
ΓΟΝΙΔΙΑΚΗ...
31
1 ΓΟΝΙΔΙΑΚΗ ΑΠΟΣΙΩΠΗΣΗ Αγγελίδης Χαράλαμπος, Αν. Καθηγητής Βιολογίας, 2010. Γονιδιακή σίγηση ή αποσιώπηση είναι ένας γενικός όρος που περιγράφει επιγενετικές διαδικασίες της γονιδιακής ρύθμισης. Ο όρος γονιδιακή αποσιώπηση χρησιμοποιείται γενικά για να περιγράψει το «σβήσιμο» ενός γονιδίου από ένα μηχανισμό διαφορετικό από τη γενετική τροποποίηση. Δηλαδή, ένα γονίδιο το οποίο θα εκφραζόταν (ενεργοποιημένο) υπό κανονικές συνθήκες είναι απενεργοποιημένο από τις μηχανές στο κύτταρο. Τα γονίδια ρυθμίζονται είτε σε μεταγραφικό και μετα-μεταγραφικό επίπεδο. Η μεταγραφική σίγηση γονιδίων είναι το αποτέλεσμα των τροποποιήσεων των ιστονών, δημιουργώντας ένα περιβάλλον ετεροχρωματίνης γύρω από ένα γονίδιο που το καθιστά απρόσιτο σε μεταγραφική μηχανή (RNA πολυμεράση, μεταγραφικούς παράγοντες, κ.λπ.). Η μετα- μεταγραφική σίγηση γονιδίων είναι το αποτέλεσμα του mRNA ενός συγκεκριμένου γονιδίου το οποίο καταστρέφεται ή μπλοκάρεται. Η καταστροφή του mRNA εμποδίζει η μετάφραση να διαμορφώσει ένα ενεργό προϊόν του γονιδίου (στις περισσότερες περιπτώσεις, μια πρωτεΐνη). Ένας κοινός μηχανισμός της μετα- μεταγραφικής σίγησης των γονιδίων είναι το RNAi. Τόσο η μεταγραφική όσο και η μετα-μεταγραφική σίγηση γονιδίων χρησιμοποιούνται για τη ρύθμιση των ενδογενών γονιδίων. Μηχανισμοί γονιδιακής σίγησης, επίσης προστατεύουν το γονιδίωμα του οργανισμού του από μεταθετά στοιχεία (τρανσποζόνια) και ιούς. Ετσι, η γονιδιακή αποσιώπηση (Gene silencing) μπορεί να είναι μέρος ενός αρχαίου ανοσοποιητικού συστήματος προστασίας από τέτοιου είδους μολυσματική DNA στοιχεία. Κατά τα τελευταία χρόνια, μόρια RNA των ~20-30 νουκλεοτιδίων έχουν αναδειχθεί ως κρίσιμης σημασίας στοιχεία στην έκφραση και τη λειτουργία των ευκαρυωτικών γονιδιωμάτων. Δύο κύριες κατηγορίες αυτών των μικρών RNAs- σύντομης παρέμβασης RNAs (Shortt interfering: siRNAs) και microRNAs (miRNAs)-ενεργούν τόσο σε σωματικά όσο και γαμετικά κύτταρα ενός ευρέως φάσματος ευκαρυωτικών ειδών για τη ρύθμιση των ενδογενών γονιδίων και για την υπεράσπιση του γονιδιώματος από νουκλεϊνικά οξέα τα οποία εισβάλλουν. Το RNA interference (RNAi) είναι ένα πανάρχαιο συντηρημένο μονοπάτι (Zamore, P.D., et al., 2000). Σήμερα, γνωρίζοντας πολλά για τον μηχανισμό αυτόν χρησιμοποιούμε στοιχεία του για να παρέμβουμε στα κύτταρα στοχοποιώντας κάποια γονίδια των οποίων η έκφραση είναι ιδιαίτερα βλαβερή για κάποιο είδος κυττάρων. Έτσι, η παρέμβαση μέσω RNA [RNA-mediated interference (RNAi)] στα κύτταρα, έχει σαν σκοπό την αναστολή της έκφρασης των γονιδίων
Transcript of ΓΟΝΙΔΙΑΚΗ...

1
ΓΟΝΙΔΙΑΚΗ ΑΠΟΣΙΩΠΗΣΗ
Αγγελίδης Χαράλαμπος, Αν. Καθηγητής Βιολογίας, 2010.
Γονιδιακή σίγηση ή αποσιώπηση είναι ένας γενικός όρος που περιγράφει
επιγενετικές διαδικασίες της γονιδιακής ρύθμισης. Ο όρος γονιδιακή αποσιώπηση
χρησιμοποιείται γενικά για να περιγράψει το «σβήσιμο» ενός γονιδίου από ένα
μηχανισμό διαφορετικό από τη γενετική τροποποίηση. Δηλαδή, ένα γονίδιο το
οποίο θα εκφραζόταν (ενεργοποιημένο) υπό κανονικές συνθήκες είναι
απενεργοποιημένο από τις μηχανές στο κύτταρο. Τα γονίδια ρυθμίζονται είτε σε
μεταγραφικό και μετα-μεταγραφικό επίπεδο. Η μεταγραφική σίγηση γονιδίων είναι
το αποτέλεσμα των τροποποιήσεων των ιστονών, δημιουργώντας ένα περιβάλλον
ετεροχρωματίνης γύρω από ένα γονίδιο που το καθιστά απρόσιτο σε μεταγραφική
μηχανή (RNA πολυμεράση, μεταγραφικούς παράγοντες, κ.λπ.). Η μετα-
μεταγραφική σίγηση γονιδίων είναι το αποτέλεσμα του mRNA ενός συγκεκριμένου
γονιδίου το οποίο καταστρέφεται ή μπλοκάρεται. Η καταστροφή του mRNA
εμποδίζει η μετάφραση να διαμορφώσει ένα ενεργό προϊόν του γονιδίου (στις
περισσότερες περιπτώσεις, μια πρωτεΐνη). Ένας κοινός μηχανισμός της μετα-
μεταγραφικής σίγησης των γονιδίων είναι το RNAi. Τόσο η μεταγραφική όσο και η
μετα-μεταγραφική σίγηση γονιδίων χρησιμοποιούνται για τη ρύθμιση των
ενδογενών γονιδίων. Μηχανισμοί γονιδιακής σίγησης, επίσης προστατεύουν το
γονιδίωμα του οργανισμού του από μεταθετά στοιχεία (τρανσποζόνια) και ιούς.
Ετσι, η γονιδιακή αποσιώπηση (Gene silencing) μπορεί να είναι μέρος ενός
αρχαίου ανοσοποιητικού συστήματος προστασίας από τέτοιου είδους μολυσματική
DNA στοιχεία.
Κατά τα τελευταία χρόνια, μόρια RNA των ~20-30 νουκλεοτιδίων έχουν
αναδειχθεί ως κρίσιμης σημασίας στοιχεία στην έκφραση και τη λειτουργία των
ευκαρυωτικών γονιδιωμάτων. Δύο κύριες κατηγορίες αυτών των μικρών RNAs-
σύντομης παρέμβασης RNAs (Shortt interfering: siRNAs) και microRNAs
(miRNAs)-ενεργούν τόσο σε σωματικά όσο και γαμετικά κύτταρα ενός ευρέως
φάσματος ευκαρυωτικών ειδών για τη ρύθμιση των ενδογενών γονιδίων και για
την υπεράσπιση του γονιδιώματος από νουκλεϊνικά οξέα τα οποία εισβάλλουν.
Το RNA interference (RNAi) είναι ένα πανάρχαιο συντηρημένο μονοπάτι
(Zamore, P.D., et al., 2000). Σήμερα, γνωρίζοντας πολλά για τον μηχανισμό
αυτόν χρησιμοποιούμε στοιχεία του για να παρέμβουμε στα κύτταρα
στοχοποιώντας κάποια γονίδια των οποίων η έκφραση είναι ιδιαίτερα βλαβερή για
κάποιο είδος κυττάρων. Έτσι, η παρέμβαση μέσω RNA [RNA-mediated interference
(RNAi)] στα κύτταρα, έχει σαν σκοπό την αναστολή της έκφρασης των γονιδίων

2
που έχουμε θέσει σαν στόχο, χρησιμοποιώντας διπλής αλυσίδας RNAs (Fire, A.,
1998). Σχετικά πρόσφατα, αποδείχθηκε ότι το siRNA (short interfering RNA) είναι
πολύ αποδοτικό για γονιδιακή αποσιώπηση στα κύτταρα των θηλαστικών
(Brummelkamp, T.R., et al., 2002).
To microRNA, συνιστά μία μεγάλη κατηγορία από εξελεκτικά συντηρημένo,
μη-κωδικοποιό, RNA που ξεκινά από μεγαλύτερα μεταγραφήματα που
χαρακτηρίζονται από ατελείς δομές φουρκέτας. Τα miRNAs είναι 19–23 nt RNAs
που προέρχονται από πρόδρομα pre-miRNA μέσω Dicer, του ίδιου ενζύμου που
επεξεργάζεται τα siRNAs
Η διαφορά μεταξύ των 2 μονοπατιών, siRNA και miRNA, για την γονιδιακή
αποσιώπηση συνίσταται στα παρακάτω. Στο siRNA μονοπάτι, το RNA προέρχεται
από εξωγενές RNA, από τρανσποζόνια ή από ϊούς, ενώ στο miRNA μονοπάτι το
RNA προέρχεται από to miRNA γονίδιο.
Οι πρόσφατες εξελίξεις έδειξαν απρόσμενη ποικιλία στα μονοπάτια
βιογένεσής και στους ρυθμιστικούς μηχανισμούς που έχουν πρόσβαση. Η
κατανόηση των μηχανισμών ρύθμισης μέσω siRNA και miRNA θα έχει άμεσες
επιπτώσεις για την θεμελιώδη βιολογία, καθώς και για την αιτιολογία νόσων και
της θεραπείας των.
siRNA (Short Interferance RNA) ή RNAi (RNA
intereference): Μικρά μόρια RNA μπορούν να αποσιωπήσουν την γονιδιακή έκφραση [Tropp B.E. (2008)]
Το 1998, οι Andrew Fire, Craig C.Mello και οι συνεργάτες τους ανακάλυψαν
έναν σημαντικό μηχανισμό ο οποίος επιτρέπει τα ευκαρυωτικά να αποσιωπήσουν
ειδικά μόρια mRNAs. Όταν ξεκίνησαν τις μελέτες τους ήταν γνωστό ότι ενίοντας
νοηματικό ή αντινοηματικό RNA μέσα σε σκώληκες του τύπου C. Elegants
μπλοκάρεται η έκφραση ειδικού γονιδίου και επίσης ότι η αναστολή εξακολουθεί
να υφίσταται και στην επόμενη γενεά. Αυτή η παρατήρηση φαινόταν
αντισυμβατική με το δεδομένο ότι τα κύτταρα ταχέως καταβολίζουν τα απλής
αλυσίδας μόρια RNA όπως το mRNA. Έτσι η προαναφερόμενη ομάδα υποψιάστηκε
ότι τα απλής αλυσίδας μόρια του RNA τα οποία χρησιμοποιήθηκαν για να ενέσουνε
τα σκουλήκια θα περιείχαν πιθανώς μερικά μόρια διπλής αλυσίδας RNA τα οποία
και θα ήταν βιολογικώς ενεργά. Αποφάσισαν να ελέγξουν την υπόθεση αυτή
μελετώντας την έκφραση ενός μη σημαντικού γονιδίου του myofilament gene του
C. Elegans μετά από ένεση μακριάς νοηματικής αλυσίδας 742 νουκλεοτιδίων, της
3
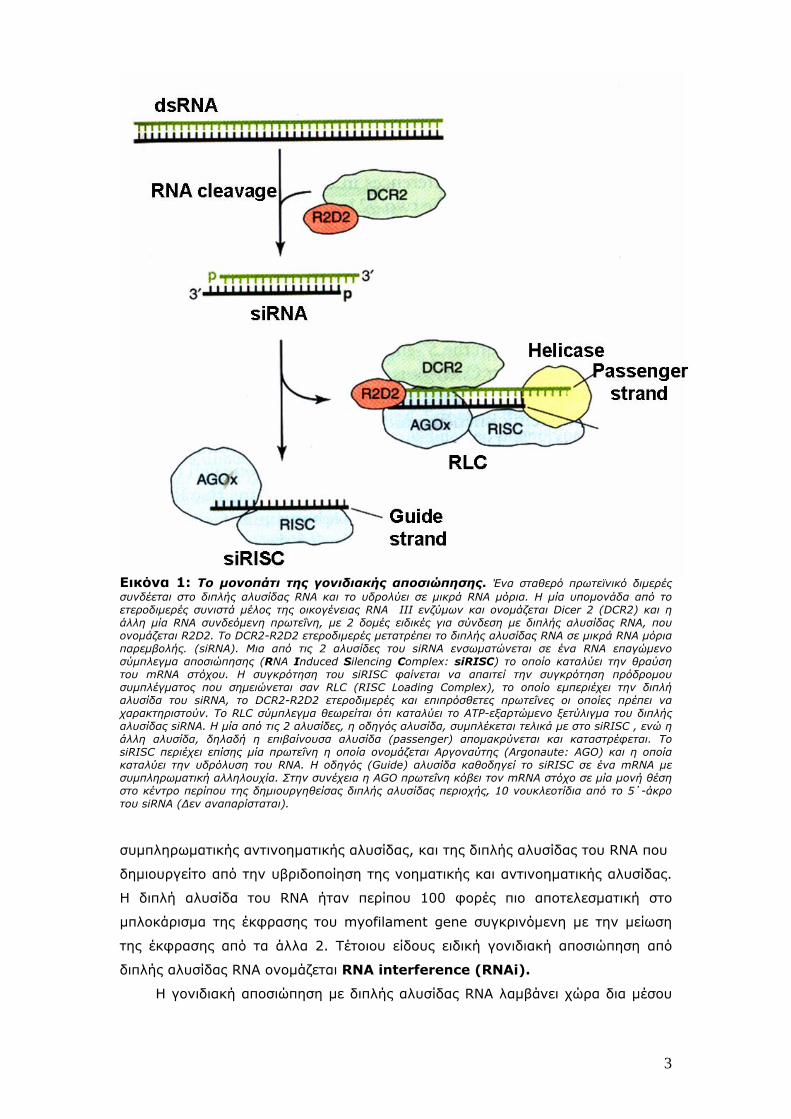
Εικόνα 1: Το μονοπάτι της γονιδιακής αποσιώπησης. Ένα σταθερό πρωτεϊνικό διμερές συνδέεται στο διπλής αλυσίδας RNA και το υδρολύει σε μικρά RNA μόρια. Η μία υπομονάδα από το ετεροδιμερές συνιστά μέλος της οικογένειας RNA III ενζύμων και ονομάζεται Dicer 2 (DCR2) και η άλλη μία RNA συνδεόμενη πρωτεΐνη, με 2 δομές ειδικές για σύνδεση με διπλής αλυσίδας RNA, που ονομάζεται R2D2. Το DCR2-R2D2 ετεροδιμερές μετατρέπει το διπλής αλυσίδας RNA σε μικρά RNA μόρια παρεμβολής. (siRNA). Mια από τις 2 αλυσίδες του siRNA ενσωματώνεται σε ένα RNA επαγώμενο σύμπλεγμα αποσιώπησης (RNA Induced Silencing Complex: siRISC) το οποίο καταλύει την θραύση του mRNA στόχου. Η συγκρότηση του siRISC φαίνεται να απαιτεί την συγκρότηση πρόδρομου συμπλέγματος που σημειώνεται σαν RLC (RISC Loading Complex), το οποίο εμπεριέχει την διπλή αλυσίδα του siRNA, το DCR2-R2D2 ετεροδιμερές και επιπρόσθετες πρωτεΐνες οι οποίες πρέπει να χαρακτηριστούν. Το RLC σύμπλεγμα θεωρείται ότι καταλύει το ATP-εξαρτώμενο ξετύλιγμα του διπλής αλυσίδας siRNA. Η μία από τις 2 αλυσίδες, η οδηγός αλυσίδα, συμπλέκεται τελικά με στο siRISC , ενώ η άλλη αλυσίδα, δηλαδή η επιβαίνουσα αλυσίδα (passenger) απομακρύνεται και καταστρέφεται. Το siRISC περιέχει επίσης μία πρωτεΐνη η οποία ονομάζεται Αργοναύτης (Argonaute: AGO) και η οποία καταλύει την υδρόλυση του RNA. H οδηγός (Guide) αλυσίδα καθοδηγεί το siRISC σε ένα mRNA με συμπληρωματική αλληλουχία. Στην συνέχεια η AGO πρωτεΐνη κόβει τον mRNA στόχο σε μία μονή θέση στο κέντρο περίπου της δημιουργηθείσας διπλής αλυσίδας περιοχής, 10 νουκλεοτίδια από το 5΄-άκρο του siRNA (Δεν αναπαρίσταται).
συμπληρωματικής αντινοηματικής αλυσίδας, και της διπλής αλυσίδας του RNA που
δημιουργείτο από την υβριδοποίηση της νοηματικής και αντινοηματικής αλυσίδας.
Η διπλή αλυσίδα του RNA ήταν περίπου 100 φορές πιο αποτελεσματική στο
μπλοκάρισμα της έκφρασης του myofilament gene συγκρινόμενη με την μείωση
της έκφρασης από τα άλλα 2. Τέτοιου είδους ειδική γονιδιακή αποσιώπηση από
διπλής αλυσίδας RNA ονομάζεται RNA interference (RNAi).
H γονιδιακή αποσιώπηση με διπλής αλυσίδας RNA λαμβάνει χώρα δια μέσου

4
ενός πολλαπλών σταδίων μονοπατιού, το οποίο είναι ακραιφνώς το ίδιο σε όλα τα
ευκαρυωτικά που έχουν μελετηθεί. Όμως, τα ονόματα που προσδίδονται σε
διάφορους πρωτεϊνικούς παράγοντες και ένζυμα τα οποία συμμετέχουν στο
μονοπάτι, διαφέρουν από είδος σε είδος. Για να αποφευχθεί το κομφούζιο της
ονοματολογίας, η παρούσα παρουσίαση περιορίζεται στο μονοπάτι της Δροσόφιλας
(Εικόνα 1). Αυτό το μονοπάτι ξεκινά από ένα σταθερό πρωτεϊνικό ετεροδιμερές το
οποίο συνδέεται στο διπλής αλυσίδας RNA που στην συνέχεια το υδρολύει σε μικρά
RNA μόρια. Μία υπομονάδα από το ετεροδιμερές συνιστά ένα μέλος από την
RNase-III ενζυμική οικογένεια που καλείται Dicer 2 (DCR2) και η άλλη είναι μία
RNA συνδεόμενη πρωτεΐνη με 2 RNA συνδεόμενες δομές και ονομάζεται R2D2.
To ετεροδιμερές DCR2/ R2D2 μετατρέπει το διπλής αλυσίδας RNA σε μικρά μόρια
RNA παρεμβολής (siRNA: short interfering RNA), τα οποία είναι μήκους 21-23
νουκλεοτιδίων διπλής αλυσίδας RNAs με συμμετρικά 2-3 νουκλεοτίδια
προεξέχοντα στα 3΄άκρα και φωσφορικές ομάδες στα 5΄άκρα. Αμφότερα, τα
DCR2 και R2D2, θα πρέπει να είναι ενεργά για να λειτουργήσει το μονοπάτι του
siRNA. Η μία από τις δύο αλυσίδες του siRNA ενσωματώνεται μέσα στο σύμπλεγμα
siRISC (RNA Induced Silencing Complex) το οποίο και καταλύει την θραύση του
στοχοποιημένου RNA. H συγκρότηση του siRISC απαιτεί προηγουμένως την
συγκρότηση του πρόδρομου συμπλέγματος RLC (RISC Loading Complex), το οποίο
περιέχει το δίκλωνο siRNA, το ετεροδιμερές DCR2/ R2D2 και επιπρόσθετες
πρωτεΐνες οι οποίες πρέπει να χαρακτηριστούν.
Θεωρείται ότι το RLC καταλύει κατά ATP-εξαρτώμενο τρόπο το ξετύλιγμα του
δίκλωνου siRNA. Η μία από τις δύο αλυσίδες, η οδηγός αλυσίδα (guide strand),
τελικά ενσωματώνεται στο σύμπλεγμα siRISC, ενώ η άλλη αλυσίδα, η επιβαίνουσα
αλυσίδα (passenger strand), απομακρύνεται και καταστρέφεται. Διαφορές στις
θερμοδυναμικές σταθερότητες του ζευγαρώματος των βάσεων στα 5΄άκρα των 2
αλυσίδων του siRNA, που η κάθε μία συνίσταται από 21 νουκλεοτίδια, καθωρίζει
για το ποια αλυσίδα siRNA θα γίνει η οδηγός αλυσίδα η οποία και ενσωματώνεται
στο siRISC. Το σύμπλεγμα siRISC περιέχει επίσης και μία πρωτεΐνη που ονομάζεται
Αργοναύτης (AGO: Argonaute) η οποία καταλύει την υδρόλυση του RNA. H
οδηγός αλυσίδα κατευθύνει το siRISC σε ένα mRNA με συμπληρωματική
αλληλουχία. Στην συνέχεια η πρωτεΐνη Αργοναύτης κόβει το στοχοποιημένο mRNA
σε μία θέση περίπου στο κέντρο της διπλής αλυσίδας, δηλαδή 10 περίπου
νουκλεοτίδια από το 5΄άκρο του siRNA (Εικόνα 2).
Η RNA παρεμβολή στα C. Elegans μπορεί να απλωθεί σε όλον τον
οργανισμό, ακόμη και όταν αυτή πυροδοτηθεί για 1 λεπτό από ποσότητα διπλής
αλυσίδας RNA. Παρόμοια συστηματική αποσιώπηση έχει επίσης παρατηρηθεί στα
φυτά αλλά όχι στα σπονδυλωτά ή στην Δροσόφιλα. Στα C. Elegans ή στα φυτά θα
5
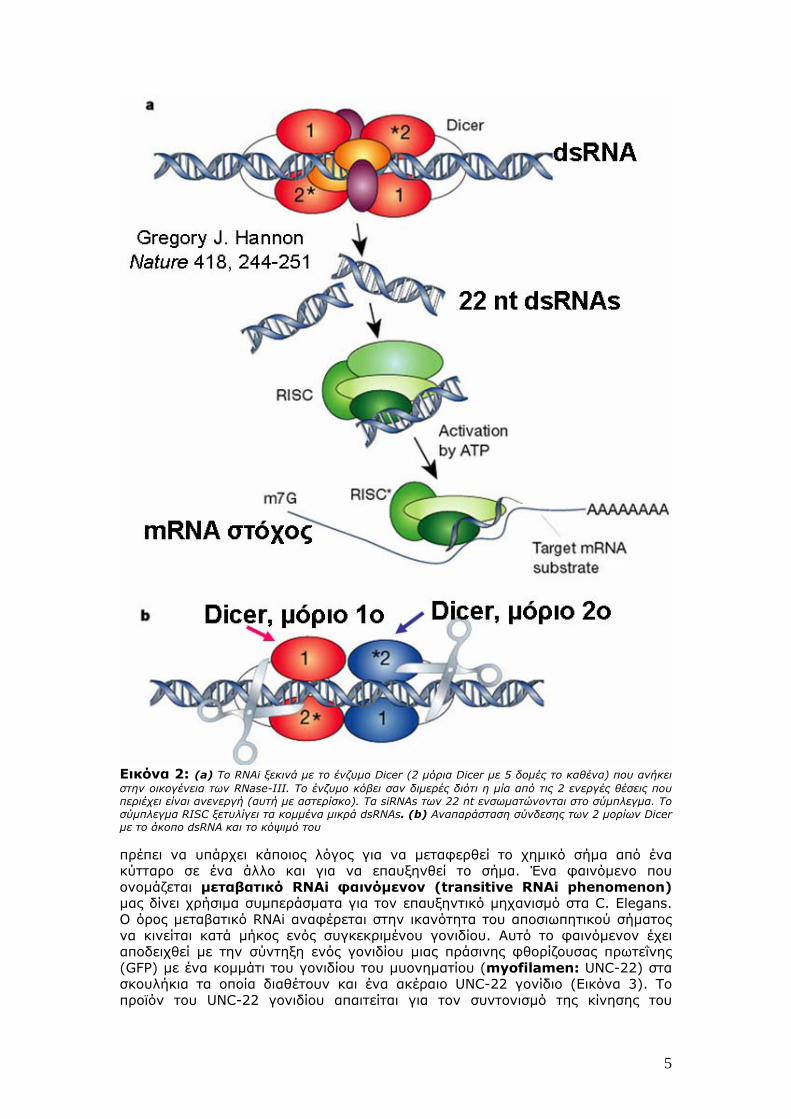
Εικόνα 2: (a) To RNAi ξεκινά με το ένζυμο Dicer (2 μόρια Dicer με 5 δομές το καθένα) που ανήκει στην οικογένεια των RNase-III. To ένζυμο κόβει σαν διμερές διότι η μία από τις 2 ενεργές θέσεις που περιέχει είναι ανενεργή (αυτή με αστερίσκο). Τα siRNAs των 22 nt ενσωματώνονται στο σύμπλεγμα. Το σύμπλεγμα RISC ξετυλίγει τα κομμένα μικρά dsRNAs. (b) Aναπαράσταση σύνδεσης των 2 μορίων Dicer με το άκοπο dsRNA και το κόψιμό του πρέπει να υπάρχει κάποιος λόγος για να μεταφερθεί το χημικό σήμα από ένα κύτταρο σε ένα άλλο και για να επαυξηνθεί το σήμα. Ένα φαινόμενο που ονομάζεται μεταβατικό RNAi φαινόμενον (transitive RNAi phenomenon) μας δίνει χρήσιμα συμπεράσματα για τον επαυξηντικό μηχανισμό στα C. Elegans. Ο όρος μεταβατικό RNAi αναφέρεται στην ικανότητα του αποσιωπητικού σήματος να κινείται κατά μήκος ενός συγκεκριμένου γονιδίου. Αυτό το φαινόμενον έχει αποδειχθεί με την σύντηξη ενός γονιδίου μιας πράσινης φθορίζουσας πρωτεΐνης (GFP) με ένα κομμάτι του γονιδίου του μυονηματίου (myofilamen: UNC-22) στα σκουλήκια τα οποία διαθέτουν και ένα ακέραιο UNC-22 γονίδιο (Εικόνα 3). To προϊόν του UNC-22 γονιδίου απαιτείται για τον συντονισμό της κίνησης του
6
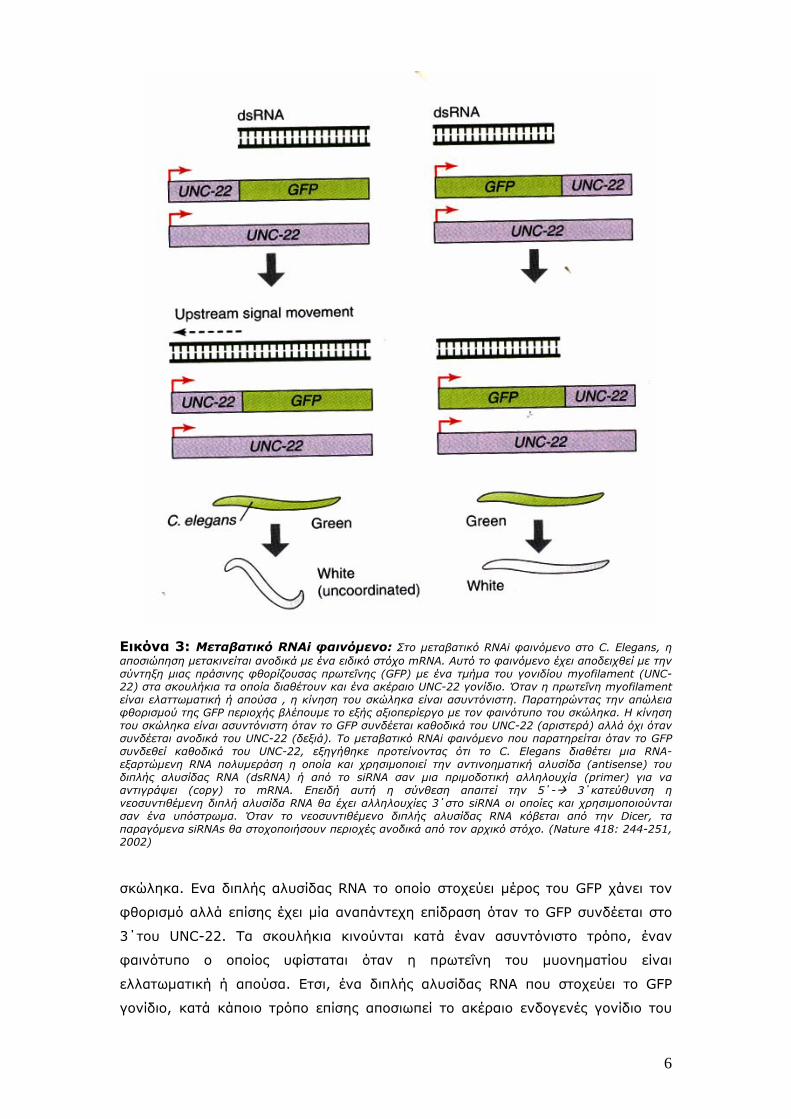
Εικόνα 3: Μεταβατικό RNAi φαινόμενο: Στο μεταβατικό RNAi φαινόμενο στο C. Elegans, η αποσιώπηση μετακινείται ανοδικά με ένα ειδικό στόχο mRNA. Αυτό το φαινόμενο έχει αποδειχθεί με την σύντηξη μιας πράσινης φθορίζουσας πρωτεΐνης (GFP) με ένα τμήμα του γονιδίου myofilament (UNC-22) στα σκουλήκια τα οποία διαθέτουν και ένα ακέραιο UNC-22 γονίδιο. Όταν η πρωτεΐνη myofilament είναι ελαττωματική ή απούσα , η κίνηση του σκώληκα είναι ασυντόνιστη. Παρατηρώντας την απώλεια φθορισμού της GFP περιοχής βλέπουμε το εξής αξιοπερίεργο με τον φαινότυπο του σκώληκα. Η κίνηση του σκώληκα είναι ασυντόνιστη όταν το GFP συνδέεται καθοδικά του UNC-22 (αριστερά) αλλά όχι όταν συνδέεται ανοδικά του UNC-22 (δεξιά). Το μεταβατικό RNAi φαινόμενο που παρατηρείται όταν το GFP συνδεθεί καθοδικά του UNC-22, εξηγήθηκε προτείνοντας ότι το C. Elegans διαθέτει μια RNA-εξαρτώμενη RNA πολυμεράση η οποία και χρησιμοποιεί την αντινοηματική αλυσίδα (antisense) του διπλής αλυσίδας RNA (dsRNA) ή από το siRNA σαν μια πριμοδοτική αλληλουχία (primer) για να αντιγράψει (copy) το mRNA. Επειδή αυτή η σύνθεση απαιτεί την 5΄- 3΄κατεύθυνση η νεοσυντιθέμενη διπλή αλυσίδα RNA θα έχει αλληλουχίες 3΄στο siRNA οι οποίες και χρησιμοποιούνται σαν ένα υπόστρωμα. Όταν το νεοσυντιθέμενο διπλής αλυσίδας RNA κόβεται από την Dicer, τα παραγόμενα siRNAs θα στοχοποιήσουν περιοχές ανοδικά από τον αρχικό στόχο. (Nature 418: 244-251, 2002)
σκώληκα. Ενα διπλής αλυσίδας RNA το οποίο στοχεύει μέρος του GFP χάνει τον
φθορισμό αλλά επίσης έχει μία αναπάντεχη επίδραση όταν το GFP συνδέεται στο
3΄του UNC-22. Τα σκουλήκια κινούνται κατά έναν ασυντόνιστο τρόπο, έναν
φαινότυπο ο οποίος υφίσταται όταν η πρωτεΐνη του μυονηματίου είναι
ελλατωματική ή απούσα. Ετσι, ένα διπλής αλυσίδας RNA που στοχεύει το GFP
γονίδιο, κατά κάποιο τρόπο επίσης αποσιωπεί το ακέραιο ενδογενές γονίδιο του
7
μυονημάτιου. Αυτό το φαινόμενο, το οποίο παρατηρείται μόνο όταν το GFP
συνδεθεί στο 3΄άκρο του UNC-22 γονιδίου, μπορεί να εξηγηθεί με το ότι το C.
Elegans διαθέτει μια RNA-εξαρτώμενη RNA πολυμεράση (την RdRp: RNA
directed RNA polymerase) η οποία και χρησιμοποιεί την αντινοηματική αλυσίδα
(antisense) του διπλής αλυσίδας RNA (dsRNA) ή από το siRNA σαν μια
πριμοδοτική αλληλουχία (primer) για να αντιγράψει (copy) το mRNA (Εικόνα 4) .
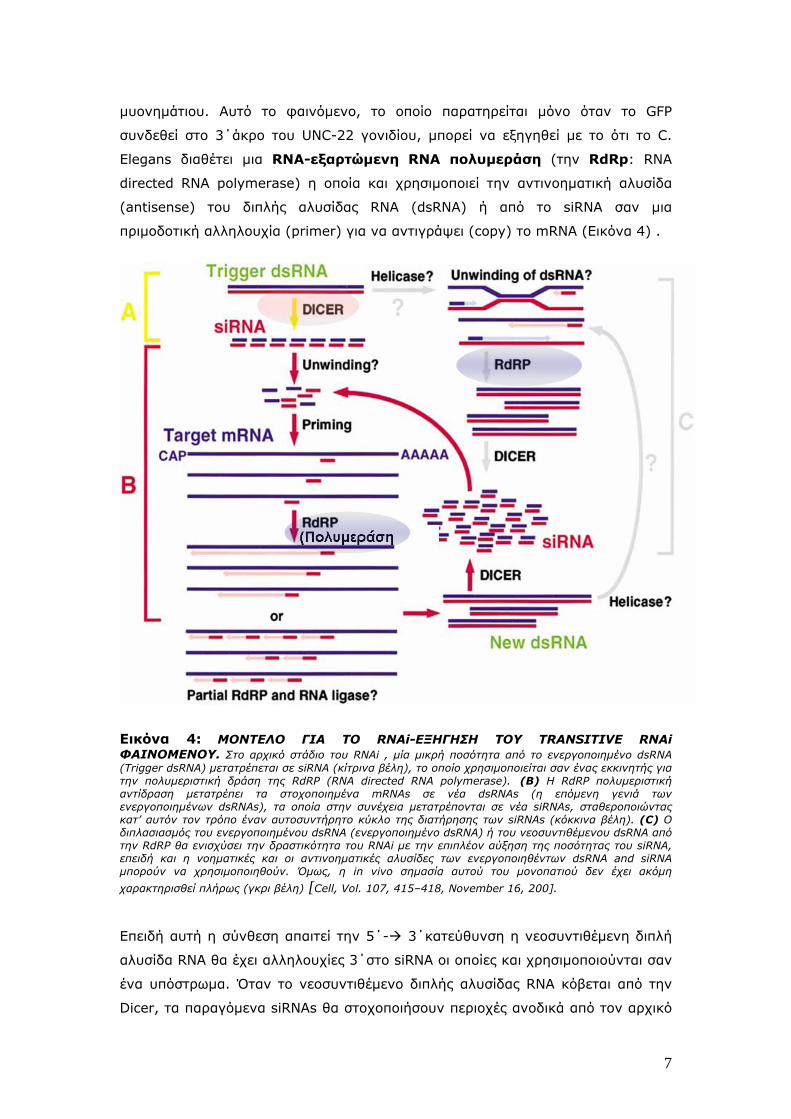
Εικόνα 4: MONTEΛΟ ΓΙΑ ΤΟ RNAi-ΕΞΗΓΗΣΗ ΤΟΥ TRANSITIVE RNAi ΦΑΙΝΟΜΕΝΟΥ. Στο αρχικό στάδιο του RNAi , μία μικρή ποσότητα από το ενεργοποιημένο dsRNA (Trigger dsRNA) μετατρέπεται σε siRNA (κίτρινα βέλη), το οποίο χρησιμοποιείται σαν ένας εκκινητής για την πολυμεριστική δράση της RdRP (RNA directed RNA polymerase). (B) Η RdRP πολυμεριστική αντίδραση μετατρέπει τα στοχοποιημένα mRNAs σε νέα dsRNAs (η επόμενη γενιά των ενεργοποιημένων dsRNAs), τα οποία στην συνέχεια μετατρέπονται σε νέα siRNAs, σταθεροποιώντας κατ’ αυτόν τον τρόπο έναν αυτοσυντήρητο κύκλο της διατήρησης των siRNAs (κόκκινα βέλη). (C) Ο διπλασιασμός του ενεργοποιημένου dsRNA (ενεργοποιημένο dsRNA) ή του νεοσυντιθέμενου dsRNA από την RdRP θα ενισχύσει την δραστικότητα του RNAi με την επιπλέον αύξηση της ποσότητας του siRNA, επειδή και η νοηματικές και οι αντινοηματικές αλυσίδες των ενεργοποιηθέντων dsRNA and siRNA μπορούν να χρησιμοποιηθούν. Όμως, η in vivo σημασία αυτού του μονοπατιού δεν έχει ακόμη
χαρακτηρισθεί πλήρως (γκρι βέλη) [Cell, Vol. 107, 415–418, November 16, 200].
Επειδή αυτή η σύνθεση απαιτεί την 5΄- 3΄κατεύθυνση η νεοσυντιθέμενη διπλή
αλυσίδα RNA θα έχει αλληλουχίες 3΄στο siRNA οι οποίες και χρησιμοποιούνται σαν
ένα υπόστρωμα. Όταν το νεοσυντιθέμενο διπλής αλυσίδας RNA κόβεται από την
Dicer, τα παραγόμενα siRNAs θα στοχοποιήσουν περιοχές ανοδικά από τον αρχικό

8
στόχο. (Nature 418: 244-251, 2002). Επειδή τα νεοδημιουργούμενα siRNA επίσης
θα συνδεθούν και με τα μεταγραφήματα του φυσιολογικού γονιδίου του
μυονηματίου, αυτό το γονίδιο επίσης θα αποσιωπηθεί. RNA-εξαρτώμενες RNA
πολυμεράσες έχουν αναφερθεί στα φυτά και τους σκώληκες όχι όμως στα
σπονδυλωτά. Η παρουσία της RNA-εξαρτώμενης RNA πολυμεράσης στους
σκώληκες επίσης εξηγεί πως το σήμα επεκτείνεται και είναι ικανό να υφίσταται
ακόμη και στην επόμενη γενεά. Το μεταβατικό RNAi φαινόμενο λαμβάνει μέρος
επίσης και στα φυτά. Όμως, το σήμα εμφανίζεται να είναι ικανό να μετακινείται και
ανοδικά και καθοδικά από τον αρχικό στόχο. Μέχρι στιγμής δεν έχει ακόμη ευρεθεί
μία αποδεκτή εξήγηση για την εκατέρωθεν κίνηση.
Το RNAi είναι ένα πολύ δυναμικό εργαλείο. Στις πρόωρες περιόδους της
Μοριακής Βιολογίας, μεταλλαγμένοι φαινότυποι χρησιμοποιήθηκαν για την
ταυτοποίηση γονιδίων. Σήμερα το θέμα έχει αντιστραφεί. Χάρις στην ανάπτυξη της
τεχνολογίας για το διάβασμα του DNA (DNA sequencing) χιλιάδες γονίδια έχουν
ταυτοποιηθεί χωρίς όμως να γνωρίζουμε τις λειτουργίες τους. Μέχρι πρόσφατα, ο
μόνος δρόμος για να καθορίσουμε την λειτουργία αυτών των γονιδίων ήταν να
χρησιμοποιήσουμε τον γενετικό ανασυνδυασμό (genetic recombination) για να
αδρανοποιήσουμε τα γονίδια που μας ενδιαφέρουν. Αυτή η προσέγγιση δουλεύει
καλά σε μονοκύτταρους οργανισμούς όπως οι ζυμομήκυτες (yeast), υπάρχει όμως
μεγάλο πρόβλημα όταν μελετάμε ανώτερους οργανισμούς τέτοιους όπως τα
θηλαστικά επειδή οι μέθοδοι που χρειάζεται για να αδρανοποιήσουμε ένα ειδικό
γονίδιο (knock out) που μας ενδιαφέρει είναι πολύπλοκο και χρονοβόρο θέμα και
πολλές φορές δεν μπορεί να επιτευχθεί όταν υπάρχουν πολλά αντίγραφα ενός
γονιδίου μέσα στο γένωμα. Επίσης πολλές φορές δεν μπορούν να αδρανοποιηθούν
κάποια βασικά γονίδια διότι δεν επιβιώνει το κύτταρο. Ετσι το RNAi προσφέρει μία
εναλλακτική μέθοδο για την μελέτη της λειτουργίας ενός γονιδίου. Αυτό που
χρειάζεται είναι να συνθέσεις ένα διπλής αλυσίδας RNA με μία αλληλουχία ίδια με
το mRNA στόχο και στην συνέχεια να το εισαγάγεις μέσα στον υπό μελέτη
οργανισμό. To Dicer θα μετατρέψει το διπλής αλυσίδας RNA σε siRNA, το οποίο θα
ενσωματωθεί στο siRISC που θα στοχοποιήσει το ειδικό mRNA προς καταστροφή.
Η Julie Ahringer και οι συνεργάτες της έχουν αναπτύξει την τεχνική RNAi
για να βρίσκουν τους φαινότυπους στον σκώληκα C. Elegans για γονίδια των
οποίων η λειτουργία είναι άγνωστη. Η προσέγγισή τους στηρίζεται στο
πλεονέκτημα του ότι ο C. Elegans τρέφεται με E. Coli για την εισαγωγή διπλής
αλυσίδας RNA στους σκώληκες. Ετσι κατασκεύασαν 16.757 βακτηριακά στελέχη,
που το κάθε ένα εκφράζει διπλής αλυσίδας RNA με στόχο ένα διαφορετικό γονίδιο
του σκώληκα. Αυτά τα στελέχη συνιστούν το 86% των 19.427 γνωστών γονιδίων
του σκώληκα. Το διπλής αλυσίδας RNA που παράγεται από τα βακτήρια αποσιωπεί

9
το γονίδιο στόχο στον σκώληκα , επιτρέποντας έτσι στον ερευνητή να βρεί τον
φαινότυπο που οφείλεται στην αδρανοποίηση του γονιδίου στόχου. Σήμερα η
ομάδα αυτή έχει ήδη μελετήσει μεταλλαγμένους φαινότυπους για 1722 γονίδια
που μελετήθηκαν (έτος 2008).Τα δύο τρίτα από αυτά τα γονίδια δεν έχουν ένα
φαινότυπο που να έχει προηγούμενα χαρακτηριστεί και συσχετιστεί με αυτά. Αυτή
η προσέγγιση είναι πολύ υποσχόμενη επειδή πολλά από γονίδια των σκωλήκων
είναι ομόλογα με αυτά του ανθρώπου. Ετσι άπαξ και γνωρίσουμε τους
φαινοτύπους στον σκώληκα θα μπορούμε να υποθέσουμε και για την λειτουργία
των ανθρώπινων γονιδίων.
Η ειδική μέθοδος για την εισαγωγή του διπλής αλυσίδας RNA μέσα στους
οργανισμούς πρέπει να φτιαχθεί για τον οργανισμό. Για παράδειγμα διπλής
αλυσίδας RNAs μεγαλύτερα των 30 νουκλεοτιδίων δεν ενεργοποιεί το RNAi
μονοπάτι στους ανθρώπους αλλά αντίθετα επάγει την απόκριση της ιντερφερόνης.
Η ιντερφερόνη, μια μικρή πρωτεΐνη η οποία βοηθά τα κύτταρα να αντισταθούν σε
ιϊκή μόλυνση, ενεργοποιεί την 2΄-5΄oligoadenylate synthase, η οποία στην
συνέχεια μετατρέπει το ΑΤΡ σε 2΄-5΄Α oligonucleotides. Στην συνέχεια τα 2΄-5΄Α
oligonucleotides ερεθίζουν με την σειρά τους την RNaseI για να κόψουν διάφορα
είδη RNAs μεταξύ αυτών και τα ριβοσωμικά RNAs. Η κοπή αυτή προκαλεί την μη-
ειδική αναστολή της μετάφρασης. Μία μέθοδος για την αποφυγή του προβλήματος
των μακριών διπλής αλυσίδας RNA είναι η κατασκευή πλασμιδίου που το σύστημα
έκφρασης να στηρίζεται στην χρήση του υποκινητή της RNA πολυμεράσης-III η
οποία είναι ειδική για την παραγωγή μικρών RNAs τα οποία δεν ενεργοποιούν την
απόκριση της ιντερφερόνης.Στα λίγα χρόνια που πέρασαν η RNAi τεχνολογία
βοήθησε στον καθορισμό της λειτουργίας γονιδίων των οποίων η λειτουργία ήταν
άγνωστη.
Γενετική ανάλυση έδειξε ότι 10 ή περισσότερα γονίδια πιθανόν να
χρειάζονται για την σύνθεση των πρωτεϊνών οι οποίες παρίστανται στο siRNA
μονοπάτι. Όμως, τα ευκαρυωτικά σπανίως αντιμετωπίζουν υψηλές συγκεντρώσεις
διπλής αλυσίδας RNA με παρόμοιες αλληλουχίες με αυτές των γονιδίων τους που
κωδικοποιούν για πρωτεΐνες. Κάτω από φυσιολογικές συνθήκες. Ετσι προκύπτει η
ερώτηση γιατί το μονοπάτι του siRNA είναι συντηρημένο στα ευκαρυωτικά τόσο
διαφορετικών ειδών όπως τα φυτά, οι μύγες, τα σκουλήκια και οι άνθρωποι.
Ενας πιθανόν λόγος είναι διότι αυτό βοηθά για την προστασία των
οργανισμών ενάντια σε εισβολή νουκλεϊκών οξέων. Στους σκώληκες ελαττώματα
στο RNAi μονοπάτι προκαλεί την ενεργοποίηση των τρανσποζονίων (transposons).
Το RNAi μονοπάτι φαίνεται ενεργητικά να καταστέλει την διάδοση των
τρανσποζονίων μέσα στο γένωμα εμποδίζοντας έτσι την αφύσικη παραγωγή
γονιδιακών προϊόντων ή την αφύσικη γονιδιακή έκφραση. Το πρόδρομο διπλής

10
αλυσίδας RNA σε siRNA φαίνεται να καθοδηγείται από τις μεταγραφικά τελικές
ανάστροφες επαναλήψεις των τρανσποζονίων. Το RNAi μονοπάτι φαίνεται επίσης
να προσφέρει ανοσία ενάντια σε ιούς οι οποίοι παράγουν διπλής αλυσίδας RNΑ σε
κάποια περίοδο του κύκλου ζωής των. Το Dicer αναγνωρίζει αυτό το διπλής
αλυσίδας ιϊκό RNA και το κόβει για να δημιουργηθεί siRNA το οποίο ενσωματώνεται
σε siRISC, το οποίο στην συνέχεια καταβολίζει το ιϊκό mRNA σημαντικό για την
επιτυχή ιϊκή μόλυνση.
ΣΥΣΤΗΜΙΚΟ RNAi
Στα φυτά και τους νηματώδεις σκώληκες, το RNAi μπορεί να έχει
συστημική επίπτωση στην έκφραση των γονιδίων, έτσι ώστε το γονιδιακό knockout
να επεκτείνεται σε όλον τον οργανισμό. Η βάση του παρόντος φαινομένου
Εικόνα 5:. Λειτουργικά στάδια της σίγηση γονιδίων με double-stranded RNA (dsRNA) σε κύτταρα των κατώτερων ζώων. Το σχήμα δείχνει τα βήματα που εμπλέκονται στην τοπική και συστημική γονιδιακή αποσιώπηση. Εξωγενές dsRNA εισάγεται στα κύτταρα, επεξεργάζεται από το Dicer σε μικρά RNA παρεμβολής (siRNA 21 nt) και συναρμολογούνται με την πρωτεΐνη Αργοναύτη στο σύμπλεγμα RISC (RNA-induced silencing complex). Το RISC σύμπλεγμα στοχοποιεί και καταβολίζει τα ειδικά mRNAs στηριζόμενο στην αλληλουχία siRNA. Συστημική RNAi επίδραση επιτυγχάνεται μέσω της παραγωγής νέων dsRNAs από RNA εξαρτώμενη RNA πολυμεράση (RdRP), η οποία χρησιμοποιεί το στόχοποιημνένο RNA σαν υπόστρωμα και τις υβριδοποιημένες siRNA αλυσίδες σαν υποκινητές (primers). Τα γονιδιακά ονόματα με πλάγια γράμματα έχουν εντοπιστεί στη Drosophila melanogaster. Οι πρωτεΐνες των μεταφορών SID-1 και SID-2 έχουν εντοπιστεί σε Caenorhabditis elegans, όπως και το ένζυμο RdRP. Οι μηχανισμοί του περάσματος μπορεί να διαφέρουν μεταξύ των διαφόρων οργανισμών.

11
πιστεύεται ότι βρίσκεται στην παρουσία μιας RNA καθοδηγούμενη RNA
πολυμεράση (RdRP) η οποία είναι σε θέση να αλληλεπιδράσει με το σύμπλεγμα
RISC και να δημιουργηθούν νέα dsRNA με βάση του μερικώς καταβολισμένου
στοχοποιημένου υποστρώματος χρησιμοποιώντας τις υβριδοποιημένες siRNA
αλυσίδες σαν υποκινητές (primers). To συντιθέμενο dsRNA είναι στη συνέχεια
ενεργεί με τα Dicer ένζυμα για την δημιουργία νέων siRNAs (δευτερογενή siRNAs),
λειτουργώντας έτσι ως ένα ενισχυτικό στάδιο. Με τον τρόπο αυτό, άπαξ και το
dsRNA εισαχθεί σε ένα κύτταρο, το αποτέλεσμά του μπορεί να παραμείνει κατά την
διάρκεια της ανάπτυξης. Επιπλέον, το dsRNAs μπορεί να εξαχθεί σε γειτονικά
κύτταρα και, συνεπώς, η γονιδιακή επίδραση “knock-down” να εξαπλωθεί σε όλον
τον οργανισμό (Εικόνα 5)
miRNA (MicroRNA)
Το miRNA μονοπάτι μπλοκάρει την μετάφραση του mRNA ή προκαλεί τον καταβολισμό του mRNA
Μία πιθανή εξήγηση για την συντήρηση του siRNA μονοπατιού είναι ότι
πολλά από τα ένζυμα και οι πρωτεϊνικοί παράγοντες που παρευρίσκονται σε αυτό
το μονοπάτι είναι παρόμοιοι δομικά ή λειτουργικά με τα ένζυμα και τους
πρωτεϊνικούς παράγοντες οι οποίοι παρευρίσκονται σε ρυθμιστικά μονοπάτια, τα
οποία χρησιμοποιούν ενδογενοί ατελούς φουρκέτας δομών RNA τα οποία
ονομάζονται micro RNA (miRNA) μόρια για να ρυθμίσουν την μετάφραση του
mRNA. (Eικόνα 6).
Εικόνα 6: miRNAs: Πολυάριθμη ομάδα ενδογενών μικρών μη κωδικοποιών RNAs που ρυθμίζουν αρνητικά την γονιδιακή έκφραση. Παράγονται σαν πρόδρομα τα οποία και επεξεργάζονται σε ώριμα μήκους που κυμαίνεται από 21-23 νουκλεοτίδια.

12
Ο Victor Ambros και οι συνεργάτες του ανακάλυψαν για πρώτη φορά το
miRNA το 1993 καθώς μελετούσαν τα πρώϊμα στάδια ανάπτυξης της λάρβας του C.
Elegans. Κατά την διάρκεια αυτών των μελετών, απομόνωσαν μία μεταλλαγμένη
μορφή, η οποία ονομάστηκε lin-4 (lineage abnormal 4) η οποία δεν κάνει την
μετάπτωση από το πρώτο στο δεύτερο στάδιο της λάρβας. Στην αρχή θεώρησαν
ότι το lin-4 μεταγράφημα μεταφράζεται για να φτιάξει μία πρωτεΐνη η οποία
χρειάζεται για την φυσιολογική ανάπτυξη της λάρβας. Όμως περαιτέρω πειράματα
τους οδήγησαν στο συμπέρασμα το lin-4 μεταγράφημα δεν μεταφράζεται αλλά
λειτουργεί σαν ένα πρόδρομο των 22-νουκλεοτιδίων RNA το οποίο λειτουργεί σαν
αρνητικός ρυθμιστής για την έκφραση δύο γονιδίων που κωδικοποιούν για
πρωτεΐνη, το lin-14 και lin-28.Το διάβασμα των αλληλουχιών έδειξε ότι το lin-4
RNA των 22 νουκλεοτιδίων περιέχει μία ατελή ομολογία σε ειδικές περιοχές της
3΄-αμετάφραστης περιοχής (3΄-UTR) των lin-14 και lin-28 mRNA (εικόνα 7).
Εικόνα 7:.Παραδείγματα προτεινόμενων αντιδράσεων μεταξύ της Caenorhabditis elegans lin-4 microRNA και ενός στόχου mRNA.To 22-νουκλεοτιδίων RNA που προέρχεται από το lineage-4 (lin-4) μεταγράφημα προτείνεται ότι αντιδρά με ζευγάρωμα βάσεων με επτά θέσεις στην 3΄-αμετάφραστη περιοχή (UTR) του lineage-14 (lin-14). H παραπάνω εικόνα δείχνει τις πρώτες δύο από τις επτά θέσεις στο 3΄-UTR του lin-14 mRNA. Το C με κόκκινο χρώμα προβλέπεται να παραγεμίζει την αλληλουχία σε 4 από τις 7 θέσεις των αντιδράσεων με το lin-14. Το C μεταλλάσεται σε U με ταυτόχρονη χάσιμο της λειτουργίας της lin-4 μεταλλαγής. lin-14

13
Το lin-4 γονίδιο παρέμενε μέχρι το 2.000 ένα ενδιαφέρον επιστημονικό
περίεργο όταν ο Gary Ruvkan και οι συνεργάτες του έδειξαν ότι το C. Elegans
απαιτεί ένα δεύτερο 22-νουκλεοτιδίων RNA που αυτή την φορά κωδικοποιείται από
το let-7 (lethal-7) γονίδιο, για να προχωρήσει από το τελευταίο στάδιο της λάρβας
στο ενήλικο στάδιο. Περαιτέρω μελέτη έδειξε ότι το 22-νουκλεοτιδίων let-7 RNA
αναστέλλει την έκφραση του lin-41 και lin-42 δύο γονιδίων που κωδικοποιούν για
πρωτεΐνες. Επιπλέον, το 22-νουκλεοτιδίων let-7 RNA περιέχει ατελείς ομολογίες με
ειδικές περιοχές στα 3’-UTRs των lin-41 και lin-42 mRNA. O Ruvkin και οι
συνεργάτες του απέδειξαν στην συνέχεια ότι το let-7 είναι συντηρημένο και
εκφράζεται σε μία μεγάλη ποικιλία ζώων συμπεριλαμβανομένων των πτηνών, των
ποντικών, των κοτόπουλων και των ανθρώπων. Με αυτήν την απόδειξη, τα 22-
νουκλεοτιδίων RNAs τοποθετήθηκαν στο κέντρο της μοριακής Βιολογίας σαν ένα
σπουδαίο και ακραίο ερευνητικό πρόβλημα. Εξ’ αιτίας της κοινής λειτουργίας τους
στην ρύθμιση του χρόνου και της ανάπτυξης της μετάπτωσης, τα lin-4 και lin-7
RNAs , αρχικά ονομάστηκαν μικρά παροδικά RNAs, αλλά αυτό το όνομα αργότερα
άλλαξε σε microRNAs ή στο συντετμημένο miRNA.
Στηριζόμενοι σε πειραματικά ή υπολογιστικά αποτελέσματα , το ανθρώπινο
γένομα φαίνεται να διαθέτει περίπου 200-250 microRNA γονίδια, ενώ τα γενώματα
των Δροσόφιλα (Drosophila) και νηματέλμινθα σκώληκα (C. Elegans) διαθέτουν
περίπου τον μισό αριθμό. Ετσι, τα γονίδια των miRNA συνιστούν περίπου το 1%
των γνωστών γονιδίων σε έναν οργανισμό και είναι παρόντα με τον ίδιο περίπου
αριθμό σαν γονίδια τα οποία κωδικοποιούν για μεταγραφικούς ενεργοποιητές.
Σήμερα, έχει γίνει πρόβλεψη για 3’-UTR περιοχές στα μεταγραφήματα των
ανθρώπινων γονιδίων για 218 miRNAs των θηλαστικών. Περίπου 2.000 ανθρώπινα
γονίδια με περιοχές στόχους για τα miRNAs έχουν χαρακτηριστεί
(http://www.microrna.org).
Τα γονίδια για την πλειονότητα των miRNAs είναι παρόντα σε μοναχικά
στοιχεία στο γένομα, τοποθετημένα σε περιοχές που άλλοτε θεωρούνταν να μην
περιέχουν γονίδια. Όμως, μερικά miRNA γονίδια μπορεί να βρεθούν ανάμεσα σε
ιντρόνια από γονίδια που κωδικοποιούν για πρωτεΐνη ή σε πολυσιστρονικές
περιοχές (π.χ η δέσμη γονιδίων mir-17–92). Aν και η RNA πολυμεράση ΙΙ φαίνεται
να είναι υπεύθυνη για την σύνθεση του RNA το οποίο επεξεργάζεται για να
δημιουργήσει miRNA, η RNA πολυμεράση ΙΙΙ μπορεί επίσης να συνεισφέρει στην
miRNA σύνθεση.
Το τρέχων μοντέλο για την δημιουργία του miRNA και την λειτουργία του
φαίνεται στην εικόνα 8. Το πρώτο στάδιο είναι το κόψιμο του πρωταρχικού RNA
μεταγραφήματος (pri-miRNA) , το οποίο ελευθερώνει ένα ολιγονουκλεοτίδιο

14
μήκους 60-70 με ενδιάμεση δομή στεφάνης-θηλειάς (stem-loop), που
υποδεικνύεται σαν miRNA πρόδρομο ή pre-miRNA. Αυτή η επεξεργασία
πραγματοποιείται από την ενδονουκλεάση RNase III σε ένα σύμπλεγμα., το οποίο
περιέχει πρωτεΐνες οι οποίες ονομάζονται Drosha και Pasha. To pre-miRNA
ενεργητικά μεταφέρεται από τον πυρήνα στο κυτταρόπλασμα από την RAN-GTP
και τον υποδοχέα εξόδου Exportin-5. Απαξ και μεταφερθεί στο κυτταρόπλασμα, το
pre-miRNA κόβεται από την Dicer. Στην Δροσόφιλα, αυτό το κόψιμο καταλύεται
από την Dicer 1 (DCR1), η οποία εκτελεί την ίδια λειτουργία σαν DCR2 στο
Εικόνα 8: Το μονοπάτι του miRNA. Η συγκρότηση του miRISC αρχίζει στον πυρήνα , όπου το γονίδιο που κωδικοποιεί για το miRNA μεταγράφεται σε πρωταρχικό miRNA (Pri-mRNA) μεταγράφημα. Το Pri-mRNA επεξεργάζεται σε ένα πρόδρομο miRNA (Pre-mRNA).από το Drosha (πράσινο) και Pasha (κόκκινο) στο Microprocessor σύμπλεγμα και στην συνέχεια εξέρχεται στο κυτταρόπλασμα από την Exportin-5 (Μωβ) για ωρίμανση σε miRNA από την κυτταροπλασματική Dicer,DCR1 (πράσινη). Στην συνέχεια το miRNA στο miRNA-DCR1 σύμπλεγμα συγκροτείται σε miRISC. Το ερωτηματικό δείχνει αβεβαιότητες περί της σύστασης ή της λειτουργίες ενός πρωτεϊνικού συμπλέγματος.
μονοπάτι του siRNA. Το διπλής αλυσίδας RNA θραύσμα είναι πιθανόν ξετυλιγμένο
από το miRNA RISC σύμπλεγμα φορτώματος (miRLC: micro RNA Loading
Complex) και το απλής αλυσίδας οδηγό RNA (Guide) τελικά ενσωματώνονται σε
miRISC σύμπλεγμα το οποίο εμπεριέχει μία πρωτεΐνη που ονομάζεται Αργοναύτης
(AGO: Argonaute). Το miRISC σύμπλεγμα εμφανίζεται να έχει τουλάχιστον 2
τρόπους δράσης. Όταν υπάρχει πλήρης ομολογία μεταξύ του οδηγού RNA και της
περιοχής του mRNA στόχου, το οδηγό RNA κατευθύνει την πρωτεΐνη AGO να
κόψει το mRNA στόχο (Εικόνα 9Α).Όταν όμως υπάρχει ατελής ομολογία, το οδηγό
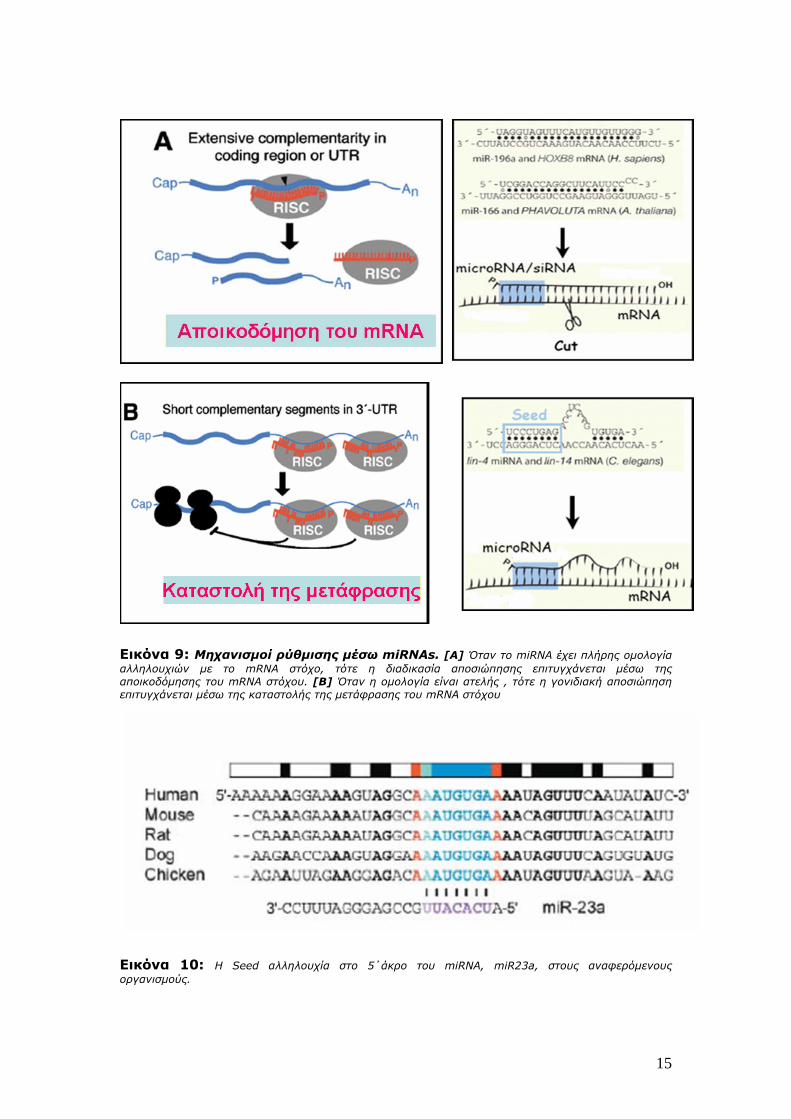
15
Εικόνα 9: Μηχανισμοί ρύθμισης μέσω miRNAs. [A] Όταν το miRNA έχει πλήρης ομολογία αλληλουχιών με το mRNA στόχο, τότε η διαδικασία αποσιώπησης επιτυγχάνεται μέσω της αποικοδόμησης του mRNA στόχου. [Β] Όταν η ομολογία είναι ατελής , τότε η γονιδιακή αποσιώπηση επιτυγχάνεται μέσω της καταστολής της μετάφρασης του mRNA στόχου
Εικόνα 10: Η Seed αλληλουχία στο 5΄άκρο του miRNA, miR23a, στους αναφερόμενους οργανισμούς.
16
mRNA συνδέεται στο 3΄-UTR του mRNA στόχου και μπλοκάρει την μετάφραση
(Εικόνα 9Β). Αν και ακόμη δεν είναι γνωστό το πώς δηλαδή η αντίδραση με το 3΄-
UTR μπλοκάρει την μετάφραση, η αναστολή λαμβάνει μέρος μετά την σύνδεση
των ριβοσωματίων με το mRNA με σκοπό την δημιουργία των πολυριβοσωμάτων.
Στην αναγνώριση του mRNA στόχου συνεισφέρει μία αλληλουχία, η Seed
αλληλουχία, που συνιστά μία αλληλουχία 8 nt στο 5’ άκρο του miRNA με ακριβές
ταίριασμα στο mRNA (Εικόνα 10).
Κάτω από αυτές τις συνθήκες, λίγες είναι οι διαφορές που χαρακτηρίζουν
τα miRNA και siRNA μονοπάτια (Εικόνα 11).
Εικόνα 11: Διαφορές μεταξύ siRNAs και mRNAs.
ΑΛΛΟΙ ΤΡΟΠΟΙ ΓΟΝΙΔΙΑΚΗΣ ΑΠΟΣΙΩΠΗΣΗΣ esiRNA
To esiRNA ή τα Endoribonuclease-προετοιμασμένα siRNAs συνιστούν ένα
μίγμα από siRNAs ολιγονουκλεοτίδια τα οποία προέρχονται από κόψιμο μακριών
διπλής αλυσίδας RNA (dsRNA) με μία ενδοριβονουκλεάση τέτοια όπως η
Escherichia coli RNase III ή dicer.
Ενας εναλλακτικός τρόπος στην χρήση των χημικά συντιθέμενων siRNAs
για την RNA παρεμβολή (RNAi) είναι η ενζυματική επώαση μακριών διπλής
αλυσίδας RNAs in vitro. Σε αυτήν την περίπτωση ένα cDNA υπόστρωμα
επαυξάνεται με PCR και ενσωματώνεται σε 2 υποκινητικές αλληλουχίες του

17
Βακτηριοφάγου. Στην συνέχεια η RNA πολυμεράση χρησιμοποιείται για την
παραγωγή μακριών διπλής αλυσίδας RNA το οποίο είναι ομόλογο του cDNA
γονιδίου στόχου. Στην συνέχεια αυτό το RNA επωάζεται με RNase III της E. coli
για την δημιουργία μικρών επικαλλυπτόμενων θραυσμάτων siRNAs με μήκος
μεταξύ 18-25 ζευγών βάσεων. Αυτό μίγμα από διπλής αλυσίδας RNAs είναι
παρόμοιο με το μίγμα που δημιουργείται από το κόψιμο της Dicer in vivo και έτσι
ονομάζεται ενδοριβονουκλεάσης-προετοιμασία siRNA ή μικρό esiRNA. Το esiRNA
είναι ετερογενές μίγμα από siRNAs τα οποία όλα στοχοποιούν την ίδια mRNA
αλληλουχία. Αυτά τα πολλαπλά αποσιωπητικά ερεθίσματα οδηγούν σε ειδική και
πολύ αποδοτική αποσιώπηση του γονιδίου (Εικόνα 12) (Yang, D., et al. 2002,
Kittler, R., et al. 2004).
Ιοειδή (Viroid)
Τα ϊοειδή είναι φυτικά παθογόνα τα οποία συνίστανται από κοντές
αλληλουχίες (ολίγων εκατοντάδων νουκλεοβάσεις) υψηλής συμπληρωματικότητας,
κυκλικών, απλής αλυσίδας RNA χωρίς πρωτεϊνική κάψα η οποία είναι τυπική για
τους ϊούς (Εικόνα 8). Το μικρότερο που ανακαλύφθηκε είναι ένα 220
Εικόνα :
Εικόνα 12: Σχηματική αναπαράσταση της παρασκευής των esiRNA. Τα στάδια που ακολουθούνται για την δημιουργία μιας ομάδας λειτουργικών siRNAs παρουσιάζονται.
Ιοειδή (Viroid)
Τα ϊοειδή είναι φυτικά παθογόνα τα οποία συνίστανται από κοντές
αλληλουχίες (ολίγων εκατοντάδων νουκλεοβάσεις) υψηλής συμπληρωματικότητας,
κυκλικών, απλής αλυσίδας RNA χωρίς πρωτεϊνική κάψα η οποία είναι τυπική για
τους ϊούς (Εικόνα 13). Το μικρότερο που ανακαλύφθηκε είναι ένα 220
νουκλεοβάσεων scRNA (small cytoplasmic RNA) που συσχετίζεται με το κίτρινο
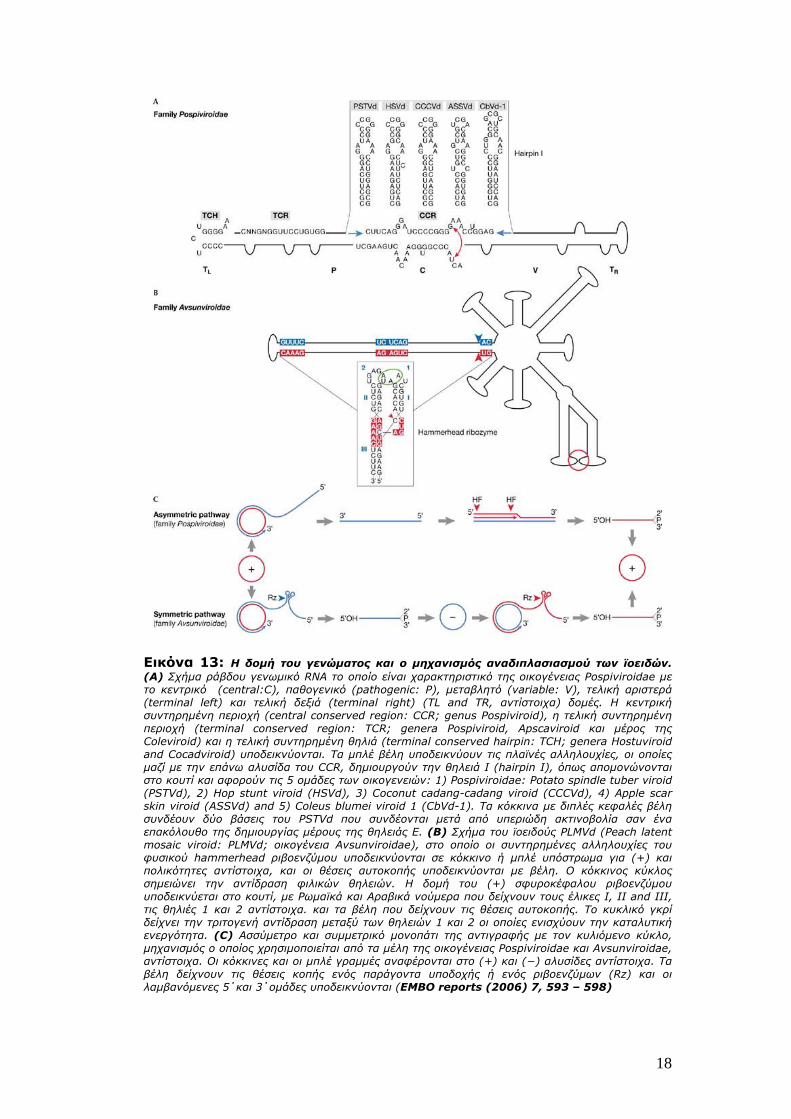
18
Εικόνα 13: Η δομή του γενώματος και ο μηχανισμός αναδιπλασιασμού των ϊοειδών. (Α) Σχήμα ράβδου γενωμικό RNA το οποίο είναι χαρακτηριστικό της οικογένειας Pospiviroidae με το κεντρικό (central:C), παθογενικό (pathogenic: P), μεταβλητό (variable: V), τελική αριστερά (terminal left) και τελική δεξιά (terminal right) (TL and TR, αντίστοιχα) δομές. Η κεντρική συντηρημένη περιοχή (central conserved region: CCR; genus Pospiviroid), η τελική συντηρημένη περιοχή (terminal conserved region: TCR; genera Pospiviroid, Apscaviroid και μέρος της Coleviroid) και η τελική συντηρημένη θηλιά (terminal conserved hairpin: TCH; genera Hostuviroid and Cocadviroid) υποδεικνύονται. Τα μπλέ βέλη υποδεικνύουν τις πλαϊνές αλληλουχίες, οι οποίες μαζί με την επάνω αλυσίδα του CCR, δημιουργούν την θηλειά Ι (hairpin I), όπως απομονώνονται στο κουτί και αφορούν τις 5 ομάδες των οικογενειών: 1) Pospiviroidae: Potato spindle tuber viroid (PSTVd), 2) Hop stunt viroid (HSVd), 3) Coconut cadang-cadang viroid (CCCVd), 4) Apple scar skin viroid (ASSVd) and 5) Coleus blumei viroid 1 (CbVd-1). Τα κόκκινα με διπλές κεφαλές βέλη συνδέουν δύο βάσεις του PSTVd που συνδέονται μετά από υπεριώδη ακτινοβολία σαν ένα επακόλουθο της δημιουργίας μέρους της θηλειάς Ε. (B) Σχήμα του ϊοειδούς PLMVd (Peach latent mosaic viroid: PLMVd; οικογένεια Avsunviroidae), στο οποίο οι συντηρημένες αλληλουχίες του φυσικού hammerhead ριβοενζύμου υποδεικνύονται σε κόκκινο ή μπλέ υπόστρωμα για (+) και πολικότητες αντίστοιχα, και οι θέσεις αυτοκοπής υποδεικνύονται με βέλη. Ο κόκκινος κύκλος σημειώνει την αντίδραση φιλικών θηλειών. Η δομή του (+) σφυροκέφαλου ριβοενζύμου υποδεικνύεται στο κουτί, με Ρωμαϊκά και Αραβικά νούμερα που δείχνουν τους έλικες I, II and III, τις θηλιές 1 και 2 αντίστοιχα. και τα βέλη που δείχνουν τις θέσεις αυτοκοπής. Το κυκλικό γκρί δείχνει την τριτογενή αντίδραση μεταξύ των θηλειών 1 και 2 οι οποίες ενισχύουν την καταλυτική ενεργότητα. (C) Ασσύμετρο και συμμετρικό μονοπάτι της αντιγραφής με τον κυλιόμενο κύκλο, μηχανισμός ο οποίος χρησιμοποιείται από τα μέλη της οικογένειας Pospiviroidae και Avsunviroidae, αντίστοιχα. Οι κόκκινες και οι μπλέ γραμμές αναφέρονται στο (+) και (−) αλυσίδες αντίστοιχα. Τα βέλη δείχνουν τις θέσεις κοπής ενός παράγοντα υποδοχής ή ενός ριβοενζύμων (Rz) και οι λαμβανόμενες 5΄και 3΄ομάδες υποδεικνύονται (EMBO reports (2006) 7, 593 – 598)

19
ρύζι και ονομάζεται mottle sobemovirus (RYMV). Σε σύγκριση, το γένομα των
μικρότερων γνωστών ϊών που είναι σε θέση να προκαλέσουν μία μόλυνση από
μόνοι τους, είναι μεγέθους 2.000 kb. Το ανθρώπινο παθογόνο Ηπατίτιδα-D
(hepatitis D) είναι παρόμοιο με τα ϊοειδή.
Τα ϊοειδή ανακαλύφθηκαν και δόθηκε το όνομά τους από τον Theodor Otto
Diener, έναν φυτοπαθολόγο στο Agricultural Research Service στο Maryland των
ΗΠΑ, το 1971. Τα ϊοειδή RNA δεν κωδικοποιούν για καμία πρωτεΐνη. Ο μηχανισμός
της αντιγραφής περιλαμβάνει την RNA πολυμεράση ΙΙ, ένα ένζυμο που
φυσιολογικά συσχετίζεται με την σύνθεση του mRNA από το DNA, το οποίο αντί
αυτού καταλύει την σύνθεση με τον τρόπο του κυλιόμενου κύκλου (Rolling
Circles) νέων RNAs χρησιμοποιώντας τα ϊοειδή RNAs σαν υπόστρωμα. Μερικά από
τα ϊοειδή είναι ριβοένζυμα , έχοντας καταλυτικές ιδιότητες οι οποίες επιτρέπουν το
κόψιμο του εαυτού τους και την ένωση θραυσμάτων του γενώματος.
Υπάρχει ένα μεγάλο μπλέξιμο στο πως τα ϊοειδή είναι ικανά να προκαλούν
συμπτώματα στα φυτά χωρίς να κωδικοποιούν για καμιά πρωτεΐνη με τις
αλληλουχίες τους. Σήμερα υπάρχουν ενδείξεις ότι η γονιδιακή αποσιώπηση
εντάσσεται στην διαδικασία. Οι ενδείξεις αυτές δείχνουν ότι όταν τα ϊοειδή
διπλασιάζονται μέσω ενός διπλής αλυσίδας ενδιάμεσου RNA, στοχοποιούνται από
ένα Dicer ένζυμο και κόβεται σε siRNAs τα οποία στην συνέχεια φορτώνονται στα
RNA επαγώμενα αποσιωπητικά συμπλέγματα. Τα ϊοειδή siRNAs στην
πραγματικότητα περιέχουν αλληλουχίες ικανές για συμπληρωματικό ζευγάρωμα
βάσεων με των φυτών τα mRNAs και την επαγωγή του καταβολισμού τους ή της
αναστολής της μετάφρασής τους. Αυτά συνιστούν και το αίτιο για τα κλασικά
συμπτώματα των ϊοειδών.
Mεταγραφική αποσιώπηση (Transcriptional
silencing) Συστατικά του μονοπατιού του RNAi χρησιμοποιούνται επίσης σε πολλά
ευκαρυωτικά κύτταρα για τη διατήρηση της οργάνωσης και της δομής των
γονιδιωμάτων τους. Η τροποποίηση των ιστονών και η η συσχετιζόμενη επαγωγή
της δημιουργίας ετεροχρωματίνης χρησιμεύει για την αρνητική ρύθμιση των
γονιδίων προ-μεταγραφικά, η διαδικασία αυτή αναφέρεται ως RNA-επαγόμενη
μεταγραφική σίγηση (RNA-Induced Transcriptional Silencing: RITS), και
πραγματοποιείται από ένα σύμπλεγμα πρωτεϊνών που ονομάζεται σύνθετη RITS
(Eικόνα 14). Σε ζυμομύκητες αυτό το σύμπλεγμα περιλαμβάνει την πρωτεΐνη
Αργοναύτη (argonaute),την πρωτεΐνη Chp1 (chromodomain), και μια πρωτεΐνη
που ονομάζεται Tas3 άγνωστης λειτουργίας. Κατά συνέπεια, η εισαγωγή και
διάδοση των ετεροχρωματινικών περιοχών απαιτεί την πρωτεΐνη Αργοναύτη και

20
RdRP (RNA directed RNA polymerase) πρωτεϊνών. Πράγματι, η διαγραφή αυτών
των γονιδίων του ζυμομύκητα S. pombe διαταράσσει την μεθυλίωση των ιστονών
και τον σχηματισμό του κεντρομεριδίου, με αποτέλεσμα η στασιμότητα της
ανάφασης κατά την κυτταρική διαίρεση.
Εικόνα 14: Το ένζυμο Dicer φροντίζει έτσι ώστε τα διπλής αλυσίδας RNAs, να δημιουργούν μικρά RNAs παρεμβολής (siRNAs) ή microRNA. Αυτά τα RNAs ενσωματώνονται μέσα στο σύμπλεγμα RISC (RNA-induced silencing complex) το οποίο στοχοποιεί το mRNA για να εμποδίσει την μετάφραση[WIKIPEDIA]
Σε ορισμένες περιπτώσεις, παρόμοιες διαδικασίες που συνδέονται με την
τροποποίηση των ιστονών έχει παρατηρηθεί ότι ρυθμίζουν θετικά την μεταγραφή
γονιδίων.
Εφαρμογές της τεχνολογίας siRNA σε ανθρώπινα κύτταρα (HeLa cells), φυτά (plants), σκώληκα (worms) και Δροσόφιλα (Drosophila)
Σε πολλούς οργανισμούς, η εισαγωγή διπλής αλυσίδας RNA έχει αποδειχθεί
σαν ένα ισχυρό όπλο για την καταστολή της γονιδιακής έκφρασης δια μέσου ενός
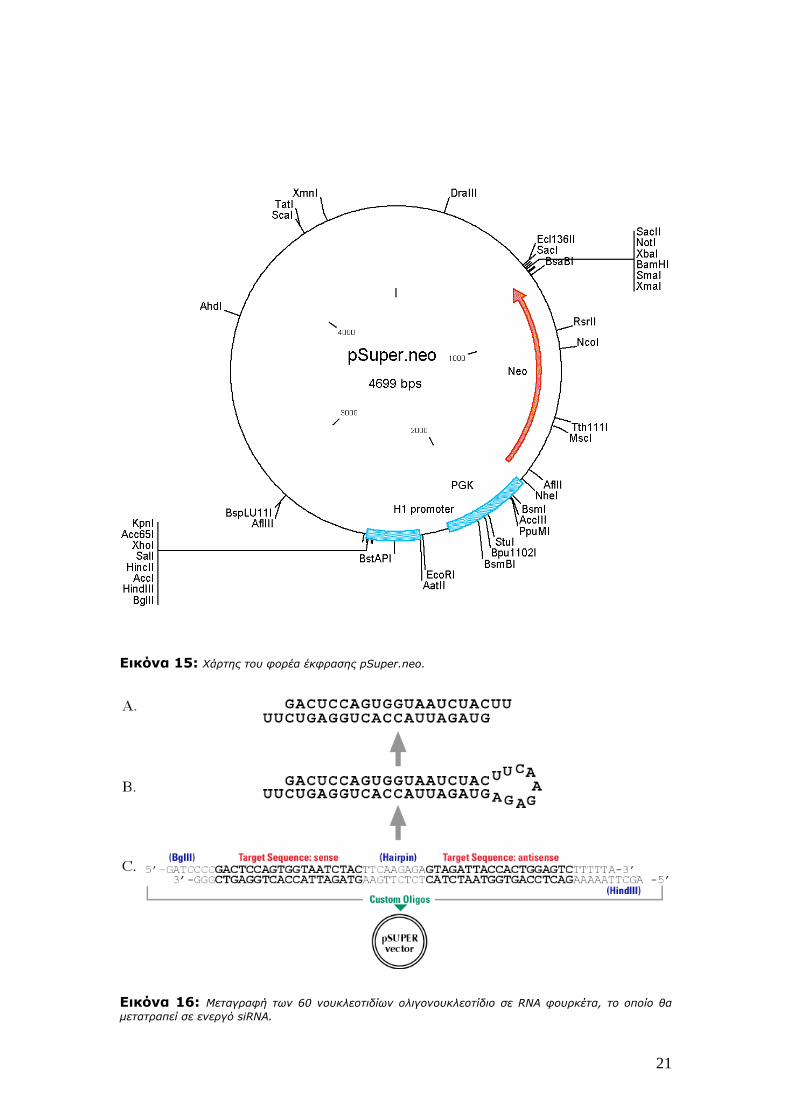
21
Εικόνα 15: Χάρτης του φορέα έκφρασης pSuper.neo.
Εικόνα 16: Μεταγραφή των 60 νουκλεοτιδίων ολιγονουκλεοτίδιο σε RNA φουρκέτα, το οποίο θα μετατραπεί σε ενεργό siRNA.
22
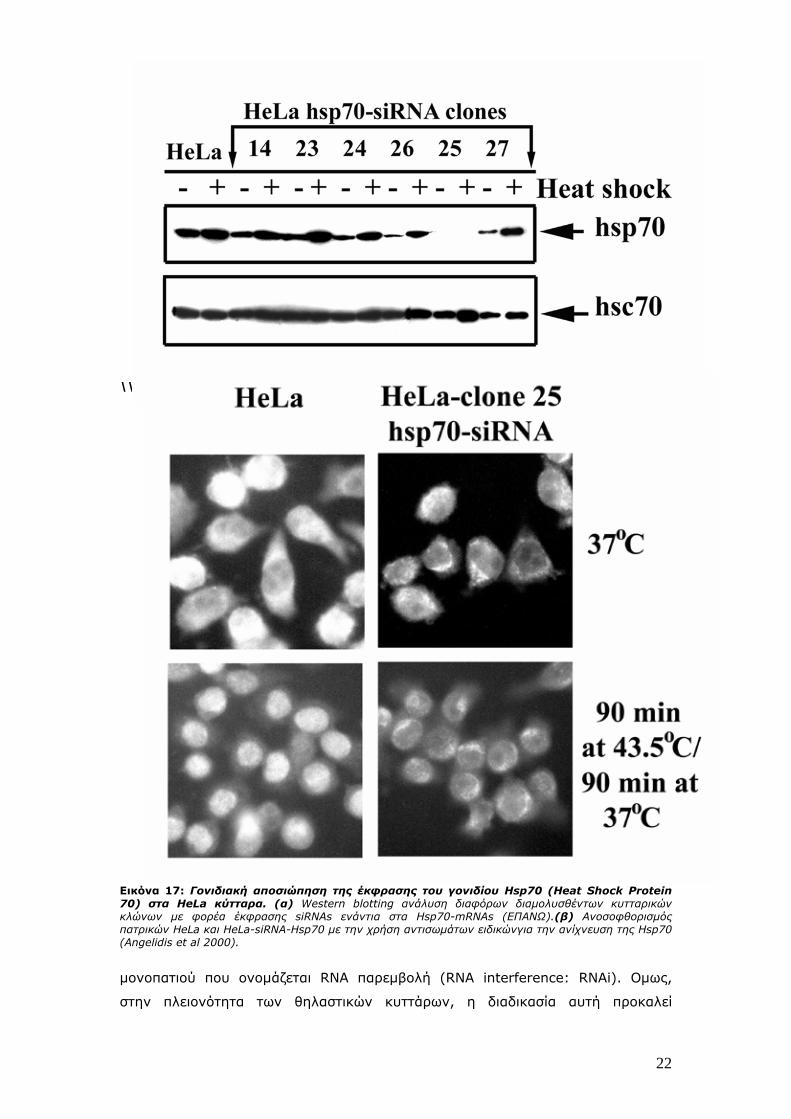
\\\\\\\\\\\\ Εικόνα 17: Γονιδιακή αποσιώπηση της έκφρασης του γονιδίου Hsp70 (Heat Shock Protein 70) στα HeLa κύτταρα. (α) Western blotting ανάλυση διαφόρων διαμολυσθέντων κυτταρικών κλώνων με φορέα έκφρασης siRNAs ενάντια στα Hsp70-mRNAs (ΕΠΑΝΩ).(β) Ανοσοφθορισμός πατρικών HeLa και HeLa-siRNA-Hsp70 με την χρήση αντισωμάτων ειδικώνγια την ανίχνευση της Hsp70 (Angelidis et al 2000).
μονοπατιού που ονομάζεται RNA παρεμβολή (RNA interference: RNAi). Oμως,
στην πλειονότητα των θηλαστικών κυττάρων, η διαδικασία αυτή προκαλεί
23
τοξικότητα. Αυτή η μη-ειδική επίδραση μπορεί να αντιμετωπιστεί με την χρήση
συνθετικών μικρών [21-22 νουκλεοτιδίων] παρεμβολικών RNAs (siRNAs), τα
οποία μπορούν να προκαλέσουν ισχυρή και ειδική καταστολή της γονιδιακής
έκφρασης. Όμως, αυτή η καταστολή είναι παροδική και για αυτό περιορίζει αρκετά
τις εφαρμογές. Για την υπερπήδηση αυτού του περιορισμού, χρησιμοποιήθηκαν
ειδικές υποκινητικές περιοχές (promoters) για την χρήση RNA πολυμερασών
ειδικών για μικρά μεταγραφήματα (RNAs). Μεταξύ αυτών και ο RNA πολυμεράσης
ΙΙΙ υποκινητής του γονιδίου του H1-RNA (το RNA συστατικό της ανθρώπινης
RNase : ενώ είναι RNA δρά σαν πρωτεϊνικό ένζυμο, κόβοντας έξτρα το πρόδρομο
μόριο των t-RNAs) Για παράδειγμα ο pSUPER φορέας περιέχει τον υποκινητή
αυτόν και ταυτόχρονα φέρει ένα σήμα λήξης από 5 συνεχόμενες θυμιδίνες (Τ5).
Το κόψιμο του μεταγραφήματος στην θέση λήξης γίνεται μετά από την δεύτερη
Ουριδίνη (U) δημιουργώντας έτσι ένα μεταγράφημα που μοιάζουν με τα συνθετικά
siRNAs τα οποία περιέχουν 2 προεξέχοντα T ή U νουκλεοτίδια (Εικόνες 15, 16). Οι
φορείς έκφρασης αυτού ή παρόμοιων τύπων προκαλούν αποδοτική και ειδική
καταστολή της γονιδιακής έκφρασης, καταλήγοντας σε λειτουργική αδράνεια
γονιδίων που έχουν στοχοποιηθεί. Με την σταθερή έκφραση των siRNAs
χρησιμοποιόντας αυτούς τους φορείς (Σταθερά διαμολυσμένα κύτταρα: stable
transfected cells), προκαλούμε σταθερή καταστολή της γονιδιακής έκφρασης,
επιτρέποντας έτσι την ανάλυση του χαμένου λειτουργικά φαινοτύπου ο οποίος
αναπτύσσεται σε μεγάλους χρονικά περιόδους. Παράδειγμα γονιδιακής
αποσιώπησης αποτελεί η αποσιώπηση του γονιδίου Hsp70 στα ανθρώπινα HeLa
κύτταρα (Εικόνα 17).
Εφαρμογές του RNAi στην θεραπευτική
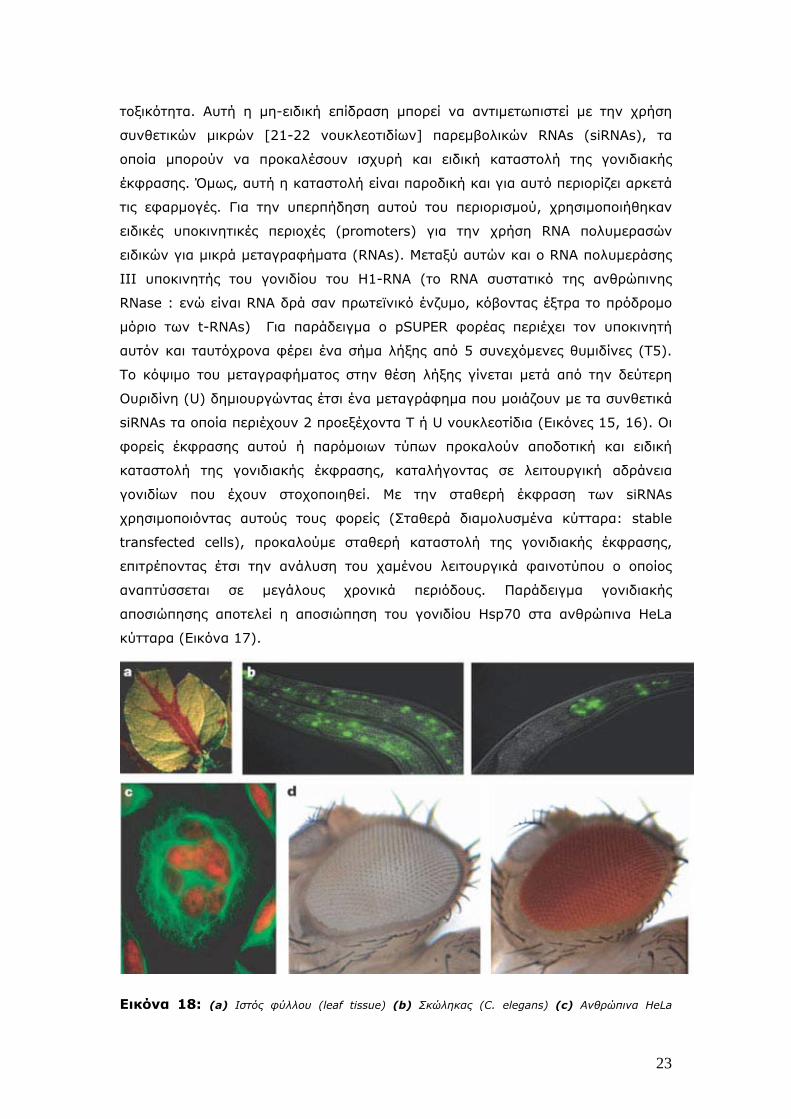
Εικόνα 18: (a) Ιστός φύλλου (leaf tissue) (b) Σκώληκας (C. elegans) (c) Ανθρώπινα HeLa

24
κύτταρα (HeLa cells) (d) Ενήλικη Δροσόφιλα (Adult Drosophila).
Στα φυτά η αποσιώπηση μπορεί να ερεθισθεί, για παράδειγμα από
τροποποιημένους RNA ιούς. Στους σκώληκες, η αποσιώπηση μπορεί να
ενεργοποιηθεί με ένεση ή με διατροφή τους με βακτηρίδια που φέρουν dsRNA. Και
στα δύο συστήματα, η αποσιώπηση είναι συστημική και εξαπλώνεται σε όλον τον
οργανισμό. Ετσι, ένα αποσιωπητικό σήμα μετακινείται από τις φλέβες (vein) στον
ιστό του φύλλου (Εικόνα 18-a). Η αλλαγή χρώματος οφείλεται στην καταστροφή
του GFP μεταγραφήματος (πράσινο) και στην εμφάνιση της χλωροφύλλης
(κόκκινο) καθώς εξαφανίζεται το GFP. Το ίδιο φαινόμενο παρουσιάζεται και σε
διαγονιδιακούς σκώληκες C. Elegance που έχουν κατασκευασθεί να εκφράζουν την
GFP στους πυρήνες των κυττάρων (Εικόνα 18, b-αριστερά). Mετά από μόλυνσή
τους με dsRNA ειδικό για την GFP, ο ανοσοφθορισμός της GFP αρχίζει να
εξαφανίζεται (Εικόνα 18, b-δεξιά). Σε HeLa κύτταρα τα οποία έχουν μολυνθεί με
ORC6 siRNA, εάν βάψουμε για tubulin (πράσινο) και DNA (κόκκινο) τότε θα δούμε
ότι η έλλειψη της ORC6 προκαλεί την συσσώρευση πολυπύρηνων κυττάρων
(Εικόνα 18 c). Εάν σε Δροσόφιλα με άσπρα μάτια χρησιμοποιήσουμε dsRNA ειδικό
για το άσπρο γονίδιο (Εικόνα 12 d-αριστερά).τότε μπορούμε να πάρουμε τα
κόκκινα μάτια (Εικόνα 18 d-δεξιά).
Η χρήση της τεχνολογίας του RNAi αφορά στην αντιμετώπιση των ασθενειών
στις οποίες η σίγηση ενός ή ορισμένων γονιδίων θα μπορούσε να επιβραδύνει ή
ακόμη και να σταματήσει την πρόοδο της ασθένειας στα συγκεκριμένα κύτταρα τα
οποία εκδηλώνουν την ανωμαλία που συνδέεται με την ασθένεια. Πολλοί
Επιστήμονες θεωρούν σήμερα ότι το RNAi συνιστά μία από τη μεγαλύτερη
επανάσταση στη θεραπευτική μετά την χρήση των αντισωμάτων. Η θεραπεία μέσω
του RNAi στηρίζεται στην αποσιώπηση μη βασικών φυσιολογικών γονιδίων των
οποίων η μετάλλαξη προκαλεί τη νόσο. Οι στόχοι της θεραπευτικής χρήσης της
τεχνολογίας του RNAi περιλαμβάνουν ένα ευρύ φάσμα πρωτεϊνών οι οποίες
σχετίζονται με ένα ανάλογο φάσμα νόσων και κυτταρικών διαταραχών μεταξύ των
οποίων ο καρκίνος, οι βακτηριακές λοιμώξεις και οι ασθένειες που οφείλονται σε
ιούς.
1. Καρκίνος: Ο καρκίνος αποτελεί μια πολυγονιδιακή ασθένεια η οποία
χαρακτηρίζεται από έξι βασικές μεταβολές της φυσιολογίας των κυττάρων: (1)
έλλειψη ανταπόκρισης σε αντι-αυξητικά σήματα, (2) κυτταρική αυτάρκεια σε
αυξητικούς παράγοντες, (3) απεριόριστη δυνατότητα πολλαπλασιασμού, (4)
αποφυγή της απόπτωσης, (5) ικανότητα αγγειογένεσης, και (6) ικανότητα
διήθησης και μετάστασης στους ιστούς. Ο στόχος λοιπόν της θεραπευτικής χρήσης
του RNAi είναι να αναστείλει τη μεταγραφή ενός ή πολλών γονιδίων ταυτόχρονα
τα οποία σχετίζονται με τα προαναφερόμενα φαινόμενα. Μια σημαντική

25
παράμετρος που θα πρέπει να ληφθεί υπόψη είναι αυτή της επιλεκτικής στόχευσης
του siRNA στα καρκινικά κύτταρα. Για την επι\εκτική καταστροφή των καρκινικών
κυττάρων χωρίς να υποστούν βλάβη τα φυσιολογικά κύτταρα, θα πρέπει το RNAi
να στοχεύει σε γονίδιο το οποίο εμπλέκεται αποκλειστικά στην ανάπτυξη ή την
επιβίωση του καρκινικού κυττάρου. Το πρόβλημα της στόχευσης των καρκινικών
κυττάρων in vivo αποτελεί τη μεγαλύτερη τροχοπέδη στην εφαρμογή της
τεχνολογίας του RNAi σε θεραπευτικό επίπεδο.
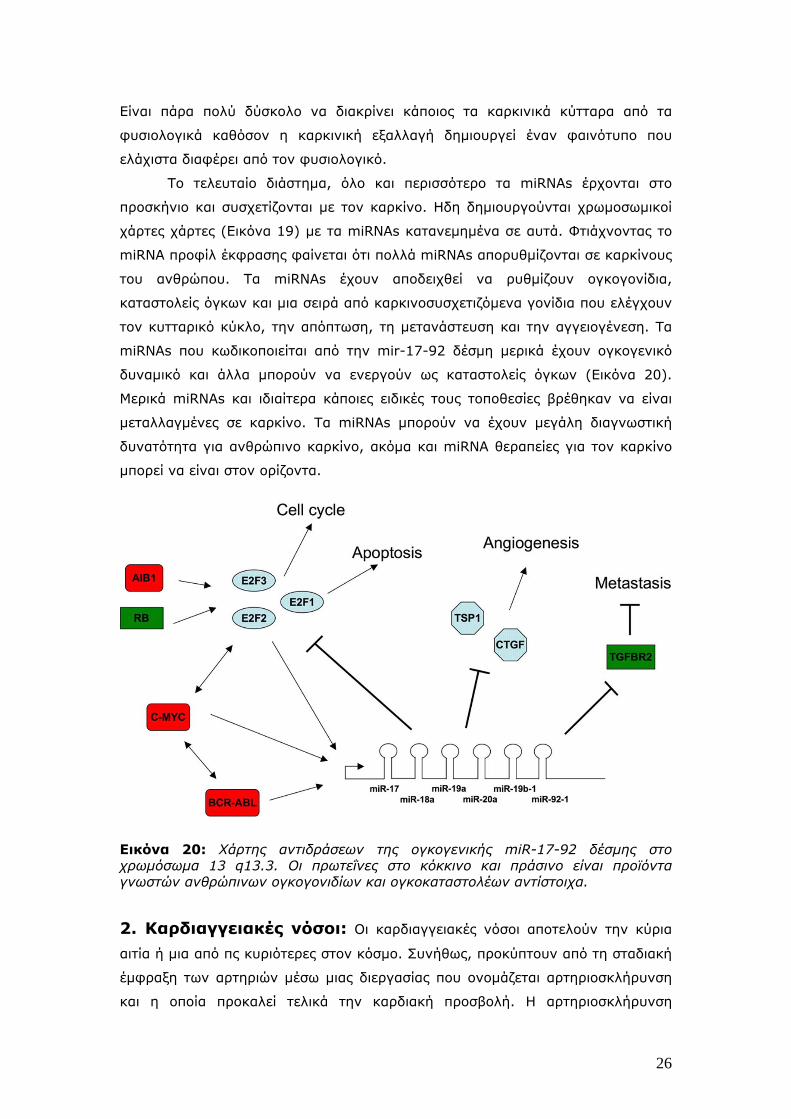
Εικόνα 19: Ενας miRNA καρκινικός χάρτης. Οι χρωμοσωμικές θέσεις των miRNAs εμπλέκονται με τον ανθρώπινο καρκίνο φαίνονται σαν χρωματισμένες κηλίδες. Τα χρώματα αναφέρονται σε ειδικούς τύπους ιστικών όγκων όπως φαίνεται πάνω δεξιά. . Μόνο τα miRNAs, των οποίων τα επίπεδα έκφρασης βρέθηκαν να αλλάζουν σημαντικά σε 2 τουλάχιστον διαφορετικούς όγκους παρουσιάζονται. Τα miRNAs τα οποία ανιχνεύτηκαν σε 4 διαφορετικούς όγκους επίσης παρουσιάζονται με τα ονόματά τους. Η γνώση συγκεντρώθηκε από μελέτες που αφορούσαν πρωτογενείς ανθρώπινους όγκους από ορθό (colorectum), θυροειδή (thyroid), στήθος (breast), πνεύμονα (lung), ήπαρ (liver), πάγκρεας (pancreas), μήτρα (uterus), χρόνια λεμφοκυτταρική λευχαιμία (chronic lymphocytic leukaemia: CLL), γλιοβλάστωμα (glioblastoma) και το νευροβλάστωμα (neuroblastoma).
26
Είναι πάρα πολύ δύσκολο να διακρίνει κάποιος τα καρκινικά κύτταρα από τα
φυσιολογικά καθόσον η καρκινική εξαλλαγή δημιουργεί έναν φαινότυπο που
ελάχιστα διαφέρει από τον φυσιολογικό.
Το τελευταίο διάστημα, όλο και περισσότερο τα miRNAs έρχονται στο
προσκήνιο και συσχετίζονται με τον καρκίνο. Ηδη δημιουργούνται χρωμοσωμικοί
χάρτες χάρτες (Εικόνα 19) με τα miRNAs κατανεμημένα σε αυτά. Φτιάχνοντας το
miRNA προφίλ έκφρασης φαίνεται ότι πολλά miRNAs απορυθμίζονται σε καρκίνους
του ανθρώπου. Τα miRNAs έχουν αποδειχθεί να ρυθμίζουν ογκογονίδια,
καταστολείς όγκων και μια σειρά από καρκινοσυσχετιζόμενα γονίδια που ελέγχουν
τον κυτταρικό κύκλο, την απόπτωση, τη μετανάστευση και την αγγειογένεση. Τα
miRNAs που κωδικοποιείται από την mir-17-92 δέσμη μερικά έχουν ογκογενικό
δυναμικό και άλλα μπορούν να ενεργούν ως καταστολείς όγκων (Eικόνα 20).
Μερικά miRNAs και ιδιαίτερα κάποιες ειδικές τους τοποθεσίες βρέθηκαν να είναι
μεταλλαγμένες σε καρκίνο. Τα miRNAs μπορούν να έχουν μεγάλη διαγνωστική
δυνατότητα για ανθρώπινο καρκίνο, ακόμα και miRNA θεραπείες για τον καρκίνο
μπορεί να είναι στον ορίζοντα.
Eικόνα 20: Χάρτης αντιδράσεων της ογκογενικής miR-17-92 δέσμης στο χρωμόσωμα 13 q13.3. Οι πρωτεΐνες στο κόκκινο και πράσινο είναι προϊόντα γνωστών ανθρώπινων ογκογονιδίων και ογκοκαταστολέων αντίστοιχα.
2. Καρδιαγγειακές νόσοι: Οι καρδιαγγειακές νόσοι αποτελούν την κύρια
αιτία ή μια από πς κυριότερες στον κόσμο. Συνήθως, προκύπτουν από τη σταδιακή
έμφραξη των αρτηριών μέσω μιας διεργασίας που ονομάζεται αρτηριοσκλήρυνση
και η οποία προκαλεί τελικά την καρδιακή προσβολή. Η αρτηριοσκλήρυνση

27
περιλαμβάνει γεγονότα, όπως είναι η τοπική παραγωγή κυτοκινών που σχετίζονται
με τη φλεγμονή, προσέλκυση μακροφάγων κυττάρων και τέλος απόπτωση των
λείων μυϊκών κυττάρων του αγγειακού συστήματος. Είναι πιθανόν, λοιπόν, να
μπορέσουμε να παρέμβουμε στη διαδικασία της αρτηριοσκλήρυνσης μειώνοντας τη
βλάβη του καρδιακού ιστού. Τέτοια παρεμβολή μπορεί να επιτευχθεί ή με siRNA
που καταστείλουν την υπερέκφραση κυτοκινών που σχετίζονται με πς φλεγμονές ή
ενζύμων των οποίων η παρατεταμένη λειτουργία συμβάλλει στα
αρτηριοσκληρωτικά φαινόμενα.
3. Νευροεκφυλιστικές διαταραχές: Μεταξύ των νευροεκφυλιστικών
νόσων που σχετίζονται με την ηλικία, οι νόσοι Alzheimer, Parkinson και
Huntington αποτελούν χαρακτηριστικά παραδείγματα. Κάθε μία από τις διαταραχές
αυτές χαρακτηρίζεται από τη δυσλειτουργία και τον θάνατο συγκεκριμένου
πληθυσμού νευρώνων. Στις νόσους Alzheimer και Parkinson, ένα μικρό ποσοστό
των περιπτώσεων οφείλεται σε συγκεκριμένες γενετικές μεταλλάξεις, ενώ η νόσος
Huntington οφείλεται αποκλειστικά σε. μεταλλάξεις στην πρωτεΐνη χαντικτίνη
(huntigtin). Μελέτες των νόσων αυτών σε ανθρώπους, καθώς και σε ζωικά
μοντέλα έχουν αποκάλυψα κοινές βιοχημικές οδούς οι οποίες προκαλούν τον
θάνατο των νευρώνων. Αυτοί περιλαμβάνουν το αυξημένο οξειδωτικό στρες, τη
δυσλειτουργία της κυτταρικής ομοιόστασης του ασβεστίου και την απόπτωση.
Εξαιτίας λοιπόν των προαναφερθέντων, οι περισσότερες προσπάθειες έχουν
επικεντρωθεί είτε στα αρχικά γεγονότα που προκαλούν τη νευροεκφυλιοτική
διαδικασία ή στα καταληκτικά γεγονότα στον νευροδιαβιβασπκό καταρράκτη
μεταγωγής μηνυμάτων.
4. Λοιμώδεις νόσοι: Πολλές ασθένειες που προέρχονται από ιούς ή βακτήρια
συνεχίζουν να αποτελούν κύριες αιτίες θανάτου παγκοσμίως. Παράλληλα, η
συνεχής εμφάνιση νέων, ανθεκτικών στελεχών και η πιθανή χρήση λοιμωδών
παθογόνων σε εμπόλεμες ή τρομοκρατικές ενέργειες προβληματίζουν τους
επιστήμονες. Ο ιός HIV, οι ιοί της ηπατίτιδας και της γρίπης, καθώς και ορισμένες
βακτηριακές λοιμώξεις που μπορούν να προκαλέσουν θάνατο, όπως η πνευμονία,
αποτελούν χαρακτηριστικά παραδείγματα. Η ικανότητα της μεθόδου του RNAi να
αναστέλλει τον πολλαπλασιασμό ή την είσοδο στο κύτταρο ιών και άλλων
παθογόνων, έχει ήδη αποδειχτεί σε in vitro πειράματα. Τα αποτελέσματα αυτά είναι
ενθαρρυντικά και για τον λόγο αυτό η τεχνολογία του RNAi υπόσχεται πολλά στον
τομέα της θεραπείας. Οι μελέτες αυτές περιλαμβάνουν τον περιορισμό της
ικανότητας του ιού HIV να προσβάλλει τα κύτταρα που έχουν επιμολυνθεί
προηγουμένως με siRNAs εναντίον της αντίστροφης μεταγραφάσης του, καθώς και
την προστασία του ήπατος των ποντικών με τη χρήση siRNAs εναντίον του
αποπτωτικού υποδοχέα Fas".

28
Τεχνικά θέματα στη χρήση του RNAi Για να τεθεί σε εφαρμογή η σίγηση ενός γονιδίου με τη χρήση του RNAi
απαιτείται πολύ προσεκτικός σχεδιασμός, καθώς και η επιλογή της κατάλληλης
στρατηγικής. Δύο στρατηγικές χρησιμοποιούνται σήμερα ευρέως. Η πρώτη αφορά
στον σχεδιασμό και την κατασκευή συνθετικών δίκλωνων ολιγονουκλεοτιδίων RNA
και την εισαγωγή τους στα κύτταρα με τη μέθοδο της επιμόλυνσης. Η τεχνική αυτή
είναι η πιο διαδεδομένη για κύτταρα σε καλλιέργεια. Πρόσφατες εργαστηριακές
μελέτες δείχνουν ότι η πιθανότητα επιτυχίας επιλεκτικής σίγηοης μπορεί να
αυξηθεί αν «στοχεύσουν» τα siRNAs σε συγκεκριμένες περιοχές του mRNA. Η
δεύτερη στρατηγική αφορά στην εισαγωγή στο εκάστοτε κύτταρο οχημάτων
μεταφοράς ή γονιδιωμάτων από ιούς που εκφράζουν siRNA με
συμπληρωματτκότητα στην αλληλουχία που επιθυμούμε να οδηγήσουμε σε
μεταγραφική σίγηση. Η επιλογή της μεθόδου που θα χρησιμοποιηθεί εξαρτάται από
την εφαρμογή της. Η επιμόλυνση με συνθετικά siRNAs εξασφαλίζει μόνο
προσωρινή σίγηση του γονιδίου-σιόχου. Ωστόσο, η μέθοδος αυτή μας επιτρέπει να
μελετήσουμε μια σειρά από βιολογικές παραμέτρους του κυττάρου οε σχέση με το
ποσοστό υποέκφρασης του γονιδίου- στόχου. Η μέθοδος αυτή χρησιμοποιείται
σήμερα σε ένα ευρύ φάσμα εφαρμογών. Αντιθέτως, η σταθερή ενδογενής
έκφραση των siRNAs από ενσωματωμένα ή επισωμικά αναπαραγώγιμα οχήματα
μεταφοράς μπορεί να προκαλέσει μακροχρόνια σίγηση της γοντδιακής έκφρασης,
γεγονός πολύ σημαντικό για τη χρήση της σε θεραπευτικές εφαρμογές, όπως η
ανπμετώπιοη χρόνιων ιικών λοιμοίξεων.
Αρκετοί τρόποι μεταφοράς του siRNΑ βρίσκονται υπό έρευνα. Ανάμεσα σε
αυτούς περιλαμβάνονται η άμεση μεταφορά siRNΑ μέσα σε φυσιολογικό ορό, η
ενθυλάκωση του σε λιποσώματα (liposomes) και λιποπλέγματα (lipoplexes), η
σύζευξη του με αντισώματα, πεπτίδια, απταμερή (aptamers) ή άλλα μόρια, και
επίσης ο σχη-ματισμός συμπλόκων με χημικά και βιολογικά πολυμερή. Κάθε είδος
μεταφοράς έχει τα προτερήματα της αλλά και συγκεκριμένες προκλήσεις8.
Μελλοντικές εφαρμογές του RNAi στις Επιστήμες Ζωής (Βιολογία και Ιατρική).
Αν και βρισκόμαστε σε πρώιμο στάδιο της κατανόησης των μηχανισμών της
τεχνολογίας του RNAi και της ανάπτυξης των μεθόδων με τις οποίες θα μπορέσει
να πραγματοποιηθεί η αποσιώπηση των γονιδίων, είναι πλέον φανερό πως στο
μέλλον το RNAi θα αποτελέσει ένα πειραμαπκό εργαλείο ευρείας χρήσης για τον
καθορισμό: (1) της λειτουργίας των γονιδίων.και (2) της γονιδιακής
θεραπείας.

29
Ηδη κατά την ανάπτυξη του κειμένου για την Ιστορία του siRNA
αναφερθήκαμε στην μεθοδολογία για την εύρεση της λειτουργίας των γονιδιακών
προϊόντων με την δημιουργία 16.757 βακτηριακών στελεχών (συνιστούν περίπου
το 86% των γνωστών γονιδίων στον σκώληκα C. Elegance), που το κάθε ένα
εκφράζει διπλής αλυσίδας RNA με στόχο ένα διαφορετικό γονίδιο του σκώληκα. Το
διπλής αλυσίδας RNA που παράγεται από τα βακτήρια αποσιωπεί το γονίδιο στόχο
στον σκώληκα , επιτρέποντας έτσι στον ερευνητή να βρεί τον φαινότυπο που
οφείλεται στην αδρανοποίηση του γονιδίου στόχου. Ωστόσο, υπάρχουν ακόμη
αρκετά προβλήματα που θα πρέπει να αντιμετωπιστούν, με πρώτο και κυριότερο
την εφαρμογή των μεθόδων του RNAi σε in vivo μελέτες. Η πολύ αργή, σε σχέση
με το αναμενόμενο, πρόοδος που παρατηρείται στην εφαρμογή της γονιδιακής
θεραπείας έχει αναγάγει τη μέθοδο του RNAi σε ιδιαίτερα σημαντική ελπίδα για
την ανπμετώπιση πολλών ασθενειών. Είναι βέβαια δύσκολο στην παρούσα φάση
να οριστεί με σαφήνεια το ακριβές φάσμα των επιδράσεων των siRNA σε γονίδια-
στόχους. Το ακριβές ποσοστό σίγησης γονιδίων-στόχων αποτελεί ένα ακόμη
αντικείμενο έρευνας. Πιθανότατα η παρεμβολή RNA θα χρησιμοποιηθεί σε
συνδυασμό με άλλες μεθόδους οι οποίες θα βοηθήσουν οτον εντοπισμό του siRNA
στους κατάλληλους ιστούς και στην αξιοποίηση του αποπελέσματος σίγησης για τη
μεταβολή της κυτταρικής φυσιολογίας προς την επιθυμητή κατεύθυνση.
ΣΥΜΠΕΡΑΣΜΑΤΙΚΑ Τα κύτταρα διαθέτουν έναν ειδικό μηχανισμό για την καταστολή της
έκφρασης ομόλογων γονιδίων μέσω αναγνώρισης και παραγωγής δίκλωνων RNAs.
Ο μηχανισμός αυτός μπορεί να χρησιμοποιεί δίκλωνο RNA που εισέρχεται στο
κύτταρο καθώς και δίκλωνο RNA που δημιουργείται εντός του κυττάρου (Εικόνα
21). Η σωστή ανάπτυξη ενός οργανισμού καθώς και η σωστή λειτουργία των
κυττάρων και των ιστών εξαρτώνται από έναν άθικτο-υγιή RNAi μηχανισμό. Στην
περίπτωση κυρίως των φυτών και λιγότερο στην περίπτωση των ζώων, ο
συγκεκριμένος μηχανισμός μπορεί να προστατεύει από ιϊκές μολύνσεις αλλά και
από ξένης προέλευσης στοιχεία που έχουν ενσωματωθεί στο γονιδίωμα και τα
οποία παραμένουν αποσιωποιημένα. Είναι επίσης ιδιαίτερα σημαντικό να αναφερθεί
πως ο συγκεκριμένος μηχανισμός πολλές φορές αναφέρεται σαν μια knockdown
μέθοδος έτσι ώστε να διαχωρίζεται από τις αμιγώς knockout μεθόδους. Και αυτό
διότι μέσω του RNAi είναι πιθανό να μην επιτευχθεί ολική γονιδιακή αποσιώπηση.
Το αποτέλεσμα του εξαρτάται από πολλούς παράγοντες μεταξύ των οποίων
αποτελούν οι πειραματικές συνθήκες, ο οργανισμός στον οποίο θα εφαρμοστεί ο
RNAi μηχανισμός αλλά και το πώς θα επιλέξει ο εκάστοτε ερευνητής να τον
εφαρμόσει ανάλογα με το τι μελετάει. Έτσι μπορεί να έχουμε ολική ή μερική
30
γονιδιακή αποσιώπηση και της οποίας τα αποτελέσματα να διαρκούν είτε για μια
περίοδο περίπου εφτά ημερών ή και να είναι σταθερά.
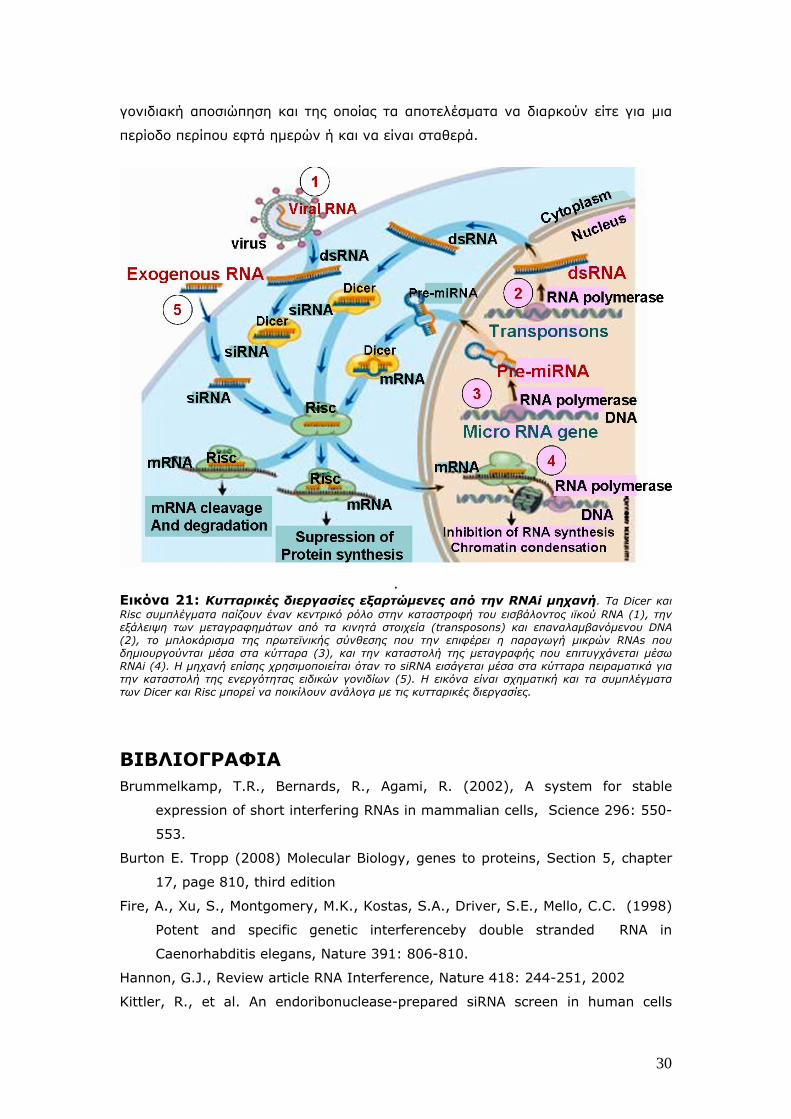
. Εικόνα 21: Κυτταρικές διεργασίες εξαρτώμενες από την RNAi μηχανή. Τα Dicer και Risc συμπλέγματα παίζουν έναν κεντρικό ρόλο στην καταστροφή του εισβάλοντος ιϊκού RNA (1), την εξάλειψη των μεταγραφημάτων από τα κινητά στοιχεία (transposons) και επαναλαμβανόμενου DNA (2), το μπλοκάρισμα της πρωτεϊνικής σύνθεσης που την επιφέρει η παραγωγή μικρών RNAs που δημιουργούνται μέσα στα κύτταρα (3), και την καταστολή της μεταγραφής που επιτυγχάνεται μέσω RNAi (4). Η μηχανή επίσης χρησιμοποιείται όταν το siRNA εισάγεται μέσα στα κύτταρα πειραματικά για την καταστολή της ενεργότητας ειδικών γονιδίων (5). Η εικόνα είναι σχηματική και τα συμπλέγματα των Dicer και Risc μπορεί να ποικίλουν ανάλογα με τις κυτταρικές διεργασίες.
ΒΙΒΛΙΟΓΡΑΦΙΑ Brummelkamp, T.R., Bernards, R., Agami, R. (2002), A system for stable
expression of short interfering RNAs in mammalian cells, Science 296: 550-
553.
Burton E. Tropp (2008) Molecular Biology, genes to proteins, Section 5, chapter
17, page 810, third edition
Fire, A., Xu, S., Montgomery, M.K., Kostas, S.A., Driver, S.E., Mello, C.C. (1998)
Potent and specific genetic interferenceby double stranded RNA in
Caenorhabditis elegans, Nature 391: 806-810.
Hannon, G.J., Review article RNA Interference, Nature 418: 244-251, 2002
Kittler, R., et al. An endoribonuclease-prepared siRNA screen in human cells

31
identifies genes essential for cell division. Nature, 432, 1036-40 (2004).
Tang, G., (2005) siRNA and miRNA: an insight into RISCs, TRENDS BIOCHEM
SCI. 30: 106-114
Yang, D., et al. Short RNA duplexes produced by hydrolysis with Escherichia coli
RNase III mediate effective RNA interference in mammalian cells. Proc. Natl.
Acad. Sci. U.S.A., 99, 9942-7 (2002). Zamore, P.D., Tuschi, T., Sharp, P.A., Bartels, D.P. (2000), RNAi: double-
stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals Cell 101: 25-33.


















