
Draft - University of Toronto T-Space › bitstream › 1807 › ... · Draft 3 Homocysteine and...
19
Draft Connecting homocysteine and obesity through pyroptosis, gut microbiome, epigenetics, peroxisome proliferator- activator receptor γ (PPARγ) and zinc finger protein 407 (Zfp407) Journal: Canadian Journal of Physiology and Pharmacology Manuscript ID cjpp-2018-0037.R1 Manuscript Type: Review Date Submitted by the Author: 29-Mar-2018 Complete List of Authors: Laha , Anwesha ; University of Louisville School of Medicine, Physiology Majumder, Avisek; University of Louisville School of Medicine, Physiology Singh, Mahavir; University of Louisville School of Medicine, Physiology Tyagi , Suresh C. ; University of Louisville School of Medicine, Physiology Is the invited manuscript for consideration in a Special Issue: IACS EU Section Keyword: PGC-1α, Zinc finger protein, DNA methylation, Lipopolysaccharide, Fatty acid metabolism https://mc06.manuscriptcentral.com/cjpp-pubs Canadian Journal of Physiology and Pharmacology
Transcript of Draft - University of Toronto T-Space › bitstream › 1807 › ... · Draft 3 Homocysteine and...

Draft
Connecting homocysteine and obesity through pyroptosis,
gut microbiome, epigenetics, peroxisome proliferator-activator receptor γ (PPARγ) and zinc finger protein 407
(Zfp407)
Journal: Canadian Journal of Physiology and Pharmacology
Manuscript ID cjpp-2018-0037.R1
Manuscript Type: Review
Date Submitted by the Author: 29-Mar-2018
Complete List of Authors: Laha , Anwesha ; University of Louisville School of Medicine, Physiology Majumder, Avisek; University of Louisville School of Medicine, Physiology Singh, Mahavir; University of Louisville School of Medicine, Physiology Tyagi , Suresh C. ; University of Louisville School of Medicine, Physiology
Is the invited manuscript for consideration in a Special
Issue: IACS EU Section
Keyword: PGC-1α, Zinc finger protein, DNA methylation, Lipopolysaccharide, Fatty
acid metabolism
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
1
Page 1 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Connecting homocysteine and obesity through pyroptosis, gut microbiome, epigenetics,
peroxisome proliferator@Hactivator receptor γ (PPARγ) and zinc finger protein 407 (Zfp407)
Anwesha Laha, Avisek Majumder, Mahavir Singh, Suresh C.Tyagi
Department of Physiology, University of Louisville, Louisville, Kentucky.
Corresponding authors: Anwesha Laha ([email protected])
or Mahavir Singh ([email protected])

Draft
2
Abstract: Although homocysteine (Hcy), a part of the epigenome, contributes to cell death
by pyroptosis and decreases PPARγ levels, the mechanisms are unclear. Hcy is found in
high concentrations in the sera of obese individuals which can elicit an immune response as
well by hypermethylating CpG islands of specific gene promoters, a marker of epigenetics.
Hcy has also been established to chelate divalent metal ions like Cu+2 and Zn+2, but this role
of Hcy has not been established in relationship with obesity. It has been known for a while
that PPARγ dysregulation results in various metabolic disorders including glucose and lipid
metabolism. Recently, zinc finger protein 407 (Zfp407) is reported to regulate PPARγ target
gene expression without affecting PPARγ transcript and protein levels by synergistically
working with PPARγ. However, the mechanism(s) of this synergy, as well as other factors
contributing to or inhibiting this synergism have not been proven. This review suggests that
Hcy contributes to pyroptosis; changes gut microbiome, alter PPARγ-dependent
mechanism(s) via Zfp407 mediated upregulated adipogenesis and misbalanced fatty acid
metabolism leading that can predispose to obesity and consequently the obesity-related
metabolic disorders.
Keywords: PGC-1α, zinc finger protein, DNA methylation, lipopolysaccharide, fatty acid
metabolism, type 2 diabetes, insulin.
Page 2 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
3
Homocysteine and obesity
Obesity is one of the leading causes of health complications like type 2 diabetes (T2D),
stroke, hypertension, cardiovascular diseases, and even some cancers. As of 2014, the
World Health Organization (WHO) stated that 1 in 5 individuals in the United States died due
to obesity-related health disorders. Per Center for Disease Control, 20% of the population in
every state in the U.S. is obese (WHO, Centre for Disease Control). Obesity is responsible
for causing the insulin resistance leading to dysregulated lipid metabolism by reducing
adipogenesis and increasing free fatty acids (FFA) in the blood (Harmancey and Smih 2007;
Kim et al. 2000; Odegaard and Chawla 2013) .
Homocysteine is a non-protein amino acid that is generated during methionine metabolism.
The fate of homocysteine is either remethylation to methionine or conversion to cysteine
through the transsulfuration pathway. Folic acid works as the cofactor essential for
remethylation of homocysteine to methionine (Barshop 2012). The two essential enzymes
required for Hcy to cysteine transsulfuration are cystathionine β-synthase (CBS) and
cystathionine γ-lyase (CSE) (Figure 1). In the rate-limiting step, CBS catalyzes the synthesis
of the intermediate, cystathionine from Hcy and CSE converts cystathionine to cysteine.
Hyperhomocysteinemia (HHcy) is a medical condition that is characterized by the
homocysteine's level which is more than 15µmol/L homocysteine (Hcy) in the blood due to
the body’s inability to metabolize homocysteine properly (Guo et al. 2009). HHcy is
responsible for a number of pathophysiological conditions like atherosclerosis, hypertension,
vascular dysfunction leading to coronary heart diseases and myocardial infarction (Sen et al.
2010). Deficiencies in folic acid, vitamins B6 and B12 and mutations in CBS gene have been
attributed to HHcy in infants and adults (Huemer et al. 2015). Dietary supplementation of
Folic acid, Vitamin B6 (pyridoxal-5’-phosphate) and Vitamin 12 (Cobalamine) (Maron and
Loscalzo 2009) are used as therapy for HHcy, a defined therapy for congenital HHcy has not
yet been established.
HHcy is also one of the major contributing factors in obesity. Excess Hcy suppresses
preadipocyte differentiation (Mentese et al. 2016) and lipolysis in primary adipocytes (Wang
et al. 2011). Decreased lipolysis in adipocytes results in reduced levels of FFA for energy
expenditure and accumulation of triglycerides in adipose tissues. Concurrently, plasma
homocysteine levels in obese individuals with hypertension have been found to be much
more than in non-obese individuals with hypertension (Konukoglu et al. 2003). Obese
individuals with high Hcy levels are predisposed to cryptogenic strokes (Vaya et al. 2011)
and insulin resistance (Vaya et al. 2012). Although it is well established that Hcy is a
contributing factor to obesity and obesity-related health risks, the exact role of Hcy in altering
the related molecular mechanisms are not yet studied. We are therefore providing avenues
Page 3 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
4
for uncovering these intricate molecular mechanisms involving the role of Hcy in obesity
paradigm.
Pyroptosis is an inflammation-mediated programmed cell death characterized by cell
swelling and membrane disruption. Pyroptosis occurs upon activation of Caspase 1 followed
by the assembly and formation of the NLRP3 inflammasome (Lamkanfi 2011). Pyroptosis
mediated inflammatory response has been reported to be elicited not only by infections via
pathogens but also by the overexpression of certain macromolecules. High Hcy levels can
elicit pyroptosis in the presence and absence of lipopolysaccharide (LPS) (Xi et al. 2016).
LPS which is usually present in the membranes of Gram-negative bacteria and can elicit an
immune response. This is found in high concentrations in the plasma of obese individuals
consuming a high-fat diet (Erridge et al. 2007) (Troseid et al. 2013). Obese adipocytes have
been reported to overexpress NLRP3 inflammasome and undergo stress-induced pyroptosis
(Giordano et al. 2013). It is also predicted that pyroptosis may be a mode of cell death in
HHcy mediated age-related macular degeneration (AMD), a disorder also prevalent among
obese individuals (Adams et al. 2011) (Singh and Tyagi 2017).
Interestingly, the gut microbiome has been shown to play a major role in the healthy living of
an individual. The gut microbiome consists of innumerable bacteria and virus and their
numbers, their genomes, and the environment are dynamic with respect to an individual's
diet. This normal gut microbiota composition is heavily altered due to perpetual consumption
of high-fat diet and that can contribute to obesity and type 2 diabetes (Baothman et al. 2016;
Wolf and Lorenz 2012). This allows an increased number of Gram-negative bacteria
expressing LPS and thus, capable of eliciting pyroptosis. The exact mechanism of obese
adipocyte stress induced pyroptosis is not clear allowing us to hypothesize that Hcy/LPS is
mediating pyroptosis in the obese adipocytes due to the gut microbiota imbalance. A
schematic representation of this hypothesis is shown in figure 2. There are several studies
which suggest that gut microbiota transplant (GMT) will be an effective therapy for obesity
and obesity related disorders, there are no concrete evidence of the effectiveness of GMT.
PPARγ, PGC-1α and their relationship with homocysteine through epigenetics
Peroxisome proliferator-activator receptor-gamma (PPARγ) is a master regulator of lipid
metabolism and adipogenesis. It belongs to the nuclear hormone receptor superfamily and is
highly expressed in adipose tissues (Chawla et al. 1994; Tontonoz et al. 1995). Its
heterodimerization with Retinoic X Receptor (RXR) regulates target gene expressions. In the
absence of a ligand, co-repressors are bound to the PPARγ-RXR complex keeping it
transcriptionally inactive. When specific ligands bind to the ligand binding domain of PPARγ,
a conformational change releases the bound corepressors. This allows co-activators like
PGC-1α and other coactivators to be recruited to the target genes promoting PPARγ-
Page 4 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
5
mediated transcription (Costa et al. 2010; Murphy and Holder 2000). Lack of PPARγ
functions have been shown to reduce preadipocyte differentiation and adipogenesis both in
vitro and in vivo (Murphy and Holder 2000). Conversely, adipogenesis has been shown to be
downregulated when its expression is reduced, implying that it is essential for normal
adipogenesis facilitating normal lipid metabolism (Kim et al. 2001; Rosen et al. 1999).
Although numerous studies reveal the various pathways and regulators of PPARγ mediated
target gene transcription, several regulators and their roles in transcription are still unknown.
Therefore, it is essential to elucidate the mechanisms by which other regulators of PPARγ
mediated transcription affects the expression of target genes. PPARγ interacts with many
coactivators to carry out the transcription of its target genes, such as coactivator-1α (PGC-
1α) which is a key PPAR coactivator regulating the target gene expressions. The LXXLL
domain of activated PGC-1α directly interacts with parts of PPARγ’s DNA binding domain
and hinge domain (Puigserver et al. 1998). This allows the recruitment of histone
acetyltransferase (HATs) containing proteins maintaining PPARγ’s transcriptional activity
(Puigserver 2005). PGC-1α has been reported to play a vital role in regulating essential
biological processes like mitochondrial biogenesis and fatty acid β-oxidation (Vega et al.
2000)
Homocysteine has been established to play a role in epigenetics, especially DNA
hypermethylation due to increased concentration of S-adenosylmethionine (SAM), a methyl
group donor (Figure 1) (Majumder et al. 2017). This eventually contributes to abnormal
silencing of specific genes involved in essential cellular processes and leads HHcy related
pathophysiologies (Kalani et al. 2013). Previous reports have shown that elevated
concentrations of Hcy results in hypermethylation of the CpG islands of the PPARγ promoter
regions resulting in reduced gene expression (Yideng et al. 2008). Similarly, high Hcy levels
also contribute to cardiomyopathy by attenuating PPARγ gene expression and this effect is
alleviated by its agonists and ligands (Mishra et al. 2010). From these and other reports, we
believe that Hcy is behaving as a PPARγ antagonist. This antagonism could dysregulate
fatty acid metabolism through PPARγ governed genes and hence could promote obesity-
related pathophysiological conditions. This hypothesis will open various avenues for
researchers to identify these factors contributing to the antagonism between PPARγ and
Hcy.
Chromatin remodeling and epigenetics play an important role in the inherent functioning of
both, PPARγ and PGC-1α. PPARγ has been reported to go through extensive chromatin
remodeling during the early stages of adipogenesis (Lee and Ge 2014). Also,
CCAAT/Enhancer Binding Proteins (C/EBPs) are a group of leucine zipper transcrition
factors that have been associated with adipogenesis in the presence of PPARγ (Lane et al.
1999) (Rosen et al. 2002). Evidence suggests that the association of C/EBPα with the
Page 5 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
6
PPARγ promoter allows increased transciption and that inhibitors disrupt this association.
This in turn stops PPARγ transcription through the recuitment of histone deacetylases
(HDACs) (Zuo et al. 2006) These suggested pathways of chromatin remodelling of the
PPARγ promoter allow transcription factors to interact with it allowing transcription of genes.
Like PPARγ, PGC-1α too has a negative correlation with Hcy. Enhanced PGC-1α
expression upregulates genes involved in Hcy metabolism (Li et al. 2009). Likewise, Hcy
contributes to attenuated PGC-1α expression leading to mitochondrial dysfunction (Veeranki
et al. 2015). It is thus clear that the synergism of PPARγ and PGC-1α is antagonized by
HHcy. Just as the PPARγ-PGC-1α synergism may be regulated by other unknown factors,
the same unknown factors may be antagonized by HHcy and these correlations require
further analyses.
PGC-1α expression too has been shown to be epigenetically regulated at its promoter.
Histone modifications can either be transcriptionally activating (H3K4me3, H3K27ac) or
repressing (H3K27me3) for their targets. In a recent study, PGC-1α promoter b (PGC-1α-b)
has been shown to have increased H3K4me3 in mice skeletal muscles upon exercise
(Lochmann et al. 2015). There is also evidence that exercise can mitigate the effects of Hcy
in skeletal muscles and white adipocytes (Winchester et al. 2014). Therefore, Hcy could
potentially silence PGC-1α expression through epigenetic histone remodeling and exercise
may possibly mitigate this effect of Hcy thereby allowing enhanced PGC-1α expression
however this would further work to investigate these possibilities. Both PGC-1α and PPARγ
have been reported to work in tandem and show close relationships as demonstrated in a
study wherein their expression levels were decreased in conditions such as obesity and
insulin resistance. Interestingly, physical training in form of exercise could restore their levels
proving the beneficial effects of physical activity in humans (Ruschke et al. 2010).
PPARγ and Zfp407
In a recent study, PPARγ target genes critical for lipid metabolism were shown to be
regulated by Zinc Finger Protein 407 (Zfp407) (Buchner et al. 2015). Knockdown of Zfp407
decreased expression of target genes. This was shown in an RNA-seq experiment where
expression of several PPARγ regulated genes involved in lipid metabolism were analyzed
after the knockdown of Zfp407 (Buchner et al. 2015). However, when PPARγ levels were
analyzed after the knockdown of Zfp407, there was no change in either the transcript or
protein levels, indicating that Zfp407 does not regulate PPARγ expression. In a luciferase
reporter assay when HEK239T cells were co-transfected with PPARγ and Zfp407, luciferase
activity of the co-transfected cells increased 2-fold compared to PPARγ or Zfp407
transfections alone suggesting that Zfp407 synergistically works with PPARγ. However, the
underlying mechanisms of this regulation remain unknown. Although PPARγ and Zfp407
Page 6 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
7
have been shown to work synergistically, other factors may influence their activities
(Buchner et al. 2015) and these other factors are yet to be elucidated. Further studies can
help us better understand the mechanism of Zfp407 to regulate PPARγ target gene
expression and determine the key players involved in regulating their activities. This can
eventually lead to the identification of potential drug targets for diabetes and obesity-related
diseases.
Potential functional similarity between Zfp407 and PRDM16
Positive Regulatory Domain Containing 16 (PRDM16) is a zinc finger-containing protein with
10 zinc finger domains that has been shown to interact with both PPARγ and PGC-1α
(Hondares et al. 2011; Kajimura et al. 2009; Seale et al. 2008). Together they form a
complex triggering the PPAR-mediated transcriptional program required for brown
adipogenesis (Fruhbeck et al. 2009). PRDM16 also binds to the PGC-1α promoter to induce
PGC-1α transcription under increased levels of active PPARα in brown adipocytes
(Hondares et al. 2011). Thus, PRDM16 performs a dual function of a transcription
coactivator for PPARs and a transcription factor for PGC-1α in brown adipogenesis.
Although the mechanisms and proteins involved in white adipocyte differentiation have been
extensively studied, many interacting partners of PPARγ and PGC-1α in the transcriptional
program of white adipogenesis still remain unknown. We hypothesize that Zfp407 could
potentially play the role of PRDM16, interacting with PPARγ and PGC-1α during
adipogenesis along with inducing PGC-1α transcription during adipogenesis in white adipose
tissue. Zfp407 could also possibly induce PGC-1α expression and interacting with PPARγ
and PGC-1α together, playing a role in their complex formation to maintain PPARγ’s
transcriptional activity which needs further analyses (Figure 3). Zfp407, transcribed by
ZNF407 on chromosome 18, belongs to the C2H2 class of zinc finger proteins and consists
of 22 zinc finger domains (Kambouris et al. 2014; Ren et al. 2013). The C2H2 (Cys-Cys-His-
His) class of zinc finger proteins are DNA binding proteins. They generally work as
transcription factors due to their higher binding affinity to DNA than to RNA. We therefore
believe that Zfp407 may also function as a transcription factor by directly interacting with
DNA through its C2H2 domain resulting in PPARγ target gene expression (Iuchi 2001; Laity
et al. 2001). The sequences could potentially be the promoters of PPARγ coactivators or
enhancers of the PPARγ target genes and Zfp407 is directly interacting with DNA sequences
that could possibly be associated with transcriptional activity. However, there is no concrete
evidence of Zfp407’s interaction with DNA or the nucleotide sequences interacting with
Zfp407 and we therefore pave the way for researchers to identify these DNA sequences
(Figure 3)
Page 7 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
8
Homocysteine can potentially regulate zinc finger proteins and contribute to obesity
It has been reported earlier and well established that the divalent cation copper (Cu+2) can
form a complex with Hcy during HHcy. This results in copper homeostasis imbalance due to
an increase in blood Cu+2 concentration and decreased intracellular Cu+2 concentration
(Dong et al. 2013). Zinc, like Cu+2 is a divalent metal ion and an essential mineral which is a
part of a zinc finger proteins working as transcription factors and involved in numerous other
cellular processes. Zn+2’ deficiency has been attributed to DNA instability also (Jurowski et
al. 2014) and can potentially result in their subsequent protein synthesis inhibition. Zn+2
deficiency could also lead to the instability and disintegration of zinc finger proteins since Zn
bonds with cysteine and histidine. As mentioned earlier, cysteine is a product of Hcy
transsulfuration metabolism. Therefore, Zn and cysteine deficiency together can exacerbate
the stability of zinc finger proteins. Similar to Cu+2, previous studies showed that serum Zn+2
and Hcy concentrations are positively correlated and serum Hcy and tissue Zn+2
concentrations are negatively correlated (Hong et al. 2000) (Jing et al. 2015). It is therefore
possible that excess serum Hcy is forcing Zn+2 to conjugate with excess Hcy thereby
reducing the intracellular Zn+2 concentrations. This hinders synthesis of Zn+2 containing
proteins including transcription factors like Zfp407 and PRDM16. In this context, it is not
surprising that HHcy can therefore contribute and hence promote obesity by suppressing
PPARγ target genes by chelating cytosolic Zn+2. This could eventually downregulate Zfp407
expression and that in turn could repress genes which are involved in insulin sensitivity and
fatty acid metabolism. We are providing a platform to researchers to establish further that
Hcy is an inhibiting factor to zinc finger protein functioning. In fact, studies revealed that a
sizable number of zinc finger proteins could function as key transcriptional regulators that
are involved in the process of adipogenesis and as a result of this observations they are
being considered as promising human obesity drug targets (Wei et al. 2013) (Figure 4).
Concluding statement
In our current review, we have proposed different mechanisms by which elevated Hcy levels
can contribute to obesity and obesity-related disorders. We have suggested three different
routes of Hcy to exhibit different obesity like phenotypes. The first route is obesity-related
inflammation through pyroptosis aided by Hcy, the second is Hcy mediated DNA methylation
and therefore, silencing of genes essential for normal FA metabolism and the third route is
by modulating cellular Zn+2 concentration to inhibit PPARγ target genes involved in lipid
metabolism. All three mechanisms either can work in tandem or individually. In this review,
we have opened different avenues for researchers to uncover the mode of actions of Hcy in
obesity and its related disorders as well as design drugs to alleviate these important
metabolic disorders.
Page 8 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
9
Conflict of interest: None
Acknowledgements: We would like to thank all the members of the laboratory for their
continuous support and encouragement. Part of this work was supported by grants from the
National Institute of Health (Heart, Lung, and Blood Institute; No. HL-74815 and HL-139047).
References
Adams, M.K., Simpson, J.A., Aung, K.Z., Makeyeva, G.A., Giles, G.G., English, D.R., Hopper, J.,
Guymer, R.H., Baird, P.N., and Robman, L.D. 2011. Abdominal obesity and age-related macular
degeneration. Am J Epidemiol 173(11): 1246-1255. doi: 10.1093/aje/kwr005.
Baothman, O.A., Zamzami, M.A., Taher, I., Abubaker, J., and Abu-Farha, M. 2016. The role of Gut
Microbiota in the development of obesity and Diabetes. Lipids Health Dis 15: 108. doi:
10.1186/s12944-016-0278-4.
Barshop, B.A. 2012. 216 - Homocystinuria and Hyperhomocysteinemia A2 - Goldman, Lee. In
Goldman's Cecil Medicine (Twenty-Fourth Edition). Edited by A.I. Schafer. W.B. Saunders,
Philadelphia. pp. 1361-1363.
Buchner, D.A., Charrier, A., Srinivasan, E., Wang, L., Paulsen, M.T., Ljungman, M., Bridges, D., and
Saltiel, A.R. 2015. Zinc finger protein 407 (ZFP407) regulates insulin-stimulated glucose uptake and
glucose transporter 4 (Glut4) mRNA. J Biol Chem 290(10): 6376-6386. doi:
10.1074/jbc.M114.623736.
Chawla, A., Schwarz, E.J., Dimaculangan, D.D., and Lazar, M.A. 1994. Peroxisome proliferator-
activated receptor (PPAR) gamma: adipose-predominant expression and induction early in adipocyte
differentiation. Endocrinology 135(2): 798-800. doi: 10.1210/endo.135.2.8033830.
Costa, V., Gallo, M.A., Letizia, F., Aprile, M., Casamassimi, A., and Ciccodicola, A. 2010. PPARG: Gene
Expression Regulation and Next-Generation Sequencing for Unsolved Issues. PPAR Res 2010. doi:
10.1155/2010/409168.
Dong, D., Wang, B., Yin, W., Ding, X., Yu, J., and Kang, Y.J. 2013. Disturbance of copper homeostasis is
a mechanism for homocysteine-induced vascular endothelial cell injury. PLoS One 8(10): e76209.
doi: 10.1371/journal.pone.0076209
PONE-D-13-19955 [pii].
Erridge, C., Attina, T., Spickett, C.M., and Webb, D.J. 2007. A high-fat meal induces low-grade
endotoxemia: evidence of a novel mechanism of postprandial inflammation. Am J Clin Nutr 86(5):
1286-1292. doi: 86/5/1286 [pii].
Fruhbeck, G., Sesma, P., and Burrell, M.A. 2009. PRDM16: the interconvertible adipo-myocyte
switch. Trends Cell Biol 19(4): 141-146. doi: 10.1016/j.tcb.2009.01.007.
Giordano, A., Murano, I., Mondini, E., Perugini, J., Smorlesi, A., Severi, I., Barazzoni, R., Scherer, P.E.,
and Cinti, S. 2013. Obese adipocytes show ultrastructural features of stressed cells and die of
pyroptosis. J Lipid Res 54(9): 2423-2436. doi: 10.1194/jlr.M038638
jlr.M038638 [pii].
Guo, H., Chi, J., Xing, Y., and Wang, P. 2009. Influence of folic acid on plasma homocysteine levels &
arterial endothelial function in patients with unstable angina. Indian J Med Res 129(3): 279-284.
Harmancey, R., and Smih, F. 2007. Response to Comment on: Harmancey et al. (2007)
Adrenomedullin Inhibits Adipogenesis Under Transcriptional Control of Insulin: Diabetes 56:553 563.
Diabetes 56(10): e18. doi: 10.2337/db07-1003.
Page 9 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
10
Hondares, E., Rosell, M., Diaz-Delfin, J., Olmos, Y., Monsalve, M., Iglesias, R., Villarroya, F., and Giralt,
M. 2011. Peroxisome proliferator-activated receptor alpha (PPARalpha) induces PPARgamma
coactivator 1alpha (PGC-1alpha) gene expression and contributes to thermogenic activation of
brown fat: involvement of PRDM16. J Biol Chem 286(50): 43112-43122. doi:
10.1074/jbc.M111.252775.
Hong, K.H., Keen, C.L., Mizuno, Y., Johnston, K.E., and Tamura, T. 2000. Effects of dietary zinc
deficiency on homocysteine and folate metabolism in rats. J Nutr Biochem 11(3): 165-169. doi:
S0955-2863(99)00089-3 [pii].
Huemer, M., Kozich, V., Rinaldo, P., Baumgartner, M.R., Merinero, B., Pasquini, E., Ribes, A., and
Blom, H.J. 2015. Newborn screening for homocystinurias and methylation disorders: systematic
review and proposed guidelines. J Inherit Metab Dis 38(6): 1007-1019. doi: 10.1007/s10545-015-
9830-z.
Iuchi, S. 2001. Three classes of C2H2 zinc finger proteins. Cell Mol Life Sci 58(4): 625-635.
Jing, M., Rech, L., Wu, Y., Goltz, D., Taylor, C.G., and House, J.D. 2015. Effects of zinc deficiency and
zinc supplementation on homocysteine levels and related enzyme expression in rats. J Trace Elem
Med Biol 30: 77-82. doi: 10.1016/j.jtemb.2014.10.013.
Jurowski, K., Szewczyk, B., Nowak, G., and Piekoszewski, W. 2014. Biological consequences of zinc
deficiency in the pathomechanisms of selected diseases. Journal of biological inorganic chemistry :
JBIC : a publication of the Society of Biological Inorganic Chemistry 19(7): 1069-1079. doi:
10.1007/s00775-014-1139-0.
Kajimura, S., Seale, P., Kubota, K., Lunsford, E., Frangioni, J.V., Gygi, S.P., and Spiegelman, B.M. 2009.
Initiation of myoblast to brown fat switch by a PRDM16-C/EBP-beta transcriptional complex. Nature
460(7259): 1154-1158. doi: 10.1038/nature08262.
Kalani, A., Kamat, P.K., Tyagi, S.C., and Tyagi, N. 2013. Synergy of homocysteine, microRNA, and
epigenetics: a novel therapeutic approach for stroke. Mol Neurobiol 48(1): 157-168. doi:
10.1007/s12035-013-8421-y.
Kambouris, M., Maroun, R.C., Ben-Omran, T., Al-Sarraj, Y., Errafii, K., Ali, R., Boulos, H., Curmi, P.A.,
and El-Shanti, H. 2014. Mutations in zinc finger 407 [ZNF407] cause a unique autosomal recessive
cognitive impairment syndrome. Orphanet J Rare Dis 9: 80. doi: 10.1186/1750-1172-9-80.
Kim, H.S., Liang, L., Dean, R.G., Hausman, D.B., Hartzell, D.L., and Baile, C.A. 2001. Inhibition of
preadipocyte differentiation by myostatin treatment in 3T3-L1 cultures. Biochem Biophys Res
Commun 281(4): 902-906. doi: 10.1006/bbrc.2001.4435.
Kim, J.K., Gavrilova, O., Chen, Y., Reitman, M.L., and Shulman, G.I. 2000. Mechanism of insulin
resistance in A-ZIP/F-1 fatless mice. J Biol Chem 275(12): 8456-8460.
Konukoglu, D., Serin, O., Ercan, M., and Turhan, M.S. 2003. Plasma homocysteine levels in obese and
non-obese subjects with or without hypertension; its relationship with oxidative stress and copper.
Clin Biochem 36(5): 405-408.
Laity, J.H., Lee, B.M., and Wright, P.E. 2001. Zinc finger proteins: new insights into structural and
functional diversity. Curr Opin Struct Biol 11(1): 39-46.
Lamkanfi, M. 2011. Emerging inflammasome effector mechanisms. Nat Rev Immunol 11(3): 213-220.
doi: 10.1038/nri2936
nri2936 [pii].
Lane, M.D., Tang, Q.Q., and Jiang, M.S. 1999. Role of the CCAAT enhancer binding proteins (C/EBPs)
in adipocyte differentiation. Biochem Biophys Res Commun 266(3): 677-683. doi:
10.1006/bbrc.1999.1885.
Lee, J.E., and Ge, K. 2014. Transcriptional and epigenetic regulation of PPARgamma expression
during adipogenesis. Cell Biosci 4: 29. doi: 10.1186/2045-3701-4-29.
Li, S., Arning, E., Liu, C., Vitvitsky, V., Hernandez, C., Banerjee, R., Bottiglieri, T., and Lin, J.D. 2009.
Regulation of homocysteine homeostasis through the transcriptional coactivator PGC-1alpha. Am J
Physiol Endocrinol Metab 296(3): E543-548. doi: 10.1152/ajpendo.90719.2008.
Page 10 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
11
Lochmann, T.L., Thomas, R.R., Bennett, J.P., Jr., and Taylor, S.M. 2015. Epigenetic Modifications of
the PGC-1α Promoter during Exercise Induced Expression in Mice. PLOS ONE 10(6): e0129647. doi:
10.1371/journal.pone.0129647.
Majumder, A., Behera, J., Jeremic, N., and Tyagi, S.C. 2017. Hypermethylation: Causes and
Consequences in Skeletal Muscle Myopathy. Journal of cellular biochemistry 118(8): 2108-2117. doi:
10.1002/jcb.25841.
Maron, B.A., and Loscalzo, J. 2009. The treatment of hyperhomocysteinemia. Annual review of
medicine 60: 39-54. doi: 10.1146/annurev.med.60.041807.123308.
Mentese, A., Alver, A., Sumer, A., and Demir, S. 2016. Effects of homocysteine on adipocyte
differentiation and CD36 gene expression in 3T3-L1 adipocytes. J Cell Commun Signal 10(1): 55-60.
doi: 10.1007/s12079-015-0316-4.
Mishra, P.K., Tyagi, N., Sen, U., Joshua, I.G., and Tyagi, S.C. 2010. Synergism in
hyperhomocysteinemia and diabetes: role of PPAR gamma and tempol. Cardiovasc Diabetol 9: 49.
doi: 10.1186/1475-2840-9-49.
Murphy, G.J., and Holder, J.C. 2000. PPAR-gamma agonists: therapeutic role in diabetes,
inflammation and cancer. Trends Pharmacol Sci 21(12): 469-474.
Odegaard, J.I., and Chawla, A. 2013. Pleiotropic actions of insulin resistance and inflammation in
metabolic homeostasis. Science 339(6116): 172-177. doi: 10.1126/science.1230721.
Puigserver, P. 2005. Tissue-specific regulation of metabolic pathways through the transcriptional
coactivator PGC1-alpha. Int J Obes (Lond) 29 Suppl 1: S5-9. doi: 10.1038/sj.ijo.0802905.
Puigserver, P., Wu, Z., Park, C.W., Graves, R., Wright, M., and Spiegelman, B.M. 1998. A cold-
inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92(6): 829-839.
Ren, C.M., Liang, Y., Wei, F., Zhang, Y.N., Zhong, S.Q., Gu, H., Dong, X.S., Huang, Y.Y., Ke, H., Son,
X.M., Tang, D., and Chen, Z. 2013. Balanced translocation t(3;18)(p13;q22.3) and points mutation in
the ZNF407 gene detected in patients with both moderate non-syndromic intellectual disability and
autism. Biochim Biophys Acta 1832(3): 431-438. doi: 10.1016/j.bbadis.2012.11.009.
Rosen, E.D., Hsu, C.H., Wang, X., Sakai, S., Freeman, M.W., Gonzalez, F.J., and Spiegelman, B.M.
2002. C/EBPalpha induces adipogenesis through PPARgamma: a unified pathway. Genes Dev 16(1):
22-26. doi: 10.1101/gad.948702.
Rosen, E.D., Sarraf, P., Troy, A.E., Bradwin, G., Moore, K., Milstone, D.S., Spiegelman, B.M., and
Mortensen, R.M. 1999. PPAR gamma is required for the differentiation of adipose tissue in vivo and
in vitro. Mol Cell 4(4): 611-617.
Ruschke, K., Fishbein, L., Dietrich, A., Kloting, N., Tonjes, A., Oberbach, A., Fasshauer, M., Jenkner, J.,
Schon, M.R., Stumvoll, M., Bluher, M., and Mantzoros, C.S. 2010. Gene expression of PPARgamma
and PGC-1alpha in human omental and subcutaneous adipose tissues is related to insulin resistance
markers and mediates beneficial effects of physical training. European journal of endocrinology
162(3): 515-523. doi: 10.1530/eje-09-0767.
Seale, P., Bjork, B., Yang, W., Kajimura, S., Chin, S., Kuang, S., Scime, A., Devarakonda, S., Conroe,
H.M., Erdjument-Bromage, H., Tempst, P., Rudnicki, M.A., Beier, D.R., and Spiegelman, B.M. 2008.
PRDM16 controls a brown fat/skeletal muscle switch. Nature 454(7207): 961-967. doi:
10.1038/nature07182.
Sen, U., Mishra, P.K., Tyagi, N., and Tyagi, S.C. 2010. Homocysteine to hydrogen sulfide or
hypertension. Cell Biochem Biophys 57(2-3): 49-58. doi: 10.1007/s12013-010-9079-y.
Singh, M., and Tyagi, S.C. 2017. Hyperhomocysteinemia and Age-related Macular Degeneration: Role
of Inflammatory Mediators and Pyroptosis; A Proposal. Med Hypotheses 105: 17-21. doi:
10.1016/j.mehy.2017.06.012.
Tontonoz, P., Hu, E., and Spiegelman, B.M. 1995. Regulation of adipocyte gene expression and
differentiation by peroxisome proliferator activated receptor gamma. Curr Opin Genet Dev 5(5):
571-576.
Page 11 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
12
Troseid, M., Nestvold, T.K., Rudi, K., Thoresen, H., Nielsen, E.W., and Lappegard, K.T. 2013. Plasma
lipopolysaccharide is closely associated with glycemic control and abdominal obesity: evidence from
bariatric surgery. Diabetes Care 36(11): 3627-3632. doi: 10.2337/dc13-0451
dc13-0451 [pii].
Vaya, A., Ejarque, I., Tembl, J., Corella, D., and Laiz, B. 2011. Hyperhomocysteinemia, obesity and
cryptogenic stroke. Clin Hemorheol Microcirc 47(1): 53-58. doi: 10.3233/CH-2010-1365.
Vaya, A., Rivera, L., Hernandez-Mijares, A., de la Fuente, M., Sola, E., Romagnoli, M., Alis, R., and
Laiz, B. 2012. Homocysteine levels in morbidly obese patients: its association with waist
circumference and insulin resistance. Clin Hemorheol Microcirc 52(1): 49-56. doi: 10.3233/CH-2012-
1544.
Veeranki, S., Winchester, L.J., and Tyagi, S.C. 2015. Hyperhomocysteinemia associated skeletal
muscle weakness involves mitochondrial dysfunction and epigenetic modifications. Biochimica et
biophysica acta 1852(5): 732-741. doi: 10.1016/j.bbadis.2015.01.008.
Vega, R.B., Huss, J.M., and Kelly, D.P. 2000. The coactivator PGC-1 cooperates with peroxisome
proliferator-activated receptor alpha in transcriptional control of nuclear genes encoding
mitochondrial fatty acid oxidation enzymes. Mol Cell Biol 20(5): 1868-1876.
Wang, Z., Pini, M., Yao, T., Zhou, Z., Sun, C., Fantuzzi, G., and Song, Z. 2011. Homocysteine
suppresses lipolysis in adipocytes by activating the AMPK pathway. Am J Physiol Endocrinol Metab
301(4): E703-712. doi: 10.1152/ajpendo.00050.2011.
Wei, S., Zhang, L., Zhou, X., Du, M., Jiang, Z., Hausman, G.J., Bergen, W.G., Zan, L., and Dodson, M.V.
2013. Emerging roles of zinc finger proteins in regulating adipogenesis. Cellular and molecular life
sciences : CMLS 70(23): 4569-4584. doi: 10.1007/s00018-013-1395-0.
Winchester, L., Veeranki, S., Givvimani, S., and Tyagi, S.C. 2014. Exercise mitigates the adverse
effects of hyperhomocysteinemia on macrophages, MMP-9, skeletal muscle, and white adipocytes.
Can J Physiol Pharmacol 92(7): 575-582. doi: 10.1139/cjpp-2014-0059.
Wolf, K.J., and Lorenz, R.G. 2012. Gut Microbiota and Obesity. Curr Obes Rep 1(1): 1-8. doi:
10.1007/s13679-011-0001-8.
Xi, H., Zhang, Y., Xu, Y., Yang, W.Y., Jiang, X., Sha, X., Cheng, X., Wang, J., Qin, X., Yu, J., Ji, Y., Yang, X.,
and Wang, H. 2016. Caspase-1 Inflammasome Activation Mediates Homocysteine-Induced Pyrop-
Apoptosis in Endothelial Cells. Circ Res 118(10): 1525-1539. doi: 10.1161/CIRCRESAHA.116.308501
CIRCRESAHA.116.308501 [pii].
Yideng, J., Zhihong, L., Jiantuan, X., Jun, C., Guizhong, L., and Shuren, W. 2008. Homocysteine-
mediated PPARalpha,gamma DNA methylation and its potential pathogenic mechanism in
monocytes. DNA Cell Biol 27(3): 143-150. doi: 10.1089/dna.2007.0658.
Zuo, Y., Qiang, L., and Farmer, S.R. 2006. Activation of CCAAT/enhancer-binding protein (C/EBP)
alpha expression by C/EBP beta during adipogenesis requires a peroxisome proliferator-activated
receptor-gamma-associated repression of HDAC1 at the C/ebp alpha gene promoter. J Biol Chem
281(12): 7960-7967. doi: 10.1074/jbc.M510682200.
List of abbreviations
AMD: age-related macular degeneration
Cu+2: copper
C/EBPs: CCAAT/enhancer binding proteins
CBS: cystathionine β-synthase
CSE: cystathionine γ-lyase
Page 12 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
13
DNMT: DNA methyl transferase
FFA: free fatty acid
GMT: gut Microbiota transplant
HATs: histone acetyltransferases
Hcy: homocysteine
HHcy: hyperhomocysteinemia
LPS: lipopolysaccharide
NLRP3: nucleotide-binding domain, leucine-rich-containing family, pyrin domain-
containing-3
PPAR: peroxisome proliferator activator receptor:
RXR: retinoic X receptor
T2D: type 2 diabetes
Zn+2’: Zinc
Zfp: zinc finger protein
Page 13 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
14
Figure legends
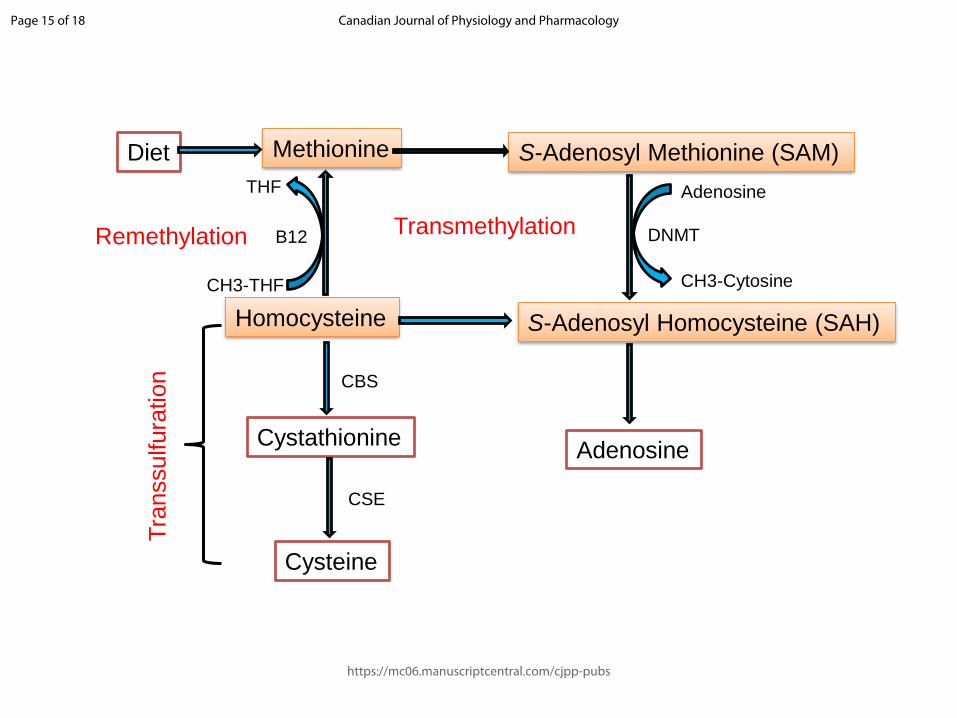
Fig. 1. A schematic representation of methionine metabolism depicting the transmethylation,
transsulfuration and remethylation pathways. Dietary methionine is converted to s-adenosyl
methionine (SAM) which is further converted to s-adenosyl homocysteine (SAH) in the
presence of DNMT. SAH is converted to homocysteine (Hcy) which is further remethylated
to methionine in the presence of folic acid (Vitamin B12) and the cycle continues. Under low
concentrations of cysteine, Hcy is transsulfurated to cysteine where CBS and CSE act as
essential catalysts.
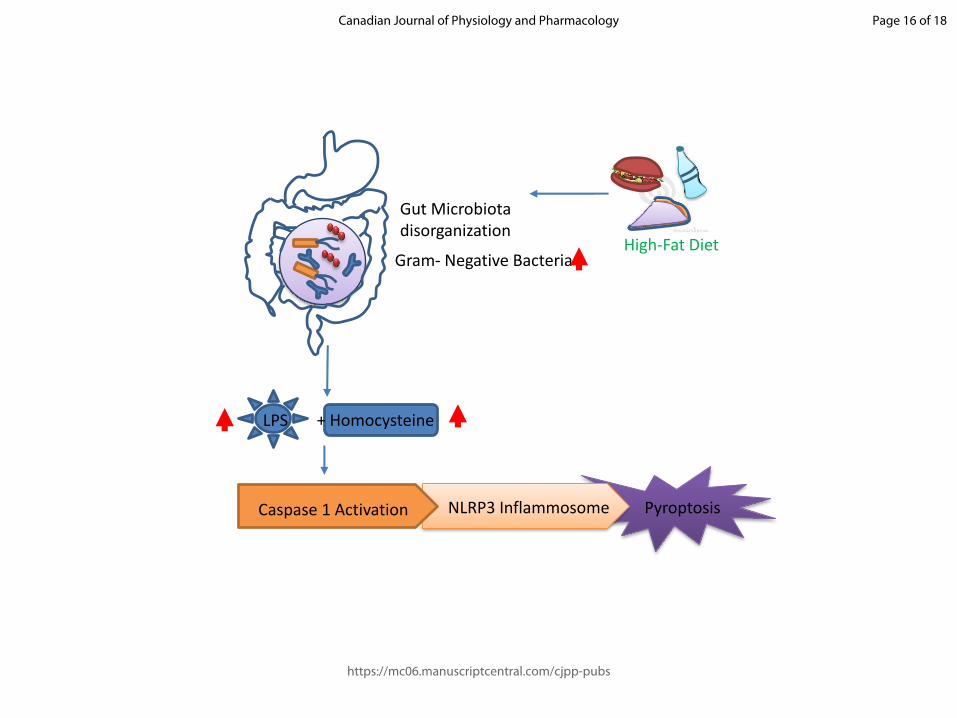
Fig. 2. Proposed pathway representing elevated homocysteine levels contributing to the
obesity-related dysbiosis and pyroptosis. Perpetual consumption of high fat diet leads to
dysbiosys of the gut microbiota. This increases the number of Gram-negative bacteria,
expressing LPS on their membrane, in the gut. The high LPS concentration together with
elevated Hcy levels elicit an immune response causing activation of Caspase 1 and
consequently NLRP3 inflammasome assembly. This induces cellular death by pyroptosis.
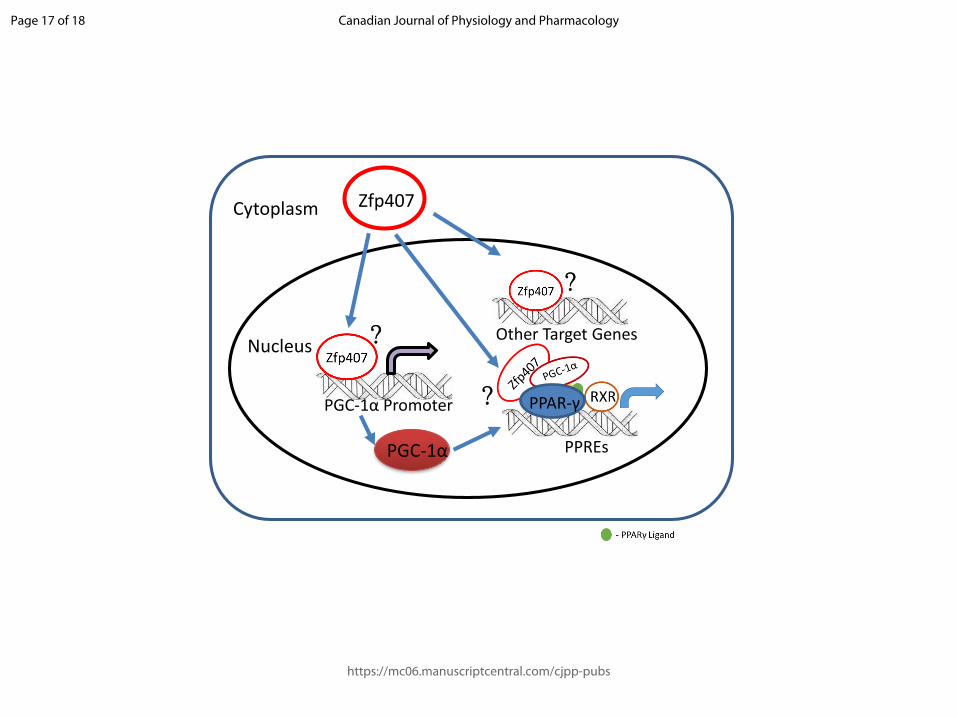
Fig. 3. A model of zinc finger protein 407 (Zpf407) regulating PPAR-γ target gene expression
under normal conditions. Zfp407, upon maturation and entry into the nucleus, may interact
with either the PPAR-γ coactivator-1α (PGC-1α) promoter to increase PGC-1α transcription
which can regulate PPAR-γ target gene expression. Zfp407 can also interact with PPAR-γ
directly or by scaffolding proteins to form a PPAR-γ-PGC-1α complex on PPAR-γ responsive
elements (PPREs) and regulate PPAR-γ target gene expression. In a third proposed
mechanism, Zfp407 itself can work as a transcription factor and interact with its own target
genes which are yet to be studied.
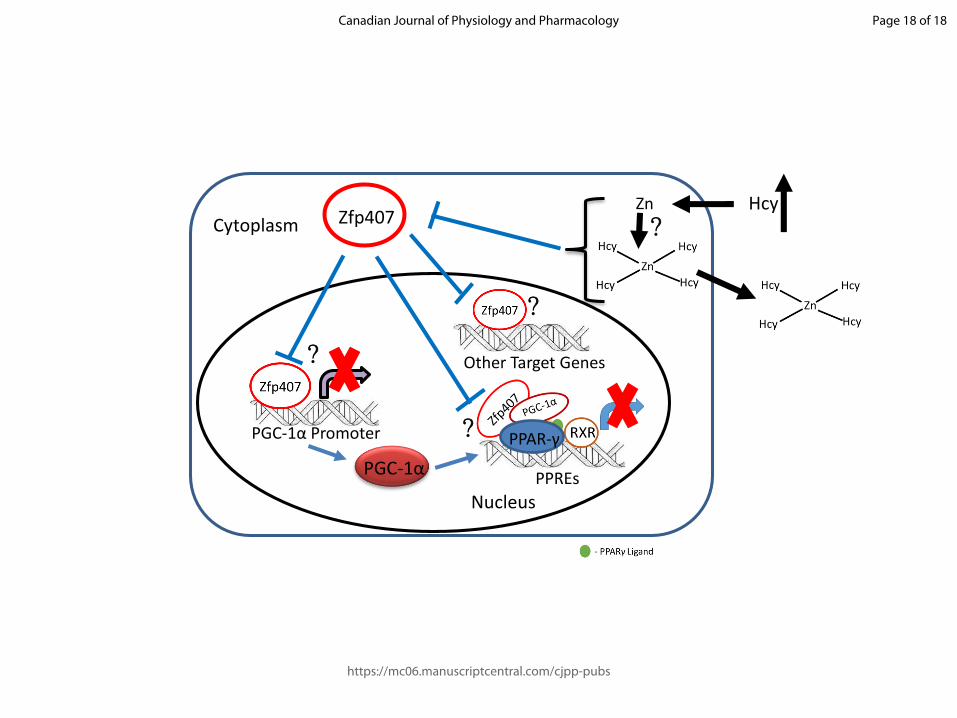
Fig. 4. Model representing the mechanistic aspects of Zfp407 regulation of PPARγ target genes under high serum homocysteine concentrations via Zn+2 sequestration. High Hcy concentration in the serum can sequester free Zn+2 ions forming a Zn-Hcy complex which can be transported to the extracellular milieu. Thus, the Zn+2 ions necessary to be incorporated into the Zfp407 is unavailable and the synthesis of Zfp407 is inhibited through its pre-mature degradation. This could potentially reduce the proposed gene regulatory mechanisms of Zfp407 either by reducing PGC-1α synthesis or weakening the interaction between PPAR-γ-PGC-1α complex.
Page 14 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
Methionine
Homocysteine
S-Adenosyl Methionine (SAM)
S-Adenosyl Homocysteine (SAH)
Diet
THF
B12
CH3-THF
Cystathionine
Cysteine
CSE
DNMT
CH3-Cytosine
CBS
Adenosine
Adenosine
Remethylation
Tra
nssulfura
tion
Transmethylation
Page 15 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
Gut Microbiota disorganization
LPS
Gram- Negative Bacteria
+ Homocysteine
Caspase 1 Activation NLRP3 Inflammosome Pyroptosis
High-Fat Diet
Page 16 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
Zfp407
PPREs
Other Target Genes
PGC-1α
PGC-1α Promoter
Cytoplasm
Nucleus
PPAR-γ?
?
?
Page 17 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
Zfp407
PPREs
Other Target Genes
PGC-1α
PGC-1α Promoter
Cytoplasm
Nucleus
HcyZn
PPAR-γ?
?
?
?
Page 18 of 18
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology


















