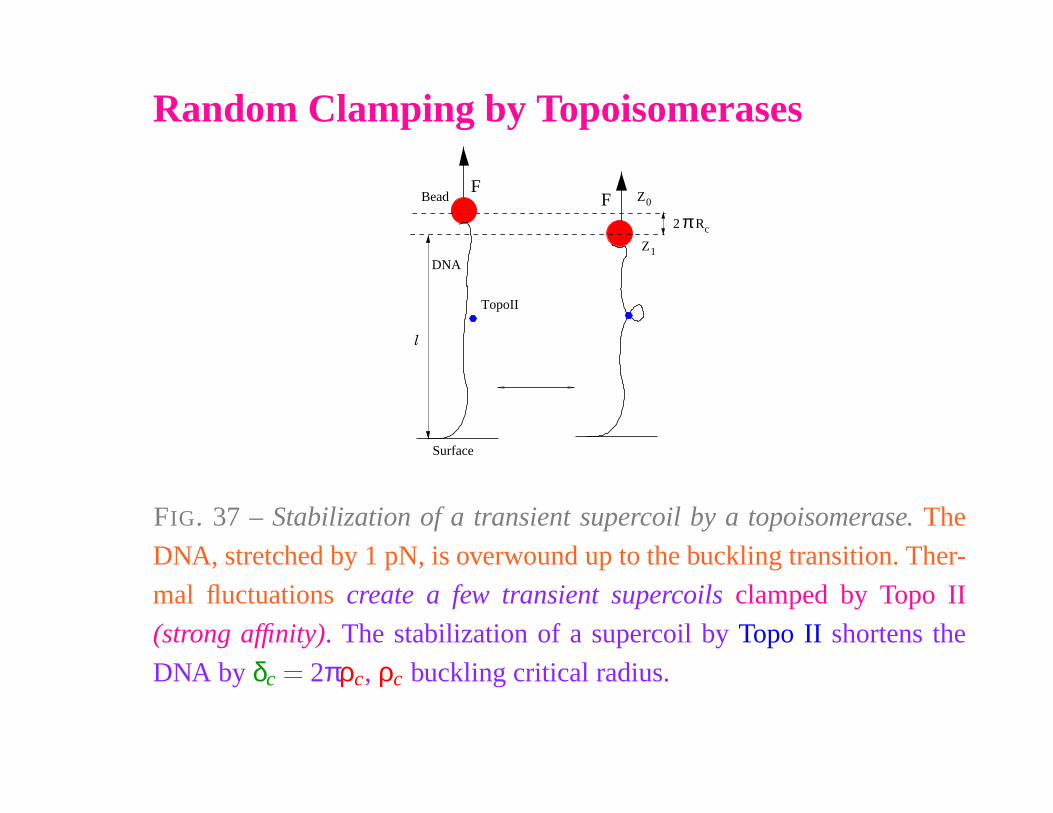
Random Clamping by Topoisomerases
Z0
Z1
cR2 π
Surface
l
F
TopoII
Bead
DNA
F
FIG. 37 – Stabilization of a transient supercoil by a topoisomerase. The
DNA, stretched by 1 pN, is overwound up to the buckling transition. Ther-
mal fluctuations create a few transient supercoils clamped by Topo II
(strong affinity). The stabilization of a supercoil by Topo II shortens the
DNA by δc
� 2πρc, ρc buckling critical radius.
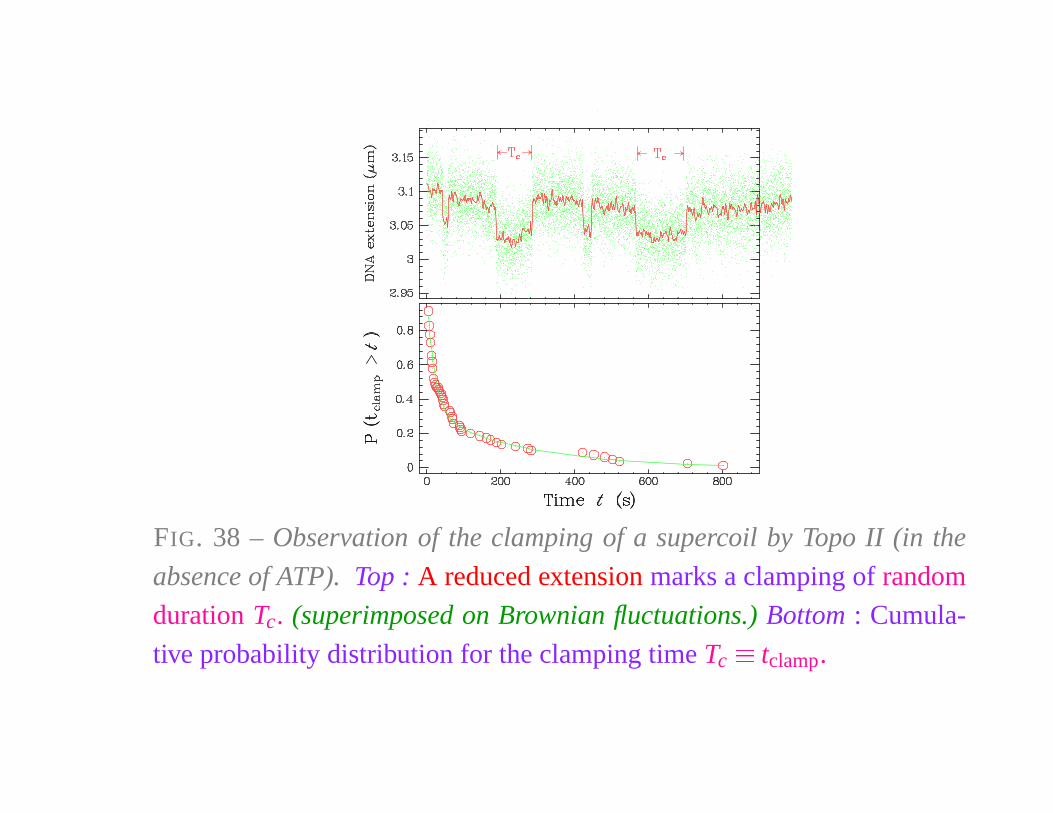
FIG. 38 – Observation of the clamping of a supercoil by Topo II (in the
absence of ATP). Top : A reduced extension marks a clamping of random
duration Tc. (superimposed on Brownian fluctuations.) Bottom : Cumula-
tive probability distribution for the clamping time Tc
� tclamp.
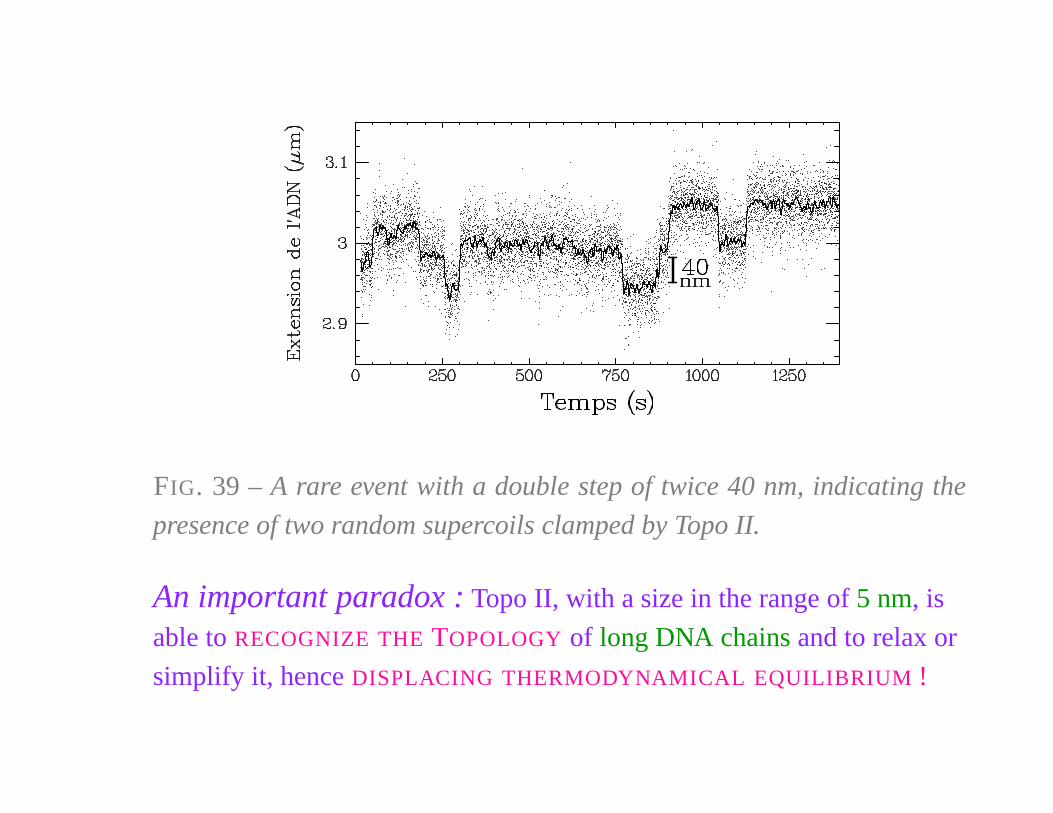
FIG. 39 – A rare event with a double step of twice 40 nm, indicating the
presence of two random supercoils clamped by Topo II.
An important paradox : Topo II, with a size in the range of 5 nm, is
able to RECOGNIZE THE TOPOLOGY of long DNA chains and to relax or
simplify it, hence DISPLACING THERMODYNAMICAL EQUILIBRIUM !

III. RNA UNFOLDING
&
FLUCTUATION RELATIONS
POLYMER PHYSICS & DNA
Troisième Cycle de la physique en Suisse romande
École Polytechnique Fédérale de Lausanne
12 March 2009
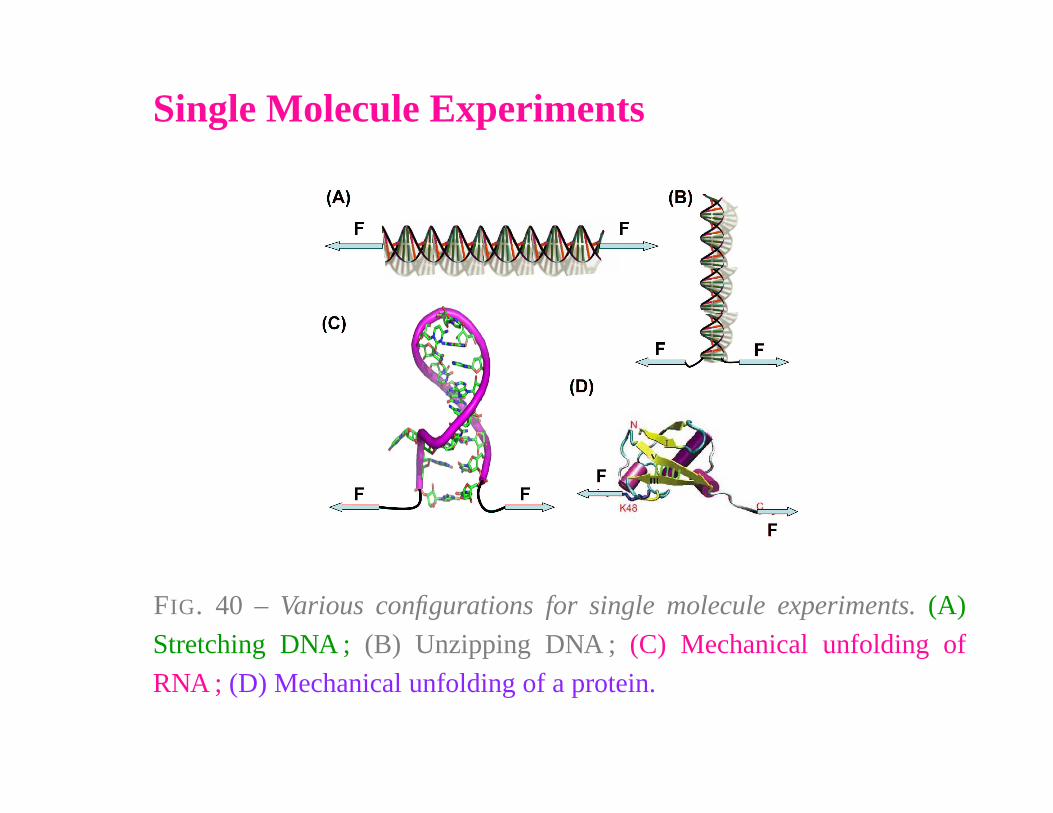
Single Molecule Experiments
FIG. 40 – Various configurations for single molecule experiments. (A)
Stretching DNA ; (B) Unzipping DNA ; (C) Mechanical unfolding of
RNA ; (D) Mechanical unfolding of a protein.
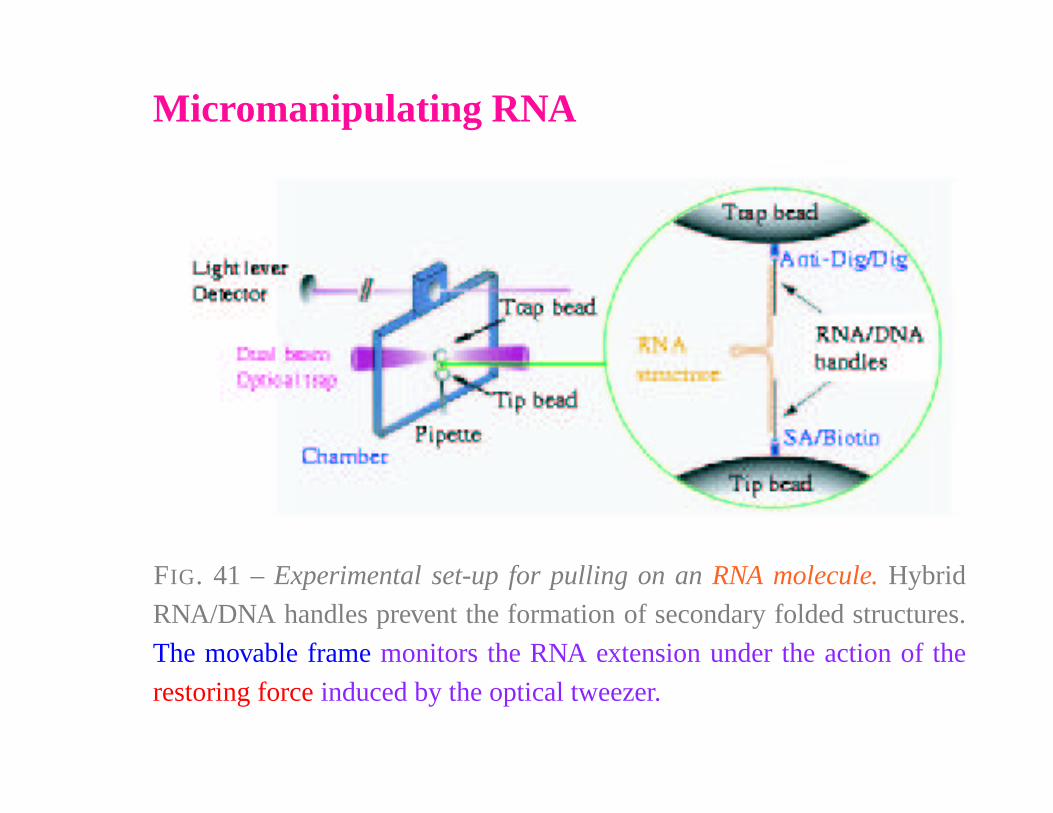
Micromanipulating RNA
FIG. 41 – Experimental set-up for pulling on an RNA molecule. Hybrid
RNA/DNA handles prevent the formation of secondary folded structures.
The movable frame monitors the RNA extension under the action of the
restoring force induced by the optical tweezer.
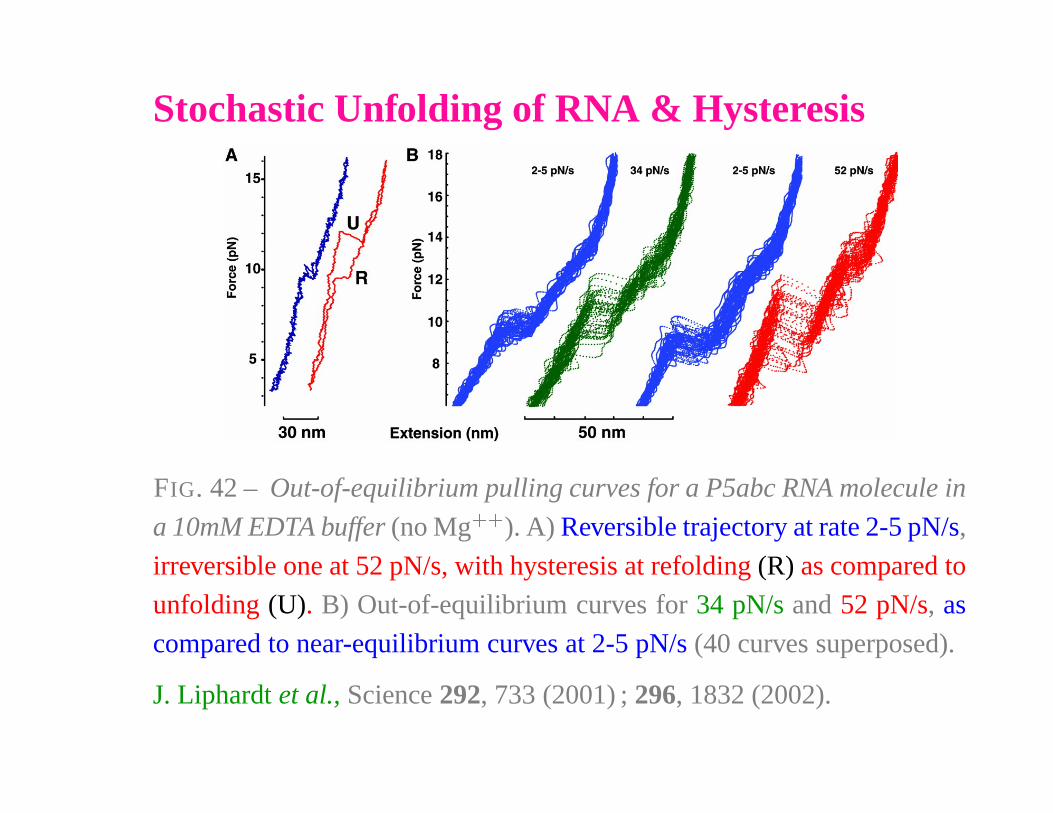
Stochastic Unfolding of RNA & Hysteresis
FIG. 42 – Out-of-equilibrium pulling curves for a P5abc RNA molecule in
a 10mM EDTA buffer (no Mg� �
). A) Reversible trajectory at rate 2-5 pN/s,
irreversible one at 52 pN/s, with hysteresis at refolding (R) as compared to
unfolding (U). B) Out-of-equilibrium curves for 34 pN/s and 52 pN/s, as
compared to near-equilibrium curves at 2-5 pN/s (40 curves superposed).
J. Liphardt et al., Science 292, 733 (2001) ; 296, 1832 (2002).
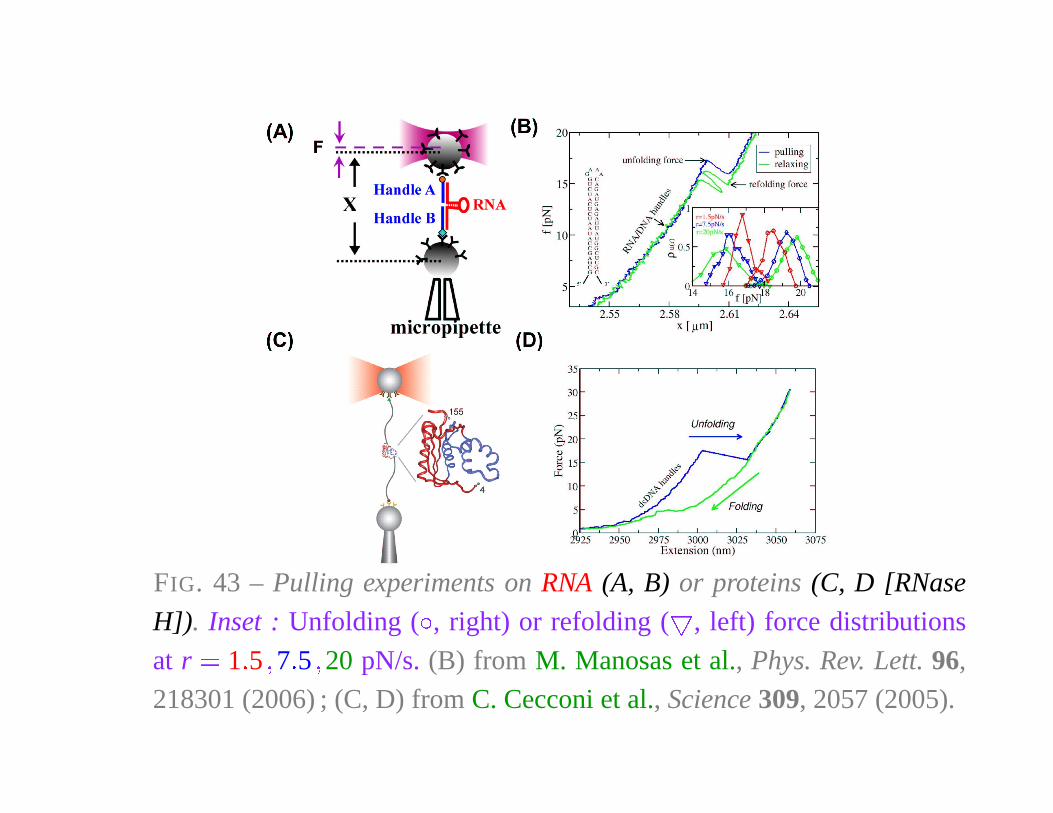
FIG. 43 – Pulling experiments on RNA (A, B) or proteins (C, D [RNase
H]). Inset : Unfolding ( �, right) or refolding ( � , left) force distributions
at r � 1 � 5 � 7 � 5 � 20 pN/s. (B) from M. Manosas et al., Phys. Rev. Lett. 96,
218301 (2006) ; (C, D) from C. Cecconi et al., Science 309, 2057 (2005).

JARZYNSKI EQUALITY
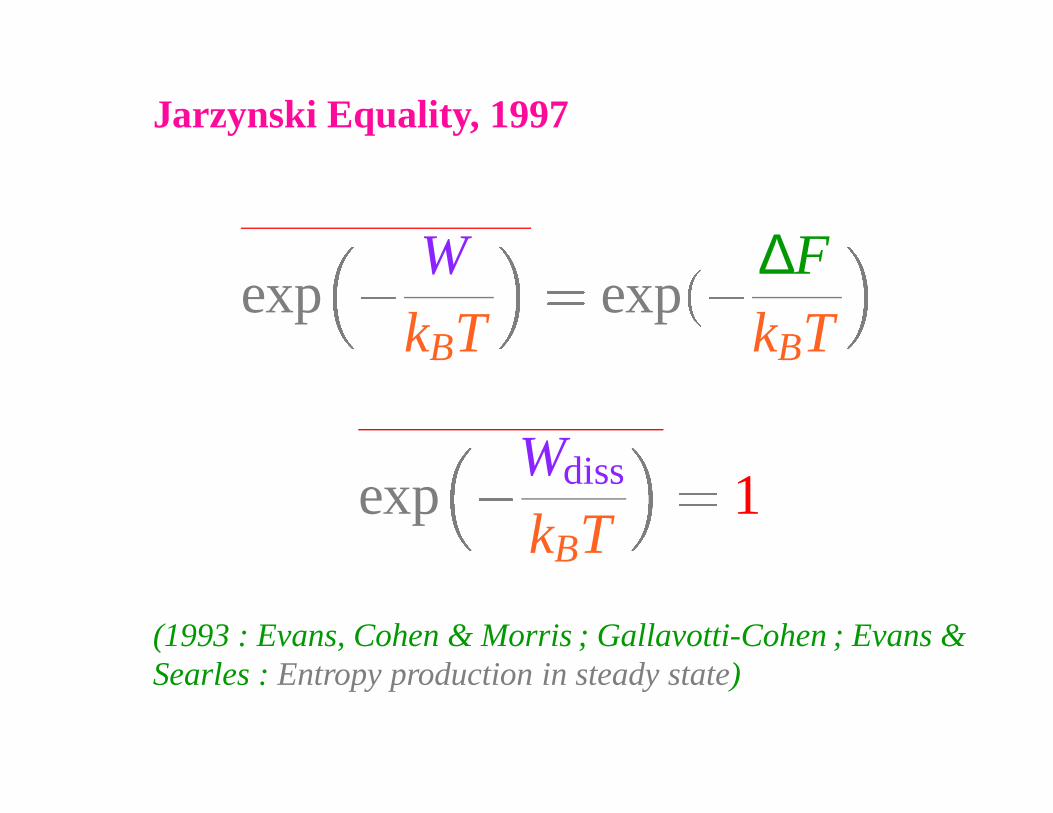
Jarzynski Equality, 1997
exp �
WkBT
� exp �∆FkBT
exp �
Wdiss
kBT
� 1
(1993 : Evans, Cohen & Morris ; Gallavotti-Cohen ; Evans &Searles : Entropy production in steady state)
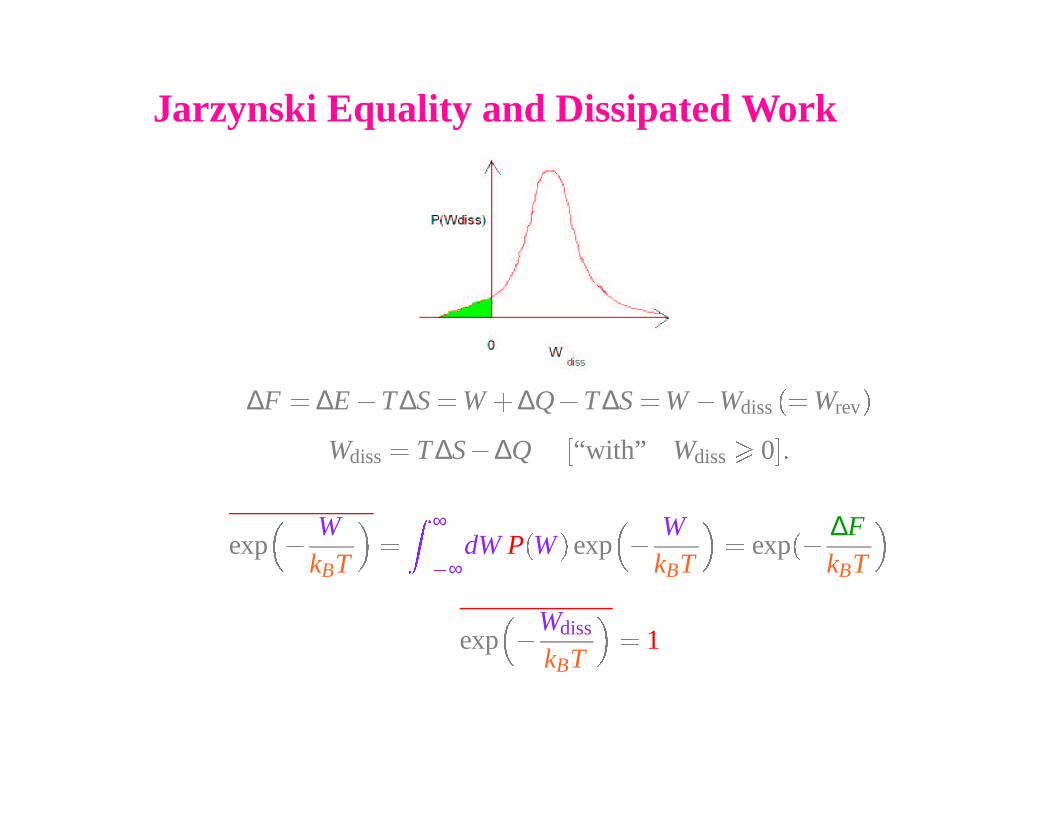
Jarzynski Equality and Dissipated Work
∆F � ∆E � T∆S � W
� ∆Q � T ∆S � W � Wdiss
�� Wrev
�
Wdiss
� T ∆S � ∆Q�
“with” Wdiss
�
0
��
exp
��
WkBT
�
�
∞
� ∞dW P
�
W
�
exp
��
WkBT
�
� exp
��
∆FkBT
�
exp
��
Wdiss
kBT
�
� 1
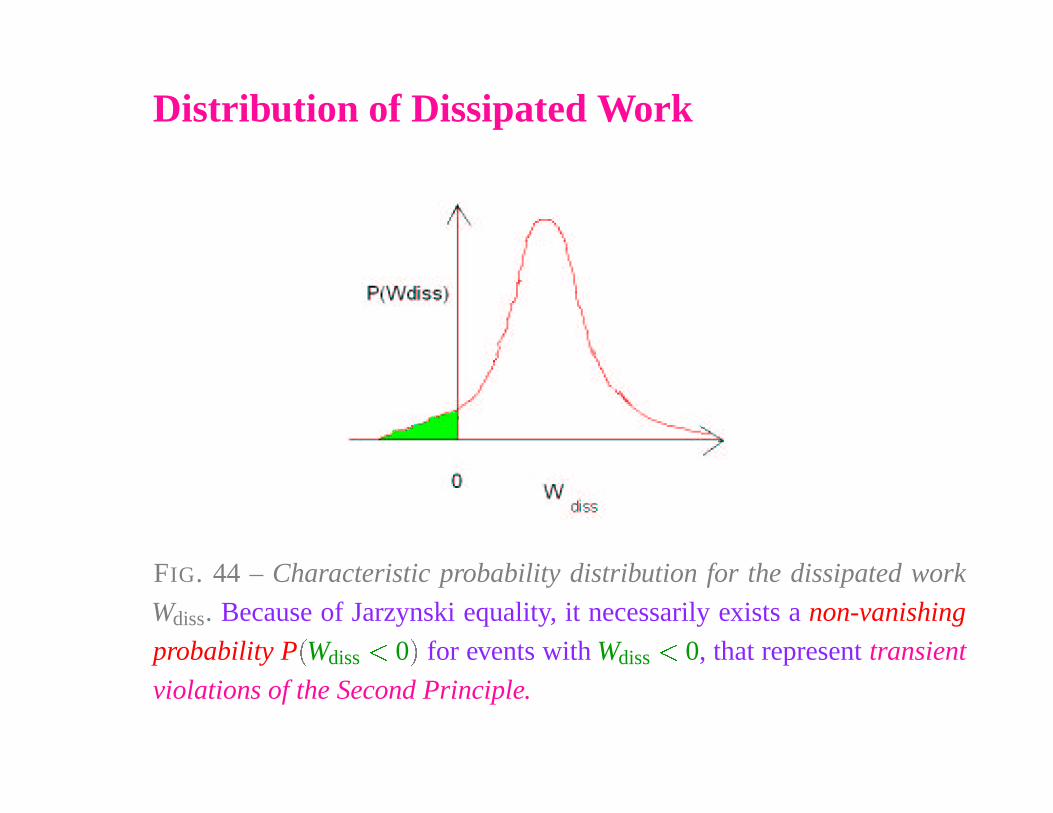
Distribution of Dissipated Work
FIG. 44 – Characteristic probability distribution for the dissipated work
Wdiss. Because of Jarzynski equality, it necessarily exists a non-vanishing
probability P
�
Wdiss� 0
�
for events with Wdiss
� 0, that represent transient
violations of the Second Principle.
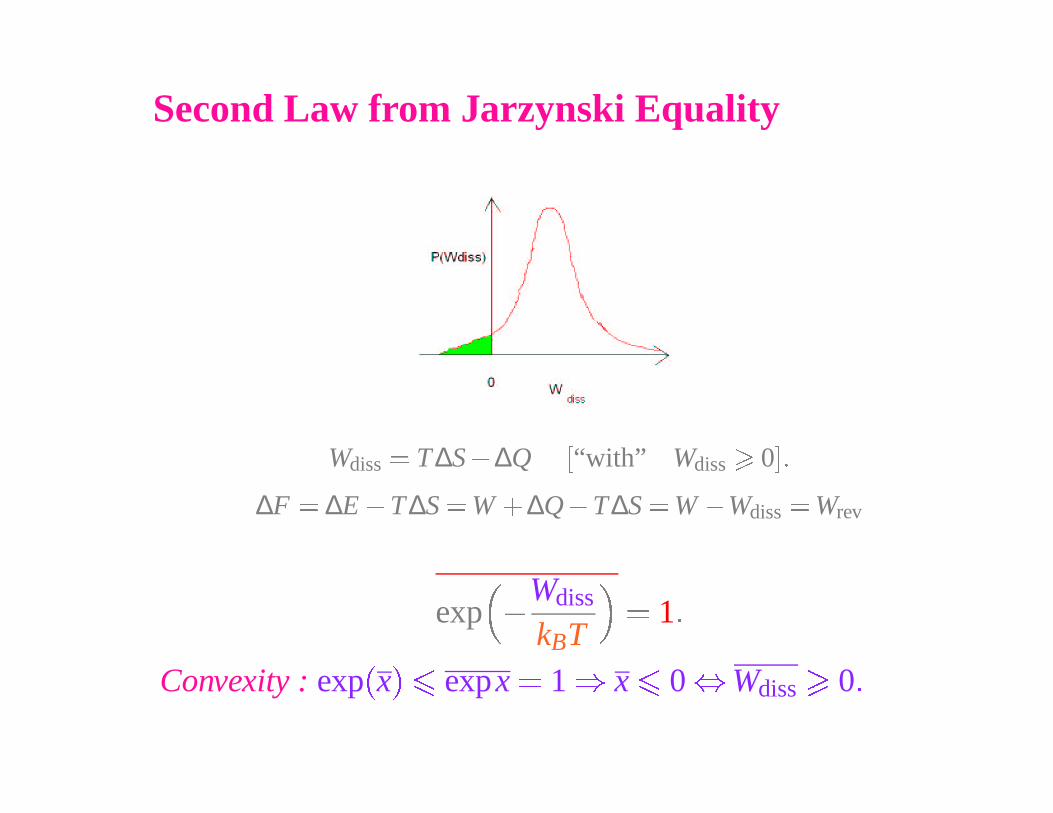
Second Law from Jarzynski Equality
Wdiss
� T ∆S � ∆Q�
“with” Wdiss
�
0
��
∆F � ∆E � T ∆S � W� ∆Q � T∆S � W � Wdiss
� Wrev
exp �
Wdiss
kBT
� 1 �
Convexity : exp�
x� �
expx � 1 � x
�
0 � Wdiss
�
0 �
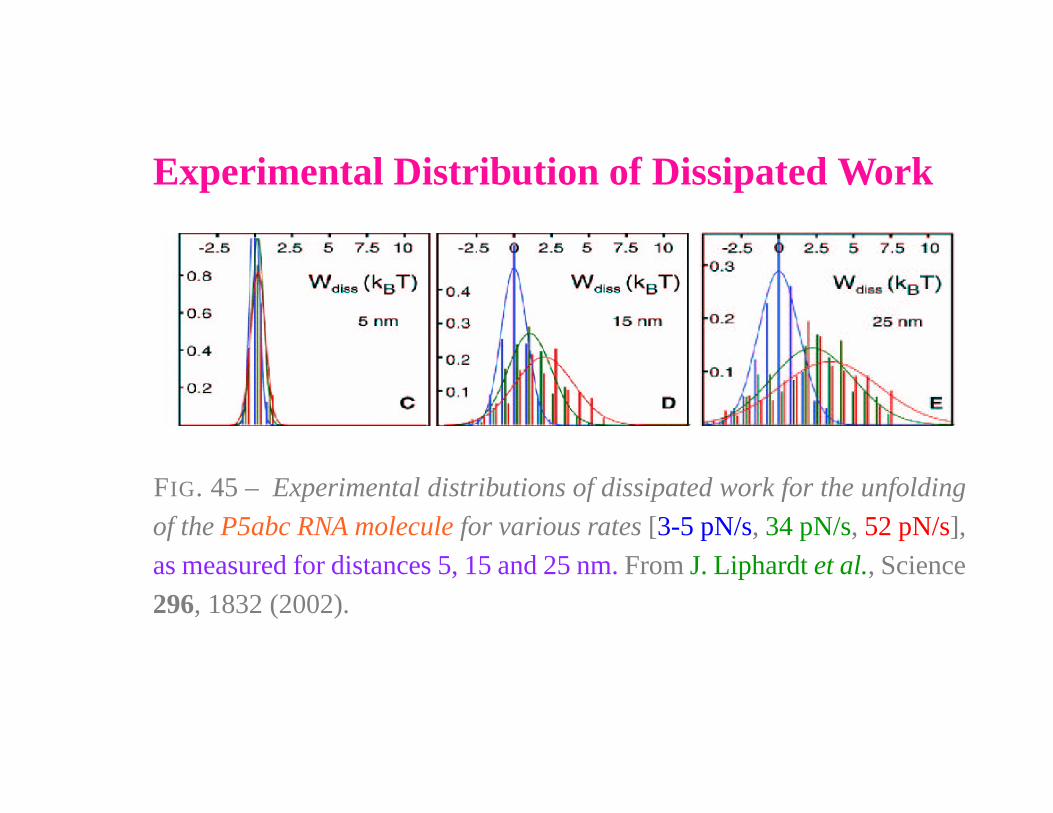
Experimental Distribution of Dissipated Work
FIG. 45 – Experimental distributions of dissipated work for the unfolding
of the P5abc RNA molecule for various rates [3-5 pN/s, 34 pN/s, 52 pN/s],
as measured for distances 5, 15 and 25 nm. From J. Liphardt et al., Science
296, 1832 (2002).
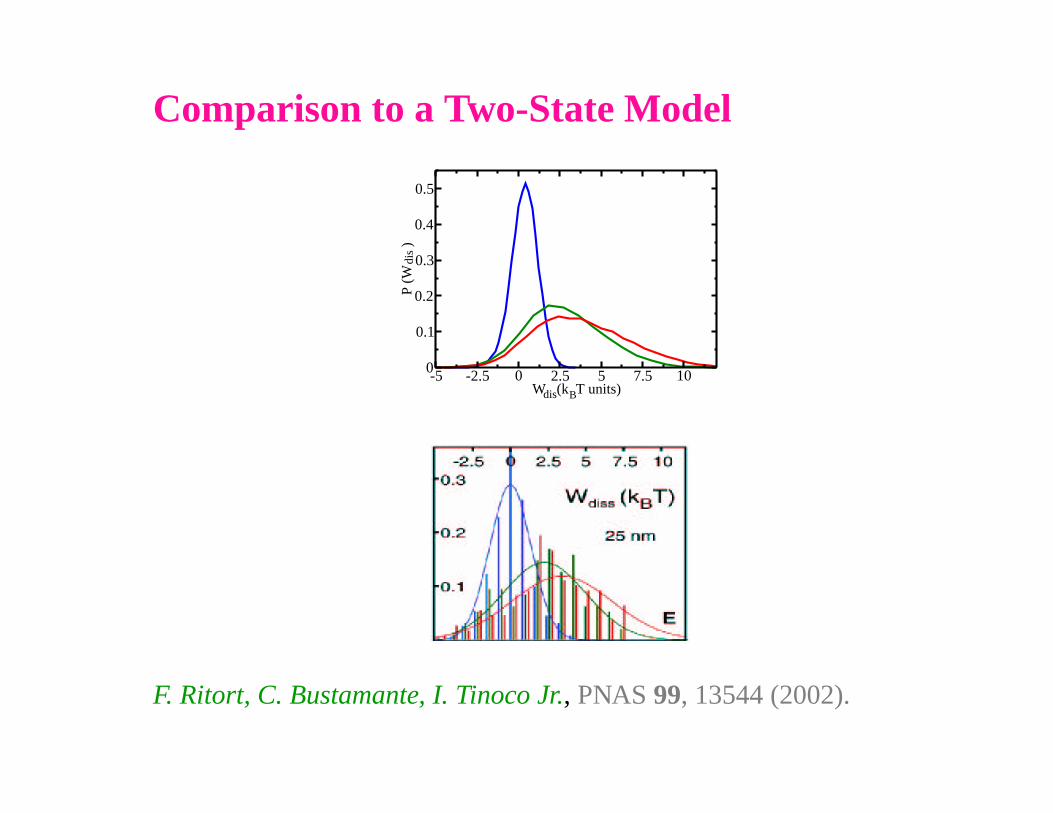
Comparison to a Two-State Model
-5 -2.5 0 2.5 5 7.5 10W (k T units)
0
0.1
0.2
0.3
0.4
0.5
P (W
)
dis
dis B
F. Ritort, C. Bustamante, I. Tinoco Jr., PNAS 99, 13544 (2002).

Convergence with the Number of Pulls
BJE � � ln exp �
WkBT �
Npulls
�
∆FkBT
� � ln exp �Wdiss
kBT �
Npulls
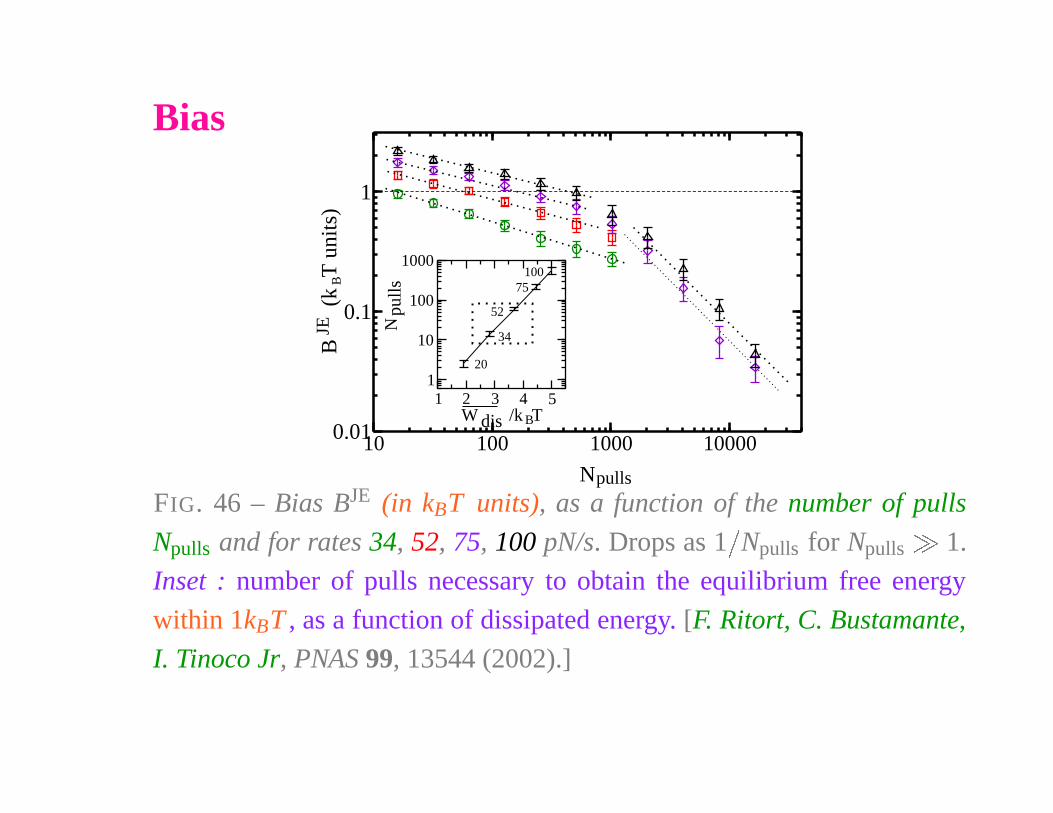
Bias
10 100 1000 10000
N
0.01
0.1
1
B
(k
T u
nits
)
1 2 3 4 5W /k T
1
10
100
1000
N
pulls
B
JE
dis
pulls
20
34
52
75100
B
FIG. 46 – Bias BJE (in kBT units), as a function of the number of pulls
Npulls and for rates 34, 52, 75, 100 pN/s. Drops as 1
�
Npulls for Npulls
�
1.
Inset : number of pulls necessary to obtain the equilibrium free energy
within 1kBT , as a function of dissipated energy. [F. Ritort, C. Bustamante,
I. Tinoco Jr, PNAS 99, 13544 (2002).]

CROOKS
FLUCTUATION THEOREM
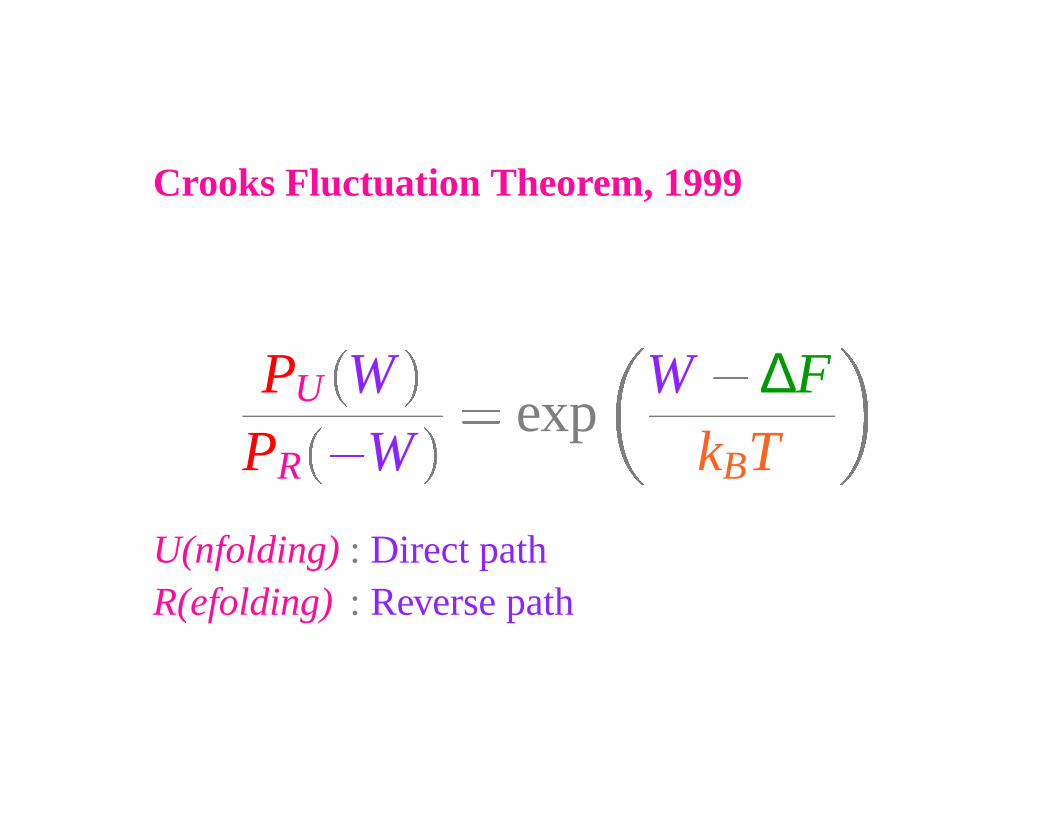
Crooks Fluctuation Theorem, 1999
PU WPR
�W
� expW � ∆F
kBT
U(nfolding) : Direct pathR(efolding) : Reverse path
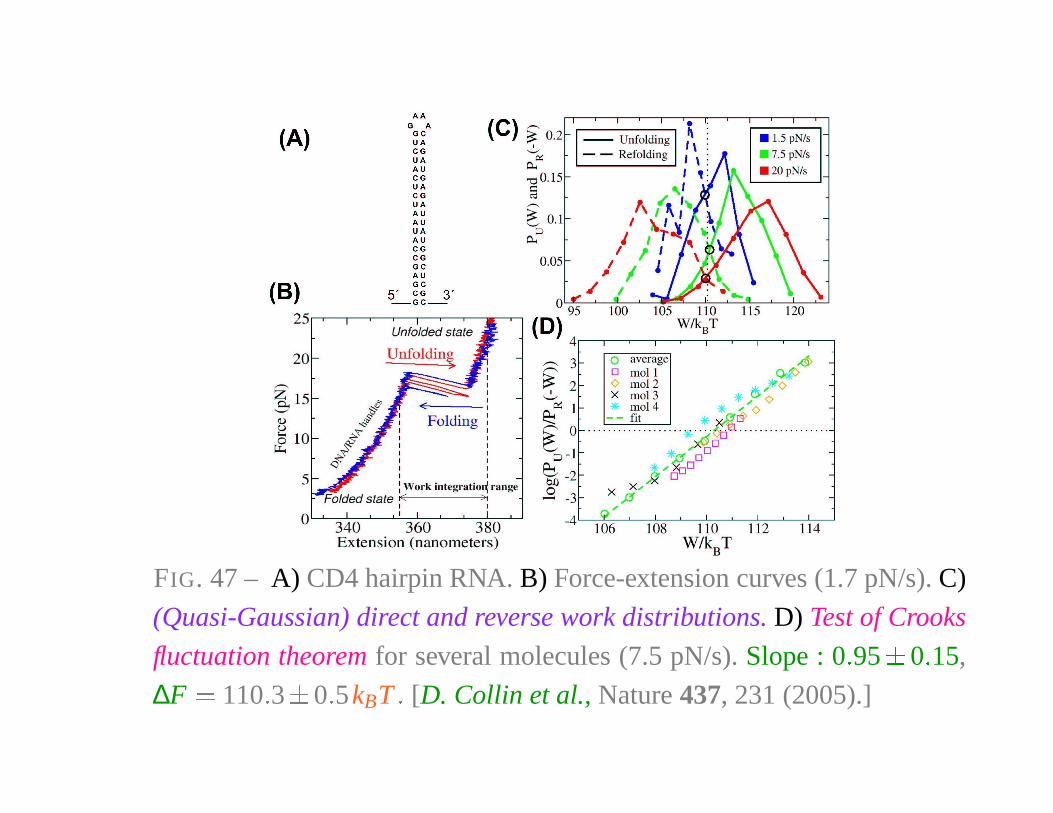
FIG. 47 – A) CD4 hairpin RNA. B) Force-extension curves (1.7 pN/s). C)
(Quasi-Gaussian) direct and reverse work distributions. D) Test of Crooks
fluctuation theorem for several molecules (7.5 pN/s). Slope : 0 � 95
�
0 � 15,
∆F � 110 � 3
�
0 � 5kBT � [D. Collin et al., Nature 437, 231 (2005).]
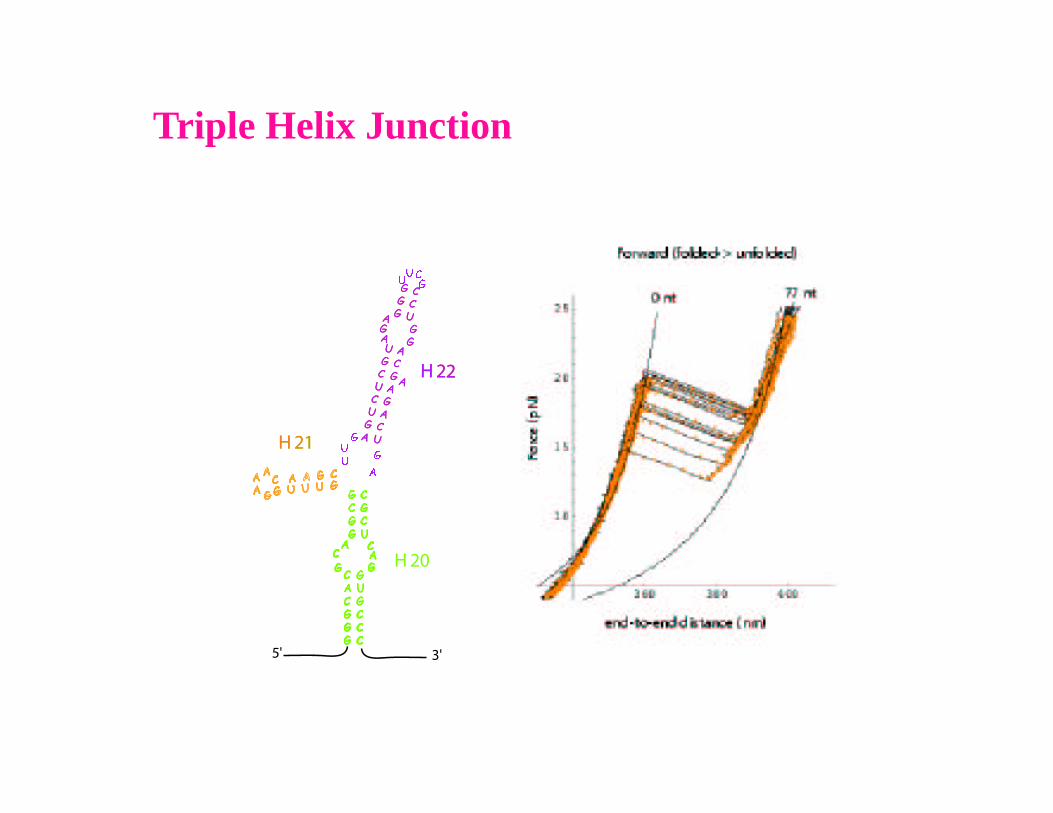
Triple Helix Junction
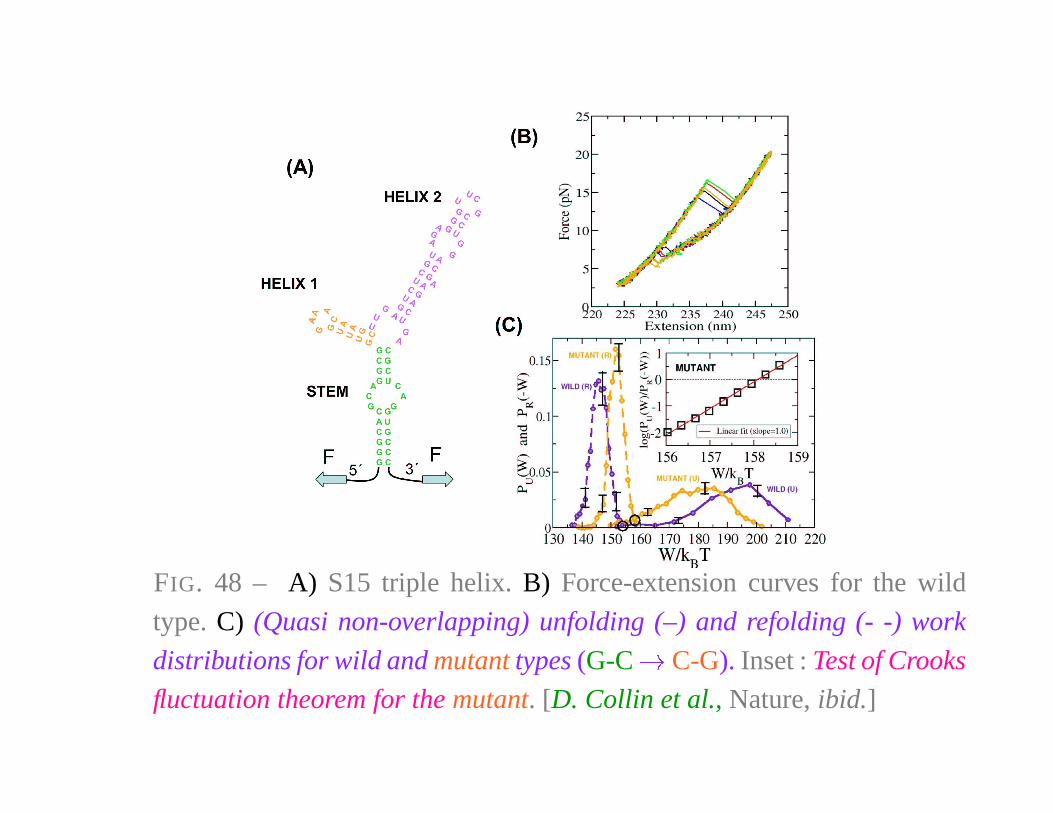
FIG. 48 – A) S15 triple helix. B) Force-extension curves for the wild
type. C) (Quasi non-overlapping) unfolding (–) and refolding (- -) work
distributions for wild and mutant types (G-C � C-G). Inset : Test of Crooks
fluctuation theorem for the mutant. [D. Collin et al., Nature, ibid.]


















