
Thymosin β4 in humanem Speichel: Quellen und Einflussfaktoren · Meinem Bruder. Μὴ δήτει...
136
Aus dem Institut für Biochemie der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg Lehrstuhl für Biochemie und Pathobiochemie Direktor: Prof. Dr. M. Wegner und Aus der Zahnklinik 1 – Zahnerhaltung und Parodontologie der Friedrich-Alexander-Universität Erlangen-Nürnberg Direktor: Prof. Dr. med. dent. A. Petschelt Thymosin β4 in humanem Speichel: Quellen und Einflussfaktoren Inaugural-Dissertation zur Erlangung der Doktorwürde der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg Vorgelegt von Moritz Zimmermann aus Sulzbach-Rosenberg
Transcript of Thymosin β4 in humanem Speichel: Quellen und Einflussfaktoren · Meinem Bruder. Μὴ δήτει...

Aus dem Institut für Biochemie der Medizinischen Fakultät
der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Lehrstuhl für Biochemie und Pathobiochemie
Direktor: Prof. Dr. M. Wegner
und
Aus der Zahnklinik 1 – Zahnerhaltung und Parodontologie
der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. med. dent. A. Petschelt
Thymosin β4 in humanem Speichel:
Quellen und Einflussfaktoren
Inaugural-Dissertation
zur Erlangung der Doktorwürde
der Medizinischen Fakultät
der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Vorgelegt von
Moritz Zimmermann
aus
Sulzbach-Rosenberg

Gedruckt mit Erlaubnis der Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Dekan: Prof. Dr. J. Schüttler
Referent: Prof. Dr. E. Hannappel
Korreferent: Prof. Dr. M. Pelka
Tag der mündlichen Prüfung: 30. März 2011

Meinem Bruder.
Μὴ δήτει τὰ γινόμενα γίνεσθαι ὡς θέλεις, ἀλλὰ θέλε τὰ γινόμενα ὡς γίνεται καὶ εὐροήσεις.
-
Verlange nicht, dass das, was geschieht, so geschieht, wie du es wünschst, sondern wünsche,
dass es so geschieht, wie es geschieht, und dein Leben wird heiter dahinströmen.
Epiktet, Handbüchlein der Moral, Kapitel 8

Thymosin β4 in humanem Speichel:
Quellen und Einflussfaktoren
Moritz Zimmermann
ZUSAMMENFASSUNG ........................................................................ 1
1. EINLEITUNG ................................................................................... 5
1.1. Thymus........................................................................................................................ 5 1.1.1. Die Rolle des Thymus im Immunsystem ............................................................. 5
1.1.2. Geschichte der Thymusforschung und die Thymosin Fraktion 5 ........................ 5
1.1.3. α, β und γ-Thymosine .......................................................................................... 7
1.2. Thymosine ................................................................................................................... 8 1.2.1. Vertreter der α-Thymosine ................................................................................... 8
1.2.2. Vertreter der β-Thymosine ................................................................................... 9
1.3. Thymosin β4 ............................................................................................................. 10 1.3.1. Chemische Eigenschaften .................................................................................. 10
1.3.2. Struktur .............................................................................................................. 11
1.3.3. Vorkommen ....................................................................................................... 11
1.3.4. Hormonelle Funktionen ..................................................................................... 12
1.3.5. Intrazelluläre Funktionen ................................................................................... 13
1.3.6. Extrazelluläre Funktionen .................................................................................. 14
1.3.7. Thymosin β4 in Körperflüssigkeiten .................................................................. 15
1.4. Mundhöhle ................................................................................................................ 16 1.4.1. Topografie und Gliederung ................................................................................ 16
1.5. Drüsenapparat .......................................................................................................... 17 1.5.1. Topografie .......................................................................................................... 17
1.5.2. Histologischer Aufbau der Speicheldrüsen und Mechanismus der Speichel-
sekretion ............................................................................................................. 19
1.5.3. Speichelzusammensetzung ................................................................................. 21
1.5.4. Speichelfunktionen............................................................................................. 22
1.6. Zahnhalteapparat .................................................................................................... 23 1.6.1. Anatomischer Aufbau ........................................................................................ 23
1.6.2. Gingiva ............................................................................................................... 24
1.6.2.1. Anatomischer Aufbau .................................................................................... 24
1.6.2.2. Saumepithel .................................................................................................... 25
1.6.2.3. Bindegewebe .................................................................................................. 26
1.6.2.4. Gingivaler Sulkus und Sulkusfluid ................................................................ 26
1.6.3. Zement ............................................................................................................... 29
1.6.4. Desmodont ......................................................................................................... 30
1.6.5. Alveolarknochen ................................................................................................ 30

1.7. Parodontopathien ..................................................................................................... 31 1.7.1. Allgemeines ....................................................................................................... 31
1.8. Gingivitis und Parodontitis ..................................................................................... 32 1.8.1. Definition ........................................................................................................... 32
1.8.2. Ätiologie ............................................................................................................ 32
1.8.2.1. Plaque ............................................................................................................. 32
1.8.2.2. Abwehrreaktionen des Wirtsorganismus ....................................................... 34
1.8.2.3. Lokale und systemische Faktoren .................................................................. 36
1.8.3. Pathogenese ........................................................................................................ 36
1.8.3.1. Initiale Läsion ................................................................................................ 37
1.8.3.2. Frühe Läsion .................................................................................................. 37
1.8.3.3. Etablierte Läsion ............................................................................................ 38
1.8.3.4. Fortgeschrittene Läsion .................................................................................. 39
2. PROBLEMSTELLUNG ................................................................ 40
2.1. Stand der Forschung ................................................................................................ 40 2.1.1. Extrazelluläre Funktionen des Thymosin β4 ...................................................... 40
2.1.2. Nachweis von oralem Thymosin β4 ................................................................... 40
2.1.3. Funktionelle Bedeutung des oralen Thymosin β4 .............................................. 40
2.2. Zielsetzung der Arbeit ............................................................................................. 40
3. KONZEPT DER STUDIE ............................................................. 42
3.1. Gruppeneinteilung der Probanden ......................................................................... 42
3.2. Beobachtungsparameter .......................................................................................... 43
4. MATERIAL UND METHODEN .................................................. 44
4.1. Material ..................................................................................................................... 44 4.1.1. Chemikalien und Reagenzien ............................................................................. 44
4.1.2. Geräte und Hilfsmittel ........................................................................................ 44
4.1.2.1. Analytische bzw. präparative RP-HPLC ........................................................ 44
4.1.2.2. Sonstiges ........................................................................................................ 45
4.1.3. Pufferlösungen ................................................................................................... 45
4.2. Methoden .................................................................................................................. 45 4.2.1. Probengewinnung............................................................................................... 45
4.2.1.1. Gesamtspeichel (Gruppe A und B) ................................................................ 46
4.2.1.2. Gesamtspeichel (Gruppe C) ........................................................................... 46
4.2.1.3. Drüsenspeichel (Gruppe C) ............................................................................ 46
4.2.1.4. Sulkusfluid parodontal gesunder Probanden (Gruppe C) .............................. 47
4.2.1.5. Sulkusfluid parodontal erkrankter Probanden (Gruppe D) ............................ 47
4.2.1.6. Sulkusfluid unter verstärkter okklusaler Belastung (Gruppe E) .................... 47
4.2.2. Bestimmung der sulkulären Taschentiefen ........................................................ 48
4.2.3. Bestimmung des API und SBI ........................................................................... 48

4.2.4. Probenvorbereitung ............................................................................................ 50
4.2.4.1. Gesamtspeichel und Drüsenspeichel .............................................................. 50
4.2.4.2. Sulkusfluid ..................................................................................................... 50
4.2.5. Proteinbestimmung mittels Biuretreaktion ........................................................ 51
4.2.5.1. Prinzip Fotometer ........................................................................................... 51
4.2.5.2. Prinzip Biuretreaktion .................................................................................... 52
4.2.5.3. Probenanalyse ................................................................................................ 52
4.2.6. Thymosin β4 Bestimmung mittels analytischer RP-HPLC ............................... 53
4.2.6.1. Prinzip der RP-HPLC..................................................................................... 53
4.2.6.2. Analytische RP-HPLC mit Fluram-Nachsäulenderivatisierung .................... 53
4.2.6.3. Probenanalyse ................................................................................................ 56
4.2.6.4. Präparative RP-HPLC mit UV-Detektion ...................................................... 57
4.2.7. Peptidaufkonzentrierung und Entsalzung mit RP-18-Säule ............................... 57
4.2.8. Massenspektrometrie der aufgereinigten Proteine ............................................. 58
4.2.9. Stabilität von Thymosin β4 im Gesamtspeichel................................................. 58
5. ERGEBNISSE ................................................................................. 60
5.1. Thymosin β4 Konzentration im Gesamtspeichel................................................... 60 5.1.1. Probanden der Gruppe A .................................................................................... 61
5.1.2. Probanden der Gruppe B .................................................................................... 63
5.2. Proteinkonzentration im Gesamtspeichel .............................................................. 65 5.2.1. Probanden der Gruppe A .................................................................................... 65
5.2.2. Probanden der Gruppe B .................................................................................... 67
5.3. Quellen für orales Thymosin β4 .............................................................................. 69 5.3.1. Thymosin β4 Konzentration im Gesamtspeichel ............................................... 70
5.3.2. Thymosin β4 Konzentration im Speichel der gl. parotis ................................... 72
5.3.3. Thymosin β4 Konzentration im Speichel der gl. submandibularis/sublingualis 72
5.3.4. Sulkusfluid als Quelle für orales Thymosin β4 .................................................. 73
5.4. Thymosin β4 im Sulkusfluid parodontal gesunder Probanden ........................... 74 5.4.1. Sulkusfluidvolumen ........................................................................................... 74
5.4.2. Thymosin β4 Konzentration im Sulkusfluid ...................................................... 76
5.4.3. Bestimmung der sulkulären Taschentiefen ........................................................ 77
5.4.4. Bestimmung des API und SBI ........................................................................... 78
5.4.5. Überblick über die Einflussfaktoren .................................................................. 79
5.5. Thymosin β4 im Sulkusfluid parodontal erkrankter Probanden ........................ 80 5.5.1. Sulkusfluidvolumen ........................................................................................... 81
5.5.2. Thymosin β4 Konzentration im Sulkusfluid ...................................................... 83
5.5.3. Bestimmung der sulkulären Taschentiefen ........................................................ 84
5.5.4. Bestimmung des API und SBI ........................................................................... 85
5.5.5. Überblick über die Einflussfaktoren .................................................................. 86
5.6. Thymosin β4 im Sulkusfluid unter verstärkter okklusaler Belastung ................ 87 5.6.1. Sulkusfluidvolumen ........................................................................................... 87
5.6.2. Thymosin β4 Konzentration im Sulkusfluid ...................................................... 88
5.7. Elutionsversuche mit Papierspitzen ....................................................................... 90

5.8. Massenspektrometrie ............................................................................................... 91
5.9. Stabilität von Thymosin β4 im Gesamtspeichel .................................................... 92
6. DISKUSSION .................................................................................. 95
6.1. Thymosin β4 Konzentration im Gesamtspeichel – Vergleich Gruppe A
und B ......................................................................................................................... 95
6.2. Proteinkonzentration im Gesamtspeichel – Vergleich Gruppe A und B ............ 96
6.3. Korrelation Thymosin β4 Konzentration und Proteinkonzentration im
Gesamtspeichel (Gruppe A und B) ......................................................................... 97
6.4. Korrelation Thymosin β4 Konzentration im Sulkusfluid und im Gesamt-
speichel (Gruppe C) ................................................................................................. 99
6.5. Thymosin β4 im Sulkusfluid (Gruppe C) – Übersicht über die Einfluss-
faktoren ................................................................................................................... 100
6.6. Thymosin β4 im Sulkusfluid (Gruppe D) – Übersicht über die Einfluss-
faktoren ................................................................................................................... 102
6.7. Thymosin β4 Konzentration im Sulkusfluid – Vergleich Gruppe C und D...... 103
6.8. Korrelation Sulkusfluidvolumen und Thymosin β4 Konzentration im
Sulkusfluid (Gruppe C und D) .............................................................................. 105
6.9. Thymosin β4 im Sulkusfluid – Freisetzung infolge verstärkter okklusaler
Belastung ................................................................................................................. 106
6.10. Stabilität von Thymosin β4 im Gesamtspeichel .................................................. 107
6.11. Vergleich der Ergebnisse mit dem aktuellen Stand der Forschung .................. 107
6.12. Mechanismus der Anreicherung von Thymosin β4 im Sulkusfluid .................. 109
6.13. Mögliche Rolle von Thymosin β4 bei Parodontopathien .................................... 110
6.14. Thymosin β4 als mögliches Diagnostiktool für Parodontopathien .................... 111
LITERATURVERZEICHNIS ........................................................... 112
ABKÜRZUNGSVERZEICHNIS ...................................................... 121
ABBILDUNGSVERZEICHNIS ........................................................ 124
TABELLENVERZEICHNIS ............................................................. 125

DIAGRAMMVERZEICHNIS ........................................................... 126
DANKSAGUNG .................................................................................. 127
LEBENSLAUF .................................................................................... 128

1
Zusammenfassung
Hintergrund und Ziele
Die oralen Flüssigkeiten Speichel und Sulkusfluid stellen vielversprechende Quellen für die
Untersuchung von Thymosin β4 dar, da sie schnell und vor allem nicht invasiv gewonnen
werden können. Das Auftreten von Thymosin β4 in humanem Speichel konnte in einer kürz-
lich veröffentlichten Studie nachgewiesen werden. Weitere Studien befassten sich mit einer
möglichen Funktion des oralen Thymosin β4, das unter anderem die Dentinogenese und die
Entwicklung der Mundhöhle beeinflusst. Wenngleich orales Thymosin β4 in letzter Zeit ver-
stärkt Gegenstand der Forschung gewesen ist, so herrscht dennoch weiterhin Unklarheit über
mögliche Quellen und Einflussfaktoren für das Auftreten von Thymosin β4 in humanem
Speichel. Ein möglicher Freisetzungsmechanismus des Peptids und ein eventueller Zusam-
menhang des Auftretens von oralem Thymosin β4 mit oralen Erkrankungen wurden eben-
falls noch nicht analysiert.
In dieser Arbeit werden demnach verschiedene Quellen und Einflussfaktoren für das Auftre-
ten von Thymosin β4 in humanem Speichel untersucht, mit dem Ziel, die funktionelle Be-
deutung dieses Peptids innerhalb der Mundhöhle genauer zu verstehen.
Methoden
Der Nachweis und die quantitative Bestimmung von oralem Thymosin β4 erfolgte mittels
RP-HPLC und MALDI-TOF. Zunächst wurden die Proteinkonzentration und die Konzentra-
tion von Thymosin β4 in humanem Speichel in Abhängigkeit von der Zeit in zwei unter-
schiedlichen Altersgruppen untersucht. Anschließend wurden verschiedene Quellen für ora-
les Thymosin β4 analysiert. Danach erfolgte die Untersuchung der Thymosin β4 Konzentra-
tion im Sulkusfluid bei parodontal gesunden und erkrankten Probanden. Zuletzt wurde der
Einfluss einer verstärkten okklusalen Belastung auf die Thymosin β4 Konzentration im Sul-
kusfluid untersucht.
Ergebnisse
Die Thymosin β4 Konzentration im Gesamtspeichel variierte unabhängig vom Alter der Pro-
banden sehr stark und war über die Zeit nicht konstant. Die durchschnittliche Thymosin β4
Konzentration war bei älteren Probanden größer als bei jüngeren Probanden. Das Sulkusfluid
konnte als Hauptquelle für das orale Thymosin β4 ausgemacht werden. Drüsenspeichel ent-
hielt keine nachweisbaren Konzentrationen an Thymosin β4. Bei der Analyse der Thymosin
β4 Konzentration im Sulkusfluid parodontal erkrankter und gesunder Probanden ergaben
sich neben den bereits beobachteten interindividuellen zusätzlich noch unterschiedlich starke

2
intraindividuelle Schwankungen. Bei parodontal erkrankten Probanden wurden bei generell
erhöhten Volumina an Sulkusfluid durchschnittlich geringere Konzentrationen an Thymosin
β4 nachgewiesen. Eine verstärkte okklusale Belastung führte zu keiner deutlichen Verände-
rung der Thymosin β4 Konzentration im Sulkusfluid.
Praktische Schlussfolgerungen
In dieser Arbeit konnte nachgewiesen werden, dass das Sulkusfluid die Hauptquelle des ora-
len Thymosin β4 darstellt und dass dessen Konzentration von Einflussfaktoren wie dem Zeit-
punkt der Probennahme, dem Alter und dem Parodontalstatus des Probanden abhängig ist.
Die Ergebnisse legen zudem nahe, dass es besondere Mechanismen der lokalen Anreicher-
ung von Thymosin β4 im gingivalen Bindegewebe und eine dementsprechende Freisetzung
in das Sulkusfluid geben muss. Während die Produktion des Sulkusfluids von Mediatoren
der vaskulären Permeabilität abhängig ist, könnte der Übertritt von Thymosin β4 in den gin-
givalen Sulkus von bisher nicht identifizierten spezifischen Mediatoren gesteuert werden. In
anderen Geweben oder Flüssigkeiten, wie beispielsweise im Blut, wurden bisher wesentlich
geringere Thymosin β4 Konzentrationen nachgewiesen. Deshalb sollte das in erstaunlich
hohen Konzentrationen auftretende orale Thymosin β4 von großer funktioneller Bedeutung
sein.

3
Summary
Background and objectives
The oral fluids saliva and gingival crevicular fluid seem to be very promising sources for the
investigation of thymosin β4 as they are readily available. Recent investigations revealed
thymosin β4 to be present in whole saliva and provided meaningful insights into the possible
function of oral thymosin β4 reported to influence dentinogenesis and the development of the
oral cavity. The data mentioned reflect an increasing interest for oral thymosin β4, but its
source in the oral cavity, the influences for its secretion, the secretion process itself, its
function and its possible diagnostic value for diseases of the oral cavity are far from being
understood. We therefore investigated possible sources and influences for the secretion of
thymosin β4 trying to find an answer for its possible functions in the oral cavity.
Methods
Determination and identification of thymosin β4 was carried out by RP-HPLC and MALDI-
TOF. At first we studied the protein concentration and the thymosin β4 concentration in
whole saliva as a function of time in two different age groups. We then tried to find out
different possible sources for thymosin β4 in the oral fluids. Afterwards we investigated the
thymosin β4 concentration in the gingival crevicular fluid from periodontal diseased patients
and compared it to healthy test persons. Concerning a possible release of thymosin β4 from
cells into the gingival crevicular fluid we tested the influence of increased occlusal contact
on the concentration of thymosin β4 present in the gingival crevicular fluid.
Results
At first whole saliva was analyzed: independent of the test persons´ age presence of oral thy-
mosin β4 varied considerably even showing significant changes when new specimens were
taken after two weeks. The mean concentration of thymosin β4 was determined to be higher
in older than in younger test persons. We revealed the gingival crevicular fluid to be the
main source of oral thymosin β4 by performing separate analysis of whole saliva, glandular
saliva and gingival crevicular fluid. The thymosin β4 concentration in both healthy and di-
seased test persons varied not only interindividually as yet observed in whole saliva but also
intraindividually. The mean concentration of thymosin β4 was determined to be lower in
periodontal diseased patients whose mean volume of the gingival crevicular fluid was deter-
mined to be higher. Finally we did not find any significant differences of the thymosin β4

4
concentration in samples of gingival crevicular fluid obtained with or without increased
occlusal contact.
Conclusions
We revealed the gingival crevicular fluid to be the main source for oral thymosin β4 and de-
monstrated that its concentration is influenced by factors like the time of sample collection
and the test persons´ age and periodontal status. Our findings suggest that there have to be
special mechanisms to ensure the local enrichment of thymosin β4 in the gingival connective
tissue and the gingival crevicular fluid. Microvascular leakage and direct systemic migration
of thymosin β4 into the gingival crevicular fluid cannot explain the high extracellular con-
centrations of thymosin β4 as they are significant higher than in whole blood. While the
influx of the gingival crevicular fluid depends on mediators of vascular permeability, the
emigration of thymosin β4 into the gingival crevice might be influenced by specific media-
tors yet unidentified. Due to these high concentrations, thymosin β4 might fulfill essential
functions in the oral environment.

5
1. Einleitung
1.1. Thymus
1.1.1. Die Rolle des Thymus im Immunsystem
Der Thymus wurde bereits zu Zeiten der Griechen entdeckt (thymos (gr.) = Herz, Seele).
Wenngleich man damals noch nichts über die wichtige Bedeutung des Thymus für das Im-
munsystem wusste, erahnte man jedoch bereits dessen besondere Funktion, denn man sah in
ihm eine Quelle der Gesundheit [95].
Der Thymus ist ein von einer Kapsel umschlossenes, kegelförmiges Organ mit drüsenähn-
lichem Aufbau und zählt neben dem Knochenmark zu den primären lymphatischen Organen.
Er besteht aus zwei untereinander verbundenen Lappen und befindet sich im oberen Medias-
tinum [96]. Im Laufe der Entwicklung unterliegt der Thymus einer umfangreichen morpho-
logischen Veränderung. Diese wird als Thymusinvolution bezeichnet und ist funktionell be-
dingt. Der Thymus entwickelt sich zunächst während der Embryogenese als lymphoepitheli-
ales Organ aus dem Epithel der dritten Schlundtasche und erreicht seine größte Ausdehnung
zwischen dem neunten und zwölften Lebensjahr [96]. In dieser Zeit übernimmt der Thymus
seine besondere Funktion im Rahmen der spezifischen zellulären Immunabwehr. Im Kno-
chenmark gebildete Lymphozyten gelangen über periphere Blutgefäße in den Thymus und
wandern durch Rinde und Mark ins Zentrum. 90 % der eingewanderten Zellen werden elimi-
niert, die restlichen Zellen reifen zu immunkompetenten T-Lymphozyten, indem sie mit spe-
zifischen Oberflächenproteinen ausgestattet werden. Diese ermöglichen den T-Lymphozyten
zwischen körperfremden und körpereigenen Strukturen zu unterscheiden. Nach der immuno-
logischen Prägung verlassen die T-Lymphozyten den Thymus. Sie zirkulieren dann entweder
im Blut oder besiedeln größtenteils die sekundären lymphatischen Organe, wie beispielswei-
se die Milz oder die Lymphknoten, wo sie im Rahmen der spezifischen zellulären Immunab-
wehr an der Elimination körperfremder Antigene beteiligt sind. Nachdem die spezifische Im-
munabwehr mit der Ausbildung immunkompetenter T-Lymphozyten etabliert wurde und
demzufolge die angeborene unspezifische Immunabwehr sinnvoll unterstützen kann, bildet
sich der Thymus schließlich im Rahmen der Thymusinvolution nach der Pubertät zu einem
retrosternalen Fettkörper zurück [96].
1.1.2. Geschichte der Thymusforschung und die Thymosin Fraktion 5
Die ersten Studien zur immunologischen Funktion des Thymus gehen auf das Jahr 1961 zu-
rück [80]. Miller erkannte, dass bei Mäusen nach einer Thymektomie sowohl in lymphati-
schen Organen als auch im Blut weniger Lymphozyten vorhanden waren. Gleichzeitig ent-

6
deckte er, dass das Ausmaß dieses Mangels an Lymphozyten vom Zeitpunkt der Entfernung
des Thymus abhängig ist. Kurze Zeit später kamen dann Burnet et al. zu der noch heute gül-
tigen Ansicht, dass der Thymus der Ort der Proliferation und Differenzierung von Lympho-
zyten mit definierter immunologischer Funktion ist [20]. Daraufhin begann man mit der
Suche nach Molekülen, die für eben jene immunologischen Effekte des Thymus verantwort-
lich sind [68]. Im Jahr 1966 gelang es der Gruppe um Goldstein et al. erstmals mittels
mehrerer Reinigungsschritte einen lymphozytopoetischen Faktor aus dem Thymus eines
Kalbs zu isolieren. Dieser Faktor wurde in Anlehnung an die Isolation aus dem Ursprungs-
organ Thymus als Thymosin bezeichnet [40]. Im weiteren Verlauf der Forschung konzen-
trierte man sich auf die durch fünf Reinigungsschritte gewonnene Thymosin Fraktion 5, von
der man anfangs dachte, dass sie aus nur einem einzigen Polypeptid mit einer Masse von 14
kDa besteht [38]. Diese Fraktion erwies sich in zahlreichen Versuchen als biologisch wirk-
sam. So konnte man beispielsweise bei Mäusen, deren Thymus entfernt wurde, nach Injek-
tion jener Fraktion 5 eine Reetablierung der zellvermittelten Immunität beobachten [70].
Aber auch entsprechende Versuche in vitro waren vielversprechend [37]. Die Abbildung 1
zeigt die zur Herstellung der Thymosin Fraktion 5 benötigten Schritte.
Abbildung 1: Reinigungsschritte zur Herstellung der Thymosin Fraktion 5 (nach Goldstein,
2007)
Thymusgewebe
14000 x g
(Fraktion 1)
80 °C Erhitzen
(Fraktion 2)
Zugabe von Aceton
(Fraktion 3)
Zugabe 25 – 50 %
(NH4)2SO4
(Fraktion 4)
Ultrafiltration
Sephadex G-25
(Fraktion 5)

7
1.1.3. α, β und γ-Thymosine
Durch weitere Analyse fand man heraus, dass die Thymosin Fraktion 5 aus mindestens 40
verschiedenen Hauptpeptiden mit einem Molekulargewicht von 1 – 15 kDa besteht. Eine
Einteilung der verschiedenen Peptide erfolgte nach ihrem isoelektrischen Punkt (pI) [57].
Die Abbildung 2 verdeutlicht deren Nomenklatur auf der Grundlage der isoelektrischen Fo-
kussierung. Demnach werden Thymosine mit einem isoelektrischen Punkt kleiner als pH 5
als α-Thymosine, zwischen pH 5 und pH 7 als β-Thymosine und über pH 7 als γ-Thymosine
bezeichnet [39]. Die weitere Benennung der Thymosine erfolgte mittels tiefgestellter Zahl
gemäß der chronologischen Reihenfolge ihrer Isolierung.
Abbildung 2: Isoelektrische Fokussierung und Nomenklatur der Thymosine (nach Goldstein,
2007)
Im weiteren Verlauf galt es nun, einzelne Vertreter der Thymosinfamilie zu isolieren, zu
sequenzieren und deren genaue Funktion zu untersuchen.

8
1.2. Thymosine
1.2.1. Vertreter der α-Thymosine
Die ersten zwei Thymosine, die isoliert und deren Aminosäuresequenz bestimmt werden
konnte, waren Thymosin α1 und β1 [76]. Thymosin α1 hat sich im Laufe der Jahre als ein
wichtiges immunmodulatorisches Molekül herausgestellt [36] und wird derzeit in über 37
Ländern zur Behandlung von Hepatitis B und C und als Hilfsmittel bei einer Vielzahl von
Erkrankungen angewendet [35]. Thymosin α1 umfasst die 28 ersten Aminosäurereste von
Prothymosin α und wird während der Isolierung von Thymosin Fraktion 5 generiert [32]. Die
Abbildung 3 gibt einen kurzen Überblick über die gegenwärtig bekannten Funktionen des
Thymosin α1.
Abbildung 3: Biologische Eigenschaften des Thymosin α1 (nach Goldstein, 2007)
Thymosin α1 blieb bisher jedoch der einzige Vertreter außerhalb der Gruppe der β-Thymo-
sine, dem eine gewisse funktionelle Relevanz nachgewiesen werden konnte. Aus diesem
Grund konzentrierte sich die weitere Forschung vor allem auf die Gruppe der β-Thymosine,
deren Vertreter eine stark ausgeprägte biologische Aktivität zeigten.
Thymosin
α1
Aktivierung der p38
MAPK und NFkB
SignalwegeZunahme der Differenzie-
rung von Toll-Like
Rezeptoren
Zunahme der Expression von MHC-I Molekülen
Blockade der Steroid-
induzierten Apoptose in Thymozyten
Wiederher-stellung der Immunant-
wort immun-supprimier-
ter Tiere
Stimulation von IL-2,
IFN-α, IL-6, IL-7

9
1.2.2. Vertreter der β-Thymosine
Thymosin β1 wurde zeitgleich mit Thymosin α1 isoliert und sequenziert [76]. Wenngleich
Thymosin β1 das prominenteste Polypeptid in der Thymosin Fraktion 5 ist, zeigte es in bio-
logischen Versuchen jedoch keine Aktivität. Es wurde schließlich als ein aus 74 Aminosäu-
ren bestehendes N-terminales Fragment von Ubiquitin mit einem fehlenden Glycin-Glycin
Rest am C-Terminus identifiziert [103]. Nach der Isolation von Thymosin β2 und β3, deren
Sequenzen allerdings nicht publiziert sind, wurde Thymosin β4 mittels Ionenaustauschchro-
matografie und Gelfiltration isoliert und anschließend sequenziert [74]. Thymosin β4 stellte
sich schnell als der mengenmäßig und funktionell bedeutendste Vertreter der β-Thymosine in
Säugerzellen heraus. Auf die genauen Eigenschaften und Funktionen dieses Peptids wird im
nächsten Abschnitt detailliert eingegangen. Neben Thymosin β4 wurden zahlreiche Thymo-
sin β4 ähnliche Peptide, die sich nur in wenigen Aminosäuren von der Sequenz des Thymo-
sin β4 unterscheiden, in anderen Spezies gefunden [30], wie beispielsweise Thymosin β9
[49] und Thymosin β10 [31]. Thymosin β10 wurde häufig nur zusammen in Gegenwart von
Thymosin β4 in Gewebe gefunden und demnach als dessen Begleitpeptid bezeichnet. Thy-
mosin β10 besitzt vielfältige Funktionen und wird beispielsweise verstärkt in Nierenkarzi-
nomzellen [45] und Melanomzellen [122] exprimiert. Das Peptid soll zudem eine wichtige
Rolle bei der Entwicklung des menschlichen Gehirns spielen [46]. Im Jahr 1992 wurde
schließlich mit Thymosin β12 der erste Vertreter der β-Thymosine bei Nichtsäugern entdeckt
[75]. Wenige Jahre später konnte mit Thymosin β15, das einen potentiellen Marker für diver-
se Krebsarten darstellt, der bisher letzte Vertreter der β-Thymosin Familie ausgemacht
werden [10]. Die Tatsache, dass in einer Spezies bis zu drei verschiedene β-Thymosine je
nach Entwicklungszustand exprimiert werden, deutet daraufhin, dass sich die weiteren ho-
mologen β-Thymosine jeweils in ihren biologischen Eigenschaften unterscheiden.
Generell handelt es sich bei den β-Thymosinen um eine Familie polarer, etwa 5 kDa großer
Peptide mit 40 – 45 Aminosäuren [61]. Alle β-Thymosine sind am N-Terminus acetyliert,
wobei sich die Aminosäuresequenzen häufig nur geringfügig unterscheiden. Gerade diese
über einen langen Evolutionszeitraum konservierte Aminosäuresequenz der β-Thymosine
lässt eine zentrale biologische Rolle dieser Peptide vermuten. Die Abbildung 4 zeigt bei-
spielhaft die Aminosäuresequenzen einzelner Mitglieder der β-Thymosin Familie.

10
Abbildung 4: Aminosäuresequenzen einzelner β-Thymosine (nach Huff, 2001)
1.3. Thymosin β4
1.3.1. Chemische Eigenschaften
Thymosin β4 wurde aus der Thymosin Fraktion 5 im Jahr 1981 isoliert und anschließend se-
quenziert [74]. Es handelt sich dabei um ein am N-Terminus acetyliertes Peptid mit 43
Aminosäuren (11 saure, 9 basische, pI: 4,6) und einem Molekulargewicht von 4963,5 Da
[73]. Durch Oxidation des an der sechsten Position befindlichen Methionins mit millimola-
ren Konzentrationen an H2O2 lässt sich Thymosin β4 in ein entsprechend polareres Thymo-
sin β4 Sulfoxid umwandeln. Diese Umwandlung hat entsprechende funktionelle Konsequen-
zen, wie beispielsweise ein geändertes Bindungsverhalten des Peptids mit Aktin [59]. Bei
der Untersuchung der mRNA von Thymosin β4 wurde festgestellt, dass im primären transla-
tierten Produkt kein Signalpeptid, sondern vielmehr nur ein zusätzlicher Methioninrest vor-
handen ist. Dieser Aminosäurerest stellt das Startcodon für die Translation dar [123]. Thy-
mosin β4 scheint demnach kein sezernierbares Peptid zu sein. Alle β-Thymosine, und damit
auch Thymosin β4, besitzen zudem wichtige Strukturelemente, die eine spätere chemische
Nachweisreaktion ermöglichen. So kann Thymosin β4 aufgrund seiner chemischen Struktur
im UV-Bereich (ca. 210 nm) oder mittels chemischer Derivatisation nachgewiesen werden
[48]. Außerdem sind alle β-Thymosine in 4 M PCA oder in 6 M Guanidiniumhydrochlorid-
lösung löslich und stabil.

11
1.3.2. Struktur
Die Sekundärstruktur von Thymosin β4 wurde mithilfe der Kernresonanzspektroskopie
(NMR) bestimmt. Demnach liegt das Peptid in wässriger Lösung unstrukturiert vor [25],
nimmt jedoch in 60 % Trifluorethanol durch die Ausbildung zweier helikaler Strukturen eine
geordnete Raumordnung an [125]. Die Abbildung 5 verdeutlicht schematisch die Lage dieser
beiden potentiellen α-Helices zwischen den Aminosäureresten 4 – 17 und 30 – 40.
Abbildung 5: Struktur des Thymosin β4 (nach Hannappel, 2007)
Eine weitere strukturelle Besonderheit wurde im Rahmen der Bindung von Thymosin β4 an
G-Aktin beobachtet. Thymosin β4 besitzt ein zu anderen Aktin-bindenden Proteinen ähnlich-
es Aktinbindungsmotiv (LKKTET) [119]. Durch Interaktion mit G-Aktin verhindert es die
Salz-induzierte Polymerisation des G-Aktins zum F-Aktin aus sterisch-geometrischen Grün-
den [48]. Bei dieser Interaktion von Thymosin β4 mit G-Aktin werden die beiden in fluorier-
ten Alkoholen nachweisbaren α-Helices ausgebildet. Da Thymosin β4 erst bei der Interaktion
mit G-Aktin und möglicherweise auch mit anderen Proteinen spezifische Sekundärstrukturen
ausbilden kann, gibt es die Möglichkeit, dass das Peptid mit vielen verschiedenen Bindungs-
partnern interagieren kann. Dieser Prozess wird im englischen Sprachgebrauch als „moon-
lighting“ bezeichnet [65].
1.3.3. Vorkommen
Schon kurz nach der Isolierung von Thymosin β4 aus Kalbsthymus erkannte man, dass das
Peptid nicht allein im Thymus, sondern vielmehr ubiquitär im Gewebe vorkommt [51]. So
konnte Thymosin β4 zunächst in vielen Organgeweben der Maus und der Ratte nachgewie-

12
sen werden (bis zu 0,4 mM), vor allem aber in den Makrophagen. Gleichzeitig fand man he-
raus, dass der Thymus nicht die Quelle dieses Thymosin β4 sein konnte, da Mäuse ohne
Thymus sogar erhöhte Konzentrationen an Thymosin β4 aufwiesen [51]. Im weiteren Ver-
lauf wurde Thymosin β4 in nahezu allen Zellen nachgewiesen [41]. Die einzige Ausnahme
bildeten die Erythrozyten im Blut [50]. Die bisher höchsten Konzentrationen für Thymosin
β4 wurden in polymorphkernigen Leukozyten nachgewiesen. Wie in der Tabelle 1 veran-
schaulicht, tritt Thymosin β4 fast immer gemeinsam mit einem anderen β-Thymosin auf. In
Säugerzellen ist dieses Begleitpeptid in der Regel Thymosin β10 [31]. Die Konzentration
von Thymosin β4 beträgt in diesen Zellen gewöhnlicherweise ca. 70 – 80 % des gesamten β-
Thymosin Gehalts [48]. Thymosin β4 ist generell sowohl im Zellkern als auch im Zytoplas-
ma vorhanden [120], wenngleich es zwischen beiden Kompartimenten nicht zum Austausch
durch einfache Diffusion kommen kann [126].
Spezies Erstes Peptid Zweites Peptid Drittes Peptid
Mensch Tβ4 Tβ10 Tβ15
Ratte Tβ4 Tβ10 Tβ15
Maus, Katze Tβ4 Tβ10
Kalb Tβ4 Tβ9
Kaninchen Tβ4Ala
Tβ10
Pferd, Huhn Tβ4
Xenopus laevis Tβ4Xen
Wal Tβ4
Seeigel Tβ4Seeigel
Tabelle 1: Spezies-spezifisches Vorkommen der β-Thymosine (nach Huff, 2001)
1.3.4. Hormonelle Funktionen
Zu Beginn der Erforschung der Funktionen von Thymosin β4 ging man noch vom hormonel-
len Charakter der Thymosine aus. So waren die ersten beschriebenen Funktionen die Induk-
tion der Expression der terminalen Desoxynukleotidtransferase in T-Lymphozyten [74] und
die Stimulation der Freisetzung des Lutein-Releasing-Faktors aus dem Hypothalamus [93].
Außerdem konnte nachgewiesen werden, dass Thymosin β4 die Makrophagenmigration inhi-
biert [115].

13
1.3.5. Intrazelluläre Funktionen
Gerade wegen der hohen intrazellulären Konzentration und des ubiquitären Vorkommens
von Thymosin β4 wurde bereits früh spekuliert, dass das Peptid wichtige intrazelluläre Funk-
tionen im Rahmen des Zytoskeletts ausüben könnte [41]. Diese Vermutung wurde dann im
Jahr 1990 bestätigt, als Safer et al. in humanen Thrombozyten erstmals einen Aktin-seques-
trierenden Faktor mit einer Masse von 5 kDa entdeckten [101]. Dieser zunächst Fx genannte
Faktor ging über ein spezifisches Bindungsmotiv eine 1 : 1 Bindung mit unpolymerisierten
G-Aktin ein und verhinderte dadurch dessen Salz-induzierte Polymerisation zum F-Aktin.
Kurze Zeit später stellte sich heraus, dass die Sequenz dieses Faktors Fx identisch mit jener
von Thymosin β4 ist [100]. Da das intrazelluläre Zytoskelett an einer Vielzahl zellulärer Ak-
tivitäten wie Lokomotion, Zytokinese, intrazelluläre Transportprozesse, Phagozytose und
Exozytose beteiligt ist [113], war Thymosin β4 als das bedeutendste intrazelluläre Aktin-se-
questrierende Peptid ab sofort von großer Bedeutung. Es gewährleistete durch die Bereitstel-
lung eines großen Pools an monomerem G-Aktin erst jene zelltypische Aktindynamik. Die
Abbildung 6 zeigt schematisch die Aktin-sequestrierende Rolle des Thymosin β4.
Abbildung 6: Thymosin β4 als Haupt-Aktin-sequestrierendes Protein (nach Huff, 2001)
Im weiteren Verlauf wurde die Interaktion von Thymosin β4 und Aktin noch detaillierter
untersucht. Man fand heraus, dass Thymosin β4 nicht an F-Aktin bindet [121], dass alle β-
Thymosine ebenfalls Aktin sequestrieren, wenngleich mit unterschiedlichen Kd-Werten [48],
dass die Oxidation von Thymosin β4 zum Sulfoxid zu einer 18fach erhöhten Dissoziations-
konstante des Aktin-Thymosin β4-Komplexes führt [59] und dass DNaseI mit Thymosin β4
um die Bindung an G-Aktin konkurriert [9].

14
1.3.6. Extrazelluläre Funktionen
In den folgenden Jahren konzentrierte man sich nun auf die Erforschung möglicher extrazel-
lulärer Wirkungen von Thymosin β4 und machte dabei eine Vielzahl von Entdeckungen.
Thymosin β4 scheint extrazellulär zytokinähnliche Eigenschaften zu besitzen. Extrazellulär
appliziertes Thymosin β4 führt zu einer verstärkten Anheftung von Endothelzellen an Ma-
trixkomponenten [81]. Es stimuliert die Migration von Endothelzellen der Nabelschnurvene
[77] und dient außerdem als Glutaminylsubstrat für Transglutaminasen [58]. Diese Transfe-
rasen vernetzen Thymosin β4 selektiv mit Fibrin oder Kollagen, was funktionell von großer
Bedeutung sein kann. Damit könnte beispielsweise erklärt werden, warum Thymosin β4 als
kleines, extrem gut wasserlösliches Peptid im Extrazellularraum funktionell wirksame Kon-
zentrationen erreichen kann, ohne z.B. durch den Blutstrom sofort verdünnt zu werden.
Nachdem mit diesen ersten Studien das chemotaktische und angiogene Wirkungspotential
des Thymosin β4 nachgewiesen werden konnte, entdeckte Young eine anti-inflammatorische
Wirkung eines Thymosin β4 Derivats. Er zeigte, dass das Sulfoxid die Chemotaxis von neu-
trophilen Granulozyten verhindert und bezeichnete das Sulfoxid folglich als extrazellulär ak-
tive Form des Thymosin β4 [124]. Die Oxidation scheint damit eine Möglichkeit zur Regula-
tion der extrazellulären Funktion von Thymosin β4 zu sein. Wenig später wurde Thymosin
β4 als bedeutender Faktor für die Wundheilung identifiziert, indem nachgewiesen werden
konnte, dass das Peptid den Wundheilungsverlauf über zahlreiche Mechanismen positiv be-
einflusst [78]. Somit wurde Thymosin β4 zum ersten Mal für medikamentöse Anwendungen
in der Klinik interessant. Thymosin β4 induziert auch die Expression von Matrixmetallopro-
teinasen im Bindegewebe [90] und ist an der Wundheilung der Kornea beteiligt, indem es
zahlreiche Entzündungsmediatoren beeinflusst [109]. Die anti-inflammatorische Wirkung
von Thymosin β4 auf Zytokine und Chemokine wurde in weiteren Studien noch eingehender
untersucht und bestätigt [110] [8]. Wenn Thymosin β4 endogen überexprimiert wird, stimu-
liert es jedoch auch das Tumorwachstum und die Metastasenbildung durch Induktion der
Zellmigration und einer VEGF vermittelten Angiogenese [21]. Es konnte des Weiteren nach-
gewiesen werden, dass Thymosin β4 das Haarwachstum durch Induktion der Stammzellpro-
liferation fördert [89] und zudem kardioprotektive Eigenschaften durch die Förderung der
Migration und der Reparatur von Kardiomyozyten nach einem Myokardinfarkt besitzt [111].
Es ist mit großer Sicherheit zu erwarten, dass durch zukünftige Forschung noch weitere inte-
ressante extrazelluläre Funktionen des Thymosin β4 entdeckt werden, und dass damit wei-
tere klinische Anwendungsmöglichkeiten für Thymosin β4 entwickelt werden. Es ist jedoch
nach wie vor unklar wie intrazellulär vorkommendes Thymosin β4 in den extrazellulären
Raum gelangen kann, da Thymosin β4 wie bereits erwähnt kein Signalpeptid besitzt [123].
Möglicherweise gibt es keinen spezifischen Weg der Thymosinfreisetzung aus Zellen. Eine
Freisetzung könnte jedoch durch eine Störung des Zytoskeletts [19] oder einfach durch Zell-

15
lyse bedingt sein. Die Abbildung 7 zeigt zusammenfassend eine Auswahl der gegenwärtig
bekannten intra- und extrazellulären Funktionen des Thymosin β4.
Abbildung 7: Biologische Eigenschaften des Thymosin β4 (nach Goldstein, 2007)
1.3.7. Thymosin β4 in Körperflüssigkeiten
Blut wurde als erste Körperflüssigkeit auf den Gehalt an Thymosin β4 untersucht [50]. Die
Konzentration von Thymosin β4 im Blut beträgt durchschnittlich 15 mg/l, wohingegen Se-
rum mit 0,45 – 1,1 µg/l weniger als 0,01 % der Thymosin β4 Menge des Gesamtblutes ent-
hält. Plasma enthält mit 0,03 – 0,4 mg/l hingegen eine deutlich höhere Konzentration an
Thymosin β4. Die Konzentration von freiem Thymosin β4 im Blut beträgt somit insgesamt
ca. 10 – 200 nM (Plasma und Serum). Dieser Wert ist verglichen mit Insulin (bis zu 140 pM)
oder Glukagon (bis zu 60 pM) sehr hoch [79]. Die Art der Freisetzung von Thymosin β4 aus
den Blutzellen ist jedoch nach wie vor unklar. Thymosin β4 könnte aus Thrombozyten nach
Aktivierung mit Thrombin freigesetzt werden und mittels Faktor XIIIa an Fibrin oder Kolla-
gen geknüpft werden [62]. Die Tabelle 2 gibt einen Überblick über die gefundenen Konzen-
trationen an Thymosin β4 im Blut.
Thymosin
β4
Schutz des Mykords und
Förderung der Gefäß-neubildung
Stimulation der Entwick-
lung und Regeneration von Nerven-
fasern
Aktinse-questrierung, Beschleuni-
gung von Wundheilungund Angio-
genese
Blockade der Apoptose
Modulation von Zyto-kinen und
Chemokinen
Hochregula-tion der
Expression bestimmter
Gene
Chemopro-tektive und
anti-mikrobi-elle Eigen-
schaften
Modulation spezifischer Proteasen

16
Thymosin β4
Probe Wertebereich [mg/l] Wertebereich
[fg/Zelle]
Gesamtblut 12,0 – 19,5
Plasma 0,026 – 0,387
Mononukleäre Leukozyten 183 – 380
Polymorphkernige Leukozyten 269 – 564
Thrombozyten 6,9 – 31,7
Erythrozyten Nicht nachweisbar
Tabelle 2: Thymosin β4 Konzentrationen im Blut (nach Hannappel, 2007)
Des Weiteren wurde Thymosin β4 noch in Tränenflüssigkeit, im Gesamtspeichel [7], und in
Wundflüssigkeit [16] mittels eines ELISA-Tests nachgewiesen und quantifiziert. Tränenflüs-
sigkeit enthielt durchschnittlich 0,5 – 7 µg/ml und Wundflüssigkeit 20 mg/l Thymosin β4. Es
konnten jedoch keine eindeutigen Schlüsse aus den gefundenen Konzentrationen gezogen
werden. Aufgrund der teilweise sehr hohen Konzentrationen bleibt jedoch zu vermuten, dass
dem Thymosin β4 in Körperflüssigkeiten eine funktionell entscheidende Rolle zukommt.
1.4. Mundhöhle
1.4.1. Topografie und Gliederung
Die Mundhöhle ist Bestandteil des Kopfdarmes, der sich funktionell in drei Abschnitte glie-
dert. Von kranial nach kaudal besteht dieser aus dem Nasopharynx (pars nasalis pharyngis),
dem Mesopharynx (pars oralis pharyngis) und dem Hypopharynx (pars laryngea pharyngis)
[98]. Die Mundhöhle liegt ventral des Mesopharynx und ist von ihm durch den weichen
Gaumen, die tonsilla palatina, die raphe pterygomandibulare und die papillae circumvalla-
tae der Zunge abgegrenzt [69]. Anatomische Bestandteile der Mundhöhle sind die Lippen,
die gingivobuccale Mukosa, die oberen und unteren Alveolarfortsätze mit den Zähnen, die
mandibula, die vorderen zwei Drittel der Zunge und der harte Gaumen. Die Abbildung 8
veranschaulicht die Gliederung des Kopfdarmes.
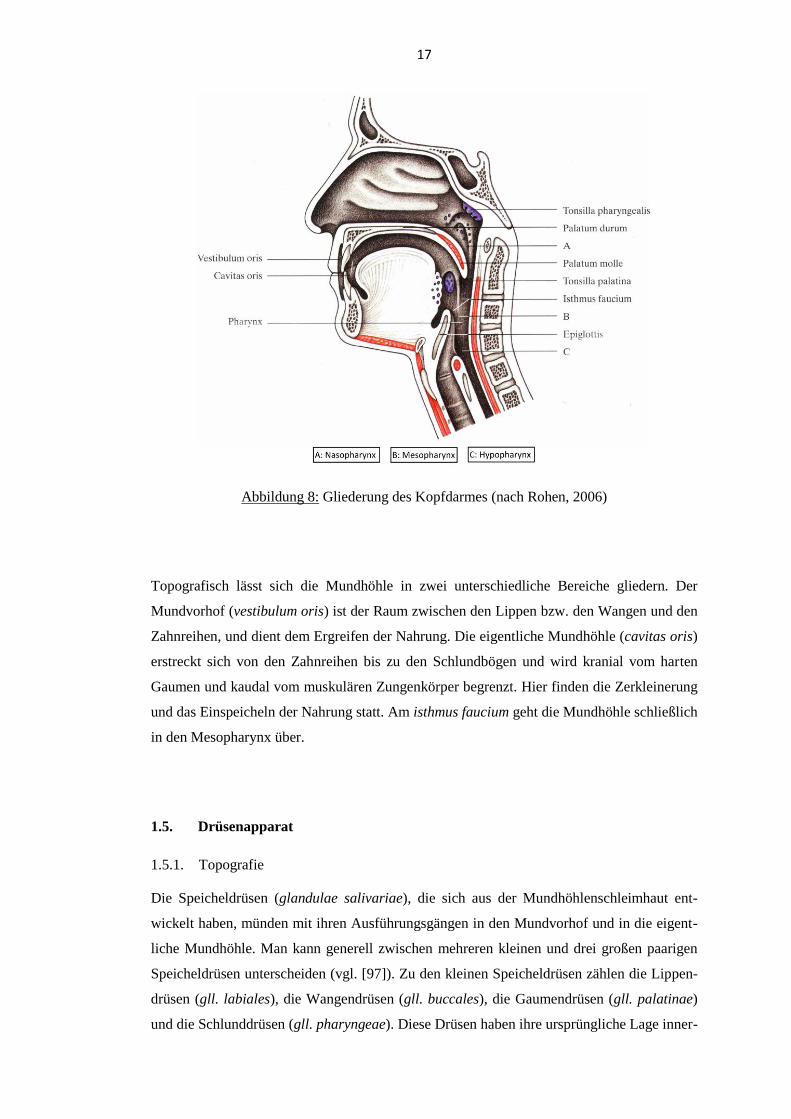
17
Abbildung 8: Gliederung des Kopfdarmes (nach Rohen, 2006)
Topografisch lässt sich die Mundhöhle in zwei unterschiedliche Bereiche gliedern. Der
Mundvorhof (vestibulum oris) ist der Raum zwischen den Lippen bzw. den Wangen und den
Zahnreihen, und dient dem Ergreifen der Nahrung. Die eigentliche Mundhöhle (cavitas oris)
erstreckt sich von den Zahnreihen bis zu den Schlundbögen und wird kranial vom harten
Gaumen und kaudal vom muskulären Zungenkörper begrenzt. Hier finden die Zerkleinerung
und das Einspeicheln der Nahrung statt. Am isthmus faucium geht die Mundhöhle schließlich
in den Mesopharynx über.
1.5. Drüsenapparat
1.5.1. Topografie
Die Speicheldrüsen (glandulae salivariae), die sich aus der Mundhöhlenschleimhaut ent-
wickelt haben, münden mit ihren Ausführungsgängen in den Mundvorhof und in die eigent-
liche Mundhöhle. Man kann generell zwischen mehreren kleinen und drei großen paarigen
Speicheldrüsen unterscheiden (vgl. [97]). Zu den kleinen Speicheldrüsen zählen die Lippen-
drüsen (gll. labiales), die Wangendrüsen (gll. buccales), die Gaumendrüsen (gll. palatinae)
und die Schlunddrüsen (gll. pharyngeae). Diese Drüsen haben ihre ursprüngliche Lage inner-

18
halb der Schleimhaut beibehalten, wohingegen sich die großen Speicheldrüsen teilweise weit
von ihrem Entstehungsort in die Umgebung verlagert haben und ihr Sekret über lange Aus-
führungsgänge in die Mundhöhle abgeben.
Die paarige Ohrspeicheldrüse (gl. parotis), als die größte und wichtigste Speicheldrüse, liegt
auf dem musculus masseter und schiebt sich vor dem Ohr hinter den ramus mandibulae bis
in die Tiefe der fossa retromandibularis. Ihr Ausführungsgang (ductus parotideus, Stensen-
Gang) überkreuzt lateral den musculus masseter, durchdringt den musculus buccinator sowie
die Wangenschleimhaut und endet gegenüber des zweiten oberen Molaren im vestibulum
oris. Die gl. parotis wird von einer derben Faszie umhüllt und von Ästen des nervus facialis
in Form des plexus intraparotideus durchzogen.
Die Unterkieferdrüse (gl. submandibularis) liegt kaudal auf dem diaphragma oris in dem
Raum zwischen Unterkiefer und den beiden Bäuchen des musculus digastricus. Der Ausfüh-
rungsgang (ductus submandibularis) biegt um den hinteren Rand des musculus mylohyoideus
herum und endet unter der Zunge auf der caruncula sublingualis, die sich seitlich beidseits
des Zungenbändchens befindet.
Die dritte große paarige Speicheldrüse ist die Unterzungendrüse (gl. sublingualis). Diese
liegt kranial auf dem diaphragma oris und wölbt sich als plica sublingualis in die Mund-
höhle vor. Der Drüsenkörper besteht im hinteren Abschnitt aus zahlreichen Einzeldrüsen, die
eigene auf der plica sublingualis ausmündende kleine Ausführungsgänge (ductus sublingu-
ales minores) besitzen. Der vordere Abschnitt hingegen leitet sein Sekret über einen Ausfüh-
rungsgang (ductus sublingualis major), der ebenfalls auf der caruncula sublingualis mündet,
in die Mundhöhle. Die gl. sublingualis und gl. submandibularis drainieren ihr Sekret jedoch
auch häufig über einen gemeinsamen Ausführungsgang (Wharton-Gang). Dies kann eine
drüsenspezifische Speichelgewinnung erschweren. Die Abbildung 9 gibt eine Übersicht über
die topografische Lage der Speicheldrüsen.

19
Abbildung 9: Topografie des Drüsenapparates (nach Rohen, 2006)
1.5.2. Histologischer Aufbau der Speicheldrüsen und Mechanismus der Speichelsekretion
Speicheldrüsen besitzen generell einen charakteristischen histologischen Aufbau. Ihr Drü-
senparenchym wird durch Bindegewebssepten in Läppchen gegliedert und ist aus zwei ana-
tomisch und funktionell unterschiedlichen Anteilen zusammengesetzt, in denen der Speichel
auf unterschiedliche Art gebildet wird (vgl. [118]).
In den Endstücken findet der erste Schritt der Speichelbildung statt. Diese bestehen aus un-
tereinander verbundenen Azinuszellen, die in der basolateralen Membran verschiedenste Io-
nentransporter und in der lumenwärts gelegenen Membran unterschiedliche Ionenkanäle be-
sitzen. Unter dem Einfluss von Neurotransmittern des Parasympathikus bzw. Sympathikus
[14] wird von den Azinuszellen aus der interstitiellen Flüssigkeit zunächst ein Primärspei-
chel gebildet. Die wasserdurchlässigen Azinuszellen sorgen mittels ihrer Ionentransportme-
chanismen für eine quantitativ vollständige Flüssigkeitssekretion und eine plasmaisotone Zu-
sammensetzung des Primärspeichels [66]. Dabei werden vor allem Na+- und Cl
--Ionen als
Elektrolyte sezerniert. Die Azinuszellen produzieren und sezernieren zudem 85 – 90 % der
Proteine im Speichel [13]. Man unterscheidet aufgrund des zellulären Aufbaus zwischen se-
rösen und mukösen Azinuszellen. Seröse Azinuszellen produzieren einen dünnflüssigen fer-
mentreichen Speichel, muköse Azinuszellen hingegen einen dickflüssigen muzinreichen
Speichel, der die Gleitfähigkeit der Nahrung sicherstellen soll. Da bei den verschiedenen

20
Speicheldrüsen oft eine Art von Azinuszellen überwiegt, teilt man die Speicheldrüsen ent-
sprechend in seröse, muköse oder gemischte Drüsen, bei denen entweder die serösen („sero-
mukös“) oder mukösen („muko-serös“) Endstücke überwiegen, ein. Nach dieser Einteilung
bezeichnet man die kleinen Speicheldrüsen allesamt als mukös, die gl. parotis als rein serös,
die gl. submandibularis als sero-mukös und die gl. sublingualis als muko-serös [28].
An die Endstücke schließt sich das Ausführungsgangsystem an. Dieses besteht aus Schalt-
stücken und Streifenstücken und endet in einem Ausführungsgang, der das Sekret schließlich
in die Mundhöhle drainiert. Der von den Endstücken gebildete Primärspeichel wird beim
Durchfluss durch das Ausführungsgangsystem in seiner Zusammensetzung mittels verschie-
denster Ionentransportmechanismen sekretorisch und resorptiv modifiziert [118], sodass der
Sekundärspeichel hypoton wird. Die Gangepithelzellen resorbieren bei relativ geringer Was-
serpermeabilität Na+- und Cl
--Ionen aus dem Lumen und sezernieren kleine Mengen an K
+-
und HCO3--Ionen [118]. Es werden zudem noch zusätzliche Proteine sezerniert. Die Abbil-
dung 10 veranschaulicht die Ionentransportprozesse in den Azinus- und Gangepithelzellen.
Abbildung 10: Ionenaustauschprozesse in der Azinuszelle (A) und in der Gangepithel-
zelle (B) (nach Thews, 2005)
21
1.5.3. Speichelzusammensetzung
Speichel besteht zu 99 % aus Wasser. Organische und anorganische Moleküle bilden das
restliche eine Prozent [71]. Der pH-Wert von Speichel liegt normalerweise zwischen 6 und
7, kann aber auch Werte von 5,3 bei niedriger und 7,8 bei hoher Speichelfließrate annehmen
[63]. Es werden täglich zwischen 1000 und 1500 ml Speichel sezerniert. Das Gesamtvolu-
men ist jedoch sehr variabel und kann durch diverse Stimuli beeinflusst werden [63]. Die
einzelnen Speicheldrüsen tragen außerdem in unterschiedlichem Ausmaß zum Gesamtspei-
chelvolumen bei. Insgesamt werden ca. 93 % des Volumens von den großen Speicheldrüsen
und 7 % von den zahlreichen kleinen Speicheldrüsen produziert [71]. Im Ruhezustand pro-
duziert die gl. submandibularis 70 %, die gl. parotis 25 % und die gl. sublingualis 5 % des
Gesamtspeichels. Nach einer entsprechenden Stimulation der Drüsen verändern sich diese
Werte zu 63 %, 34 % bzw. zu 3 % [114]. Demzufolge muss man in quantitativer aber auch in
qualitativer Hinsicht zwischen Ruhespeichel und Reizspeichel unterscheiden. Reizspeichel
wird infolge einer Beeinflussung des autonomen Nervensystems durch mechanische, gusta-
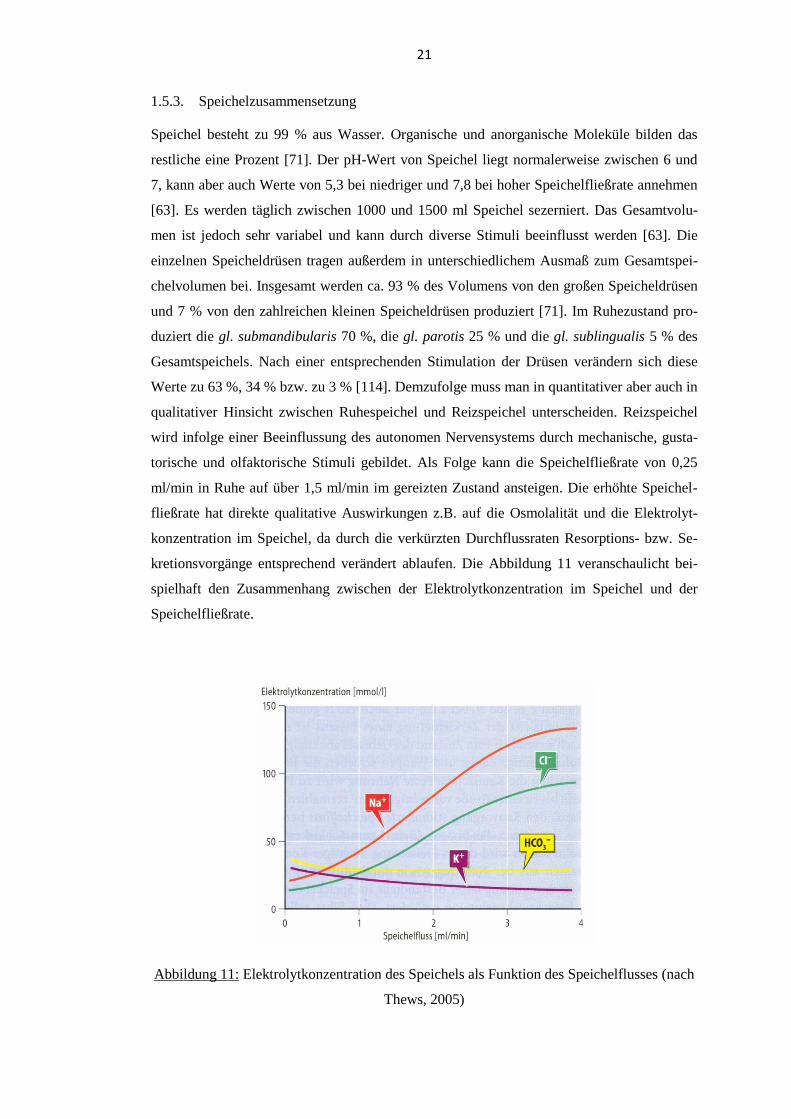
torische und olfaktorische Stimuli gebildet. Als Folge kann die Speichelfließrate von 0,25
ml/min in Ruhe auf über 1,5 ml/min im gereizten Zustand ansteigen. Die erhöhte Speichel-
fließrate hat direkte qualitative Auswirkungen z.B. auf die Osmolalität und die Elektrolyt-
konzentration im Speichel, da durch die verkürzten Durchflussraten Resorptions- bzw. Se-
kretionsvorgänge entsprechend verändert ablaufen. Die Abbildung 11 veranschaulicht bei-
spielhaft den Zusammenhang zwischen der Elektrolytkonzentration im Speichel und der
Speichelfließrate.
Abbildung 11: Elektrolytkonzentration des Speichels als Funktion des Speichelflusses (nach
Thews, 2005)

22
Generell gilt jedoch, dass nicht nur die Elektrolytkonzentration, sondern auch die Konzentra-
tion der von den Speicheldrüsen sezernierten Makromoleküle vom Grad des Speichelflusses
abhängt. Zu den Makromolekülen zählen beispielsweise α-Amylase, Glykoproteine, Muzine,
Lysozym, Laktoferrin, Immunglobulin A, Haptocorrine und häufig auch Blutgruppenantige-
ne [114]. Die einzelnen Speicheldrüsen zeigen zudem Unterschiede im Sekretionsverhalten
dieser Makromoleküle. So wird beispielsweise α-Amylase eher von der gl. parotis, Muzine
dagegen bevorzugt von der gl. sublingualis und gl. submandibularis sezerniert [114]. Damit
ergeben sich für die Analyse von Speichel wichtige Konsequenzen, da sich Drüsenspeichel
von Gesamtspeichel und Ruhespeichel von Reizspeichel in der Zusammensetzung unter-
scheiden. Des Weiteren unterscheiden sich Speichelfließraten interindividuell. Es gibt aber
auch intraindividuelle Unterschiede, je nach den Bedingungen, unter denen Speichel gewon-
nen wird [27]. So müssen Parameter wie die Jahreszeit [104], die Tageszeit [26] und die
Körperposition [105] beachtet werden, da sie alle Einfluss auf die Speichelsekretion haben.
Es ist daher von großer Wichtigkeit eine Methode zur Speichelprobengewinnung zu standar-
disieren [84]. Zuletzt gilt es noch zu beachten, dass Gesamtspeichel auch Nichtspeichelbe-
standteile wie desquamierte Epithelzellen, Nahrungsbestandteile, Bakterien, Sulkusfluid und
Leukozyten enthalten kann [84].
1.5.4. Speichelfunktionen
Für die Funktionen des Speichels sind sowohl dessen Quantität als auch dessen Qualität von
entscheidender Bedeutung (vgl. [66]). Insgesamt gesehen erfüllt der Speichel zunächst eine
Reihe mechanischer Funktionen. Er erleichtert das Sprechen durch Feuchthalten der Mund-
höhle, leitet mittels Quellungsvorgängen und Einspeicheln bzw. Gleitfähigmachen der Nah-
rungsbestandteile die Verdauung ein und bildet das Lösungsmittel für Geschmacksstoffe.
Speichel dient aber auch der Selbstreinigung der Mundhöhle, indem Speisereste ausgespült
werden. Dieser Effekt wird mit dem Begriff „oral sugar clearance“ umschrieben [27]. Spei-
chel ist aber auch in fermentativer Hinsicht von Bedeutung, da in ihm enthaltene Enzyme
(z.B. α-Amylase) die Spaltung von Kohlenhydraten bereits in der Mundhöhle einleiten.
Einer Reihe von Bestandteilen im Speichel kommen außerdem spezifische biologische Funk-
tionen zu. So haben Lysozym, Laktoferrin und Immunglobulin A eine anti-bakterielle Wir-
kung. Bikarbonat, Phosphate und Proteine gewährleisten eine große Pufferkapazität und da-
mit eine gewisse Schutzfunktion für die Zähne und die orale Mukosa. Der hohe Gehalt an
Kalzium- und Phosphationen im Speichel ermöglicht zudem bis zu einem gewissen Ausmaß
eine Remineralisation eines durch Karies demineralisierten Zahnschmelzes. Die Tabelle 3

23
gibt einen zusammenfassenden Überblick über die Funktionen des Speichels und die dafür
verantwortlichen Speichelbestandteile.
Funktion Bestandteile
Gleitfähigmachen des Speisebolus Muzin, Prolin-reiche Glykoproteine, Wasser
Anti-mikrobielle Wirkung Lysozym, Laktoferrin, Laktoperoxidase, Ig A
Aufrechterhaltung der
Schleimhautintegrität
Muzin, Elektrolyte, Wasser
Spülfunktion Wasser
Pufferkapazität und Remineralisation Bikarbonat, Phosphat, Proteine, Fluoride,
Kalzium
Verdauung Amylase, Lipase, Protease, Wasser, Muzin
Geschmack Wasser, Gustin
Phonetik Wasser, Muzin
Tabelle 3: Speichelfunktionen und Speichelbestandteile (nach Llena-Puy, 2006)
1.6. Zahnhalteapparat
1.6.1. Anatomischer Aufbau
Der Zahnhalteapparat wird auch als Parodont oder Parodontium bezeichnet (par (gr.) = he-
rum; odontos (gr.) = Zahn) (vgl. [53]). Er stellt ein funktionelles System unterschiedlicher
Gewebegruppen dar und bettet den Zahn strukturell im Alveolarknochen des Kiefers ein.
Das Parodont setzt sich im Einzelnen aus dem Zahnfleisch (Gingiva), dem Zement, der Wur-
zelhaut (Desmodont oder Periodont) und dem angrenzenden Alveolarknochen zusammen.
Jeder dieser Komponenten besitzt eine spezielle Struktur, die wiederum unmittelbar dessen
Funktion bestimmt. Die korrekte physiologische Funktion des Parodonts wird insgesamt je-
doch erst durch das Zusammenspiel der einzelnen Komponenten untereinander und der Auf-
rechterhaltung der strukturellen Integrität des Systems erreicht [11]. Die Hauptaufgaben des
Parodonts sind die Verankerung der Zähne im Knochen, die Dämpfung der auftretenden
Kaukräfte, die Abwehr äußerer Noxen und die Trennung zwischen Mundhöhlenmilieu und
Zahnwurzel [53]. Die Abbildung 12 zeigt eine schematische Übersicht über die Bestandteile
des Parodonts.

24
Abbildung 12: Aufbau des Parodonts (nach Hellwig, 2007)
Die Kenntnis des Aufbaus und der Funktion der einzelnen Bestandteile des Parodonts ist
eine wichtige Voraussetzung für das Verständnis pathologischer Veränderungen und deren
Therapie, weswegen die einzelnen Bestandteile zunächst genauer dargestellt werden sollen.
1.6.2. Gingiva
1.6.2.1. Anatomischer Aufbau
Makroskopisch lässt sich die Gingiva in die freie marginale Gingiva und die befestigte Gin-
giva einteilen (vgl. [53]). Die freie Gingiva verläuft girlandenförmig als 0,8 – 2,5 mm breiter
Saum koronal der Schmelz-Zement-Grenze. Sie besitzt eine blassrosa Farbe, eine gestippelte
Oberfläche und geht vestibulär in Höhe der Schmelz-Zement-Grenze an der gingivalen Fur-
che in die 1 – 9 mm breite befestigte Gingiva über. Diese ist über Bindegewebsfasern fest
mit dem Alveolarknochen und dem Wurzelzement verbunden und demnach nicht gegen ihre
Unterlage verschiebbar. Wie die freie Gingiva besitzt sie ebenfalls eine blassrosa Farbe und
eine gestippelte Oberfläche. Die befestigte Gingiva geht schließlich an der linea girlandifor-
mis in die das Vestibulum auskleidende dunkelrote, nicht keratinisierte Mukosa über. Mi-
kroskopisch betrachtet kann man verschiedene Arten des Gingivaepithels unterscheiden (vgl.
[53]). Das der Mundhöhle zugewandte orale Epithel und das dem Zahn zugewandte orale
Sulkusepithel sind mehrschichtig verhornte Epithelien, die zapfenartig mit dem darunter lie-
genden Bindegewebe verbunden sind. Die Zellumsatzrate dieser beiden Epithelien beträgt
zehn bis zwölf Tage. Das orale Sulkusepithel geht apikal in das Saumepithel über. Dieses
unverhornte Epithel liegt kragenförmig um den Zahn herum und reicht vom Boden des Gin-
givasulkus bis zur Schmelz-Zement-Grenze. Es ist mit dem benachbarten Bindegewebe nicht
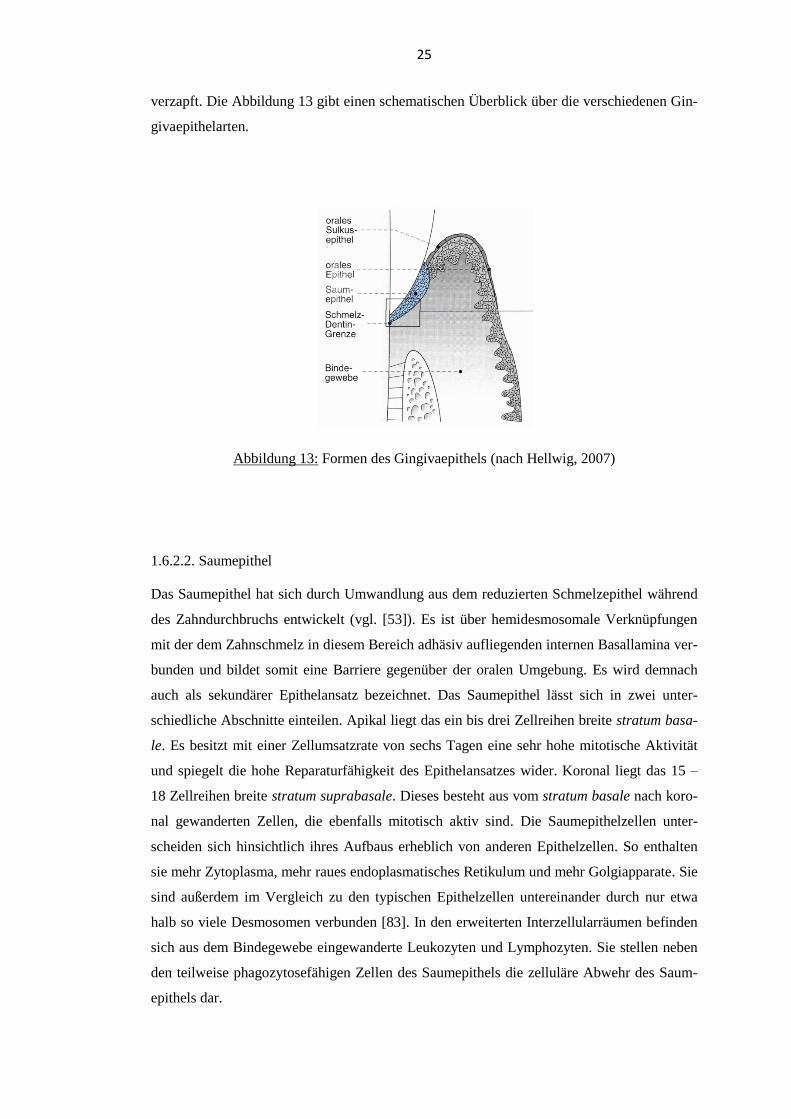
25
verzapft. Die Abbildung 13 gibt einen schematischen Überblick über die verschiedenen Gin-
givaepithelarten.
Abbildung 13: Formen des Gingivaepithels (nach Hellwig, 2007)
1.6.2.2. Saumepithel
Das Saumepithel hat sich durch Umwandlung aus dem reduzierten Schmelzepithel während
des Zahndurchbruchs entwickelt (vgl. [53]). Es ist über hemidesmosomale Verknüpfungen
mit der dem Zahnschmelz in diesem Bereich adhäsiv aufliegenden internen Basallamina ver-
bunden und bildet somit eine Barriere gegenüber der oralen Umgebung. Es wird demnach
auch als sekundärer Epithelansatz bezeichnet. Das Saumepithel lässt sich in zwei unter-
schiedliche Abschnitte einteilen. Apikal liegt das ein bis drei Zellreihen breite stratum basa-
le. Es besitzt mit einer Zellumsatzrate von sechs Tagen eine sehr hohe mitotische Aktivität
und spiegelt die hohe Reparaturfähigkeit des Epithelansatzes wider. Koronal liegt das 15 –
18 Zellreihen breite stratum suprabasale. Dieses besteht aus vom stratum basale nach koro-
nal gewanderten Zellen, die ebenfalls mitotisch aktiv sind. Die Saumepithelzellen unter-
scheiden sich hinsichtlich ihres Aufbaus erheblich von anderen Epithelzellen. So enthalten
sie mehr Zytoplasma, mehr raues endoplasmatisches Retikulum und mehr Golgiapparate. Sie
sind außerdem im Vergleich zu den typischen Epithelzellen untereinander durch nur etwa
halb so viele Desmosomen verbunden [83]. In den erweiterten Interzellularräumen befinden
sich aus dem Bindegewebe eingewanderte Leukozyten und Lymphozyten. Sie stellen neben
den teilweise phagozytosefähigen Zellen des Saumepithels die zelluläre Abwehr des Saum-
epithels dar.

26
Aus der histologischen Struktur des Saumepithels ergeben sich funktionell wichtige Konse-
quenzen. Zunächst gewährleistet die Intaktheit des Saumepithels den physiologisch gesun-
den Zustand des Parodonts. Parodontalerkrankungen treten dann auf, wenn die Struktur des
Saumepithels zu zerfallen beginnt [83]. Die lockere Struktur des Saumepithels ist zudem ent-
scheidend für die schnelle Penetration sowohl von externen Noxen als auch von am Sulkus-
boden austretendem Sulkusfluid.
1.6.2.3. Bindegewebe
Das gingivale Bindegewebe setzt sich hauptsächlich aus Bindegewebsfasern, Fibroblasten
und Proteoglykanen zusammen (vgl. [53]). Die Bindegewebsfasern ordnen sich zu Faserbün-
deln an, die entsprechend ihrer Verlaufsrichtung unterschieden werden können. So gibt es
dentogingivale, dentoperiostale, zirkuläre, semizirkuläre, transseptale, transgingivale, inter-
gingivale, alveologingivale und interpapilläre Fasern. Neben Fibroblasten gibt es im Binde-
gewebe noch viele Abwehrzellen, wie z.B. polymorphkernige Granulozyten, Monozyten und
Lymphozyten.
Das gingivale Bindegewebe zeigt je nach dem von ihm unterstützten Epithel strukturelle Un-
terschiede [83]. Das dem Saumepithel benachbarte Bindegewebe besitzt als Besonderheit
eine zellreiche Zone mit zahlreichen Makrophagen und zudem einen ausgeprägten anasto-
mosierenden Gefäßplexus, der die Quelle des Sulkusfluids darstellt. Aus diesem Plexus tre-
ten ständig Entzündungszellen aus, die anschließend durch das Saumepithel in Richtung Sul-
kus wandern. Selbst unter normalen klinischen Bedingungen ist das dem Saumepithel be-
nachbarte Bindegewebe leicht entzündet [6]. Bei entzündlichen Reaktionen wird der Plexus
äußerst permeabel.
1.6.2.4. Gingivaler Sulkus und Sulkusfluid
Die Vertiefung zwischen dem oralen Sulkusepithel und der Zahnoberfläche wird als gingiva-
ler Sulkus bezeichnet (vgl. [53]). Der Sulkus wird apikal durch die freie Oberfläche des
Saumepithels begrenzt und hat bei einem physiologisch intakten Parodont eine Tiefe von 0,1
– 0,5 mm. Am Sulkusboden tritt eine klare Flüssigkeit aus, die als Sulkusflüssigkeit oder
Sulkusfluid bezeichnet wird. Obwohl man von der Existenz des Sulkusfluids seit mehr als
100 Jahren weiß [15], wurde dessen Ursprung und Zusammensetzung jahrelang kontrovers
diskutiert.
27
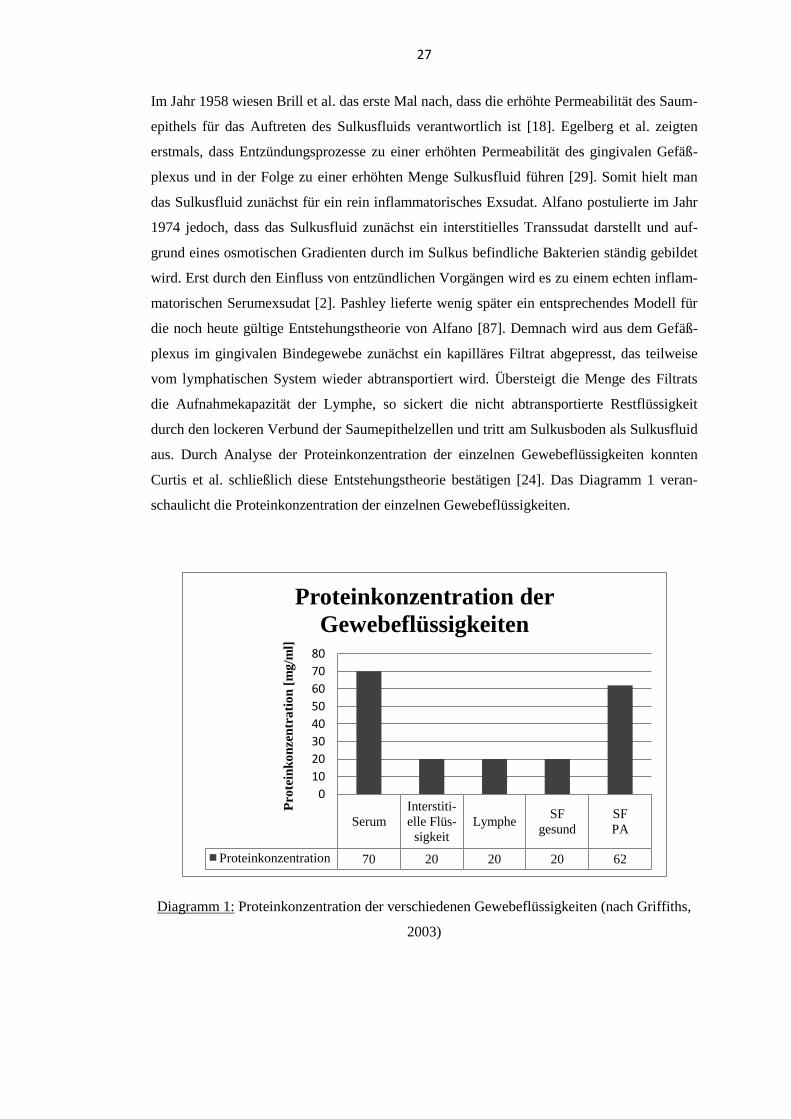
Im Jahr 1958 wiesen Brill et al. das erste Mal nach, dass die erhöhte Permeabilität des Saum-
epithels für das Auftreten des Sulkusfluids verantwortlich ist [18]. Egelberg et al. zeigten
erstmals, dass Entzündungsprozesse zu einer erhöhten Permeabilität des gingivalen Gefäß-
plexus und in der Folge zu einer erhöhten Menge Sulkusfluid führen [29]. Somit hielt man
das Sulkusfluid zunächst für ein rein inflammatorisches Exsudat. Alfano postulierte im Jahr
1974 jedoch, dass das Sulkusfluid zunächst ein interstitielles Transsudat darstellt und auf-
grund eines osmotischen Gradienten durch im Sulkus befindliche Bakterien ständig gebildet
wird. Erst durch den Einfluss von entzündlichen Vorgängen wird es zu einem echten inflam-
matorischen Serumexsudat [2]. Pashley lieferte wenig später ein entsprechendes Modell für
die noch heute gültige Entstehungstheorie von Alfano [87]. Demnach wird aus dem Gefäß-
plexus im gingivalen Bindegewebe zunächst ein kapilläres Filtrat abgepresst, das teilweise
vom lymphatischen System wieder abtransportiert wird. Übersteigt die Menge des Filtrats
die Aufnahmekapazität der Lymphe, so sickert die nicht abtransportierte Restflüssigkeit
durch den lockeren Verbund der Saumepithelzellen und tritt am Sulkusboden als Sulkusfluid
aus. Durch Analyse der Proteinkonzentration der einzelnen Gewebeflüssigkeiten konnten
Curtis et al. schließlich diese Entstehungstheorie bestätigen [24]. Das Diagramm 1 veran-
schaulicht die Proteinkonzentration der einzelnen Gewebeflüssigkeiten.
Diagramm 1: Proteinkonzentration der verschiedenen Gewebeflüssigkeiten (nach Griffiths,
2003)
Serum
Interstiti-
elle Flüs-
sigkeit
LympheSF
gesund
SF
PA
Proteinkonzentration 70 20 20 20 62
0
10
20
30
40
50
60
70
80
Pro
tein
ko
nze
ntr
ati
on
[m
g/m
l]
Proteinkonzentration der
Gewebeflüssigkeiten
28
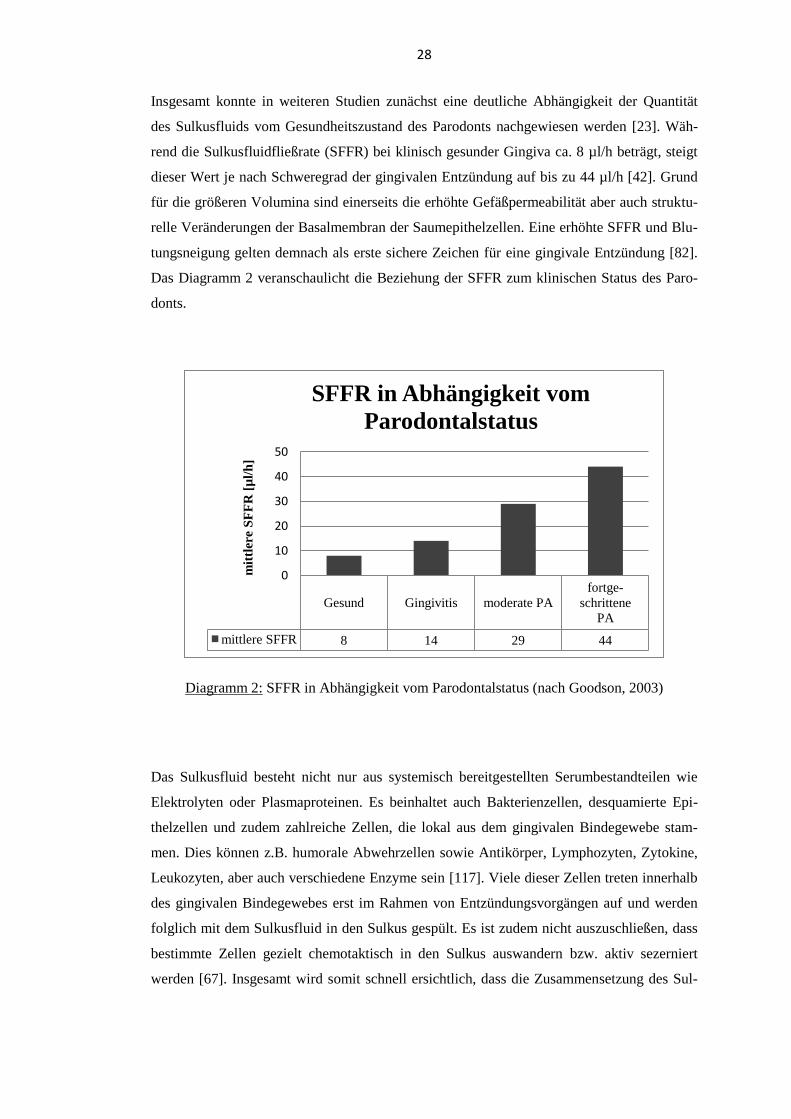
Insgesamt konnte in weiteren Studien zunächst eine deutliche Abhängigkeit der Quantität
des Sulkusfluids vom Gesundheitszustand des Parodonts nachgewiesen werden [23]. Wäh-
rend die Sulkusfluidfließrate (SFFR) bei klinisch gesunder Gingiva ca. 8 µl/h beträgt, steigt
dieser Wert je nach Schweregrad der gingivalen Entzündung auf bis zu 44 µl/h [42]. Grund
für die größeren Volumina sind einerseits die erhöhte Gefäßpermeabilität aber auch struktu-
relle Veränderungen der Basalmembran der Saumepithelzellen. Eine erhöhte SFFR und Blu-
tungsneigung gelten demnach als erste sichere Zeichen für eine gingivale Entzündung [82].
Das Diagramm 2 veranschaulicht die Beziehung der SFFR zum klinischen Status des Paro-
donts.
Diagramm 2: SFFR in Abhängigkeit vom Parodontalstatus (nach Goodson, 2003)
Das Sulkusfluid besteht nicht nur aus systemisch bereitgestellten Serumbestandteilen wie
Elektrolyten oder Plasmaproteinen. Es beinhaltet auch Bakterienzellen, desquamierte Epi-
thelzellen und zudem zahlreiche Zellen, die lokal aus dem gingivalen Bindegewebe stam-
men. Dies können z.B. humorale Abwehrzellen sowie Antikörper, Lymphozyten, Zytokine,
Leukozyten, aber auch verschiedene Enzyme sein [117]. Viele dieser Zellen treten innerhalb
des gingivalen Bindegewebes erst im Rahmen von Entzündungsvorgängen auf und werden
folglich mit dem Sulkusfluid in den Sulkus gespült. Es ist zudem nicht auszuschließen, dass
bestimmte Zellen gezielt chemotaktisch in den Sulkus auswandern bzw. aktiv sezerniert
werden [67]. Insgesamt wird somit schnell ersichtlich, dass die Zusammensetzung des Sul-
Gesund Gingivitis moderate PA
fortge-
schrittene
PA
mittlere SFFR 8 14 29 44
0
10
20
30
40
50
mit
tler
e S
FF
R [
µl/
h]
SFFR in Abhängigkeit vom
Parodontalstatus

29
kusfluids in besonderer Weise den parodontalen Zustand widerspiegelt und dass das Sulkus-
fluid folglich als Marker für Parodontalerkrankungen dienen kann.
Zahlreiche Studien zeigten, dass sowohl quantitative als auch qualitative Unterschiede beste-
hen, wenn Sulkusfluidproben von verschiedenen Sulki einer Person analysiert werden [107].
Die einzelnen Parameter sind oft über die Zeit gesehen nicht konstant, sondern schwanken in
ihren Werten beträchtlich. Zu den zahlreichen Erklärungsversuchen für dieses Phänomen
zählen die Art der Probengewinnung, das Problem der exakten Volumenbestimmung, der
Zahnseiten-spezifische Verlauf von Parodontalerkrankungen, die unterschiedliche Anatomie
der Sulki und eine mögliche Kontamination mit Speichel, Blut oder Plaque [44].
Das Sulkusfluid hat viele verschiedene funktionelle Bedeutungen. So besitzt es infolge sei-
nes beständigen Flusses zunächst eine mechanische Spülfunktion [17]. Es stellt zudem ein
Substrat für die Mikroorganismen der subgingivalen Plaque dar und hat aufgrund der enthal-
tenen Abwehrzellen auch eine anti-mikrobielle Wirkung.
1.6.3. Zement
Zement ist mit einem Mineralgehalt von 65 Gew. % die am geringsten mineralisierte Zahn-
hartsubstanz (vgl. [52]). Die organische Komponente beträgt 23 Gew. %, der Rest besteht
aus Wasser. Damit ist Zement dem Knochen strukturell sehr ähnlich. Zement wird zeitlebens
gebildet und liegt als dünne Schicht auf dem Dentin der Zahnwurzel auf. Hauptaufgabe des
Zements ist die Verankerung der aus dem Desmodont einstrahlenden Fasern und damit wie-
derum die Verankerung der Zähne in der Alveole. Man kann aufgrund der histologischen
Struktur zwei verschiedene Zementarten unterscheiden.
Der azelluläre-fibrilläre Zement (Primärzement) enthält keine Zellen, jedoch zahlreiche min-
eralisierte kollagene Fibrillen, die die Fortsetzung der vom Desmodont einstrahlenden Fasern
darstellen. Diese werden im Zement als Sharpey-Fasern bezeichnet und verlaufen senkrecht
zur Dentinoberfläche. Der Primärzement befindet sich vorwiegend im koronalen Drittel der
Zahnwurzel.
Im apikalen Bereich der Zahnwurzel und im Bereich von Bi- und Trifurkationen findet man
mit dem zellulären-fibrillären Zement die zweite Zementart vor (Sekundärzement). Er ist
ebenfalls von senkrecht zur Dentinoberfläche verlaufenden Faserbündeln durchzogen, die
auch als Fortsetzung der desmodontalen Fasern anzusehen sind. Diese sind hier jedoch
dicker und weniger mineralisiert. Zusätzlich gibt es bei dieser Zementart mit den in Lakunen
liegenden Zementozyten auch eine zelluläre Komponente.

30
1.6.4. Desmodont
Das Desmodont wird auch als Periodont oder Wurzelhaut bezeichnet. Es stellt ein zell- und
faserreiches Bindegewebe dar und füllt den 0,25 mm breiten sanduhrförmigen Desmodontal-
spalt zwischen Wurzeloberfläche und Alveolarknochen aus (vgl. [53]). Es endet 1 – 2 mm
unterhalb der Schmelz-Zement-Grenze und geht kontinuierlich in das Bindegewebe der be-
festigten Gingiva über. Das Desmodont ermöglicht mittels zahlreicher Fasern eine syndes-
motische Verankerung der Zähne in den knöchernen Alveolen.
Es wird grundsätzlich zwischen primären und sekundären Fasern unterschieden. Die mehr-
heitlich kollagenen primären Faserbündel verlaufen vom Alveolarknochen senkrecht zum
Wurzelzement. Der im Hartgewebe verlaufende Teil dieser Fasern wird als Sharpey-Fasern
bezeichnet. Sie sind nicht dehnbar, verlaufen jedoch in gewellter Form und ermöglichen da-
durch eine gewisse Zahnmobilität. Als sekundäre Fasern werden die im Desmodont unge-
bündelt in zufälliger Ausrichtung liegenden kollagenen und elastischen Fasern bezeichnet.
Beide Faserarten sind bei funktionell belasteten Zähnen generell dichter und dicker als bei
funktionslosen.
Neben den Fasern kommen im Desmodont noch zahlreiche Zellen wie Fibroblasten, Osteo-
blasten, Osteoklasten, Zementoblasten, undifferenzierte Mesenchymzellen, Makrophagen
und Monozyten vor, die eine außergewöhnlich hohe Umbaubereitschaft des Desmodonts er-
möglichen [83].
Das Desmodont ist zudem von einem dichten anastomosierenden Blutgefäßsystem durchzo-
gen, das funktionell eine Art Puffersystem bei Zahnbelastung darstellt. Apikal und interradi-
kulär gibt es als Besonderheit mit den Wedl-Gefäßknäueln Direktverbindungen zwischen
Arteriolen und Venolen. Das Desmodont wird außerdem von zahlreichen Fasern des nervus
trigeminus innerviert und übernimmt demzufolge auch wichtige sensorische Funktionen.
1.6.5. Alveolarknochen
Der Alveolarfortsatz ist der Teil des Kieferknochens, der die Knochenfächer (Alveolen) für
die Zähne bereitstellt (vgl. [53]). Er liegt dem Knochen der mandibula bzw. maxilla auf und
endet 1 – 2 mm apikal der Schmelz-Zement-Grenze. Der Alveolarknochen besteht aus einer
oralen und vestibulären Kortikalisschicht und ist zentral aus Spongiosa aufgebaut. Die dem
Zahn zugewandte Seite der Alveole ist mit einer dünnen Alveoleninnenkortikalis ausgeklei-
det (lamina cribriformis). Diese besitzt zahlreiche Öffnungen (Volkmann-Kanäle), durch die
Blutgefäße und Nerven zum Desmodont ziehen. Wegen der vielen einstrahlenden primären
Fasern aus dem Desmodont, die hier ebenfalls Sharpey-Fasern genannt werden, wird der Al-
veolarknochen an dieser Stelle auch Faserknochen genannt. Wie Zement und Desmodont ist

31
auch der Alveolarknochen eine funktionell adaptive Substanz. Er unterliegt demnach einer
ständigen Remodellation durch Osteoklasten bzw. Osteoblasten und reagiert auf physiologi-
sche funktionelle Reize mit einem entsprechenden Aufbau. Bei Zahnverlust bildet er sich
entsprechend zurück.
1.7. Parodontopathien
1.7.1. Allgemeines
Parodontopathien sind entzündlich oder nicht-entzündlich bedingte Erkrankungen des Paro-
donts. Sie manifestieren sich als pathologische Veränderungen der parodontalen Gewebebe-
standteile und führen im Endstadium zum Zahnverlust [55]. Erste Beobachtungen zu Paro-
dontopathien wurden bereits vor 5000 Jahren gemacht [34], doch ihre Ätiologie und Patho-
genese waren bis ins 19. Jahrhundert nur wenig erforscht. Man erkannte jedoch schnell, dass
sich sowohl die Ätiologie als auch die Pathogenese teilweise stark unterscheiden, weshalb
man Mitte des 20. Jahrhunderts folgerte, dass es viele unterschiedliche Arten der Parodonto-
pathien geben muss [85]. Dies machte es notwendig eine allgemein gültige Klassifikation für
die zahlreichen Parodontopathien einzuführen. Diese Nomenklatur wurde in zahlreichen Dis-
kursen über die Jahre hinweg mehrfach modifiziert, wobei die derzeit gültige Form auf eine
Konsensuskonferenz der American Academy of Periodontology (AAP) aus dem Jahr 1999
zurückgeht [4]. Die Tabelle 4 zeigt eine stark vereinfachte Zusammenstellung dieser Nomen-
klatur.
I. Gingivale Erkrankungen
a) Plaque-induziert
b) Nicht-Plaque-induziert
II. Chronische Parodontitis
a) Lokalisiert
b) Generalisiert
III. Aggressive Parodontitis
a) Lokalisiert
b) Generalisiert
IV. Parodontitis als Manifestation einer Systemerkrankung
V. Nekrotisierende Parodontalerkrankungen
VI. Parodontalabszesse
VII. Paro-Endo-Läsionen
VIII. Entwicklungsbedingte Zustände oder Deformitäten
Tabelle 4: AAP Nomenklatur der Parodontopathien (nach Armitage, 1999)

32
Trotz des mittlerweile profunden Verständnisses für das Wesen der Parodontopathien, er-
folgt die Diagnose und damit die Klassifizierung der Parodontopathien heute immer noch
fast ausschließlich mittels traditioneller klinischer Parameter [3]. Diese sind jedoch keine
präzisen Indikatoren wie beispielsweise biochemische Parameter. Da diese klinischen Krite-
rien oftmals auch unterschiedlich definiert werden [91], kommt es bei der Definition der ein-
zelnen Parodontopathien häufig zu Ungenauigkeiten und Überschneidungen. Eine genaue
Analyse der Ätiologie und Pathogenese von entzündlichen Parodontalerkrankungen muss
sich demzufolge zwangsläufig auf eine allgemeine Betrachtungsweise der beiden Hauptver-
treter Gingivitis und Parodontitis beschränken.
1.8. Gingivitis und Parodontitis
1.8.1. Definition
Gingivitis und Parodontitis sind Entzündungsreaktionen des parodontalen Gewebes, die in
Folge einer bakteriellen Infektion des Zahnes auftreten [22]. Als Gingivitis wird eine akute
oder chronische Entzündung der Gingiva bezeichnet, wobei das Ausmaß der Gewebezerstö-
rung gering und zudem vollständig reversibel ist. Bei einer länger andauernden bakteriellen
Exposition kann sich aus der Gingivitis entsprechend eine Parodontitis entwickeln. Diese ist
als entzündliche Erkrankung des gesamten Zahnhalteapparates definiert. Im Gegensatz zur
Gingivitis kommt es hierbei zu einem destruktiven Abbau des parodontalen Gewebes mit zu-
sätzlichem Alveolarknochenabbau [55].
1.8.2. Ätiologie
Gingivitis und Parodontitis haben generell eine multifaktorielle Ätiologie [83]. So ist die Ur-
sache für ihr Auftreten stets eine Kombination aus bakterieller Exposition und der vom
Wirtsorganismus daraufhin ausgelösten Reaktion. Schließlich wird dieser Prozess noch
durch lokale und systemische Faktoren beeinflusst.
1.8.2.1. Plaque
In der Mundhöhle gibt es über 500 Bakterienarten [88]. Diese kommen entweder in einer
planktonischen Phase frei schwimmend vor, oder sie lagern sich in Form eines Biofilmes
ortsständig auf die Zahnoberfläche auf. Dieser strukturierte, mit Wasserspray nicht entfern-

33
bare Zahnbelag wird als Plaque bezeichnet (vgl. [55]). Koronal des Gingivasaumes liegende
Plaque wird als supragingivale Plaque bezeichnet. Diese besteht zu 75 % aus grampositiven,
fakultativ anaeroben Kokken und Stäbchen. Plaque, die sich im gingivalen Sulkus ausbreitet
wird als subgingivale Plaque bezeichnet und unterscheidet sich hinsichtlich ihrer Zusammen-
setzung und Struktur von der supragingivalen Plaque. Das Vorhandensein von Plaque ist für
die Entstehung von entzündlichen Parodontopathien eine notwendige Voraussetzung. Ging
man anfangs noch davon aus, dass alleine die Quantität der Plaque die Entzündung hervor-
ruft (unspezifische Plaquehypothese), so erkannte man bald, dass es sich bei Gingivitis und
Parodontitis um spezifische Infektionen handelt, die mit einer entsprechenden Veränderung
in der Plaquezusammensetzung einhergehen (spezifische Plaquehypothese). Socransky et al.
teilten die Bakterien der Plaque entsprechend ihrer Pathogenität in sechs verschiedene Kom-
plexe ein [108]. Während Bakterien des grünen und gelben Komplexes keine Parodontopa-
thien verursachen, treten solche des Aa Komplexes vor allem bei aggressiven Entzündungs-
erscheinungen auf. Die Bakterien des orangen und vor allem des roten Komplexes wurden
als hauptverantwortlich für das Auftreten von Parodontalerkrankungen ausgemacht. Die
Abbildung 14 veranschaulicht vereinfacht die Komplexeinteilung oraler Mikroorganismen.
Abbildung 14: Mikrobielle Komplexe der Plaque (nach Socransky, 1992)
Heutzutage werden entzündliche Parodontopathien als opportunistische Infektionen angese-
hen, bei denen es neben der Anwesenheit pathogener Keime zusätzlich auf ein für die Ver-
mehrung der Keime günstiges Milieu ankommt [55].
Bakterien können einen direkten oder indirekten pathologischen Effekt auf das parodontale
Gewebe ausüben (vgl. [102]). Stoffwechselprodukte oder Enzyme, die während des Metabo-
lismus von Bakterien freigesetzt werden, können direkt zu einer Entzündung bzw. zu einem
destruktiven Abbau des Gewebes führen. Zudem werden von Bakterien Endo- bzw. Exotox-
ine gebildet, die indirekt durch Aktivierung von Abwehrreaktionen im Wirtsorganismus zu
einer Schädigung des parodontalen Gewebes führen. Als Endotoxine werden dabei Lipopo-
Aa Komplex
• A. actinomy-cetemcomitans
Roter Komplex
• P. gingivalis
• T. forsythensis
• T. denticola
Oranger Komplex
• P. intermedia
• P. nigrescens
• P. micros
• F. nucleatum
Orange assoziierter
Komplex
• C. rectus
• E. nodatum
Gelber Komplex
• S. sanguis
• S. mitis
• S. oralis
Grüner Komplex
• E. corrodens
• C. species

34
lysaccharide aus der Wand gramnegativer Bakterien bezeichnet, Exotoxine sind die von
Bakterien sezernierten Antigene [55].
Man fand jedoch heraus, dass die Anwesenheit parodontal pathogener Keime nicht zwangs-
läufig bei jedem Menschen zu einer entzündlichen Parodontopathie führt. Diese bilden sich
vielmehr individuell in unterschiedlichem Ausmaß aus und sind zudem von der jeweiligen
Prädisposition des Wirtsorganismus abhängig [83]. Somit ist klar, dass den Abwehrreaktio-
nen des Wirtsorganismus im Rahmen der Ätiologie entzündlicher Parodontopathien eine ent-
scheidende Rolle zukommt.
1.8.2.2. Abwehrreaktionen des Wirtsorganismus
Die Abwehrreaktionen des Wirtsorganismus haben generell die Elimination des mikrobiellen
Erregers zum Ziel und werden demzufolge nach einer entsprechenden bakteriellen Expositi-
on ausgelöst. Die dabei ablaufenden Mechanismen tragen neben der direkten bakteriellen Pa-
thogenität jedoch auch zu einer Zerstörung des Parodonts bei. Zu den Abwehrreaktionen
zählen einerseits das Auslösen von Entzündungsreaktionen und andererseits das Aktivieren
von Prozessen der unspezifischen und spezifischen Immunabwehr (vgl. [55]).
Entzündungen manifestieren sich zunächst lokal als Gefäßbindegewebsreaktionen mit einer
erhöhten Gefäßpermeabilität für Blutplasma und Blutzellen. Es kommt zu einer Transmigra-
tion von Abwehrzellen wie Monozyten, Leukozyten und Makrophagen aus dem Blut und zu
einer Proliferation ortsständiger Zellen. Die fünf typischen Entzündungszeichen wurden be-
reits in der Antike von Galen als calor, dolor, rubor, tumor und functio laesa beschrieben.
Die Entzündungsreaktionen werden durch ein komplexes Netzwerk von Mediatoren gesteu-
ert. Die wichtigsten Mediatoren sind neben den Arachidonsäurederivaten Prostaglandin und
Leukotrien die Zytokine. Zu ihnen werden unter anderem die Interleukine, Interferone, Che-
mokine und bestimmte Wachstumsfaktoren gezählt [92]. Zytokine werden nach entsprechen-
der Stimulation durch andere Zytokine, aber auch durch bakterielle Endotoxine, aus den ver-
schiedensten Wirtszellen freigesetzt. Sie beeinflussen das Verhalten bzw. die Eigenschaften
anderer Zellen und haben unterschiedlichste immunregulatorische Funktionen [56]. Einige
Zytokine wie Interleukin-1β und Tumor-Nekrose-Faktor-α haben pro-inflammatorische, an-
dere wie Interleukin-10 und Transforming-Growth-Faktor-β anti-inflammatorische Wirkun-
gen (vgl. [12]). Zytokine beeinflussen jedoch auch Matrixmetalloproteinasen (MMPs) [99]
oder Osteoklasten [83], die den Kollagen- und Knochenabbau steuern. Somit ist offensicht-
lich, dass Zytokine je nach vorliegender Konzentration entweder einen stabilisierenden oder
zerstörenden Einfluss auf das Parodont haben können. Die Abbildung 15 gibt einen schema-

35
tischen Überblick über die verschiedenen Zytokinwirkungen infolge einer mikrobiellen Ex-
position und die damit verbundenen pathologischen parodontalen Veränderungen.
Abbildung 15: Zytokinwirkungen im Parodont (nach Bartold, 2006)
Im Rahmen von allgemeinen Entzündungsreaktionen kommt es auch zur Auslösung von Im-
munreaktionen der unspezifischen und spezifischen Abwehr (vgl. [55]). Die Immunreaktion-
en werden von Bestandteilen des Blutes vermittelt und laufen entsprechend den Entzündun-
gsreaktionen im gingivalen Bindegewebe ab.
Die Phagozytose körperfremder Substanzen zählt zur zellulären unspezifischen Abwehr. Sie
wird bei entzündlichen Parodontopathien vorwiegend von chemotaktisch lokal angereicher-
ten polymorphkernigen neutrophilen Granulozyten übernommen. Mit dem Komplementsys-
tem, einem Komplex von 17 verschiedenen Serumproteinen, gibt es zudem eine humorale
Komponente der unspezifischen Abwehr zur Elimination von Antigenen. Da die unspezifi-
sche Abwehr nicht gegen alle antigenen Substanzen wirksam ist, wird sie meist von der spe-
zifischen Abwehr ergänzt. Deren humorale Komponente ist die gezielte Produktion von An-
tikörpern, das heißt Immunglobulinen, durch aktivierte B-Lymphozyten. T-Lymphozyten un-
terstützen als T-Helferzellen einerseits die humorale Immunantwort der B-Lymphozyten, an-
Mikrobielle Exposition
Gewebezellen
Zytokinfreisetzung
Osteoklasten
Bindegewebszerfall und Knochenresorption
Fibroblasten, Monozyten
MMP-Freisetzung

36
dererseits bilden sie als T-Killerzellen durch das Auslösen der Apoptose körperfremder Zel-
len die zelluläre Komponente der spezifischen Abwehr.
1.8.2.3. Lokale und systemische Faktoren
Lokale und systemische Faktoren können alleine keine entzündlichen Parodontopathien aus-
lösen. Sie beeinflussen jedoch deren Verlauf (vgl. [55]). Lokale Faktoren wie die Zahnanato-
mie, Zahnstellung, okklusale Kräfte aber auch Mundatmung, Ernährung und Speichel kön-
nen beispielsweise die Retention der Plaque beeinflussen und damit den Verlauf einer Gingi-
vitis bzw. Parodontitis begünstigen.
Zu den systemischen Faktoren zählen zahlreiche Risikofaktoren, die in der Regel ebenfalls
das Auftreten einer entzündlichen Parodontopathie begünstigen. Als Risikofaktoren gelten
beispielsweise Rauchen, Diabetes und Osteoporose [86]. In jüngster Zeit wurden zudem ge-
netische Faktoren, die die Abwehrreaktionen des Patienten steuern, als Risikofaktoren ent-
deckt. Das bekannteste Beispiel ist der Interleukin-1 Gen-Polymorphismus [106]. Hierbei
kommt es infolge einer Basenpaarvertauschung in den Genclustern des Zytokins bei einer
mikrobiellen Exposition zu einer Überproduktion von Interleukin-1 mit einer entsprechend
intensiveren und umfangreicheren Abwehrreaktion.
1.8.3. Pathogenese
Die Pathogenese entzündlich bedingter Parodontopathien läuft histologisch in vier Schritten
ab (vgl. [55] [102]). Man unterscheidet zwischen der initialen, der frühen, der etablierten und
der fortgeschrittenen Läsion. Die initiale und frühe Läsion beschreiben den Zustand einer
klinisch manifesten akuten Gingivitis, die etablierte Läsion den Zustand einer chronischen
Gingivitis. Die fortgeschrittene Läsion stellt den Übergang von der chronischen Gingivitis in
eine Parodontitis dar. Eine chronische Gingivitis muss jedoch nicht zwangsläufig in eine Pa-
rodontitis übergehen.
Die unterschiedlichen Läsionstypen können innerhalb eines Gebisses gleichzeitig nebenein-
ander auftreten, da Parodontopathien generell „site-specific“ verlaufen [22]. Zusätzlich kann
man zwischen sich abwechselnden aktiven und inaktiven Phasen der parodontalen Erkran-
kung unterscheiden.

37
1.8.3.1. Initiale Läsion
Die initiale Läsion tritt innerhalb von zwei bis vier Tagen nach einer Plaqueneubildung auf.
Dieser Zustand ist vollständig reversibel und lässt klinisch noch keine Entzündungszeichen
erkennen. Er ist demnach als physiologisch anzusehen. Kennzeichen sind eine akut entzünd-
liche Reaktion des gingivalen Gefäßplexus. Vasoaktive Mediatoren führen zu einer erhöhten
Permeabilität und einer Dilatation der Gefäße. Es kommt in der Folge zu einer entzündlich-
ödematösen Schwellung der Gingiva und zu einer erhöhten SFFR. Des Weiteren kann eine
verstärkte Migration von neutrophilen Granulozyten in das Saumepithel und den Sulkus, ein
Auftreten von Serumproteinen im Sulkus, ein Abbau perivaskulären Kollagens und eine
Auflockerung des koronalen Saumepithels mit einer partiellen Auflösung des dortigen Epi-
thelansatzes beobachtet werden. Die Abbildung 16 verdeutlicht die Entwicklung der frühen
Läsion aus einer klinisch gesunden Gingiva.
Abbildung 16: Histologie der gesunden Gingiva (a) und der initialen Läsion (b) (nach
Hellwig, 2007)
1.8.3.2. Frühe Läsion
Aus der initialen Läsion kann sich innerhalb von 14 Tagen die frühe Läsion entwickeln. Die-
se zeigt ähnliche Kennzeichen wie ihre Vorstufe, jedoch in stärkerer Ausprägung. Es kommt
zusätzlich zu einer Anreicherung von Abwehrzellen im Interstitium, wobei vor allem T-
Lymphozyten und Makrophagen nachgewiesen werden können. Neben einer zytopathischen
Veränderung der ortsständigen Fibroblasten und einem weiteren Kollagenverlust beginnt das
Saumepithel unter Ausbildung epithelialer Reteleisten lateral ins Bindegewebe zu proliferie-
ren. Die Abbildung 17 verdeutlicht die histologischen Veränderungen der frühen Läsion.

38
Abbildung 17: Histologie der frühen Läsion (c) (nach Hellwig, 2007)
1.8.3.3. Etablierte Läsion
Die etablierte Läsion tritt wenige Wochen nach der frühen Läsion auf. Sie ist wie diese ein
vollständig reversibler Zustand und immer mit subgingival auftretender Plaque verbunden.
Die akut entzündlichen Kennzeichen sind weiterhin vorhanden und entsprechend verstärkt
ausgeprägt. Des Weiteren treten extravaskuläre Immunglobuline im Bindegewebe und
Saumepithel auf, wobei nun B-Lymphozyten dominieren. Das gingivale Stützgewebe löst
sich nahezu vollständig auf und das Saumepithel proliferiert weiter nach apikal und lateral,
wobei es zu einem keratinisierten Taschenepithel umgewandelt wird. Der gingivale Sulkus
hat sich in der Folge zu einer Zahnfleischtasche mit einer Tiefe von 2 – 3 mm umgewandelt.
Die Abbildung 18 verdeutlicht die histologischen Veränderungen der etablierten Läsion.
Abbildung 18: Histologie der etablierten Läsion (d) (nach Hellwig, 2007)

39
1.8.3.4. Fortgeschrittene Läsion
Die fortgeschrittene Läsion ist ein destruktiver Prozess mit Perioden der Exazerbation und
Stagnation. Neben den weiter fortbestehenden Kennzeichen der etablierten Läsion, dehnt
sich die Läsion hier auf den Alveolarknochen und das Desmodont aus. Es kommt zu einem
Knochenabbau, einem anhaltenden Kollagenverlust unterhalb des Saum- bzw. Taschenepi-
thels und zur Ausbildung einer parodontalen Tasche. Generell handelt es sich um eine ausge-
dehnte entzündliche und immunologische Gewebereaktion, die durch alleinige Mundhy-
gienemaßnahmen nicht mehr zu einer „restitutio ad integrum“ führt. Die Abbildung 19 ver-
deutlicht die histologischen Vorgänge der fortgeschrittenen Läsion.
Abbildung 19: Histologie der fortgeschrittenen Läsion (e) (nach Hellwig, 2007)

40
2. Problemstellung
2.1. Stand der Forschung
2.1.1. Extrazelluläre Funktionen des Thymosin β4
In den letzten Jahren konzentrierte sich die Forschung verstärkt auf das extrazellulär vor-
kommende Thymosin β4, das zytokinähnliche Eigenschaften aufweist [48]. Es wirkt unter
anderem chemotaktisch, es induziert bzw. reprimiert die Expression verschiedener Gene und
hat angiogene und anti-inflammatorische Eigenschaften. Thymosin β4 wurde in verschieden-
en Körperflüssigkeiten wie Blut, Wundflüssigkeit, Tränenflüssigkeit und Speichel nachge-
wiesen. Es ist jedoch unklar, wie Thymosin β4 als intrazelluläres Peptid ohne Signalsequenz
in den extrazellulären Raum gelangen kann.
2.1.2. Nachweis von oralem Thymosin β4
Gerade orales Thymosin β4 wurde in vergangener Zeit verstärkt untersucht. Die Erforschung
von Thymosin β4 in der Mundhöhle begann im Jahr 2007 als Badamchian et al. das Peptid in
der Tränenflüssigkeit und im Gesamtspeichel mittels der ELISA-Methode nachweisen konn-
ten [7]. Die gefundenen Konzentrationen schwankten zwischen 0,2 und 3,6 µg/ml und waren
sowohl vom Alter als auch vom Geschlecht der Testpersonen abhängig. Die höchsten Kon-
zentrationen wurden in der Gruppe der 25- bis 35-jährigen, die niedrigsten in der Gruppe der
über 50-jährigen Probanden entdeckt. Männliche Testpersonen zeigten generell erhöhte Kon-
zentrationen an Thymosin β4.
2.1.3. Funktionelle Bedeutung des oralen Thymosin β4
Akhter et al. entdeckten im Jahr 2005 als Erste eine mögliche Funktion von Thymosin β4 in
der Mundhöhle [1]. Sie berichteten, dass Thymosin β4 während der Zahnkeimentwicklung
bei Mäusen verstärkt exprimiert wird und schrieben dem Peptid demnach eine funktionelle
Bedeutung bei der Initiation, dem Wachstum und der Differenzierung der Zahnkeime zu.
2.2. Zielsetzung der Arbeit
Die bisherigen Studien über das orale Thymosin β4 zeigen, dass Thymosin β4 in der Mund-
höhle vorkommt und dass ein Nachweis des Peptids im Gesamtspeichel möglich ist. Da sich

41
Gesamtspeichel jedoch aus verschiedenen Komponenten zusammensetzt, ist nicht klar, wo-
her das nachgewiesene Thymosin β4 stammt. Es herrschen zudem Unklarheiten über mög-
liche Abhängigkeiten und Mechanismen der Freisetzung von Thymosin β4 in die Mund-
höhle.
Ziel der vorliegenden Arbeit ist demnach, mögliche Quellen und Einflussfaktoren für das
Auftreten von Thymosin β4 im menschlichen Speichel zu untersuchen.
Zunächst wird die Thymosin β4 Konzentration im Gesamtspeichel in Abhängigkeit vom Al-
ter der Probanden und als eine Funktion der Zeit untersucht. Anschließend werden durch
Analyse von Gesamtspeichel-, Drüsenspeichel- und Sulkusfluidproben verschiedene mög-
liche Quellen für das orale Thymosin β4 analysiert. Zuletzt werden Thymosin β4 Konzentra-
tionen im Sulkusfluid parodontal gesunder und erkrankter Patienten und ein möglicher Zu-
sammenhang zu den Mundhygiene- bzw. Entzündungsindizes API und SBI untersucht. Zu-
dem wird geprüft, ob eine verstärkte okklusale Belastung einen möglichen Freisetzungsme-
chanismus für Thymosin β4 in das Sulkusfluid darstellt.

42
3. Konzept der Studie
3.1. Gruppeneinteilung der Probanden
Die Probennahmen erfolgten gemäß ethischer Prinzipien mit dem Einverständnis und einer
entsprechenden Aufklärung der Probanden. Das Probandenkollektiv bestand aus Studenten
der Universität Erlangen-Nürnberg, Mitarbeitern des biochemischen Instituts Erlangen und
Patienten der Abteilung für Zahnerhaltung und Parodontologie der Zahnklinik 1 Erlangen.
Die Bestimmung von Thymosin β4 mittels Reverse-Phase High Pressure Liquid Chromato-
graphy (RP-HPLC) und anschließender Matrix-Assisted Laser Desorption/Ionisation - Time
of Flight (MALDI-TOF) Analyse erfolgte ebenso wie die Beobachtung der Stabilität von
Thymosin β4 im Gesamtspeichel durch Analyse von Proben einzelner gezielt ausgewählter
Probanden. Das Hauptaugenmerk der Arbeit liegt jedoch auf einer kollektiven Betrachtung
von Thymosin β4 in oralen Flüssigkeiten durch einen entsprechenden Nachweis mittels RP-
HPLC. Hierfür wurden die Probanden entsprechend der jeweiligen zu untersuchenden Para-
meter anhand verschiedener Kriterien in unterschiedliche Gruppen A, B, C, D und E einge-
teilt. Die Probanden der Gruppen A und B wurden aufgrund ihres Alters ohne Berücksichti-
gung des Parodontalstatus eingeteilt. Das Kriterium für die Probanden der Gruppe C war pa-
rodontale Gesundheit, die durch Taschenmessung mit einer PCP 12 Sonde im Anschluss an
die entsprechende Probengewinnung sichergestellt wurde. Die Taschentiefen betrugen
durchschnittlich 1,9 mm. Die Gruppe D bestand aus Patienten, die im Rahmen einer PA-Re-
call Therapie in der Abteilung für Zahnerhaltung und Parodontologie an der Zahnklinik 1 Er-
langen behandelt wurden. Sie waren alle an unterschiedlichen Formen der Parodontopathien
erkrankt. Die Taschentiefen betrugen im Durchschnitt 3,4 mm. Die Tabelle 5 gibt eine Über-
sicht über die Parameter und Einteilungskriterien für die Gruppen A, B, C und D.
Gruppe Parameter Einteilungskriterium
A 22 – 30 Jahre, n = 7 Alter
B 46 – 60 Jahre, n = 8 Alter
C 22 – 26 Jahre, n = 11 Parodontal gesund
D 48 – 71 Jahre, n = 8 Parodontal erkrankt
Tabelle 5: Einteilungskriterien der untersuchten Gruppen Teil 1
Zuletzt wurde noch eine Gruppe E gebildet, die aus vier zufällig ausgewählten Probanden
aus der Gruppe C bestand. Die Taschentiefen betrugen im Durchschnitt weniger als 3 mm.
Die Tabelle 6 gibt eine Übersicht über die Zusammensetzung der Gruppe E.

43
Gruppe Parameter Einteilungskriterium
E 23 – 25 Jahre, n = 4 Parodontal gesund
Tabelle 6: Einteilungskriterien der untersuchten Gruppen Teil 2
3.2. Beobachtungsparameter
Von den Probanden der oben genannten Gruppen wurden die Speichel- bzw. Sulkusfluid-
proben mit verschiedenen Methoden und unter unterschiedlichen Bedingungen (siehe Punkt
4.2.1.) gewonnen und auf die relevanten Parameter hin untersucht. Die Tabelle 7 gibt eine
Übersicht über die dabei jeweils analysierten Parameter.
Gruppe Analysierte Parameter
A Tβ4 und Protein im Gesamtspeichel
B Tβ4 und Protein im Gesamtspeichel
C Tβ4 im Gesamtspeichel, Drüsenspeichel, Sulkusfluid
D Tβ4 im Sulkusfluid
E Tβ4 im Sulkusfluid unter verstärkter okklusaler Belastung
Tabelle 7: Gruppen-spezifisch analysierte Parameter
Die Konzentrationen an Thymosin β4 in den Speichel- bzw. Sulkusfluidproben wurden mit-
tels RP-HPLC (siehe Punkt 4.2.6.) bestimmt. Die Angabe der Werte für die Gruppen A – E
erfolgte dabei in µg/ml. Die Proteinkonzentration in den Gruppen A und B wurde mittels
Biuretreaktion ermittelt und in g/l angegeben.

44
4. Material und Methoden
4.1. Material
4.1.1. Chemikalien und Reagenzien
Aceton Roth
Acetonitril (ACN) Roth
Borsäure Roth
Biuret-Reagenz: Merck
12 mmol/l Kupfersulfat
32 mmol/l Kaliumnatriumtartrat
30 mmol/l Kaliumiodid
in 0,2 mol NaOH/l
Ethanol Roth
Fluorescamine (Fluram) Serva
LiChroprep RP-18 Merck
Lithiumhydroxid Johnson Matthey
Methanol Merck
Perchlorsäure (PCA) Merck
Trifluoressigsäure Uvasol (TFA) Merck
Waschlösung für Papierspitzen: Merck
10 ml 6 M Guanidiniumhydrochlorid (GuaHCL)
50 µl Thiodiglycol (TD)
Wasserstoffperoxid 30 % (H2O2) Merck
4.1.2. Geräte und Hilfsmittel
4.1.2.1. Analytische bzw. präparative RP-HPLC
Derivatisierungspumpe: 655A-13 Reaction Pump Merck / Hitachi
Fluorometer: FL-Detektor L-7480 Merck / Hitachi
HPLC-Controller: L-6200 LC Controller Merck / Hitachi
Interface: Interface D-7000 Merck / Hitachi
Lösungsmittelpumpe: 655A-12 Liquid Chromatograph Merck / Hitachi
Trennsäule: ODS Ultrasphere RP-18 Beckman Coulter
(5 µm, 4,6 250 mm)
UV-Detektor: L-4000 Merck / Hitachi

45
4.1.2.2. Sonstiges
Aufbisssilikon: SilaPlast Detax Dental
Erythrosinpellets: Erythrosinpellets Henry Schein
Fotometer: Novaspec II Visible Spectrometer Pharmacia Biotech
Heidemann-Instrument: Heidemann-Instrument mittel Henry Schein
Küvette: Ultramikro Quarzküvette 100 µl Merck
MALDI-TOF-MS: BiflexTM
Bruker Daltonics
Mixer: Vortexer REAX 2000 Heidolph
Papierspitzen: Paper Cones ISO 40 Henry Schein
Parodontalsonde: Parodontalsonde PCP-12 Hu-Friedy
Pipetten: Mikropipette 20 – 200 µl Corning Lambda
Mikropipette 100 – 1000 µl Corning Lambda
Pipettenspitzen: Pipettenspitze (gelb) 200 µl Sarstedt
Pipettenspitze (blau) 1000 µl Sarstedt
Rüttler: Unimax 1010 Heidolph
Speichelsaugapparatur: Glaskapillare 200 µl und Gummisauger Brand
Ultrazentrifuge: Centrifuge 5417 R Eppendorf
Watterollen: Watterollen Henry Schein
Zentrifuge: Omnifuge 2,0 RS Heraeus Sepatech
4.1.3. Pufferlösungen
Boratpuffer H3BO3 (0,4 M) mit Lithiumhydroxid (gesättigt) auf pH 9,3
Fluramlösung Fluorescamine in Aceton (200 mg/l)
Puffer A 0,1 % TFA in H2O
Puffer B 0,1 % TFA in 50 % Acetonitril
4.2. Methoden
4.2.1. Probengewinnung
Die Speichel- bzw. Sulkusfluidproben wurden mittels verschiedener Methoden und unter
verschiedenen Bedingungen auf die bereits genannten Beobachtungsparameter (siehe Punkt
3.2.) untersucht. Die Speichelfließrate wird durch eine Vielzahl verschiedener Faktoren be-
einflusst und ist dementsprechend variabel [84]. Da sich die Zusammensetzung des entspre-
chend stimuliert, bzw. unstimuliert gewonnenen Speichels unterscheidet [28], wurde die Me-

46
thode zur Speichelprobengewinnung standardisiert. Da die Methode der Sulkusfluidproben-
gewinnung ebenfalls die Zusammensetzung des Sulkusfluids beeinflussen kann [44], wurde
zur Probengewinnung ebenfalls ein standardisiertes Verfahren angewendet.
4.2.1.1. Gesamtspeichel (Gruppe A und B)
Die Probengewinnung erfolgte zur Mittagszeit. Vor der Probengewinnung wurde die Mund-
höhle des Probanden mit reinem Wasser kurz gespült um Speisereste bzw. Restspeichel aus-
zuwaschen. Der Gesamtspeichel in den Gruppen A und B wurde unstimuliert durch einfa-
ches Spucken in einen kleinen Glasbehälter über einen Zeitraum von fünf Minuten gewon-
nen. Die erhaltenen Volumina betrugen 1 – 5 ml. Die Probennahme in den Gruppen A und B
wurde nach zwei Wochen unter entsprechenden Bedingungen wiederholt.
4.2.1.2. Gesamtspeichel (Gruppe C)
Die Probengewinnung des Gesamtspeichels in der Gruppe C erfolgte ebenfalls zur Mittags-
zeit und nach entsprechendem Auswaschen der Mundhöhle mit Wasser. Im Unterschied zu
obigen Gruppen wurde der Speichel jedoch mit einer Speichelsaugapparatur von dem im Be-
handlungsstuhl sitzenden Probanden aus dem Bereich des anterioren Mundbodens gewon-
nen, nachdem der Proband für wenige Minuten Speichel unstimuliert in der Mundhöhle ge-
sammelt hatte. Die Apparatur bestand aus einer einfachen 200 µl Glaskapillare an deren En-
de ein Gummisauger angebracht war. Mit dieser Methode wurden jeweils mindestens 500 µl
Gesamtspeichel gewonnen.
4.2.1.3. Drüsenspeichel (Gruppe C)
Die Probengewinnung des Drüsenspeichels erfolgte direkt nach der Gewinnung des Gesamt-
speichels, diesmal vom im Behandlungsstuhl liegenden Probanden. Der Speichel wurde mit
der gleichen Speichelsaugapparatur, wie unter Punkt 4.2.1.2. beschrieben, direkt von den
Ausführungsgängen der einzelnen Speicheldrüsen gewonnen. Es wurden von jedem
Probanden drei Proben genommen: „gl. parotis links“, „gl. parotis rechts“ und „gl. subman-
dibularis/sublingualis“. Eine Kontamination der einzelnen Proben wurde durch entsprechen-
des Erneuern der Glaskapillare vor jeder neuen Probennahme und einem Trockenlegen des
Drüsenausführungsganges mit Watterollen sichergestellt. Es konnten somit jeweils mindes-
tens 100 µl drüsenspezifischer Speichel ohne Kontamination gewonnen werden.

47
4.2.1.4. Sulkusfluid parodontal gesunder Probanden (Gruppe C)
Nach der Gewinnung der oben genannten Gesamt- und Drüsenspeichelproben wurden an-
schließend von dem im Behandlungsstuhl liegenden Probanden noch Sulkusfluidproben ab-
genommen. Dazu wurden Papierspitzen der Größe ISO 40 verwendet, deren Gewicht vor
und nach der Probennahme in einem verschlossenen Eppendorfcup bestimmt wurde. Es
wurde insgesamt jeweils eine Sulkusfluidprobe an sechs verschiedenen Zähnen genommen.
In der Gruppe C waren dies die Zähne 13 – 23. Zur Vermeidung einer Speichelkontaminati-
on wurden die Zähne mit Watterollen trockengelegt und für 20 Sekunden mit dem Luftpuster
leicht getrocknet um bereits gebildetes Sulkusfluid zu verblasen. Dann wurde die Papier-
spitze mit einem Heidemann-Instrument unter sanftem Druck zirkulär in den gingivalen Sul-
kus eingebracht. Nach fünf Minuten wurde die Papierspitze wieder entfernt und sofort in das
entsprechende Eppendorfcup gegeben, um eine Verdunstung zu vermeiden. Dieses wurde
anschließend ausgewogen und das in der Papierspitze aufgesaugte Volumen an Sulkusfluid
durch entsprechendes Subtrahieren des vorher ermittelten Ausgangsgewichtes berechnet.
4.2.1.5. Sulkusfluid parodontal erkrankter Probanden (Gruppe D)
Das Sulkusfluid parodontal erkrankter Probanden wurde nach dem gleichen Prinzip, wie
unter Punkt 4.2.1.4. beschrieben, gewonnen und quantifiziert. Es wurde ebenfalls je eine Sul-
kusfluidprobe an sechs verschiedenen Zähnen genommen. Die Auswahl der Zähne erfolgte
diesmal jedoch aufgrund der größten Taschentiefen, wie sie in der letzten PA-Recall Sitzung
bestimmt worden waren.
4.2.1.6. Sulkusfluid unter verstärkter okklusaler Belastung (Gruppe E)
Das Sulkusfluid wurde nach dem gleichen Prinzip, wie unter Punkt 4.2.1.4. beschrieben, ge-
wonnen und quantifiziert. Es wurden jedoch nur die Zähne 13 und 23 betrachtet, von denen
insgesamt fünf Proben genommen wurden. Zunächst wurde jeweils eine Probe unter norma-
len Bedingungen gewonnen („vor Aufbiss rechts“, „vor Aufbiss links“). Dann wurde ein ver-
tikal ca. 3 cm hoher einseitiger Aufbisswall aus Silikon für den rechten oberen Eckzahn her-
gestellt. Anschließend erfolgte ca. fünf Minuten nach der ersten Probennahme eine erneute
Probengewinnung an diesem einen Eckzahn unter repetitivem kräftigem Zubeißen des Pro-
banden über die gesamte Zeit der Probennahme („Aufbiss rechts“). Danach wurde der einsei-
tige Aufbisswall entfernt und nach ca. fünf Minuten nochmals an beiden Eckzähnen Sulkus-

48
fluidproben unter normalen Bedingungen gewonnen („nach Aufbiss rechts“, „nach Aufbiss
links“).
4.2.2. Bestimmung der sulkulären Taschentiefen
Die sulkuläre Taschentiefe ist definiert als der Abstand des marginalen Gingivasaumes zum
Sulkusboden [5]. Eine Taschentiefe von 1 – 2 mm gilt als physiologisch. Die Messung der
Taschentiefen ist von großer klinischer Bedeutung in der Diagnose von Parodontopathien, da
diese mit einer apikalen Wanderung des Saumepithels und damit einer Vergrößerung der Ta-
schentiefen einhergehen. Durch Messung der Taschentiefen lassen sich somit parodontal ge-
sunde von parodontal erkrankten Probanden unterscheiden. Die Taschentiefen wurden in der
Gruppe C mesial und distal und in der Gruppe D mesial, distal, buccal und oral an allen
Zähnen mittels einer in 3 mm Schritten graduierten PCP 12 Sonde ermittelt. Die Messung
der Taschentiefen erfolgte jeweils nach der Gewinnung der einzelnen Sulkusfluidproben, um
eine mögliche Kontamination mit Blut zu vermeiden.
4.2.3. Bestimmung des API und SBI
Der (modifizierte) Approximale Plaque Index (API) wurde von Lange im Jahr 1986 einge-
führt (vgl. [54]). Hierbei wird nach Anfärben der Plaque mit Erythrosin und anschließendem
Ausspülen der Mundhöhle mit Wasser in einer Ja/Nein Entscheidung beurteilt, ob im
Approximalraum Plaque vorhanden ist. Die Beurteilung erfolgt beim API im ersten und
dritten Quadranten oral und im zweiten und vierten Quadranten vestibulär. Das Ergebnis
wird nach folgender Formel in Prozent ausgedrückt:
Die Werte des API dienen dabei zur Beurteilung der Mundhygienesituation beim Patienten.
Dabei gelten folgende Richtwerte:

49
API Wert Mundhygienezustand
< 25 % Optimal
25 – 39 % Gut
40 – 70 % Mäßig
> 70 % Unzureichend
Tabelle 8: Korrelation API Werte und Mundhygienezustand
Der (modifizierte) Sulkus Blutungs Index (SBI) wurde von Mühlemann 1971 entwickelt
(vgl. [54]) und ist ein Gingiva- bzw. Entzündungsindex. Man geht hierbei davon aus, dass
der Entzündungsgrad der Gingiva mit einer Blutungsneigung des Gingivasulkus nach dem
stumpfen Sondieren mit einer Parodontalsonde korreliert. Es wird demnach das Vorhanden-
sein einer Blutung nach Sondierung in Form einer Ja/Nein Entscheidung beurteilt. Die Erhe-
bung erfolgt im ersten und dritten Quadranten vestibulär und im zweiten und vierten Qua-
dranten oral. Das Ergebnis wird nach folgender Formel in Prozent ausgedrückt:
Die Werte des SBI dienen dabei zur Beurteilung des Entzündungszustandes des Parodonts
beim Patienten. Dabei gelten folgende Richtwerte:
SBI Wert Entzündungszustand
< 10 % Klinisch normal
10 – 20 % Schwach
21 – 50 % Mittelschwer
> 50 % Stark und generalisiert
Tabelle 9: Korrelation SBI Werte und Entzündungszustand
Der SBI wird oft in Kombination mit dem API erhoben, da somit leicht zu überprüfen ist, ob
der Patient dauerhaft eine gute Mundhygiene betreibt (niedriger SBI und niedriger API) oder
nur kurz vor der entsprechenden Untersuchung (hoher SBI und niedriger API). Der API und
SBI wurden nach oben geschilderten Kriterien in den Gruppen C und D direkt nach den ent-
sprechenden Probennahmen ermittelt.

50
4.2.4. Probenvorbereitung
Nachdem die Speichel- und Sulkusfluidproben, wie unter Punkt 4.2.1. geschildert, von den
entsprechenden Probanden gewonnen wurden, wurden sie für die entsprechende Analyse
vorbereitet. Die Vorgehensweisen waren dabei für Speichel- und Sulkusfluidproben unter-
schiedlich.
4.2.4.1. Gesamtspeichel und Drüsenspeichel
Das Vorgehen für Gesamtspeichel- und Drüsenspeichelproben war identisch und entspricht
der von Hannappel entwickelten Methode [47]. Unmittelbar nach der Probengewinnung wur-
den die Eppendorfcups bei 20000 g für fünf Minuten bei 4 °C zentrifugiert um Zellen ab-
zutrennen, die sich im Speichel befinden. Der Überstand wurde vorsichtig abgenommen und
mit 1/10 Volumen 4 M PCA versetzt. Die Probe wurde gemischt, für 30 Minuten auf Eis ge-
stellt und wiederum bei 20000 g für fünf Minuten bei 4 °C zentrifugiert. PCA führt bei
Proteinen zu einer Protonierung und einem damit verbundenen Konformationsverlust. Das
gebildete positiv geladene Proteinkation kann mit dem anwesenden Säureanion entweder ein
lösliches oder ein unlösliches Salz bilden. PCA führt zur Ausfällung der meisten Proteine,
Thymosin β4 bleibt jedoch aufgrund seiner geringen molaren Masse löslich und kann im
Überstand bestimmt werden [47]. Nach dem Zentrifugieren wurde der Überstand vorsichtig
abgenommen und mittels RP-HPLC analysiert. Das durch PCA ausgefällte Protein der Ge-
samtspeichelproben wurde mit Hilfe der Biuretreaktion quantitativ bestimmt.
4.2.4.2. Sulkusfluid
Unmittelbar nach der Volumenbestimmung des Sulkusfluids durch Auswiegen des Eppen-
dorfcups, wurde in das Eppendorfcup 100 µl einer speziellen Waschlösung (siehe Punkt 5.7.)
zugegeben. Das Eppendorfcup wurde anschließend für zwei Stunden bei Raumtemperatur
geschüttelt. Danach wurden die 100 µl Waschlösung abgenommen und in jeweils separaten
Eppendorfcups unter Eiskühlung gesammelt. Der Waschprozess wurde dann zweimal nach
dem gleichen Schema wiederholt (siehe Punkt 5.7.). Am Ende standen 300 µl Waschlösung
zur Analyse mit Hilfe der RP-HPLC bereit.
Die Waschlösung, die für diese Arbeit speziell entwickelt wurde, enthielt 10 ml 6 M Guani-
diniumhydrochlorid (GuaHCL) und 50 µl Thiodiglycol (TD). Im Wiederfindungsexperiment
(siehe Punkt 5.7.) konnte gezeigt werden, dass durch Zugabe von GuaHCL und TD im Ge-
gensatz zu anderen Waschlösungen wie 0,1 % TFA oder 4 M PCA eine fast vollständige

51
Wiederfindung von in Papierspitzen aufgesaugtem Thymosin β4 möglich ist. TD wurde da-
bei als Oxidationsschutz zugegeben.
4.2.5. Proteinbestimmung mittels Biuretreaktion
4.2.5.1. Prinzip Fotometer
Fotometer dienen der Konzentrationsmessung von Substanzen mit Chromophoren und beste-
hen grundsätzlich aus einer Lichtquelle (L), einem Monochromator (P) sowie einer Fotozel-
le. Der Aufbau ist in der Abbildung 20 dargestellt. Nach Durchtritt durch die Küvette (K),
die die zu analysierende Substanz enthält, trifft das monochromatische Licht auf einen licht-
empfindlichen Fotoempfänger (E). Dieser misst die entsprechend abgeschwächte Lichtinten-
sität, wandelt sie in ein proportionales elektrisches Signal um, das dann wiederum nach ent-
sprechender Verstärkung (V) auf der Anzeigeeinheit (A) abgelesen werden kann. Die Anzei-
ge der Messwerte erfolgt als Extinktion [E] oder als Transmission [T].
Abbildung 20: Aufbau eines Fotometers
Die Extinktion, also die Abnahme der Lichtintensität, beruht auf der Fähigkeit von Stoffen
elektromagnetische Strahlung zu absorbieren. Die Extinktion einer gelösten Substanz hängt
bei monochromatischem Licht einer bestimmten Wellenlänge (λ) von ihrer Konzentration (c)
und der Schichtdicke (d) der Küvette ab. Unter Berücksichtigung eines Temperatur und Lö-
sungsmittel abhängigen molaren Extinktionskoeffizienten (ε) ergibt sich das Lambert-
Beer´sche Gesetz, das nach entsprechender Umformung zur Konzentrationsberechnung ver-
wendet wird:

52
4.2.5.2. Prinzip Biuretreaktion
Die Biuretreaktion ist ein seit langer Zeit bekanntes, einfaches Verfahren zur quantitativen
Bestimmung von Proteinen [43]. Die Reaktion beruht auf einer Komplexbildung mit Cu2+
-
Ionen, die einen blauvioletten Farbumschlag bedingt. Der Grund für dieses Verhalten ist der
elektronenanziehende Charakter der Peptidbindungscarbonylgruppe, der dem benachbarten
–NH2 bzw. >NH eine gewisse Azidität verleiht, sodass bei der Reaktion die Protonen durch
Cu2+
-Ionen ersetzt werden können. Durch eine entsprechende fotometrische Analyse von mit
Biuret-Reagenz versetzten Peptidlösungen lässt sich demnach unter Anwendung des Lam-
bert-Beer´schen Gesetzes die Proteinkonzentration berechnen.
4.2.5.3. Probenanalyse
Das durch Fällung mit PCA erhaltene Präzipitat der Gesamtspeichelproben wurde zur Be-
stimmung der Proteinkonzentration der jeweiligen Probe genutzt. Zum Präzipitat wurden je-
weils 500 µl Biuret-Reagenz zugegeben. Die Lösung wurde über Nacht bei mittlerer Intensi-
tät auf dem Rüttler geschüttelt, bis sich das Präzipitat vollständig gelöst hatte. Anschließend
wurden jeweils 100 µl abgenommen, und es wurde in einer Ultramikro-Quarzküvette die Ex-
tinktion bei 546 nm gegen einen Leerwert bestimmt. Die quantitative Bestimmung der Prote-
inkonzentration erfolgte unter Berücksichtigung der jeweiligen Verdünnung (V: Gesamtvo-
lumen, v: Probenvolumen) mittels des Lambert-Beer´schen Gesetzes:
Bei den für diese Versuche geltenden Bedingungen betrug der Wellenlängen abhängige Ex-
tinktionskoeffizient (ε) 0,277 l g-1 cm
-1. Da Proteine in der Zelle sehr variable Massen
haben, erfolgte die Angabe von (ε) jedoch nicht wie sonst üblich in der Einheit des molaren
Extinktionskoeffizienten. Die Schichtdicke (d) der Küvette entsprach 1 cm.

53
4.2.6. Thymosin β4 Bestimmung mittels analytischer RP-HPLC
Für die Bestimmung von Thymosin β4 wurden für Speichelproben der nach PCA Fällung
gewonnene Überstand und für Sulkusfluidproben die entsprechend gesammelten Waschlö-
sungen ohne weitere Manipulation verwendet. Der Nachweis erfolgte mittels der Reverse-
Phase High Pressure Liquid Chromatography (RP-HPLC).
4.2.6.1. Prinzip der RP-HPLC
Die RP-HPLC ist ein besonderes Verfahren der Verteilungschromatografie, das wegen seiner
hohen Trennschärfe zum Nachweis und zur Aufreinigung von Peptiden verwendet wird [72].
Es beruht prinzipiell auf der unterschiedlichen Verteilung von Substanzen infolge ihrer che-
mischen Eigenschaften zwischen zwei nicht miteinander mischbaren Phasen. Als stationäre
Phase dienen Kieselgelpartikel mit kovalent gebundenen hydrophoben Alkylketten (meist 18
C-Atome), die in eine Stahlsäule gefüllt werden. Die mobile Phase besteht aus einer polaren
wässrigen Lösung mit definiertem pH-Wert und einem organischen Lösungsmittel (z.B.
Acetonitril), dessen Konzentration kontinuierlich erhöht wird. Substanzen, die in wässriger
Lösung auf die Säule aufgebracht werden, binden zunächst über hydrophobe Wechselwir-
kungen an die Alkylketten der stationären Matrix. Durch steigende Konzentration eines ge-
eigneten organischen Lösungsmittels in der mobilen Phase werden die Substanzen dann ab-
hängig von ihrer Hydrophobizität von der Säule zu unterschiedlichen Zeitpunkten eluiert und
somit getrennt. Der Nachweis der eluierten Peptide erfolgt meist durch einfache Messung der
UV-Absorption bei 280 oder 208 – 210 nm oder durch Derivatisierung funktioneller Grup-
pen zu fluoreszierenden Produkten (Nachsäulenderivatisierung).
4.2.6.2. Analytische RP-HPLC mit Fluram-Nachsäulenderivatisierung
Die verwendete Methode zur Trennung und zur quantitativen Bestimmung von Thymosin β4
entspricht der von Huff et al. [60]. Ein Fluoreszenznachweis der mit Fluram derivatisierten
Peptide bietet generell eine hohe Sensitivität und ermöglicht zudem die Verwendung von or-
ganischen Lösungsmitteln, die unterhalb von 240 nm absorbieren.
Fluram wurde vor über 30 Jahren eingeführt [116] und sein chemisches Verhalten entspre-
chend ausführlich untersucht [33]. Fluram reagiert mit primären Amino- nicht aber mit Am-
moniumgruppen zu fluoreszierenden Derivaten. Alleine ist es, genauso wie sein Hydrolyse-
produkt, nicht fluoreszierend. Da Fluram in wässriger Lösung innerhalb von 10 – 20 Sekun-
den hydrolysiert, muss es in Aceton oder Acetonitril gelöst werden. Zusätzlich muss der Lö-

54
sung Boratpuffer (pH = 9,4) zugesetzt werden, da die Reaktion nur bei einem pH-Wert von
oberhalb 9 ablaufen kann. In Abbildung 21 sind die Reaktionen von Fluram dargestellt.
Abbildung 21: Reaktionsmechanismus von Fluram
Das eigentliche RP-HPLC-System besteht aus einem von Stein und Moschera entwickelten
Peptidanalysegerät [112]. Dieses ist aus einem RP-HPLC-Trennungssystem und einem Deri-
vatisierungs- und Detektionssystem zusammengesetzt.
Das konventionelle RP-HPLC-Trennungssystem besteht aus einer 500 µl Probenschleife und
einer RP-18-Trennsäule (ODS Ultrasphere; 5 µm; 4,6 250 mm; Beckmann Coulter). Als
Fließmittel wurde Puffer A (0,1 % TFA in H2O) verwendet. Die Peptide wurden mit Puffer
B (0,1 % TFA in 50 % ACN) bei einem Fluss von 0,75 ml/min eluiert. Der Anteil des Puf-
fers B an der mobilen Phase wurde im Rahmen dieser Arbeit für das biologische Probenma-
terial optimiert und dabei während der Analysedauer folgendermaßen verändert:

55
Zeit Anteil Puffer B
5 min 46 %
20 min 49 %
25 min 100 %
30 min 100 %
31 min 0 %
Tabelle 10: Gradient der RP-HPLC
Im Anschluss an die Säule schloss sich das Derivatisierungssystem an, dessen Aufbau in der
Abbildung 22 schematisch dargestellt ist.
Abbildung 22: Aufbau des RP-HPLC-Systems
Das Säuleneluat wird im Derivatisierungssystem mit dem von weiteren Pumpen bereitge-
stellten Borat und Fluram vermischt. Zunächst wird der Boratpuffer mit einer Fließge-
schwindigkeit von 0,22 ml/min zugesetzt, um den für die Fluramreaktion nötigen pH-Wert
einzustellen. Die Fluramlösung wird anschließend mit einer Fließgeschwindigkeit von 0,22
ml/min zugepumpt. Danach durchläuft das Reaktionsgemisch eine Derivatisierungsschleife
(DS), um dem Fluram die nötige Zeit von etwa 15 Sekunden zur Reaktion mit den primären
Aminen zu geben. Die Fluoreszenz wird anschließend in einem angeschlossenen Fluorezenz-
spektrofotometer (FM) (λEX = 390 nm, λEM = 475 nm) detektiert. Das entsprechende Signal
wird über ein Interface digitalisiert und mit einer entsprechenden Software ausgewertet.

56
4.2.6.3. Probenanalyse
Zur Analyse der Konzentration von Thymosin β4 wurden die entsprechend vorbereiteten
Proben verwendet (siehe Punkt 4.2.4.). Bei den Speichelproben wurden 25 µl und bei den
Sulkusfluidproben die gesamten 300 µl Elutionslösung zunächst auf ein Volumen von 500 µl
durch Zugabe von 0,1 % TFA verdünnt und anschließend vollständig in die RP-HPLC inji-
ziert.
Die qualitative und quantitative Identifizierung von Thymosin β4 erfolgte mittels eines vor-
her analysierten Standards mit einer definierten Menge an Thymosin β4 (200 ng), indem die
Flächen unterhalb der Peaks integriert wurden. Es ergab sich folgende Formel zur Berech-
nung der Thymosin β4 Menge:
β
Zusätzlich wurde noch bei einigen Proben durch Oxidation und durch Zugabe kleiner Men-
gen Thymosin β4 sichergestellt, dass der Elutionspeak auch dem für Thymosin β4 entsprach.
Für die Oxidation wurden die Proben mit 1/10 des Volumens an 30 % H2O2 versetzt und für
eine Stunde bei Raumtemperatur inkubiert. Dies führte durch Oxidation des Methioninrestes
im Thymosin β4 zur Bildung eines Sulfoxids, das aufgrund seines polareren Charakters ent-
sprechend den hier angewendeten Bedingungen ca. fünf Minuten früher von der Säule eluier-
te. Die Nachweisgrenze bei diesem Vorgehen lag bei einer Thymosin β4 Konzentration von
0,05 µg/ml. Die Abbildung 23 zeigt typische Chromatogramme für einen Thymosin β4/β10
Standard und eine Sulkusfluidprobe.
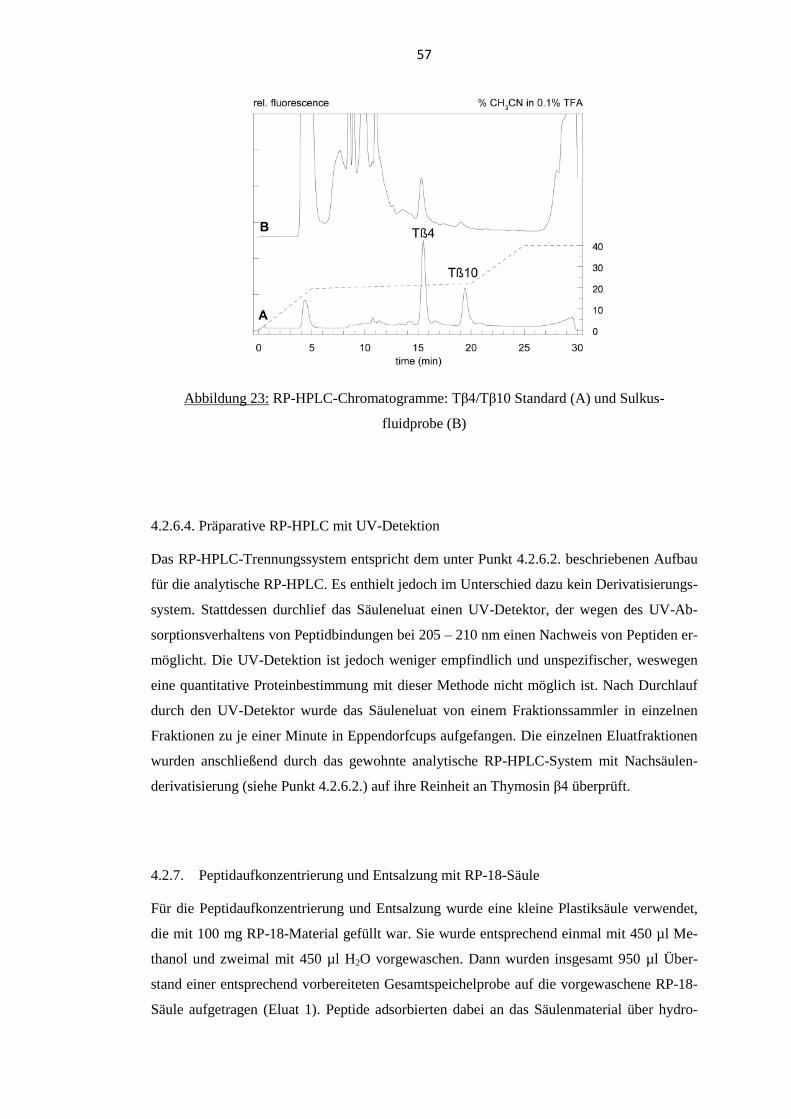
57
Abbildung 23: RP-HPLC-Chromatogramme: Tβ4/Tβ10 Standard (A) und Sulkus-
fluidprobe (B)
4.2.6.4. Präparative RP-HPLC mit UV-Detektion
Das RP-HPLC-Trennungssystem entspricht dem unter Punkt 4.2.6.2. beschriebenen Aufbau
für die analytische RP-HPLC. Es enthielt jedoch im Unterschied dazu kein Derivatisierungs-
system. Stattdessen durchlief das Säuleneluat einen UV-Detektor, der wegen des UV-Ab-
sorptionsverhaltens von Peptidbindungen bei 205 – 210 nm einen Nachweis von Peptiden er-
möglicht. Die UV-Detektion ist jedoch weniger empfindlich und unspezifischer, weswegen
eine quantitative Proteinbestimmung mit dieser Methode nicht möglich ist. Nach Durchlauf
durch den UV-Detektor wurde das Säuleneluat von einem Fraktionssammler in einzelnen
Fraktionen zu je einer Minute in Eppendorfcups aufgefangen. Die einzelnen Eluatfraktionen
wurden anschließend durch das gewohnte analytische RP-HPLC-System mit Nachsäulen-
derivatisierung (siehe Punkt 4.2.6.2.) auf ihre Reinheit an Thymosin β4 überprüft.
4.2.7. Peptidaufkonzentrierung und Entsalzung mit RP-18-Säule
Für die Peptidaufkonzentrierung und Entsalzung wurde eine kleine Plastiksäule verwendet,
die mit 100 mg RP-18-Material gefüllt war. Sie wurde entsprechend einmal mit 450 µl Me-
thanol und zweimal mit 450 µl H2O vorgewaschen. Dann wurden insgesamt 950 µl Über-
stand einer entsprechend vorbereiteten Gesamtspeichelprobe auf die vorgewaschene RP-18-
Säule aufgetragen (Eluat 1). Peptide adsorbierten dabei an das Säulenmaterial über hydro-

58
phobe Wechselwirkungen. Durch das zweimalige Waschen mit je 450 µl H2O wurden Salze
entfernt (Eluat 2 und 3). Danach wurden die gebundenen Peptide durch zweimalige Zugabe
von 450 µl 40 % ACN in H2O von der Säule eluiert (Eluat 4 und 5). Die Analyse der Wasch-
und Eluatfraktionen erfolgte anschließend über die RP-HPLC-Auftrennung und Fluramde-
tektion.
4.2.8. Massenspektrometrie der aufgereinigten Proteine
Die Massenspektrometrie ist eine schnelle und äußerst genaue Methode zur Bestimmung der
molaren Masse eines Moleküls. Es gibt generell verschiedene Prinzipien der Massenspektro-
metrie. Die Massenbestimmung von Thymosin β4 wurde nach dem MALDI-TOF-Prinzip
mit einem Biflex™ III Massenspektrometer durchgeführt. Das MALDI-Verfahren (Matrix-
Assisted Laser Desorption/Ionisation) dient dabei der Ionisierung des Peptids. Dazu werden
etwa 100 ng des entsprechend lypophylisierten Peptids mit α-Cyano-4-hydroxy-Zimtsäure in
0,1 % TFA und 33 % ACN vermischt und auf ein Stahltarget aufgetragen. Beim Trocknen
cokristallisieren organische Moleküle der Matrix mit denen des Peptids. Nach Einschießen
eines Laserpuls (λ = 337 nm) im Hochvakuum kommt es zu einer Freisetzung von ionisier-
ten Matrix- und Proteinmolekülen in die Gasphase. Das TOF-Verfahren (Time Of Flight)
dient nun der Trennung und Analyse des ionisierten Peptids. Hierbei werden die durch den
Laser verdampften positiven Ionen durch eine angelegte Spannung von 19 kV beschleunigt,
reflektiert und schließlich nach entsprechender Flugzeit detektiert. Diese ist proportional
zum Quotienten aus Masse und Ladung (m/z). Zur Kalibrierung dient ein externer Standard
mit bekannten Molekülmassen.
Für eine Massenspektrometrie-Analyse von Thymosin β4 wurden zwei mit oben beschrie-
benen Methoden aufgereinigte Fraktionen einer Gesamtspeichelprobe und einer Schweine-
leukozytenprobe verwendet. Die Proben, deren Konzentration an Thymosin β4 durch vorhe-
rige analytische RP-HPLC ermittelt wurde, wurden zunächst von ca. 700 µl auf 1,5 – 3 µl in
einer SpeedVac konzentriert und anschließend wie oben beschrieben weiterverarbeitet.
4.2.9. Stabilität von Thymosin β4 im Gesamtspeichel
Anhand zweier Gesamtspeichelproben, die unterschiedliche Mengen Thymosin β4 enthielten
und denen Thymosin β4 in einer bekannten Menge zugegeben wurde, sollte die Stabilität
von Thymosin β4 im Speichel über einen längeren Zeitraum beobachtet werden. Somit sollte
ein eventueller Abbau durch Proteasen oder eine Oxidation von Thymosin β4 im Gesamt-

59
speichel ausgeschlossen werden. Die Probengewinnung erfolgte wie unter Punkt 4.2.1.1. be-
schrieben. Anschließend wurden die Proben, wie aus der Abbildung 24 ersichtlich, entspre-
chend präpariert und verarbeitet. Die weitere Probenvorbereitung und -analyse hinsichtlich
der Konzentration an Thymosin β4 entsprach den sonst ebenfalls angewandten Methoden.
Abbildung 24: Versuchsaufbau zur Messung der Stabilität von Tβ4 im Gesamtspeichel

60
5. Ergebnisse
5.1. Thymosin β4 Konzentration im Gesamtspeichel
Zunächst sollen die Ergebnisse der Probenauswertungen der Probanden aus den Gruppen A
und B, bei denen die Proteinkonzentration und die Thymosin β4 Konzentration im Gesamt-
speichel untersucht wurden, dargestellt werden. Die Gruppeneinteilung erfolgte nach den
unter Punkt 3.1. beschriebenen Kriterien. Die Tabellen 11 und 12 geben einen Überblick
über die Zusammensetzung der beiden gebildeten Gruppen A und B.
Gruppe A
Proband Geschlecht Alter
1 M 23
2 M 23
3 M 30
4 M 22
5 M 24
6 W 26
7 W 26
Tabelle 11: Zusammensetzung Gruppe A
Gruppe B
Proband Geschlecht Alter
1 M 55
2 W 54
3 M 60
4 M 49
5 W 57
6 W 56
7 W 46
8 W 51
Tabelle 12: Zusammensetzung Gruppe B
Die Gruppe A umfasste insgesamt sieben Probanden im Alter von 22 – 30 Jahren. Das
Durchschnittsalter betrug 25 ± 3 Jahre. Zwei Probanden waren weiblich, der Rest war männ-
lich.
Die Gruppe B bestand aus acht Probanden im Alter von 46 – 60 Jahren, darunter waren fünf
weiblich. Das Durchschnittsalter dieser Gruppe betrug 54 ± 5 Jahre.
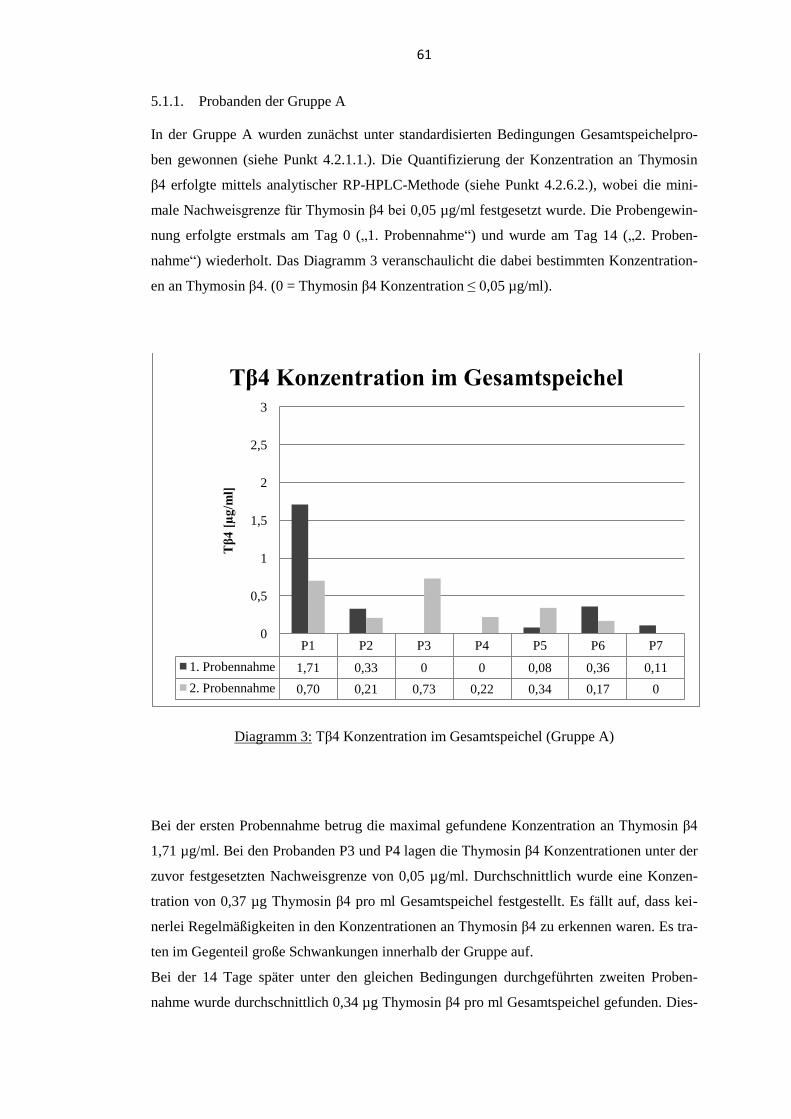
61
5.1.1. Probanden der Gruppe A
In der Gruppe A wurden zunächst unter standardisierten Bedingungen Gesamtspeichelpro-
ben gewonnen (siehe Punkt 4.2.1.1.). Die Quantifizierung der Konzentration an Thymosin
β4 erfolgte mittels analytischer RP-HPLC-Methode (siehe Punkt 4.2.6.2.), wobei die mini-
male Nachweisgrenze für Thymosin β4 bei 0,05 µg/ml festgesetzt wurde. Die Probengewin-
nung erfolgte erstmals am Tag 0 („1. Probennahme“) und wurde am Tag 14 („2. Proben-
nahme“) wiederholt. Das Diagramm 3 veranschaulicht die dabei bestimmten Konzentration-
en an Thymosin β4. (0 = Thymosin β4 Konzentration ≤ 0,05 µg/ml).
Diagramm 3: Tβ4 Konzentration im Gesamtspeichel (Gruppe A)
Bei der ersten Probennahme betrug die maximal gefundene Konzentration an Thymosin β4
1,71 µg/ml. Bei den Probanden P3 und P4 lagen die Thymosin β4 Konzentrationen unter der
zuvor festgesetzten Nachweisgrenze von 0,05 µg/ml. Durchschnittlich wurde eine Konzen-
tration von 0,37 µg Thymosin β4 pro ml Gesamtspeichel festgestellt. Es fällt auf, dass kei-
nerlei Regelmäßigkeiten in den Konzentrationen an Thymosin β4 zu erkennen waren. Es tra-
ten im Gegenteil große Schwankungen innerhalb der Gruppe auf.
Bei der 14 Tage später unter den gleichen Bedingungen durchgeführten zweiten Proben-
nahme wurde durchschnittlich 0,34 µg Thymosin β4 pro ml Gesamtspeichel gefunden. Dies-
P1 P2 P3 P4 P5 P6 P7
1. Probennahme 1,71 0,33 0 0 0,08 0,36 0,11
2. Probennahme 0,70 0,21 0,73 0,22 0,34 0,17 0
0
0,5
1
1,5
2
2,5
3
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im Gesamtspeichel

62
mal lag die Thymosin β4 Konzentration nur bei dem Probanden P7 unterhalb der Nachweis-
grenze. Der Maximalwert der Thymosin β4 Konzentration fiel dieses Mal mit 0,73 µg/ml bei
dem Probanden P3 geringer aus als bei der ersten Probennahme. Wie auch bei der ersten
Probennahme schon beobachtet wurde, schwankten die gefundenen Werte für die Thymosin
β4 Konzentration innerhalb der Gruppe stark und es waren keine Regelmäßigkeiten zu er-
kennen.
Im Vergleich zur ersten Probennahme ergaben sich zahlreiche Auffälligkeiten. So lagen bei
den Probanden P3 und P4 die Thymosin β4 Konzentrationen in der ersten Probennahme un-
terhalb der Nachweisgrenze, in der zweiten Probennahme enthielt ihr Gesamtspeichel jedoch
deutlich höhere Konzentrationen an Thymosin β4. Der Proband P7 wiederum hatte in der
ersten Probennahme eine nachweisbare Konzentration an Thymosin β4 im Gesamtspeichel,
wohingegen in der zweiten Probennahme die Thymosin β4 Konzentration unterhalb der
Nachweisgrenze lag. Insgesamt gesehen kam es bei der Wiederholung der Probenanalyse
nach 14 Tagen also entweder zu einer Zunahme (P3, P4, P5) oder zu einer Abnahme (P1, P2,
P6, P7) der Konzentration an Thymosin β4. Es kam dabei zu einer maximalen Zunahme von
0,73 µg/ml bei dem Probanden P3. Die maximale Abnahme der Konzentration trat bei dem
Probanden P1 mit 1,02 µg/ml auf. Am geringsten veränderte sich die Konzentration bei dem
Probanden P2 mit einer Abnahme der Konzentration um 0,09 µg/ml. Die Konzentrationen an
Thymosin β4 im Gesamtspeichel waren somit über die Zeit betrachtet sehr variabel. Das
Diagramm 4 verdeutlicht die bei der zweiten Probennahme im Vergleich zur ersten Proben-
nahme festgestellten Veränderungen der Konzentration an Thymosin β4 im Gesamtspeichel.
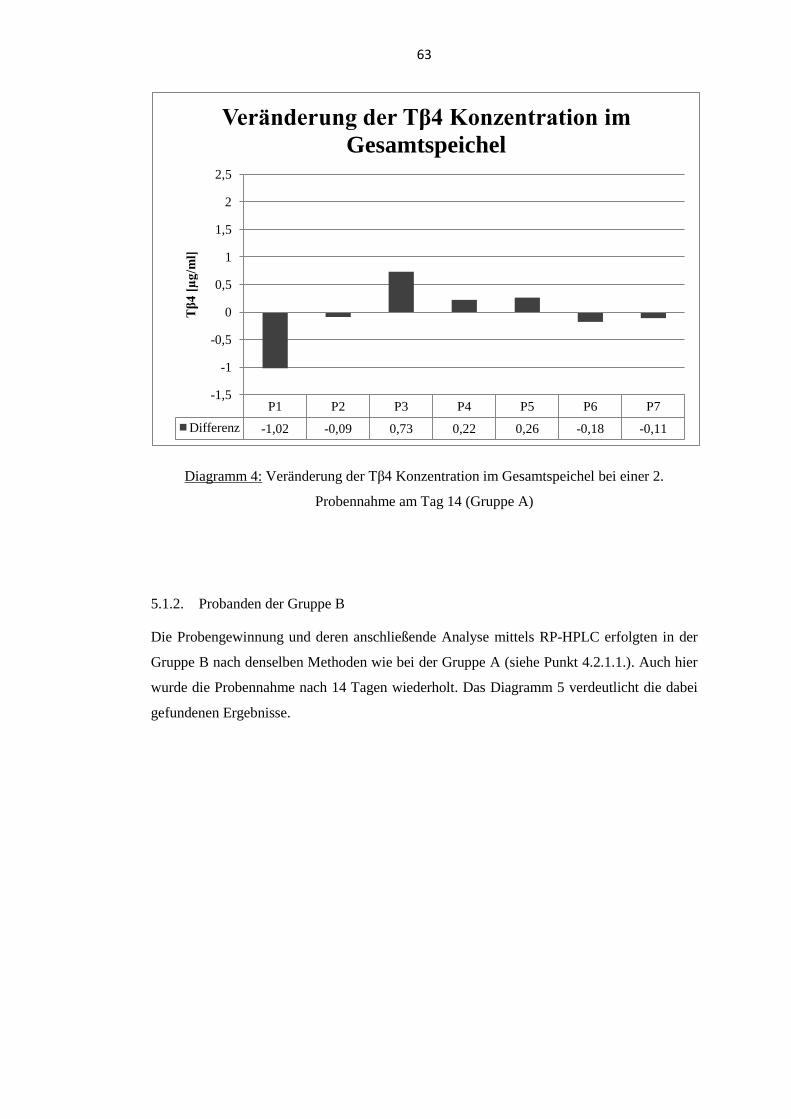
63
Diagramm 4: Veränderung der Tβ4 Konzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe A)
5.1.2. Probanden der Gruppe B
Die Probengewinnung und deren anschließende Analyse mittels RP-HPLC erfolgten in der
Gruppe B nach denselben Methoden wie bei der Gruppe A (siehe Punkt 4.2.1.1.). Auch hier
wurde die Probennahme nach 14 Tagen wiederholt. Das Diagramm 5 verdeutlicht die dabei
gefundenen Ergebnisse.
P1 P2 P3 P4 P5 P6 P7
Differenz -1,02 -0,09 0,73 0,22 0,26 -0,18 -0,11
-1,5
-1
-0,5
0
0,5
1
1,5
2
2,5T
β4
[µ
g/m
l]
Veränderung der Tβ4 Konzentration im
Gesamtspeichel
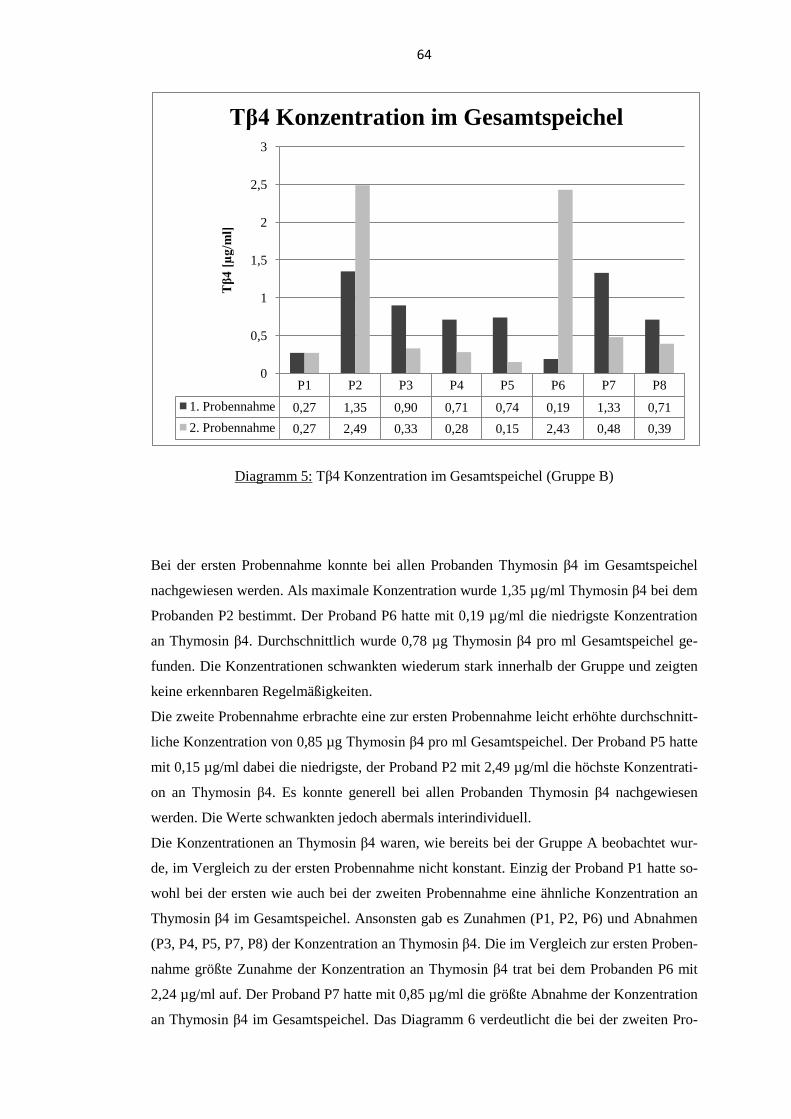
64
Diagramm 5: Tβ4 Konzentration im Gesamtspeichel (Gruppe B)
Bei der ersten Probennahme konnte bei allen Probanden Thymosin β4 im Gesamtspeichel
nachgewiesen werden. Als maximale Konzentration wurde 1,35 µg/ml Thymosin β4 bei dem
Probanden P2 bestimmt. Der Proband P6 hatte mit 0,19 µg/ml die niedrigste Konzentration
an Thymosin β4. Durchschnittlich wurde 0,78 µg Thymosin β4 pro ml Gesamtspeichel ge-
funden. Die Konzentrationen schwankten wiederum stark innerhalb der Gruppe und zeigten
keine erkennbaren Regelmäßigkeiten.
Die zweite Probennahme erbrachte eine zur ersten Probennahme leicht erhöhte durchschnitt-
liche Konzentration von 0,85 µg Thymosin β4 pro ml Gesamtspeichel. Der Proband P5 hatte
mit 0,15 µg/ml dabei die niedrigste, der Proband P2 mit 2,49 µg/ml die höchste Konzentrati-
on an Thymosin β4. Es konnte generell bei allen Probanden Thymosin β4 nachgewiesen
werden. Die Werte schwankten jedoch abermals interindividuell.
Die Konzentrationen an Thymosin β4 waren, wie bereits bei der Gruppe A beobachtet wur-
de, im Vergleich zu der ersten Probennahme nicht konstant. Einzig der Proband P1 hatte so-
wohl bei der ersten wie auch bei der zweiten Probennahme eine ähnliche Konzentration an
Thymosin β4 im Gesamtspeichel. Ansonsten gab es Zunahmen (P1, P2, P6) und Abnahmen
(P3, P4, P5, P7, P8) der Konzentration an Thymosin β4. Die im Vergleich zur ersten Proben-
nahme größte Zunahme der Konzentration an Thymosin β4 trat bei dem Probanden P6 mit
2,24 µg/ml auf. Der Proband P7 hatte mit 0,85 µg/ml die größte Abnahme der Konzentration
an Thymosin β4 im Gesamtspeichel. Das Diagramm 6 verdeutlicht die bei der zweiten Pro-
P1 P2 P3 P4 P5 P6 P7 P8
1. Probennahme 0,27 1,35 0,90 0,71 0,74 0,19 1,33 0,71
2. Probennahme 0,27 2,49 0,33 0,28 0,15 2,43 0,48 0,39
0
0,5
1
1,5
2
2,5
3
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im Gesamtspeichel
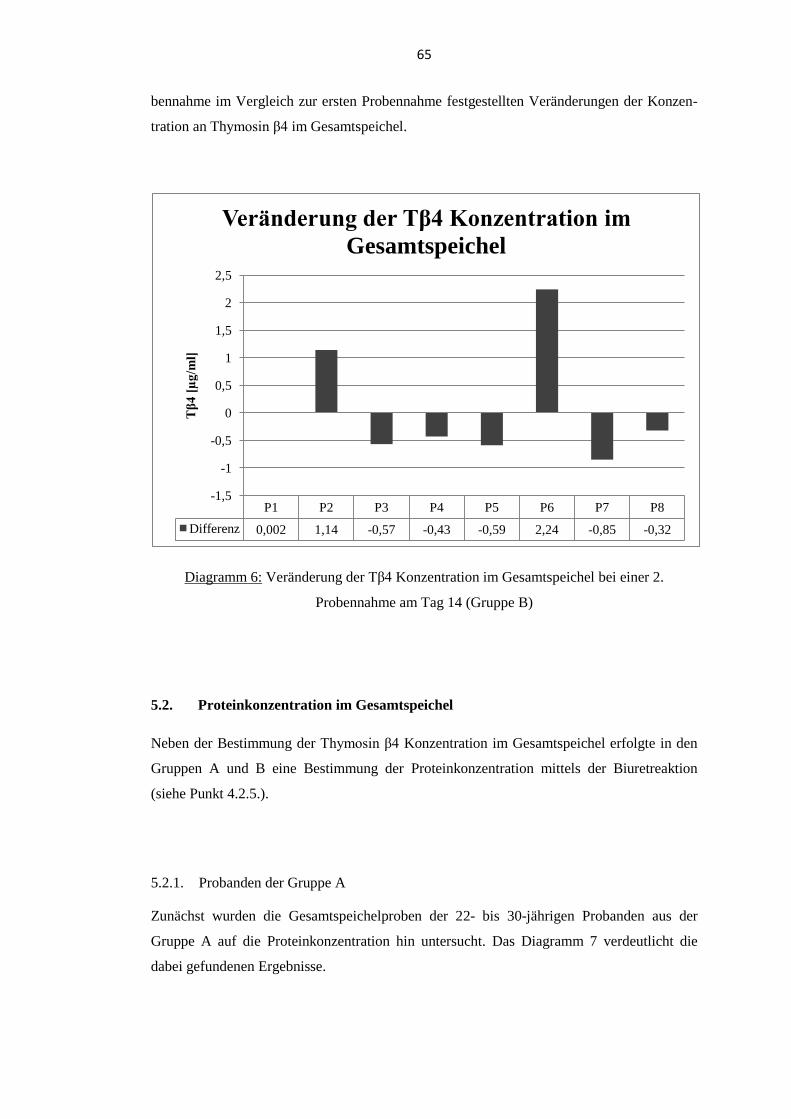
65
bennahme im Vergleich zur ersten Probennahme festgestellten Veränderungen der Konzen-
tration an Thymosin β4 im Gesamtspeichel.
Diagramm 6: Veränderung der Tβ4 Konzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe B)
5.2. Proteinkonzentration im Gesamtspeichel
Neben der Bestimmung der Thymosin β4 Konzentration im Gesamtspeichel erfolgte in den
Gruppen A und B eine Bestimmung der Proteinkonzentration mittels der Biuretreaktion
(siehe Punkt 4.2.5.).
5.2.1. Probanden der Gruppe A
Zunächst wurden die Gesamtspeichelproben der 22- bis 30-jährigen Probanden aus der
Gruppe A auf die Proteinkonzentration hin untersucht. Das Diagramm 7 verdeutlicht die
dabei gefundenen Ergebnisse.
P1 P2 P3 P4 P5 P6 P7 P8
Differenz 0,002 1,14 -0,57 -0,43 -0,59 2,24 -0,85 -0,32
-1,5
-1
-0,5
0
0,5
1
1,5
2
2,5
Tβ
4 [
µg
/ml]
Veränderung der Tβ4 Konzentration im
Gesamtspeichel
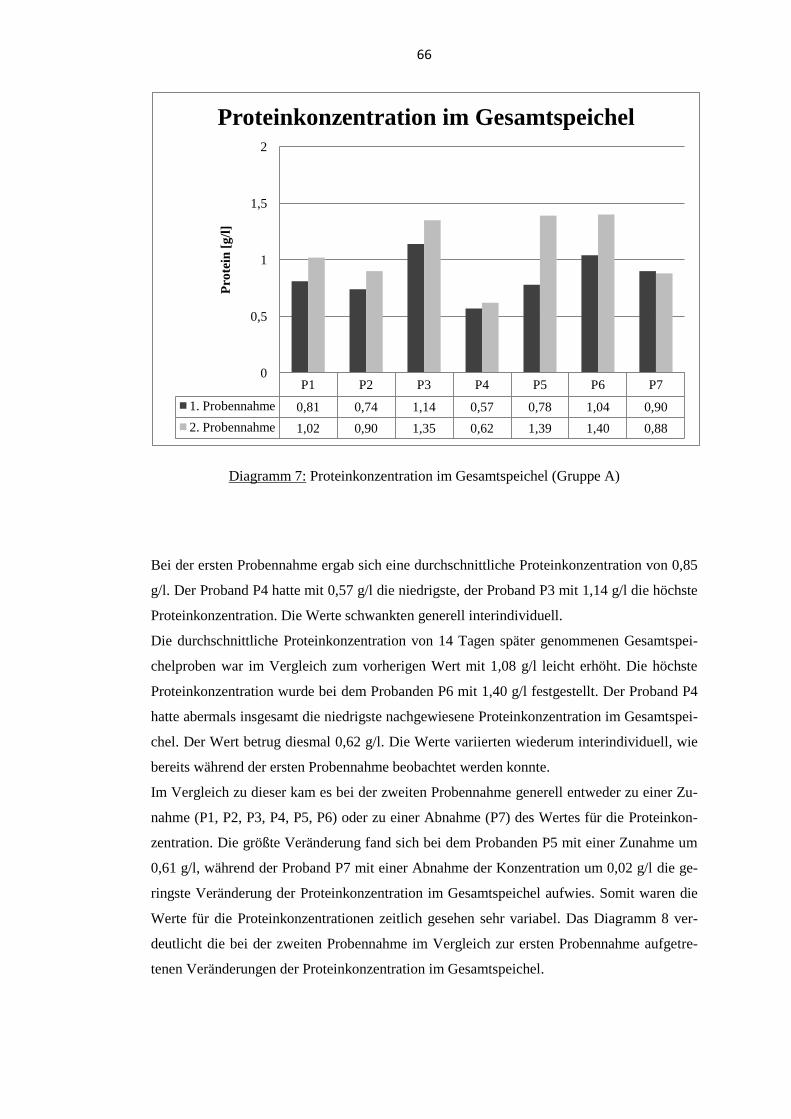
66
Diagramm 7: Proteinkonzentration im Gesamtspeichel (Gruppe A)
Bei der ersten Probennahme ergab sich eine durchschnittliche Proteinkonzentration von 0,85
g/l. Der Proband P4 hatte mit 0,57 g/l die niedrigste, der Proband P3 mit 1,14 g/l die höchste
Proteinkonzentration. Die Werte schwankten generell interindividuell.
Die durchschnittliche Proteinkonzentration von 14 Tagen später genommenen Gesamtspei-
chelproben war im Vergleich zum vorherigen Wert mit 1,08 g/l leicht erhöht. Die höchste
Proteinkonzentration wurde bei dem Probanden P6 mit 1,40 g/l festgestellt. Der Proband P4
hatte abermals insgesamt die niedrigste nachgewiesene Proteinkonzentration im Gesamtspei-
chel. Der Wert betrug diesmal 0,62 g/l. Die Werte variierten wiederum interindividuell, wie
bereits während der ersten Probennahme beobachtet werden konnte.
Im Vergleich zu dieser kam es bei der zweiten Probennahme generell entweder zu einer Zu-
nahme (P1, P2, P3, P4, P5, P6) oder zu einer Abnahme (P7) des Wertes für die Proteinkon-
zentration. Die größte Veränderung fand sich bei dem Probanden P5 mit einer Zunahme um
0,61 g/l, während der Proband P7 mit einer Abnahme der Konzentration um 0,02 g/l die ge-
ringste Veränderung der Proteinkonzentration im Gesamtspeichel aufwies. Somit waren die
Werte für die Proteinkonzentrationen zeitlich gesehen sehr variabel. Das Diagramm 8 ver-
deutlicht die bei der zweiten Probennahme im Vergleich zur ersten Probennahme aufgetre-
tenen Veränderungen der Proteinkonzentration im Gesamtspeichel.
P1 P2 P3 P4 P5 P6 P7
1. Probennahme 0,81 0,74 1,14 0,57 0,78 1,04 0,90
2. Probennahme 1,02 0,90 1,35 0,62 1,39 1,40 0,88
0
0,5
1
1,5
2
Pro
tein
[g
/l]
Proteinkonzentration im Gesamtspeichel
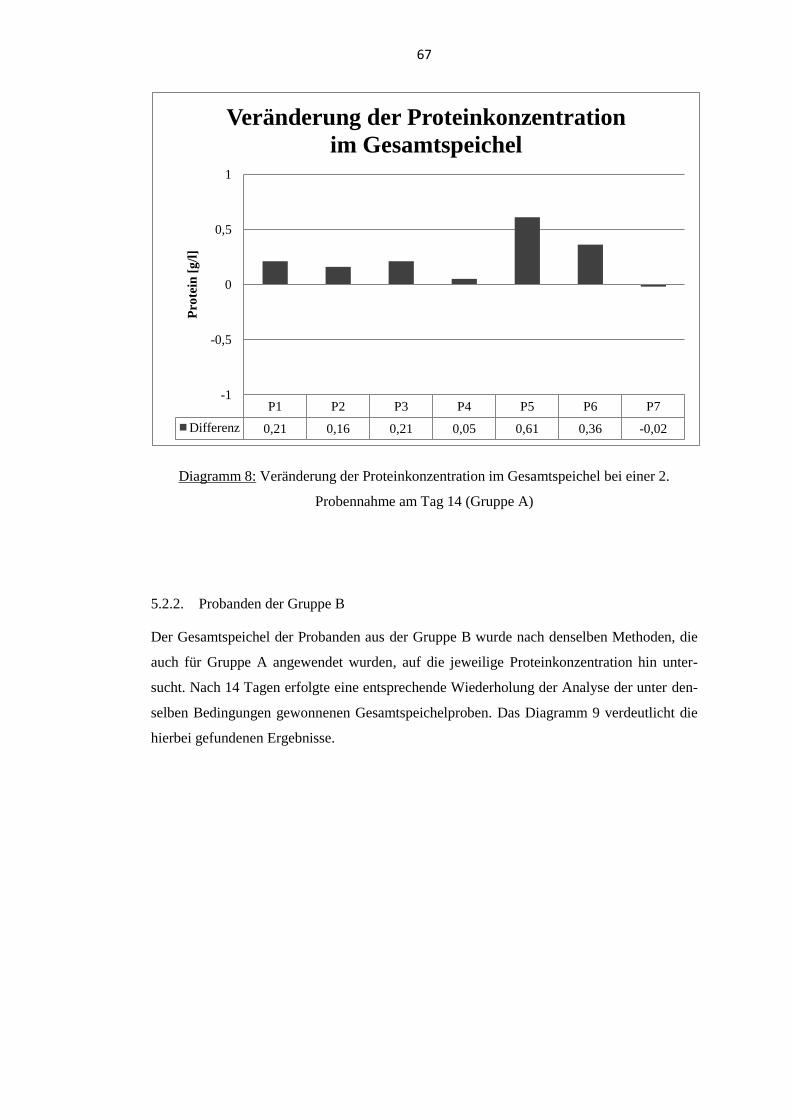
67
Diagramm 8: Veränderung der Proteinkonzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe A)
5.2.2. Probanden der Gruppe B
Der Gesamtspeichel der Probanden aus der Gruppe B wurde nach denselben Methoden, die
auch für Gruppe A angewendet wurden, auf die jeweilige Proteinkonzentration hin unter-
sucht. Nach 14 Tagen erfolgte eine entsprechende Wiederholung der Analyse der unter den-
selben Bedingungen gewonnenen Gesamtspeichelproben. Das Diagramm 9 verdeutlicht die
hierbei gefundenen Ergebnisse.
P1 P2 P3 P4 P5 P6 P7
Differenz 0,21 0,16 0,21 0,05 0,61 0,36 -0,02
-1
-0,5
0
0,5
1
Pro
tein
[g
/l]
Veränderung der Proteinkonzentration
im Gesamtspeichel
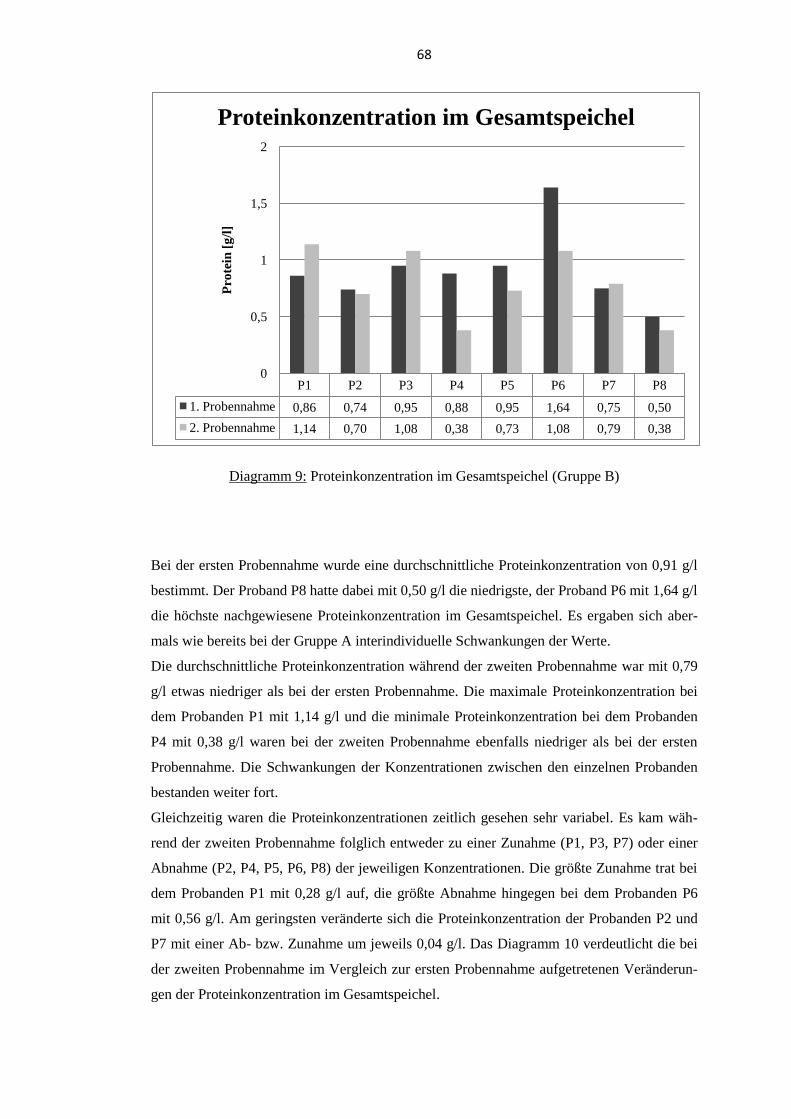
68
Diagramm 9: Proteinkonzentration im Gesamtspeichel (Gruppe B)
Bei der ersten Probennahme wurde eine durchschnittliche Proteinkonzentration von 0,91 g/l
bestimmt. Der Proband P8 hatte dabei mit 0,50 g/l die niedrigste, der Proband P6 mit 1,64 g/l
die höchste nachgewiesene Proteinkonzentration im Gesamtspeichel. Es ergaben sich aber-
mals wie bereits bei der Gruppe A interindividuelle Schwankungen der Werte.
Die durchschnittliche Proteinkonzentration während der zweiten Probennahme war mit 0,79
g/l etwas niedriger als bei der ersten Probennahme. Die maximale Proteinkonzentration bei
dem Probanden P1 mit 1,14 g/l und die minimale Proteinkonzentration bei dem Probanden
P4 mit 0,38 g/l waren bei der zweiten Probennahme ebenfalls niedriger als bei der ersten
Probennahme. Die Schwankungen der Konzentrationen zwischen den einzelnen Probanden
bestanden weiter fort.
Gleichzeitig waren die Proteinkonzentrationen zeitlich gesehen sehr variabel. Es kam wäh-
rend der zweiten Probennahme folglich entweder zu einer Zunahme (P1, P3, P7) oder einer
Abnahme (P2, P4, P5, P6, P8) der jeweiligen Konzentrationen. Die größte Zunahme trat bei
dem Probanden P1 mit 0,28 g/l auf, die größte Abnahme hingegen bei dem Probanden P6
mit 0,56 g/l. Am geringsten veränderte sich die Proteinkonzentration der Probanden P2 und
P7 mit einer Ab- bzw. Zunahme um jeweils 0,04 g/l. Das Diagramm 10 verdeutlicht die bei
der zweiten Probennahme im Vergleich zur ersten Probennahme aufgetretenen Veränderun-
gen der Proteinkonzentration im Gesamtspeichel.
P1 P2 P3 P4 P5 P6 P7 P8
1. Probennahme 0,86 0,74 0,95 0,88 0,95 1,64 0,75 0,50
2. Probennahme 1,14 0,70 1,08 0,38 0,73 1,08 0,79 0,38
0
0,5
1
1,5
2
Pro
tein
[g
/l]
Proteinkonzentration im Gesamtspeichel
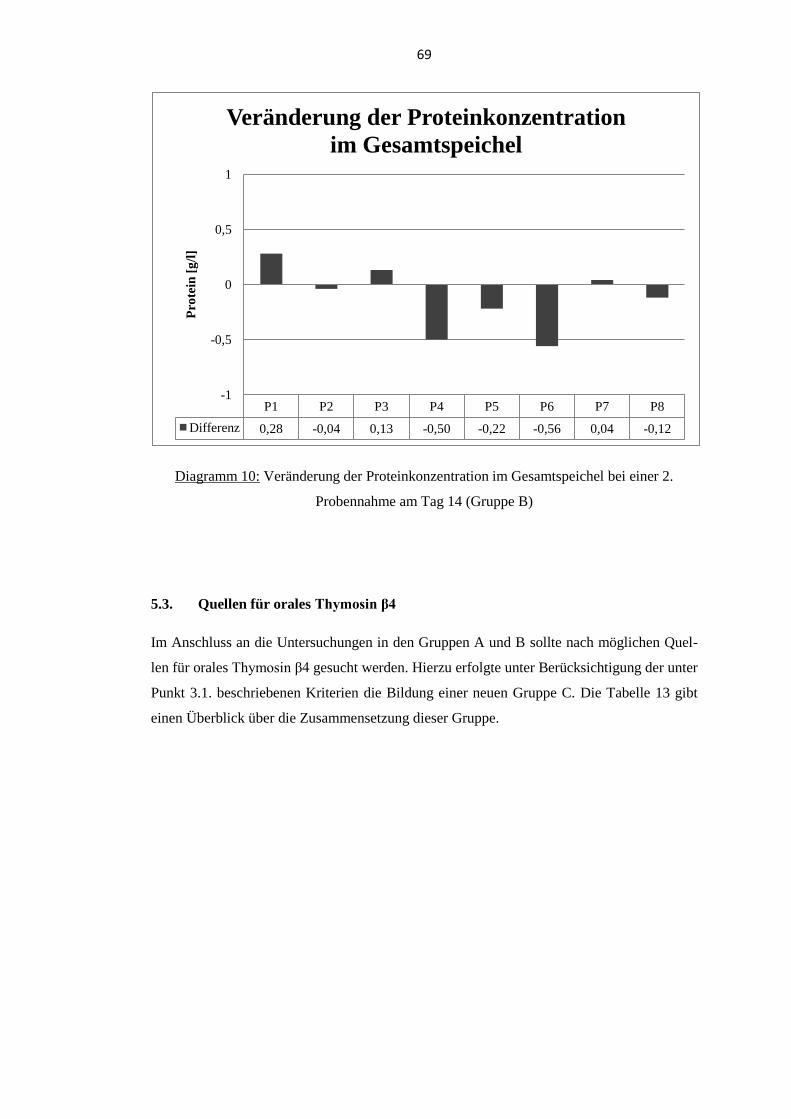
69
Diagramm 10: Veränderung der Proteinkonzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe B)
5.3. Quellen für orales Thymosin β4
Im Anschluss an die Untersuchungen in den Gruppen A und B sollte nach möglichen Quel-
len für orales Thymosin β4 gesucht werden. Hierzu erfolgte unter Berücksichtigung der unter
Punkt 3.1. beschriebenen Kriterien die Bildung einer neuen Gruppe C. Die Tabelle 13 gibt
einen Überblick über die Zusammensetzung dieser Gruppe.
P1 P2 P3 P4 P5 P6 P7 P8
Differenz 0,28 -0,04 0,13 -0,50 -0,22 -0,56 0,04 -0,12
-1
-0,5
0
0,5
1P
rote
in [
g/l
]
Veränderung der Proteinkonzentration
im Gesamtspeichel

70
Gruppe C
Proband Geschlecht Alter
1 W 26
2 W 22
3 M 25
4 M 22
5 W 22
6 M 24
7 W 23
8 M 26
9 M 22
10 M 23
11 M 23
Tabelle 13: Zusammensetzung Gruppe C
Die Gruppe C umfasste insgesamt elf Probanden im Alter von 22 – 26 Jahren. Das Durch-
schnittsalter betrug 23 ± 2 Jahre. Vier Probanden waren weiblich, der Rest war männlich.
Kriterium für die Zugehörigkeit zu dieser Gruppe war parodontale Gesundheit.
In dieser Gruppe wurde im Rahmen der Suche nach möglichen Quellen für orales Thymosin
β4 eine Reihe verschiedener Untersuchungen durchgeführt. So wurden nacheinander, drü-
senspezifischer Speichel und schließlich Sulkusfluid auf den Gehalt an Thymosin β4 unter-
sucht.
5.3.1. Thymosin β4 Konzentration im Gesamtspeichel
Von den Probanden der Gruppe C wurden zunächst Gesamtspeichelproben abgenommen
(siehe Punkt 4.2.1.2.). Die Methode der Probengewinnung unterschied sich dabei jedoch von
der bisher in den Gruppen A und B angewandten Methode (siehe Punkt 4.2.1.1.). Die Analy-
se der Proben erfolgte nach entsprechender Probenvorbereitung mittels RP-HPLC. Ein typi-
sches Chromatogramm für eine Gesamtspeichelprobe und einen Thymosin β4 Standard ist
unter der Abbildung 25 dargestellt.
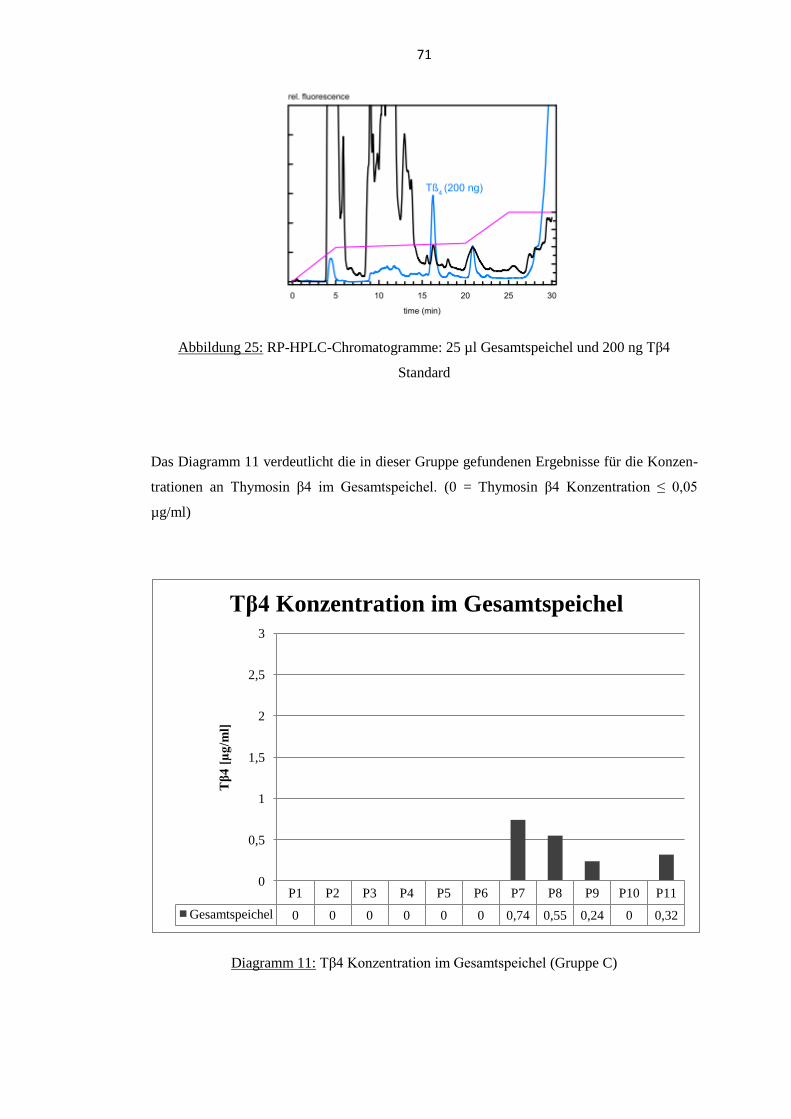
71
Abbildung 25: RP-HPLC-Chromatogramme: 25 µl Gesamtspeichel und 200 ng Tβ4
Standard
Das Diagramm 11 verdeutlicht die in dieser Gruppe gefundenen Ergebnisse für die Konzen-
trationen an Thymosin β4 im Gesamtspeichel. (0 = Thymosin β4 Konzentration ≤ 0,05
µg/ml)
Diagramm 11: Tβ4 Konzentration im Gesamtspeichel (Gruppe C)
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11
Gesamtspeichel 0 0 0 0 0 0 0,74 0,55 0,24 0 0,32
0
0,5
1
1,5
2
2,5
3
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im Gesamtspeichel
72
Insgesamt konnte in den Gesamtspeichelproben sieben Mal kein Thymosin β4 in nachweis-
baren Mengen detektiert werden (P1, P2, P3, P4, P5, P6, P10). Bei den übrigen vier Proban-
den (P7, P8, P9, P11) wurde durchschnittlich 0,46 µg/ml Thymosin β4 nachgewiesen. Der
Proband P7 hatte dabei mit 0,74 µg Thymosin β4 pro ml Gesamtspeichel den höchsten Wert,
der Proband P9 mit 0,24 µg/ml den niedrigsten Wert. Die Werte zeigten interindividuelle
Schwankungen, die jedoch keiner erkennbaren Regelmäßigkeit unterlagen.
5.3.2. Thymosin β4 Konzentration im Speichel der gl. parotis
Nach der entsprechenden Probengewinnung und Probenvorbereitung (siehe Punkt 4.2.1.3.
und Punkt 4.2.4.1.) wurde je Proband eine Probe der linken und rechten gl. parotis mittels
RP-HPLC untersucht. Dabei konnten bei keinem Probanden relevante Mengen an Thymosin
β4 detektiert werden. Die jeweiligen Chromatogramme glichen alle einander und zeigten
keine Peaks bei 15 Minuten. Die Abbildung 26 stellt exemplarisch die Chromatogramme
einer gl. parotis Speichelprobe und eines Thymosin β4 Standards dar.
Abbildung 26: RP-HPLC-Chromatogramme: 25 µl Speichel gl. parotis und 200 ng
Tβ4 Standard
5.3.3. Thymosin β4 Konzentration im Speichel der gl. submandibularis/sublingualis
Von jedem Probanden wurde zudem eine Probe eines Speichelgemisches aus gl. submandi-
bularis und gl. sublingualis entnommen (siehe Punkt 4.2.1.3.). Diese wurde entsprechend
vorbereitet (siehe Punkt 4.2.4.1.) und mittels RP-HPLC auf den Gehalt an Thymosin β4 un-
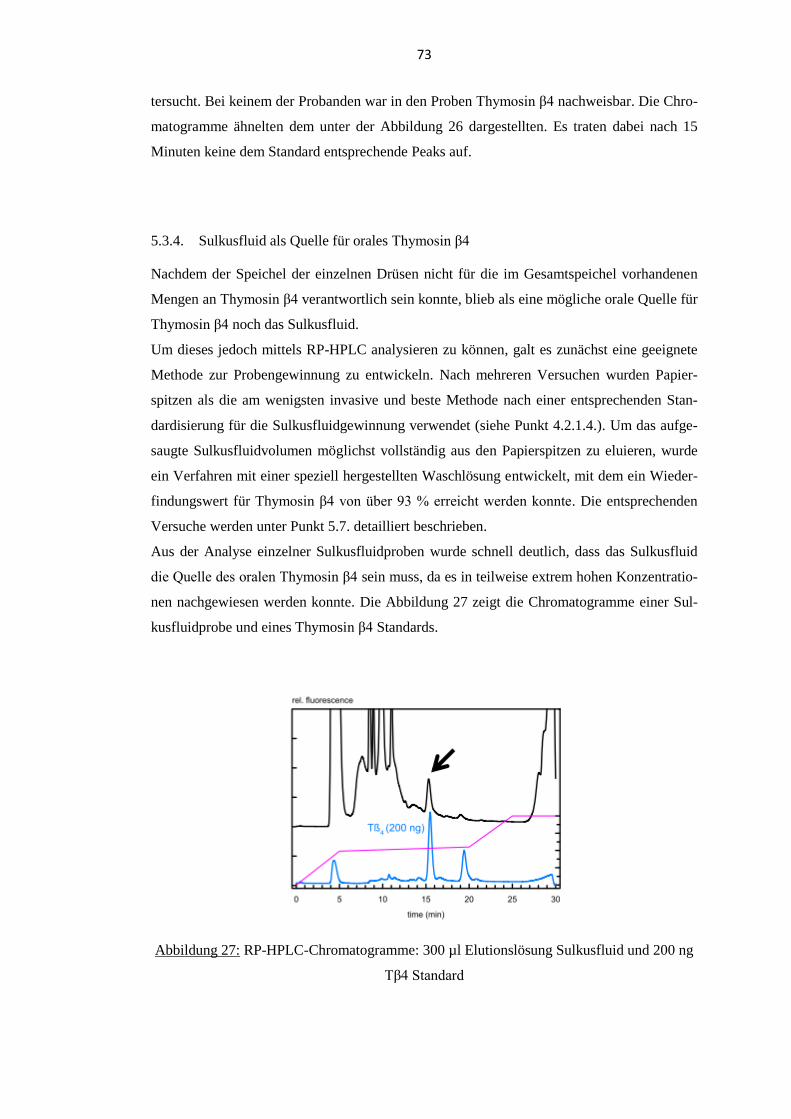
73
tersucht. Bei keinem der Probanden war in den Proben Thymosin β4 nachweisbar. Die Chro-
matogramme ähnelten dem unter der Abbildung 26 dargestellten. Es traten dabei nach 15
Minuten keine dem Standard entsprechende Peaks auf.
5.3.4. Sulkusfluid als Quelle für orales Thymosin β4
Nachdem der Speichel der einzelnen Drüsen nicht für die im Gesamtspeichel vorhandenen
Mengen an Thymosin β4 verantwortlich sein konnte, blieb als eine mögliche orale Quelle für
Thymosin β4 noch das Sulkusfluid.
Um dieses jedoch mittels RP-HPLC analysieren zu können, galt es zunächst eine geeignete
Methode zur Probengewinnung zu entwickeln. Nach mehreren Versuchen wurden Papier-
spitzen als die am wenigsten invasive und beste Methode nach einer entsprechenden Stan-
dardisierung für die Sulkusfluidgewinnung verwendet (siehe Punkt 4.2.1.4.). Um das aufge-
saugte Sulkusfluidvolumen möglichst vollständig aus den Papierspitzen zu eluieren, wurde
ein Verfahren mit einer speziell hergestellten Waschlösung entwickelt, mit dem ein Wieder-
findungswert für Thymosin β4 von über 93 % erreicht werden konnte. Die entsprechenden
Versuche werden unter Punkt 5.7. detailliert beschrieben.
Aus der Analyse einzelner Sulkusfluidproben wurde schnell deutlich, dass das Sulkusfluid
die Quelle des oralen Thymosin β4 sein muss, da es in teilweise extrem hohen Konzentratio-
nen nachgewiesen werden konnte. Die Abbildung 27 zeigt die Chromatogramme einer Sul-
kusfluidprobe und eines Thymosin β4 Standards.
Abbildung 27: RP-HPLC-Chromatogramme: 300 µl Elutionslösung Sulkusfluid und 200 ng
Tβ4 Standard

74
Die weitere Untersuchung des oralen Thymosin β4 beschränkte sich demnach fortan auf die
Analyse von Sulkusfluidproben der jeweiligen Probanden, die je nach Fragestellung in unter-
schiedliche Gruppen eingeteilt wurden. Dabei sollte zunächst eine mögliche Abhängigkeit
der Konzentration an Thymosin β4 im Sulkusfluid von Parametern wie dem Parodontalstatus
untersucht werden, bzw. nach möglichen Freisetzungsmechanismen des Thymosin β4 in das
Sulkusfluid gesucht werden.
5.4. Thymosin β4 im Sulkusfluid parodontal gesunder Probanden
Es wurden zunächst Sulkusfluidproben von den Probanden aus der Gruppe C untersucht.
Diese hatten zum Zeitpunkt der Untersuchung entsprechend der unter Punkt 3.1. beschrie-
benen Kriterien keine parodontalen Erkrankungen.
5.4.1. Sulkusfluidvolumen
Die Sulkusfluidproben wurden zunächst mittels standardisierter Methode von den Probanden
gewonnen (siehe Punkt 4.2.1.4.). Das Volumen des in der Papierspitze aufgesaugten Sulkus-
fluids wurde durch entsprechendes Auswiegen der Eppendorfcups vor bzw. nach der Proben-
nahme bestimmt. Das Diagramm 12 verdeutlicht die hierbei gefundenen Ergebnisse.
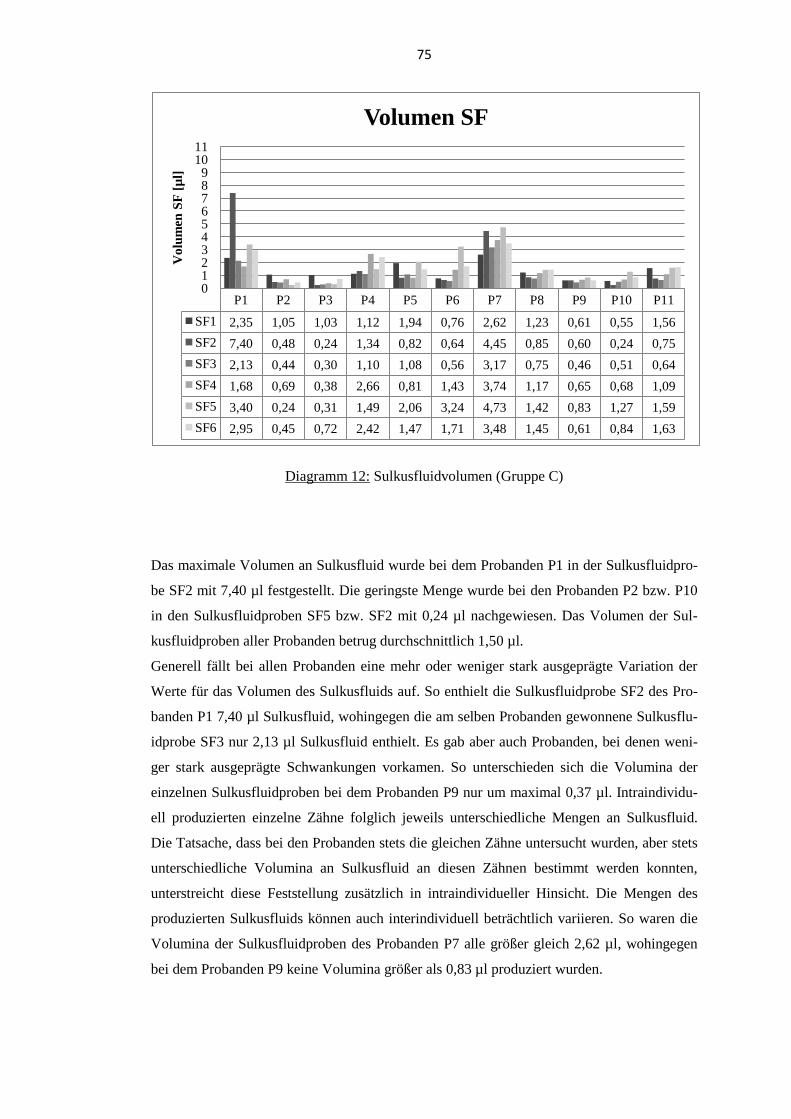
75
Diagramm 12: Sulkusfluidvolumen (Gruppe C)
Das maximale Volumen an Sulkusfluid wurde bei dem Probanden P1 in der Sulkusfluidpro-
be SF2 mit 7,40 µl festgestellt. Die geringste Menge wurde bei den Probanden P2 bzw. P10
in den Sulkusfluidproben SF5 bzw. SF2 mit 0,24 µl nachgewiesen. Das Volumen der Sul-
kusfluidproben aller Probanden betrug durchschnittlich 1,50 µl.
Generell fällt bei allen Probanden eine mehr oder weniger stark ausgeprägte Variation der
Werte für das Volumen des Sulkusfluids auf. So enthielt die Sulkusfluidprobe SF2 des Pro-
banden P1 7,40 µl Sulkusfluid, wohingegen die am selben Probanden gewonnene Sulkusflu-
idprobe SF3 nur 2,13 µl Sulkusfluid enthielt. Es gab aber auch Probanden, bei denen weni-
ger stark ausgeprägte Schwankungen vorkamen. So unterschieden sich die Volumina der
einzelnen Sulkusfluidproben bei dem Probanden P9 nur um maximal 0,37 µl. Intraindividu-
ell produzierten einzelne Zähne folglich jeweils unterschiedliche Mengen an Sulkusfluid.
Die Tatsache, dass bei den Probanden stets die gleichen Zähne untersucht wurden, aber stets
unterschiedliche Volumina an Sulkusfluid an diesen Zähnen bestimmt werden konnten,
unterstreicht diese Feststellung zusätzlich in intraindividueller Hinsicht. Die Mengen des
produzierten Sulkusfluids können auch interindividuell beträchtlich variieren. So waren die
Volumina der Sulkusfluidproben des Probanden P7 alle größer gleich 2,62 µl, wohingegen
bei dem Probanden P9 keine Volumina größer als 0,83 µl produziert wurden.
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11
SF1 2,35 1,05 1,03 1,12 1,94 0,76 2,62 1,23 0,61 0,55 1,56
SF2 7,40 0,48 0,24 1,34 0,82 0,64 4,45 0,85 0,60 0,24 0,75
SF3 2,13 0,44 0,30 1,10 1,08 0,56 3,17 0,75 0,46 0,51 0,64
SF4 1,68 0,69 0,38 2,66 0,81 1,43 3,74 1,17 0,65 0,68 1,09
SF5 3,40 0,24 0,31 1,49 2,06 3,24 4,73 1,42 0,83 1,27 1,59
SF6 2,95 0,45 0,72 2,42 1,47 1,71 3,48 1,45 0,61 0,84 1,63
0123456789
1011
Vo
lum
en S
F [
µl]
Volumen SF
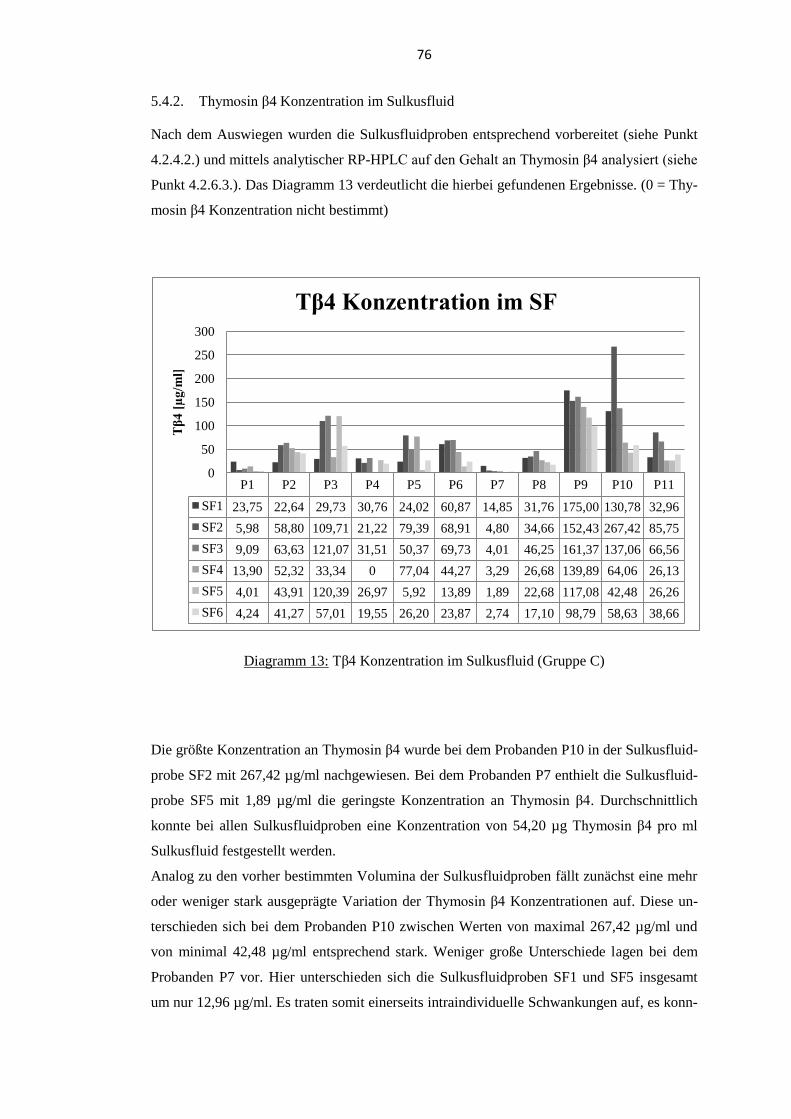
76
5.4.2. Thymosin β4 Konzentration im Sulkusfluid
Nach dem Auswiegen wurden die Sulkusfluidproben entsprechend vorbereitet (siehe Punkt
4.2.4.2.) und mittels analytischer RP-HPLC auf den Gehalt an Thymosin β4 analysiert (siehe
Punkt 4.2.6.3.). Das Diagramm 13 verdeutlicht die hierbei gefundenen Ergebnisse. (0 = Thy-
mosin β4 Konzentration nicht bestimmt)
Diagramm 13: Tβ4 Konzentration im Sulkusfluid (Gruppe C)
Die größte Konzentration an Thymosin β4 wurde bei dem Probanden P10 in der Sulkusfluid-
probe SF2 mit 267,42 µg/ml nachgewiesen. Bei dem Probanden P7 enthielt die Sulkusfluid-
probe SF5 mit 1,89 µg/ml die geringste Konzentration an Thymosin β4. Durchschnittlich
konnte bei allen Sulkusfluidproben eine Konzentration von 54,20 µg Thymosin β4 pro ml
Sulkusfluid festgestellt werden.
Analog zu den vorher bestimmten Volumina der Sulkusfluidproben fällt zunächst eine mehr
oder weniger stark ausgeprägte Variation der Thymosin β4 Konzentrationen auf. Diese un-
terschieden sich bei dem Probanden P10 zwischen Werten von maximal 267,42 µg/ml und
von minimal 42,48 µg/ml entsprechend stark. Weniger große Unterschiede lagen bei dem
Probanden P7 vor. Hier unterschieden sich die Sulkusfluidproben SF1 und SF5 insgesamt
um nur 12,96 µg/ml. Es traten somit einerseits intraindividuelle Schwankungen auf, es konn-
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11
SF1 23,75 22,64 29,73 30,76 24,02 60,87 14,85 31,76 175,00 130,78 32,96
SF2 5,98 58,80 109,71 21,22 79,39 68,91 4,80 34,66 152,43 267,42 85,75
SF3 9,09 63,63 121,07 31,51 50,37 69,73 4,01 46,25 161,37 137,06 66,56
SF4 13,90 52,32 33,34 0 77,04 44,27 3,29 26,68 139,89 64,06 26,13
SF5 4,01 43,91 120,39 26,97 5,92 13,89 1,89 22,68 117,08 42,48 26,26
SF6 4,24 41,27 57,01 19,55 26,20 23,87 2,74 17,10 98,79 58,63 38,66
0
50
100
150
200
250
300
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im SF
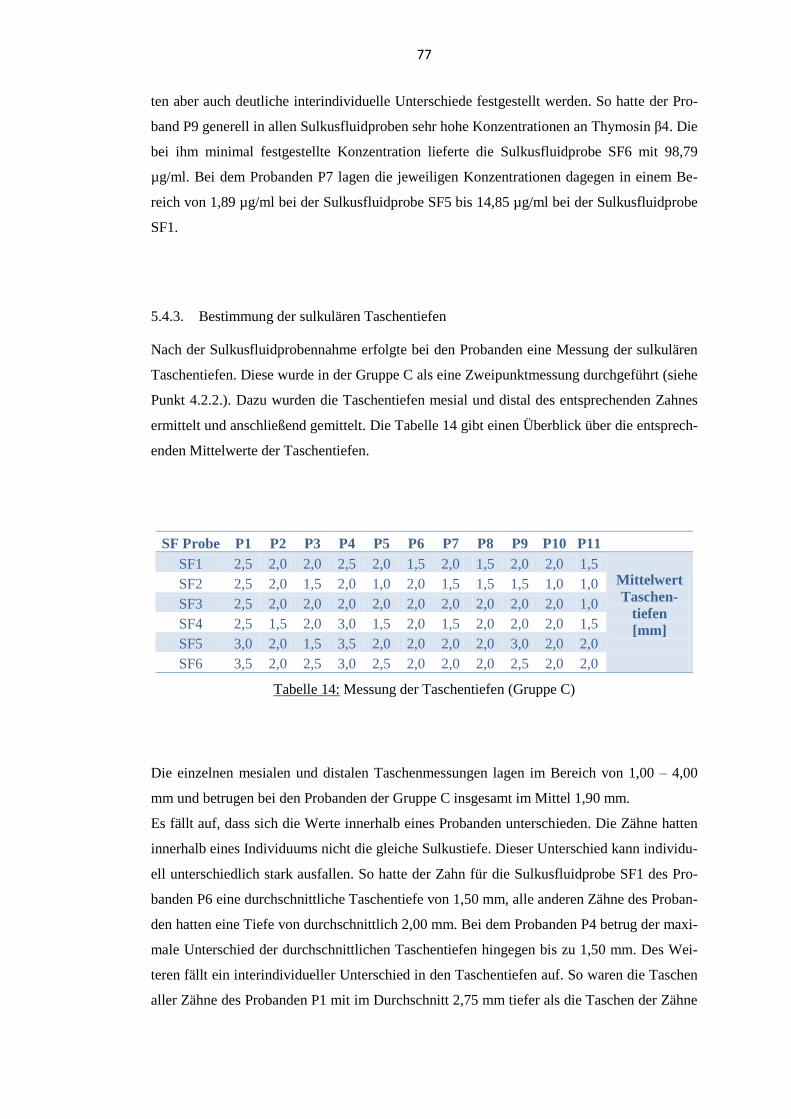
77
ten aber auch deutliche interindividuelle Unterschiede festgestellt werden. So hatte der Pro-
band P9 generell in allen Sulkusfluidproben sehr hohe Konzentrationen an Thymosin β4. Die
bei ihm minimal festgestellte Konzentration lieferte die Sulkusfluidprobe SF6 mit 98,79
µg/ml. Bei dem Probanden P7 lagen die jeweiligen Konzentrationen dagegen in einem Be-
reich von 1,89 µg/ml bei der Sulkusfluidprobe SF5 bis 14,85 µg/ml bei der Sulkusfluidprobe
SF1.
5.4.3. Bestimmung der sulkulären Taschentiefen
Nach der Sulkusfluidprobennahme erfolgte bei den Probanden eine Messung der sulkulären
Taschentiefen. Diese wurde in der Gruppe C als eine Zweipunktmessung durchgeführt (siehe
Punkt 4.2.2.). Dazu wurden die Taschentiefen mesial und distal des entsprechenden Zahnes
ermittelt und anschließend gemittelt. Die Tabelle 14 gibt einen Überblick über die entsprech-
enden Mittelwerte der Taschentiefen.
SF Probe P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11
SF1 2,5 2,0 2,0 2,5 2,0 1,5 2,0 1,5 2,0 2,0 1,5 Mittelwert
Taschen-
tiefen
[mm]
SF2 2,5 2,0 1,5 2,0 1,0 2,0 1,5 1,5 1,5 1,0 1,0
SF3 2,5 2,0 2,0 2,0 2,0 2,0 2,0 2,0 2,0 2,0 1,0
SF4 2,5 1,5 2,0 3,0 1,5 2,0 1,5 2,0 2,0 2,0 1,5
SF5 3,0 2,0 1,5 3,5 2,0 2,0 2,0 2,0 3,0 2,0 2,0
SF6 3,5 2,0 2,5 3,0 2,5 2,0 2,0 2,0 2,5 2,0 2,0
Tabelle 14: Messung der Taschentiefen (Gruppe C)
Die einzelnen mesialen und distalen Taschenmessungen lagen im Bereich von 1,00 – 4,00
mm und betrugen bei den Probanden der Gruppe C insgesamt im Mittel 1,90 mm.
Es fällt auf, dass sich die Werte innerhalb eines Probanden unterschieden. Die Zähne hatten
innerhalb eines Individuums nicht die gleiche Sulkustiefe. Dieser Unterschied kann individu-
ell unterschiedlich stark ausfallen. So hatte der Zahn für die Sulkusfluidprobe SF1 des Pro-
banden P6 eine durchschnittliche Taschentiefe von 1,50 mm, alle anderen Zähne des Proban-
den hatten eine Tiefe von durchschnittlich 2,00 mm. Bei dem Probanden P4 betrug der maxi-
male Unterschied der durchschnittlichen Taschentiefen hingegen bis zu 1,50 mm. Des Wei-
teren fällt ein interindividueller Unterschied in den Taschentiefen auf. So waren die Taschen
aller Zähne des Probanden P1 mit im Durchschnitt 2,75 mm tiefer als die Taschen der Zähne

78
des Probanden P11. Diese waren im Durchschnitt nur 1,50 mm tief. Innerhalb der Gruppe
konnten jedoch keine diese Unterschiede verursachenden Faktoren ausgemacht werden.
5.4.4. Bestimmung des API und SBI
Nach der Taschenmessung wurden abschließend mittels Erhebung des API und SBI der
Mundhygienestatus und der Entzündungszustand des gingivalen Bindegewebes der Proban-
den ermittelt (siehe Punkt 4.2.3.). Das Diagramm 14 gibt eine Übersicht über die dabei er-
mittelten Werte.
Diagramm 14: API und SBI (Gruppe C)
Der Proband P1 hatte mit einem API von 72 % und einem SBI von 82 % die für beide Indi-
zes jeweils maximal festgestellten Werte. Der API war mit 21 % am niedrigsten bei dem
Probanden P8, wohingegen der SBI mit 17 % am geringsten bei dem Probanden P10 ausfiel.
Der API betrug in der Gruppe C durchschnittlich 48 % und der SBI durchschnittlich 40 %.
Neben den teilweise deutlichen Unterschieden zwischen den jeweiligen Probanden sowohl
beim API als auch beim SBI, fällt vor allem auch eine unterschiedliche Korrelation der bei-
den Werte zueinander auf. So war der SBI bei dem Probanden P1 mit 82 % um 10 % größer
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11
API 72 36 67 63 42 61 46 21 52 33 35
SBI 82 32 42 42 30 46 46 33 44 17 26
0
10
20
30
40
50
60
70
80
90
100
AP
I/S
BI
[%]
API und SBI
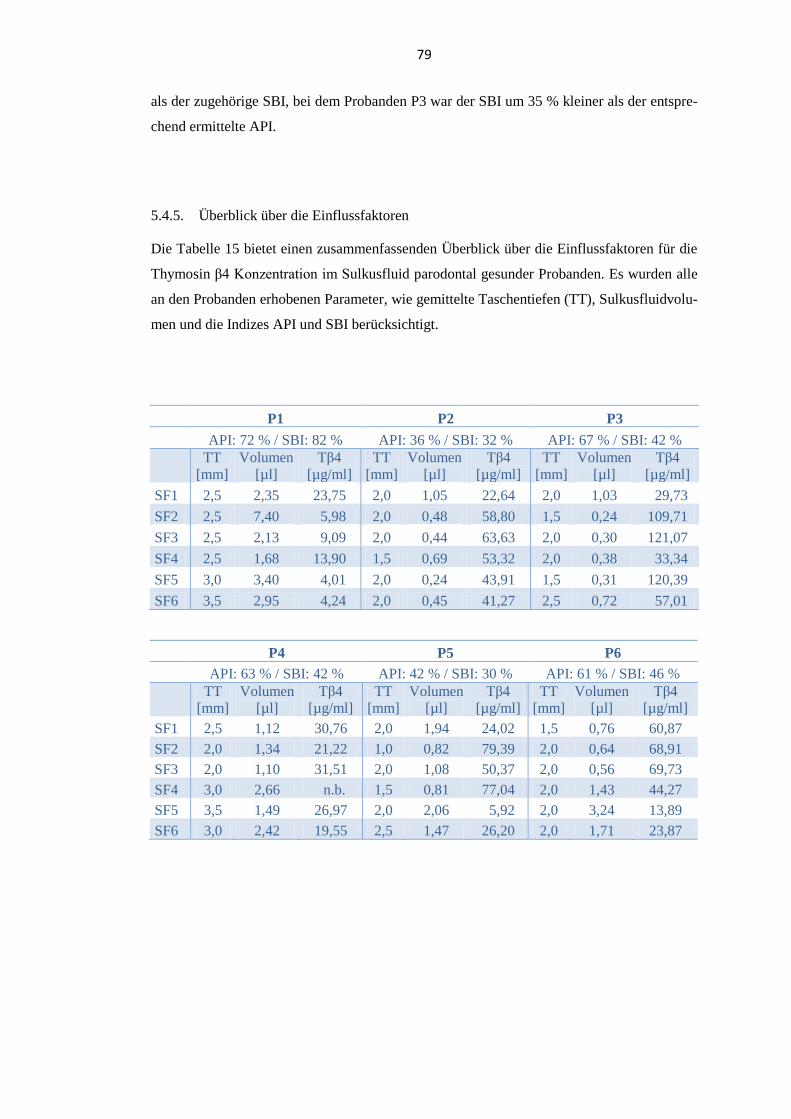
79
als der zugehörige SBI, bei dem Probanden P3 war der SBI um 35 % kleiner als der entspre-
chend ermittelte API.
5.4.5. Überblick über die Einflussfaktoren
Die Tabelle 15 bietet einen zusammenfassenden Überblick über die Einflussfaktoren für die
Thymosin β4 Konzentration im Sulkusfluid parodontal gesunder Probanden. Es wurden alle
an den Probanden erhobenen Parameter, wie gemittelte Taschentiefen (TT), Sulkusfluidvolu-
men und die Indizes API und SBI berücksichtigt.
P1 P2 P3
API: 72 % / SBI: 82 % API: 36 % / SBI: 32 % API: 67 % / SBI: 42 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 2,5 2,35 23,75 2,0 1,05 22,64 2,0 1,03 29,73
SF2 2,5 7,40 5,98 2,0 0,48 58,80 1,5 0,24 109,71
SF3 2,5 2,13 9,09 2,0 0,44 63,63 2,0 0,30 121,07
SF4 2,5 1,68 13,90 1,5 0,69 53,32 2,0 0,38 33,34
SF5 3,0 3,40 4,01 2,0 0,24 43,91 1,5 0,31 120,39
SF6 3,5 2,95 4,24 2,0 0,45 41,27 2,5 0,72 57,01
P4 P5 P6
API: 63 % / SBI: 42 % API: 42 % / SBI: 30 % API: 61 % / SBI: 46 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 2,5 1,12 30,76 2,0 1,94 24,02 1,5 0,76 60,87
SF2 2,0 1,34 21,22 1,0 0,82 79,39 2,0 0,64 68,91
SF3 2,0 1,10 31,51 2,0 1,08 50,37 2,0 0,56 69,73
SF4 3,0 2,66 n.b. 1,5 0,81 77,04 2,0 1,43 44,27
SF5 3,5 1,49 26,97 2,0 2,06 5,92 2,0 3,24 13,89
SF6 3,0 2,42 19,55 2,5 1,47 26,20 2,0 1,71 23,87
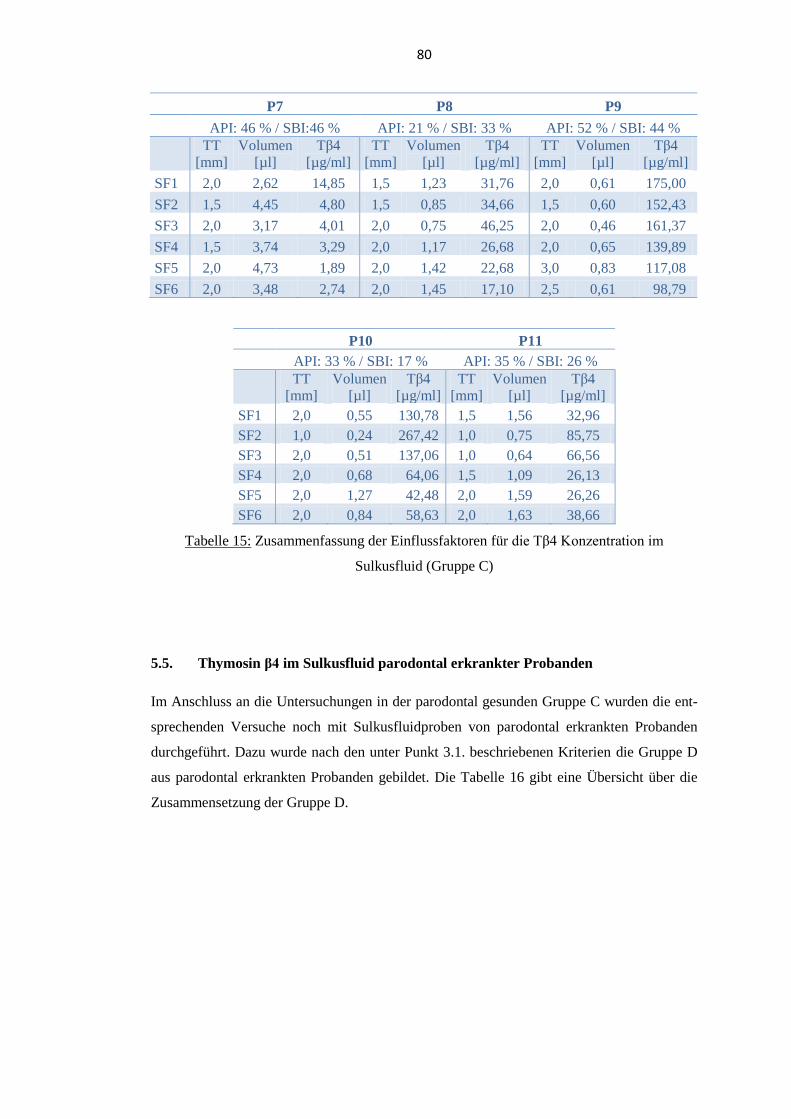
80
P7 P8 P9
API: 46 % / SBI:46 % API: 21 % / SBI: 33 % API: 52 % / SBI: 44 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 2,0 2,62 14,85 1,5 1,23 31,76 2,0 0,61 175,00
SF2 1,5 4,45 4,80 1,5 0,85 34,66 1,5 0,60 152,43
SF3 2,0 3,17 4,01 2,0 0,75 46,25 2,0 0,46 161,37
SF4 1,5 3,74 3,29 2,0 1,17 26,68 2,0 0,65 139,89
SF5 2,0 4,73 1,89 2,0 1,42 22,68 3,0 0,83 117,08
SF6 2,0 3,48 2,74 2,0 1,45 17,10 2,5 0,61 98,79
P10 P11
API: 33 % / SBI: 17 % API: 35 % / SBI: 26 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 2,0 0,55 130,78 1,5 1,56 32,96
SF2 1,0 0,24 267,42 1,0 0,75 85,75
SF3 2,0 0,51 137,06 1,0 0,64 66,56
SF4 2,0 0,68 64,06 1,5 1,09 26,13
SF5 2,0 1,27 42,48 2,0 1,59 26,26
SF6 2,0 0,84 58,63 2,0 1,63 38,66
Tabelle 15: Zusammenfassung der Einflussfaktoren für die Tβ4 Konzentration im
Sulkusfluid (Gruppe C)
5.5. Thymosin β4 im Sulkusfluid parodontal erkrankter Probanden
Im Anschluss an die Untersuchungen in der parodontal gesunden Gruppe C wurden die ent-
sprechenden Versuche noch mit Sulkusfluidproben von parodontal erkrankten Probanden
durchgeführt. Dazu wurde nach den unter Punkt 3.1. beschriebenen Kriterien die Gruppe D
aus parodontal erkrankten Probanden gebildet. Die Tabelle 16 gibt eine Übersicht über die
Zusammensetzung der Gruppe D.

81
Gruppe D
Proband Geschlecht Alter
1 M 58
2 W 71
3 W 49
4 M 68
5 M 48
6 M 53
7 W 57
8 W 57
Tabelle 16: Zusammensetzung Gruppe D
Die Gruppe D bildeten 8 Probanden zwischen 48 und 71 Jahren. Das Durchschnittsalter be-
trug 58 ± 9 Jahre. In der Gruppe befanden sich vier weibliche Probanden, der Rest war
männlich. Die Gruppe bestand aus Probanden, die zum Zeitpunkt der Untersuchung an
unterschiedlichen Formen von Parodontopathien erkrankt waren. Eine genaue Ätiologie der
jeweiligen Parodontopathie war nicht bekannt.
5.5.1. Sulkusfluidvolumen
Die Sulkusfluidproben wurden im Gegensatz zu der Gruppe C an unterschiedlichen Zähnen
der an Parodontopathien erkrankten Probanden gewonnen (siehe Punkt 4.2.1.5.). Anschlie-
ßend wurde das Volumen der Sulkusfluidproben bestimmt, bevor diese für die weitere RP-
HPLC-Analyse entsprechend vorbereitet wurden (siehe Punkt 4.2.4.2.). Das Diagramm 15
verdeutlicht die hierbei gefundenen Ergebnisse.
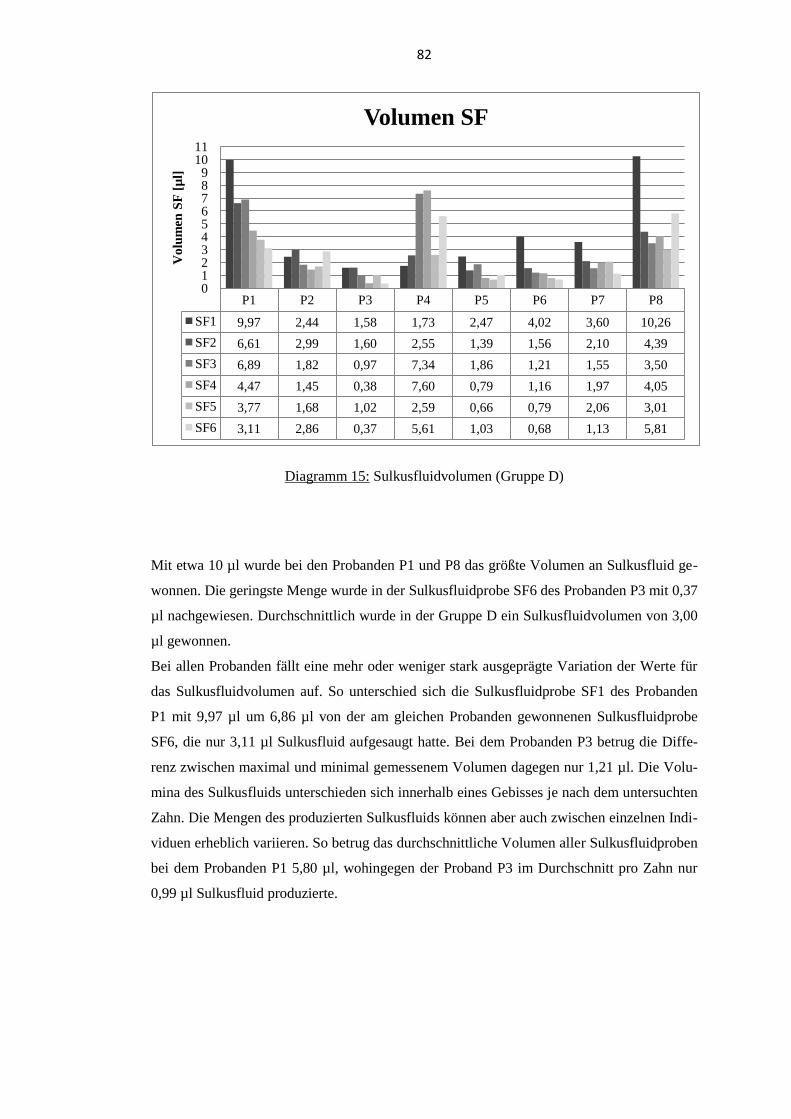
82
Diagramm 15: Sulkusfluidvolumen (Gruppe D)
Mit etwa 10 µl wurde bei den Probanden P1 und P8 das größte Volumen an Sulkusfluid ge-
wonnen. Die geringste Menge wurde in der Sulkusfluidprobe SF6 des Probanden P3 mit 0,37
µl nachgewiesen. Durchschnittlich wurde in der Gruppe D ein Sulkusfluidvolumen von 3,00
µl gewonnen.
Bei allen Probanden fällt eine mehr oder weniger stark ausgeprägte Variation der Werte für
das Sulkusfluidvolumen auf. So unterschied sich die Sulkusfluidprobe SF1 des Probanden
P1 mit 9,97 µl um 6,86 µl von der am gleichen Probanden gewonnenen Sulkusfluidprobe
SF6, die nur 3,11 µl Sulkusfluid aufgesaugt hatte. Bei dem Probanden P3 betrug die Diffe-
renz zwischen maximal und minimal gemessenem Volumen dagegen nur 1,21 µl. Die Volu-
mina des Sulkusfluids unterschieden sich innerhalb eines Gebisses je nach dem untersuchten
Zahn. Die Mengen des produzierten Sulkusfluids können aber auch zwischen einzelnen Indi-
viduen erheblich variieren. So betrug das durchschnittliche Volumen aller Sulkusfluidproben
bei dem Probanden P1 5,80 µl, wohingegen der Proband P3 im Durchschnitt pro Zahn nur
0,99 µl Sulkusfluid produzierte.
P1 P2 P3 P4 P5 P6 P7 P8
SF1 9,97 2,44 1,58 1,73 2,47 4,02 3,60 10,26
SF2 6,61 2,99 1,60 2,55 1,39 1,56 2,10 4,39
SF3 6,89 1,82 0,97 7,34 1,86 1,21 1,55 3,50
SF4 4,47 1,45 0,38 7,60 0,79 1,16 1,97 4,05
SF5 3,77 1,68 1,02 2,59 0,66 0,79 2,06 3,01
SF6 3,11 2,86 0,37 5,61 1,03 0,68 1,13 5,81
0123456789
1011
Vo
lum
en S
F [
µl]
Volumen SF

83
5.5.2. Thymosin β4 Konzentration im Sulkusfluid
Nach dem Auswiegen wurden die Sulkusfluidproben entsprechend vorbereitet (siehe Punkt
4.2.4.2.) und mittels RP-HPLC auf den Gehalt an Thymosin β4 analysiert (siehe Punkt
4.2.6.3.). Das Diagramm 16 verdeutlicht die hierbei gefundenen Ergebnisse. (0 = Thymosin
β4 Konzentration nicht bestimmt)
Diagramm 16: Tβ4 Konzentration im Sulkusfluid (Gruppe D)
Die Sulkusfluidprobe SF5 des Probanden P5 enthielt mit 82,52 µg/ml die größte Konzentra-
tion an Thymosin β4. Die niedrigste Konzentration wurde in der Sulkusfluidprobe SF1 des
Probanden P1 mit 2,16 µg/ml nachgewiesen. Durchschnittlich wurde bei allen Sulkusfluid-
proben eine Konzentration von 28,30 µg/ml gemessen.
Wie auch bei den parodontal gesunden Probanden der Gruppe C fällt eine mehr oder weniger
stark ausgeprägte Variation der Thymosin β4 Konzentration sowohl zwischen den einzelnen
Probanden aber auch innerhalb eines Probanden auf. Bei dem Probanden P5 wiesen zwei
Sulkusfluidproben (SF4, SF5) Thymosin β4 Konzentrationen von größer als 80 µg/ml auf,
drei Sulkusfluidproben (SF2, SF3, SF6) hatten Thymosin β4 Konzentrationen zwischen 30
und 53 µg/ml und eine Sulkusfluidprobe (SF1) nur eine Thymosin β4 Konzentration von
5,64 µg/ml. Diese intraindividuellen Unterschiede fielen am geringsten bei dem Probanden
P1 P2 P3 P4 P5 P6 P7 P8
SF1 2,16 31,80 20,06 33,69 5,64 6,20 53,45 17,45
SF2 0 15,41 13,98 24,42 30,05 21,27 13,45 28,93
SF3 6,06 19,96 4,68 0 52,22 29,24 74,81 44,77
SF4 12,78 21,94 7,89 0 82,29 26,29 29,86 15,30
SF5 11,83 21,86 8,20 22,46 82,52 64,08 34,76 22,70
SF6 21,00 9,41 53,49 6,29 41,76 70,18 43,75 12,21
0
50
100
150
200
250
300
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im SF

84
P1 aus. Hier betrug der Unterschied zwischen der größten und kleinsten nachgewiesenen
Konzentration an Thymosin β4 10,62 µg/ml. Es konnten auch deutliche interindividuelle Un-
terschiede festgestellt werden. So schien die Thymosin β4 Konzentration bei dem Probanden
P5 mit im Durchschnitt 49,08 µg/ml generell höher zu sein, als die in den Sulkusfluidproben
des Probanden P1. Diese betrug durchschnittlich 10,77 µg/ml.
5.5.3. Bestimmung der sulkulären Taschentiefen
Nach der Sulkusfluidprobennahme erfolgte bei den Probanden noch eine Messung der sulku-
lären Taschentiefen. Diese wurde als eine Vierpunktmessung (siehe Punkt 4.2.2.) durchge-
führt. Dazu wurden am zu untersuchenden Zahn jeweils buccal, oral, mesial und distal ent-
sprechende Werte für die Taschentiefe ermittelt und anschließend gemittelt. Die Tabelle 17
gibt einen Überblick über die entsprechenden Mittelwerte der Taschentiefen.
SF Probe P1 P2 P3 P4 P5 P6 P7 P8
SF1 3,50 2,75 3,75 3,00 6,25 5,50 3,75 5,50 Mittelwert
Taschen-
tiefen
[mm]
SF2 3,25 2,25 5,00 4,50 3,25 4,50 2,00 3,75
SF3 2,75 2,00 6,25 2,75 2,75 5,00 2,75 2,25
SF4 2,50 2,25 3,00 2,50 2,50 3,75 2,00 4,50
SF5 2,25 2,75 3,75 3,50 2,50 3,25 2,25 2,25
SF6 3,00 2,50 2,75 3,50 3,00 3,50 2,00 5,50
Tabelle 17: Messung der Taschentiefen (Gruppe D)
Die einzelnen Taschenmessungen lagen in einem Bereich zwischen 2,00 und 12,00 mm. Die
durchschnittliche Taschentiefe betrug bei allen Probanden 3,40 mm.
Es fällt auf, dass sich die gemittelten Werte intraindividuell zum Teil stark unterschieden. So
hatte der für die Sulkusfluidprobe SF1 untersuchte Zahn des Probanden P5 eine durch-
schnittliche Taschentiefe von 6,25 mm, wohingegen der am gleichen Patienten für die Sul-
kusfluidprobe SF4 untersuchte Zahn durchschnittlich nur eine Tiefe von 2,50 mm besaß.
Dieser Unterschied fiel bei dem Probanden P2 mit einer maximalen Differenz der durch-
schnittlichen Taschentiefen von 0,75 mm entsprechend geringer aus. Es gab des Weiteren
auch einen interindividuellen Unterschied bei den Taschentiefen der Probanden. Bei dem
Probanden P8 hatten alle untersuchten Zähne eine durchschnittliche Taschentiefe von 3,96
mm. Die Taschentiefen bei dem Probanden P2 waren hingegen generell weniger ausgeprägt
und hatten einen Durchschnittswert von 2,42 mm.

85
5.5.4. Bestimmung des API und SBI
Nach Messung der Taschentiefen wurde in der Gruppe D abschließend noch der Mundhy-
gienestatus anhand des API und SBI ermittelt (siehe Punkt 4.2.3.). Das Diagramm 17 gibt
eine Übersicht über die dabei ermittelten Werte.
Diagramm 17: API und SBI (Gruppe D)
Mit 92 % hatte der Proband P6 den höchsten und mit 35 % der Proband P7 den niedrigsten
API Wert. Für den SBI Wert wurde mit 67 % das Maximum bei dem Probanden P5 ermittelt.
Der kleinste Wert trat bei dem Probanden P2 mit 29 % auf. Der API betrug durchschnittlich
67 %, der SBI 47 %.
Die Probanden wiesen teilweise deutliche Unterschiede in den API und SBI Werten auf.
Aber auch die Beziehung der beiden Indizes zueinander zeigte deutliche Unterschiede. So
hatte der Proband P6 einen sehr hohen API Wert (92 %), aber einen eher kleinen SBI Wert
(31 %). Bei dem Probanden P5 waren beide Werte hingegen annähernd gleich groß (API: 60
%, SBI: 67 %).
P1 P2 P3 P4 P5 P6 P7 P8
API 63 72 60 85 60 92 35 91
SBI 46 29 30 59 67 31 51 60
0
10
20
30
40
50
60
70
80
90
100
AP
I/S
BI
[%]
API und SBI
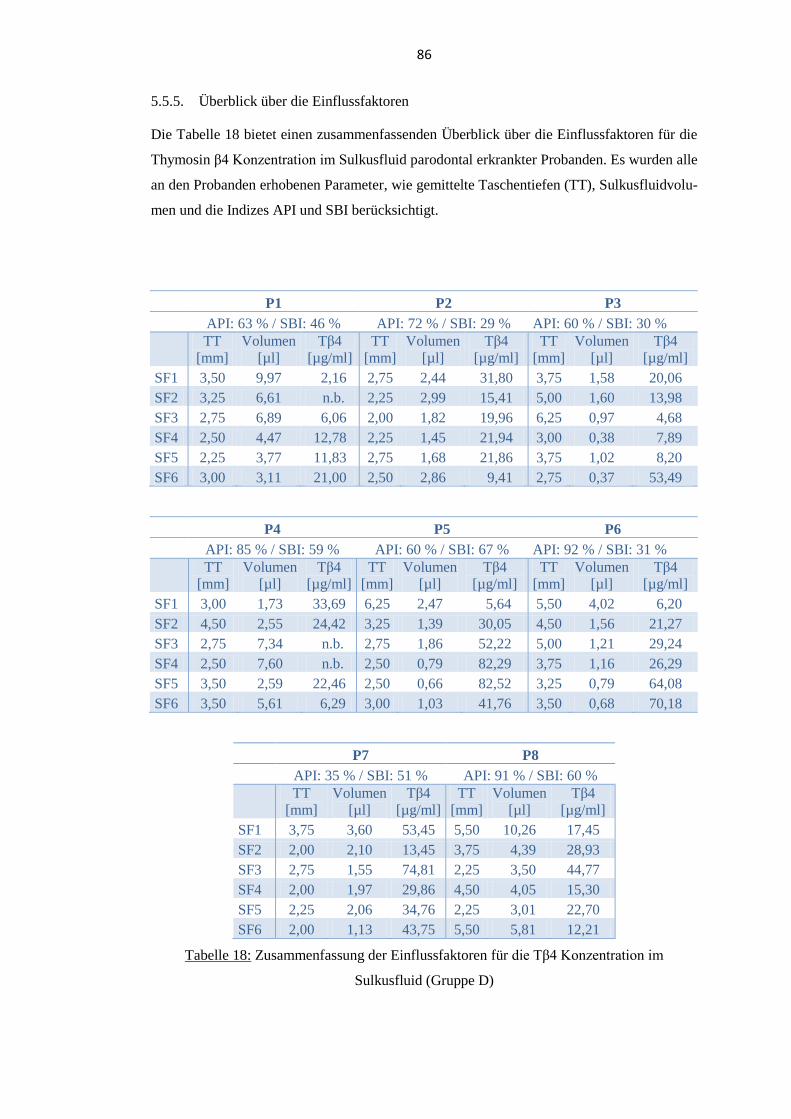
86
5.5.5. Überblick über die Einflussfaktoren
Die Tabelle 18 bietet einen zusammenfassenden Überblick über die Einflussfaktoren für die
Thymosin β4 Konzentration im Sulkusfluid parodontal erkrankter Probanden. Es wurden alle
an den Probanden erhobenen Parameter, wie gemittelte Taschentiefen (TT), Sulkusfluidvolu-
men und die Indizes API und SBI berücksichtigt.
P1 P2 P3
API: 63 % / SBI: 46 % API: 72 % / SBI: 29 % API: 60 % / SBI: 30 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 3,50 9,97 2,16 2,75 2,44 31,80 3,75 1,58 20,06
SF2 3,25 6,61 n.b. 2,25 2,99 15,41 5,00 1,60 13,98
SF3 2,75 6,89 6,06 2,00 1,82 19,96 6,25 0,97 4,68
SF4 2,50 4,47 12,78 2,25 1,45 21,94 3,00 0,38 7,89
SF5 2,25 3,77 11,83 2,75 1,68 21,86 3,75 1,02 8,20
SF6 3,00 3,11 21,00 2,50 2,86 9,41 2,75 0,37 53,49
P4 P5 P6
API: 85 % / SBI: 59 % API: 60 % / SBI: 67 % API: 92 % / SBI: 31 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 3,00 1,73 33,69 6,25 2,47 5,64 5,50 4,02 6,20
SF2 4,50 2,55 24,42 3,25 1,39 30,05 4,50 1,56 21,27
SF3 2,75 7,34 n.b. 2,75 1,86 52,22 5,00 1,21 29,24
SF4 2,50 7,60 n.b. 2,50 0,79 82,29 3,75 1,16 26,29
SF5 3,50 2,59 22,46 2,50 0,66 82,52 3,25 0,79 64,08
SF6 3,50 5,61 6,29 3,00 1,03 41,76 3,50 0,68 70,18
P7 P8
API: 35 % / SBI: 51 % API: 91 % / SBI: 60 %
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
TT
[mm]
Volumen
[µl]
Tβ4
[µg/ml]
SF1 3,75 3,60 53,45 5,50 10,26 17,45
SF2 2,00 2,10 13,45 3,75 4,39 28,93
SF3 2,75 1,55 74,81 2,25 3,50 44,77
SF4 2,00 1,97 29,86 4,50 4,05 15,30
SF5 2,25 2,06 34,76 2,25 3,01 22,70
SF6 2,00 1,13 43,75 5,50 5,81 12,21
Tabelle 18: Zusammenfassung der Einflussfaktoren für die Tβ4 Konzentration im
Sulkusfluid (Gruppe D)

87
5.6. Thymosin β4 im Sulkusfluid unter verstärkter okklusaler Belastung
Im Anschluss an die Untersuchungen der Konzentration von Thymosin β4 im Sulkusfluid
von Probanden mit unterschiedlichem Parodontalstatus, wurde der Einfluss einer verstärkten
okklusalen Belastung auf das Volumen des Sulkusfluids und die Thymosin β4 Konzentration
im Sulkusfluid untersucht. Dazu wurde nach den unter Punkt 3.1. beschriebenen Kriterien
die Gruppe E gebildet. Die Tabelle 19 gibt einen Überblick über die Zusammensetzung die-
ser Gruppe.
Gruppe E
Proband Geschlecht Alter
1 M 25
2 M 24
3 W 23
4 M 23
Tabelle 19: Zusammensetzung Gruppe E
Die Gruppe E bestand aus vier Probanden zwischen 23 und 25 Jahren. Das Durchschnitts-
alter betrug 24 ± 1 Jahre. Darunter war ein Proband weiblich, drei waren männlich. Alle Pro-
banden waren zum Zeitpunkt der Untersuchung parodontal gesund.
5.6.1. Sulkusfluidvolumen
Das Sulkusfluid wurde an den beiden Eckzähnen nach dem unter Punkt 4.2.1.6. beschrieben-
en Vorgehen gewonnen. Es wurden je Proband insgesamt fünf Proben genommen. „Vor
Aufbiss rechts“ und „vor Aufbiss links“ bezeichnen dabei die Proben, die vom rechten bzw.
linken oberen Eckzahn des Probanden vor einer okklusalen Belastung gewonnen wurden.
„Aufbiss rechts“ kennzeichnet die Sulkusfluidprobe, die während des Aufbisses auf einen
Silikonwall vom rechten oberen Eckzahn gewonnen wurde. Die Sulkusfluidproben „nach
Aufbiss rechts“ und „nach Aufbiss links“ wurden wie die ersten beiden Proben nur eben
nach erfolgter okklusaler Belastung gewonnen. Vor Analyse der Thymosin β4 Konzentration
mittels RP-HPLC wurde zunächst das Volumen des Sulkusfluids der einzelnen Proben be-
stimmt. Das Diagramm 18 gibt einen Überblick über die dabei gefundenen Ergebnisse.
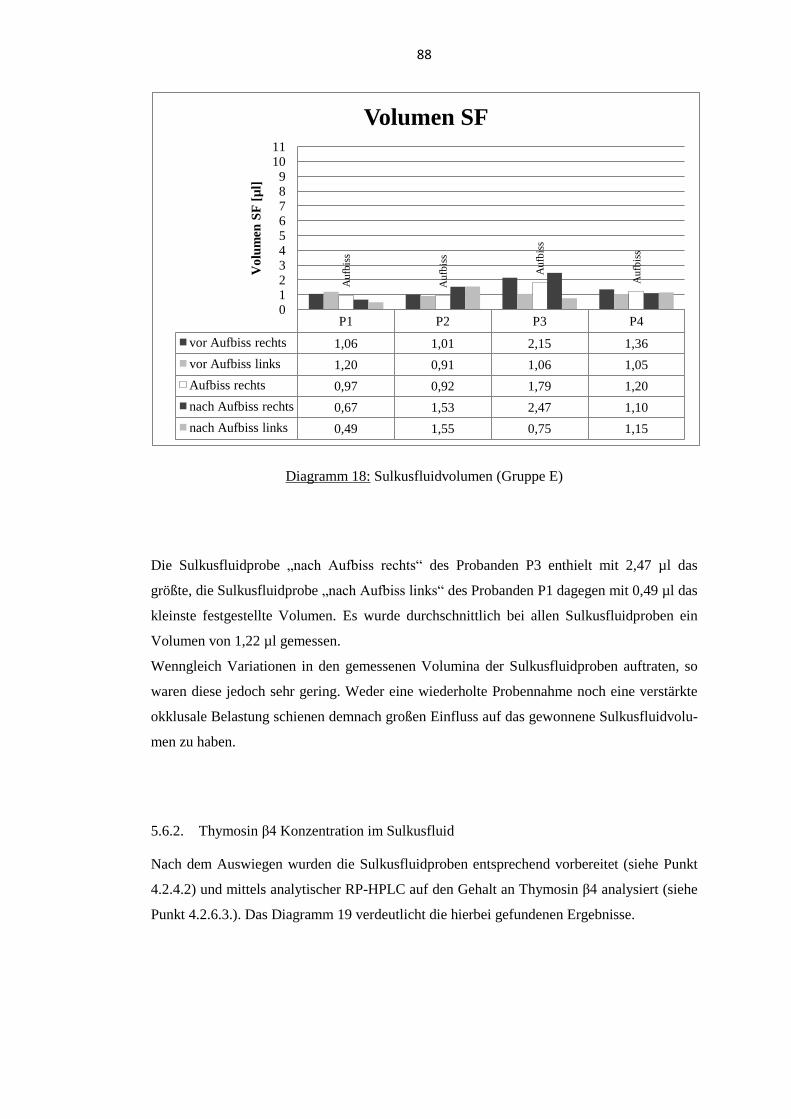
88
Diagramm 18: Sulkusfluidvolumen (Gruppe E)
Die Sulkusfluidprobe „nach Aufbiss rechts“ des Probanden P3 enthielt mit 2,47 µl das
größte, die Sulkusfluidprobe „nach Aufbiss links“ des Probanden P1 dagegen mit 0,49 µl das
kleinste festgestellte Volumen. Es wurde durchschnittlich bei allen Sulkusfluidproben ein
Volumen von 1,22 µl gemessen.
Wenngleich Variationen in den gemessenen Volumina der Sulkusfluidproben auftraten, so
waren diese jedoch sehr gering. Weder eine wiederholte Probennahme noch eine verstärkte
okklusale Belastung schienen demnach großen Einfluss auf das gewonnene Sulkusfluidvolu-
men zu haben.
5.6.2. Thymosin β4 Konzentration im Sulkusfluid
Nach dem Auswiegen wurden die Sulkusfluidproben entsprechend vorbereitet (siehe Punkt
4.2.4.2) und mittels analytischer RP-HPLC auf den Gehalt an Thymosin β4 analysiert (siehe
Punkt 4.2.6.3.). Das Diagramm 19 verdeutlicht die hierbei gefundenen Ergebnisse.
P1 P2 P3 P4
vor Aufbiss rechts 1,06 1,01 2,15 1,36
vor Aufbiss links 1,20 0,91 1,06 1,05
Aufbiss rechts 0,97 0,92 1,79 1,20
nach Aufbiss rechts 0,67 1,53 2,47 1,10
nach Aufbiss links 0,49 1,55 0,75 1,15
Au
fbis
s
Au
fbis
s
Au
fbis
s
Au
fbis
s
0123456789
1011
Vo
lum
en S
F [
µl]
Volumen SF
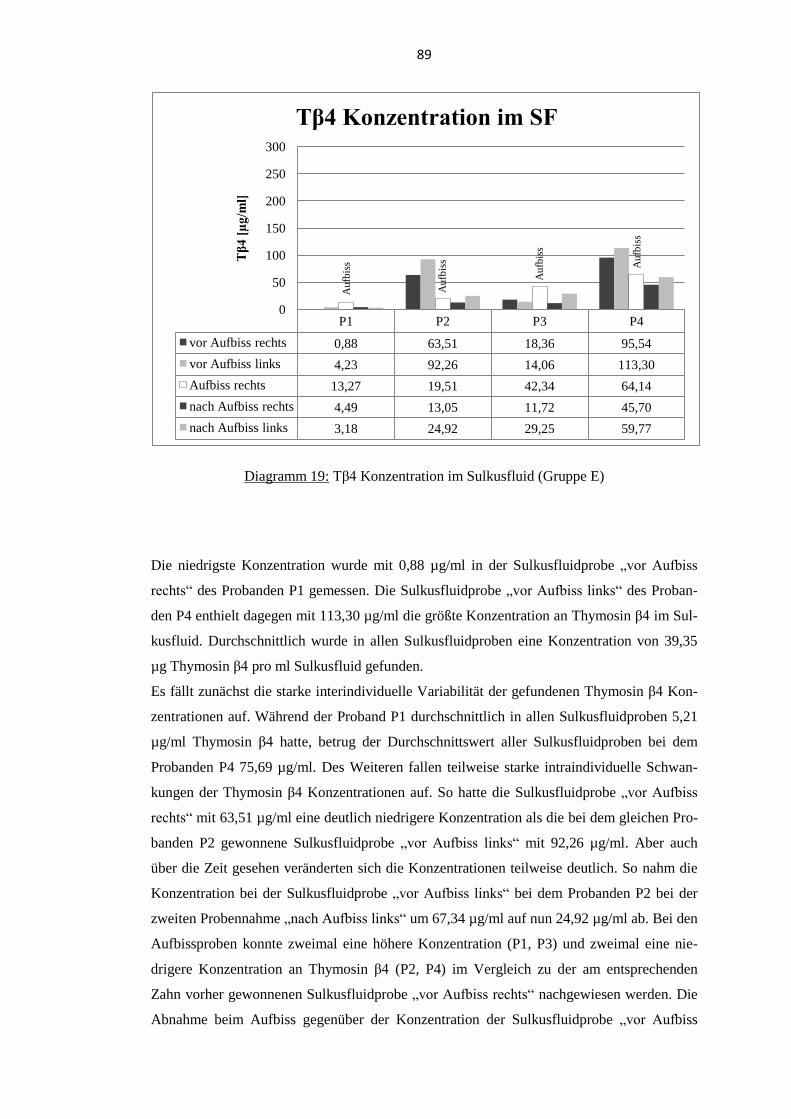
89
Diagramm 19: Tβ4 Konzentration im Sulkusfluid (Gruppe E)
Die niedrigste Konzentration wurde mit 0,88 µg/ml in der Sulkusfluidprobe „vor Aufbiss
rechts“ des Probanden P1 gemessen. Die Sulkusfluidprobe „vor Aufbiss links“ des Proban-
den P4 enthielt dagegen mit 113,30 µg/ml die größte Konzentration an Thymosin β4 im Sul-
kusfluid. Durchschnittlich wurde in allen Sulkusfluidproben eine Konzentration von 39,35
µg Thymosin β4 pro ml Sulkusfluid gefunden.
Es fällt zunächst die starke interindividuelle Variabilität der gefundenen Thymosin β4 Kon-
zentrationen auf. Während der Proband P1 durchschnittlich in allen Sulkusfluidproben 5,21
µg/ml Thymosin β4 hatte, betrug der Durchschnittswert aller Sulkusfluidproben bei dem
Probanden P4 75,69 µg/ml. Des Weiteren fallen teilweise starke intraindividuelle Schwan-
kungen der Thymosin β4 Konzentrationen auf. So hatte die Sulkusfluidprobe „vor Aufbiss
rechts“ mit 63,51 µg/ml eine deutlich niedrigere Konzentration als die bei dem gleichen Pro-
banden P2 gewonnene Sulkusfluidprobe „vor Aufbiss links“ mit 92,26 µg/ml. Aber auch
über die Zeit gesehen veränderten sich die Konzentrationen teilweise deutlich. So nahm die
Konzentration bei der Sulkusfluidprobe „vor Aufbiss links“ bei dem Probanden P2 bei der
zweiten Probennahme „nach Aufbiss links“ um 67,34 µg/ml auf nun 24,92 µg/ml ab. Bei den
Aufbissproben konnte zweimal eine höhere Konzentration (P1, P3) und zweimal eine nie-
drigere Konzentration an Thymosin β4 (P2, P4) im Vergleich zu der am entsprechenden
Zahn vorher gewonnenen Sulkusfluidprobe „vor Aufbiss rechts“ nachgewiesen werden. Die
Abnahme beim Aufbiss gegenüber der Konzentration der Sulkusfluidprobe „vor Aufbiss
P1 P2 P3 P4
vor Aufbiss rechts 0,88 63,51 18,36 95,54
vor Aufbiss links 4,23 92,26 14,06 113,30
Aufbiss rechts 13,27 19,51 42,34 64,14
nach Aufbiss rechts 4,49 13,05 11,72 45,70
nach Aufbiss links 3,18 24,92 29,25 59,77
Au
fbis
s
Au
fbis
s
Au
fbis
s
Au
fbis
s
0
50
100
150
200
250
300
Tβ
4 [
µg
/ml]
Tβ4 Konzentration im SF

90
rechts“ war dabei bei dem Probanden P2 mit 44,00 µg/ml am größten. Die größte Zunahme
im Vergleich zur Sulkusfluidprobe „vor Aufbiss rechts“ fand sich bei dem Probanden P3 mit
23,98 µg/ml. Eine verstärkte okklusale Belastung des Zahnes führte folglich nicht immer zu
einer entsprechend verstärkt freigesetzten Thymosin β4 Menge im Sulkusfluid. Betrachtet
man die Sulkusfluidproben, die am entsprechend belasteten Zahn nach dessen Belastung als
Sulkusfluid „nach Aufbiss rechts“ gewonnen wurden, so lassen sich ebenfalls Auffälligkei-
ten erkennen. So war bei allen Probanden, die in den Sulkusfluidproben „nach Aufbiss
rechts“ gefundene Konzentration an Thymosin β4 im Vergleich zu der in den Aufbissproben
stets verringert. Der größte Unterschied trat bei dem Probanden P3 mit einer Abnahme um
30,62 µg/ml, der kleinste bei dem Probanden P1 mit einer Abnahme um 8,78 µg/ml auf.
5.7. Elutionsversuche mit Papierspitzen
Für die Elutionsversuche von in Papierspitzen aufgesaugtem Thymosin β4 wurden diese für
eine Minute in ein Eppendorfcup mit einem Thymosin β4 Standard bekannter Konzentration
gestellt. Das während dieser Zeit aufgesaugte Volumen wurde durch Auswiegen bestimmt
und die daraus zu erwartende Thymosin β4 Menge berechnet. Anhand der anschließend aus
dem Eluat mittels RP-HPLC bestimmten tatsächlichen Thymosin β4 Menge wurde dann für
die jeweilige Methode ein entsprechender Wiederfindungswert (Wwert) bestimmt. Es wur-
den insgesamt elf verschiedene Methoden zur Elution entwickelt. Die Tabellen 20 und 21
zeigen die Prinzipien dieser Methoden und die dabei jeweils wiedergefundenen Konzentrati-
onen an Thymosin β4 in Prozent.
Methode Prinzip Wwert
1 1 100 µl TFA, 1 min rütteln 9,2 %
2 2 200 µl TFA, je 1 min rütteln 26,0 %
3 4 100 µl TFA, je 2 min rütteln 35,0 %
4
6 100 µl TFA, je 2 h rütteln, Proben nach:
2 h
4 h
6 – 12 h
11,2 %
7,3 %
0 – 5 %
5 1 100 µl TFA + TD, 1 min rütteln 22,2 %
6 1 100 µl GuaHCL, 3 100 µl TFA, je 1 min
rütteln, je 5 min Ultraschall Oxidiert
7 1 100 µl GuaHCL, 1 min rütteln 43,9 %
8 1 100 µl GuaHCL, 6 h rütteln 73,0 %
9 1 100 µl GuaHCL + TD, 1 min rütteln 72,0 %
Tabelle 20: Methoden zur Elution von Tβ4 aus Papierspitzen Teil 1

91
Mit der unter Methode 9 beschriebenen Kombination aus 10 ml 6 M GuaHCL und 50 µl TD
schien im Vergleich zu einer alleinigen Elution der Papierspitzen mit 0,1 % TFA ein guter
und effizienter Ansatz gemacht worden zu sein. Durch weitere Versuche mit dieser Waschlö-
sung konnte die Ausbeute an Thymosin β4 aus Papierspitzen weiter gesteigert werden.
Methode Prinzip Wwert
10 2 100 µl GuaHCL + TD, je 1 h rütteln, Proben nach:
1 h
2 h
75,7 %
19,8 %
11 3 100 µl GuaHCL + TD, je 2 h rütteln 93,2 %
Tabelle 21: Methoden zur Elution von Tβ4 aus Papierspitzen Teil 2
Es stellte sich heraus, dass die Anwendung der Waschlösung aus GuaHCL und TD am effek-
tivsten nach der Methode 11 funktioniert. Mit dem unter Methode 11 beschriebenen Prinzip
konnten bis zu 93,2 % der aufgesaugten Menge an Thymosin β4 aus einer Papierspitze elu-
iert werden. Diese Werte wurden durch eine entsprechende Wiederholung der Elutionsme-
thode bestätigt (Daten nicht gezeigt). Die Methode 11 kam demnach zur Elution von Sulkus-
fluid aus Papierspitzen nach einer entsprechenden Standardisierung zum Einsatz.
5.8. Massenspektrometrie
Nachdem mit denen unter Punkt 4.2.6.2. beschriebenen Methoden eine eindeutige Identifi-
zierung und Quantifizierung von Thymosin β4 durch die analytische RP-HPLC-Analyse
sichergestellt worden war, sollte exemplarisch die molare Masse des Thymosin β4 in einer
Gesamtspeichelprobe analysiert werden. Als Methode kam das MALDI-TOF-Verfahren zur
Anwendung (siehe Punkt 4.2.8.).
Die Gesamtspeichelprobe eines 53-jährigen männlichen Probanden wurde zunächst mit dem
RP-18-Säulen-Verfahren aufgereinigt (siehe Punkt 4.2.7.). Nachdem mittels analytischer RP-
HPLC nachgewiesen wurde, dass sich das entsprechend aufgereinigte Thymosin β4 in der
Elutionsfraktion 4 mit einer Konzentration von 5,75 µg/ml befand, wurde diese mittels prä-
parativem RP-HPLC-Verfahren fraktioniert (siehe Punkt 4.2.6.4.). Dabei enthielt Fraktion 24
die gesamte Thymosin β4 Menge, die nun eine Konzentration von 1,91 µg/ml besaß. Parallel
dazu wurde eine entsprechend aufbereitete Schweineleukozytenprobe nach analogem Vorge-
hen fraktioniert. Hier enthielt ebenfalls die Fraktion 24 die gesamte Thymosin β4 Menge.
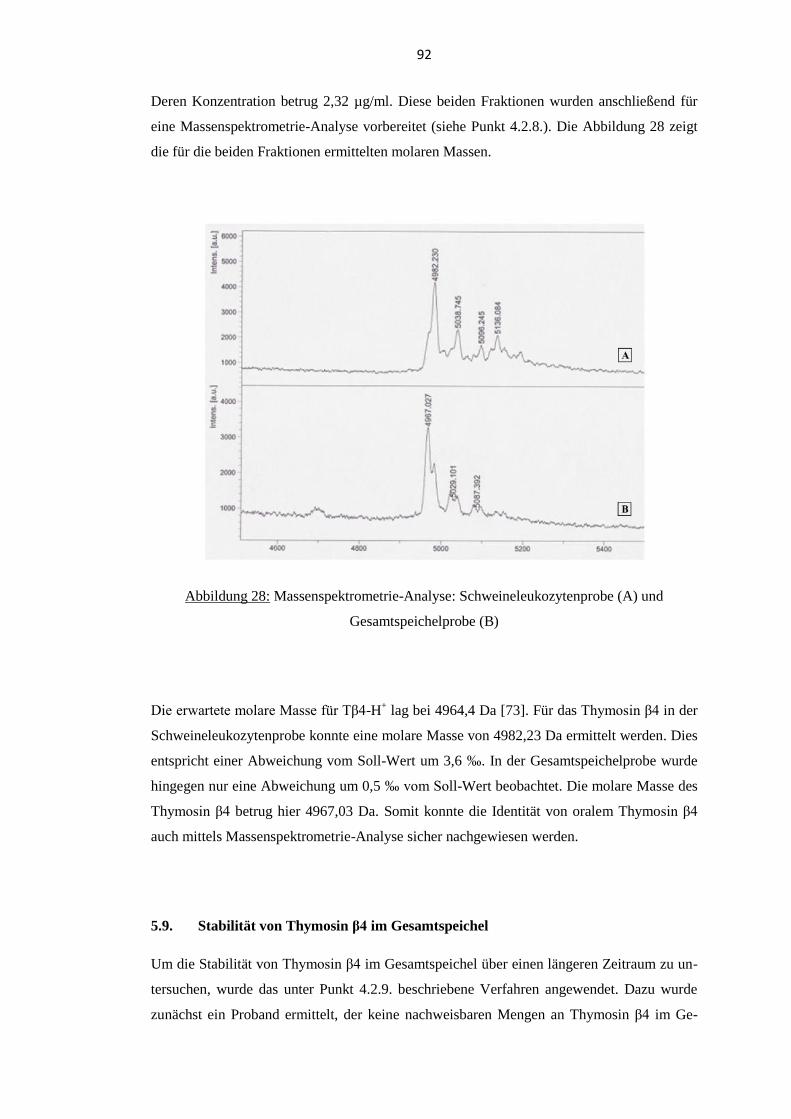
92
Deren Konzentration betrug 2,32 µg/ml. Diese beiden Fraktionen wurden anschließend für
eine Massenspektrometrie-Analyse vorbereitet (siehe Punkt 4.2.8.). Die Abbildung 28 zeigt
die für die beiden Fraktionen ermittelten molaren Massen.
Abbildung 28: Massenspektrometrie-Analyse: Schweineleukozytenprobe (A) und
Gesamtspeichelprobe (B)
Die erwartete molare Masse für Tβ4-H+ lag bei 4964,4 Da [73]. Für das Thymosin β4 in der
Schweineleukozytenprobe konnte eine molare Masse von 4982,23 Da ermittelt werden. Dies
entspricht einer Abweichung vom Soll-Wert um 3,6 ‰. In der Gesamtspeichelprobe wurde
hingegen nur eine Abweichung um 0,5 ‰ vom Soll-Wert beobachtet. Die molare Masse des
Thymosin β4 betrug hier 4967,03 Da. Somit konnte die Identität von oralem Thymosin β4
auch mittels Massenspektrometrie-Analyse sicher nachgewiesen werden.
5.9. Stabilität von Thymosin β4 im Gesamtspeichel
Um die Stabilität von Thymosin β4 im Gesamtspeichel über einen längeren Zeitraum zu un-
tersuchen, wurde das unter Punkt 4.2.9. beschriebene Verfahren angewendet. Dazu wurde
zunächst ein Proband ermittelt, der keine nachweisbaren Mengen an Thymosin β4 im Ge-
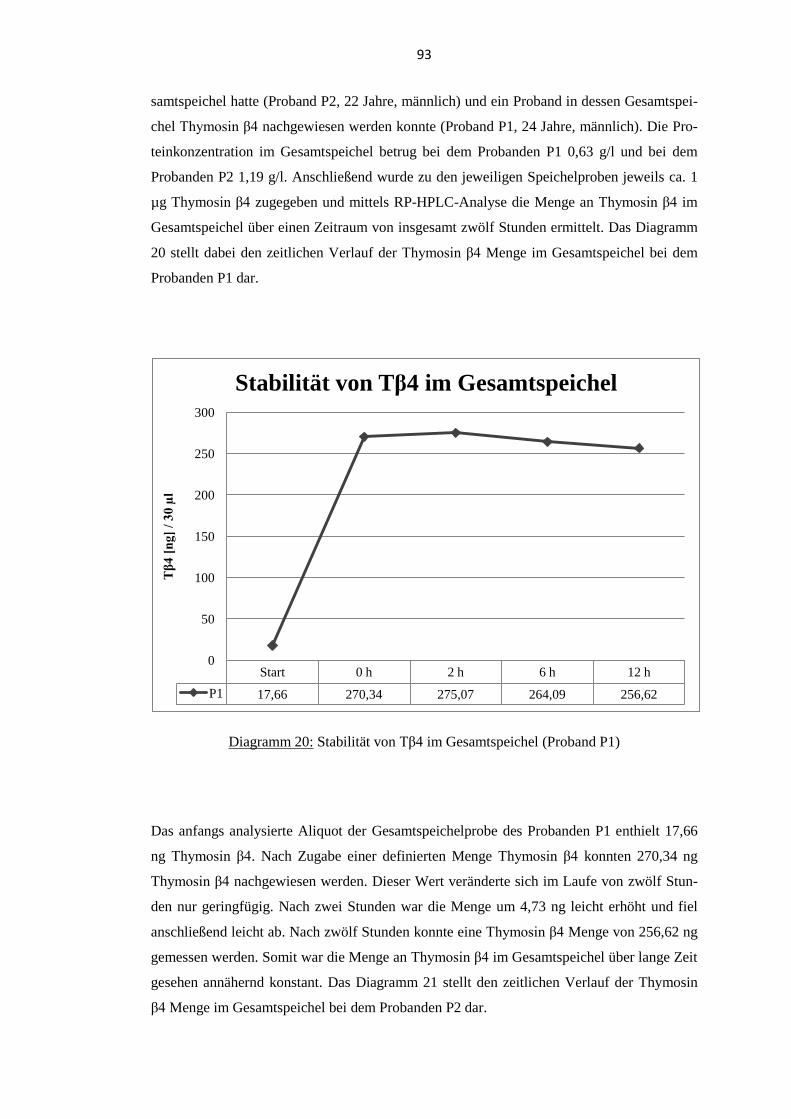
93
samtspeichel hatte (Proband P2, 22 Jahre, männlich) und ein Proband in dessen Gesamtspei-
chel Thymosin β4 nachgewiesen werden konnte (Proband P1, 24 Jahre, männlich). Die Pro-
teinkonzentration im Gesamtspeichel betrug bei dem Probanden P1 0,63 g/l und bei dem
Probanden P2 1,19 g/l. Anschließend wurde zu den jeweiligen Speichelproben jeweils ca. 1
µg Thymosin β4 zugegeben und mittels RP-HPLC-Analyse die Menge an Thymosin β4 im
Gesamtspeichel über einen Zeitraum von insgesamt zwölf Stunden ermittelt. Das Diagramm
20 stellt dabei den zeitlichen Verlauf der Thymosin β4 Menge im Gesamtspeichel bei dem
Probanden P1 dar.
Diagramm 20: Stabilität von Tβ4 im Gesamtspeichel (Proband P1)
Das anfangs analysierte Aliquot der Gesamtspeichelprobe des Probanden P1 enthielt 17,66
ng Thymosin β4. Nach Zugabe einer definierten Menge Thymosin β4 konnten 270,34 ng
Thymosin β4 nachgewiesen werden. Dieser Wert veränderte sich im Laufe von zwölf Stun-
den nur geringfügig. Nach zwei Stunden war die Menge um 4,73 ng leicht erhöht und fiel
anschließend leicht ab. Nach zwölf Stunden konnte eine Thymosin β4 Menge von 256,62 ng
gemessen werden. Somit war die Menge an Thymosin β4 im Gesamtspeichel über lange Zeit
gesehen annähernd konstant. Das Diagramm 21 stellt den zeitlichen Verlauf der Thymosin
β4 Menge im Gesamtspeichel bei dem Probanden P2 dar.
Start 0 h 2 h 6 h 12 h
P1 17,66 270,34 275,07 264,09 256,62
0
50
100
150
200
250
300
Tβ
4 [
ng
] /
30
µl
Stabilität von Tβ4 im Gesamtspeichel
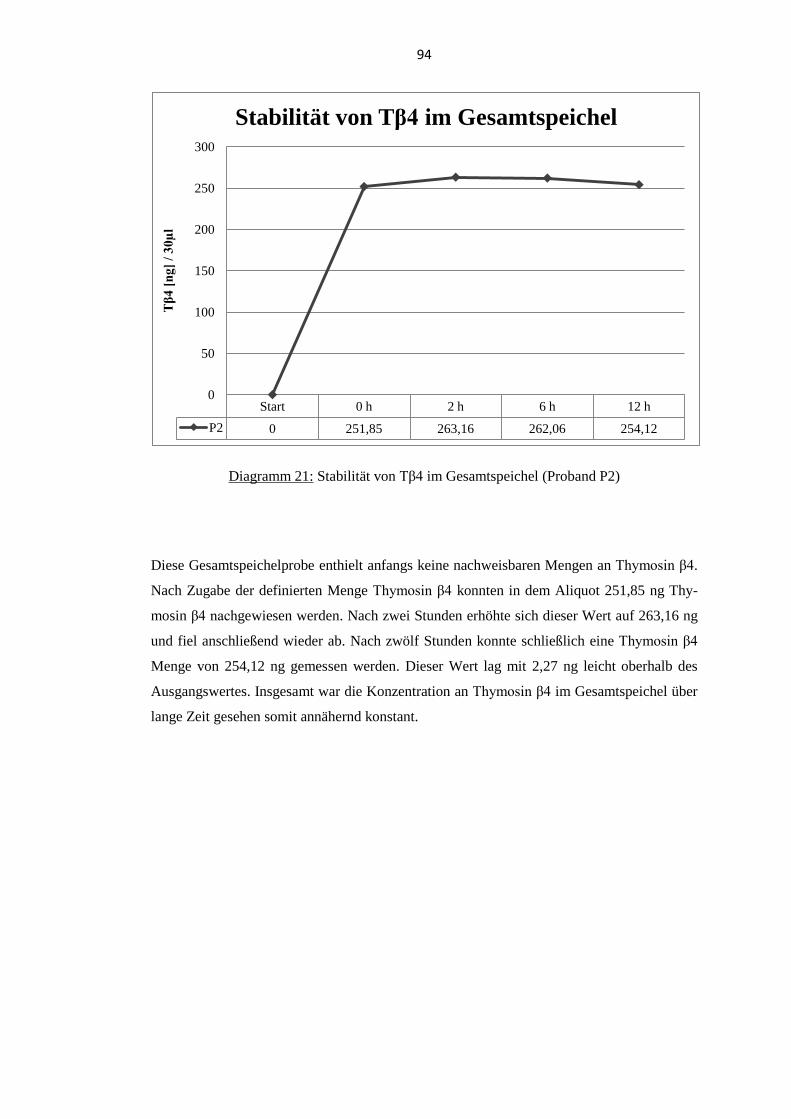
94
Diagramm 21: Stabilität von Tβ4 im Gesamtspeichel (Proband P2)
Diese Gesamtspeichelprobe enthielt anfangs keine nachweisbaren Mengen an Thymosin β4.
Nach Zugabe der definierten Menge Thymosin β4 konnten in dem Aliquot 251,85 ng Thy-
mosin β4 nachgewiesen werden. Nach zwei Stunden erhöhte sich dieser Wert auf 263,16 ng
und fiel anschließend wieder ab. Nach zwölf Stunden konnte schließlich eine Thymosin β4
Menge von 254,12 ng gemessen werden. Dieser Wert lag mit 2,27 ng leicht oberhalb des
Ausgangswertes. Insgesamt war die Konzentration an Thymosin β4 im Gesamtspeichel über
lange Zeit gesehen somit annähernd konstant.
Start 0 h 2 h 6 h 12 h
P2 0 251,85 263,16 262,06 254,12
0
50
100
150
200
250
300
Tβ
4 [
ng
] /
30
µl
Stabilität von Tβ4 im Gesamtspeichel

95
6. Diskussion
6.1. Thymosin β4 Konzentration im Gesamtspeichel – Vergleich Gruppe A und B
Innerhalb der jeweiligen Gruppen A und B konnte keine Regelmäßigkeit für die jeweils auf-
tretenden Konzentrationen an Thymosin β4 im Gesamtspeichel erkannt werden. Die Werte
schwankten interindividuell in beiden Gruppen teilweise sehr stark (siehe Punkt 5.1.1. Dia-
gramm 3 und Punkt 5.1.2. Diagramm 5). Vergleicht man jedoch die beiden Gruppen mitein-
ander, so lassen sich einige Feststellungen treffen. Die Tabelle 22 gibt einen Überblick über
die gefundenen Konzentrationen an Thymosin β4 im Gesamtspeichel in den Gruppen A
(Durchschnittsalter 25 ± 3 Jahre) und B (Durchschnittsalter 54 ± 5 Jahre).
Tβ4 Konzentration [µg/ml]
Gruppe Mittelwert Maximum Minimum
A Tag 0
Tag 14
0,37
0,34
1,71
0,73
≤ 0,05
≤ 0,05
B Tag 0
Tag 14
0,78
0,85
1,35
2,49
0,19
0,15
Tabelle 22: Tβ4 Konzentration im Gesamtspeichel – Vergleich Gruppe A (n = 7)
und B (n = 8)
Bei dieser Betrachtung fällt zunächst auf, dass die für Tag 0 und Tag 14 gefundenen Mittel-
werte der Thymosin β4 Konzentration für die jeweiligen Gruppen in einem ähnlichen Be-
reich liegen. In der Gruppe A liegt der Mittelwert der ersten Probennahme bei 0,37 µg/ml.
Der Mittelwert bei der zweiten Probennahme beträgt 0,34 µg/ml. In Gruppe B sind die Mit-
telwerte dagegen mit 0,78 µg/ml bei der ersten und 0,85 µg/ml bei der zweiten Probennahme
größer. Zudem haben ältere Probanden im Gegensatz zu jüngeren Probanden stets gut mess-
bare Konzentrationen an Thymosin β4 im Gesamtspeichel. In Gruppe A liegen die Konzen-
trationen für Thymosin β4 hingegen bei der Probennahme am Tag 0 bei zwei und bei der
Probennahme am Tag 14 bei einem Probanden unterhalb der Nachweisgrenze von 0,05
µg/ml. Ältere Probanden scheinen im Mittel also höhere Konzentrationen an Thymosin β4
im Gesamtspeichel zu haben.
Als Gemeinsamkeit kann sowohl in der Gruppe A als auch in der Gruppe B festgestellt
werden, dass die Konzentrationen zeitlich gesehen variabel sind (siehe Punkt 5.1.1. Dia-
gramm 4 und Punkt 5.1.2. Diagramm 6). Verglichen mit der ersten Probennahme treten bei
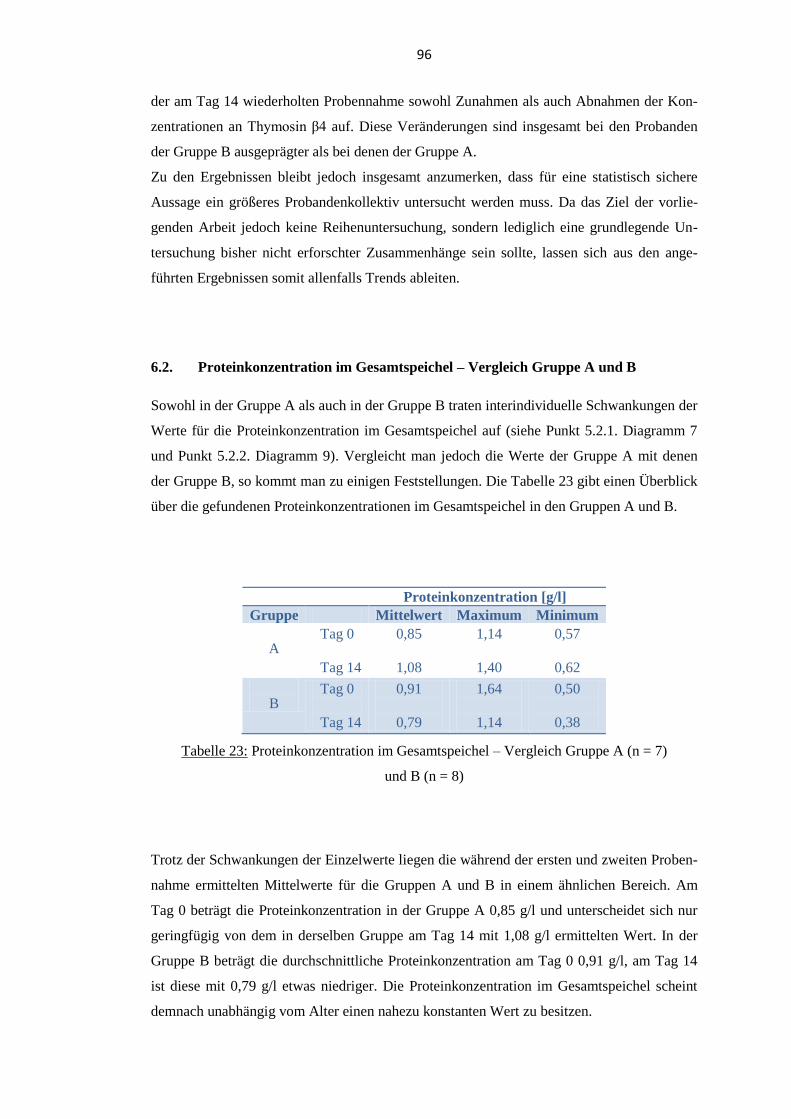
96
der am Tag 14 wiederholten Probennahme sowohl Zunahmen als auch Abnahmen der Kon-
zentrationen an Thymosin β4 auf. Diese Veränderungen sind insgesamt bei den Probanden
der Gruppe B ausgeprägter als bei denen der Gruppe A.
Zu den Ergebnissen bleibt jedoch insgesamt anzumerken, dass für eine statistisch sichere
Aussage ein größeres Probandenkollektiv untersucht werden muss. Da das Ziel der vorlie-
genden Arbeit jedoch keine Reihenuntersuchung, sondern lediglich eine grundlegende Un-
tersuchung bisher nicht erforschter Zusammenhänge sein sollte, lassen sich aus den ange-
führten Ergebnissen somit allenfalls Trends ableiten.
6.2. Proteinkonzentration im Gesamtspeichel – Vergleich Gruppe A und B
Sowohl in der Gruppe A als auch in der Gruppe B traten interindividuelle Schwankungen der
Werte für die Proteinkonzentration im Gesamtspeichel auf (siehe Punkt 5.2.1. Diagramm 7
und Punkt 5.2.2. Diagramm 9). Vergleicht man jedoch die Werte der Gruppe A mit denen
der Gruppe B, so kommt man zu einigen Feststellungen. Die Tabelle 23 gibt einen Überblick
über die gefundenen Proteinkonzentrationen im Gesamtspeichel in den Gruppen A und B.
Proteinkonzentration [g/l]
Gruppe Mittelwert Maximum Minimum
A Tag 0
Tag 14
0,85
1,08
1,14
1,40
0,57
0,62
B Tag 0
Tag 14
0,91
0,79
1,64
1,14
0,50
0,38
Tabelle 23: Proteinkonzentration im Gesamtspeichel – Vergleich Gruppe A (n = 7)
und B (n = 8)
Trotz der Schwankungen der Einzelwerte liegen die während der ersten und zweiten Proben-
nahme ermittelten Mittelwerte für die Gruppen A und B in einem ähnlichen Bereich. Am
Tag 0 beträgt die Proteinkonzentration in der Gruppe A 0,85 g/l und unterscheidet sich nur
geringfügig von dem in derselben Gruppe am Tag 14 mit 1,08 g/l ermittelten Wert. In der
Gruppe B beträgt die durchschnittliche Proteinkonzentration am Tag 0 0,91 g/l, am Tag 14
ist diese mit 0,79 g/l etwas niedriger. Die Proteinkonzentration im Gesamtspeichel scheint
demnach unabhängig vom Alter einen nahezu konstanten Wert zu besitzen.

97
Im Gegensatz zu den im Gesamtspeichel bestimmten Thymosin β4 Konzentrationen sind die
Proteinkonzentrationen im Gesamtspeichel sowohl in der Gruppe A als auch in der Gruppe B
zeitlich gesehen annähernd konstant (siehe Punkt 5.2.1. Diagramm 8 und Punkt 5.2.2. Dia-
gramm 10). Verglichen mit der ersten Probennahme treten zwar auch hier bei der am Tag 14
wiederholten Probennahme sowohl Zunahmen als auch Abnahmen der ermittelten Konzen-
trationen auf, das Ausmaß dieser Veränderungen ist jedoch wesentlich geringer als bei den
im Gesamtspeichel ermittelten Thymosin β4 Konzentrationen.
6.3. Korrelation Thymosin β4 Konzentration und Proteinkonzentration im Gesamt-
speichel (Gruppe A und B)
Vergleicht man die Thymosin β4 Konzentrationen im Gesamtspeichel der Probanden mit den
entsprechenden Proteinkonzentrationen, so fällt zunächst auf, dass die Proteinkonzentratio-
nen im Gegensatz zu den Thymosin β4 Konzentrationen scheinbar nicht vom Alter abhängig
sind. Während sich die Proteinkonzentrationen im Gesamtspeichel zwischen älteren und jün-
geren Probanden kaum unterscheiden (siehe Punkt 6.2. Tabelle 23), haben ältere Probanden
durchschnittlich höhere Thymosin β4 Konzentrationen im Gesamtspeichel als jüngere Pro-
banden (siehe Punkt 6.1. Tabelle 22).
Setzt man die Thymosin β4 Konzentrationen in Relation zu den jeweiligen Proteinkonzentra-
tionen im Gesamtspeichel, so lässt sich keine direkte Korrelation erkennen. Die Thymosin
β4 Konzentration scheint sowohl in der Gruppe A als auch in der Gruppe B unabhängig von
der jeweiligen Proteinkonzentration im Gesamtspeichel zu sein (siehe Diagramm 22 und 23).
In den Diagrammen 22 und 23 wurden jeweils alle Wertepaare der Ergebnisse aus den bei-
den Probennahmen in den Gruppen A und B berücksichtigt, es sei denn die Werte für die
Thymosin β4 Konzentration lagen unterhalb der Nachweisgrenze von 0,05 µg/ml.
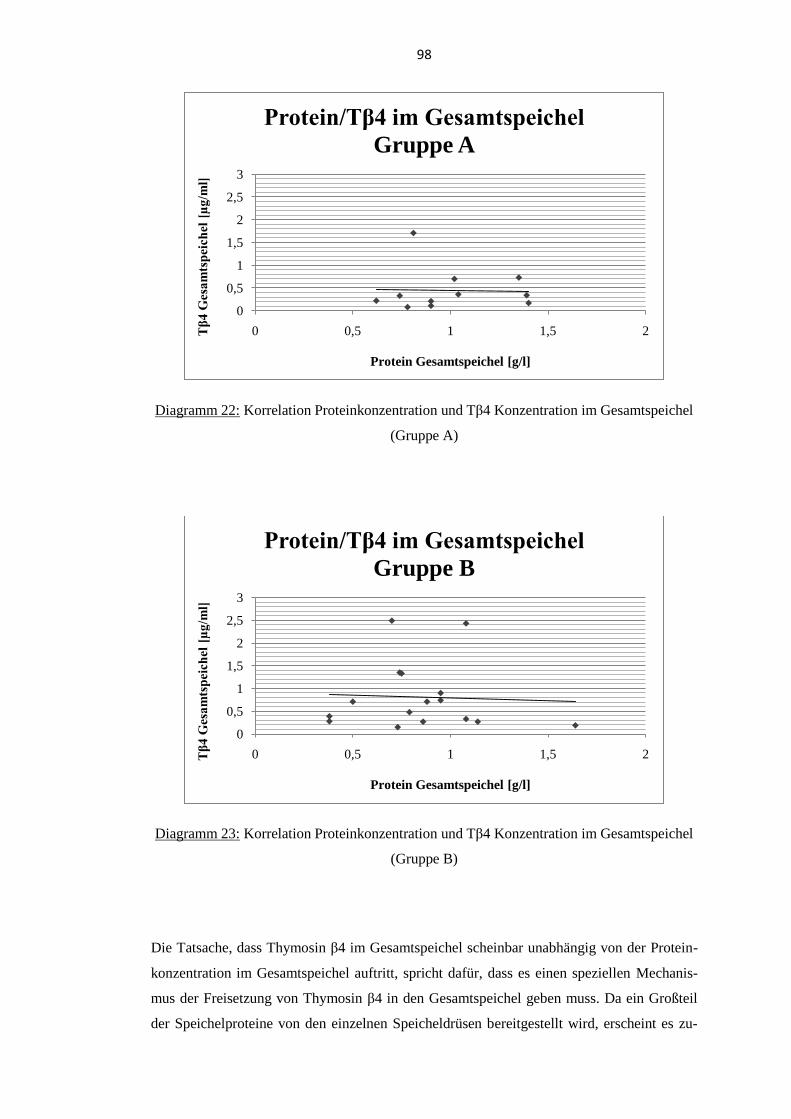
98
Diagramm 22: Korrelation Proteinkonzentration und Tβ4 Konzentration im Gesamtspeichel
(Gruppe A)
Diagramm 23: Korrelation Proteinkonzentration und Tβ4 Konzentration im Gesamtspeichel
(Gruppe B)
Die Tatsache, dass Thymosin β4 im Gesamtspeichel scheinbar unabhängig von der Protein-
konzentration im Gesamtspeichel auftritt, spricht dafür, dass es einen speziellen Mechanis-
mus der Freisetzung von Thymosin β4 in den Gesamtspeichel geben muss. Da ein Großteil
der Speichelproteine von den einzelnen Speicheldrüsen bereitgestellt wird, erscheint es zu-
0
0,5
1
1,5
2
2,5
3
0 0,5 1 1,5 2Tβ
4 G
esa
mts
pei
chel
[µ
g/m
l]
Protein Gesamtspeichel [g/l]
Protein/Tβ4 im Gesamtspeichel
Gruppe A
0
0,5
1
1,5
2
2,5
3
0 0,5 1 1,5 2Tβ
4 G
esa
mts
pei
chel
[µ
g/m
l]
Protein Gesamtspeichel [g/l]
Protein/Tβ4 im Gesamtspeichel
Gruppe B

99
dem als folgerichtig, dass die einzelnen Speicheldrüsen keine Quelle für orales Thymosin β4
darstellen. Ansonsten müsste eine hohe Proteinkonzentration im Gesamtspeichel mit einer
ebenfalls erhöhten Thymosin β4 Konzentration im Gesamtspeichel einhergehen. Wie aus den
beiden Diagrammen ersichtlich ist, ist dies ist aber eben nicht der Fall.
6.4. Korrelation Thymosin β4 Konzentration im Sulkusfluid und im Gesamtspeichel
(Gruppe C)
Bei einer genaueren Analyse der Ergebnisse aus der Gruppe C fallen einige Besonderheiten
auf. In der Gruppe C liegen die Thymosin β4 Konzentrationen im Gesamtspeichel bei sieben
der insgesamt elf untersuchten Proben unter der Nachweisgrenze von 0,05 µg/ml (siehe
Punkt 5.3.1. Diagramm 11). Dies steht im Gegensatz zu den Ergebnissen aus den Gruppen A
und B, bei denen nur in drei von insgesamt dreißig Gesamtspeichelproben kein Thymosin β4
in nachweisbaren Mengen bestimmt werden konnte (siehe Punkt 5.1.1. Diagramm 3 und
Punkt 5.1.2. Diagramm 5). Dieser Unterschied mag auf die im Vergleich zu den Gruppen A
und B veränderte Methode der Gesamtspeichelprobengewinnung zurückzuführen sein. Wäh-
rend die Probanden der Gruppen A und B „aktiv“ in ein Glas spuckten, wurde der Gesamt-
speichel in Gruppe C „passiv“ abgenommen, nachdem er sich in der Mundhöhle gesammelt
hatte. Angesichts des in dieser Arbeit erzielten Ergebnisses, dass das Sulkusfluid die Quelle
des oralen Thymosin β4 darstellt, liegt somit die Vermutung nahe, dass bei der „aktiven“
Methode der Probengewinnung in den Gruppen A und B ein größerer Anteil an Sulkusfluid
in den Gesamtspeichel gelangt ist als bei der „passiven“ Methode der Probengewinnung in
der Gruppe C.
Insofern sollte es auch als äußerst schwierig bezeichnet werden, eine direkte Korrelation
zwischen der Thymosin β4 Konzentration im Gesamtspeichel und im jeweiligen Sulkusfluid
herzustellen. Es gibt zu viele unbekannte beeinflussende und verfälschende Faktoren für die
Thymosin β4 Konzentration im Gesamtspeichel. So kann beispielsweise nicht genau vorher-
gesagt werden, welche Menge an Sulkusfluid in den Speichel fließt. Zudem kann eine er-
höhte Speichelfließrate die Konzentration des Sulkusfluids und damit des Thymosin β4 im
Gesamtspeichel verdünnen. Das Diagramm 24 stellt einen Zusammenhang zwischen der
Konzentration an Thymosin β4 im Sulkusfluid und der Thymosin β4 Konzentration in den
jeweiligen Gesamtspeichelproben dar. Es wurden dabei nur die Probanden aus der Gruppe C
berücksichtigt, bei denen Thymosin β4 im Gesamtspeichel nachweisbar war.
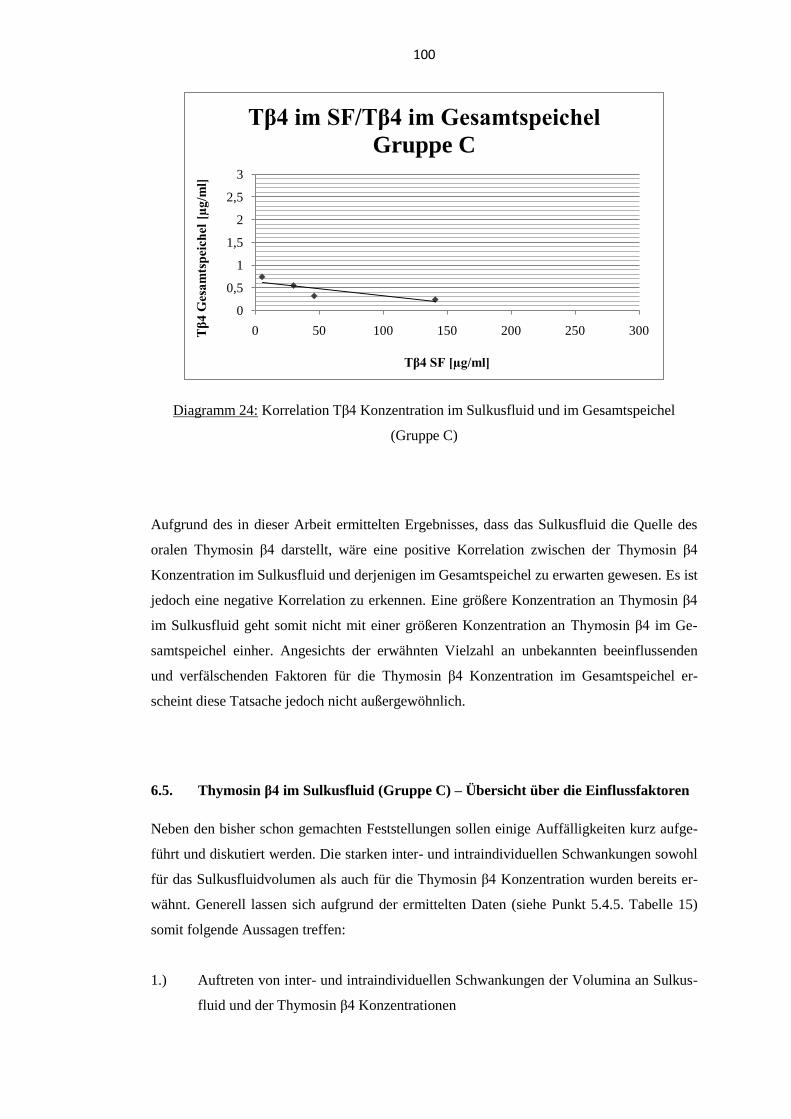
100
Diagramm 24: Korrelation Tβ4 Konzentration im Sulkusfluid und im Gesamtspeichel
(Gruppe C)
Aufgrund des in dieser Arbeit ermittelten Ergebnisses, dass das Sulkusfluid die Quelle des
oralen Thymosin β4 darstellt, wäre eine positive Korrelation zwischen der Thymosin β4
Konzentration im Sulkusfluid und derjenigen im Gesamtspeichel zu erwarten gewesen. Es ist
jedoch eine negative Korrelation zu erkennen. Eine größere Konzentration an Thymosin β4
im Sulkusfluid geht somit nicht mit einer größeren Konzentration an Thymosin β4 im Ge-
samtspeichel einher. Angesichts der erwähnten Vielzahl an unbekannten beeinflussenden
und verfälschenden Faktoren für die Thymosin β4 Konzentration im Gesamtspeichel er-
scheint diese Tatsache jedoch nicht außergewöhnlich.
6.5. Thymosin β4 im Sulkusfluid (Gruppe C) – Übersicht über die Einflussfaktoren
Neben den bisher schon gemachten Feststellungen sollen einige Auffälligkeiten kurz aufge-
führt und diskutiert werden. Die starken inter- und intraindividuellen Schwankungen sowohl
für das Sulkusfluidvolumen als auch für die Thymosin β4 Konzentration wurden bereits er-
wähnt. Generell lassen sich aufgrund der ermittelten Daten (siehe Punkt 5.4.5. Tabelle 15)
somit folgende Aussagen treffen:
1.) Auftreten von inter- und intraindividuellen Schwankungen der Volumina an Sulkus-
fluid und der Thymosin β4 Konzentrationen
0
0,5
1
1,5
2
2,5
3
0 50 100 150 200 250 300Tβ
4 G
esa
mts
pei
chel
[µ
g/m
l]
Tβ4 SF [µg/ml]
Tβ4 im SF/Tβ4 im Gesamtspeichel
Gruppe C

101
2.) Auftreten von inter- und intraindividuellen Schwankungen der sulkulären Taschen-
tiefen
3.) Tiefere Taschen bedeuten nicht immer mehr Sulkusfluidvolumen und eine höhere
Thymosin β4 Konzentration
4.) Mehr Sulkusfluidvolumen bedeutet nicht immer eine höhere Thymosin β4 Kon-
zentration
5.) Höhere API/SBI Werte bedeuten nicht immer mehr Sulkusfluidvolumen und eine
höhere Thymosin β4 Konzentration
6.) Höhere API/SBI Werte bedeuten nicht immer tiefere Taschen
Man kann zahlreiche Gründe für diese Feststellungen anführen. So sind die Ursachen gene-
rell zunächst in der Physiologie der Gegebenheiten zu suchen. Wie in der Einleitung bereits
erwähnt, wurde in der Literatur bisher in zahlreichen Studien berichtet, dass sowohl quantita-
tive als auch qualitative Unterschiede bestehen, wenn Sulkusfluidproben von verschiedenen
Sulki einer Person bzw. von Sulki unterschiedlicher Personen hinsichtlich verschiedener Pa-
rameter analysiert werden [107]. Die Parameter zeigen zudem oft keine zeitliche Konstanz,
sondern schwanken in ihren Werten. Somit entsprechen die in dieser Arbeit festgestellten
inter- und intraindividuellen Schwankungen sowohl des Sulkusfluidvolumens als auch der
Thymosin β4 Konzentration den bisher gemachten Beobachtungen.
Es gibt zahlreiche Erklärungsversuche für dieses Phänomen (vgl. [44]). Zunächst besteht ein
Problem in der Art der Probengewinnung. So könnte das Einbringen der Papierspitzen zu ei-
ner unbemerkten Traumatisierung bzw. Schädigung des Saumepithels geführt haben. Dies
wiederum hätte je nach Ausmaß der Schädigung qualitative und quantitative Veränderungen
der Sulkusfluidmenge und deren Zusammensetzung zur Folge.
Des Weiteren besteht ein Problem in der exakten Volumenbestimmung des Sulkusfluids und
damit wiederum ein Problem in der exakten Konzentrationsbestimmung von im Sulkusfluid
enthaltenen Substanzen. Da teilweise nur sehr geringe Volumina an Sulkusfluid gewonnen
werden konnten, hat eine geringe Verfälschung in der Volumenbestimmung eine große Aus-
wirkung auf die damit ermittelte Thymosin β4 Konzentration. So konnte in dieser Arbeit
durch zahlreiche Wiegeversuche zwar eine annähernd exakte Volumenbestimmung erreicht
werden (Daten nicht aufgeführt), das Problem der Verdunstung und damit der Aufkonzen-
trierung von Substanzen in der Papierspitze kann jedoch nicht vollständig ausgeschlossen
werden.
Eine mögliche Fehlerquelle, die zu Ungenauigkeiten führen kann, könnte darüber hinaus in
der Elution des von den Papierspitzen aufgesaugten Sulkusfluids liegen. Da die Wiederfin-
dungswerte jedoch durchschnittlich bei ca. 93 % liegen (siehe Punkt 5.7.), mag dieser Faktor
für eine exakte Bestimmung der Thymosin β4 Konzentration im Sulkusfluid jedoch wesent-

102
lich weniger ins Gewicht fallen, wie ein eventueller Fehler bei der Volumenbestimmung des
Sulkusfluids.
Zudem ergeben sich generell zahnspezifische Besonderheiten in der Anatomie der gingiva-
len Sulki, die sich generell sowohl inter- bzw. intraindividuell unterscheiden. Bei manchen
Probanden fiel das Einbringen der Papierspitzen sehr leicht, die Ausbeute an Sulkusfluid war
dann in der Regel dementsprechend höher. Bei anderen Probanden, bei denen nur ein kleiner
Teil der Papierspitze eingebracht werden konnte, war die Saugkraft der Papierspitze entspre-
chend limitiert und demnach waren die Volumina eher geringer.
Wenngleich bei der Probennahme der Sulkusfluidproben auf die Vermeidung einer mögli-
chen Kontamination mit Blut, Plaque und Speichel geachtet wurde, kann dennoch nicht aus-
geschlossen werden, dass eventuell unbemerkte Verunreinigungen und damit Verfälschun-
gen des bestimmten Sulkusfluidvolumens und damit der Thymosin β4 Konzentration bei ein-
zelnen Proben aufgetreten sind.
Des Weiteren sei noch angemerkt, dass die erhobenen klinischen Parameter wie Taschentiefe
und Mundhygiene- bzw. Entzündungsindex eher subjektive Parameter darstellen und die
klinische Situation in ihren vielen komplexen Facetten nicht exakt abbilden können.
6.6. Thymosin β4 im Sulkusfluid (Gruppe D) – Übersicht über die Einflussfaktoren
In Analogie zu den parodontal gesunden Probanden der Gruppe C waren in der Gruppe D die
wichtigsten Beobachtungen ebenfalls zunächst die inter- und intraindividuellen Schwankun-
gen des Sulkusfluidvolumens, der Thymosin β4 Konzentration im Sulkusfluid und der Ta-
schentiefen (siehe Punkt 5.5.5. Tabelle 18). Es gelten generell auch hier dieselben sechs Beo-
bachtungen und entsprechenden Erklärungsversuche, die auch bei der parodontal gesunden
Gruppe C gemacht werden konnten (siehe Punkt 6.5.) Die Erklärungsversuche für diese Phä-
nomene müssen bei den parodontal erkrankten Probanden jedoch um einige Aspekte erwei-
tert werden.
Wie in der Einleitung bereits erwähnt, konnte in zahlreichen Studien gezeigt werden, dass
eine deutliche Abhängigkeit zwischen der Quantität des Sulkusfluids und dem Gesundheits-
zustand des Parodonts besteht [23]. So kann die SFFR je nach Schweregrad der gingivalen
Entzündung bis um den Faktor dreißig variieren [42]. Des Weiteren ist aus der Literatur be-
kannt, dass Parodontopathien generell „site-specific“ verlaufen [22]. Das bedeutet, dass die
unterschiedlichen typischen Läsionstypen mit ihren unterschiedlichen Eigenschaften inner-
halb eines Gebisses gleichzeitig nebeneinander auftreten können. Zudem muss man bei Paro-
dontopathien generell zwischen aktiven und inaktiven Phasen unterscheiden. Des Weiteren
unterscheiden sich Parodontopathien in ihrem Verlauf und Ausmaß individuell teilweise sehr

103
stark. Grund hierfür ist die bei verschiedenen Individuen unterschiedlich stark ausgebildete
Fähigkeit zum Einleiten von Abwehrreaktionen auf die die Parodontopathien auslösende
Plaque, aber auch die Tatsache, dass zahlreiche lokale und systemische Faktoren die Patho-
genese einer Parodontopathie beeinflussen können. Zuletzt gibt es noch zahlreiche unter-
schiedliche Formen der Parodontopathien, die wiederum Unterschiede in der Pathogenese
zur Folge haben können.
Es wird somit schnell klar, dass man bei Betrachtung der ausgewerteten Ergebnisse für paro-
dontal erkrankte Probanden zahlreiche Faktoren berücksichtigen muss und zwangsläufig von
erheblichen intra- und interindividuellen Schwankungen der Thymosin β4 Konzentration und
des Sulkusfluidvolumens ausgehen muss. Die intraindividuellen Schwankungen der Thymo-
sin β4 Konzentrationen ergeben sich geradezu zwangsläufig aus dem „site-specific“ und zeit-
lich unterschiedlichen Verlauf der Parodontopathien. Die Tatsache, dass Individuen generell
unterschiedlich auf Erkrankungen des Parodonts ansprechen, lässt hingegen die interindivi-
duellen Unterschiede als logische Konsequenz erscheinen.
6.7. Thymosin β4 Konzentration im Sulkusfluid – Vergleich Gruppe C und D
Die genaue Bedeutung der für die parodontal erkrankten Probanden gefundenen Werte wird
jedoch erst durch einen Vergleich mit der parodontal gesunden Gruppe C deutlich. Sowohl
in der Gruppe C als auch in der Gruppe D traten intra- und interindividuelle Schwankungen
der Werte des Sulkusfluidvolumens, der Thymosin β4 Konzentration und der Taschentiefen
auf. Vergleicht man jedoch die Werte der Gruppe C mit denen der Gruppe D, so kommt man
zu einigen Beobachtungen. Die Tabelle 24 gibt einen Überblick über die gefundenen Sulkus-
fluidvolumina, die Thymosin β4 Konzentrationen und die Taschentiefen in den Gruppen C
(parodontal gesunde Probanden) und D (parodontal erkrankte Probanden).
Gruppe Taschentiefe [mm] Volumen SF [µl] Tβ4 [µg/ml]
C Wertebereich
Mittelwert
1,00 – 4,00
1,90
0,24 – 7,40
1,50
1,89 – 267,42
54,20
D Wertebereich
Mittelwert
2,00 – 12,00
3,40
0,37 – 10,26
3,00
2,16 – 82,52
28,30
Tabelle 24: Taschentiefe, Sulkusfluidvolumen und Tβ4 Konzentration im Sulkusfluid –
Vergleich Gruppe C (n = 11) und D (n = 8)

104
Die ersten Unterschiede zwischen den beiden Gruppen ergeben sich bei den ermittelten Ta-
schentiefen, die als Kriterium für die Zuteilung der Probanden in die jeweilige Gruppe ver-
wendet wurden. Parodontal gesunde Probanden der Gruppe C haben durchschnittlich Ta-
schentiefen von 1,90 mm. Die Einzelmessungen der Taschen liegen in einem Bereich von
1,00 – 4,00 mm. Die Taschentiefen parodontal erkrankter Probanden sind mit durchschnitt-
lich 3,40 mm dagegen größer. Die Einzelmessungen liegen in einem Bereich von 2,00 –
12,00 mm.
Die Sulkusfluidvolumina für parodontal gesunde Probanden der Gruppe C liegen in einem
Bereich von 0,24 – 7,40 µl. Durchschnittlich wurde ein Volumen von 1,50 µl gefunden. Im
Gegensatz dazu ist das durchschnittliche Sulkusfluidvolumen bei parodontal erkrankten Pro-
banden in der Gruppe D mit 3,00 µl doppelt so groß. Die einzelnen Volumina der Sulkus-
fluidproben liegen in einem Bereich von 0,37 – 10,26 µl. Parodontal erkrankte Probanden
haben demnach neben tieferen Taschen auch durchschnittlich größere Volumina an Sulkus-
fluid als parodontal gesunde Probanden.
Die Thymosin β4 Konzentrationen im Sulkusfluid parodontal gesunder Probanden liegen
zwischen 1,89 µg/ml und 267,42 µg/ml. Die durchschnittliche Konzentration beträgt 54,20
µg/ml. Damit ist sie fast doppelt so groß wie die durchschnittliche Konzentration an Thymo-
sin β4 in der Gruppe D mit 28,30 µg/ml. Die Konzentrationen der parodontal erkrankten Pro-
banden in der Gruppe D liegen in einem Bereich von 2,16 – 82,52 µg/ml. Somit lässt sich
insgesamt bei den parodontal erkrankten Probanden verglichen mit den parodontal gesunden
Probanden durchschnittlich ein doppelt so großes Sulkusfluidvolumen bei einer halb so
großen Konzentration an Thymosin β4 feststellen. Man kann daraus schließen, dass die Thy-
mosin β4 Konzentration bei den parodontal erkrankten Probanden infolge der verstärkt pro-
duzierten Sulkusfluidmenge entsprechend verdünnt wird.
In Analogie zu den Beobachtungen in den Gruppen A und B muss auch bei diesen Ergebnis-
sen angemerkt werden, dass für eine statistisch sichere Aussage ein größeres Probandenkol-
lektiv untersucht werden muss. Aus den angeführten Ergebnissen lassen sich somit allenfalls
Trends erkennen. Eine zusätzliche Schwierigkeit stellt die Einteilung der Probanden in die
jeweilige Gruppe dar. So könnten sich unter den Probanden der Gruppe C auch solche mit
entzündlichen gingivalen Veränderungen, bei denen noch keine Taschenvertiefung aufgetre-
ten ist, befinden. Andererseits könnten sich auch als parodontal erkrankt eingestufte Proban-
den gerade in einer Regenerations- oder Ruhephase der Parodontopathie befinden und nicht
die für einen akuten Verlauf typischen entzündlichen und immunologischen Symptome auf-
weisen.
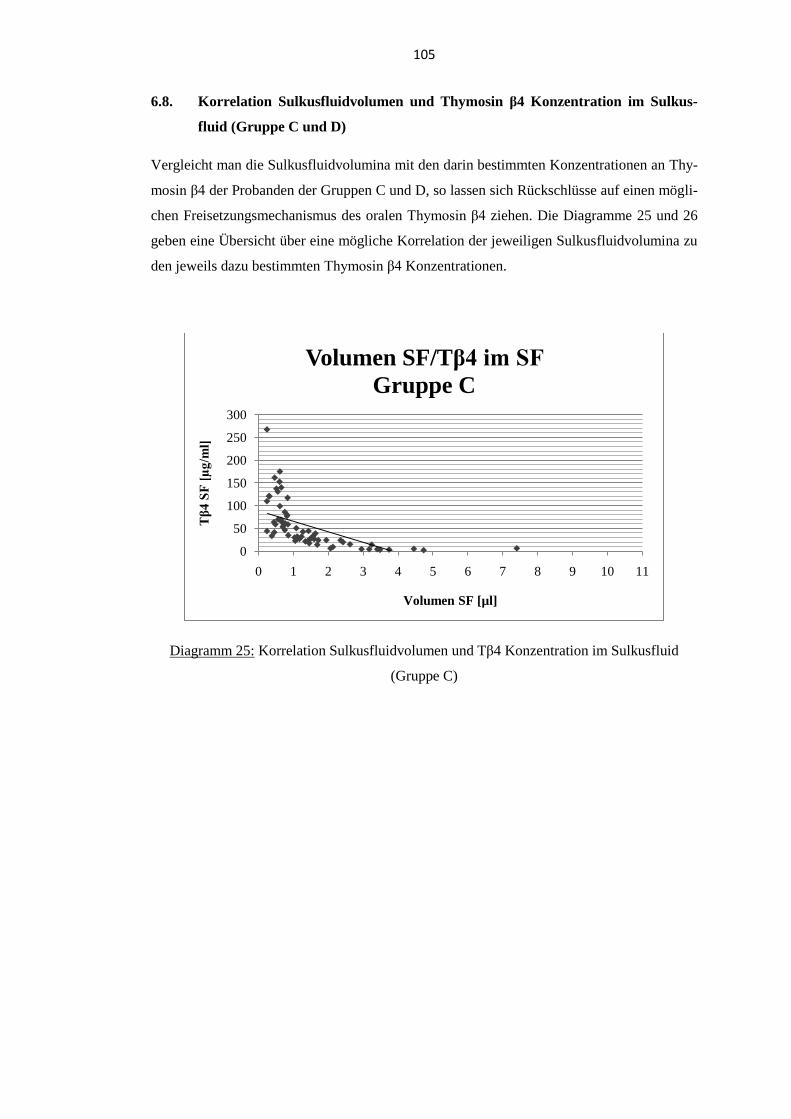
105
6.8. Korrelation Sulkusfluidvolumen und Thymosin β4 Konzentration im Sulkus-
fluid (Gruppe C und D)
Vergleicht man die Sulkusfluidvolumina mit den darin bestimmten Konzentrationen an Thy-
mosin β4 der Probanden der Gruppen C und D, so lassen sich Rückschlüsse auf einen mögli-
chen Freisetzungsmechanismus des oralen Thymosin β4 ziehen. Die Diagramme 25 und 26
geben eine Übersicht über eine mögliche Korrelation der jeweiligen Sulkusfluidvolumina zu
den jeweils dazu bestimmten Thymosin β4 Konzentrationen.
Diagramm 25: Korrelation Sulkusfluidvolumen und Tβ4 Konzentration im Sulkusfluid
(Gruppe C)
0
50
100
150
200
250
300
0 1 2 3 4 5 6 7 8 9 10 11
Tβ
4 S
F [
µg
/ml]
Volumen SF [µl]
Volumen SF/Tβ4 im SF
Gruppe C
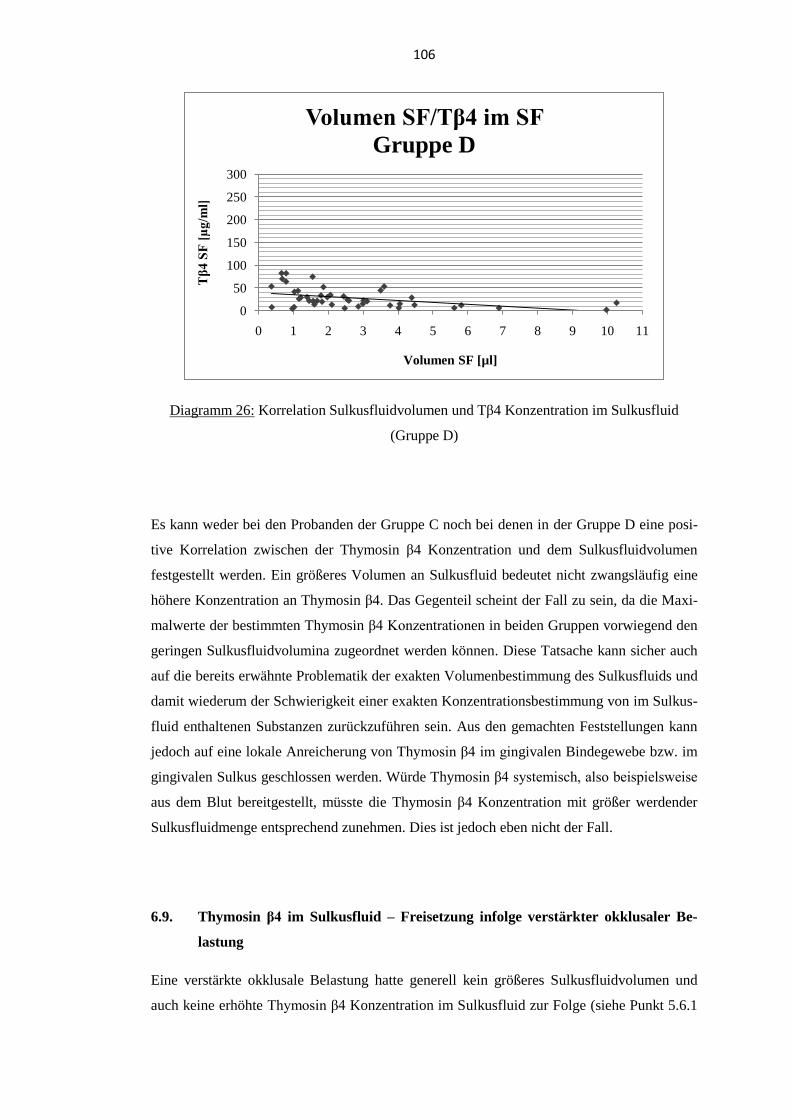
106
Diagramm 26: Korrelation Sulkusfluidvolumen und Tβ4 Konzentration im Sulkusfluid
(Gruppe D)
Es kann weder bei den Probanden der Gruppe C noch bei denen in der Gruppe D eine posi-
tive Korrelation zwischen der Thymosin β4 Konzentration und dem Sulkusfluidvolumen
festgestellt werden. Ein größeres Volumen an Sulkusfluid bedeutet nicht zwangsläufig eine
höhere Konzentration an Thymosin β4. Das Gegenteil scheint der Fall zu sein, da die Maxi-
malwerte der bestimmten Thymosin β4 Konzentrationen in beiden Gruppen vorwiegend den
geringen Sulkusfluidvolumina zugeordnet werden können. Diese Tatsache kann sicher auch
auf die bereits erwähnte Problematik der exakten Volumenbestimmung des Sulkusfluids und
damit wiederum der Schwierigkeit einer exakten Konzentrationsbestimmung von im Sulkus-
fluid enthaltenen Substanzen zurückzuführen sein. Aus den gemachten Feststellungen kann
jedoch auf eine lokale Anreicherung von Thymosin β4 im gingivalen Bindegewebe bzw. im
gingivalen Sulkus geschlossen werden. Würde Thymosin β4 systemisch, also beispielsweise
aus dem Blut bereitgestellt, müsste die Thymosin β4 Konzentration mit größer werdender
Sulkusfluidmenge entsprechend zunehmen. Dies ist jedoch eben nicht der Fall.
6.9. Thymosin β4 im Sulkusfluid – Freisetzung infolge verstärkter okklusaler Be-
lastung
Eine verstärkte okklusale Belastung hatte generell kein größeres Sulkusfluidvolumen und
auch keine erhöhte Thymosin β4 Konzentration im Sulkusfluid zur Folge (siehe Punkt 5.6.1
0
50
100
150
200
250
300
0 1 2 3 4 5 6 7 8 9 10 11
Tβ
4 S
F [
µg
/ml]
Volumen SF [µl]
Volumen SF/Tβ4 im SF
Gruppe D

107
Diagramm 18 und Punkt 5.6.2. Diagramm 19). Eine verstärkte okklusale Belastung stellt so-
mit keinen Freisetzungsmechanismus von intrazellulärem Thymosin β4 dar. Während der
Versuchsabfolge konnten jedoch weiterhin unterschiedlich starke inter- und intraindividuelle
Schwankungen der beiden Parameter Sulkusfluidvolumen und Thymosin β4 Konzentration
beobachtet werden.
Ein Teil der Schwankungen könnte hierbei aber zusätzlich noch methodisch bedingt sein. Da
die entsprechenden Sulkusfluidproben an den gleichen Zähnen in kurzen Abständen hinter-
einander genommen wurden, blieb unter Umständen nicht genügend Zeit für das Sulkusfluid
um in den gingivalen Sulkus nach einer vorher erfolgten Probennahme nachzufließen.
6.10. Stabilität von Thymosin β4 im Gesamtspeichel
In den Versuchen zur Stabilität von Thymosin β4 wurde untersucht, ob sich die Menge an
exogen zugegebenem Thymosin β4 in Gesamtspeichelproben über die Zeit hinweg verän-
dert. Insgesamt war die Konzentration an Thymosin β4 im Gesamtspeichel über lange Zeit
gesehen annähernd konstant, unabhängig davon ob der Proband vor der Zugabe von Thymo-
sin β4 bereits nachweisbare Mengen an Thymosin β4 im Speichel hatte oder nicht (siehe
Punkt 5.9. Diagramm 20 und 21). Damit lässt sich ausschließen, dass eventuell in der Mund-
höhle vorhandene Proteasen für die stark auftretenden interindividuellen Schwankungen der
Konzentrationen für orales Thymosin β4 verantwortlich sind. Gleichzeitig lässt sich eine
eventuelle Oxidation des Thymosin β4 im Gesamtspeichel ausschließen.
6.11. Vergleich der Ergebnisse mit dem aktuellen Stand der Forschung
Zum Abschluss der Diskussion, sollen die in dieser Arbeit gefundenen Ergebnisse mit denen
aus aktuellen Arbeiten verglichen werden. Badamchian et al. wiesen im Jahr 2007 erstmals
Thymosin β4 im Gesamtspeichel mit der ELISA-Methode nach [7]. Die gefundenen Thymo-
sin β4 Konzentrationen schwankten zwischen 0,2 und 3,6 µg/ml und waren sowohl vom Al-
ter als auch vom Geschlecht der Probanden abhängig. Die Probanden zwischen 25 und 35
Jahren hatten dabei die höchsten, die Probanden über 50 Jahre die niedrigsten Thymosin β4
Konzentrationen im Gesamtspeichel. Männliche Testpersonen hatten generell höhere Thy-
mosin β4 Konzentrationen im Gesamtspeichel.
Die in dieser Arbeit im Gesamtspeichel nachgewiesenen Thymosin β4 Konzentrationen
lagen in einem ähnlichen Bereich, wie die von Badamchian et al. zuvor bestimmten. Analog
zu Badamchian et al. schwankten die Thymosin β4 Konzentrationen interindividuell unter-

108
schiedlich stark, wobei die durchschnittlichen Thymosin β4 Konzentrationen kleiner waren
als bei Badamchian et al. Dies ist eventuell auf die angewendete ELISA-Nachweismethode
zurückzuführen, bei der auch Thymosin β4 Fragmente detektiert werden könnten. Unter-
schiede zu Badamchian et al. ergaben sich bei Betrachtung der Thymosin β4 Konzentration
in Abhängigkeit vom Alter. Im Gegensatz zu Badamchian et al. hatten in dieser Arbeit jün-
gere Probanden durchschnittlich geringere Thymosin β4 Konzentrationen als ältere Proban-
den. Badamchian et al. wiesen hingegen erhöhte Konzentrationen in der jüngeren Proban-
dengruppe nach. Im Gegensatz zu Badamchian et al. war zudem keine Geschlechtsabhängig-
keit der Thymosin β4 Konzentration nachweisbar.
Während des Verfassens der Arbeit gelang es der Gruppe um Inzitari et al. im Jahr 2009
ebenfalls das Sulkusfluid als Hauptquelle des oralen Thymosin β4 zu identifizieren [64].
Zum Nachweis des Thymosin β4 wurde hierbei die HPLC-ESI-MS-Methode angewendet.
Es wurden zwei Gruppen mit jeweils neun Probanden zwischen 21 und 45 Jahren untersucht.
Die Speichelproben wurden drüsenspezifisch mittels spezifischer Apparaturen gewonnen,
bzw. als Gesamtspeichel im vorderen Bereich des Mundbodens abgesaugt. Die Sulkusfluid-
proben wurden jeweils am unteren ersten Molar mittels Papierspitzen gewonnen. Die Sul-
kusfluidvolumina betrugen zwischen 1,5 und 3,7 µl und wurden durch Auswiegen bestimmt.
Eine mit 0,2 % TFA hergestellte Elutionslösung der Papierspitze wurde anschließend mittels
HPLC-ESI-MS-Methode auf den Gehalt an Thymosin β4 analysiert. Während im Speichel
der einzelnen Drüsen vernachlässigbar geringe Thymosin β4 Konzentrationen nachgewiesen
wurden, konnten im Sulkusfluid der Probanden Konzentrationen zwischen 55 und 145 µg/ml
quantifiziert werden. Zudem konnte stets, wenn auch in sehr geringen Mengen, Thymosin β4
Sulfoxid und Thymosin β10 nachgewiesen werden. Dabei ergab sich ein Verhältnis von Thy-
mosin β4 zu Thymosin β10 von 5 : 1. Desweitereren berichteten Inzitari et al. von einer line-
aren Korrelation zwischen der Thymosin β4 Konzentration im Gesamtspeichel und im Sul-
kusfluid. Zudem stellte die Gruppe eine lineare Korrelation zwischen der Thymosin β4 Kon-
zentration und der Thymosin β10 Konzentration im Sulkusfluid fest. Des Weiteren wurde
Speicheldrüsengewebe immunhistochemisch mit gegen Thymosin β4 gerichteten Antikör-
pern gefärbt, wobei jedoch nur eine geringe Immunreaktivität nachgewiesen werden konnte.
Die in dieser Arbeit ermittelten Werte für die Thymosin β4 Konzentrationen im Sulkusfluid
lagen in ähnlichen Bereichen wie von Inzitari et al. berichtet, wobei der Bereich jedoch we-
sentlich größer war als bei Inzitari et al.. Im Unterschied zu Inzitari et al. konnte keine Kor-
relation der Thymosin β4 Konzentration im Gesamtspeichel und im Sulkusfluid nachgewie-
sen werden. Thymosin β10 konnte nicht nachgewiesen werden.

109
6.12. Mechanismus der Anreicherung von Thymosin β4 im Sulkusfluid
Eine verstärkte okklusale Belastung stellt wie dargelegt keinen Freisetzungsmechanismus für
intrazelluläres Thymosin β4 in das Sulkusfluid dar. Aufgrund der in den Gruppen C und D
gemachten Beobachtungen kann jedoch über den Mechanismus der Anreicherung von Thy-
mosin β4 im Sulkusfluid unter Berücksichtigung der folgenden Parameter spekuliert werden.
Die Produktion des Sulkusfluids wird durch den Übertritt von Flüssigkeit aus den Kapillaren
des gingivalen Gefäßplexus in das gingivale Bindegewebe und dessen anschließende Wie-
deraufnahme durch lymphatische Gefäße geregelt (siehe Punkt 1.6.2.4.). Das Sulkusfluidvo-
lumen und die SFFR repräsentieren demnach die momentane Gefäßpermeabilität. Sie sind
folglich keine konstanten Parameter, sondern unterscheiden sich sowohl intra- als auch inter-
individuell. Es muss im Sulkusfluid grundsätzlich zwischen Substanzen unterschieden wer-
den, die systemisch bereitgestellt werden und solchen, die lokal im Bindegewebe angerei-
chert werden. Letztere werden dann mit dem Sulkusfluid in den Sulkus gespült und dort an-
gereichert. Ein solcher Mechanismus der lokalen Anreicherung kann beispielsweise auf dem
Prinzip der Zelllyse oder der Mediator-vermittelten Freisetzung der entsprechenden Substanz
aus Zellen basieren. Wenn eine Substanz systemisch bereitgestellt wird, müsste deren Kon-
zentration im Sulkusfluid mit einer erhöhten SFFR folglich steigen. Würde die Substanz
durch lokale Anreicherung im Bindegewebe bereitgestellt, würde eine erhöhte SFFR entspre-
chend zu einer Erniedrigung der Konzentration der Substanz im Sulkusfluid führen. Im Falle
des im Sulkusfluid nachgewiesenen Thymosin β4 scheint letzteres Prinzip zu gelten. Grund
für diese Behauptung ist die Tatsache, dass bei parodontal erkrankten Probanden bei einer im
Vergleich zu parodontal gesunden Probanden durchschnittlich doppelten Menge an Sulkus-
fluid die Konzentration an Thymosin β4 durchschnittlich halbiert war (siehe Punkt 6.7.
Tabelle 24). Die größere Menge an Sulkusfluid hatte die Thymosin β4 Konzentration ent-
sprechend verdünnt. Die Ergebnisse legen demzufolge nahe, dass es besondere Mechanis-
men zur lokalen Anreicherung von Thymosin β4 im gingivalen Bindegewebe und folglich
im Sulkusfluid geben muss. Eine erhöhte mikrovaskuläre Permeabilität des gingivalen Ge-
fäßplexus und eine dadurch bedingte systemische Freisetzung von Thymosin β4 aus dem
Blut kann die hohen extrazellulären Konzentrationen an Thymosin β4 im Sulkusfluid nicht
erklären, zumal diese wesentlich größer sind als im Blut [50]. Während die Produktion des
Sulkusfluids von Mediatoren der vaskulären Permeabilität abhängig ist, könnte die Freisetz-
ung von Thymosin β4 in den Sulkus von spezifischen noch nicht identifizierten Mediatoren
abhängen. Dies würde jedoch bedeuten, dass Thymosin β4 zuvor in den Zellen durch aktive
Aufnahme angereichert werden müsste. Aufgrund der vergleichsweise eher geringen Kon-
zentrationen von intrazellulärem Thymosin β4 würde zum Erreichen der im Sulkusfluid
nachgewiesenen Thymosin β4 Konzentrationen ansonsten eine sehr große Anzahl an Zellen
nötig sein. Unter Umständen lassen sich diese Besonderheiten demnach aufgrund des spe-

110
ziellen histologischen Aufbaus der den gingivalen Sulkus auskleidenden Saumepithelzellen
erklären. Dieser unterscheidet sich wie dargelegt grundlegend von dem anderer Bindege-
webs- und Epithelzellen.
6.13. Mögliche Rolle von Thymosin β4 bei Parodontopathien
Die Konzentrationen an Thymosin β4 im Sulkusfluid waren sowohl bei parodontal gesunden
als auch bei parodontal erkrankten Probanden außergewöhnlich hoch. Bei parodontal erkran-
kten Probanden wurden durchschnittlich niedrigere Thymosin β4 Konzentrationen nachge-
wiesen als bei parodontal gesunden (siehe Punkt 6.7. Tabelle 24). Diese Beobachtungen und
die Tatsache, dass einige Funktionen von Thymosin β4 eng mit der Pathogenese von Paro-
dontopathien verbunden zu sein scheinen, legen die Vermutung nahe, dass Thymosin β4 eine
mögliche Rolle bei Parodontopathien spielen könnte. Dies könnte wiederum sowohl von
diagnostischer oder aber auch von therapeutischer Bedeutung sein. Thymosin β4 könnte da-
bei entweder ein parodontal pathogenes oder ein parodontal protektives Potential besitzen.
Die Tatsache, dass Thymosin β4 die Expression von Matrixmetalloproteinasen (MMPs) in
Endothelzellen induziert [90], spricht für eine mögliche pathogene Wirkung von Thymosin
β4 bei Parodontopathien. Diese MMPs sind nach einer entsprechenden Aktivierung haupt-
verantwortlich für den destruktiven Abbau des gingivalen Bindegewebes [99].
Orales Thymosin β4 könnte aber auch parodontal protektive Wirkungen besitzen. So könnte
Thymosin β4 im Rahmen seiner Funktion der Gefäßneubildung durch verstärkte Expression
des VEGF dazu beitragen, dass am Ort der Entzündung bei einer Parodontopathie vermehrt
Abwehrzellen aus dem Blut bereitgestellt werden [21]. Zudem wird berichtet, dass das Sulf-
oxid von Thymosin β4 anti-inflammatorische Eigenschaften besitzt [124]. Dies geschieht
durch Beeinflussung der Expression von verschiedenen Zytokinen bzw. der Beeinflussung
von Vorgängen wie der Chemotaxis von Abwehrzellen. Die Tatsache, dass nur die oxidierte
Form des Thymosin β4 diese Funktion ausüben soll, ließe sich damit erklären, dass Thymo-
sin β4 durch die bei Parodontopathien entsprechend vorhandene oxidative Umgebung der
Apoptose zunächst aktiviert wird. Die Oxidation könnte aber auch durch die bei Parodonto-
pathien stets gebildeten reaktiven Sauerstoffspezies geschehen.
Wenngleich einige dieser Vermutungen noch spekulativ sind und aktiv erforscht werden
müssen, so lässt sich ein direkter Zusammenhang zwischen Thymosin β4 und der Physiolo-
gie des Parodonts nicht bestreiten. Eine erst im Verlauf des Verfassens dieser Arbeit ver-
öffentlichte Studie von Reti et al. aus dem Jahr 2008 berichtete erstmals von einem zytopro-
tektiven und anti-inflammatorischen in vitro Effekt von Thymosin β4 auf humane Gingivafi-
broblasten [94]. Die Gruppe wies außerdem nach, dass Lipopolysaccharide zu einer verstärk-

111
ten Freisetzung von Thymosin β4 durch humane Gingivafibroblasten führen. Berücksichtigt
man die Tatsache, dass Lipopolysaccharide als bakterielle Endotoxine vorzugsweise in der
dentalen Plaque auftreten und hauptverantwortlich für die Pathogenese und die Etablierung
einer Gingivitis bzw. Parodontitis sind, so wird das weite Feld der therapeutischen Möglich-
keiten des oralen Thymosin β4 offensichtlich. So kamen Reti et al. damals bereits zu der
Schlussfolgerung, dass Thymosin β4 zwar wahrscheinlich ungeeignet ist, eine Lipopoly-
saccharid-induzierte Entzündung im Anfangsstadium einer Gingivitis bzw. Parodontitis zu
unterdrücken, es jedoch durchaus dafür geeignet sein kann, den durch pro-inflammatorische
Zytokine im Verlauf der Parodontalerkrankung ausgelösten intrazellulären Schaden zu be-
grenzen. Somit wäre ein möglicher Therapieansatz für orales Thymosin β4 im Rahmen einer
Parodontalerkrankung durch die Regulation der spezifischen Entzündungsmediatoren gege-
ben. Thymosin β4 hätte aufgrund seiner anti-mikrobiellen, anti-inflammatorischen, anti-
apoptotischen und zytoprotektiven Eigenschaften folglich ein äußerst großes Anwendungs-
gebiet als orales Medikament.
6.14. Thymosin β4 als mögliches Diagnostiktool für Parodontopathien
Neben einer möglichen therapeutischen Verwendung von Thymosin β4 bei Parodontopa-
thien, könnte es auch in diagnostischer Hinsicht von großer Bedeutung sein. Da Thymosin
β4 im Sulkusfluid in erstaunlich hohen Konzentrationen auftritt und somit leicht nachweisbar
ist, könnte es eventuell als ein Diagnostiktool für die Diagnose und Prognose von Parodonto-
pathien verwendet werden.
Die Suche nach einem Marker für Parodontopathien ergibt sich generell aus der Notwendig-
keit exaktere Diagnosen und erfolgreichere Therapieschemata für Parodontopathien zu ent-
wickeln. Die bisher angewendeten traditionellen klinischen Parameter wie Messung der Ta-
schentiefe und des Knochen- bzw. des Attachmentverlustes liefern hierfür zu ungenaue Er-
gebnisse. Da in zahlreichen Studien inflammatorische Zytokine im Sulkusfluid nachgewie-
sen werden konnten, dienten diese zunächst als Indikator des lokalen immunregulatorischen
und inflammatorischen Zustandes. Neben diesen Mediatoren wurden bisher Wirts-stämmige
Enzyme und Gewebszerfallsprodukte als mögliche Marker untersucht. Insgesamt wurden bis
heute mehr als 65 einzelne Marker analysiert. Nur wenige lieferten jedoch vielversprechende
und reproduzierbare Ergebnisse. Thymosin β4 könnte aufgrund seiner hohen Konzentration
und der nachweislichen Korrelation zum Parodontalstatus des Probanden als Diagnostiktool
zwar dazu verwendet werden um parodontal erkrankte von parodontal gesunden Probanden
zu unterscheiden, es ist jedoch zu bezweifeln, dass Thymosin β4 als Diagnostiktool sensitiv
genug für die Erkennung der unterschiedlichen Formen der Parodontopathien wäre.

112
Literaturverzeichnis
1. Akhter, M., I. Kobayashi, T. Kiyoshima, K. Matsuo, H. Yamaza, H. Wada, J.Y.
Honda, X. Ming, and H. Sakai, (2005), Possible functional involvement of thymosin
beta 4 in developing tooth germ of mouse lower first molar, Histochem Cell Biol,
124(3-4): p. 207-213.
2. Alfano, M.C., (1974), The origin of gingival fluid, J Theor Biol, 47(1): p. 127-136.
3. Armitage, G.C., (1995), Clinical evaluation of periodontal diseases, Periodontol
2000, 7: p. 39-53.
4. Armitage, G.C., (1999), Development of a classification system for periodontal
diseases and conditions, Ann Periodontol, 4(1): p. 1-6.
5. Armitage, G.C., (2003), Diagnosis of periodontal diseases, J Periodontol, 74(8): p.
1237-1247.
6. Attstrom, R., (1970), Presence of leukocytes in crevices of healthy and chronically
inflamed gingivae, J Periodontal Res, 5(1): p. 42-47.
7. Badamchian, M., A.A. Damavandy, H. Damavandy, S. Wadhwa, B. Katz, and A.L.
Goldstein, (2007), Identification and Quantification of Thymosin 4 in Human Saliva
and Tears, Ann N Y Acad Sci, 1112: p. 458-465.
8. Badamchian, M., M.O. Fagarasan, R.L. Danner, A.F. Suffredini, H. Damavandy,
and A.L. Goldstein, (2003), Thymosin 4 reduces lethality and down-regulates
inflammatory mediators in endotoxin-induced septic shock, Int Immunopharmacol,
3(8): p. 1225-1233.
9. Ballweber, E., E. Hannappel, T. Huff, and H.G. Mannherz, (1997), Mapping the
binding site of thymosin beta4 on actin by competition with G-actin binding proteins
indicates negative co-operativity between binding sites located on opposite
subdomains of actin, Biochem J, 327(Pt 3): p. 787-793.
10. Bao, L., M. Loda, and B.R. Zetter, (1998), Thymosin beta15 expression in tumor cell
lines with varying metastatic potential, Clin Exp Metastasis, 16(3): p. 227-233.
11. Bartold, P.M., (2006), Periodontal tissues in health and disease: introduction,
Periodontol 2000, 40: p. 7-10.
12. Bartold, P.M. and A.S. Narayanan, (2006), Molecular and cell biology of healthy
and diseased periodontal tissues, Periodontol 2000, 40: p. 29-49.
13. Baum, B.J., (1989), Salivary gland fluid secretion during aging, J Am Geriatr Soc,
37(5): p. 453-458.
14. Baum, B.J., (1993), Principles of saliva secretion, Ann N Y Acad Sci, 694: p. 17-23.
15. Black, G., (1899), The fibers and glands of the peridental membrane, Dent Cosmos,
41: p. 101-122.
16. Bodendorf, S., G. Born, and E. Hannappel, (2007), Determination of thymosin beta4
and protein in human wound fluid after abdominal surgery, Ann N Y Acad Sci,
1112: p. 418-424.

113
17. Brill, N., (1959), Removal of particles and bacteria from gingival pockets by tissue
fluid, Acta Odontol Scand, 17: p. 431-440.
18. Brill, N., Krasse B., (1958), The passage of tissue fluid into the clinically healthy
gingival pocket, Acta Odontol Scand, 16: p. 233-245.
19. Bubb, M.R., (2003), Thymosin β-4 interactions. Vitamins and Hormones, 66: p. 297-
316.
20. Burnet, M., (1962), Role of the thymus and related organs in immunity, Br Med J,
2(5308): p. 807-811.
21. Cha, H.J., M.J. Jeong, and H.K. Kleinman, (2003), Role of thymosin beta4 in tumor
metastasis and angiogenesis, J Natl Cancer Inst, 95(22): p. 1674-1680.
22. Champagne, C.M., W. Buchanan, M.S. Reddy, J.S. Preisser, J.D. Beck, and S.
Offenbacher, (2003), Potential for gingival crevice fluid measures as predictors of
risk for periodontal diseases, Periodontol 2000, 31: p. 167-180.
23. Cimasoni, G., (1983), Crevicular fluid updated, Monogr Oral Sci, 12: p. III-VII, 1-
152.
24. Curtis, M.A., G.S. Griffiths, S.J. Price, S.K. Coulthurst, and N.W. Johnson, (1988),
The total protein concentration of gingival crevicular fluid. Variation with sampling
time and gingival inflammation, J Clin Periodontol, 15(10): p. 628-632.
25. Czisch, M., M. Schleicher, S. Horger, W. Voelter, and T.A. Holak, (1993),
Conformation of thymosin beta 4 in water determined by NMR spectroscopy, Eur J
Biochem, 218(2): p. 335-344.
26. Dawes, C., (1974), Rhythms in salivary flow rate and composition, Int J Chronobiol,
2(3): p. 253-279.
27. Dawes, C., (1987), Physiological factors affecting salivary flow rate, oral sugar
clearance, and the sensation of dry mouth in man, J Dent Res, 66 Spec No: p. 648-
653.
28. Edgar, W.M., (1992), Saliva: its secretion, composition and functions, Br Dent J,
172(8): p. 305-312.
29. Egelberg, J., (1966), Permeability of the dento-gingival blood vessels. 3. Chronically
inflamed gingivae, J Periodontal Res, 1(4): p. 287-296.
30. Erickson-Viitanen, S., S. Ruggieri, P. Natalini, and B.L. Horecker, (1983),
Distribution of thymosin beta 4 in vertebrate classes, Arch Biochem Biophys,
221(2): p. 570-576.
31. Erickson-Viitanen, S., S. Ruggieri, P. Natalini, and B.L. Horecker, (1983), Thymosin
beta 10, a new analog of thymosin beta 4 in mammalian tissues, Arch Biochem
Biophys, 225(2): p. 407-413.
32. Franco, F.J., C. Diaz, M. Barcia, and M. Freire, (1992), Thymosin alpha 1 is a native
peptide in several tissues, Biochim Biophys Acta, 1120(1): p. 43-48.

114
33. Frei, R.W., L. Michel, and W. Santi, (1976), Post-column fluorescence
derivatization of peptides. Problems and potential in high-performance liquid
chromatography, J Chromatogr, 126: p. 665-677.
34. Gold, S.I., (1985), Periodontics. The past. Part (I). Early sources, J Clin
Periodontol, 12(2): p. 79-97.
35. Goldstein, A.L., (2007), History of the discovery of the thymosins, Ann N Y Acad
Sci, 1112: p. 1-13.
36. Goldstein, A.L. and M. Badamchian, (2004), Thymosins: chemistry and biological
properties in health and disease, Expert Opin Biol Ther, 4(4): p. 559-573.
37. Goldstein, A.L., A. Guha, M.L. Howe, and A. White, (1971), Ontogenesis of cell-
mediated immunity in murine thymocytes and spleen cells and its acceleration by
thymosin, a thymic hormone, J Immunol, 106(3): p. 773-780.
38. Goldstein, A.L., A. Guha, M.M. Zatz, M.A. Hardy, and A. White, (1972),
Purification and biological activity of thymosin, a hormone of the thymus gland,
Proc Natl Acad Sci U S A, 69(7): p. 1800-1803.
39. Goldstein, A.L., T.L. Low, M. McAdoo, J. McClure, G.B. Thurman, J. Rossio, C.Y.
Lai, D. Chang, S.S. Wang, C. Harvey, A.H. Ramel, and J. Meienhofer, (1977),
Thymosin alpha1: isolation and sequence analysis of an immunologically active
thymic polypeptide, Proc Natl Acad Sci U S A, 74(2): p. 725-729.
40. Goldstein, A.L., F.D. Slater, and A. White, (1966), Preparation, assay, and partial
purification of a thymic lymphocytopoietic factor (thymosin), Proc Natl Acad Sci U
S A, 56(3): p. 1010-1017.
41. Goodall, G.J., J.I. Morgan, and B.L. Horecker, (1983), Thymosin beta 4 in cultured
mammalian cell lines, Arch Biochem Biophys, 221(2): p. 598-601.
42. Goodson, J.M., (2003), Gingival crevice fluid flow, Periodontol 2000, 31: p. 43-54.
43. Gornall, A.G., C.J. Bardawill, and M.M. David, (1949), Determination of serum
proteins by means of the biuret reaction, J Biol Chem, 177(2): p. 751-766.
44. Griffiths, G.S., (2003), Formation, collection and significance of gingival crevice
fluid, Periodontol 2000, 31: p. 32-42.
45. Hall, A.K., (1991), Differential expression of thymosin genes in human tumors and
in the developing human kidney, Int J Cancer, 48(5): p. 672-677.
46. Hall, A.K., J. Hempstead, and J.I. Morgan, (1990), Thymosin beta 10 levels in
developing human brain and its regulation by retinoic acid in the HTB-10
neuroblastoma, Brain Res Mol Brain Res, 8(2): p. 129-135.
47. Hannappel, E., (1986), One-step procedure for the determination of thymosin beta 4
in small tissue samples and its separation from other thymosin beta 4-like peptides
by high-pressure liquid chromatography, Anal Biochem, 156(2): p. 390-396.
48. Hannappel, E., (2007), beta-Thymosins, Ann N Y Acad Sci, 1112: p. 21-37.

115
49. Hannappel, E., S. Davoust, and B.L. Horecker, (1982), Thymosins beta 8 and beta 9:
two new peptides isolated from calf thymus homologous to thymosin beta 4, Proc
Natl Acad Sci U S A, 79(6): p. 1708-1711.
50. Hannappel, E. and M. van Kampen, (1987), Determination of thymosin beta 4 in
human blood cells and serum, J Chromatogr, 397: p. 279-285.
51. Hannappel, E., G.J. Xu, J. Morgan, J. Hempstead, and B.L. Horecker, (1982),
Thymosin beta 4: a ubiquitous peptide in rat and mouse tissues, Proc Natl Acad Sci
U S A, 79(7): p. 2172-2175.
52. Hellwig, E., Klimek, J., Attin, T., Einführung in der Zahnerhaltung, 4. Auflage,
Urban und Fischer Verlag München, 2007: p. 10-11.
53. Hellwig, E., Klimek, J., Attin, T., Einführung in die Zahnerhaltung, 4. Auflage,
Urban und Fischer Verlag München, 2007: p. 361-370.
54. Hellwig, E., Klimek, J., Attin, T., Einführung in die Zahnerhaltung, 4. Auflage,
Urban und Fischer Verlag München, 2007: p. 391-398.
55. Hellwig, E., Klimek, J., Attin, T., Einführung in die Zahnerhaltung, 4. Auflage,
Urban und Fischer Verlag München, 2007: p. 371-390.
56. Henderson, B., S. Poole, and M. Wilson, (1996), Microbial/host interactions in
health and disease: who controls the cytokine network? , Immunopharmacology,
35(1): p. 1-21.
57. Hooper, J.A., M.C. McDaniel, G.B. Thurman, G.H. Cohen, R.S. Schulof, and A.L.
Goldstein, (1975), Purification and properties of bovine thymosin, Ann N Y Acad
Sci, 249: p. 125-144.
58. Huff, T., E. Ballweber, A. Humeny, T. Bonk, C. Becker, C.S. Muller, H.G.
Mannherz, and E. Hannappel, (1999), Thymosin beta(4) serves as a glutaminyl
substrate of transglutaminase. Labeling with fluorescent dansylcadaverine does not
abolish interaction with G-actin, FEBS Lett, 464(1-2): p. 14-20.
59. Huff, T. and E. Hannappel, (1997), Oxidation and reduction of thymosin beta(4) and
its influence on the interaction with G-actin studied by reverse-phase HPLC and
post-column derivatization with fluorescamine, Analytica Chimica Acta, 352(1-3): p.
249-255.
60. Huff, T., C. Muller, and E. Hannappel, (1997), HPLC and post-column
derivatization with fluorescamine: isolation of actin-sequestering beta-thymosins by
reversed-phase HPLC, Analytica Chimica Acta, 352(1-3): p. 239-248.
61. Huff, T., C.S. Muller, A.M. Otto, R. Netzker, and E. Hannappel, (2001), beta-
Thymosins, small acidic peptides with multiple functions, Int J Biochem Cell Biol,
33(3): p. 205-220.
62. Huff, T., A.M. Otto, C.S. Muller, M. Meier, and E. Hannappel, (2002), Thymosin
beta4 is released from human blood platelets and attached by factor XIIIa
(transglutaminase) to fibrin and collagen, Faseb J, 16(7): p. 691-696.
63. Humphrey, S.P. and R.T. Williamson, (2001), A review of saliva: normal
composition, flow, and function, J Prosthet Dent, 85(2): p. 162-169.

116
64. Inzitari, R., T. Cabras, E. Pisano, C. Fanali, B. Manconi, E. Scarano, A. Fiorita, G.
Paludetti, A. Manni, S. Nemolato, G. Faa, M. Castagnola, and I. Messana, (2009),
HPLC-ESI-MS analysis of oral human fluids reveals that gingival crevicular fluid is
the main source of oral thymosins beta(4) and beta(10), J Sep Sci, 32(1): p. 57-63.
65. Jeffery, C.J., (1999), Moonlighting proteins, Trends Biochem Sci, 24(1): p. 8-11.
66. Kaplan, M.D. and B.J. Baum, (1993), The functions of saliva, Dysphagia, 8(3): p.
225-229.
67. Kinane, D.F., D.F. Lappin, O. Koulouri, and A. Buckley, (1999), Humoral immune
responses in periodontal disease may have mucosal and systemic immune features,
Clin Exp Immunol, 115(3): p. 534-541.
68. Klein, J.J., A.L. Goldstein, and A. White, (1965), Enhancement of in Vivo
Incorporation of Labeled Precursors into DNA and Total Protein of Mouse Lymph
Nodes after Administration of Thymic Extracts, Proc Natl Acad Sci U S A, 53: p.
812-817.
69. Laine, F.J. and W.R. Smoker, (1995), Oral cavity: anatomy and pathology, Semin
Ultrasound CT MR, 16(6): p. 527-545.
70. Law, L.W., A.L. Goldstein, and A. White, (1968), Influence of thymosin on
immunological competence of lymphoid cells from thymectomized mice, Nature,
219(5161): p. 1391-1392.
71. Llena-Puy, C., (2006), The role of saliva in maintaining oral health and as an aid to
diagnosis, Med Oral Patol Oral Cir Bucal, 11(5): p. E449-455.
72. Löffler, G., Petrides, P., Heinrich, P., Biochemie und Pathobiochemie, 8. Auflage,
Springer Medizin Verlag Heidelberg, 2007: p. 59-62.
73. Low, T.L. and A.L. Goldstein, (1982), Chemical characterization of thymosin 4, J
Biol Chem, 257(2): p. 1000-1006.
74. Low, T.L., S.K. Hu, and A.L. Goldstein, (1981), Complete amino acid sequence of
bovine thymosin beta 4: a thymic hormone that induces terminal deoxynucleotidyl
transferase activity in thymocyte populations, Proc Natl Acad Sci U S A, 78(2): p.
1162-1166.
75. Low, T.L., D.T. Liu, and J.H. Jou, (1992), Primary structure of thymosin beta 12, a
new member of the beta-thymosin family isolated from perch liver, Arch Biochem
Biophys, 293(1): p. 32-39.
76. Low, T.L., G.B. Thurman, M. McAdoo, J. McClure, J.L. Rossio, P.H. Naylor, and
A.L. Goldstein, (1979), The chemistry and biology of thymosin. I. Isolation,
characterization, and biological activities of thymosin alpha1 and polypeptide beta1
from calf thymus, J Biol Chem, 254(3): p. 981-986.
77. Malinda, K.M., A.L. Goldstein, and H.K. Kleinman, (1997), Thymosin beta 4
stimulates directional migration of human umbilical vein endothelial cells, Faseb J,
11(6): p. 474-481.

117
78. Malinda, K.M., G.S. Sidhu, H. Mani, K. Banaudha, R.K. Maheshwari, A.L.
Goldstein, and H.K. Kleinman, (1999), Thymosin beta4 accelerates wound healing,
J Invest Dermatol, 113(3): p. 364-368.
79. Mannherz, H.G. and E. Hannappel, (2009), The beta-thymosins: intracellular and
extracellular activities of a versatile actin binding protein family, Cell Motil
Cytoskeleton, 66(10): p. 839-851.
80. Miller, J.F., (1961), Immunological function of the thymus, Lancet, 2(7205): p. 748-
749.
81. Moscinski, L.C., P.H. Naylor, J. Oliver, and A.L. Goldstein, (1993), Thymosin beta
4 synergizes with human granulocyte-macrophage colony-stimulating factor in
maintaining bone marrow proliferation, Immunopharmacology, 26(1): p. 83-92.
82. Muhlemann, H.R. and S. Son, (1971), Gingival sulcus bleeding--a leading symptom
in initial gingivitis, Helv Odontol Acta, 15(2): p. 107-113.
83. Nanci, A. and D.D. Bosshardt, (2006), Structure of periodontal tissues in health and
disease, Periodontol 2000, 40: p. 11-28.
84. Navazesh, M., (1993), Methods for collecting saliva, Ann N Y Acad Sci, 694: p. 72-
77.
85. Orban, B., (1942), Classification and nomenclature of periodontal diseases, J
Periodontol, 13: p. 88-91.
86. Page, R.C. and J.D. Beck, (1997), Risk assessment for periodontal diseases, Int Dent
J, 47(2): p. 61-87.
87. Pashley, D.H., (1976), A mechanistic analysis of gingival fluid production, J
Periodontal Res, 11(2): p. 121-134.
88. Paster, B.J., S.K. Boches, J.L. Galvin, R.E. Ericson, C.N. Lau, V.A. Levanos, A.
Sahasrabudhe, and F.E. Dewhirst, (2001), Bacterial diversity in human subgingival
plaque, J Bacteriol, 183(12): p. 3770-3783.
89. Philp, D., A.L. Goldstein, and H.K. Kleinman, (2004), Thymosin 4 promotes
angiogenesis, wound healing, and hair follicle development, Mech Ageing Dev,
125(2): p. 113-115.
90. Philp, D., B. Scheremeta, K. Sibliss, M. Zhou, E.L. Fine, M. Nguyen, L. Wahl, M.P.
Hoffman, and H.K. Kleinman, (2006), Thymosin beta4 promotes matrix
metalloproteinase expression during wound repair, J Cell Physiol, 208(1): p. 195-
200.
91. Preshaw, P.M., (2009), Definitions of periodontal disease in research, J Clin
Periodontol, 36(1): p. 1-2.
92. Pschyrembel, Klinisches Wörterbuch, 261. Auflage, Walter de Gruyter Verlag Ber-
lin, 2007: p. 2121-2122.
93. Rebar, R.W., A. Miyake, T.L. Low, and A.L. Goldstein, (1981), Thymosin
stimulates secretion of luteinizing hormone-releasing factor, Science, 214(4521): p.
669-671.

118
94. Reti, R., E. Kwon, P. Qiu, M. Wheater, and G. Sosne, (2008), Thymosin beta4 is
cytoprotective in human gingival fibroblasts, Eur J Oral Sci, 116(5): p. 424-430.
95. Ribatti, D., E. Crivellato, and A. Vacca, (2006), Miller's seminal studies on the role
of thymus in immunity, Clin Exp Immunol, 144(3): p. 371-375.
96. Rohen, J.W., Lütjen-Drecoll, E., Funktionelle Anatomie des Menschen, 11. Auflage,
Schattauer Verlag Stuttgart, 2006: p. 147-148.
97. Rohen, J.W., Lütjen-Drecoll, E., Funktionelle Anatomie des Menschen, 11. Auflage,
Schattauer Verlag Stuttgart, 2006: p. 112-113.
98. Rohen, J.W., Lütjen-Drecoll, E., Funktionelle Anatomie des Menschen, 11. Auflage,
Schattauer Verlag Stuttgart, 2006: p. 98-99.
99. Ryan, M.E., S. Ramamurthy, and L.M. Golub, (1996), Matrix metalloproteinases
and their inhibition in periodontal treatment, Curr Opin Periodontol, 3: p. 85-96.
100. Safer, D., M. Elzinga, and V.T. Nachmias, (1991), Thymosin beta 4 and Fx, an
actin-sequestering peptide, are indistinguishable, J Biol Chem, 266(7): p. 4029-
4032.
101. Safer, D., R. Golla, and V.T. Nachmias, (1990), Isolation of a 5-kilodalton actin-
sequestering peptide from human blood platelets, Proc Natl Acad Sci U S A, 87(7):
p. 2536-2540.
102. Schenkein, H., (1999), The pathogenesis of periodontal diseases, J Periodontol,
70(4): p. 457-470.
103. Schlesinger, D.H. and G. Goldstein, (1975), Molecular conservation of 74 amino
acid sequence of ubiquitin between cattle and man, Nature, 255(5507): p. 42304.
104. Shannon, I.L., (1966), Climatological effects on human parotid gland function, Arch
Oral Biol, 11(4): p. 451-453.
105. Shannon, R.L., R.P. Feller, and R.P. Suddick, (1972), Effects of body position, blood
pressure and sleep on human parotid gland function, Ann Dent, 31(4): p. 81-86.
106. Shirodaria, S., J. Smith, I.J. McKay, C.N. Kennett, and F.J. Hughes, (2000),
Polymorphisms in the IL-1A gene are correlated with levels of interleukin-1alpha
protein in gingival crevicular fluid of teeth with severe periodontal disease, J Dent
Res, 79(11): p. 1864-1869.
107. Smith, Q.T. and S.J. Geegan, (1991), Repeated measurement of crevicular fluid
parameters at different sites, J Clin Periodontol, 18(3): p. 171-176.
108. Socransky, S.S. and A.D. Haffajee, (1992), The bacterial etiology of destructive
periodontal disease: current concepts, J Periodontol, 63(4 Suppl): p. 322-331.
109. Sosne, G., C.C. Chan, K. Thai, M. Kennedy, E.A. Szliter, L.D. Hazlett, and H.K.
Kleinman, (2001), Thymosin 4 promotes corneal wound healing and modulates
inflammatory mediators in vivo, Exp Eye Res, 72(5): p. 605-608.

119
110. Sosne, G., E.A. Szliter, R. Barrett, K.A. Kernacki, H. Kleinman, and L.D. Hazlett,
(2002), Thymosin beta 4 promotes corneal wound healing and decreases
inflammation in vivo following alkali injury, Exp Eye Res, 74(2): p. 293-299.
111. Srivastava, D., A. Saxena, J. Michael Dimaio, and I. Bock-Marquette, (2007),
Thymosin beta4 is cardioprotective after myocardial infarction, Ann N Y Acad Sci,
1112: p. 161-170.
112. Stein, S. and J. Moschera, (1981), High-performance liquid chromatography and
picomole-level detection of peptides and proteins, Methods Enzymol, 79(Pt B): p. 7-
16.
113. Theriot, J.A. and T.J. Mitchison, (1991), Actin microfilament dynamics in
locomoting cells, Nature, 352(6331): p. 126-131.
114. Thews, G., Vaupel, P., Vegetative Physiologie, 5. Auflage, Springer Medizin Verlag
Heidelberg, 2005: p. 381-384.
115. Thurman, G.B., C. Seals, T.L. Low, and A.L. Goldstein, (1984), Restorative effects
of thymosin polypeptides on purified protein derivative--dependent migration
inhibition factor production by the peripheral blood lymphocytes of adult
thymectomized guinea pigs, J Biol Response Mod, 3(2): p. 160-173.
116. Udenfriend, S., S. Stein, P. Bohlen, W. Dairman, W. Leimgruber, and M. Weigele,
(1972), Fluorescamine: a reagent for assay of amino acids, peptides, proteins, and
primary amines in the picomole range, Science, 178(63): p. 871-872.
117. Uitto, V.J., (2003), Gingival crevice fluid--an introduction, Periodontol 2000, 31: p.
9-11.
118. Ulfig, N., Kurzlehrbuch Histologie, 2. Auflage, Georg Thieme Verlag Suttgart,
2005: p. 129-131.
119. Vancompernolle, K., J. Vandekerckhove, M.R. Bubb, and E.D. Korn, (1991), The
interfaces of actin and Acanthamoeba actobindin. Identification of a new actin-
binding motif, J Biol Chem, 266(23): p. 15427-15431.
120. Watts, J.D., P.D. Cary, P. Sautiere, and C. Crane-Robinson, (1990), Thymosins: both
nuclear and cytoplasmic proteins, Eur J Biochem, 192(3): p. 643-651.
121. Weber, A., V.T. Nachmias, C.R. Pennise, M. Pring, and D. Safer, (1992),
Interaction of thymosin beta 4 with muscle and platelet actin: implications for actin
sequestration in resting platelets, Biochemistry, 31(27): p. 6179-6185.
122. Weterman, M.A., G.N. van Muijen, D.J. Ruiter, and H.P. Bloemers, (1993),
Thymosin beta-10 expression in melanoma cell lines and melanocytic lesions: a new
progression marker for human cutaneous melanoma, Int J Cancer, 53(2): p. 278-
284.
123. Wodnar-Filipowicz, A., U. Gubler, Y. Furuichi, M. Richardson, E.F. Nowoswiat,
M.S. Poonian, and B.L. Horecker, (1984), Cloning and sequence analysis of cDNA
for rat spleen thymosin beta 4, Proc Natl Acad Sci U S A, 81(8): p. 2295-2297.

120
124. Young, J.D., A.J. Lawrence, A.G. MacLean, B.P. Leung, I.B. McInnes, B. Canas,
D.J. Pappin, and R.D. Stevenson, (1999), Thymosin beta 4 sulfoxide is an anti-
inflammatory agent generated by monocytes in the presence of glucocorticoids, Nat
Med, 5(12): p. 1424-1427.
125. Zarbock, J., H. Oschkinat, E. Hannappel, H. Kalbacher, W. Voelter, and T.A. Holak,
(1990), Solution conformation of thymosin beta 4: a nuclear magnetic resonance
and simulated annealing study, Biochemistry, 29(34): p. 7814-7821.
126. Zoubek, R.E. and E. Hannappel, (2007), Subcellular distribution of thymosin beta4,
Ann N Y Acad Sci, 1112: p. 442-450.

121
Abkürzungsverzeichnis
AAP American Academy of Periodontology
ACN Acetonitril
API Approximaler Plaque Index
c Konzentration
°C Grad Celsius
C-Terminus Carboxy-Terminus
d Schichtdicke
Da Dalton
DNase Desoxyribonuklease
ε Extinktionskoeffizient
E Extinktion
ELISA Enzyme Linked Immunoabsorbent Assay
F-Aktin Filamentäres Aktin
fg Femtogramm
g Erdbeschleunigung
g Gramm
G-Aktin Globuläres Aktin
gl. Glandula
gll. Glandulae
GuaHCL Guanidiniumhydrochlorid
h Stunde
HPLC-ESI-MS High Pressure Liquid Chromatography - Electrospray-Ionisation -
Mass Spectrometry
Ig A Immunglobulin A
IFN Interferon
IL Interleukin
Kd Dissoziationskonstante
kDa Kilodalton
kV Kilovolt
λ Wellenlänge

122
l Liter
M Mol pro Liter
MALDI-TOF Matrix-Assisted Laser Desorption/Ionization - Time Of Flight Mass
Spectrometry
MAPK Mitogen-Activated Protein Kinase
mg Milligramm
MHC Major Histocompatibility Complex
min Minute
mM Millimolar
mm Millimeter
MMP Matrixmetalloproteinase
mRNA Messenger Ribonucleic Acid
NFkB Nuclear Factor kappa-light-chain-enhancer of activated B-cells
ng Nanogramm
nM Nanomolar
nm Nanometer
NMR Nuclear Magnetic Resonance
N-Terminus Amino-Terminus
P Proband
PA Parodontopathie
PCA Perchlorsäure
pH Pondus Hydrogenii
pI Isoelektrischer Punkt
pM Pikomolar
RP-HPLC Reverse-Phase High Pressure Liquid Chromatography
SBI Sulkus Blutungs Index
SF Sulkusfluid
SFFR Sulcus Fluid Flow Rate
Tβ4 Thymosin β4
TD Thiodiglycol
TFA Trifluoressigsäure

123
TT Taschentiefe
µg Mikrogramm
µl Mikroliter
UV Ultraviolettes Licht
VEGF Vascular Endothelian Growth Factor

124
Abbildungsverzeichnis
Abbildung 1: Reinigungsschritte zur Herstellung der Thymosin Fraktion 5 (nach
Goldstein, 2007) ............................................................................................. 6
Abbildung 2: Isoelektrische Fokussierung und Nomenklatur der Thymosine
(nach Goldstein, 2007) ................................................................................... 7
Abbildung 3: Biologische Eigenschaften des Thymosin α1 (nach Goldstein, 2007) ........... 8
Abbildung 4: Aminosäuresequenzen einzelner β-Thymosine (nach Huff, 2001) .............. 10
Abbildung 5: Struktur des Thymosin β4 (nach Hannappel, 2007) ..................................... 11
Abbildung 6: Thymosin β4 als Haupt-Aktin-sequestrierendes Protein (nach Huff, 2001) 13
Abbildung 7: Biologische Eigenschaften des Thymosin β4 (nach Goldstein, 2007) ......... 15
Abbildung 8: Gliederung des Kopfdarmes (nach Rohen, 2006) ........................................ 17
Abbildung 9: Topografie des Drüsenapparates (nach Rohen, 2006) .................................. 19
Abbildung 10: Ionenaustauschprozesse in der Azinuszelle (A) und in der Gangepithel-
zelle (B) (nach Thews, 2005) ....................................................................... 20
Abbildung 11: Elektrolytkonzentration des Speichels als Funktion des Speichel-
flusses (nach Thews, 2005) .......................................................................... 21
Abbildung 12: Aufbau des Parodonts (nach Hellwig, 2007) ................................................ 24
Abbildung 13: Formen des Gingivaepithels (nach Hellwig, 2007) ...................................... 25
Abbildung 14: Mikrobielle Komplexe der Plaque (nach Socransky, 1992) ......................... 33
Abbildung 15: Zytokinwirkungen im Parodont (nach Bartold, 2006) ................................. 35
Abbildung 16: Histologie der gesunden Gingiva (a) und der initialen Läsion (b) (nach
Hellwig, 2007) .............................................................................................. 37
Abbildung 17: Histologie der frühen Läsion (c) (nach Hellwig, 2007) ............................... 38
Abbildung 18: Histologie der etablierten Läsion (d) (nach Hellwig, 2007) ......................... 38
Abbildung 19: Histologie der fortgeschrittenen Läsion (e) (nach Hellwig, 2007) ............... 39
Abbildung 20: Aufbau eines Fotometers .............................................................................. 51
Abbildung 21: Reaktionsmechanismus von Fluram ............................................................. 54
Abbildung 22: Aufbau des RP-HPLC-Systems ................................................................... 55
Abbildung 23: RP-HPLC-Chromatogramme: Tβ4/Tβ10 Standard (A) und Sulkus-
fluidprobe (B) ............................................................................................... 57
Abbildung 24: Versuchsaufbau zur Messung der Stabilität von Tβ4 im Gesamtspeichel ... 59
Abbildung 25: RP-HPLC-Chromatogramme: 25 µl Gesamtspeichel und 200 ng
Tβ4 Standard ................................................................................................ 71
Abbildung 26: RP-HPLC-Chromatogramme: 25 µl Speichel gl. parotis und 200 ng
Tβ4 Standard ................................................................................................ 72
Abbildung 27: RP-HPLC-Chromatogramme: 300 µl Elutionslösung Sulkusfluid und
200 ng Tβ4 Standard .................................................................................... 73
Abbildung 28: Massenspektrometrie-Analyse: Schweineleukozytenprobe (A) und
Gesamtspeichelprobe (B) ............................................................................. 92

125
Tabellenverzeichnis
Tabelle 1: Spezies-spezifisches Vorkommen der β-Thymosine (nach Huff, 2001) ........... 12
Tabelle 2: Thymosin β4 Konzentrationen im Blut (nach Hannappel, 2007) ...................... 16
Tabelle 3: Speichelfunktionen und Speichelbestandteile (nach Llena-Puy, 2006) ............ 23
Tabelle 4: AAP Nomenklatur der Parodontopathien (nach Armitage, 1999)..................... 31
Tabelle 5: Einteilungskriterien der untersuchten Gruppen Teil 1 ...................................... 42
Tabelle 6: Einteilungskriterien der untersuchten Gruppen Teil 2 ...................................... 43
Tabelle 7: Gruppen-spezifisch analysierte Parameter ........................................................ 43
Tabelle 8: Korrelation API Werte und Mundhygienezustand ............................................ 49
Tabelle 9: Korrelation SBI Werte und Entzündungszustand .............................................. 49
Tabelle 10: Gradient der RP-HPLC ..................................................................................... 55
Tabelle 11: Zusammensetzung Gruppe A ............................................................................ 60
Tabelle 12: Zusammensetzung Gruppe B ............................................................................ 60
Tabelle 13: Zusammensetzung Gruppe C ............................................................................ 70
Tabelle 14: Messung der Taschentiefen (Gruppe C) ............................................................ 77
Tabelle 15: Zusammenfassung der Einflussfaktoren für die Tβ4 Konzentration im
Sulkusfluid (Gruppe C) ..................................................................................... 80
Tabelle 16: Zusammensetzung Gruppe D ............................................................................ 81
Tabelle 17: Messung der Taschentiefen (Gruppe D) ............................................................ 84
Tabelle 18: Zusammenfassung der Einflussfaktoren für die Tβ4 Konzentration im
Sulkusfluid (Gruppe D) ..................................................................................... 86
Tabelle 19: Zusammensetzung Gruppe E ............................................................................. 87
Tabelle 20: Methoden zur Elution von Tβ4 aus Papierspitzen Teil 1 .................................. 90
Tabelle 21: Methoden zur Elution von Tβ4 aus Papierspitzen Teil 2 .................................. 91
Tabelle 22: Tβ4 Konzentration im Gesamtspeichel – Vergleich Gruppe A (n = 7)
und B (n = 8) ..................................................................................................... 95
Tabelle 23: Proteinkonzentration im Gesamtspeichel – Vergleich Gruppe A (n = 7)
und B (n = 8) ..................................................................................................... 96
Tabelle 24: Taschentiefe, Sulkusfluidvolumen und Tβ4 Konzentration im Sulkusfluid –
Vergleich Gruppe C (n = 11) und D (n = 8) .................................................... 103

126
Diagrammverzeichnis
Diagramm 1: Proteinkonzentration der verschiedenen Gewebeflüssigkeiten (nach
Griffiths, 2003) ............................................................................................. 27
Diagramm 2: SFFR in Abhängigkeit vom Parodontalstatus (nach Goodson, 2003) .......... 28
Diagramm 3: Tβ4 Konzentration im Gesamtspeichel (Gruppe A) ..................................... 61
Diagramm 4: Veränderung der Tβ4 Konzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe A) .......................................................... 63
Diagramm 5: Tβ4 Konzentration im Gesamtspeichel (Gruppe B) ..................................... 64
Diagramm 6: Veränderung der Tβ4 Konzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe B) ........................................................... 65
Diagramm 7: Proteinkonzentration im Gesamtspeichel (Gruppe A) ................................. 66
Diagramm 8: Veränderung der Proteinkonzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe A) .......................................................... 67
Diagramm 9: Proteinkonzentration im Gesamtspeichel (Gruppe B) .................................. 68
Diagramm 10: Veränderung der Proteinkonzentration im Gesamtspeichel bei einer 2.
Probennahme am Tag 14 (Gruppe B) ........................................................... 69
Diagramm 11: Tβ4 Konzentration im Gesamtspeichel (Gruppe C) ..................................... 71
Diagramm 12: Sulkusfluidvolumen (Gruppe C) .................................................................. 75
Diagramm 13: Tβ4 Konzentration im Sulkusfluid (Gruppe C) ............................................ 76
Diagramm 14: API und SBI (Gruppe C) .............................................................................. 78
Diagramm 15: Sulkusfluidvolumen (Gruppe D) .................................................................. 82
Diagramm 16: Tβ4 Konzentration im Sulkusfluid (Gruppe D) ........................................... 83
Diagramm 17: API und SBI (Gruppe D) .............................................................................. 85
Diagramm 18: Sulkusfluidvolumen (Gruppe E) .................................................................. 88
Diagramm 19: Tβ4 Konzentration im Sulkusfluid (Gruppe E) ............................................ 89
Diagramm 20: Stabilität von Tβ4 im Gesamtspeichel (Proband P1) ................................... 93
Diagramm 21: Stabilität von Tβ4 im Gesamtspeichel (Proband P2) ................................... 94
Diagramm 22: Korrelation Proteinkonzentration und Tβ4 Konzentration im Gesamt-
speichel (Gruppe A) ..................................................................................... 98
Diagramm 23: Korrelation Proteinkonzentration und Tβ4 Konzentration im Gesamt-
speichel (Gruppe B) ..................................................................................... 98
Diagramm 24: Korrelation Tβ4 Konzentration im Sulkusfluid und im Gesamtspeichel
(Gruppe C).................................................................................................. 100
Diagramm 25: Korrelation Sulkusfluidvolumen und Tβ4 Konzentration im Sulkus-
fluid (Gruppe C) ......................................................................................... 105
Diagramm 26: Korrelation Sulkusfluidvolumen und Tβ4 Konzentration im Sulkus-
fluid (Gruppe D) ......................................................................................... 106

127
Danksagung
Mein besonderer Dank gilt Herrn Prof. Dr. Ewald Hannappel für die Überlassung des
Themas sowie für seine ständige Betreuung und Förderung dieser Arbeit.
Herrn Prof. Dr. med. dent. Matthias Pelka danke ich für die intensive fachliche Beratung zu
zahnmedizinischen Aspekten sowie für seine aufmerksame Begleitung der Arbeit. Allen
Mitarbeitern des Instituts für Biochemie danke ich für ihre allzeit freundliche Unterstützung
bei der Durchführung der experimentellen Arbeiten. Dank geht nicht zuletzt auch an alle
diejenigen Personen ohne deren stetige Motivation und Anteilnahme diese Arbeit nicht
zustande gekommen wäre. Insbesondere Herrn Prof. em. Dr. Werner Loewe sowie meiner
Familie möchte ich in diesem Zusammenhang meinen Dank aussprechen.

128
Lebenslauf
Name: Moritz Lutz Zimmermann
Anschrift: Eichenstr. 19
91217 Hersbruck
E-Mail: [email protected]
Geboren am: 08.06.1986 in Sulzbach-Rosenberg
Eltern: Dr. Hans-Peter Zimmermann Zahnarzt
Heidrun Zimmermann Realschullehrerin
Geschwister: Philipp Zimmermann, geboren am 08.06.1986
Schulbildung: September 1992 – Juli 1996 Grundschule Altensittenbach
September 1996 – Juni 2005 Paul-Pfinzing Gymnasium Hersbruck
Studium Oktober 2005 – Dezember 2010 Zahnmedizinstudium an der Universität
Erlangen-Nürnberg
Abiturfächer: Leistungskurse: Mathematik, Französisch
Grundkurs: Latein
Colloquium: Geschichte
Facharbeit: Les relations franco-allemands face à la guerre en Irak.
Sprachen: Englisch 7 Jahre auf dem Paul-Pfinzing Gymnasium Hersbruck
Latein 7 Jahre auf dem Paul-Pfinzing Gymnasium Hersbruck
Französisch 5 Jahre auf dem Paul-Pfinzing Gymnasium Hersbruck
Spanisch 2 Jahre auf dem Paul-Pfinzing Gymnasium Hersbruck
Italienisch 1,5 Jahre an der Sprachenschule der Universität Erlangen-
Nürnberg
Sonstiges: 2000 Praktikum in der Bayerischen Landesbank in Nürnberg
2001 Teilnahme am deutsch-französischen Schüleraustausch in Les Sables
d´Olonne
2001 und 2002 Teilnahme am Planspiel Börse
2003 Teilnahme am deutsch-englischen Schüleraustausch in Exeter
2004 Teilnahme am Projekt Business@school der Boston Consulting Group
2005 Teilnahme am Auswahlverfahren der Studienstiftung des deutschen
Volkes
2009 Teilnahme am Auswahlverfahren der Bayerischen EliteAkademie


















