
ωρίμανση του M rna
24
ΕΡΓΑΣΙΑ 6 η Θέμα 1 Να περιγραφούν τα κυριότερα στάδια της ωρίμανσης του mRNA των ευκαρυωτικών οργανισμών. Μικρό πυρηνικό RNA, snRNA Τα μικρά πυρηνικά RNAs συναντώνται στον πυρήνα των ευκαρυωτικών κυττάρων και αποτελούνται περίπου από 107- 210 νουκλεοτίδια. Συνθέτονται από την RNA πολυμεράση II ή III. Συμμετέχουν στην ωρίμανση του πρόδρομου m RNA, όπου αφαιρούν τα ιντρόνια, και στη διατήρηση των τελομερών. Ανακαλύφθηκαν αρχικά το 1966 και το 1982 προσδιορίστηκε η δομή τους. Ο Phillip Sharp και ο Richard J. Roberts πήραν το βραβείο Nobel το 1993 για την ανακάλυψη των ιντρονίων και της διαδικασίας ωρίμανσης. Εικόνες 1,2,3 5 snRNAs U1, U2, U4, U5, και U6 σχηματίζουν το σωμάτιο συναρμογής (spliceosomes), υπεύθυνο για την αφαίρεση των ιντρονίων. Τα snRNAs συνδέονται με 6-10 πρωτείνες και σχηματίζουν τα ριβονουκλεοπρωτεινικά σύμπλοκα snRNP. Η δευτεροταγής δομή τους είναι υψηλά συντηρητική σε όλους τους οργανισμούς. Ο Thomas Cech και Sydney Altman ανακάλυψαν ότι τα μόρια RNA μπορούν να έχουν καταλύτική δράση, γεγονός που άλλαξε τον προσανατολισμό στη μοριακή εξέλιξη. Εικόνες 4,5,6 Λυρατζόπουλος Εμμανουήλ Βιολόγος 1
Transcript of ωρίμανση του M rna

ΕΡΓΑΣΙΑ 6 η
Θέμα 1
Να περιγραφούν τα κυριότερα στάδια της ωρίμανσης του mRNA των
ευκαρυωτικών οργανισμών.
Μικρό πυρηνικό RNA, snRNA
Τα μικρά πυρηνικά RNAs συναντώνται στον πυρήνα των ευκαρυωτικών κυττάρων
και αποτελούνται περίπου από 107-210 νουκλεοτίδια. Συνθέτονται από την RNA
πολυμεράση II ή III. Συμμετέχουν στην ωρίμανση του πρόδρομου m RNA, όπου
αφαιρούν τα ιντρόνια, και στη διατήρηση των τελομερών. Ανακαλύφθηκαν αρχικά
το 1966 και το 1982 προσδιορίστηκε η δομή τους.
Ο Phillip Sharp και ο Richard J. Roberts πήραν το βραβείο Nobel το 1993 για την
ανακάλυψη των ιντρονίων και της διαδικασίας ωρίμανσης. Εικόνες 1,2,3
5 snRNAs U1, U2, U4, U5, και U6 σχηματίζουν το σωμάτιο συναρμογής
(spliceosomes), υπεύθυνο για την αφαίρεση των ιντρονίων. Τα snRNAs συνδέονται
με 6-10 πρωτείνες και σχηματίζουν τα ριβονουκλεοπρωτεινικά σύμπλοκα snRNP.
Η δευτεροταγής δομή τους είναι υψηλά συντηρητική σε όλους τους οργανισμούς.
Ο Thomas Cech και Sydney Altman ανακάλυψαν ότι τα μόρια RNA μπορούν να έχουν
καταλύτική δράση, γεγονός που άλλαξε τον προσανατολισμό στη μοριακή εξέλιξη.
Εικόνες 4,5,6
Στα ιντρόνια το 5΄άκρο έχει συνήθως τις βάσεις GU και στο 3΄άκρο AG.
Εικόνες 7,8
Ο μηχανισμός έχει τα χαρακτηριστικά ριβόζυμου.
Το εναλλακτικό μάτισμα (Alternative splicing) είναι ένας βασικός μηχανισμός
γενετικής ποικιλότητας στους ευκαρυωτικούς και αιτιολογεί και το μικρό αριθμό
γονιδίων στο ανθρώπινο γονιδίωμα.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 1

Τα περισσότερα γονίδια στα ανώτερα ευκαρυωτικά αποτελούνται από εξώνια και
ιντρόνια. Τα ιντρόνια πρέπει να εξαχθούν και τα εξώνια να συνδεθούν για να
σχηματιστεί το τελικό mRNA με μια διεργασία που ονομάζεται μάτισμα (splicing).
Το μάτισμα πρέπει να είναι εξαιρετικά ευαίσθητο: μια ολίσθηση έστω και ενός
νουκλεοτιδίου στο σημείο ματίσματος θα μετατόπιζε το πλαίσιο ανάγνωσης στην
3΄-πλευρά του ματίσματος, δίνοντας μια τελείως διαφορετική αλληλουχία
αμινοξέων. Επομένως, η σωστή θέση ματίσματος πρέπει να είναι καθαρά
σημειωμένη.
Στα ευκαρυωτικά, από τους ζυμομύκητες έως τα θηλαστικά, οι αλληλουχίες αυτές
έχουν ένα κοινό δομικό πρότυπο:
η αλληλουχία βάσεων ενός ιντρονίου αρχίζει με GU και τελειώνει με AG.
Η ομόφωνη αλληλουχία στο 5΄-άκρο του ιντρονίου σε σπονδυλωτά είναι
AGGUAAGU.
Στο 3΄-άκρο ενός ιντρονίου η ομόφωνη αλληλουχία είναι ένα τμήμα 10 πυριμιδινών
(U ή C), ακολουθούμενο από οποιαδήποτε βάση και μετά από μια C, η οποία
τελειώνει με την αμετάβλητη αλληλουχία AG. Τα ιντρόνια έχουν επίσης μια
σημαντική εσωτερική θέση που τοποθετείται μεταξύ 20 και 50 νουκλεοτιδίων
ανοδικά της θέσης ματίσματος 3΄. Το σημείο αυτό ονομάζεται θέση διακλάδωσης.
Στους ζυμομύκητες, η θέση διακλάδωσης σχεδόν πάντοτε είναι UACUAAC, ενώ στα
θηλαστικά απαντά μια ποικιλία αλληλουχιών.
Τμήματα των ιντρονίων, διαφορετικά από τις θέσεις ματίσματος 5΄ και 3΄
καθώς και τη θέση διακλάδωσης, είναι λιγότερο σημαντικά για τον καθορισμό του
σημείου όπου θα γίνει το μάτισμα. Το μήκος των ιντρονίων ποικίλλει από 50 μέχρι
10.000 νουκλεοτίδια. Το μεγαλύτερο μέρος ενός ιντρονίου μπορεί να απαλειφθεί
χωρίς να αλλάζει η θέση ή η αποτελεσματικότητα του ματίσματος.
Παρομοίως, το μάτισμα δεν επηρεάζεται από την εισαγωγή μεγάλων τμημάτων
DNA στα ιντρόνια γονιδίων. Επιπλέον, χιμαιρικά ιντρόνια που κατασκευάστηκαν με
μεθόδους ανασυνδυασμένου DNA από το 5΄-άκρο ενός ιντρονίου και το 3΄-άκρο
ενός τελείως διαφορετικού ιντρονίου ματίζονται κανονικά, εφόσον δεν έχουν
αλλαχθεί οι θέσεις ματίσματος και διακλάδωσης. Αντίθετα, μεταλλάξεις σε
οποιαδήποτε από αυτές τις τρεις κρίσιμες περιοχές οδηγούν σε μη φυσιολογικό
μάτισμα.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 2

Το μη φυσιολογικό μάτισμα προκαλεί μερικές μορφές θαλασσαιμίας, μιας
ομάδας κληρονομικών αναιμιών που χαρακτηρίζονται από την ελαττωματική
σύνθεση της αιμοσφαιρίνης. Σε έναν ασθενή, μια μετάλλαξη G σε A 19 νουκλεοτίδια
μακριά από την κανονική θέση ματίσματος 3΄ του πρώτου ιντρονίου δημιούργησε
μια νέα θέση ματίσματος. Εικόνα 9
Το mRNA που προέκυψε περιείχε μια σειρά κωδικονίων που δεν υπάρχουν στα
φυσιολογικά άτομα. Το έκτο κωδικόνιο μετά το μάτισμα είναι ένα σήμα
τερματισμού της σύνθεσης πρωτεϊνών, και έτσι η έκτροπη πρωτεΐνη τελειώνει
πρώιμα. Μεταλλάξεις που επηρεάζουν τις θέσεις ματίσματος έχει υπολογιστεί ότι
προκαλούν το 15% των γενετικών νόσων.
Tο μάτισμα αποτελείται από δύο αντιδράσεις τρανσεστεροποίησης.
Το μάτισμα αρχίζει με τη διάσπαση του φωσφοδιεστερικού δεσμού μεταξύ του
ανοδικού εξωνίου (εξώνιο 1) και του 5΄-άκρου του ιντρονίου.
Η προσβάλλουσα ομάδα στην αντίδραση αυτή είναι η 2΄-υδροξυλική ομάδα ενός
καταλοίπου αδενυλικού στη θέση διακλάδωσης.
Ένας 2΄,5΄-φωσφοδιεστερικός δεσμός σχηματίζεται μεταξύ αυτού του
καταλοίπου Α και της φωσφορικής ομάδας του 5΄-άκρου του ιντρονίου. Αυτή η
αντίδραση είναι μια τρανσεστεροποίηση.
Επισημαίνεται ότι το συγκεκριμένο κατάλοιπο αδενυλικού συνδέεται επίσης με
άλλα δύο νουκλεοτίδια διά μέσου κανονικών 3΄,5΄-φωσφοδιεστερικών δεσμών.
Επομένως, μια διακλάδωση δημιουργείται στη θέση αυτή και σχηματίζεται ένα
ενδιάμεσο δομής βρόχου.
Στη συνέχεια, το 3΄-ΟΗ άκρο του εξωνίου 1 προσβάλλει τον φωσφοδιεστερικό
δεσμό μεταξύ του ιντρονίου και του εξωνίου 2.
Τα εξώνια 1 και 2 συνδέονται και το ιντρόνιο ελευθερώνεται με τη μορφή βρόχου.
Και στην περίπτωση αυτή η αντίδραση είναι μια τρανσεστεροποίηση.
Επομένως, το μάτισμα επιτυγχάνεται με δύο αντιδράσεις τρανσεστεροποίησης και
όχι με υδρόλυση ακολουθούμενη από ανασύνδεση.
Η πρώτη αντίδραση δημιουργεί μια ελεύθερη 3΄-υδροξυλική ομάδα στο 3΄-άκρο του
εξωνίου 1, και η δεύτερη αντίδραση συνδέει την ομάδα αυτή με την 5΄-φωσφορική
ομάδα του εξωνίου 2.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 3

Ο αριθμός των φωσφοδιεστερικών δεσμών παραμένει ο ίδιος κατά τη διάρκεια
αυτών των βημάτων, γεγονός εξαιρετικά κρίσιμο διότι επιτρέπει στην αντίδραση
ματίσματος να προχωρά χωρίς πηγή ενέργειας όπως η ΑΤΡ και η GTP. Mικρά
πυρηνικά μόρια RNA στα σωμάτια ματίσματος καταλύουν το μάτισμα πρόδρομων
μορίων mRNA.
Ο πυρήνας περιέχει πολλά είδη μικρών μορίων RNA με λιγότερα από 300
νουκλεοτίδια, τα οποία αναφέρονται ως μικρά πυρηνικά μόρια RNA (small nuclear
RNA, snRNA). Μερικά από αυτά —τα οποία συμβολίζονται ως U1, U2, U4, U5 και U6
— είναι ουσιώδη για το μάτισμα πρόδρομων mRNA. Οι δευτεροταγείς δομές αυτών
των μορίων RNA είναι εξαιρετικά συντηρημένες σε οργανισμούς που εκτείνονται
από τους ζυμομύκητες μέχρι τον άνθρωπο. Αυτά τα μόρια RNA συνδέονται με
ειδικές πρωτεΐνες σχηματίζοντας σύμπλοκα που ονομάζονται μικρά πυρηνικά
ριβονουκλεοπρωτεϊνικά σωμάτια (small nuclear ribonucleoprotein particles, snRNP).
Οι ερευνητές συχνά τα αναφέρουν ως «σναρπς» (snurps). Τα σωμάτια ματίσματος
είναι μεγάλα (60 S) δυναμικά συγκροτήματα αποτελούμενα από μόρια snRNP,
άλλες πρωτεΐνες που ονομάζονται παράγοντες ματίσματος και το πρόδρομο μόριο
mRNA που υφίσταται την επεξεργασία.
Στα κύτταρα των θηλαστικών, το μάτισμα αρχίζει με την αναγνώριση της θέσης
ματίσματος 5΄ από το snRNP U1.
Πράγματι, το snRNP U1 περιέχει μια εξαιρετικά συντηρημένη αλληλουχία έξι
νουκλεοτιδίων που σχηματίζουν ζεύγη βάσεων με τη θέση ματίσματος 5΄ του προ-
mRNA. Αυτή η δέσμευση αρχίζει τη συναρμολόγηση στο μόριο του προ-mRNA.
Στη συνέχεια, το snRNP U2 δεσμεύεται στη θέση διακλάδωσης στο ιντρόνιο διά
μέσου ζευγών βάσεων μεταξύ της εξαιρετικά συντηρημένης αλληλουχίας του
snRNP U2 και του προ-mRNA. Η δέσμευση του snRNP U2 χρειάζεται υδρόλυση ΑΤΡ.
Ένα προσυναρμολογημένο σύμπλοκο U4-U5-U6 συνδέεται με αυτό το σύμπλοκο
U1, U2 και του πρόδρομου μορίου mRNA για να σχηματιστεί το πλήρες σωμάτιο
ματίσματος. Αυτή η σύνδεση επίσης χρειάζεται υδρόλυση ΑΤΡ.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 4

Πρώτα, το U5 αλληλεπιδρά με αλληλουχίες του εξωνίου στη θέση ματίσματος 5΄ και
μετά με το εξώνιο 3΄. Στη συνέχεια, το U6 αποσυνδέεται από το U4 και υφίσταται
μια ενδομοριακή αναδιάταξη η οποία επιτρέπει το ζευγάρωμα των βάσεων με το U2
και εκτοπίζει το U1 από το σωμάτιο ματίσματος αλληλεπιδρώντας με το 5΄-άκρο του
ιντρονίου. Η έλικα U2•U6 είναι απολύτως αναγκαία για το μάτισμα, γεγονός που
υποδηλώνει ότι τα μόρια snRNA U2 και U6 πιθανώς σχηματίζουν το ενεργό κέντρο
του σωματίου ματίσματος. Το U4 χρησιμεύει ως αναστολέας ο οποίος επισκιάζει το
U6 μέχρις ότου οι ειδικές θέσεις ματίσματος ευθυγραμμιστούν.
Αυτές οι ανακατατάξεις έχουν ως αποτέλεσμα την πρώτη αντίδραση
τρανσεστεροποίησης, δημιουργώντας το ενδιάμεσο δομής βρόχου και το
διαχωρισμένο εξώνιο 5΄.
Περαιτέρω ανακατατάξεις του RNA στο σωμάτιο ματίσματος διευκολύνουν
τη δεύτερη τρανσεστεροποίηση. Οι ανακατατάξεις αυτές ευθυγραμμίζουν το
ελεύθερο εξώνιο 5΄ με το εξώνιο 3΄ ούτως ώστε η 3΄-υδροξυλική ομάδα του εξωνίου
5΄ τοποθετείται κατάλληλα για να μπορεί να προσβάλλει πυρηνοφιλικά τη θέση
ματίσματος 3΄ και να δημιουργήσει το ματισμένο προϊόν. Τα U2, U5 και U6 που
είναι δεσμευμένα στο εκτεμνόμενο ιντρόνιο δομής βρόχου ελευθερώνονται
συμπληρώνοντας την αντίδραση ματίσματος.
Πολλά από τα βήματα στη διεργασία του ματίσματος χρειάζονται υδρόλυση ΑΤΡ.
Πώς η ελεύθερη ενέργεια που προέρχεται από την υδρόλυση της ΑΤΡ
χρησιμοποιείται για να ωθήσει την υδρόλυση; Για να επιτευχθούν οι
καλοδιαταγμένες ανακατατάξεις που είναι αναγκαίες για το μάτισμα, RNA-ελικάσες
που ωθούνται από την υδρόλυση ΑΤΡ πρέπει να ξετυλίξουν τις έλικες
του RNA και να επιτρέψουν τον σχηματισμό εναλλακτικών ζευγών βάσεων.
Επομένως, αξίζει να σημειωθούν δύο χαρακτηριστικά της διεργασίας ματίσματος.
Πρώτον, μόρια RNA παίζουν καθοριστικό ρόλο στην καθοδήγηση της
ευθυγράμμισης των θέσεων ματίσματος και στη διεξαγωγή της κατάλυσης.
Δεύτερον, ελικάσες ωθούμενες από την υδρόλυση της ΑΤΡ ξετυλίγουν δίκλωνα
ενδιάμεσα μόρια RNA, γεγονός που διευκολύνει την κατάλυση και επάγει την
ελευθέρωση των μορίων snRNP από το mRNA. Εικόνες 10-20
Λυρατζόπουλος Εμμανουήλ Βιολόγος 5

Mερικά μόρια προ-mRNA μπορούν να ματίζονται με εναλλακτικούς τρόπους
δίνοντας διαφορετικά mRNA.
Το εναλλακτικό μάτισμα είναι ένας αρκετά διαδεδομένος μηχανισμός για τη
δημιουργία πρωτεϊνικής ποικιλομορφίας. Συνιστά τη διαφορική συμμετοχή
εξωνίων σε ένα ώριμο RNA. Το εναλλακτικό μάτισμα μπορεί να ρυθμίζεται για να
παράγει διαφορετικές μορφές μιας πρωτεΐνης για ειδικούς ιστούς ή στάδια της
ανάπτυξης. Πρόσφατοι υπολογισμοί δείχνουν ότι τα προϊόντα RNA του 30% των
ανθρώπινων γονιδίων ματίζονται εναλλακτικά. Το εναλλακτικό μάτισμα αποτελεί
έναν ισχυρό μηχανισμό που επεκτείνει την ποικιλομορφία των γονιδιωματικών
αλληλουχιών. Εικόνα 21
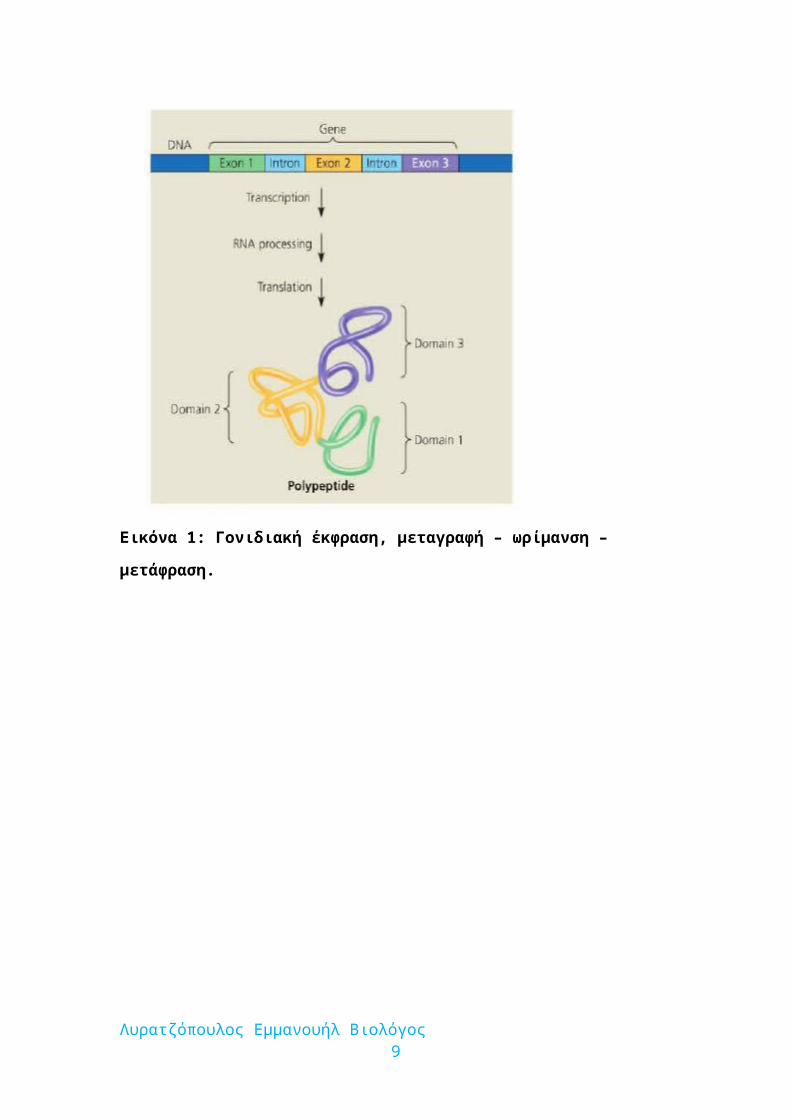
Εικόνα 1: Γονιδιακή έκφραση, μεταγραφή – ωρίμανση – μετάφραση.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 6

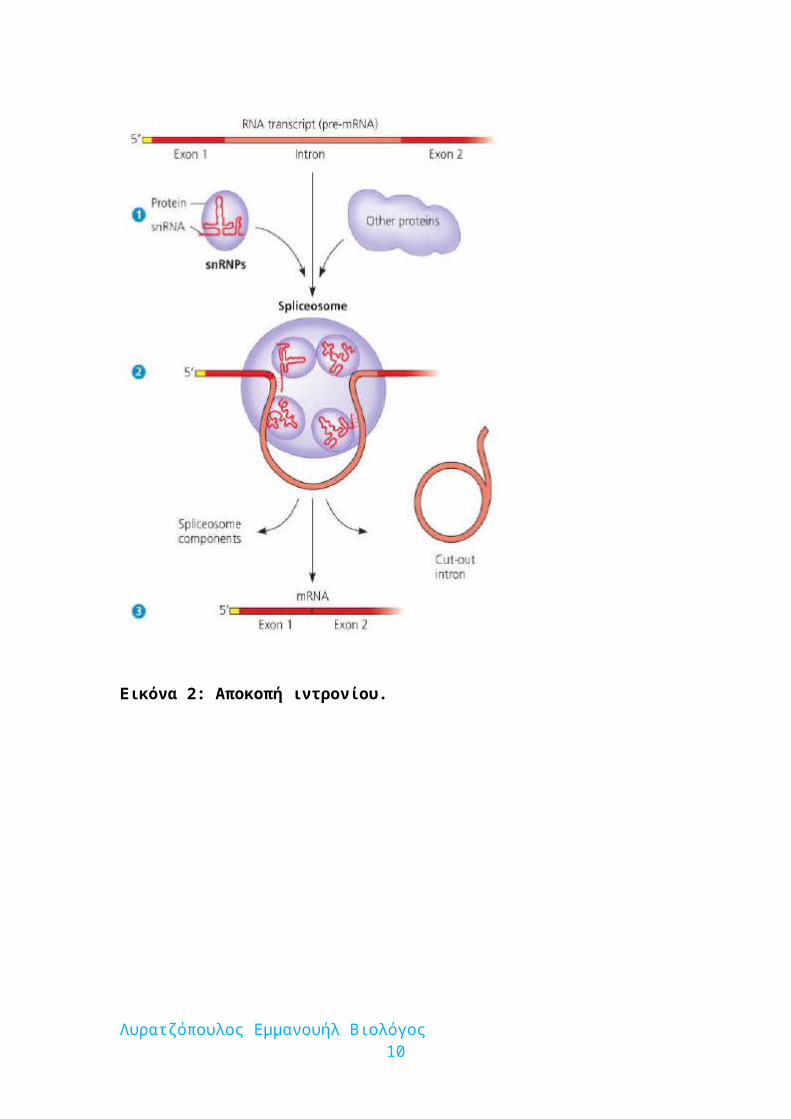
Εικόνα 2: Αποκοπή ιντρονίου.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 7

Εικόνα 3: Το μάτισμα είναι μια διαδικασία ωρίμανσης του pre-mRNA κατά την
οποία τα ιντρόνια (introns) του pre-RNA αποκόπτονται και το υπόλοιπο μόριο
(εξώνια) επανασυνδέεται ώστε να σχηματιστεί ένα ενιαίο ώριμο μετάγραφο
μόριο. Το μάτισμα γίνεται ταυτόχρονα με τη μεταγραφή.
Εικόνα 4: Μόρια snRNPs.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 8
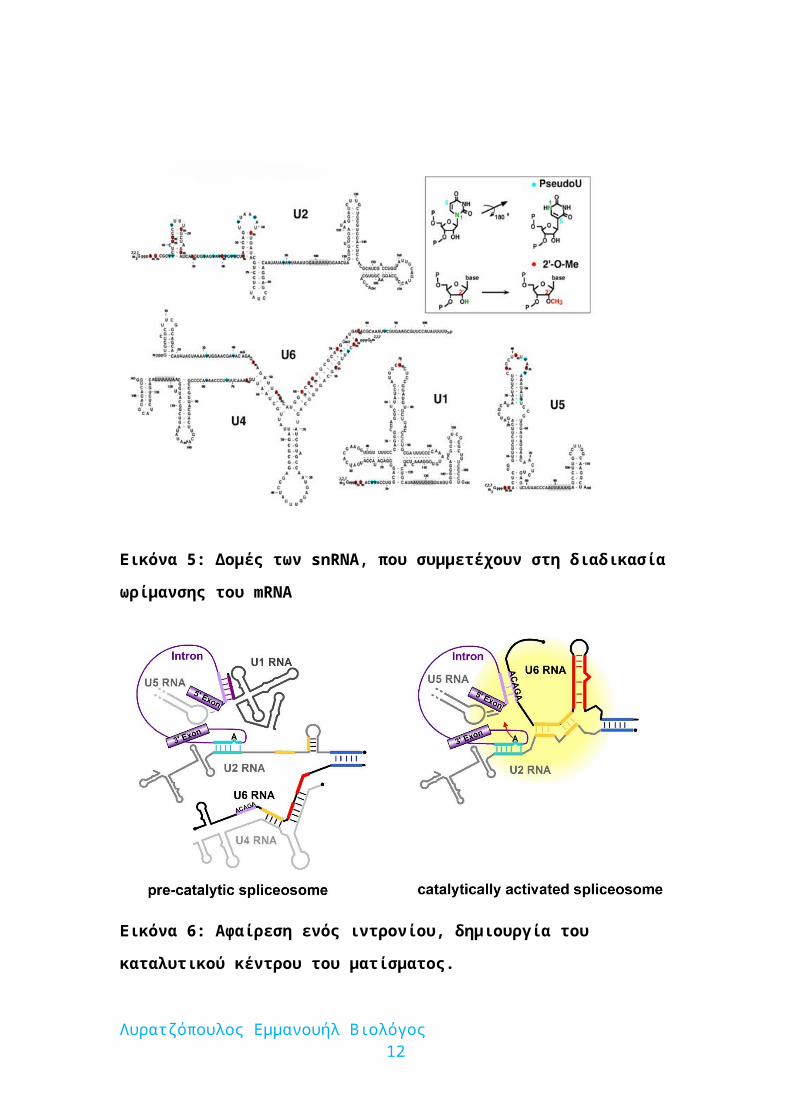
Εικόνα 5: Δομές των snRNA, που συμμετέχουν στη διαδικασία ωρίμανσης του
mRNA
Εικόνα 6: Αφαίρεση ενός ιντρονίου, δημιουργία του καταλυτικού κέντρου του
ματίσματος.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 9
Εικόνα 7: Θέσεις splicing.
Εικόνα 8: Θέσεις splicing.
Εικόνα 9: Μετάλλαξη σε μια βάση σε ένα ιντρόνιο του γονιδίου της β-σφαιρίνης
οδηγεί σε β θαλασσαιμία. Η μετάλλαξη δημιουργεί μια νέα θέση ματίσματος.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 10

Εικόνα 10: Μηχανισμός ματίσματος. Η 2-ΟΗ της Α διακλάδωσης εστεροποιείται με
τη 5΄θέση ματίσματος, δημιουργώντας έναν νέο 3΄-5΄φωσφοδιεστερικό δεσμό και
ένα νέο ελεύθερο 3΄ΟΗ στο άκρο του ανοδικού εξωνίου. Το άκρο αυτό
εστεροποιείται με τη Ρ της 3΄θέσης ματίσματος δημιουργώντας έναν νέο 3΄-
5΄φωσφοδιεστερικό δεσμό μεταξύ των δύο εξωνίων και ένα ελεύθερο βρόγχο
ιντρονίου.
Εικόνα 11: Μηχανισμός τρανσεστεροποίησης.
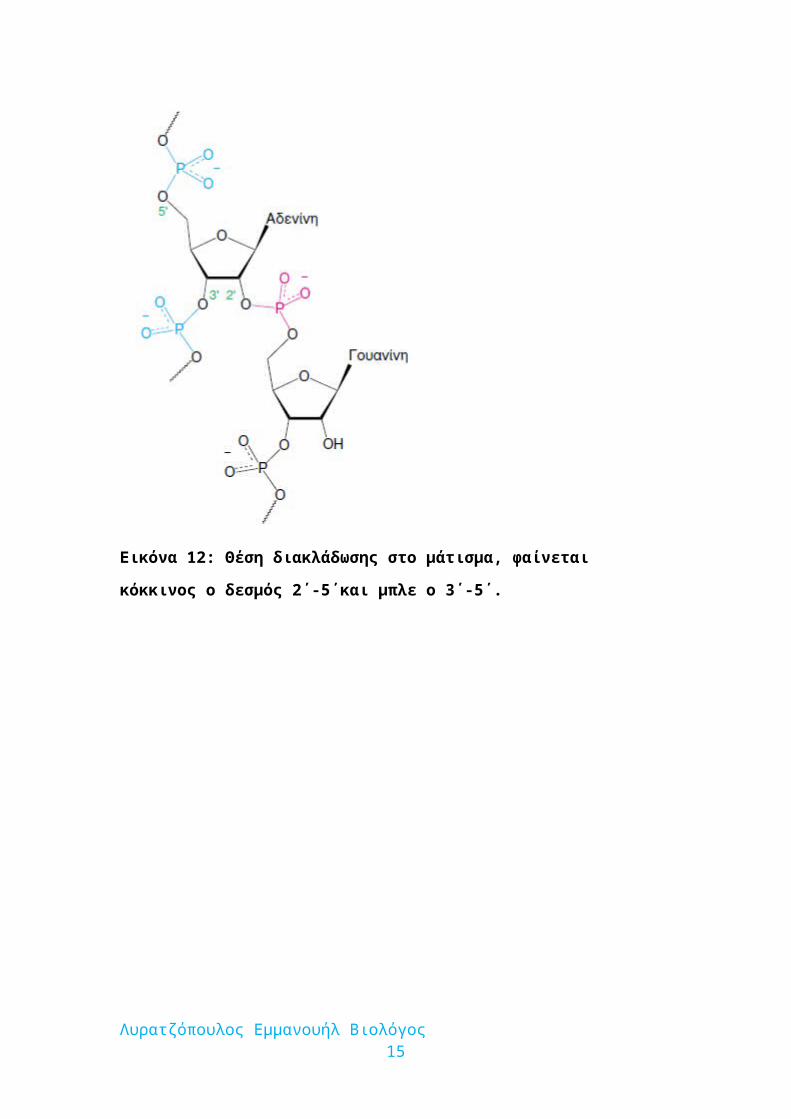
Εικόνα 12: Θέση διακλάδωσης στο μάτισμα, φαίνεται κόκκινος ο δεσμός 2΄-5΄και
μπλε ο 3΄-5΄.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 11
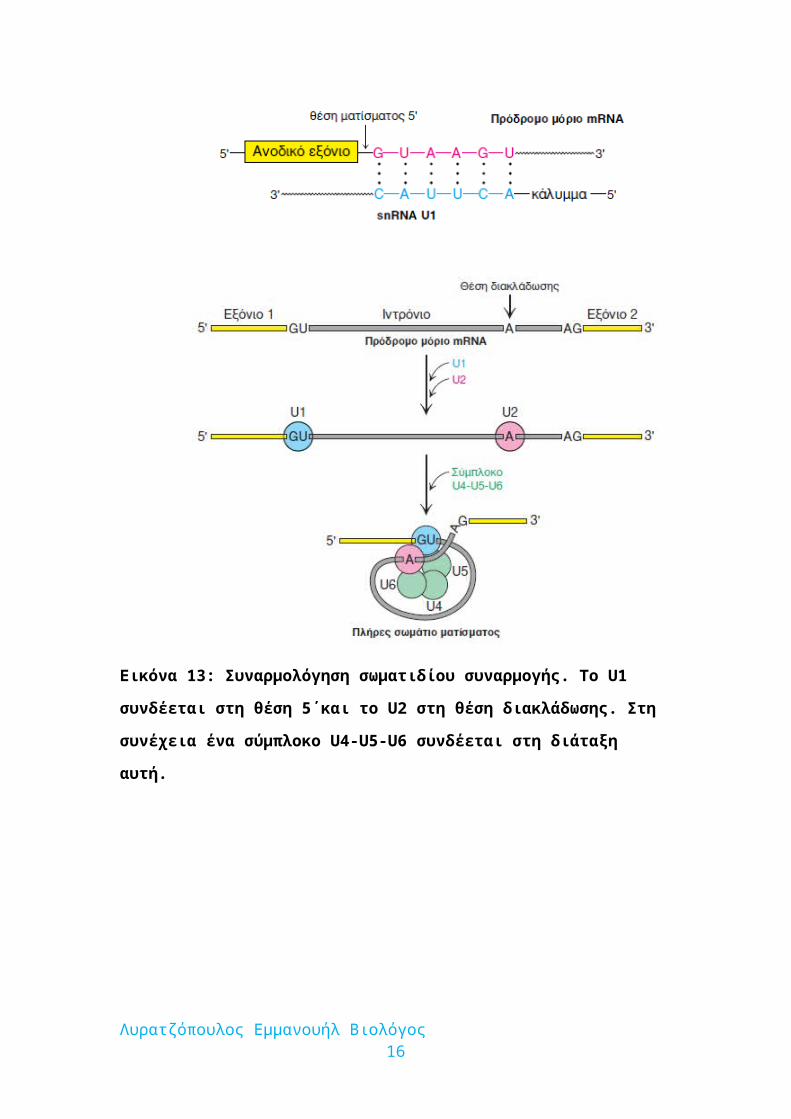
Εικόνα 13: Συναρμολόγηση σωματιδίου συναρμογής. Το U1 συνδέεται στη θέση
5΄και το U2 στη θέση διακλάδωσης. Στη συνέχεια ένα σύμπλοκο U4-U5-U6
συνδέεται στη διάταξη αυτή.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 12

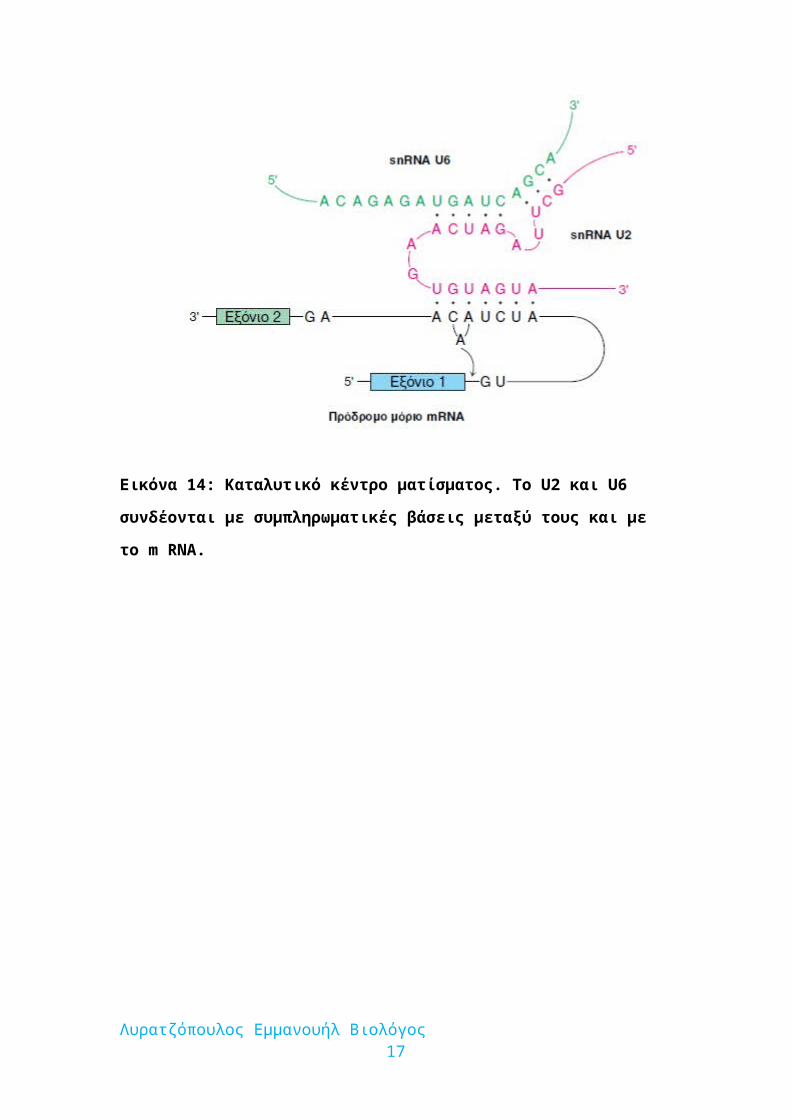
Εικόνα 14: Καταλυτικό κέντρο ματίσματος. Το U2 και U6 συνδέονται με
συμπληρωματικές βάσεις μεταξύ τους και με το m RNA.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 13

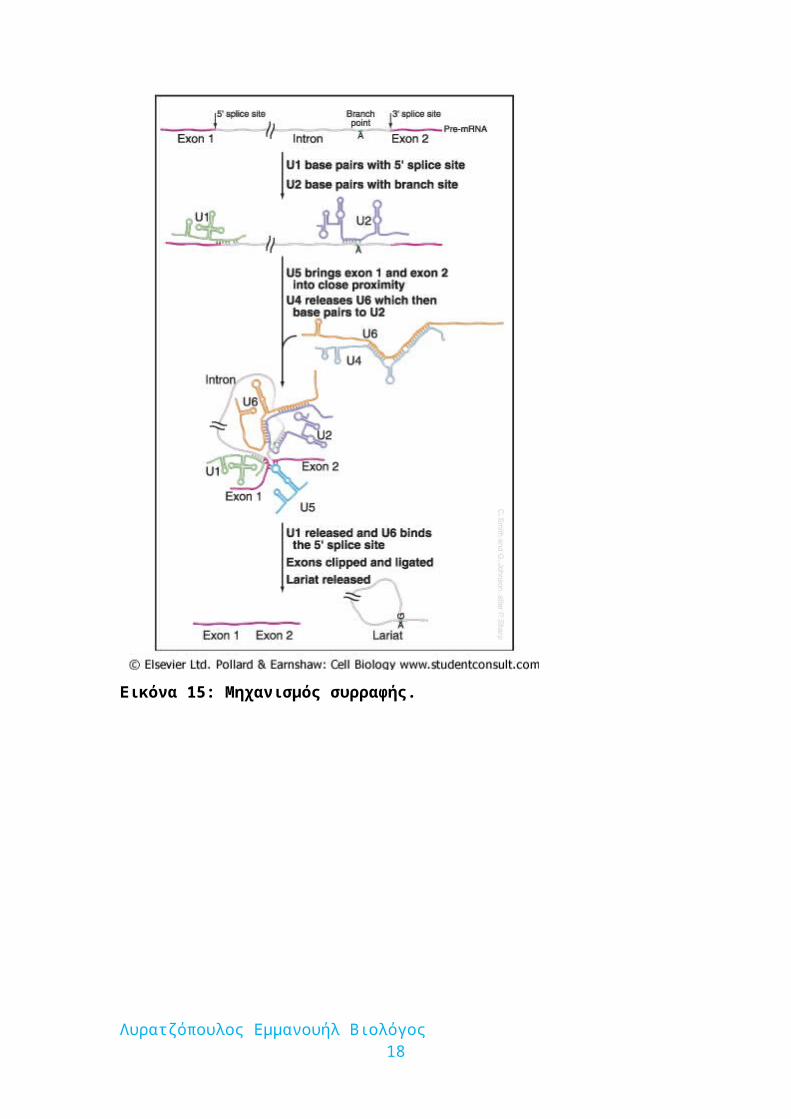
Εικόνα 15: Μηχανισμός συρραφής.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 14
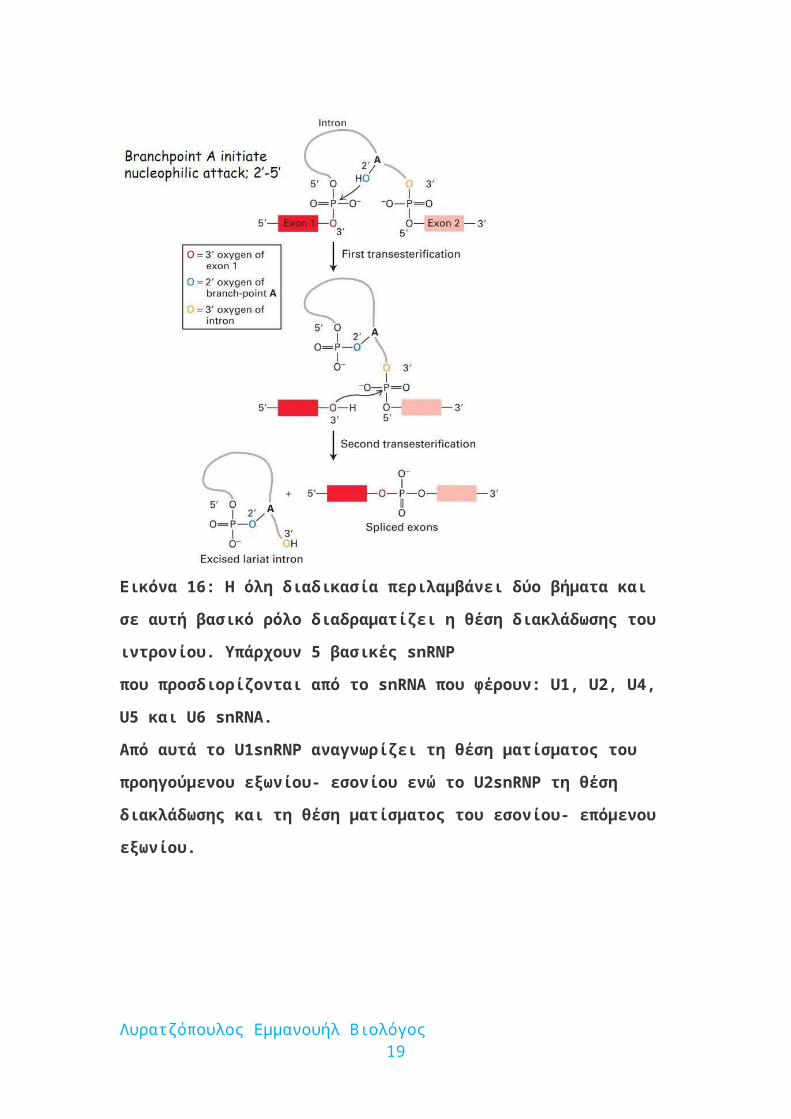
Εικόνα 16: Η όλη διαδικασία περιλαμβάνει δύο βήματα και σε αυτή βασικό ρόλο
διαδραματίζει η θέση διακλάδωσης του ιντρονίου. Υπάρχουν 5 βασικές snRNP
που προσδιορίζονται από το snRNA που φέρουν: U1, U2, U4, U5 και U6 snRNA.
Από αυτά το U1snRNP αναγνωρίζει τη θέση ματίσματος του προηγούμενου
εξωνίου- εσονίου ενώ το U2snRNP τη θέση διακλάδωσης και τη θέση ματίσματος
του εσονίου- επόμενου εξωνίου.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 15
Εικόνα 17: Η αποκοπή και η συρραφή ενός συγκεκριμένου πρόδρομου mRNA
γίνεται με συγκρότημα του συμπλόκου πάνω στο πρόδρομο mRNA. Συγκεκριμένα
το πρώτο βήμα στη συγκρότηση της μηχανής αποκοπής και συρραφής του mRNA
είναι η δέσμευση του U1 snRNP στο 5' σημείο αποκοπής του ιντρονίου. Η
δέσμευση οφείλεται στη συμπληρωματικότητα του U1 snRNP με το πρόδρομο
mRNA. Κατόπιν δεσμεύεται το U2 snRNP με παρόμοιο μηχανισμό και ακολουθεί η
δέσμευση στο U2 snRNP, του προσχηματισμένου συμπλόκου U4, U5, U6 με
ταυτόχρονη απελευθέρωση του U1 snRNP. Στο σημείο αυτό γίνονται διάφορες
αναδιατάξεις και διασυνδέσεις μεταξύ των επιμέρους συμπλόκων με τελικό
αποτέλεσμα την αποκοπή του ιντρονίου στο 5' και στο 3' άκρο του με τη μορφή
θηλιάς. Το συστατικό που προκαλεί δύο τομές στο πρόδρομο mRNA που
απαιτούνται για την απελευθέρωση του ιντρονίου είναι το U6 RNA. Όμως
φαίνεται ότι η μηχανή αποκοπής και συρραφής του πρόδρομου mRNA δεν
αναγνωρίζει τις αλληλουχίες των ιντρονίων, αλλά μάλλον των εξωνίων. Επιπλέον
Λυρατζόπουλος Εμμανουήλ Βιολόγος 16
καθοριστικό ρόλο στην επιλογή των θέσεων αποκοπής πρέπει να έχει και μια
άλλη κατηγορία πρωτεϊνών γνωστών ως SR - πρωτεΐνες επειδή περιέχουν συχνά
το διπεπτίδιο S (σερίνη) και R (αργινίνη).
Εικόνα 18: Μετάλλαξη παρεμποδίζει τη σύνδεση του U1 snRNP.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 17
Εικόνα 19: Τρανσεστεροποιήσεις στο μηχανισμό του ματίσματος.
Αρχικά το U1snRNP προσδένεται στην περιοχή Ε1-Ι, στην περιοχή διακλάδωσης
συνδέεται ο SF1 και στην περιοχή Ι-Ε2 ο U2 AF. Ο τελευταίος προσελκύει το
U2snRNP και απομακρύνεται ο SF1.
Το spliceosome ολοκληρώνεται με την προσθήκη και των υπολοίπων snRNPs
(U4/U6/U5). Το spliceosome ενεργοποιείται με την απομάκρυνση των U1, U4
ώστε να ολοκληρωθεί η διπλή αντίδραση transεστεροποίησης. Το spliceosome
λύεται και το εσόνιο (δομή λάσου) απομακρύνεται.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 18
Εικόνα 20: Σύνδεση U1 snRNP και U2 snRNP με τη βοήθεια των πρωτεινών SR.
Εικόνα 21: Εναλλακτικό μάτισμα στο καθορισμό φύλου στη Drosophila.
Λυρατζόπουλος Εμμανουήλ Βιολόγος 19

















![Introduction Abstract - Neurology...medulla oblongata were dissected [27] and were stored in RNA later solution for RNA isolation. Whole brain (n = 5 per group) weighing 80-90mg) and](https://static.fdocument.org/doc/165x107/5f7aaac355c0bb44193d6438/introduction-abstract-neurology-medulla-oblongata-were-dissected-27-and.jpg)