

Fusobacterium nucleatum and human beta defe nsins...
40
1 Fusobacterium nucleatum and human beta defensins modulate the 1 release of antimicrobial chemokine CCL20/Mip-3α 2 3 4 Santosh K. Ghosh, Sanhita Gupta, Bin Jiang and Aaron Weinberg* 5 Department of Biological Sciences, School of Dental Medicine, Case Western Reserve 6 University, Cleveland, OH 44106 7 8 9 Running title: Fusobacterium nucleatum modulate the release of CCL20 10 Key words: Fusobacterium nucleatum, CCL20, Mip-3α, Human beta defensins, Oral 11 epithelial cells. 12 13 14 15 *Correspondence: 16 Case Western Reserve School of Dental Medicine 17 2124 Cornell Road 18 Cleveland, OH 44106-4905 19 phone: 216-368-6729 20 fax: 216-368-0145 21 E-mail: [email protected] 22 23 Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Infect. Immun. doi:10.1128/IAI.05586-11 IAI Accepts, published online ahead of print on 12 September 2011 on June 18, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Fusobacterium nucleatum and human beta defe nsins...

1
Fusobacterium nucleatum and human beta defensins modulate the 1
release of antimicrobial chemokine CCL20/Mip-3α 2
3
4
Santosh K. Ghosh, Sanhita Gupta, Bin Jiang and Aaron Weinberg* 5
Department of Biological Sciences, School of Dental Medicine, Case Western Reserve 6
University, Cleveland, OH 44106 7
8
9
Running title: Fusobacterium nucleatum modulate the release of CCL20 10
Key words: Fusobacterium nucleatum, CCL20, Mip-3α, Human beta defensins, Oral 11
epithelial cells. 12
13
14
15
*Correspondence: 16
Case Western Reserve School of Dental Medicine 17
2124 Cornell Road 18
Cleveland, OH 44106-4905 19
phone: 216-368-6729 20
fax: 216-368-0145 21
E-mail: [email protected] 22
23
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.05586-11 IAI Accepts, published online ahead of print on 12 September 2011
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2
ABSTRACT: Cells of the innate immune system regulate immune responses through the 24
production of antimicrobial peptides, chemokines and cytokines, including human beta 25
defensins (hBDs) and CCL20. In this study, we examined the kinetics of primary human 26
oral epithelial cells’ (HOECs) production of CCL20 and hBDs, in response to 27
Fusobacterium nucleatum, a commensal bacterium of the oral cavity, which we previously 28
showed promotes HOEC induction of hBDs. HOECs secrete maximal levels of CCL20 at 29
6h, following stimulation by F. nucleatum cell wall (FnCW). Kinetics of CCL20 release is 30
distinct from that of hBD-2 and -3, which peaks after 24h and 48h FnCW stimulation, 31
respectively. FnCW induced release of CCL20 by HOECs requires both transcriptional 32
and translational activation. Release of CCL20 by HOECs is inhibited by brefeldin A, 33
suggesting that it is secreted through a vesicle transport pathway. Other epithelial 34
derived agents that FnCW induces, such as hBD-2, hBD-3, TNF-α and IL-1β, are also 35
able to release CCL20. By focusing on MAPKs, we show that both ERK1/2 and p38, but 36
not JNK, are required for hBDs, TNF-α and IL-1β induced secretion of CCL20 by 37
HOECs. The ability of FnCW and its induced hBDs, to produce proinflammatory 38
cytokines and CCL20, suggests the broad role of F. nucleatum and human antimicrobial 39
peptides in primary immune responses elicited by oral epithelium. 40
41
42
43
44
45
46
47
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

3
INTRODUCTION 48
Epithelial cells are the host's first line of defense against microbial invasion and pathogenesis 49
[9, 12, 13,18]. At mucosal surfaces, including those of oral epithelia, cells have evolved as part 50
of the innate immune system, to possess antimicrobial activity and to “cross-talk” with the 51
adaptive immune system. Cells of the innate immune system regulate immune responses 52
through the production of chemokines and cytokines, including macrophage inflammatory 53
protein 3α (Mip-3α), also known as CCL20. CCL20 is a 70 amino-acid chemokine that attracts 54
immature dendritic cells and T cells via the chemokine receptor CCR6 [55], as has been 55
reported for the epithelial cell derived antimicrobial peptides (AMPs) human beta defensins 56
(hBD) 1 and -2 [60]. We and others have reported on the ability of hBDs to “cross-talk” with 57
the adaptive immune system [21,25,52] and, most recently we showed that overexpression of 58
hBD-3 is linked to oral cancer [34,35]. CCL20 has also been linked to a variety of diseases, 59
including cancer [39], rheumatoid arthritis [54] and may have an important regulatory role in 60
specific lymphocyte migration into inflamed periodontal tissues [30]. By in situ hybridization, 61
Abiko et al [2] showed that CCL20 expression in oral squamous cell carcinoma (OSCC) is 62
localized primarily at the epithelial pearls corresponding to the spinous layer. A recent study 63
[6] also suggests that CCL20 may represent a potential immunohistochemical marker for 64
prognostic prediction of OSCC. In periodontal diseased tissue, CCL20 has been shown to be 65
distributed in the basal layer of gingival epithelial cells [30]. The protein has direct 66
antibacterial, antifungal, and antiviral activities; again comparable to hBDs [11, 37, 46, 61]. 67
Therefore, CCL20, along with hBDs, may play an important role in bridging between innate 68
and adaptive immune responses. 69
Oral mucosa is in continuous contact with heterogeneous populations of diverse 70
commensal and opportunistic microorganisms and yet, for the most part, it remains healthy and 71
uninflamed. Recent research has highlighted the possibility that oral epithelium not only serves 72
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

4
a barrier function as the first physical line of defense against colonizing pathogens, but also 73
orchestrates local immune responses [14,26]. Of the 700 species of oral bacteria estimated to 74
colonize the oral cavity [1, 20, 48], a few have demonstrated the capacity to induce epithelial 75
cell innate responses, while others have been more stealth-like [23, 40, 56]. The ubiquitous oral 76
bacterium Fusobacterium nucleatum (Fn) and its cell wall extract (FnCW) induces expression 77
of hBD-2 and -3 transcript in gingival epithelial cells (GECs) in vitro [15,32,33,41]. Recently, 78
we have identified an FnCW associated peptide that induces hBD-2 in human oral epithelial 79
cells (HOECs) [29]. 80
FnCW has been shown to induce CCL20 transcript, and indeed induction of CCL20 by 81
FnCW is highest (72.7 fold in 6h) amongst all the other cytokines/chemokines studied [62]. In 82
the present communication, we further characterized F. nucleatum induced release of CCL20 83
by human oral epithelial cells (HOECs), the kinetics of beta defensin release by HOECs upon 84
stimulation with F. nucleatum, and their effect on CCL20 release by the host cells. The effects 85
of pro-inflammatory cytokines on HOEC regulation of CCL20 are also looked into. Finally, we 86
investigated the MAP kinase pathways regulating hBD and cytokine induced secretions of 87
CCL20 by HOECs. 88
89
90
91
92
93
94
95
96
97
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

5
METHODS AND MATERIALS 98
Chemicals and reagents 99
Synthetic hBD2 and hBD-3 were purchased from Peptides International [Kentucky, 100
USA]. Recombinant TNF-α and recombinant IL-1β and were purchased from Peprotech Inc 101
(NJ, USA). All the MAPK inhibitors used in this study were purchased from Calbiochem (NJ, 102
USA). Actinomycin D, brefeldin A, cyclohexamide and DMSO were purchased from Sigma 103
Aldrich (St. Louis, USA). 104
Human primary oral epithelial cells culture and stimulation 105
Healthy human gingival tissue overlying impacted third molars was obtained from oral surgery 106
clinics, in the greater Cleveland area, in accordance with the Case Western Reserve University 107
Institutional Review Board approved protocol. Primary HOECs were isolated and grown in 108
keratinocyte basal medium with supplements as previously described [10, 40, 41]. To 109
overcome interpersonal variability, epithelial cells from three donors were mixed at a ratio of 110
1:1:1 and grown to 80% confluence prior to challenge. Results were compared to HOECs 111
grown to similar confluence from single donors. To avoid the effects of changes in epithelial 112
cell physiology during growth in culture, each time point had a corresponding untreated, 113
control group. Each experimental condition was conducted in triplicate. 114
Preparation of F. nucleatum cell wall fractions. 115
Cell wall from F. nucleatum was prepared as described previously [36]. Briefly, F. nucleatum 116
(ATCC 25586) was grown anaerobically in Columbia broth (BD Biosciences) overnight. 117
Crude cell wall preparations were prepared by French pressure cell disruption of freshly 118
harvested whole cells in phosphate-buffered saline (PBS; pH 7.2) at 15,000 pounds/inch2. The 119
cell walls were recovered after low speed centrifugation (1,000 x g, 15 min), followed by high 120
speed centrifugation (138,000 x g, 30 min) of the supernatant. 121
Quantification of CCL20 in cell free supernatant and in cell lysates. 122
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

6
CCL20 was quantified in culture supernatants and in cell lysates by sandwich ELISA using 123
antibody pairs from R&D Systems (MN, USA) following the vendor’s instructions. CCL20 124
concentration was calculated from standard curves using recombinant human CCL20 (R&D 125
Systems, MN). The detection threshold was 250 pg/ml of CCL20. 126
Detection of hBD-2 and hBD-3 peptides in supernatant. 127
Human beta-defensins were measured in supernatants from FnCW stimulated HOECs 128
following our previously published protocol [27, 29]. 129
RNA preparation and PCR. 130
Cells were lysed using Trizol reagent (Invitrogen, Carlsbad, CA) and total RNA was isolated in 131
accordance with the manufacturer’s instructions. RNA concentration was measured by UV 132
absorbance using the Nanodrop 1000 spectrophotometer (Thermo Fisher Scientic inc., USA). 133
RT PCR was done with the Superscript III One Step RT PCR system (Invitrogen,USA) 134
according to the manufacturer’s protocol and the amplified product was analyzed on a 1.5% 135
agarose gel. Real-time PCR was done using the BIO-RAD (Hercules, CA) MyIQ system with 136
iScript cDNA Synthesis Kit and iQ SYBR Green Supermix according to the manufacturer’s 137
protocol. The results were analyzed to calculate fold induction of CCL20 with human keratin 5 138
(HK5) as the internal control. The following primers were used to amplify CCL20 and HK-5: 139
For CCL20 5’-AATTTATTGTGGGCTTCACACG-3’, 5’-140
ACCCAAGTCTGTTTTGGATTTG-3’ and for HK5 5’-GTC CTC TCC ATG GAC AAC 141
AAC- 3’, 5’-TGT CAA TCT CGG CTC TCA GCC-3’. 142
PathScan® MAP Kinase Multi-Target Sandwich ELISA. 143
Mixed donor (N=3) HOECs were treated by adding fresh media containing the regulator for 144
the desired time, rinsed with ice-cold PBS and lysed with ice-cold 1X lysis buffer (Cell 145
Signaling, Danvers, MA) plus 1mM phenylmethylsulfonyl fluoride (PMSF) for 10 min on ice. 146
Cells were scraped off the plate, transferred to a microfuse tube, sonicated on ice and clarified 147
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

7
by centrifugation (13,000 rpm, 4°C, 15 min). The PathScan® Multi-Target (phospho-p38, 148
phospho-ERK1/2, phospho-MEK1/2, phospho-SAPK/JNK and MEK1) ELISA was then 149
performed following the vendor’s (Cell signaling, MA) instruction on the clarified supernatant. 150
Statistical analysis 151
Statistical analyses were performed using either unpaired Student’s t-test or ANOVA test. The 152
differences were considered significant when the probability value was less than 5% (P < 0.05). 153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

8
RESULTS 173
F. nucleatum induces release of CCL20 by HOECs. 174
To determine baseline release of CCL20 by unstimulated HOECs, cell free supernatants from 175
epithelial cell cultures were collected at different time intervals and CCL20 concentration was 176
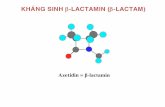
quantified by ELISA. Figure 1A demonstrates release of CCL20 protein by HOECs as early as 177
1 hr after the addition of fresh media and increases gradually with time. To examine the release 178
of CCL20 protein upon stimulation with F. nucleatum, HOECs were treated with F. nucleatum 179
cell wall (FnCW; 10 µg/ml) for 1–48h. Time course studies demonstrated a significant increase 180
in CCL20 release at 4h following F. nucleatum stimulation, with a plateauing of CCL20 181
release after 6 hr of stimulation [Fig. 1A]. We observed that the fold increase in secreted 182
CCL20 from FnCW treated cells, when compared to untreated quiescent cells, reached its 183
maximum by 6 hrs post treatment [Fig. 1B, derived from Fig 1A], and that interpersonal 184
variability was demonstrated in HOECs from different donors towards release of CCL20 upon 185
F. nucleatum challenge; i.e., from 2.8 to 8.5 fold above baseline (N=5; variation 186
coefficient, %CV= 37) [Fig. 1C] 187
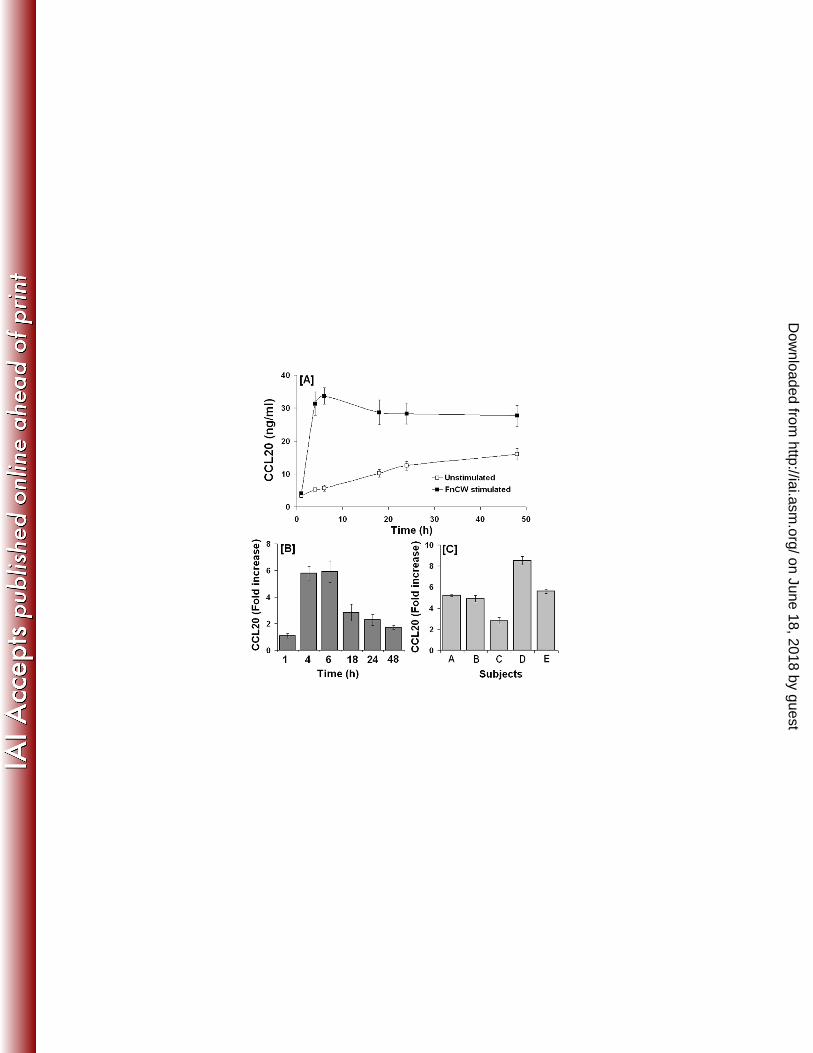
CCL20 release requires rapid de novo generation and is inhibited by brefeldin A. 188
To determine if CCL20 is synthesized by HOECs in response to FnCW, near confluent HOEC 189
cultures were pretreated with actinomycin D (10μg/ml) or cyclohexamide (10μg/ml) for 5 min. 190
HOECs were then cultured for an additional 6 h with FnCW, as that was shown to be the 191
optimal time for CCL20 release (Fig. 1A, B). Controls included HOECs with media alone (or 192
DMSO in media), with FnCW alone, or with either actinomycin D or cyclohexamide alone. 193
After 6 h of exposure, media were collected and analyzed for the presence of CCL20. As 194
shown in Figure 2A, FnCW is unable to induce the release of CCL20 by HOECs in the 195
presence of actinomycin D or cyclohexamide, indicating that the release of CCL20 by FnCW 196
is transcriptionally and translationally regulated. To examine the effect of brefeldin A (BFA), a 197
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

9
protein release inhibitor, on FnCW induced release of CCL20 by HOECs, near confluent 198
HOECs were treated with BFA (10 μg/ml) for 5 min followed by FnCW (10 μg/ml) for 6h, or 199
with either agent alone for up to 6h; supernatant and pellet samples were collected and CCL20 200
levels were measured by ELISA. BFA inhibited CCL20 release from HOECs in the presence 201
of FnCW stimulation [Fig. 2B]. In contrast, BFA alone or in combination with FnCW caused 202
an increase in intracellular CCL20 [Fig. 2C]. Comparison of CCL20 levels in supernatants and 203
in cell pellets indicates that the CCL20 is rapidly released upon synthesis. 204
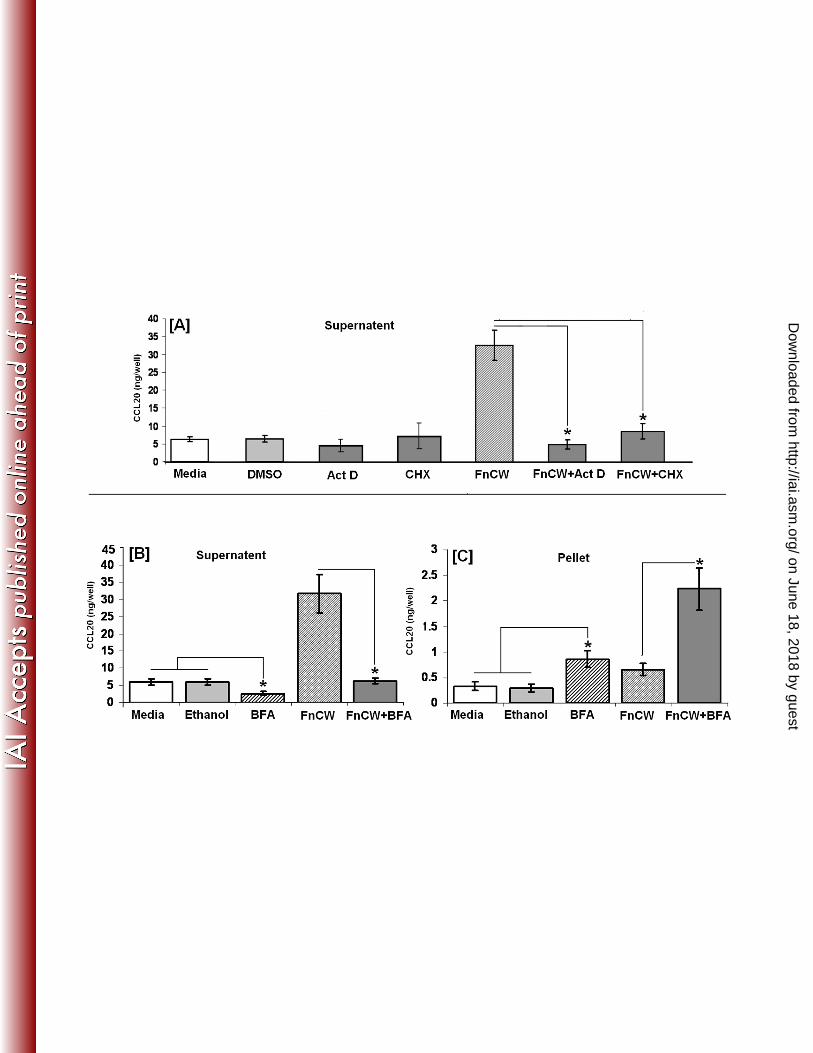
Release of beta defensins in HOECs by FnCW. 205
Cell free supernatants from FnCW (10μg/ml) treated HOECs were collected at different time 206
points (1 – 72 hr) and hBD-2 and -3 peptide concentrations were quantified by ELISA. Figures 207
3A and 3B show the kinetics of hBD-2 and -3 released from untreated vs. FnCW treated 208
HOECs over the designated time periods, respectively. When compared to the untreated cells, 209
hBD-2 fold increases of 1.1, 7.5, 13.5, 3.9 and 2.7 were observed at 4, 18, 24, 48 and 72 h, 210
respectively [Fig. 3C]. Fold increases for hBD-3 [Fig. 3E] were 1.1, 1.4, 2.3, 2.9 and 2.7 at the 211
same respective time points as for hBD-2. Interpersonal variability was evident for hBD-2 and 212
-3 release from HOECs induced by FnCW for 24h [Fig. 3D and 3F; N=5 donors; variation 213
coefficient, % CV=38.5 and 27.5, for hBD-2 and hBD-3, respectively]. 214
In contrast to hBD-2 and -3, we observed no differences in the release of hBD-1 215
between FnCW stimulated and unstimulated control cells (data not shown), supporting the 216
constitutive nature of hBD-1 expression in HOECs. However, linear accumulation of hBD-1 in 217
both stimulated and unstimulated culture media over time was observed with R2 values 0.94 218
and 0.97, respectively. 219
220
221
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

10
Induction of CCL20 by beta defensins (-2 and-3). 222
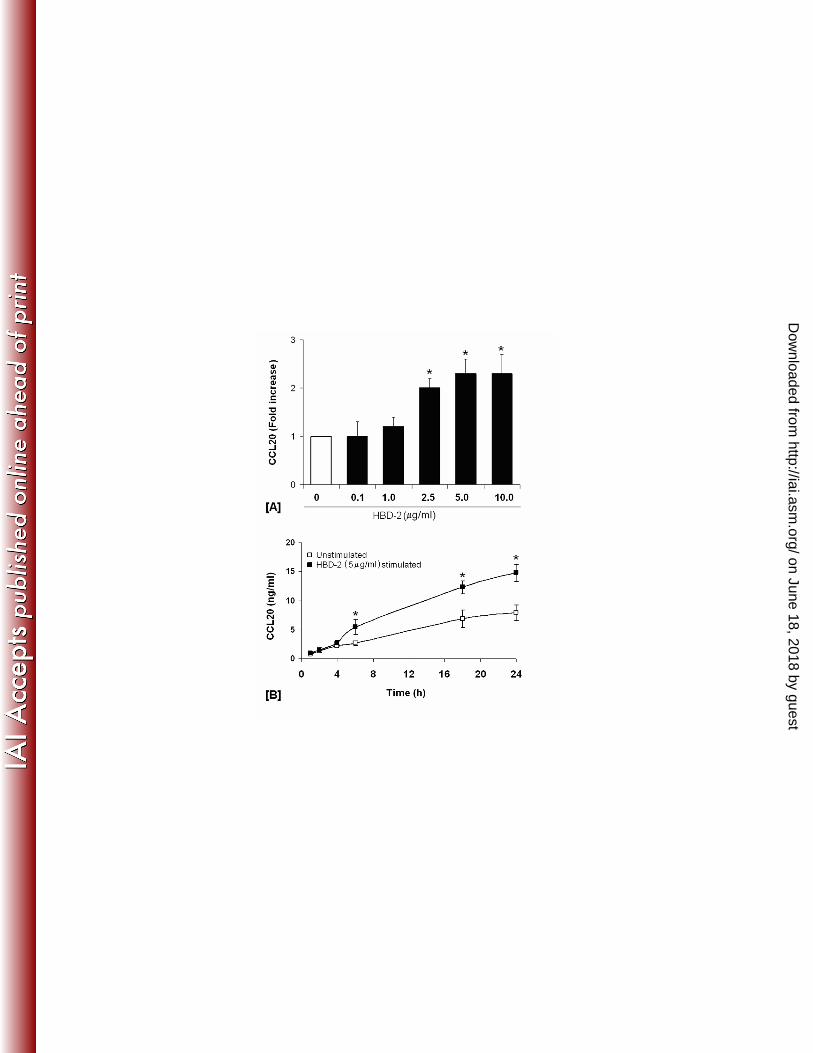
Since hBD-2 (2μg/ml) has been shown to induce CCL20 transcript in primary oral epithelial 223
cells [62], we looked for hBD-2 induced CCL20 release by HOECs. HOECs were incubated 224
with synthetic hBD-2 (0.1 to 10 μg/ml) for 6 hr. An hBD-2 dose dependent release of CCL20 225
was evidenced, with the maximum release noted with 5 μg/ml hBD-2 [Fig. 4A]. Moreover, 226
CCL20 was seen to increase in the spent supernatant over time (1-24 hrs) when hBD-2 was 227
added to HOECs at the optimal concentration of 5 μg/ml [Fig. 4B]; a kinetic profile different 228
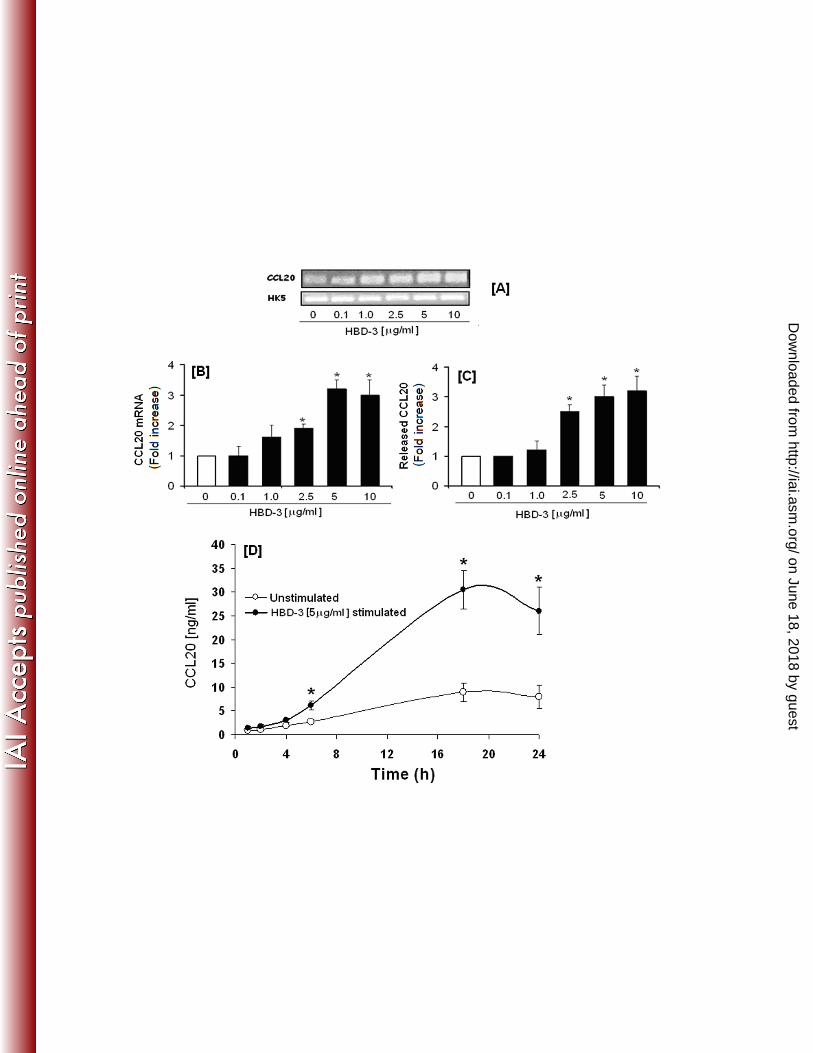
from that seen with FnCW (Fig. 1A). To investigate the inductive capacity of hBD-3 to 229
promote CCL20 expression and subsequent release from HOECs, we incubated near confluent 230
HOECs with increasing concentrations of hBD-3 (0.1 to 10 µg) for 6 hrs, followed by isolation 231
of total RNA and amplification of CCL20 mRNA by PCR and quantification of released 232
CCL20 by ELISA, respectively. The increase in CCL20 mRNA expression was also 233
determined by real time PCR. There was a direct correlation between the increase in CCL20 234
transcript expression [Fig. 5B] and protein release from HOECs with hBD-3 peptide 235
concentrations used over the 6 hr period [Fig. 5C]. CCL20 was seen to also increase in the 236
spent supernatant over time (1-24 hrs) when hBD-3 was added to HOECs at the optimal 237
concentration of 5 μg/ml, as seen with hBD-2, but different from that seen with FnCW (Fig. 238
1A). Interestingly, hBD-3 induced higher levels of released CCL20 than did hBD-2 (compare 239
Fig. 4B and 5D) and there appeared to be an approximately 6 fold increase in CCL20 in the 240
spent supernatant induced by hBD-3 at around 18 hrs of incubation (Fig. 5D). 241
No lactate dehydrogenase activity was detected in the supernatants after stimulation 242
with hBD-2 or hBD-3 (0.1 to 10 μg/ml) for 24 hrs (data not shown), confirming that cellular 243
viability was not impaired by hBD-2 or -3 at the concentration range used and time periods 244
studied. 245
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

11
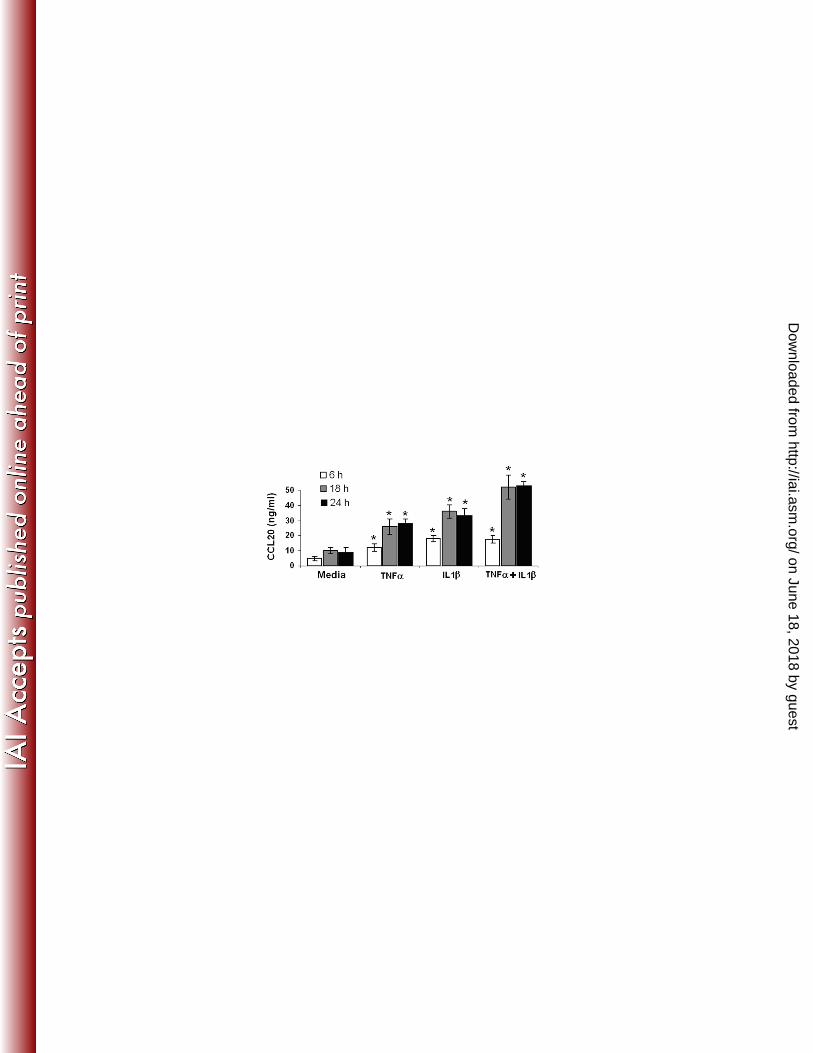
Cytokines (IL-1β, TNF-α) induce secretion of CCL20 by HOECs. 246
We and others have previously shown that FnCW induces TNF-α and IL-1β transcript 247
expression in HOECs [41,62], and that these two cytokines have been shown to induce 248
CCL20 release from several epithelial cell lines [8,49,51]. Here, we looked to see if these 249
cytokines could induce the release of CCL20 from HOECs. We incubated HOEC monolayers 250
with 10 ng/ml of either IL-1β or TNF-α for three different time periods (6, 18 and 24 hrs), 251
followed by ELISA analysis. We observed that both cytokines are capable of releasing 252
increased amounts of CCL20 compared to the untreated control [Fig. 6]. When cells were 253
stimulated with a combination of TNF-α and IL1β, an additive effect was observed in the 254
release of CCL20. Since F. nucleatum and hBD induced CCL20 production is a relatively early 255
event, it is unlikely that the production of intermediary cytokines are involved in F. nucleatum 256
or hBD induced release of CCL20. We corroborated this by using antibodies against TNF-α 257
and IL1β in our assay system, and saw that FnCW and hBDs, respectively, were not prevented 258
from inducing CCL20 in HOECs (data not shown). 259
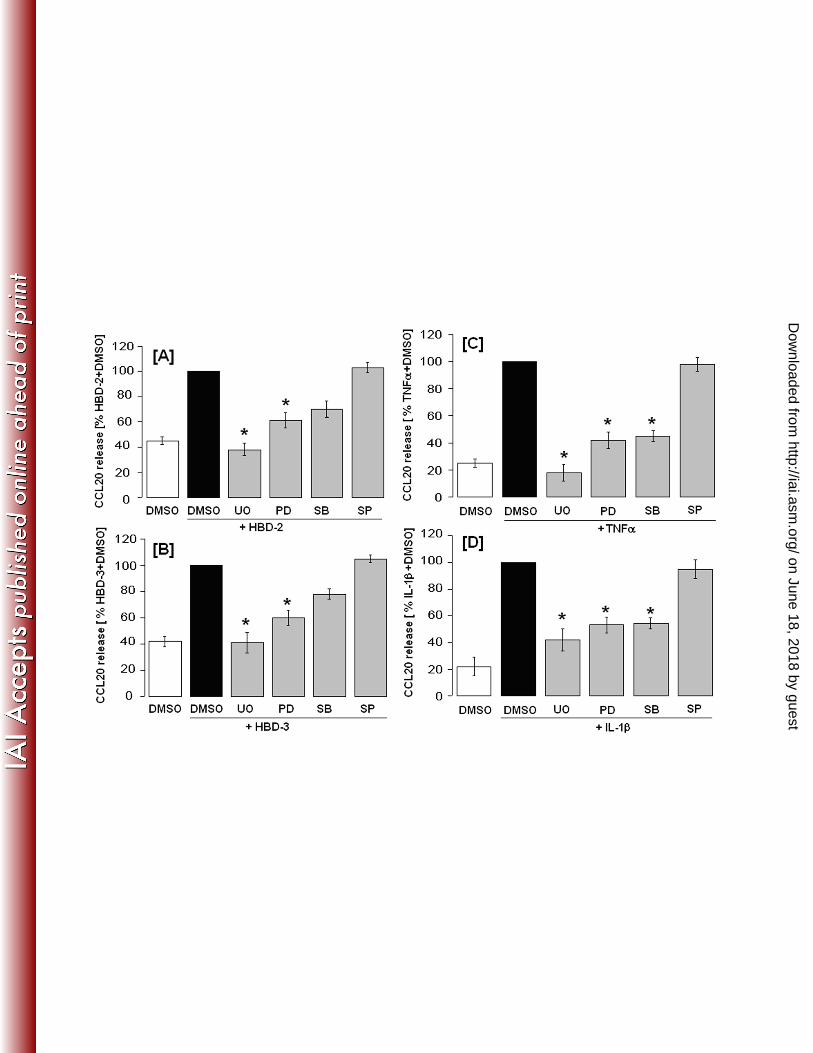
Effect of MAP kinase inhibitors on defensin and cytokine induced release of CCL20 by 260
HOECs. 261
Different MAP kinases exist and their impact on CCL20 production in each cell line is 262
different [31, 43,51,57]. Hence, the signaling pathway involved in CCL20 production in 263
HOECs needs further investigation. Since it was reported that beta defensins may activate 264
MAPK pathways [47], we used inhibitors of these pathways to see whether any are involved in 265
hBD induced release of CCL20. HOEC monolayers were pretreated for 1 hr with different 266
inhibitors before 18 hr incubation with stimulants. As shown in Fig. 7A and 7B, ERK1/2 267
activation inhibitors [UO126 (20μM) or PD98059 (20μM)] were effective in suppressing beta-268
defensin (hBD-2 or hBD-3) induced release of CCL20 by HOECs. The specific p38 mitogen 269
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

12
kinase inhibitor, SB203580 (20μM), also had a modest inhibitory effect on defensin induced 270
CCL20 release. In contrast, the JNK specific inhibitor, SP600125 (20μM), had no inhibitory 271
effect on defensin induced CCL20 release. Similar experiments were conducted to assess the 272
importance of MAPK in TNF-α or IL-1β induced release of CCL20 by HOECs. We found that, 273
like beta defensins, these cytokines induce CCL20 via ERK1/2 and p38 but not via JNK [Fig. 274
7C and 7D]. Interestingly, all four MAPK inhibitors showed little or no effect on FnCW 275
induced CCL20 release by HOECs (data not shown), suggesting that the signaling pathway(s) 276
involved for FnCW induced secretion of CCL20 is/are different from defensins and cytokine 277
induced release of CCL20 by HOECs. 278
Since the inhibitor studies showed that beta defensins ( hBD-2 or hBD-3) and cytokines 279
(TNFα or IL-1β) induced release of CCL20 via ERK1/2 and p38 but not via JNK, we used 280
PathScan® MAP Kinase Multi-Target Sandwich ELISA to compare the levels of phospho-281
ERK1/2 (Thr202/Tyr204), phospho-MEK1/2 (Ser217/221), phospho-p38 MAPK 282
(Thr180/Tyr182) and phospho-SAPK/JNK (Thr183/Tyr185) in the cell lysates of HOECs after 283
stimulating the cells with TNFα, IL-1β, hBD-2 and hBD-3, respectively. As shown in Fig. 8, 284
phosphorylation levels of ERK1/2, MEK1/2 and p38, but not of SAPK/JNK, were enhanced in 285
HOECs following 30 or 60 min of challenge with the above mentioned stimuli. 286
287
288
289
290
291
292
293
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

13
DISCUSSION 294
Commensal bacteria are usually regarded as beneficial to the host by displacing pathogens 295
from a microbial niche or providing protection by continually stimulating epithelial surfaces to 296
express and secret antimicrobial peptides (AMPs) at levels that kill opportunistic/pathogenic 297
organisms [4]. The release of antimicrobial chemokine CCL20 [51] requires stimulation for its 298
release and the discrete stimuli that lead to its up regulation may be cell specific [17, 31, 31, 299
57,58]. In these studies, we demonstrated the ability of primary oral epithelial cells to release 300
CCL20 in response to the oral commensal bacterium F. nucleatum and other epithelial derived 301
agents that it also induces; i.e., hBD-2, hBD-3, TNF-α and IL-1β. The kinetic studies of 302
CCL20 release by HOECs suggest that this is an early event subsequent to challenge by any of 303
the agents used. FnCW caused rapid release of CCL20 from HOECs; i.e., 6 hr post challenge 304
resulted in a 6 fold greater presence of CCL20 in the spent supernatant than control supernatant 305
and twice the increase of a 24 hr challenge (6 fold vs ~3 fold). This is in agreement with a 306
recent study that showed that F. nucleatum elicits rapid induction of the CCL20 gene (72.7 307
fold in 6 hr compared to 21.7 fold in 24hr [62]. Lin et al [43] also showed early release of 308
CCL20 by human mast cells in response to Pseudomonas aeruginosa, which also peaked at 6 309
hr. Adenylate and uridylate (A+U)-rich elements (AREs)-mediated mRNA turnover is an 310
important regulatory component of gene expression for innate and specific immunity [24]. The 311
transient and rapid up-regulation of CCL20 gene expression by FnCW could be due to 312
posttranscriptional regulation of mRNA stability via AREs [53,19]. Indeed, when we looked 313
for AREs within the 3’untranslated region of CCL20 mRNA, using “ARE site” 314
[http://rna.tbi.univie.ac.at/AREsite., 28], we found three ATTTA domains [Supplementary 315
Figure S1]. Another rapidly induced gene by FnCW in HOECs is CXCL5 (40.5 fold in 6h) 316
[62] and using the ARE site we could find eleven ATTTA domains within its 3’UTR 317
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

14
[Supplementary Figure S1]. We could not find any AREs within the 3’UTR of DEFB4 318
(hBD-2) or in DEFB103 (hBD-3) and, not surprisingly, these defensins are not released as 319
rapidly as CCL20 by FnCW in HOECs. 320
321
In the present study, we found that actinomycin D and cyclohexamide inhibit FnCW 322
induced release of CCL20 by HOECs, indicating that CCL20 release requires transcriptional 323
and translational activation. Brefeldin A (BFA) is a fungal metabolite able to rapidly and 324
reversibly block intracellular vesicle transport from the endoplasmic reticulum to the Golgi 325
apparatus, and therefore, inhibit protein secretion with minimal effect on protein synthesis [38]. 326
BFA blocks the release of newly synthesized proteins, but has no effect on preformed granule-327
bound proteins [45]. Although BFA has been widely used as an inhibitor of protein secretion, 328
the information concerning its effect on CCL20 release has not been reported. Our study 329
showed that BFA not only inhibited the FnCW induced release of CCL20 by HOECs, but also 330
constitutive (unstimulated) release of CCL20. The increased levels of intracellular CCL20 in 331
unstimulated and FnCW stimulated cell lysates from BFA pretreated HOECs further 332
corroborates the inhibition of CCL20 release by BFA. Given the well-characterized properties 333
of BFA, it is reasonable to conclude that CCL20 is secreted through the vesicle transport 334
pathway. 335
Certain commensal bacteria are excellent inducers of beta defensins [41,59, 22], 336
suggesting that the commensal bacterial community may act by priming innate immune 337
readiness of oral epithelia. We and others have shown that F. nucleatum can induce innate 338
response elements, such as hBDs, in HOECs [29, 32, 41, 50,]. More recently, several 339
immunoregulatory functions with the adaptive immune system have been attributed to hBDs in 340
addition to their antimicrobial capacity; i.e., their ability to cross-talk with the adaptive 341
immune system by acting as chemokines [52, 34], and by changing a cell’s phenotype, either in 342
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

15
activation as seen with antigen presenting cells [25] or antagonism, as seen with T cells [21]. 343
These findings conform to reports in the literature that innate responses usually precede, and 344
are necessary for, the establishment of adaptive immunity [44]. Upon stimulation by 345
antimicrobial peptides, different immune and inflammatory cells have been shown to produce 346
cytokines and/or chemokines. For example, the α-defensins, human neutrophil peptides-1 and -347
3 stimulate the production of IL-1, -4, -6, TNF-α, and IFN-γ in monocytes [5]. Moreover, 348
hBDs and LL-37 have been reported to enhance the generation of IL-18 in keratinocytes [47]. 349
In this study, we demonstrated that the inducible defensins, hBD-2 and hBD-3, can induce the 350
release of other AMPs/ chemokines, i.e., CCL20, by HOECs. Therefore, the ability of hBDs to 351
promote the release of the antimicrobial chemokine CCL20 suggests the broad role of human 352
antimicrobial peptides in primary immune responses by oral epithelial cells. This regulatory 353
loop could act in a complimentary manner to provide added protection against microbial 354
assault. 355
To gain initial insight into the cellular pathways through which defensins induce 356
CCL20 release by HOECs, we focused on three major downstream MAPK cascades: ERK, 357
p38 and JNK. In cells, these kinase cascades act as signal sorters for a variety of upstream 358
signals before entering the nucleus. The p38, ERK, and JNK kinase cascades have been shown 359
to be involved in a large variety of cellular activities [3, 7]. Defensins have been shown to 360
activate primary human keratinocytes through p38 and ERK1/2 [47]. By focusing on these 361
three MAPKs, we demonstrated that both ERK1/2 and p38, but not JNK are required for hBD 362
induced CCL20 secretion by HOECs. The involvement of similar MAPKs was also observed 363
in the release of CCL20 by TNF-α or IL-1β stimulated oral epithelial cells. The p38 mitogen-364
activated protein kinase (MAPK) pathway plays an important role in the post-transcriptional 365
regulation of inflammatory genes and has been found to regulate both the translation and the 366
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

16
stability of inflammatory mRNAs. The mRNAs regulated by p38 are known to have AU-rich 367
elements (ARE) present in their 3’-untranslated regions [ 16,24] and thus, the presence of ARE 368
within the 3’UTR of CCL20 supports the involvement of p38 in it’s induction by defensins and 369
cytokines. Surprisingly, ERK1/2 and p38 inhibitors were found to have a negligible effect on 370
FnCW induced release of CCL20 by HOECs. Most recently, Yin and Chung [63] found the 371
lack of involvement of p38 in F. nucleatum induced CCL20 mRNA induction. At this stage, 372
however, it is not clear why, in contrast to induction of CCL20 by defensins and cytokines, the 373
induction of CCL20 by FnCW does not occur via p38, despite the fact that FnCW is capable of 374
causing rapid and transient activation of p38 MAPK [42] in HOECs. Nevertheless, these 375
observations further corroborate the fact that the induction of CCL20 by FnCW is not 376
dependent on defensins (hBD-2 and-3), TNF-α or IL-1β. 377
Kinetics of FnCW induced CCL20 and β-defensin release are distinct; i.e., CCL20 378
release peaks at 4 to 6 h [ Fig. 1B] by FnCW, while peak time is 24 h for hBD-2 [Fig 3C] and 379
48 h for hBD-3 [ Fig 3D]. Thus, coordinated stimulation of CCL20 and β-defensins by FnCW 380
on oral epithelial cells, might suggest a complementary activity in exerting the innate nature of 381
F. nucleatum in host defense. Since FnCW is a complex mixture of proteins, it is tempting to 382
speculate that different components within the FnCW fraction could be responsible for the 383
release of β-defensins and CCL20. We have recently identified a fusobacterium associated β-384
defensin inducer peptide (FAD-I), within FnCW, which is responsible for the induction of 385
hBD-2 in HOECs [29]. When we tested FAD-I’s ability to induce CCL20 release by HOECs, 386
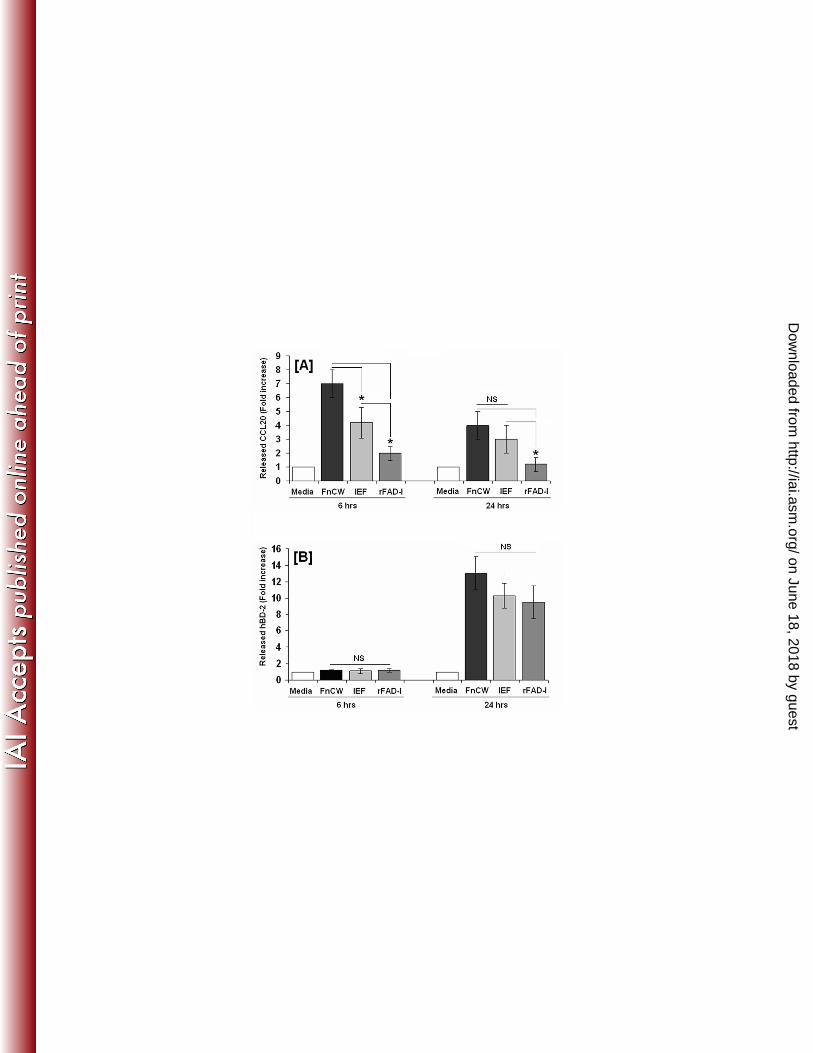
we found it to be negligible when compared to FnCW [Fig. 9A], while its ability to induce 387
hBD-2 release was comparable to that of FnCW [Fig. 9B]. However, an isoelecric focusing 388
(IEF) fraction of crude FnCW, from which FAD-I was further isolated [29], is capable of 389
inducing CCL20 release by HOECs [Fig. 9A], indicating that the CCL20 inducing factor(s) 390
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

17
is/are present in the IEF fraction. Results from Fig. 9 A, B appear to suggest this. Although the 391
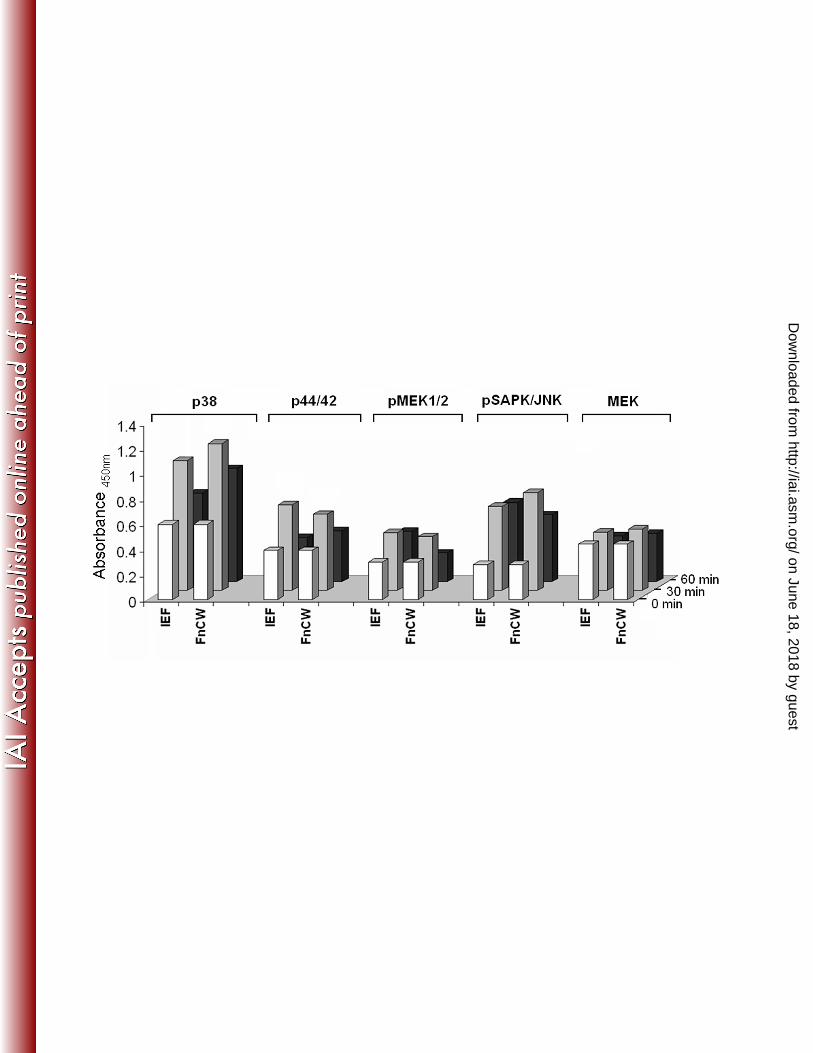
IEF fraction is a semi purified fraction from crude FnCW, the phosphorylation pattern of 392
different MAPKs elicited by the IEF fraction is similar to that elicited by crude FnCW [Fig. 393
10], and not surprisingly, the effectiveness of the IEF fraction in inducing CCL20 or hBD-2 is 394
close to that of FnCW. Isolating the CCL20 inducing factor, in conjunction with FAD-I may 395
one day be used to bolster the innate defenses of vulnerable mucosae. 396
Acknowledgement: We thank Drs. J. R. Blakemore, E. K. Schneider, W. S. Blood and F. 397
Faddoul for providing normal human oral tissue. We appreciate helpful discussions with Drs. 398
Zhimin Feng, Ge Jin and Jennifer Greene, Department of Biological Sciences. This work was 399
supported by National Institutes of Health Grants R01DE016334 (AW) and RO1DE018276 400
(AW.). 401
402 403 404 405 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

18
431 432 REFERENCES: 433 434 435
1. Aas, J.A., B.J. Paster, L.N. Stokes, I. Olsen, F.E. Dewhirst. 2005. Defining the 436
normal bacterial flora of the oral cavity. J Clin Microbiol 43: 5721–5732 437
2. Abiko, Y., M. Nishimura, K. Kusano, K. Nakashima, K. Okumura, T. Arakawa, T. 438
Takuma, and I. Mizoguchi. 2003. Expression of MIP-3alpha/CCL20, a macrophage 439
inflammatory protein in oral squamous cell carcinoma. Arch Oral Biol. 48(2):171-5. 440
3. Ballif, B. A., and J. Blenis. 2001. Molecular mechanisms mediating mammalian 441
mitogen-activated protein kinase (MAPK) kinase (MEK)-MAPK cell survival signals. 442
Cell Growth Differ. 12: 397–408 443
4. Boman, H.G. 2000. Innate immunity and the normal microflora. Immunol Rev 173:5-444
16 445
5. Chaly, Y.V., E.M. Paleolog, T.S. Kolesnikova, I.I. Tikhonov, E.V. Petratchenko, 446
and N.N. Voitenok. 2000. Neutrophil -defensin human neutrophil peptide modulates 447
cytokine production in human monocytes and adhesion molecule expression in 448
endothelial cells. Eur Cytokine Netw 11:257–266 449
6. Chang, K.P., et al. 2011. Overexpression of macrophage inflammatory protein-3a in 450
oral cavity squamous cell carcinoma is associated with nodal metastasis Oral Oncol. 451
47(2):108-113 452
7. Chang, L., and M. Karin. 2001. Mammalian MAP kinase signalling cascades. Nature 453
410: 37–40 454
8. Choi, S.C., et al. 2006 DA-9601, a standardized extract of Artemisia asiatica, blocks 455
TNF-alpha-induced IL-8 and CCL20 production by inhibiting p38 kinase and NF-456
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

19
kappaB pathways in human gastric epithelial cells. World J Gastroenterol. 12(30):4850-457
4858 458
9. Christ, A. D., and R. S. Blumberg. 1997. The intestinal epithelial cell: immunological 459
aspects. Springer Semin. Immunopathol. 18:449-461 460
10. Chung, W.O., and B.A. Dale. 2004. Innate immune response of oral and foreskin 461
keratinocytes: utilization of different signaling pathways by various bacterial 462
species.2004 72(1):352-8. 463
11. Cole, A. M., T. Ganz, A. M. Liese, M. D. Burdick, L. Liu, and R. M. Strieter. 2001. 464
Cutting edge: IFN-inducible ELR-CXC chemokines display defensin-like antimicrobial 465
activity. J. Immunol. 167:623-627 466
12. Cook, D. N., et al. 2000. CCR6 mediates dendritic cell localization, lymphocyte 467
homeostasis, and immune responses in mucosal tissue. Immunity 12:495-503 468
13. Crane-Godreau, M. A., and C. R. Wira. 2004. Effect of Escherichia coli and 469
Lactobacillus rhamnosus on macrophage inflammatory protein 3 alpha, tumor necrosis 470
factor alpha, and transforming growth factor beta release by polarized rat uterine 471
epithelial cells in culture. Infect. Immun. 72:1866-1873. 472
14. Dale, B. A. 2002 Periodontal epithelium: a newly recognized role in health and disease. 473
Periodontol 2000. 30:70-78 474
15. Dale, B. A., J.R. Kimball, S. Krisanaprakornkit, F. Roberts, M. Robinovitch, R. 475
O’Neal, E.V. Valore, T. Ganz, G.M. Anderson, and A. Weinberg. 2001. Localized 476
antimicrobial peptide expression in human gingiva. J Periodontal Res 36: 285 294 477
16. Dean, J.L., G. Sully, A.R. Clark, and J. Saklatvala. 2004. The involvement of AU-478
rich element-binding proteins in p38 mitogen-activated protein kinase pathway-479
mediated mRNA stabilization Cell Signal. 16(10):1113-1121 480
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

20
17. Dieu-Nosjean, M. C., et al. 2000. Macrophage inflammatory protein 3alpha is 481
expressed at inflamed epithelial surfaces and is the most potent chemokine known in 482
attracting Langerhans cell precursors. J. Exp. Med. 192:705–718 483
18. Eckmann, L., S.L. Reed, J.R. Smith, and M.F. Kagnoff. 1995. Entamoeba 484
histolytica trophozoites induce an inflammatory cytokine response by cultured human 485
cells through the paracrine action of cytolytically released interleukin-1 alpha. J Clin 486
Invest. 96(3):1269-1279 487
19. Fan, J., N.M. Heller, M. Gorospe, U. Atasoy, and C. Stellato. 2005.The role of post-488
transcriptional regulation in chemokine gene expression in inflammation and allergy. 489
Eur Respir J 26:933–947 490
20. Faveri, M, M.P.Mayer, M. Feres,L.C. de Figueiredo, F.E. Dewhirst, B.J. Paster. 491
2008. Microbiological diversity of generalized aggressive periodontitis by 16S rRNA 492
clonal analysis. Oral Microbiol Immunol 23:112–118. 493
21. Feng, Z. , G.R. Dubyak, M.M. Lederman, and A. Weinberg. 2006. Cutting edge: 494
human beta defensin 3--a novel antagonist of the HIV-1 coreceptor CXCR4. J Immunol. 495
177(2):782-786. 496
22. Feng, Z., and A. Weinberg. 2006. Role of bacteria in health and disease of periodontal 497
tissues. Periodontol 2000. 40:50-76. 498
23. Feucht, E.C., C.L. DeSanti, and A. Weinberg . 2003. Selective induction of human 499
beta-defensin mRNAs by Actinobacillus actinomycetemcomitans in primary and 500
immortalized oral epithelial cells Oral Microbiol Immunol. 18(6):359-63 501
24. Frevel, M.A., T. Bakheet, A.M. Silva, J.G. Hissong, K.S. Khabar, and B.R. 502
Williams.2003. p38 Mitogen-activated protein kinase-dependent and -independent 503
signaling of mRNA stability of AU-rich element-containing transcripts Mol Cell Biol. 504
23(2):425-436 505
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

21
25. Funderburg, N, M.M. Lederman, Z. Feng, M.G. Drage, J. Jadlowsky, C.V. 506
Harding , A. Weinberg , and S.F. Sieg. 2007. Human β-defensin-3 activates 507
professional antigen-presenting cells via Toll-like receptors 1 and 2. Proc Natl Acad Sci 508
U S A. 104(47):18631-18635 509
26. Ganz, T. 2003. Defensins: antimicrobial peptides of innate immunity. Nat Rev 510
Immunol 3: 710–720 511
27. Ghosh, S.K., T.A. Gerken, K.M. Schneider, Z. Feng, T.S. McCormick, and A. 512
Weinberg. 2007. Quantification of human beta-defensin-2 and -3 in body fluids: 513
application for studies of innate immunity. Clin Chem. 53(4):757-765 514
28. Gruber, A.R, J. Fallmann, F. Kratochvill, P. Kovarik, and H. L. Hofacker. 2011. 515
AREsite: a database for the comprehensive investigation of AU-rich elements Nucleic 516
Acids Res. 39 (Database issue):D66-9 517
29. Gupta, S., S.K. Ghosh, M.E. Scott, B. Bainbridge, B. Jiang, R.J. Lamont, T.S. 518
McCormick, and A. Weinberg. 2010. Fusobacterium nucleatum-associated beta-519
defensin inducer (FAD-I): identification, isolation, and functional evaluation. J Biol 520
Chem. 285 (47):36523-31. 521
30. Hosokawa, Y., T. Nakanishi, D. Yamaguchi, K. Takahashi, H. Yumoto, K. Ozaki, 522
and T. Matsuo. 2002. Macrophage inflammatory protein 3alpha-CC chemokine 523
receptor 6 interactions play an important role in CD4+ T-cell accumulation in 524
periodontal diseased tissue. Clin Exp Immunol. 128(3):548-54 525
31. Izadpanah, A., M. B. Dwinell, L. Eckmann, N. M. Varki, and M. F. Kagnoff. 2001. 526
Regulated MIP-3alpha/CCL20 production by human intestinal epithelium: mechanism 527
for modulating mucosal immunity. Am. J. Physiol. Gastrointest. Liver Physiol. 528
280:G710–G719 529
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

22
32. Ji, S, J.E. Shin, Y.S. Kim, J-E. Oh, B-M. Min, Y. Choi. 2009. Toll-like receptor 2 530
and NALP2 mediate induction of human beta-defensins by Fusobacterium nucleatum 531
in gingival epithelial cells. Infect Immun. 77(3):1044-52. 532
33. Ji, S., Y. Kim, B-M. Min, S.H. Han, Y. Choi. 2007. Innate immune responses of 533
gingival epithelial cells to nonperiodontopathic and periodontopathic bacteria. J 534
Periodont Res 2007; 42: 503–510 535
34. Jin, G., H.I. Kawsar, S.A. Hirsch, C. Zeng, X. Jia, Z. Feng, S.K. Ghosh, Q.Y. 536
Zheng, A. Zhou, T.M. McIntyre, and A. Weinberg. 2010. An antimicrobial peptide 537
regulates tumor-associated macrophage trafficking via the chemokine receptor CCR2, a 538
model for tumorigenesis. PLoS One. 5(6):e10993. 539
35. Kawsar, H.I., A. Weinberg, S.A. Hirsch, A. Venizelos, S. Howell, B. Jiang, and G. 540
Jin. 2009. Overexpression of human beta-defensin-3 in oral dysplasia: potential role 541
in macrophage trafficking. Oral Oncol. 45(8):696-702 542
36. Kennell, W., and S.C. Holt. 1990. Comparative studies of the outer membranes of 543
Bacteroides gingivalis, strains ATCC 33277, W50, W83, 381. Oral Microbiol Immunol. 544
5(3):121-130. 545
37. Kim, B. E., D. Y. M. Leung, J. E. Streib, M. Boguniewicz, Q. A. Hamid, and M. D. 546
Howell. 2007. Macrophage inflammatory protein 3alpha deficiency in atopic dermatitis 547
skin and role in innate immune response to vaccinia virus. J. Allergy Clin. Immunol. 548
119:457-463 549
38. Klausner, R.D., J.G. Donaldson, and J. Lippincott-Schwartz. 1992. Brefeldin A: 550
insights into the control of membrane traffic and organelle structure J Cell Biol. 551
116(5):1071-1080 552
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

23
39. Kleeff J., T. Kusama, D.L. Rossi, T. Ishiwata, H. Maruyama, H. Friess, M.W. 553
Büchler, A. Zlotnik, and M. Korc. 1999. Detection and localization of Mip-554
3alpha/LARC/Exodus, a macrophage proinflammatory chemokine, and its CCR6 555
receptor in human pancreatic cancer Int J Cancer. 81(4):650-7 556
40. Krisanaprakornkit, S., A. Weinberg, C.N. Perez, and B.A. Dale.1998. Expression of 557
the peptide antibiotic human beta-defensin 1 in cultured gingival epithelial cells and 558
gingival tissue Infect Immun. 66(9):4222-8. 559
41. Krisanaprakornkit, S., J.R. Kimball, A. Weinberg, R.P. Darveau, B. W. 560
Bainbridge, and B.A. Dale. 2000. Inducible expression of human ß-defensin 2 by 561
Fusobacterium nucleatum in oral epithelial cells: multiple signaling pathways and role 562
of commensal bacteria in innate immunity and the epithelial barrier. Infect Immun 68: 563
2907–2915 564
42. Krisanaprakornkit, S., J.R. Kimball, and B.A. Dale. 2002. Regulation of human 565
beta-defensin-2 in gingival epithelial cells: the involvement of mitogen-activated 566
protein kinase pathways, but not the NF-kappaB transcription factor family. J Immunol. 567
168(1):316-24 568
43. Lin, T.J., L.H. Maher, K. Gomi, J.D. McCurdy, R. Garduno, J.S. Marshall. 2003. 569
Selective early production of CCL20, or macrophage inflammatory protein 3alpha, by 570
human mast cells in response to Pseudomonas aeruginosa. Infect Immun. 71(1):365-73 571
44. Medzhitov, R., and C. A. Janeway, Jr. 1997. Innate immunity: impact on the 572
adaptive immune response. Curr. Opin. Immunol. 9:4–9 573
45. Miller, S. G., L. Carnell, and H.P.H. Moore. 1992. Post-Golgi membrane traffic: 574
brefeldin A inhibits export from distal Golgi compartments to the cell surface but not 575
recycling. J. Cell Biol. 118, 267-283 576
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

24
46. Nakayama, T., J. Shirane, K. Hieshima, M. Shibano, M. Watanabe, Z. Jin, D. 577
Nagakubo, T. Saito, Y. Shimomura, and O. Yoshie. 2006. Novel antiviral activity of 578
chemokines. Virology 350:484-492 579
47. Niyonsaba, F. H. Ushio, I. Nagaoka, K. Okumura, and H. Ogawa. 2005. The human 580
beta-defensins (-1, -2, -3, -4) and cathelicidin LL-37 induce IL-18 secretion through 581
p38 and ERK MAPK activation in primary human keratinocytes.J Immunol. 582
175(3):1776-1784 583
48. Paster, B.J., I. Olsen, J.A. Ass, F.E. Dewhirst. 2006. The breadth of bacterial 584
diversity in the human periodontal pocket and other oral sites. Periodontol 2000 42:80–585
87 586
49. Pernet, I., C. Reymermier, A. Guezennec, J.E. Branka, J. Guesnet, E. Perrier, C. 587
Dezutter-Dambuyant, D. Schmitt, and J. Viac. 2003. Calcium triggers beta-defensin 588
(hBD-2 and hBD-3) and chemokine macrophage inflammatory protein-3 alpha (MIP-589
3alpha/CCL20) expression in monolayers of activated human keratinocytes. Exp 590
Dermatol. 12(6):755-760 591
50. Peyret-Lacombe, A., G. Brunel, M. Watts, M. Charveron, and H. Duplan. 2009. 592
TLR2 sensing of F. nucleatum and S. sanguinis distinctly triggered gingival innate 593
response. Cytokine. 46(2):201-210. 594
51. Reibman, J., Y. Hsu, L.C. Chen, B. Bleck, T. Gordon. 2003. Airway epithelial cells 595
release MIP-3alpha/CCL20 in response to cytokines and ambient particulate matter Am 596
J Respir Cell Mol Biol. 28(6):648-654 597
52. Röhrl, J., D. Yang, J.J. Oppenheim, and T. Hehlgans. 2010. Human beta-defensin 2 598
and 3 and their mouse orthologs induce chemotaxis through interaction with CCR2 J 599
Immunol. 184(12):6688-6694. 600
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

25
53. Rossi, D. L., A. P. Vicari, K. Franz-Bacon, T.K. McClanahan, and A. Zlotnik. 601
1997. Identification through bioinformatics of two new macrophage proinflammatory 602
human chemokines: MIP-3alpha and MIP-3beta. J. Immunol. 158:1033–1036. 603
54. Ruth, J.H., S. Shahrara, C.C. Park, J.C. Morel , P. Kumar, S. Qin, and A.F. 604
Koch. 2003. Role of macrophage inflammatory protein-3alpha and its ligand CCR6 in 605
rheumatoid arthritis. Lab Invest. 83(4):579-588 606
55. Schutyser, E., S. Struyf, and J. Van Damme. 2003. The CC chemokine CCL20 and 607
its receptor CCR6Cytokine Growth Factor Rev. 14(5):409-426 608
56. Socransky, S. S., A.D. Haffajee, M.A. Cugini, C. Smith, and R.L. Kent, Jr. 1998. 609
Microbial complexes in subgingival plaque. J Clin Periodontol 25:134–144 610
57. Tanaka, Y., T. Imai, M. Baba, I. Ishikawa, M. Uehira, H. Nomiyama, and O. 611
Yoshie. 1999. Selective expression of liver and activation-regulated chemokine 612
(LARC) in intestinal epithelium in mice and humans. Eur. J. Immunol. 29:633–642 613
58. Tohyama, M., Y. Shirakara, K. Yamasaki, K. Sayama, and K. Hashimoto. 2001. 614
Differentiated keratinocytes are responsible for TNF-alpha regulated production of 615
macrophage inflammatory protein 3alpha/CCL20, a potent chemokine for Langerhans 616
cells. J. Dermatol. Sci. 27:130–139 617
59. Vankeerberghen, A., H. Nuytten, K. Dierickx, M. Quirynen, J.J. Cassiman, and H. 618
Cuppens. 2005. Differential Induction of Human Beta-Defensin Expression by 619
Periodontal Commensals and Pathogens in Periodontal Pocket Epithelial Cells J 620
Periodontol. 76(8):1293-303. 621
60. Yang, D., et al. 1999. Beta-defensins: linking innate and adaptive immunity through 622
dendritic and T cell CCR6. Science 286:525-528 623
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

26
61. Yang, D., Q. Chen, D.M. Hoover, P. Staley, K.D. Tucker, J. Lubkowski, and J.J. 624
2003. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity J 625
Leukoc Biol. 74(3):448-455. 626
62. Yin, L, and B.A. Dale. 2007. Activation of protective responses in oral epithelial cells 627
by Fusobacterium nucleatum and human beta-defensin-2. J Med Microbiol. 56(7):976-628
987 629
63. Yin, L, and W.O. Chung. 2011. Epigenetic regulation of human β-defensin 2 and CC 630
chemokine ligand 20 expression in gingival epithelial cells in response to oral bacteria 631
Mucosal Immunol. doi: 10.1038/mi.2010.83 632
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

27
Figure Legends: 633
Fig. 1: Kinetics of FnCW induced release of CCL20 by HOECs: [A] HOECs (mixed 634
donors; N=3) were challenged with FnCW (10 μg/ml) for indicated time periods and the 635
amount of CCL20 in cell free supernatants were measured by ELISA. (Results are the mean 636
[+SD] of three independent experiments.) [B] Fold changes in CCL20 release by HOECs 637
(mixed donors; N=3) after challenge with FnCW (10μg/ml) for indicated time periods. (Results 638
are the mean [+SD] of three independent experiments) [C] Interpersonal variability in CCL20 639
release by HOECs from five different donors after challenge with FnCW (10 μg/ml) for 6hrs. 640
(Results are the mean [+SD] of three independent ELISA measurements) 641
642 Fig. 2: [A] Effect of actinomycin D (Act D) and cyclohexamide (CHX) on FnCW induced 643
release of CCL20 by HOECs: HOEC monolayers (mixed donor; N=3) were pretreated with 644
actinomycin D or cyclohexamide for 5 min and then incubated with or without FnCW (10 645
μg/ml) for an additional 6 h. As a negative control, cells were incubated with media or media 646
with vehicle control (DMSO) for actinomycin D. CCL20 levels in cell free supernatants were 647
measured by ELISA. [B] and [C] Effect of brefeldin A (BFA) on CCL20 release by HOECs. 648
HOEC monolayers (mixed donor; N=3) were pretreated with BFA (10 μg/ml) for 5 min, 649
followed by incubation with or without FnCW (10 μg/ml) for an additional 6 h. As a negative 650
control, cells were incubated with media alone or media with vehicle control (ethanol) for BFA. 651
CCL20 levels in cell free supernatants [B] and cell lysates [C] were measured by ELISA. 652
Results are the mean (+ SD) of three independent experiments (* p<0.05). For comparison of 653
CCL20 content in supernatants and in cell pellets the levels of CCL20 is expressed as ng/well 654
instead of ng/ml. One ml of media per well was used in a six well format. 655
656
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

28
Fig. 3: Kinetics of FnCW induced release of hBD-2 and hBD-3 by HOECs: [A] and [B], 657
HOEC monolayers (mixed donor; N=3) were challenged with FnCW (10 μg/ml) for the 658
indicated time periods, and the levels of hBD-2 and hBD-3 in cell free supernatants were 659
determined by ELISA. . (Results are the mean [+SD] of three independent experiments) [C] 660
and [E] Fold changes (calculated from figures A and B) in hBD-2 and -3 release by HOECs 661
(mixed donor; N=3) after FnCW challenge for indicated time periods. (Results are the mean 662
[+SD] of three independent experiments) [D] and [F] Interpersonal variability in hBD-2 and-3 663
release by HOECs from five different donors after FnCW treatment for 24 hrs. (Results are the 664
mean [+SD] of three independent ELISA measurements) 665
Fig. 4: HBD-2 induced release of CCL20 by HOECs: [A] HOEC monolayers (mixed donor; 666
N=3) were challenged with hBD-2 (0.1 to 10 μg/ml) for 6 hrs; released CCL20 in cell-free 667
supernatants was determined by ELISA, followed by calculation of fold increase above 668
baseline. [B] HOEC monolayers (mixed donor; N=3) were challenged with hBD-2 (5 μg/ml) 669
for indicated time periods and the amount of released CCL20 in cell-free supernatants was 670
determined by ELISA. Results are the mean (+ SD) of three independent experiments [* 671
indicates p<0.05 with respect to control (no treatment)] 672
673 Fig. 5: HBD-3 induced up regulation of CCL20 transcript in HOECs ([A] and [B]). HOEC 674
monolayers (mixed donors; N=3) were treated with indicated amounts of synthetic hBD-3 for 6 675
hrs. Total RNAs from both treated and untreated cells were isolated, the hBD-3 transcripts 676
were amplified by RT-PCR [A] and the fold changes were quantified by real-time PCR [B]. 677
HK5, human keratin 5 has been used as housekeeping gene. HBD-3 induced release of 678
CCL20 by HOECs [C]: HOEC monolayers (mixed donors; N=3) were treated with indicated 679
amounts of synthetic hBD-3 for 6 hrs and the levels of CCL20 in the cell free supernatants 680
were determined by ELISA, followed by fold change determinations. Kinetics of hBD-3 681
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

29
induced CCL20 release by HOECs [D]. HOEC monolayers (mixed donors; N=3) were 682
treated with hBD-3 (5μg/ml) for indicated time periods and release of CCL20 in cell free 683
supernatants for both treated and untreated cells was determined by ELISA. Results for B-D 684
are the mean (+ SD) of three independent experiments [* = p<0.05]. 685
686 Fig. 6: TNF-α and IL-1β induced release of CCL20 by HOECs. HOEC monolayers 687
(mixed donors; N=3) were separately treated with TNF-α (10 ng/ml), IL-1β (10 ng/ml), TNF-688
α+ IL-1β (5 ng/ml each) or media alone for indicated time periods, and the levels of released 689
CCL20 in the spent supernatants were determined by ELISA. Results are the mean (+ SD) of 690
three independent experiments. [* indicates p<0.05 with respect to control (no treatment)] 691
Fig. 7: Inhibition of ERK, p38 and JNK in hBD-2, hBD-3, TNF-α and IL-1β induced 692
release of CCL20 by HOECs. HOEC monolayers (mixed donor, N=3) were pretreated with 693
inhibitors of ERK [UO126 (20μM) or PD98059 (20μM)], p38 [SB203580 (20μM)] or JNK 694
specific inhibitors [SP600125 (20μM)], respectively, for 1 hr, followed by treatment with [A] 695
hBD-2 (5 μg/ml) or [B] hBD-3 (5 μg/ml) or [C] TNFα (10 μg/ml) or [D] IL1β (10 μg/ml) 696
for an additional 18 hrs. DMSO was used as the negative control. CCL20 levels in the spent 697
supernatants were measure by ELISA. Results are the mean (+ SD) of three independent 698
experiments. (* = p<0.05) 699
Fig. 8: PathScan® MAP Kinase Multi-Target Sandwich ELISA: HOEC monolayers 700
(mixed donor, N=3) were treated with either TNFα (10 ng/ml), IL-1β (10 ng/ml), hBD-2 (5 701
μg/ml) or hBD-3 (5 μg/ml) for 0, 30 and 60 minutes, respectively. The cell lysates were 702
analyzed for the indicated target, using PathScan® MAP Kinase Multi-Target Sandwich 703
ELISA kit (Cell Signaling, MA) according to the vendor’s instructions. 704
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

30
Fig. 9: Comparison of FnCW, isoelectric focusing (IEF) fraction of FnCW and 705
recombinant FAD-I induced CCL20 and hBD-2 release. HOECs (mixed donor, n=3) were 706
treated with media alone, FnCW (10 μg/ml), IEF fraction (10 μg/ml) or recombinant FAD-I 707
(rFAD-I) (10 μg/ml) for indicated time periods. The levels of CCL20 [A] and hBD-2 [B] in 708
cell free supernatants were measured by ELISA. The IEF fraction and rFAD-I were prepared as 709
described by Gupta et al [29]. (* = p<0.05 and NS= none significant) 710
Fig. 10: PathScan® MAP Kinase Multi-Target Sandwich ELISA: HOEC monolayers 711
(mixed donor, N=3) were treated with either IEF (10 μg/ml ) or FnCW (10 μg/ml) for 0, 30 712
and 60 minutes, respectively. The cell lysates were analyzed for the indicated target, using 713
PathScan® MAP Kinase Multi-Target Sandwich ELISA kit (Cell Signaling, MA) according to 714
the vendor’s instructions. 715
716
on June 18, 2018 by guesthttp://iai.asm
.org/D
ownloaded from