Rivaroxaban Inhibits Angiotensin II-Induced Activation in ... · 544 Rivaroxaban Inhibits...
7
544 Rivaroxaban Inhibits Angiotensin II-Induced Activation in Cultured Mouse Cardiac Fibroblasts Through the Modulation of NF-κB Pathway Takehiro Hashikata, 1 MD, Minako Y amaoka-Tojo, 1,2 MD, Sayaka Namba, 1 MD, Lisa Kitasato, 1 MD, Ryo Kameda, 3 MD, Masami Murakami, 3 MD, Hiroe Niwano, 4 MD, Takao Shimohama, 3 MD, Taiki Tojo, 3 MD, and Junya Ako, 1,3 MD Summary Cell migration, proliferation, and differentiation of cardiac fibroblasts (CFs) play a central role in cardiac fibrosis. Factor Xa (FXa)-dependent protease-activated receptor (PAR)-1 and PAR-2 have been reported as important targets in proinflammatory and fibroproliferative diseases. From this viewpoint, we aimed to investigate whether treatment of ri- varoxaban, an approved oral direct FXa inhibitor, attenuates functional changes in angiotensin (Ang) II-induced mouse CFs. Confluent cultured mouse CFs were pretreated with or without rivaroxaban. Ang II-induced cell migration was de- creased by 73% in rivaroxaban induced cells. Rivaroxaban inhibited Ang II-induced cell proliferation by 27% at 0.01 μg/ mL, 69% at 0.1 μg/mL, 71% at 1 μg/mL, and 69% at 5 μg/mL. In mouse cytokine array measuring 40 cytokines, the productions of interleukin-16, TIMP-1, and tumor necrosis factor-α (TNF-α) were significantly reduced with 0.1 μg/mL of rivaroxaban pretreatment (all P < 0.05). TIMP-1 levels in the culture supernatant measured by ELISA were also de- creased by rivaroxaban pretreatment in Ang II-induced CFs (35% decrease at 0.01 μg/mL, 47% at 0.1 μg/mL, 47% at 1 μg/mL, and 57% at 5 μg/mL). In the dual reporter assay analysis, rivaroxaban inhibited various inflammatory signal pathways, including the nuclear factor-kappa B (NF-κB), active protein-1 (AP-1), and mitogen-activated protein kinase (MAPK) pathways (decreases of 82%, 78%, and 75%, respectively). These data suggest that rivaroxaban inhibits Ang II-induced functional activation in cultured mouse CFs via inhibit- ing NF-κB and MAPK/AP-1 signaling pathways, which may be a possible target of heart failure, through the antifibrotic and anti-inflammatory efficacy of rivaroxaban in Ang II-stimulated cardiac fibroblasts. (Int Heart J 2015; 56: 544-550) Key words: Factor Xa inhibitor, Cardiac fibrosis, Anticoagulant, Cell signaling, Inflammation T he pathologic changes in heart failure with preserved ejection fraction (HFpEF) occur in multiple steps: in- cluding cardiomyocyte hypertrophic remodeling and cell death, coronary microvascular rarefaction, and myocardial fibrosis caused by excessive functional activation of cardiac fi- broblasts (CFs). Many recent studies focused on the impor- tance of cardiac fibrosis in the pathogenesis of heart failure. 1-3) Modulation of cardiac fibrosis has become an important target for failing hearts in postischemic cardiac remodeling and re- generation therapy. Improvement of left ventricular systolic/di- astolic function in patients with HF was closely related to changes in myocardial collagen metabolism. 4) Cardiac fibrosis is defined as a progressive accumulation of fibrillar extracellu- lar matrix (ECM) in the myocardium. The regulation of ECM remodeling is primarily mediated by CFs, which are the main non-muscle cells in the heart. CFs modulate matrix turnover in non-pathological conditions, and their activity is greatly en- hanced after an acute cardiac event or during chronic cardio- vascular disease. 5) The phenotypic transformation of fibroblasts into osteoblasts, stimulated by the renin-angiotensin system (RAS), leads to tissue calcification. 6,7) It is known that angi- otensin II (Ang II) is the main active peptide in the RAS, regu- lating cellular and physiological responses in the cardiovascu- lar system. 8) A lot of evidence has implicated Ang II in the progression of myocardial fibrosis. Ang II has been suggested to be a potent pro-fibrotic molecule. 9) Increased serum levels of Ang II are seen in patients with cardiovascular diseases that are associated with myocardial fibrosis, including atherosclerosis, hypertension, cardiac hypertrophy, and heart failure. 9-11) Rapid myocardial cellular infiltration is evident in animals infused with Ang II, which suggests that the cellular component has an effector function in the development of myocardial fibrosis. 12) From the 1 Kitasato University Graduate School of Medical Sciences, 2 Department of Rehabilitation, Kitasato University School of Allied Health Sciences, 3 Depart- ment of Cardiovascular Medicine, Kitasato University School of Medicine, Sagamihara, and 4 Department of Education, Tamagawa University College of Education, Machida, Japan. Address for correspondence: Minako Yamaoka-Tojo, MD, Department of Rehabilitation, Kitasato University School of Allied Health Sciences, 1-15-1 Kitasato, Minami-ku, Sagamihara, Kanagawa 252-0373, Japan. E-mail: [email protected] Received for publication March 7, 2015. Revised and accepted April 6, 2015. Released in advance online on J-STAGE September 11, 2015. All rights reserved by the International Heart Journal Association.
Transcript of Rivaroxaban Inhibits Angiotensin II-Induced Activation in ... · 544 Rivaroxaban Inhibits...

544
Rivaroxaban Inhibits Angiotensin II-Induced Activation in Cultured Mouse Cardiac Fibroblasts Through the
Modulation of NF-κB PathwayTakehiro Hashikata,1 MD, Minako Yamaoka-Tojo,1,2 MD, Sayaka Namba,1 MD,
Lisa Kitasato,1 MD, Ryo Kameda,3 MD, Masami Murakami,3 MD, Hiroe Niwano,4 MD, Takao Shimohama,3 MD, Taiki Tojo,3 MD, and Junya Ako,1,3 MD
Summary
Cell migration, proliferation, and differentiation of cardiac fibroblasts (CFs) play a central role in cardiac fibrosis. Factor Xa (FXa)-dependent protease-activated receptor (PAR)-1 and PAR-2 have been reported as important targets in proinflammatory and fibroproliferative diseases. From this viewpoint, we aimed to investigate whether treatment of ri-varoxaban, an approved oral direct FXa inhibitor, attenuates functional changes in angiotensin (Ang) II-induced mouse CFs.
Confluent cultured mouse CFs were pretreated with or without rivaroxaban. Ang II-induced cell migration was de-creased by 73% in rivaroxaban induced cells. Rivaroxaban inhibited Ang II-induced cell proliferation by 27% at 0.01 μg/mL, 69% at 0.1 μg/mL, 71% at 1 μg/mL, and 69% at 5 μg/mL. In mouse cytokine array measuring 40 cytokines, the productions of interleukin-16, TIMP-1, and tumor necrosis factor-α (TNF-α) were significantly reduced with 0.1 μg/mL of rivaroxaban pretreatment (all P < 0.05). TIMP-1 levels in the culture supernatant measured by ELISA were also de-creased by rivaroxaban pretreatment in Ang II-induced CFs (35% decrease at 0.01 μg/mL, 47% at 0.1 μg/mL, 47% at 1 μg/mL, and 57% at 5 μg/mL). In the dual reporter assay analysis, rivaroxaban inhibited various inflammatory signal pathways, including the nuclear factor-kappa B (NF-κB), active protein-1 (AP-1), and mitogen-activated protein kinase (MAPK) pathways (decreases of 82%, 78%, and 75%, respectively).
These data suggest that rivaroxaban inhibits Ang II-induced functional activation in cultured mouse CFs via inhibit-ing NF-κB and MAPK/AP-1 signaling pathways, which may be a possible target of heart failure, through the antifibrotic and anti-inflammatory efficacy of rivaroxaban in Ang II-stimulated cardiac fibroblasts. (Int Heart J 2015; 56: 544-550)
Key words: Factor Xa inhibitor, Cardiac fibrosis, Anticoagulant, Cell signaling, Inflammation
T he pathologic changes in heart failure with preserved ejection fraction (HFpEF) occur in multiple steps: in-cluding cardiomyocyte hypertrophic remodeling and
cell death, coronary microvascular rarefaction, and myocardial fibrosis caused by excessive functional activation of cardiac fi-broblasts (CFs). Many recent studies focused on the impor-tance of cardiac fibrosis in the pathogenesis of heart failure.1-3) Modulation of cardiac fibrosis has become an important target for failing hearts in postischemic cardiac remodeling and re-generation therapy. Improvement of left ventricular systolic/di-astolic function in patients with HF was closely related to changes in myocardial collagen metabolism.4) Cardiac fibrosis is defined as a progressive accumulation of fibrillar extracellu-lar matrix (ECM) in the myocardium. The regulation of ECM remodeling is primarily mediated by CFs, which are the main non-muscle cells in the heart. CFs modulate matrix turnover in
non-pathological conditions, and their activity is greatly en-hanced after an acute cardiac event or during chronic cardio-vascular disease.5) The phenotypic transformation of fibroblasts into osteoblasts, stimulated by the renin-angiotensin system (RAS), leads to tissue calcification.6,7) It is known that angi-otensin II (Ang II) is the main active peptide in the RAS, regu-lating cellular and physiological responses in the cardiovascu-lar system.8) A lot of evidence has implicated Ang II in the progression of myocardial fibrosis. Ang II has been suggested to be a potent pro-fibrotic molecule.9) Increased serum levels of Ang II are seen in patients with cardiovascular diseases that are associated with myocardial fibrosis, including atherosclerosis, hypertension, cardiac hypertrophy, and heart failure.9-11) Rapid myocardial cellular infiltration is evident in animals infused with Ang II, which suggests that the cellular component has an effector function in the development of myocardial fibrosis.12)
From the 1 Kitasato University Graduate School of Medical Sciences, 2 Department of Rehabilitation, Kitasato University School of Allied Health Sciences, 3 Depart-ment of Cardiovascular Medicine, Kitasato University School of Medicine, Sagamihara, and 4 Department of Education, Tamagawa University College of Education, Machida, Japan.
Address for correspondence: Minako Yamaoka-Tojo, MD, Department of Rehabilitation, Kitasato University School of Allied Health Sciences, 1-15-1 Kitasato, Minami-ku, Sagamihara, Kanagawa 252-0373, Japan. E-mail: [email protected]
Received for publication March 7, 2015. Revised and accepted April 6, 2015.Released in advance online on J-STAGE September 11, 2015.All rights reserved by the International Heart Journal Association.

545Vol 56No 5 RIVAROXABAN INHIBITS FIBROTIC PROGRESSION IN VITRO
It has been reported that FXa-dependent protease-activat-ed receptor (PAR)-1 and PAR-2 cleavage might play a role in tissue fibrosis and remodeling.13) Previous experiments in our laboratory demonstrated that the activation of fibroblast cardiac fibrosis is stimulated by FXa signaling via activator protein (AP)-1 and the nuclear factor kappa-light-chain-enhancer of the activated B cell (NF-κB) pathway, accompanied by reac-tive oxygen species (ROS) elevation and fibrotic signal activa-tion in growth-arrested fibroblasts.14) Factor Xa (FXa) inhibi-tors are anticoagulants to prevent and treat stroke and other thromboembolic events. Rivaroxaban, an approved oral direct FXa inhibitor, has been evaluated in a number of clinical set-tings to prevent and treat venous thromboembolism and stroke prophylaxis in atrial fibrillation (AF).15-17) However, there is a limited amount of evidence to substantiate the pleiotropic ef-fects of direct FXa inhibitors. The aim of this study was to in-vestigate whether rivaroxaban treatment attenuates cell prolif-eration and migration in Ang II induced mouse cardiac derived fibroblasts.
Methods
Reagents: Rivaroxaban was obtained from Toronto Research Chemicals (Ontario, Canada). Ang II was purchased from Sig-ma-Aldrich (St. Louis, MO, USA). MTT assay kits were from Roche (Basel, Switzerland). Cell migration assay kits were ob-tained from Platypus Technologies, LLC (Madison, WI, USA). Mouse cytokine array kits and TIMP-1 ELISA kits were from R&D Systems (Minneapolis, MN, USA). Diff-Quick stain was obtained from SYSMEX (Hyogo, Japan). The Cignal Finder 45-pathway Reporter Array and Attractene were from Qiagen (Dusseldorf, Germany). The Dual-Luciferase Reporter Assay Systems were obtained from Promega Corp. (Madison, WI, USA). All other reagents used were of analytical grade.Cell cultures: Cardiac derived fibroblasts were isolated from 1-day-old C57/BL/6 mice by the methods previously de-scribed 18) with minor modification. In brief, the ventricles were minced and digested using a collagenase/pancreatin mixture at 37°C for 30 minutes. The digested fractions were filtered through a sterile 70 μm nylon mesh. The pooled cells were re-suspended in alpha-modified Eagle’s medium (α-MEM) with 10% fibroblast growth factor. The cells were incubated twice at 37°C for 30 minutes using different two flasks. The cells were maintained in α-MEM supplemented with 10% fetal calf serum (FCS). Unless stated otherwise, the cells were starved in Hank’s balanced salt solution (HBSS) and subsequently stimu-lated as described.Rivaroxaban pretreatment: Rivaroxaban was dissolved in dimethyl sulfoxide (DMSO), and diluted 100 times with α-MEM with 1% FCS and 1% streptomycin. The final concen-tration of DMSO in culture media was 1%. CFs without rivar-oxaban pretreatment (control) were treated with α-MEM with 1% FCS, 1% streptomycin, and 1% DMSO under the same conditions as rivaroxaban pretreatment.Cell migration assay: Cell migration was determined using an Oris Pro Cell migration assay kit as described previously.19) CFs were pretreated with rivaroxaban (0.1 μg/mL) for 4 hours during adherence (n = 5). The cells were fixed and stained us-ing Diff-Quick staining. Images were captured immediately and at 24 hours after Ang II stimulation (10-8 M). The valua-
tion was taken as the area which a cell occupies using an imag-ing analysis system (Image J, National Institutes of Health, Bethesda, MD, USA).Proliferation assay (MTT Assay): Cells seeded at a density of 2.5 × 104/cm2 in 96-well plates were pretreated with or without rivaroxaban (5, 1, 0.1, and 0.01 μg/mL) for 4 hours, and then stimulated by Ang II (10-8 M) in 12 and 24 hours (n = 5). Cell proliferation was determined as the indicated intervals using MTT assay as described before.20)
Mouse cytokine array: Comprehensive cytokine and chemok-ine productions were evaluated by Mouse Cytokine Array Pan-el A Array after 18 hours of Ang II stimulation, as described previously.21) CFs were plated at 2.5 × 104 cells/mL in 96-well plates, pretreated either with or without rivaroxaban (0.1 μg/mL), and stimulated by Ang II (10-8 M). After the treatment, the supernatants were harvested and analyzed using the cy-tokine panel A array according to the manufacturer’s protocol. Briefly, the array membranes were blocked with blocking buff-er at room temperature for 1 hour, and media and the detection antibody cocktail were added and incubated at 4°C overnight. After washing 3 times with 2 mL of wash buffer at room tem-perature, streptavidin-HRP was added to each membrane and incubated at room temperature for 30 minutes. After washing, the cytokines were detected by a chemiluminescence reaction. We exposed wrapped membranes to X-ray film for 4 minutes and analyzed the array image file using image analysis soft-ware.TIMP-1 ELISA: After the pretreatment with rivaroxaban (0.01, 0.1, 1, and 5 μg/mL) for 4 hours, tissue inhibitors of metalloproteinases (TIMP)-1 production were evaluated by ELISA using the supernatant of mouse CFs after incubation with Ang II (10-8 M). The ELISA immunoassay was performed according to the manufacturer’s protocol. Briefly, the wells were precoated with a monoclonal antibody specific for TIMP-1. The standards, controls, and samples were added, and the samples were incubated for 2 hours. After washing, an en-zyme-linked polyclonal antibody specific for TIMP-1 was added. After another wash, the samples were incubated with a substrate solution for 30 minutes, and the color reaction was stopped by the addition of a stop solution. The optical density of each well was measured immediately with a microplate reader at 450 nm.Signal pathway reporter assay: The signal pathways were evaluated using the signal pathway reporter assay. CFs were seeded in 96-well cell culture plates and transfected with the signal reporter array panel of transcription factors. The tran-scription factors used for the 45 signaling pathways were the following: activating transcription factor (ATF)2/3/4, androgen receptor, nuclear factor (Nrf) 2 /Nrf1, ATF6, CCAAT-enhanc-er-binding proteins, cyclic adenosine monophosphate (cAMP) response enhancer-binding proteins, E2F, p53, early growth re-sponse protein 1, CBF/NF-Y/YY1, estrogen receptor, GATA, glucocorticoid receptor, heat shock transcription factor-1, met-al regulatory transcription factor-1, Gli, hepatocyte nuclear factor-4, hypoxia inducible factor-1a, interferon regulatory fac-tor 1, signal transducer and activator of transcription (STAT)1/STAT2, STAT1, Krüppel-like factor 4, liver X receptor, serum response factor (SRF)/Elk-1, AP-1, monocyte enhancer fac-tor-2, c-Myc, Nanog, recombining binding protein Suppressor of Hairless, NF-κB, octamer-binding transcription factor 4, Pax6, FOXO, nuclear factor of activated T cells, peroxisome
546Int Heart J
September 2015HASHIKATA, ET AL
proliferator-activated receptor, progesterone receptor, retinoic acid receptor, retinoid X receptor, Sox2, SP1, STAT3, SMAD2/3/4, vitamin D receptor, TCF/LEF, and aryl hydrocar-bon receptor. In each well, a pathway-specific transcription factor was transfected along with renilla luciferase (constitu-tively expressing the renilla construct, 20:1) using Attractene. Sixteen hours after the transfection, the cells were stimulated with Ang II (10-8 M), and then dual luciferase assays were per-formed as described previously.22)
Statistical analysis: All results represented experiments from a minimum of three separate preparations. Data are expressed as the mean ± SE unless otherwise indicated. Comparisons were performed using Student’s t test or a one-way analysis of vari-ance followed by Bonferroni correction where appropriate. A P value of ≦ 0.05 was considered to be statistically signifi-cant.
Results
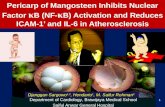
Cell migration and proliferation: Cardiac fibroblast (CF) mi-gration was evaluated 24 hours after the addition of Ang II. Only Ang II stimulation led to augmented cell migration in mouse CFs. Cell migration was significantly decreased by 73% in rivaroxaban induced cells compared to without rivar-oxaban (Figure 1A). Ribvaroxaban pretreatment by itself did not augment cell proliferation and migration (data not shown).
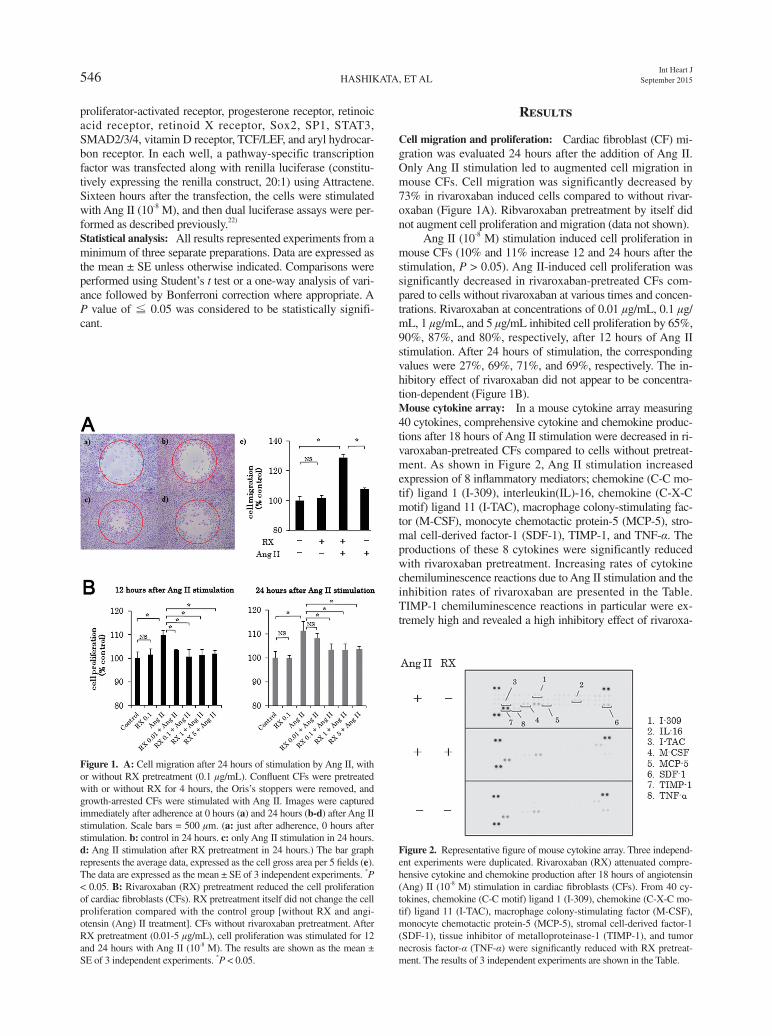
Ang II (10-8 M) stimulation induced cell proliferation in mouse CFs (10% and 11% increase 12 and 24 hours after the stimulation, P > 0.05). Ang II-induced cell proliferation was significantly decreased in rivaroxaban-pretreated CFs com-pared to cells without rivaroxaban at various times and concen-trations. Rivaroxaban at concentrations of 0.01 μg/mL, 0.1 μg/mL, 1 μg/mL, and 5 μg/mL inhibited cell proliferation by 65%, 90%, 87%, and 80%, respectively, after 12 hours of Ang II stimulation. After 24 hours of stimulation, the corresponding values were 27%, 69%, 71%, and 69%, respectively. The in-hibitory effect of rivaroxaban did not appear to be concentra-tion-dependent (Figure 1B).Mouse cytokine array: In a mouse cytokine array measuring 40 cytokines, comprehensive cytokine and chemokine produc-tions after 18 hours of Ang II stimulation were decreased in ri-varoxaban-pretreated CFs compared to cells without pretreat-ment. As shown in Figure 2, Ang II stimulation increased expression of 8 inflammatory mediators; chemokine (C-C mo-tif) ligand 1 (I-309), interleukin(IL)-16, chemokine (C-X-C motif) ligand 11 (I-TAC), macrophage colony-stimulating fac-tor (M-CSF), monocyte chemotactic protein-5 (MCP-5), stro-mal cell-derived factor-1 (SDF-1), TIMP-1, and TNF-α. The productions of these 8 cytokines were significantly reduced with rivaroxaban pretreatment. Increasing rates of cytokine chemiluminescence reactions due to Ang II stimulation and the inhibition rates of rivaroxaban are presented in the Table. TIMP-1 chemiluminescence reactions in particular were ex-tremely high and revealed a high inhibitory effect of rivaroxa-
Figure 1. A: Cell migration after 24 hours of stimulation by Ang II, with or without RX pretreatment (0.1 μg/mL). Confluent CFs were pretreated with or without RX for 4 hours, the Oris’s stoppers were removed, and growth-arrested CFs were stimulated with Ang II. Images were captured immediately after adherence at 0 hours (a) and 24 hours (b-d) after Ang II stimulation. Scale bars = 500 μm. (a: just after adherence, 0 hours after stimulation. b: control in 24 hours. c: only Ang II stimulation in 24 hours. d: Ang II stimulation after RX pretreatment in 24 hours.) The bar graph represents the average data, expressed as the cell gross area per 5 fields (e). The data are expressed as the mean ± SE of 3 independent experiments. *P < 0.05. B: Rivaroxaban (RX) pretreatment reduced the cell proliferation of cardiac fibroblasts (CFs). RX pretreatment itself did not change the cell proliferation compared with the control group [without RX and angi-otensin (Ang) II treatment]. CFs without rivaroxaban pretreatment. After RX pretreatment (0.01-5 μg/mL), cell proliferation was stimulated for 12 and 24 hours with Ang II (10-8 M). The results are shown as the mean ± SE of 3 independent experiments. *P < 0.05.
Figure 2. Representative figure of mouse cytokine array. Three independ-ent experiments were duplicated. Rivaroxaban (RX) attenuated compre-hensive cytokine and chemokine production after 18 hours of angiotensin (Ang) II (10-8 M) stimulation in cardiac fibroblasts (CFs). From 40 cy-tokines, chemokine (C-C motif) ligand 1 (I-309), chemokine (C-X-C mo-tif) ligand 11 (I-TAC), macrophage colony-stimulating factor (M-CSF), monocyte chemotactic protein-5 (MCP-5), stromal cell-derived factor-1 (SDF-1), tissue inhibitor of metalloproteinase-1 (TIMP-1), and tumor necrosis factor-α (TNF-α) were significantly reduced with RX pretreat-ment. The results of 3 independent experiments are shown in the Table.
547Vol 56No 5 RIVAROXABAN INHIBITS FIBROTIC PROGRESSION IN VITRO
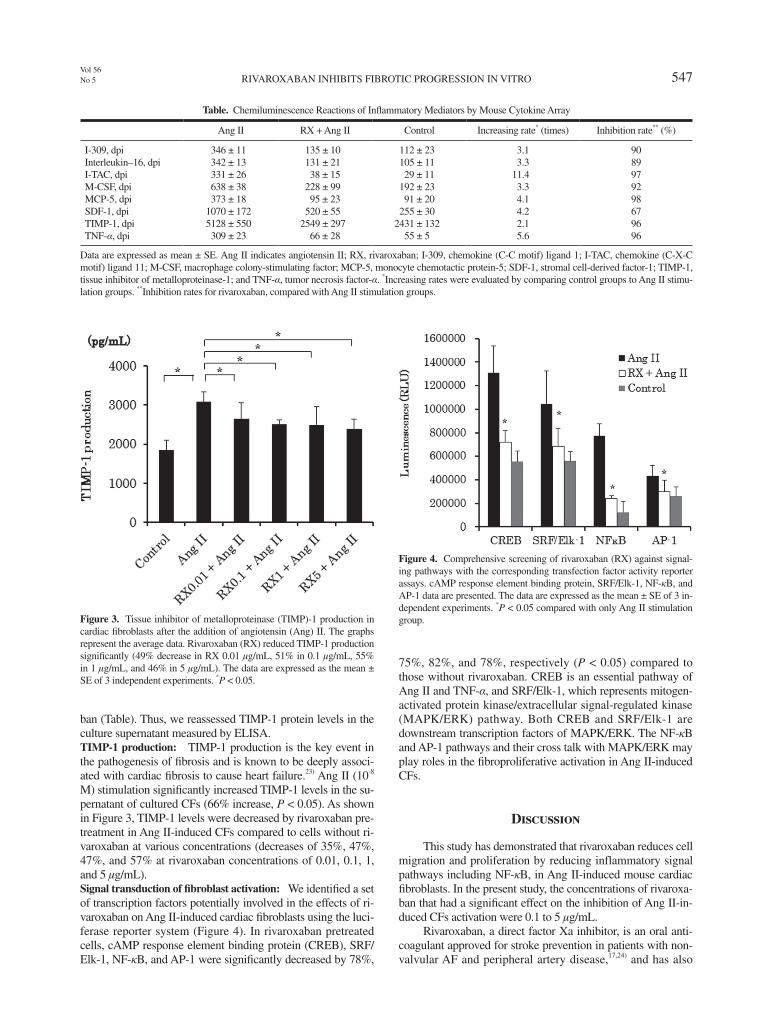
ban (Table). Thus, we reassessed TIMP-1 protein levels in the culture supernatant measured by ELISA.TIMP-1 production: TIMP-1 production is the key event in the pathogenesis of fibrosis and is known to be deeply associ-ated with cardiac fibrosis to cause heart failure.23) Ang II (10-8 M) stimulation significantly increased TIMP-1 levels in the su-pernatant of cultured CFs (66% increase, P < 0.05). As shown in Figure 3, TIMP-1 levels were decreased by rivaroxaban pre-treatment in Ang II-induced CFs compared to cells without ri-varoxaban at various concentrations (decreases of 35%, 47%, 47%, and 57% at rivaroxaban concentrations of 0.01, 0.1, 1, and 5 μg/mL).Signal transduction of fibroblast activation: We identified a set of transcription factors potentially involved in the effects of ri-varoxaban on Ang II-induced cardiac fibroblasts using the luci-ferase reporter system (Figure 4). In rivaroxaban pretreated cells, cAMP response element binding protein (CREB), SRF/Elk-1, NF-κB, and AP-1 were significantly decreased by 78%,
75%, 82%, and 78%, respectively (P < 0.05) compared to those without rivaroxaban. CREB is an essential pathway of Ang II and TNF-α, and SRF/Elk-1, which represents mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK) pathway. Both CREB and SRF/Elk-1 are downstream transcription factors of MAPK/ERK. The NF-κB and AP-1 pathways and their cross talk with MAPK/ERK may play roles in the fibroproliferative activation in Ang II-induced CFs.
Discussion
This study has demonstrated that rivaroxaban reduces cell migration and proliferation by reducing inflammatory signal pathways including NF-κB, in Ang II-induced mouse cardiac fibroblasts. In the present study, the concentrations of rivaroxa-ban that had a significant effect on the inhibition of Ang II-in-duced CFs activation were 0.1 to 5 μg/mL.
Rivaroxaban, a direct factor Xa inhibitor, is an oral anti-coagulant approved for stroke prevention in patients with non-valvular AF and peripheral artery disease,17,24) and has also
Table. Chemiluminescence Reactions of Inflammatory Mediators by Mouse Cytokine Array
Ang II RX + Ang II Control Increasing rate* (times) Inhibition rate** (%)
I-309, dpi 346 ± 11 135 ± 10 112 ± 23 3.1 90Interleukin–16, dpi 342 ± 13 131 ± 21 105 ± 11 3.3 89I-TAC, dpi 331 ± 26 38 ± 15 29 ± 11 11.4 97M-CSF, dpi 638 ± 38 228 ± 99 192 ± 23 3.3 92MCP-5, dpi 373 ± 18 95 ± 23 91 ± 20 4.1 98SDF-1, dpi 1070 ± 172 520 ± 55 255 ± 30 4.2 67TIMP-1, dpi 5128 ± 550 2549 ± 297 2431 ± 132 2.1 96TNF-α, dpi 309 ± 23 66 ± 28 55 ± 5 5.6 96
Data are expressed as mean ± SE. Ang II indicates angiotensin II; RX, rivaroxaban; I-309, chemokine (C-C motif) ligand 1; I-TAC, chemokine (C-X-C motif) ligand 11; M-CSF, macrophage colony-stimulating factor; MCP-5, monocyte chemotactic protein-5; SDF-1, stromal cell-derived factor-1; TIMP-1, tissue inhibitor of metalloproteinase-1; and TNF-α, tumor necrosis factor-α. *Increasing rates were evaluated by comparing control groups to Ang II stimu-lation groups. **Inhibition rates for rivaroxaban, compared with Ang II stimulation groups.
Figure 3. Tissue inhibitor of metalloproteinase (TIMP)-1 production in cardiac fibroblasts after the addition of angiotensin (Ang) II. The graphs represent the average data. Rivaroxaban (RX) reduced TIMP-1 production significantly (49% decrease in RX 0.01 μg/mL, 51% in 0.1 μg/mL, 55% in 1 μg/mL, and 46% in 5 μg/mL). The data are expressed as the mean ± SE of 3 independent experiments. *P < 0.05.
Figure 4. Comprehensive screening of rivaroxaban (RX) against signal-ing pathways with the corresponding transfection factor activity reporter assays. cAMP response element binding protein, SRF/Elk-1, NF-κB, and AP-1 data are presented. The data are expressed as the mean ± SE of 3 in-dependent experiments. *P < 0.05 compared with only Ang II stimulation group.

548Int Heart J
September 2015HASHIKATA, ET AL
been approved in Europe to prevent recurrent ischemic events in patients with recent acute coronary syndromes.25) It also has been reported that rivaroxaban attenuates progression and pro-motes stability of advanced atherosclerotic lesions in hyperlipi-demic apolipoprotein E-deficient mice.26)
Ang II is the main active peptide in the RAS, regulating cellular and physiological responses in the cardiovascular sys-tem.8) Tissue culture and animal experiments have demonstrat-ed a variety of mechanisms by which Ang II, an effector of the RAS, may promote atherogenesis by induction of cell growth and proliferation, oxidative stress, adhesion, and chemoattract-ant molecules and cytokines. Ang II stimulates the production of ROS by inducing nicotinamide adenine dinucleotide oxi-dase.27) The enzymatically active flavoprotein subunit gp-91phox of nicotinamide adenine dinucleotide phosphate oxi-dase present on fibroblasts is chiefly responsible for Ang II– stimulated oxidative stress in vivo.28) In addition, ROS activated coagulant tissue factor (TF).29) Active TF binds factors VIIa and Xa, and cleaves and activates PAR-2, and soluble Xa me-diates PAR1.30,31) By blocking PAR-1 and PAR-2 with rivar-oxaban, the production of several inflammatory cytokines, such as TIMP-1, I-309, IL-16, IL-2, I-TAC, M-CSF, MCP-5, SDF-1, and TNF-α, were significantly inhibited in Ang II-in-duced CFs. These inflammatory mediators are expected to ac-celerate cell migration and proliferation in activated CFs. Moreover, these mediators are also known to activate transfor-mations of CFs to various types of cells in the heart. Among these 9 cytokines, TIMP-1, IL-16, and TNF-α are reported to be deeply associated with cardiac fibrosis. It has been also re-ported that TIMP-1 and TNF-α through growth stimulating signal pathways, such as the Wnt and NF-κB pathways, are as-sociated with tissue calcification.6)
TIMP-1 is a natural glycoprotein that can inhibit the acti-vation of matrix metalloproteinases (MMPs).32) A manifesta-tion of myocardial remodeling is an increase in ECM which results from dysregulated levels of collagen synthesis, and its degradation via the delicate balance maintained between ma-trix MMPs and TIMPs.33) Increased protein levels of TIMP-1 were observed in both hypertrophied and unloaded hearts, sug-gesting the influence of changes in TIMPs in cardiac remode-ling.23) Adenoviral overexpression of TIMP-1 in cultured cardi-ac fibroblasts revealed that TIMP-1 induced fibroblast proliferation and provoked a switch to a more activated myofi-broblast phenotype.34) TIMPs may stimulate cardiac fibroblast proliferation and phenotypic differentiation into myofibroblasts at the site of tissue injury, thereby contributing to the formation of a qualitative and mature collagen matrix.35) Reduction of TIMP-1 production by rivaroxaban pretreatment may cause not only proliferative inhibition but also suppress phenotypic change in Ang II-induced CFs.
We have previously reported that the activation of cardiac fibroblasts is stimulated by FXa signaling via AP-1 and NF-κB pathways, accompanied by increasing ROS production and fi-brotic signal activation through the ERK and JNK pathways.14) Substances such as ROS are capable of activating intracellular transcription factors such as AP-1 and NF-κB. The MAPK/ERK pathways also activate AP-1 and NF-κB, eventually in-ducing TIMP-1 expression.36) Additionally, TIMP-1 is upregu-lated by transcription factors including MAPK modulation.37) In the present study, we also demonstrated that rivaroxaban in-hibits all of these inflammatory signal pathways, including AP-
1, NF-κB, and MAPK/ERK (Figure 5).IL-16 has been shown to be a key mediator of several in-
flammatory, allergic, or infectious diseases.38-40) IL-16 was one of the first cytokines characterized with chemoattractant activi-ty for human T cells and therefore was originally designated as lymphocyte chemoattractant factor. IL-16 can stimulate the synthesis of proinflammatory cytokines, including IL-1, IL-6, and TNF-α in monocytic cells.41) IL-16 has been reported to mediate cardiac inflammation leading to progressive cardiac fi-brosis and LV stiffness.42) However, the signal pathways induc-ing IL-16 still remain unclear. On the other hand, TNF-α is known to be involved in cardiac fibrosis and remodeling. TNF-α evokes inflammation and may synergically activate IL-16-related signaling cascade in CFs mainly through the NF-κB.43)
In clinical settings, in addition to its classical role in the prevention or treatment of thromboembolic diseases, rivaroxa-ban was also evaluated in a phase III clinical trial in patients stabilized after acute coronary syndromes.25) From the analysis of the ROKET AF trial, a 14% reduction in the hazard for car-diovascular death or acute coronary syndrome was observed in patients without prior myocardial infarction taking rivaroxaban compared with that of warfarin.44) This report suggests that ri-varoxaban has an effect on the early phase of development of atherosclerosis. Although no report has demonstrated the treat-ment effect of rivaroxaban in heart failure (HF) directly, it was reported the clinical outcomes of rivaroxaban were similar in patients with and without HF, suggesting the efficacy and safe-ty of rivaroxaban extend to the patient population with HF and AF.45) Our data in the present in vitro study may support these clinical effects of rivaroxaban in cardiovascular disease. In the next stage of the study, a pressure overload animal experimen-tal model of HF could be effective to confirm the cardioprotec-tive and anti-fibroproliferative effect of rivaroxaban on HFpEF. Rivaroxaban reduced angiotensin II-induced cardiac fibroblast migration and proliferation, accompanied by reductions in the
Figure 5. Proposed model of signaling pathways involved in cardiac fi-brosis in vitro. RAS indicates renin-angiotensin system; ROS, reactive ox-ygen species; TF, tissue factor; Vlla, factor VIIa; Xa, factor Xa; PAR, pro-tease activated receptor; MAPK, mitogen activated protein kinase; ATP, adenosine triphosphate; cAMP, cyclic adenosine monophosphate; PKA, protein kinase A; pathway; NFκB, nuclear factor kappa B; AP, activator protein; TNF, tumor necrosis factor; TIMP, tissue inhibitors of metallopro-teinases; and IL, interleukin.

549Vol 56No 5 RIVAROXABAN INHIBITS FIBROTIC PROGRESSION IN VITRO
production of various inflammatory cytokines. In the present study, we have demonstrated the novel pleiotropic effects of the direct Xa inhibitor rivaroxaban in mouse cardiac derived fi-broblasts.
Disclosure
Conflict of interest: Dr. Minako Yamaoka-Tojo was partly supported by international grants from MSD K.K., Bayer Pharma, Daiichi-Sankyo, and Boehringer Ingelheim. Dr. Junya Ako received speaking honorarium from Bayer Pharma. Other authors have nothing to disclose regarding this manuscript.
Acknowledgments
We appreciate the technical assistance of Ms. Kazumi Nakazato of the Department of Cardiovascular Medicine, Ki-tasato University School of Medicine.
References
1. Quercioli A, Mach F, Montecucco F. Novel treatment strategy with direct renin inhibition against heart failure. Panminerva Med 2010; 52: 41-51. (Review)
2. Chen W, Frangogiannis NG. The role of inflammatory and fibro-genic pathways in heart failure associated with aging. Heart Fail Rev 2010; 15: 415-22. (Review)
3. Shyu KG, Wang BW, Chen WJ, Kuan P, Hung CR. Mechanism of the inhibitory effect of atorvastatin on endoglin expression induced by transforming growth factor-beta1 in cultured cardiac fibrob-lasts. Eur J Heart Fail 2010; 12: 219-26.
4. Fukui M, Goda A, Komamura K, et al. Changes in collagen me-tabolism account for ventricular functional recovery following be-ta-blocker therapy in patients with chronic heart failure. Heart Vessels (in press)
5. Olson ER, Shamhart PE, Naugle JE, Meszaros JG. Angiotensin II-induced extracellular signal-regulated kinase 1/2 activation is me-diated by protein kinase Cdelta and intracellular calcium in adult rat cardiac fibroblasts. Hypertension 2008; 51: 704-11.
6. Akat K, Borggrefe M, Kaden JJ. Aortic valve calcification: basic science to clinical practice. Heart 2009; 95: 616-23. (Review)
7. Simionescu A, Simionescu DT, Vyavahare NR. Osteogenic re-sponses in fibroblasts activated by elastin degradation products and transforming growth factor-beta1: role of myofibroblasts in vascular calcification. Am J Pathol 2007; 171: 116-23.
8. Jiang B, Xu S, Hou X, Pimentel DR, Cohen RA. Angiotensin II differentially regulates interleukin-1-beta-inducible NO synthase (iNOS) and vascular cell adhesion molecule-1 (VCAM-1) expres-sion: role of p38 MAPK. J Biol Chem 2004; 279: 20363-8.
9. Kim S, Iwao H. Molecular and cellular mechanisms of angiotensin II-mediated cardiovascular and renal diseases. Pharmacol Rev 2000; 52: 11-34. (Review)
10. Hara M, Sakata Y, Nakatani D, et al. Renin-angiotensin-aldoster-one system polymorphisms and 5-year mortality in survivors of acute myocardial infarction: a report from the Osaka Acute Coro-nary Insufficiency Study. Int Heart J 2014; 55: 190-6.
11. Fujita S, Shimojo N, Terasaki F, et al. Atrial natriuretic peptide ex-erts protective action against angiotensin II-induced cardiac re-modeling by attenuating inflammation via endothelin-1/endothelin receptor A cascade. Heart Vessels 2013; 28: 646-57.
12. Mann DL. Mechanisms and models in heart failure: A combinato-rial approach. Circulation 1999; 100: 999-1008. (Review)
13. Borensztajn K, Stiekema J, Nijmeijer S, Reitsma PH, Peppelen-
bosch MP, Spek CA. Factor Xa stimulates proinflammatory and profibrotic responses in fibroblasts via protease-activated recep-tor-2 activation. Am J Pathol 2008; 172: 309-20.
14. Kitasato L, Yamaoka-Tojo M, Hashikata T, et al. Factor Xa in mouse fibroblasts may induce fibrosis more than thrombin. Int Heart J 2014; 55: 357-61.
15. Eriksson BI, Borris LC, Friedman RJ, et al. Rivaroxaban versus enoxaparin for thromboprophylaxis after hip arthroplasty. N Engl J Med 2008; 358: 2765-75.
16. Kakkar AK, Brenner B, Dahl OE, et al. Extended duration rivar-oxaban versus short-term enoxaparin for the prevention of venous thromboembolism after total hip arthroplasty: a double-blind, ran-domised controlled trial. Lancet 2008; 372: 31-9.
17. Patel MR, Mahaffey KW, Garg J, et al. Rivaroxaban versus warfa-rin in nonvalvular atrial fibrillation. N Engl J Med 2011; 365: 883-91.
18. Harada M, Itoh H, Nakagawa O, et al. Significance of ventricular myocytes and nonmyocytes interaction during cardiocyte hyper-trophy: evidence for endothelin-1 as a paracrine hypertrophic fac-tor from cardiac nonmyocytes. Circulation 1997; 96: 3737-44.
19. Hulkower KI, Herber RL. Cell migration and invasion assays as tools for drug discovery. Pharmaceutics 2011; 3: 107-24.
20. Yamaoka-Tojo M, Ushio-Fukai M, Hilenski L, et al. IQGAP1, a novel vascular endothelial growth factor receptor binding protein, is involved in reactive oxygen species--dependent endothelial mi-gration and proliferation. Circ Res 2004; 95: 276-83.
21. Zhao W, Su W, Kuang P, et al. The role of hepatic stellate cells in the regulation of T-cell function and the promotion of hepatocellu-lar carcinoma. Int J Oncol 2012; 41: 457-64.
22. McNabb DS, Reed R, Marciniak RA. Dual luciferase assay sys-tem for rapid assessment of gene expression in Saccharomyces cerevisiae. Eukaryot Cell 2005; 4: 1539-49.
23. Xu R, Lin F, Zhang S, Chen X, Hu S, Zheng Z. Signal pathways involved in reverse remodeling of the hypertrophic rat heart after pressure unloading. Int J Cardiol 2010; 143: 414-23.
24. Jones WS, Hellkamp AS, Halperin J, et al. Efficacy and safety of rivaroxaban compared with warfarin in patients with peripheral ar-tery disease and non-valvular atrial fibrillation: insights from ROCKET AF. Eur Heart J 2014; 35: 242-9.
25. Mega JL, Braunwald E, Wiviott SD, et al. Rivaroxaban in patients with a recent acute coronary syndrome. N Engl J Med 2012; 366: 9-19.
26. Zhou Q, Bea F, Preusch M, et al. Evaluation of plaque stability of advanced atherosclerotic lesions in apo E-deficient mice after treatment with the oral factor Xa inhibitor rivaroxaban. Mediators Inflamm 2011: 2011; 432080.
27. Griendling KK, Minieri CA, Ollerenshaw JD, Alexander RW. An-giotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ Res 1994; 74: 1141-8.
28. Wang HD, Xu S, Johns DG, et al. Role of NADPH oxidase in the vascular hypertrophic and oxidative stress response to angiotensin II in mice. Circ Res 2001; 88: 947-53.
29. Muller I, Klocke A, Alex M, et al. Intravascular tissue factor initi-ates coagulation via circulating microvesicles and platelets. FASEB J 2003; 17: 476-8.
30. Camerer E, Trejo J. Cryptic messages: is noncoagulant tissue fac-tor reserved for cell signaling? Proc Natl Acad Sci U S A 2006; 103: 14259-60.
31. Borensztajn K, Peppelenbosch MP, Spek CA. Factor Xa: at the crossroads between coagulation and signaling in physiology and disease. Trends Mol Med 2008; 14: 429-40. (Review)
32. Khokha R, Waterhouse P. The role of tissue inhibitor of metallo-proteinase-1 in specific aspects of cancer progression and repro-duction. J Neurooncol 1994; 18: 123-7. (Review)
33. Berry MF, Woo YJ, Pirolli TJ, et al. Administration of a tumor necrosis factor inhibitor at the time of myocardial infarction atten-uates subsequent ventricular remodeling. J Heart Lung Transplant 2004; 23: 1061-8.
34. Lovelock JD, Baker AH, Gao F, et al. Heterogeneous effects of tissue inhibitors of matrix metalloproteinases on cardiac fibrob-

550Int Heart J
September 2015HASHIKATA, ET AL
lasts. Am J Physiol Heart Circ Physiol 2005; 288: H461-8. 35. Camelliti P, Borg TK, Kohl P. Structural and functional characteri-
sation of cardiac fibroblasts. Cardiovasc Res 2005; 65: 40-51. (Re-view)
36. del Zoppo GJ, Milner R, Mabuchi T, et al. Microglial activation and matrix protease generation during focal cerebral ischemia. Stroke 2007; 38: 646-51. (Review)
37. Reunanen N, Li SP, Ahonen M, Foschi M, Han J, Kähäri VM. Ac-tivation of p38 alpha MAPK enhances collagenase-1 (matrix met-alloproteinase (MMP)-1) and stromelysin-1 (MMP-3) expression by mRNA stabilization. J Biol Chem 2002; 277: 32360-8.
38. Keates AC, Castagliuolo I, Cruickshank WW, et al. Interleukin 16 is up-regulated in Crohn’s disease and participates in TNBS colitis in mice. Gastroenterology 2000; 119: 972-82.
39. Skundric DS, Cai J, Cruikshank WW, Gveric D. Production of IL-16 correlates with CD4+ Th1 inflammation and phosphorylation of axonal cytoskeleton in multiple sclerosis lesions. J Neuroin-flammation 2006; 3: 13.
40. Hessel EM, Cruikshank WW, Van Ark I, et al. Involvement of IL-16 in the induction of airway hyper-responsiveness and up-regula-
tion of IgE in a murine model of allergic asthma. J Immunol 1998; 160: 2998-3005.
41. Mathy NL, Bannert N, Norley SG, Kurth R. Cutting edge: CD4 is not required for the functional activity of IL-16. J Immunol 2000; 164: 4429-32.
42. Tamaki S, Mano T, Sakata Y, et al. Interleukin-16 promotes cardi-ac fibrosis and myocardial stiffening in heart failure with pre-served ejection fraction. PLoS One 2013; 8: e68893.
43. Marchant DJ, Boyd JH, Lin DC, Granville DJ, Garmaroudi FS, McManus BM. Inflammation in myocardial diseases. Circ Res 2012; 110: 126-44. (Review)
44. Mahaffey KW, Stevens SR, White HD, et al. Ischaemic cardiac outcomes in patients with atrial fibrillation treated with vitamin K antagonism or factor Xa inhibition: results from the ROCKET AF trial. Eur Heart J 2014; 35: 233-41.
45. van Diepen S, Hellkamp AS, Patel MR, et al. Efficacy and safety of rivaroxaban in patients with heart failure and nonvalvular atrial fibrillation: insights from ROCKET AF. Circ Heart Fail 2013; 6: 740-7.