Syndecan-4 inhibits Wnt/β-catenin signaling through regulation of low-density-lipoprotein...
10
The International Journal of Biochemistry & Cell Biology 46 (2014) 103–112 Contents lists available at ScienceDirect The International Journal of Biochemistry & Cell Biology journal h om epage: www.elsevier.com/locate/biocel Syndecan-4 inhibits Wnt/-catenin signaling through regulation of low-density-lipoprotein receptor-related protein (LRP6) and R-spondin 3 Pablo Astudillo, Héctor Carrasco, Juan Larraín ∗ Center for Aging and Regeneration, Millennium Nucleus in Regenerative Biology, Faculty of Biological Sciences, P. Universidad Católica de Chile, Alameda 340, Santiago, Chile a r t i c l e i n f o Article history: Received 27 March 2013 Received in revised form 18 October 2013 Accepted 6 November 2013 Available online 23 November 2013 Keywords: Wnt Syndecan-4 LRP6 Fibronectin R-spondin a b s t r a c t Regulation of Wnt signaling is crucial for embryonic development and adult homeostasis. Here we study the role of Syndecan-4 (SDC4), a cell-surface heparan sulphate proteoglycan, and Fibronectin (FN), in Wnt/-catenin signaling. Gain- and loss-of-function experiments in mammalian cell lines and Xenopus embryos demonstrate that SDC4 and FN inhibit Wnt/-catenin signaling. Epistatic and biochemical exper- iments show that this inhibition occurs at the cell membrane level through regulation of LRP6. R-spondin 3, a ligand that promotes canonical and non-canonical Wnt signaling, is more prone to potentiate Wnt/- catenin signaling when SDC4 levels are reduced, suggesting a model whereby SDC4 tunes the ability of R-spondin to modulate the different Wnt signaling pathways. Since SDC4 has been previously related to non-canonical Wnt signaling, our results also suggest that this proteoglycan can be a key component in the regulation of Wnt signaling. © 2013 Elsevier Ltd. All rights reserved. 1. Introduction The Wnt (Wingless, Int1) family constitutes a large number of cysteine-rich secreted glycoproteins that regulate a variety of cel- lular processes such as development, homeostasis, regeneration, stem cell pluripotency, cell polarity and migration, through activa- tion of signaling pathways (Clevers and Nusse, 2012; Niehrs, 2012). One of these signaling pathways, known as the Wnt/-catenin (or ‘canonical’) pathway, is triggered through binding of Wnt ligands to Frizzled (FZ) and LRP5/6 receptors, which activate intracellular cascades leading to the stabilization and nuclear translocation of - catenin. Extracellular and intracellular molecules tightly regulate this signaling pathway. Among those are R-spondins (roof plate- specific spondin, RSPO), a family of secreted glycoproteins that bind to its receptors LGR4/5 (orphan leucine-rich repeat contain- ing G-protein-coupled receptor 4/5) and augments Wnt/-catenin signaling through regulation of FZ and LRP5/6 cell-surface levels (Koo et al., 2012; Hao et al., 2012; Glinka et al., 2011; de Lau et al., 2011). Syndecan-4 (SDC4; Xsdc4 in Xenopus) is a novel regulator of the non-canonical (or ‘Wnt/-catenin independent’) Wnt pathway ∗ Corresponding author at: Faculty of Biological Sciences, Pontificia Universidad Católica de Chile, Alameda 340, Santiago, Postal Code 8331150, Chile. Tel.: +56 2 686 2129; fax: +56 2 6862824. E-mail address: [email protected] (J. Larraín). during Xenopus laevis development (Mu˜ noz et al., 2006; Matthews et al., 2008; Ohkawara et al., 2011). SDC4 is a member of the Syndecan family of heparan sulphate proteoglycans (HSPGs), char- acterized by its ubiquitous expression and initially described as a component of focal adhesions because of its ability to bind Fibronectin (FN) (Woods and Couchman, 1994; Couchman, 2010). It has been demonstrated that R-spondin 3 (RSPO3) binds SDC4 and induces its clathrin-mediated endocytosis, providing a molec- ular mechanism explaining how this HSPG regulates non-canonical Wnt signaling (Ohkawara et al., 2011). In mice, SDC4 has also been involved in the regulation of the expansion of satellite stem cells, in a process dependent on FN, Frizzled-7 and Wnt7a through the non-canonical pathway in the murine muscle fiber (Bentzinger et al., 2013). More recently, we showed that Sdc4 interacts geneti- cally with Vangl2 (Strabismus), a co-receptor for the non-canonical Wnt pathway, to regulate neural tube closure in mice (Escobedo et al., 2013). Furthermore, close examination of the Sdc4 knockout mice reveals defects in the orientation of stereociliary bundles in the mechanosensory hair cells in the inner ear (Escobedo et al., 2013), a hallmark phenotype of disrupted non-canonical Wnt signaling (Montcouquiol et al., 2006). These results indicate that the regulation of the non-canonical Wnt signaling by SDC4 is a con- served feature of this HSPG. Impairment of SDC4 function has been recently associated with defects in reparative processes, such as muscle fiber repair (Bentzinger et al., 2013), differentiation of car- diac myofibroblasts after mechanical stress (Herum et al., 2013) and in bone repair after fracture (Bertrand et al., 2013), highlighting the 1357-2725/$ – see front matter © 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.biocel.2013.11.012
Transcript of Syndecan-4 inhibits Wnt/β-catenin signaling through regulation of low-density-lipoprotein...

SlR
PC3
a
ARRAA
KWSLFR
1
clstO‘tcctsbis(2
t
C2
1h
The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112
Contents lists available at ScienceDirect
The International Journal of Biochemistry& Cell Biology
journa l h om epage: www.elsev ier .com/ locate /b ioce l
yndecan-4 inhibits Wnt/�-catenin signaling through regulation ofow-density-lipoprotein receptor-related protein (LRP6) and-spondin 3
ablo Astudillo, Héctor Carrasco, Juan Larraín ∗
enter for Aging and Regeneration, Millennium Nucleus in Regenerative Biology, Faculty of Biological Sciences, P. Universidad Católica de Chile, Alameda40, Santiago, Chile
r t i c l e i n f o
rticle history:eceived 27 March 2013eceived in revised form 18 October 2013ccepted 6 November 2013vailable online 23 November 2013
a b s t r a c t
Regulation of Wnt signaling is crucial for embryonic development and adult homeostasis. Here we studythe role of Syndecan-4 (SDC4), a cell-surface heparan sulphate proteoglycan, and Fibronectin (FN), inWnt/�-catenin signaling. Gain- and loss-of-function experiments in mammalian cell lines and Xenopusembryos demonstrate that SDC4 and FN inhibit Wnt/�-catenin signaling. Epistatic and biochemical exper-iments show that this inhibition occurs at the cell membrane level through regulation of LRP6. R-spondin
eywords:nt
yndecan-4RP6
3, a ligand that promotes canonical and non-canonical Wnt signaling, is more prone to potentiate Wnt/�-catenin signaling when SDC4 levels are reduced, suggesting a model whereby SDC4 tunes the ability ofR-spondin to modulate the different Wnt signaling pathways. Since SDC4 has been previously related tonon-canonical Wnt signaling, our results also suggest that this proteoglycan can be a key component inthe regulation of Wnt signaling.
ibronectin-spondin
. Introduction
The Wnt (Wingless, Int1) family constitutes a large number ofysteine-rich secreted glycoproteins that regulate a variety of cel-ular processes such as development, homeostasis, regeneration,tem cell pluripotency, cell polarity and migration, through activa-ion of signaling pathways (Clevers and Nusse, 2012; Niehrs, 2012).ne of these signaling pathways, known as the Wnt/�-catenin (or
canonical’) pathway, is triggered through binding of Wnt ligandso Frizzled (FZ) and LRP5/6 receptors, which activate intracellularascades leading to the stabilization and nuclear translocation of �-atenin. Extracellular and intracellular molecules tightly regulatehis signaling pathway. Among those are R-spondins (roof plate-pecific spondin, RSPO), a family of secreted glycoproteins thatind to its receptors LGR4/5 (orphan leucine-rich repeat contain-
ng G-protein-coupled receptor 4/5) and augments Wnt/�-cateninignaling through regulation of FZ and LRP5/6 cell-surface levelsKoo et al., 2012; Hao et al., 2012; Glinka et al., 2011; de Lau et al.,
011).Syndecan-4 (SDC4; Xsdc4 in Xenopus) is a novel regulator ofhe non-canonical (or ‘Wnt/�-catenin independent’) Wnt pathway
∗ Corresponding author at: Faculty of Biological Sciences, Pontificia Universidadatólica de Chile, Alameda 340, Santiago, Postal Code 8331150, Chile. Tel.: +56 2 686129; fax: +56 2 6862824.
E-mail address: [email protected] (J. Larraín).
357-2725/$ – see front matter © 2013 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.biocel.2013.11.012
© 2013 Elsevier Ltd. All rights reserved.
during Xenopus laevis development (Munoz et al., 2006; Matthewset al., 2008; Ohkawara et al., 2011). SDC4 is a member of theSyndecan family of heparan sulphate proteoglycans (HSPGs), char-acterized by its ubiquitous expression and initially described asa component of focal adhesions because of its ability to bindFibronectin (FN) (Woods and Couchman, 1994; Couchman, 2010).It has been demonstrated that R-spondin 3 (RSPO3) binds SDC4and induces its clathrin-mediated endocytosis, providing a molec-ular mechanism explaining how this HSPG regulates non-canonicalWnt signaling (Ohkawara et al., 2011). In mice, SDC4 has also beeninvolved in the regulation of the expansion of satellite stem cells,in a process dependent on FN, Frizzled-7 and Wnt7a through thenon-canonical pathway in the murine muscle fiber (Bentzingeret al., 2013). More recently, we showed that Sdc4 interacts geneti-cally with Vangl2 (Strabismus), a co-receptor for the non-canonicalWnt pathway, to regulate neural tube closure in mice (Escobedoet al., 2013). Furthermore, close examination of the Sdc4 knockoutmice reveals defects in the orientation of stereociliary bundles inthe mechanosensory hair cells in the inner ear (Escobedo et al.,2013), a hallmark phenotype of disrupted non-canonical Wntsignaling (Montcouquiol et al., 2006). These results indicate thatthe regulation of the non-canonical Wnt signaling by SDC4 is a con-served feature of this HSPG. Impairment of SDC4 function has been
recently associated with defects in reparative processes, such asmuscle fiber repair (Bentzinger et al., 2013), differentiation of car-diac myofibroblasts after mechanical stress (Herum et al., 2013) andin bone repair after fracture (Bertrand et al., 2013), highlighting the
1 of Bioc
pt
phWtWdHwapoa
2
2
(pw((vwNcTX2Btma�iasarS
2
mspamgpc(
(e1tc
c
04 P. Astudillo et al. / The International Journal
hysiological relevance of SDC4 and the interest in understandinghe signaling pathways regulated by this proteoglycan.
The canonical and non-canonical Wnt pathways share com-onents and receptors, and cross-talk between both branchesas been reported. Whether SDC4 also regulates the canonicalnt pathway, has not been addressed. Furthermore, considering
hat RSPO3 regulates canonical (Wnt/�-catenin) and non-canonicalnt pathways, and that SDC4 acts as a receptor for RSPO3, we
ecided to evaluate the role of SDC4 in the Wnt/�-catenin pathway.ere we show that SDC4 and FN inhibit the Wnt/�-catenin path-ay in several models tested. This effect is exerted at the LRP6 level,
nd reduction of SDC4 levels results in increased LRP6 activity andhosphorylation. Furthermore, SDC4 seems to restrict the activityf RSPO3, since depletion of this HSPG increases Wnt/�-cateninctivity induced by RSPO3.
. Materials and methods
.1. Constructs and reagents
HEK293 cells (CRL-1573), parental mouse intestinal L-CellsCRL-2648) and their derivative, L-Cells Wnt3a (CRL-2647), wereurchased from the American Type Culture Collection. HeLa cellsere provided by Dr. A. González, and mouse embryonic fibroblasts
MEFs) were prepared as described below. Constitutive active LRP6caLRP6) and stabilized �-catenin (�-catenin*) were kindly pro-ided by Dr. E. de Robertis (Taelman et al., 2010). LRP6-FLAG, asell as �-Tp1479-LRP6 antibody, were kindly provided by Dr. C.iehrs (Davidson et al., 2005). FLAG/HA-tagged LGR4 and RSPO3onstructs were provided by Dr. H. Clevers (de Lau et al., 2011).he �HS antibody was provided by Dr. E. Brandan, and FLAG-sdc4 was previously described (Munoz et al., 2006; Carvallo et al.,010). Small interfering RNAs were purchased from Santa Cruziotechnology. A detailed description of the corresponding vec-ors and siRNAs is provided in the Supplementary Material. Rabbit
onoclonal LRP6 (2560S) and rabbit polyclonal Sp1490 (2568S)ntibodies were purchased from Cell Signaling; Rabbit polyclonal-catenin (H-102) and mouse monoclonal SDC4 (5G9) antibod-
es were purchased from Santa Cruz Biotechnology. Fibronectinnd Tubulin antibodies were purchased from Sigma. Chinese Ham-ter Ovary (CHO)-derived recombinant human Wnt3a, as wells purified RSPO3, were purchased from R&D Systems. Othereagents, unless otherwise is indicated, were purchased fromigma.
.2. Cell culture, transfection and RNA isolation
HEK293, HeLa and MEFs cells were cultured in complete DMEMedium, consisting in DMEM supplemented with 10% fetal bovine
erum (HyClone), 2 mM l-glutamine (HyClone) and 100 units/mL ofenicillin/streptomycin (HyClone). Conditioned media from L-Cellsnd L-Cells-Wnt3a was produced by culture with complete DMEMedium supplemented with 25 mM of d-glucose (Life Technolo-
ies). L-Cells Wnt3a were maintained in complete DMEM mediumlus 25 mM d-glucose and 0.4 mg/mL of G418 for selection. HEK293ells were seeded in plates coated previously with poly-l-lysineSigma).
Mouse embryonic fibroblasts (MEFs) from Syndecan-4 wild-typeSdc4 +/+) and null (Sdc4 −/−) mice were prepared from E14.5mbryos and cultured in DMEM/F-12 medium supplemented with0% FBS, l-glutamine and antibiotics, as indicated above. For MEFs
ransfection, Lipofectamine LTX and Plus Reagent were used, inultures from passages two and three.Reporter assays were performed in MEFs, HEK293 and HeLaells 72 h after transfection. Briefly, HEK293, HeLa and L-Cells were
hemistry & Cell Biology 46 (2014) 103– 112
cultured and transfected using Lipofectamine 2000 as indicatedby the manufacturer (Life Technologies). Between 10 pmol (forTOPFlash assays) and 40 pmol (for Western Blots) of siRNA weretransfected, and knockdown of the protein of interest was eval-uated 48 h after transfection. For stimulation assays, conditionedmedium from parental L-Cells or Wnt-3a expressing L-Cells, orpurified Wnt3a or RSPO3 ligands, were used. When needed,Fibronectin from bovine plasma (Sigma) was diluted in PBS 1×to a final concentration of 10 �g/mL or 25 �g/mL, and cultureplates were incubated overnight before the experiments. HEK293,HeLa and MEFs cells were transfected as needed with 0.5–1 �g ofTOPFlash reporter to monitor Wnt/�-catenin signaling (Molenaaret al., 1996) and pRenilla-firefly (10 ng) luciferase as an internalcontrol. 24 h after transfection, cells were trypsinized and platedin 96-well plates. One day later, cells were stimulated as indi-cated in the text by 24 h, lysed with passive lysis buffer (PLB1×) and processed using the Dual Luciferase Assay (Promega).TOPFlash/pRenilla luciferase ratios (relative luciferase units, Rlu)were calculated for each assay. In HEK293 and HeLa cells, eachexperiment was performed at least n = 3. For MEFs, each experimentwas performed twice.
RNA was isolated from MEFs stimulated with L-Cell CM orWnt3a CM by 8 h using the Trizol reagent (Life Technologies), andcomplementary DNA was synthesized using standard procedures.
2.3. Western Blot
For endogenous LRP6 phosphorylation, membrane-enrichedfractions from MEFs were prepared as described (Davidson et al.,2005); for each treatment, three wells from 6-well plates were col-lected in 1.5 mL of total volume of lysis buffer and processed. MEFswere stimulated with Wnt3a conditioned medium for 30 min, andconditioned medium from parental L-Cells was used as a control.For evaluation of endogenous LRP6 levels in HEK293 cells, lysateswere prepared from cells plated in 6-well plates as described(Munoz et al., 2006; Carvallo et al., 2010). For �-catenin stabiliza-tion in L-Cells (Shibamoto et al., 1998), cells were stimulated for1 h with the indicated CMs and then lysed using a hypotonic buffer(5 mM Hepes pH 6.8; 1 mM MgCl2) supplemented with proteaseand phosphatase inhibitors (10 mM Na4O7P2·10H2O; 10 mM NaF;5 mM Na2VO4; 1 mM PMSF). For SDC4, cells were lysed as described(Munoz et al., 2006; Carvallo et al., 2010), and lysates were treatedovernight with 0.5 × 10−3 units of heparitinase from Flavobacteriumheparinum (US Biologicals, USA).
To evaluate the knockdown of SDC4 (Supplementary Fig. 1A andB) and FN (Supplementary Fig. 3A and B), HEK293 cells were trans-fected with 40 pmol of the indicated siRNAs, and the effect wasevaluated 48 h after the transfection. Densitometry analysis wasperformed using ImageJ (Abràmoff et al., 2004).
2.4. Microinjection and analysis of X. laevis embryos
All mRNAs were synthesized in vitro with SP6 polymerase usingthe Message Machine kit (Ambion Inc., Austin, TX), as previouslydescribed (Munoz et al., 2006; Carvallo et al., 2010). SyntheticmRNAs encoding xwnt8-HA (5–10 pg per embryo), FLAG-xsdc4(100–1000 pg per embryo), calrp6-GFP (100 pg per embryo) andˇ-catenin* (50 pg per embryo) were microinjected as indicatedin the text, together with the TOPFlash reporter DNA (60 pg perembryo) and Renilla luciferase (20 pg per embryo). The embryoswere microinjected in the animal pole, one time in each blas-tomere at the 8-cell stage, and were grown to stage 10.5, and
then processed for TOPFlash assay. Fifteen embryos per treatmentwere pooled in triplicates (n = 5 per pool), lysed in PLB 1× and ana-lyzed, and each experiment was performed in three independentexperiments. In all cases, representative experiments are shown.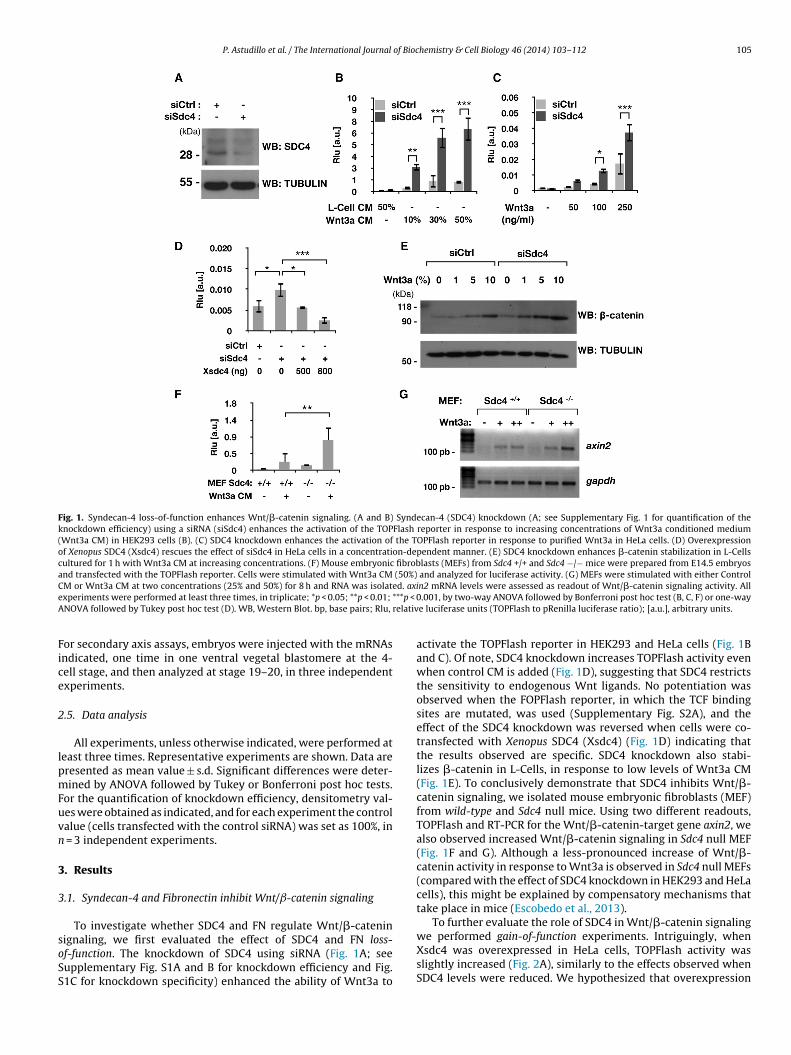
P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112 105
Fig. 1. Syndecan-4 loss-of-function enhances Wnt/�-catenin signaling. (A and B) Syndecan-4 (SDC4) knockdown (A; see Supplementary Fig. 1 for quantification of theknockdown efficiency) using a siRNA (siSdc4) enhances the activation of the TOPFlash reporter in response to increasing concentrations of Wnt3a conditioned medium(Wnt3a CM) in HEK293 cells (B). (C) SDC4 knockdown enhances the activation of the TOPFlash reporter in response to purified Wnt3a in HeLa cells. (D) Overexpressionof Xenopus SDC4 (Xsdc4) rescues the effect of siSdc4 in HeLa cells in a concentration-dependent manner. (E) SDC4 knockdown enhances �-catenin stabilization in L-Cellscultured for 1 h with Wnt3a CM at increasing concentrations. (F) Mouse embryonic fibroblasts (MEFs) from Sdc4 +/+ and Sdc4 −/− mice were prepared from E14.5 embryosand transfected with the TOPFlash reporter. Cells were stimulated with Wnt3a CM (50%) and analyzed for luciferase activity. (G) MEFs were stimulated with either ControlC ed. axie ***p < 0A relativ
Fice
2
lpmFuvn
3
3
soSS
M or Wnt3a CM at two concentrations (25% and 50%) for 8 h and RNA was isolatxperiments were performed at least three times, in triplicate; *p < 0.05; **p < 0.01;
NOVA followed by Tukey post hoc test (D). WB, Western Blot. bp, base pairs; Rlu,
or secondary axis assays, embryos were injected with the mRNAsndicated, one time in one ventral vegetal blastomere at the 4-ell stage, and then analyzed at stage 19–20, in three independentxperiments.
.5. Data analysis
All experiments, unless otherwise indicated, were performed ateast three times. Representative experiments are shown. Data areresented as mean value ± s.d. Significant differences were deter-ined by ANOVA followed by Tukey or Bonferroni post hoc tests.
or the quantification of knockdown efficiency, densitometry val-es were obtained as indicated, and for each experiment the controlalue (cells transfected with the control siRNA) was set as 100%, in
= 3 independent experiments.
. Results
.1. Syndecan-4 and Fibronectin inhibit Wnt/ˇ-catenin signaling
To investigate whether SDC4 and FN regulate Wnt/�-catenin
ignaling, we first evaluated the effect of SDC4 and FN loss-f-function. The knockdown of SDC4 using siRNA (Fig. 1A; seeupplementary Fig. S1A and B for knockdown efficiency and Fig.1C for knockdown specificity) enhanced the ability of Wnt3a ton2 mRNA levels were assessed as readout of Wnt/�-catenin signaling activity. All.001, by two-way ANOVA followed by Bonferroni post hoc test (B, C, F) or one-way
e luciferase units (TOPFlash to pRenilla luciferase ratio); [a.u.], arbitrary units.
activate the TOPFlash reporter in HEK293 and HeLa cells (Fig. 1Band C). Of note, SDC4 knockdown increases TOPFlash activity evenwhen control CM is added (Fig. 1D), suggesting that SDC4 restrictsthe sensitivity to endogenous Wnt ligands. No potentiation wasobserved when the FOPFlash reporter, in which the TCF bindingsites are mutated, was used (Supplementary Fig. S2A), and theeffect of the SDC4 knockdown was reversed when cells were co-transfected with Xenopus SDC4 (Xsdc4) (Fig. 1D) indicating thatthe results observed are specific. SDC4 knockdown also stabi-lizes �-catenin in L-Cells, in response to low levels of Wnt3a CM(Fig. 1E). To conclusively demonstrate that SDC4 inhibits Wnt/�-catenin signaling, we isolated mouse embryonic fibroblasts (MEF)from wild-type and Sdc4 null mice. Using two different readouts,TOPFlash and RT-PCR for the Wnt/�-catenin-target gene axin2, wealso observed increased Wnt/�-catenin signaling in Sdc4 null MEF(Fig. 1F and G). Although a less-pronounced increase of Wnt/�-catenin activity in response to Wnt3a is observed in Sdc4 null MEFs(compared with the effect of SDC4 knockdown in HEK293 and HeLacells), this might be explained by compensatory mechanisms thattake place in mice (Escobedo et al., 2013).
To further evaluate the role of SDC4 in Wnt/�-catenin signaling
we performed gain-of-function experiments. Intriguingly, whenXsdc4 was overexpressed in HeLa cells, TOPFlash activity wasslightly increased (Fig. 2A), similarly to the effects observed whenSDC4 levels were reduced. We hypothesized that overexpression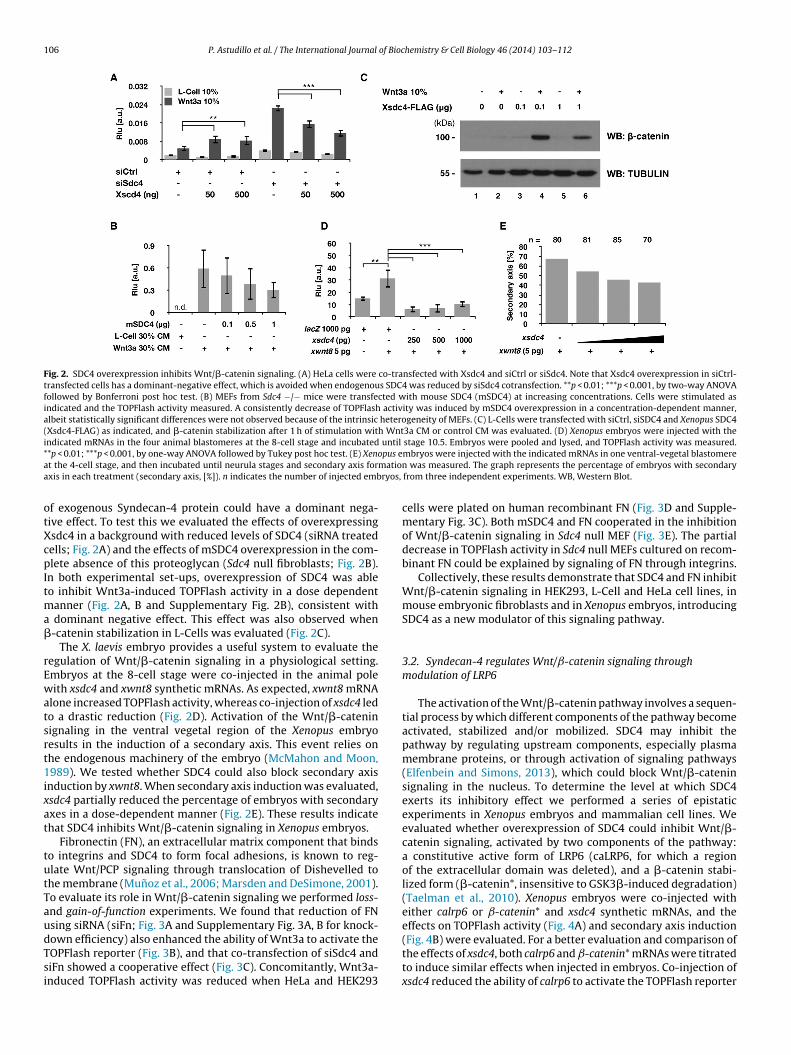
106 P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112
Fig. 2. SDC4 overexpression inhibits Wnt/�-catenin signaling. (A) HeLa cells were co-transfected with Xsdc4 and siCtrl or siSdc4. Note that Xsdc4 overexpression in siCtrl-transfected cells has a dominant-negative effect, which is avoided when endogenous SDC4 was reduced by siSdc4 cotransfection. **p < 0.01; ***p < 0.001, by two-way ANOVAfollowed by Bonferroni post hoc test. (B) MEFs from Sdc4 −/− mice were transfected with mouse SDC4 (mSDC4) at increasing concentrations. Cells were stimulated asindicated and the TOPFlash activity measured. A consistently decrease of TOPFlash activity was induced by mSDC4 overexpression in a concentration-dependent manner,albeit statistically significant differences were not observed because of the intrinsic heterogeneity of MEFs. (C) L-Cells were transfected with siCtrl, siSDC4 and Xenopus SDC4(Xsdc4-FLAG) as indicated, and �-catenin stabilization after 1 h of stimulation with Wnt3a CM or control CM was evaluated. (D) Xenopus embryos were injected with theindicated mRNAs in the four animal blastomeres at the 8-cell stage and incubated until stage 10.5. Embryos were pooled and lysed, and TOPFlash activity was measured.* pus ea matioa ryos,
otXcpItma�
rEwatsrt1ixat
tutTaudTsi
*p < 0.01; ***p < 0.001, by one-way ANOVA followed by Tukey post hoc test. (E) Xenot the 4-cell stage, and then incubated until neurula stages and secondary axis forxis in each treatment (secondary axis, [%]). n indicates the number of injected emb
f exogenous Syndecan-4 protein could have a dominant nega-ive effect. To test this we evaluated the effects of overexpressingsdc4 in a background with reduced levels of SDC4 (siRNA treatedells; Fig. 2A) and the effects of mSDC4 overexpression in the com-lete absence of this proteoglycan (Sdc4 null fibroblasts; Fig. 2B).
n both experimental set-ups, overexpression of SDC4 was ableo inhibit Wnt3a-induced TOPFlash activity in a dose dependent
anner (Fig. 2A, B and Supplementary Fig. 2B), consistent with dominant negative effect. This effect was also observed when-catenin stabilization in L-Cells was evaluated (Fig. 2C).
The X. laevis embryo provides a useful system to evaluate theegulation of Wnt/�-catenin signaling in a physiological setting.mbryos at the 8-cell stage were co-injected in the animal poleith xsdc4 and xwnt8 synthetic mRNAs. As expected, xwnt8 mRNA
lone increased TOPFlash activity, whereas co-injection of xsdc4 ledo a drastic reduction (Fig. 2D). Activation of the Wnt/�-cateninignaling in the ventral vegetal region of the Xenopus embryoesults in the induction of a secondary axis. This event relies onhe endogenous machinery of the embryo (McMahon and Moon,989). We tested whether SDC4 could also block secondary axis
nduction by xwnt8. When secondary axis induction was evaluated,sdc4 partially reduced the percentage of embryos with secondaryxes in a dose-dependent manner (Fig. 2E). These results indicatehat SDC4 inhibits Wnt/�-catenin signaling in Xenopus embryos.
Fibronectin (FN), an extracellular matrix component that bindso integrins and SDC4 to form focal adhesions, is known to reg-late Wnt/PCP signaling through translocation of Dishevelled tohe membrane (Munoz et al., 2006; Marsden and DeSimone, 2001).o evaluate its role in Wnt/�-catenin signaling we performed loss-nd gain-of-function experiments. We found that reduction of FNsing siRNA (siFn; Fig. 3A and Supplementary Fig. 3A, B for knock-
own efficiency) also enhanced the ability of Wnt3a to activate theOPFlash reporter (Fig. 3B), and that co-transfection of siSdc4 andiFn showed a cooperative effect (Fig. 3C). Concomitantly, Wnt3a-nduced TOPFlash activity was reduced when HeLa and HEK293mbryos were injected with the indicated mRNAs in one ventral-vegetal blastomeren was measured. The graph represents the percentage of embryos with secondaryfrom three independent experiments. WB, Western Blot.
cells were plated on human recombinant FN (Fig. 3D and Supple-mentary Fig. 3C). Both mSDC4 and FN cooperated in the inhibitionof Wnt/�-catenin signaling in Sdc4 null MEF (Fig. 3E). The partialdecrease in TOPFlash activity in Sdc4 null MEFs cultured on recom-binant FN could be explained by signaling of FN through integrins.
Collectively, these results demonstrate that SDC4 and FN inhibitWnt/�-catenin signaling in HEK293, L-Cell and HeLa cell lines, inmouse embryonic fibroblasts and in Xenopus embryos, introducingSDC4 as a new modulator of this signaling pathway.
3.2. Syndecan-4 regulates Wnt/ˇ-catenin signaling throughmodulation of LRP6
The activation of the Wnt/�-catenin pathway involves a sequen-tial process by which different components of the pathway becomeactivated, stabilized and/or mobilized. SDC4 may inhibit thepathway by regulating upstream components, especially plasmamembrane proteins, or through activation of signaling pathways(Elfenbein and Simons, 2013), which could block Wnt/�-cateninsignaling in the nucleus. To determine the level at which SDC4exerts its inhibitory effect we performed a series of epistaticexperiments in Xenopus embryos and mammalian cell lines. Weevaluated whether overexpression of SDC4 could inhibit Wnt/�-catenin signaling, activated by two components of the pathway:a constitutive active form of LRP6 (caLRP6, for which a regionof the extracellular domain was deleted), and a �-catenin stabi-lized form (�-catenin*, insensitive to GSK3�-induced degradation)(Taelman et al., 2010). Xenopus embryos were co-injected witheither calrp6 or ˇ-catenin* and xsdc4 synthetic mRNAs, and theeffects on TOPFlash activity (Fig. 4A) and secondary axis induction
(Fig. 4B) were evaluated. For a better evaluation and comparison ofthe effects of xsdc4, both calrp6 and ˇ-catenin* mRNAs were titratedto induce similar effects when injected in embryos. Co-injection ofxsdc4 reduced the ability of calrp6 to activate the TOPFlash reporter
P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112 107
Fig. 3. Fibronectin (FN) loss-of-function enhances Wnt/�-catenin signaling. (A and B) FN knockdown using siRNA (siFn; A) enhances TOPFlash activity in response to increasingconcentrations of Wnt3a CM (B). **p < 0.01, ***p < 0.001, by two-way ANOVA followed by Bonferroni post hoc test (B). (C) siFn and siSdc4 have a cooperative effect on TOPFlashactivity, compared with each siRNA transfected separately, in HEK293 cells. (D) Culture of cells upon recombinant FN decreases Wnt/�-catenin signaling; in HeLa cells in aconcentration-dependent manner. *p < 0.05, ***p < 0.001, by one-way ANOVA followed by Tukey post hoc test. (E) MEFs from Sdc4 −/− mice were stimulated with Wnt3a CMa ouse
t (bar
n
(c
acWlWtNl(Wrt(
c2toflt(2S
nd treated as indicated. Both culture of MEFs upon recombinant FN, or rescue of mreatments combined have a cooperative effect, further decreasing TOPFlash activityot detected. WB, Western Blot.
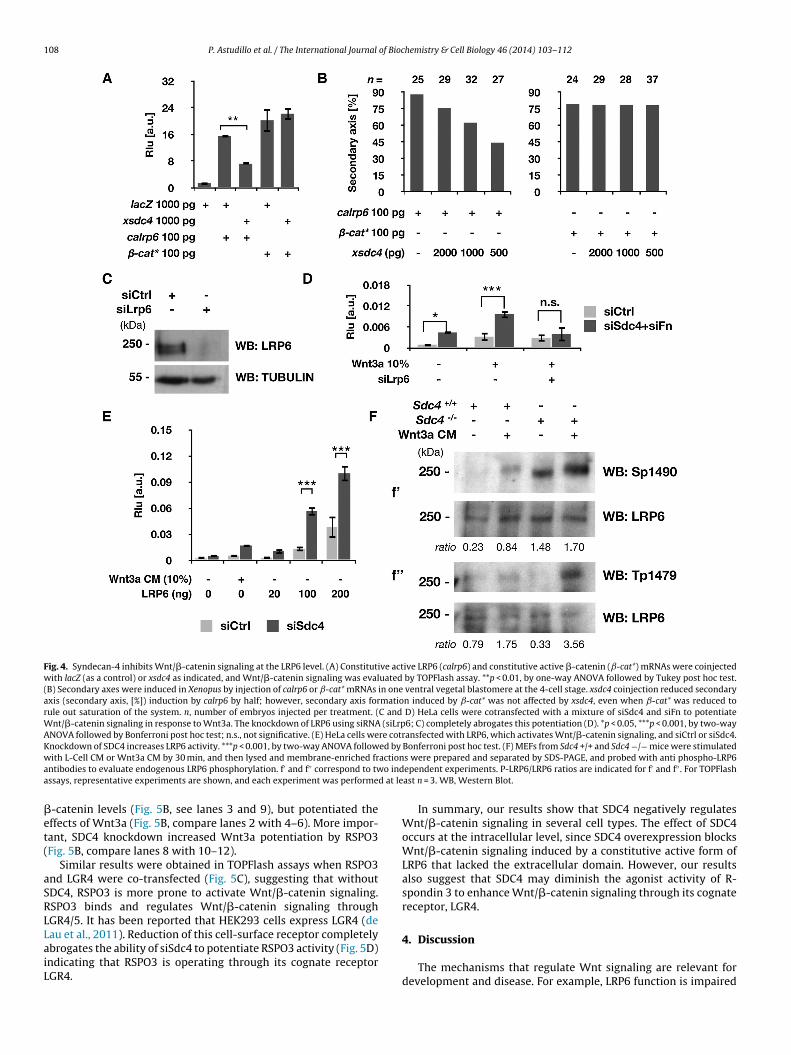
Fig. 4A), and to induce secondary axis formation (Fig. 4B), but wasompletely unable to block ˇ-catenin* activity (Fig. 4A and B).
The finding that SDC4 is able to inhibit Wnt/�-catenin signalingctivated by a truncated form of LRP6 consisting only in the intra-ellular and transmembrane domains, suggests that SDC4 regulates
nt/�-catenin through modulation of LRP6 activity at the intracel-ular level. Therefore, LRP6 should be necessary for the increase of
nt/�-catenin signaling induced by SDC4 knockdown. To directlyest this, we evaluated whether the knockdown of SDC4 using siR-As was able to potentiate Wnt/�-catenin signaling when LRP6
evels were reduced. Importantly, reduction of LRP6 levels by siRNAFig. 4C) completely abrogated the potentiation of Wnt3a-induced
nt/�-catenin signaling observed when SDC4 and FN levels wereeduced (Fig. 4D). In agreement with these observations, the reduc-ion of SDC4 levels enhanced the activity of overexpressed LRP6Fig. 4E).
LRP6 phosphorylation is a hallmark of activation of the Wnt/�-atenin signaling pathway (Davidson et al., 2005; Zeng et al.,005). To evaluate whether SDC4 regulates LRP6 phosphoryla-ion, wild-type and Sdc4 null MEFs were incubated with controlr Wnt3a conditioned medium for 30 min, and plasma-membraneractions were obtained (Davidson et al., 2005). LRP6 phosphory-ation on Serine 1490 and Threonine 1479 was evaluated. These
wo residues are phosphorylated sequentially by different kinasesGSK3� and CKI�, respectively) (Davidson et al., 2005; Zeng et al.,005). Higher levels of LRP6 phosphorylation were detected indc4 null mice even in the absence of exogenous Wnt3a (Fig. 4F).SDC4 (mSDC4) expression, decreased Wnt/�-catenin signaling (see bars 4–5). Both6). **p < 0.01, ***p < 0.001, by one-way ANOVA followed by Tukey post hoc test; n.d.,
In summary, these results strongly suggest that SDC4 inhibitsWnt/�-catenin signaling via modulation of LRP6 activity. How-ever, given the complexity of the events triggered by the activationof the Wnt/�-catenin pathway, our results do not exclude thepossibility that other mechanisms may also contribute to theinhibition of the Wnt/�-catenin signaling by SDC4 (see Section4).
3.3. Syndecan-4 regulates the ability of R-spondin 3 to activatethe Wnt/ˇ-catenin pathway
Recently, a key role for R-spondin (RSPO) in the modulation ofWnt signaling has been demonstrated. R-spondins bind to the cell-surface receptors LGR4/5 and enhances Wnt/�-catenin signaling(Glinka et al., 2011; de Lau et al., 2011). RSPO3 also binds to SDC4to regulate Wnt/PCP signaling (Ohkawara et al., 2011). Consideringthese findings, we studied whether the ability of RSPO3 to enhanceWnt/�-catenin signaling was regulated by SDC4. For this we testedthe effect of reducing SDC4 levels on the ability of RSPO3 to enhanceWnt3a activity. Addition of pure recombinant RSPO3 enhancedWnt3a activity, as described previously (Glinka et al., 2011; de Lauet al., 2011), and this increase was higher when SDC4 levels werereduced (Fig. 5A). Of note, SDC4 knockdown increased RSPO3 activ-
ity, even when exogenous Wnt3a was not added (SupplementaryFig. 4).A similar result was obtained when �-catenin stabilization inL-Cells was analyzed, since RSPO3 alone was not able to increase
108 P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112
Fig. 4. Syndecan-4 inhibits Wnt/�-catenin signaling at the LRP6 level. (A) Constitutive active LRP6 (calrp6) and constitutive active �-catenin (ˇ-cat*) mRNAs were coinjectedwith lacZ (as a control) or xsdc4 as indicated, and Wnt/�-catenin signaling was evaluated by TOPFlash assay. **p < 0.01, by one-way ANOVA followed by Tukey post hoc test.(B) Secondary axes were induced in Xenopus by injection of calrp6 or ˇ-cat* mRNAs in one ventral vegetal blastomere at the 4-cell stage. xsdc4 coinjection reduced secondaryaxis (secondary axis, [%]) induction by calrp6 by half; however, secondary axis formation induced by ˇ-cat* was not affected by xsdc4, even when ˇ-cat* was reduced torule out saturation of the system. n, number of embryos injected per treatment. (C and D) HeLa cells were cotransfected with a mixture of siSdc4 and siFn to potentiateWnt/�-catenin signaling in response to Wnt3a. The knockdown of LRP6 using siRNA (siLrp6; C) completely abrogates this potentiation (D). *p < 0.05, ***p < 0.001, by two-wayANOVA followed by Bonferroni post hoc test; n.s., not significative. (E) HeLa cells were cotransfected with LRP6, which activates Wnt/�-catenin signaling, and siCtrl or siSdc4.Knockdown of SDC4 increases LRP6 activity. ***p < 0.001, by two-way ANOVA followed by Bonferroni post hoc test. (F) MEFs from Sdc4 +/+ and Sdc4 −/− mice were stimulatedw actiona o inda d at le
�et(
aSRLLaiL
ith L-Cell CM or Wnt3a CM by 30 min, and then lysed and membrane-enriched frntibodies to evaluate endogenous LRP6 phosphorylation. f′ and f′′ correspond to twssays, representative experiments are shown, and each experiment was performe
-catenin levels (Fig. 5B, see lanes 3 and 9), but potentiated theffects of Wnt3a (Fig. 5B, compare lanes 2 with 4–6). More impor-ant, SDC4 knockdown increased Wnt3a potentiation by RSPO3Fig. 5B, compare lanes 8 with 10–12).
Similar results were obtained in TOPFlash assays when RSPO3nd LGR4 were co-transfected (Fig. 5C), suggesting that withoutDC4, RSPO3 is more prone to activate Wnt/�-catenin signaling.SPO3 binds and regulates Wnt/�-catenin signaling throughGR4/5. It has been reported that HEK293 cells express LGR4 (de
au et al., 2011). Reduction of this cell-surface receptor completelybrogates the ability of siSdc4 to potentiate RSPO3 activity (Fig. 5D)ndicating that RSPO3 is operating through its cognate receptorGR4.s were prepared and separated by SDS-PAGE, and probed with anti phospho-LRP6ependent experiments. P-LRP6/LRP6 ratios are indicated for f′ and f′′ . For TOPFlashast n = 3. WB, Western Blot.
In summary, our results show that SDC4 negatively regulatesWnt/�-catenin signaling in several cell types. The effect of SDC4occurs at the intracellular level, since SDC4 overexpression blocksWnt/�-catenin signaling induced by a constitutive active form ofLRP6 that lacked the extracellular domain. However, our resultsalso suggest that SDC4 may diminish the agonist activity of R-spondin 3 to enhance Wnt/�-catenin signaling through its cognatereceptor, LGR4.
4. Discussion
The mechanisms that regulate Wnt signaling are relevant fordevelopment and disease. For example, LRP6 function is impaired
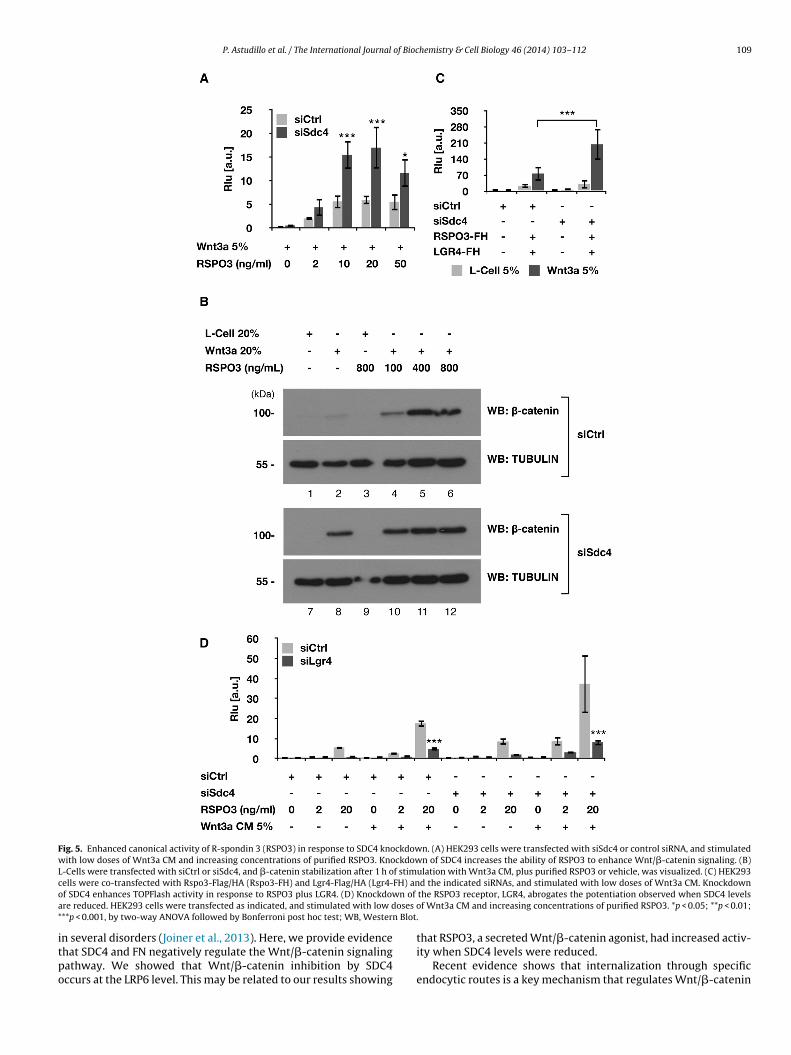
P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112 109
Fig. 5. Enhanced canonical activity of R-spondin 3 (RSPO3) in response to SDC4 knockdown. (A) HEK293 cells were transfected with siSdc4 or control siRNA, and stimulatedwith low doses of Wnt3a CM and increasing concentrations of purified RSPO3. Knockdown of SDC4 increases the ability of RSPO3 to enhance Wnt/�-catenin signaling. (B)L-Cells were transfected with siCtrl or siSdc4, and �-catenin stabilization after 1 h of stimulation with Wnt3a CM, plus purified RSPO3 or vehicle, was visualized. (C) HEK293c FH) ano n of
a oses o* n Blot.
itpo
ells were co-transfected with Rspo3-Flag/HA (Rspo3-FH) and Lgr4-Flag/HA (Lgr4-f SDC4 enhances TOPFlash activity in response to RSPO3 plus LGR4. (D) Knockdowre reduced. HEK293 cells were transfected as indicated, and stimulated with low d**p < 0.001, by two-way ANOVA followed by Bonferroni post hoc test; WB, Wester
n several disorders (Joiner et al., 2013). Here, we provide evidencehat SDC4 and FN negatively regulate the Wnt/�-catenin signalingathway. We showed that Wnt/�-catenin inhibition by SDC4ccurs at the LRP6 level. This may be related to our results showing
d the indicated siRNAs, and stimulated with low doses of Wnt3a CM. Knockdownthe RSPO3 receptor, LGR4, abrogates the potentiation observed when SDC4 levelsf Wnt3a CM and increasing concentrations of purified RSPO3. *p < 0.05; **p < 0.01;
that RSPO3, a secreted Wnt/�-catenin agonist, had increased activ-ity when SDC4 levels were reduced.
Recent evidence shows that internalization through specificendocytic routes is a key mechanism that regulates Wnt/�-catenin

110 P. Astudillo et al. / The International Journal of Biochemistry & Cell Biology 46 (2014) 103– 112
Fig. 6. Model for the regulation of Wnt signaling by SDC4. Wnt/�-catenin signaling, initiated by Wnt ligands, relies on receptors (Frizzled, FZD) and co-receptors (LRP6) atthe plasma membrane, plus synergic activities of Rspo ligands, inducing LRP6 phosphorylation, by mechanisms mediated by Dishevelled and other cytoplasmic proteins.N d by
r nally
a ay bei ion of
aaeuftimrrlicinuaipntrwntrau
t2rswi(awfs
on-canonical Wnt signaling mediated by Wnt ligands, such as Wnt5a, is initiateeceptors and Dishevelled. SDC4 participates in this process, by interacting functiolso with RSPO3 (Ohkawara et al., 2011). It is likely that the participation of SDC4 mn titration of RSPO3, or competition with intracellular proteins, resulting in inhibit
ctivation (Niehrs, 2012; Kikuchi et al., 2009). Some proteins, suchs Disabled-2 (Jiang et al., 2012), Waif1/5T4 (Kagermeier-Schenkt al., 2011) and R-spondins (Koo et al., 2012; Hao et al., 2012), reg-late either the localization of LRP6, its endocytosis or its clearancerom the plasma membrane. Hence, LRP6 is a focal point to con-rol the Wnt/�-catenin pathway, and our results suggest that SDC4nhibits the pathway by regulating LRP6 activity by an undefined
echanism. Further experiments are needed to test whether SDC4egulates LRP6 activity by regulating its internalization by specificoutes. Importantly, inhibition of LRP6 occured at the intracellularevel, because SDC4 was able to inhibit Wnt/�-catenin signalingnduced by a constitutive active form of LRP6 that lacked the extra-ellular domain (caLRP6). Our results suggest that SDC4 did notmpair LRP6 total levels; however, LRP6 phosphorylation in Sdc4ull MEFs was increased. Therefore, SDC4 could inhibit LRP6 by reg-lating the localization or activity of intracellular proteins, likelyffecting its phosphorylation status, with possible consequencesn the internalization of Wnt components. Our results exclude theossibility that SDC4 may inhibit the Wnt/�-catenin pathway at theucleus, for example by regulating the �-catenin transcription fac-or, as it has been shown for the transcription factor ATF2, which isegulated by SDC4 through modulation of the Rac1/p38/JNK path-ay (Saoncella et al., 2004). However, the results presented here doot exclude the possibility that other mechanisms, such as seques-ration of Wnt ligands through the heparan sulphate chains, oregulation of other intracellular proteins (such as Dishevelled), maylso contribute to the inhibition of the Wnt/�-catenin signalingnder specific circumstances.
Previous studies from our group and others have demonstratedhat SDC4 regulates the Wnt/PCP pathway in Xenopus (Munoz et al.,006; Matthews et al., 2008; Ohkawara et al., 2011). SDC4 mayegulate Wnt/PCP signaling by modulating intracellular proteinsuch as Dishevelled, which interacts physically and biochemicallyith SDC4 (Munoz et al., 2006; Carvallo et al., 2010), or by sensitiz-
ng cells to R-spondin ligands when these proteins are expressedOhkawara et al., 2011). We must note that these two mech-
nisms can be complementary, and likely represent redundantays to regulate Wnt/PCP signaling. Furthermore, recent resultsrom our laboratory show that SDC4 is also involved in Wnt/PCPignaling during mouse development (Escobedo et al., 2013). Sdc4
plasma membrane components and through mechanisms involving also Frizzledwith Dishevelled and Frizzled-7 (see Munoz et al., 2006; Carvallo et al., 2010), and
related with adhesion proteins, such as Fibronectin and integrins; this may result the Wnt/�-catenin signaling.
can interact genetically with Vangl2/Strabismus, which codes for aWnt/PCP co-receptor, to regulate neural tube closure during mousedevelopment. We also showed that VANGL2 regulates SDC4 lev-els, suggesting a mechanism for this interaction. In addition, it hasbeen shown recently that SDC4 is involved in non-canonical Wntsignaling induced by Wnt7a in mouse satellite stem cells, by form-ing a complex with Frizzled-7 and FN (Bentzinger et al., 2013).Collectively, this evidence supports the notion that the ability ofSDC4 to regulate the non-canonical Wnt pathway is a conservedfeature.
Both branches of the Wnt pathway (canonical and non canon-ical) establish a cross-talk and share components like Dishevelledand FZ receptors. For example, non-canonical Wnt ligands eitheractivate (He et al., 1997) or inhibit Wnt/�-catenin signaling (Torreset al., 1996) in Xenopus. This raises the possibility that proteinsregulating the non-canonical Wnt pathway could also regulateWnt/�-catenin signaling. Considering the evidence mentioned andthe results presented in this work, to date SDC4 is the only HSPGthat has an opposite effect on each branch of the Wnt signalingpathway. This differentiates SDC4 from Glypican-4/Knypek (GPC4),a HSPG from the Glypican family that has been shown to be involvedin regulation of Wnt signaling (Topczewski et al., 2001). GPC4 regu-lates both canonical and non-canonical Wnt pathways (Ohkawaraet al., 2003, Sakane et al., 2012), but two features differentiate GPC4from SDC4. First, GPC4 can promote and inhibit both Wnt pathways,depending on its localization in specific domains at the cell mem-brane (Sakane et al., 2012). Second, GPC4 regulates Wnt signalingat the extracellular level, since Glypicans are devoid from intracel-lular domains and harbors a cysteine-rich domain (CRD), which isessential for its role in regulating Wnt signaling (Ohkawara et al.,2003).
Several inhibitors of the Wnt/�-catenin pathway, such as Dick-kopfs and sFRPs, can be expressed at a given time, whereas some ofthem are also Wnt/�-catenin target genes, and are therefore con-trolled by the pathway. This suggests that redundant mechanismsmay ensure proper regulation of the pathway. Nonetheless, SDC4
could be relevant in tissues or cell types under regulation of R-spondin ligands, such as in the intestinal crypt, where R-spondinligands plays a relevant role regulating the Wnt/�-catenin pathway(Jin and Yoon, 2012), and where hyperactivation of this pathway
of Bioc
cs
fWsi2tWaWtt
acWcnbSassie(ed
wiFfo2tcfrtm
A
CiwCPPdC
A
f2
R
A
P. Astudillo et al. / The International Journal
ould lead to disease. SDC4 may assist to restrict Wnt/�-cateninignaling and ensure limited activation of the pathway.
This same role could be exerted by SDC4 during development,or example during Xenopus gastrulation, where the non-canonical
nt pathway plays a pivotal role, and where cells from the dor-al marginal zone migrate along a layer of Fibronectin through thenner roof of the blastocoele at the onset of gastrulation (Keller et al.,003). SDC4 could have a role in regulating the polarized migra-ion of cells along the FN-rich layer, and additionally in restricting
nt/�-catenin signaling. It must be noted that LRP6 also plays role in this process, since it has been described that opposesnt/PCP signaling during gastrulation (Tahinci et al., 2007), raising
he interesting possibility that plasma membrane proteins regulatehe interplay between Wnt branches.
VANGL2 (Park and Moon, 2002), ROR2 (Witte et al., 2010)nd PTK7 (Peradziryi et al., 2011), other plasma-membraneomponents of the non-canonical Wnt pathway, also inhibit
nt/�-catenin signaling. This suggests that control of the Wnt/�-atenin pathway by plasma-membrane components from theon-canonical Wnt pathway has physiological relevance and maye a common feature of Wnt/PCP components. In this context,DC4 is especially important because it possibly links cell/matrixdhesion via focal adhesions with Wnt signaling, with possible con-equences in pathological conditions. For example, recent evidencehows that Syndecans might be associated with cancer (reviewedn Choi et al., 2013), including acute myeloid leukemia (AML), a dis-ase characterized by (a) an increase of the Wnt/�-catenin signalingWang et al., 2010), and (b) the loss of expression of Sdc4 (Toyotat al., 2001) and other Wnt/�-catenin inhibitors (Hou et al., 2011),ue to hypermethylation of their promoters.
Based on our results and previous evidence we propose a modelhereby SDC4 influences each branch of the Wnt pathway, depend-
ng on the availability of specific Wnt or R-spondin ligands andrizzled receptors (Fig. 6). Furthermore, SDC4 titrates (or competesor) intracellular proteins, such as Dishevelled, due to the abilityf SDC4 to bind to this protein (Munoz et al., 2006; Carvallo et al.,010). The functional interaction of SDC4 with other Wnt/PCP pro-eins, especially VANGL2 (Escobedo et al., 2013) may increase theomplexity of the modulation of the Wnt pathway by SDC4. There-ore, the results presented in this work establish SDC4 as a newegulator of the Wnt/�-catenin pathway, and provide a novel targeto gain insights about new therapeutic approaches and develop-
ental mechanisms of Wnt/�-catenin regulation.
cknowledgments
We thank Dr. Edward de Robertis, Dr. Hans Clevers and Dr.hristof Niehrs for providing useful materials and reagents used
n this work. Also, we thank Dr. N. Escobedo for her assistanceith the Sdc4 −/− mice. This work was supported by FONDE-YT #1100471, the Millennium Nucleus for Regenerative Biology07-011-F from the Iniciativa Científica Milenio, and Basal fundingFB12/2007 (J.L.), and the “Beca de Doctorado Nacional” and “Becae Apoyo para la Realización de Tesis Doctoral AT-24100095” fromONICYT (P.A.).
ppendix A. Supplementary data
Supplementary data associated with this article can beound, in the online version, at http://dx.doi.org/10.1016/j.biocel.013.11.012.
eferences
bràmoff M, Magalhães P, Ram S. Image processing with ImageJ. Biophotonics Int2004;11:36–42.
hemistry & Cell Biology 46 (2014) 103– 112 111
Bentzinger C, Wang Y, Von Maltzahn J, Soleimani V, Yin H, Rudnicki M.Fibronectin regulates Wnt7a signaling and satellite cell expansion. Cell StemCell 2013;12:75–87.
Bertrand J, Stange R, Hidding H, Echtermeyer F, Nalesso G, Godmann L, et al. Synde-can 4 supports bone fracture repair, but not fetal skeletal development, in mice.Arthritis Rheum 2013;65:743–52.
Carvallo L, Munoz R, Bustos F, Escobedo N, Carrasco H, Olivares G, et al. Non-canonicalWnt signaling induces ubiquitination and degradation of Syndecan4. J Biol Chem2010;285:29546–55.
Choi S, Kang D-H, Oh E-S. Targeting syndecans: a promising strategy for the treat-ment of cancer. Expert Opin Ther Targets 2013;17:695–705.
Clevers H, Nusse R. Wnt/�-catenin signaling and disease. Cell 2012;149:1192–205.Couchman J. Transmembrane signaling proteoglycans. Annu Rev Cell Dev Biol
2010;26:89–114.Davidson G, Wu W, Shen J, Bilic J, Fenger U, Stannek P, et al. Casein kinase 1 gamma
couples Wnt receptor activation to cytoplasmic signal transduction. Nature2005;438:867–72.
de Lau W, Barker N, Low T, Koo B, Li V, Teunissen H, et al. Lgr5 homologues associatewith Wnt receptors and mediate R-spondin signalling. Nature 2011;476:293–7.
Elfenbein A, Simons M. Syndecan-4 signaling at a glance. J Cell Sci2013;126:3799–804.
Escobedo N, Contreras O, Munoz R, Farías M, Carrasco H, Hill C, et al. Syndecan 4interacts genetically with Vangl2 to regulate neural tube closure and planar cellpolarity. Development 2013;140:3008–17.
Glinka A, Dolde C, Kirsch N, Huang Y, Kazanskaya O, Ingelfinger D, et al. LGR4and LGR5 are R-spondin receptors mediating Wnt/�-catenin and Wnt/PCPsignalling. EMBO Rep 2011;12:1055–61.
Hao H, Xie Y, Zhang Y, Charlat O, Oster E, Avello M, et al. ZNRF3 promotesWnt receptor turnover in an R-spondin-sensitive manner. Nature 2012;485:195–200.
Herum K, Lunde I, Skrbic B, Florholmen G, Behmen D, Sjaastad I, et al. Syndecan-4signaling via NFAT regulates extracellular matrix production and cardiac myofi-broblast differentiation in response to mechanical stress. J Mol Cell Cardiol2013;54:73–81.
Hou H, Kuo Y, Liu C, Lee M, Tang J, Chen C, et al. Distinct association between aber-rant methylation of Wnt inhibitors and genetic alterations in acute myeloidleukaemia. Br J Cancer 2011;105:1927–33.
Jiang Y, He X, Howe P. Disabled-2 (Dab2) inhibits Wnt/�-catenin signalling bybinding LRP6 and promoting its internalization through clathrin. EMBO J2012;31:2336–49.
Jin Y, Yoon J. The R-spondin family of proteins: emerging regulators of WNTsignaling. Int J Biochem Cell Biol 2012;44:2278–87.
Joiner D, Ke J, Zhong Z, Xu H, Williams B. LRP5 and LRP6 in development and disease.Trends Endocrinol Metab 2013;24:31–9.
Kagermeier-Schenk B, Wehner D, Ozhan-Kizil G, Yamamoto H, Li J, Kirchner K,et al. Waif1/5T4 inhibits Wnt/�-catenin signaling and activates noncanon-ical Wnt pathways by modifying LRP6 subcellular localization. Dev Cell2011;21:1129–43.
Keller R, Davidson L, Shook D. How we are shaped: the biomechanics of gastrulation.Differentiation 2003;71:171–205.
Kikuchi A, Yamamoto H, Sato A. Selective activation mechanisms of Wnt signalingpathways. Trends Cell Biol 2009;19:119–29.
Koo B, Spit M, Jordens I, Low T, Stange D, Van de Wetering M, et al. Tumour suppressorRNF43 is a stem-cell E3 ligase that induces endocytosis of Wnt receptors. Nature2012;488:665–9.
Marsden M, DeSimone D. Regulation of cell polarity, radial intercalation andepiboly in Xenopus: novel roles for integrin and fibronectin. Development2001;128:3635–47.
Matthews H, Marchant L, Carmona-Fontaine C, Kuriyama S, Larraín J, HoltM, et al. Directional migration of neural crest cells in vivo is regulatedby Syndecan-4/Rac1 and non-canonical Wnt signaling/RhoA. Development2008;135:1771–80.
McMahon A, Moon R. Ectopic expression of the proto-oncogene int-1 in Xeno-pus embryos leads to duplication of the embryonic axis. Cell 1989;58:1075–84.
Molenaar M, Van de Wetering M, Oosterwegel M, Peterson-Maduro J, Godsave S,Korinek V, et al. XTcf-3 transcription factor mediates beta-catenin-induced axisformation in Xenopus embryos. Cell 1996;86:391–9.
Montcouquiol M, Crenshaw E, Kelley M. Noncanonical Wnt signaling and neuralpolarity. Annu Rev Neurosci 2006;29:363–86.
Munoz R, Moreno M, Oliva C, Orbenes C, Larraín J. Syndecan-4 regulates non-canonical Wnt signalling and is essential for convergent and extensionmovements in Xenopus embryos. Nat Cell Biol 2006;8:492–500.
Niehrs C. The complex world of WNT receptor signalling. Nat Rev Mol Cell Biol2012;13:767–79.
Ohkawara B, Yamamoto T, Tada M, Ueno N. Role of glypican 4 in the regulation ofconvergent extension movements during gastrulation in Xenopus laevis. Devel-opment 2003;130:2129–38.
Ohkawara B, Glinka A, Niehrs C. Rspo3 binds Syndecan 4 and induces Wnt/PCPsignaling via clathrin-mediated endocytosis to promote morphogenesis. DevCell 2011;20:303–14.
Park M, Moon R. The planar cell-polarity gene stbm regulates cell behaviour and cellfate in vertebrate embryos. Nat Cell Biol 2002;4:20–5.
Peradziryi H, Kaplan N, Podleschny M, Liu X, Wehner P, Borchers A, et al.PTK7/Otk interacts with Wnts and inhibits canonical Wnt signalling. EMBO J2011;30:3729–40.

1 of Bioc
S
S
S
T
T
T
12 P. Astudillo et al. / The International Journal
akane H, Yamamoto H, Matsumoto S, Sato A, Kikuchi A. Localization of glypican-4 in different membrane microdomains is involved in the regulation of Wntsignaling. J Cell Sci 2012;125:449–60.
aoncella S, Calautti E, Neveu W, Goetinck PF. Syndecan-4 regulates ATF-2transcriptional activity in a Rac1-dependent manner. J Biol Chem 2004;279:47172–6.
hibamoto S, Higano K, Takada R, Ito F, Takeichi M, Takada S. Cytoskeletalreorganization by soluble Wnt-3a protein signalling. Genes Cells 1998;3:659–70.
aelman V, Dobrowolski R, Plouhinec J, Fuentealba L, Vorwald P, Gumper I, et al.Wnt signaling requires sequestration of glycogen synthase kinase 3 inside mul-tivesicular endosomes. Cell 2010;143:1136–48.
ahinci E, Thorne C, Franklin J, Salic A, Christian K, Lee L, et al. Lrp6 is
required for convergent extension during Xenopus gastrulation. Development2007;134:4095–106.opczewski J, Sepich D, Myers D, Walker C, Amores A, Lele Z, et al. The zebrafish glyp-ican knypek controls cell polarity during gastrulation movements of convergentextension. Dev Cell 2001;1:251–64.
hemistry & Cell Biology 46 (2014) 103– 112
Torres M, Yang-Snyder J, Purcell S, DeMarais A, McGrew L, Moon R. Activities of theWnt-1 class of secreted signaling factors are antagonized by the Wnt-5A classand by a dominant negative cadherin in early Xenopus development. J Cell Biol1996;133:1123–37.
Toyota M, Kopecky K, Toyota M, Jair K, Willman C, Issa J. Methylation profiling inacute myeloid leukemia. Blood 2001;97:2823–9.
Wang Y, Krivtsov A, Sinha A, North T, Goessling W, Feng Z, et al. The Wnt/beta-catenin pathway is required for the development of leukemia stem cells in AML.Science 2010;327:1650–3.
Witte F, Bernatik O, Kirchner K, Masek J, Mahl A, Krejci P, et al. Negative regulationof Wnt signaling mediated by CK1-phosphorylated Dishevelled via Ror2. FASEBJ 2010;24:2417–26.
Woods A, Couchman J. Syndecan 4 heparan sulfate proteoglycan is a selec-
tively enriched and widespread focal adhesion component. Mol Biol Cell1994;5:183–92.Zeng X, Tamai K, Doble B, Li S, Huang H, Habas R, et al. A dual-kinase mecha-nism for Wnt co-receptor phosphorylation and activation. Nature 2005;438:873–7.