
Mémoire de M1 SIMS (2014)
19
Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la contractilité de cellules cardiaques isolées de souris adultes Grimaud Joaquim 2013-2014 Master 1 SIMS Sous la direction du Dr. Noireaud Jacques, Directeur de Recherche Inserm Membres du jury Nom/Prénom 1 | Fonction Nom/Prénom 2 | Fonction Nom/Prénom 3 | Fonction Nom/Prénom 4 | Fonction Nom/Prénom 5 | Fonction Nom/Prénom 6 | Fonction Soutenu le : 19 juin 2014
-
Upload
joaquim-grimaud -
Category
Documents
-
view
221 -
download
0
description
Implication de la voie du TNF-a dans les effets de microparticules macrophagiques sur la contractilité de cellules cardiaques isolées de souris adultes
Transcript of Mémoire de M1 SIMS (2014)

Implication de la voie du TNF-α dans les effets de
microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de
souris adultes
Grimaud Joaquim
2013-2014 Master 1 SIMS
Sous la direction du Dr. Noireaud Jacques,
Directeur de Recherche Inserm Membres du jury
Nom/Prénom 1 | Fonction
Nom/Prénom 2 | Fonction
Nom/Prénom 3 | Fonction
Nom/Prénom 4 | Fonction
Nom/Prénom 5 | Fonction
Nom/Prénom 6 | Fonction
Soutenu le : 19 juin 2014

L’auteur du présent document vous
autorise à le partager, reproduire,
distribuer et communiquer selon
les conditions suivantes :
Vous devez le citer en l’attribuant de la manière indiquée par l’auteur (mais pas d’une
manière qui suggérerait qu’il approuve votre utilisation de l’œuvre).
Vous n’avez pas le droit d’utiliser ce document à des fins commerciales.
Vous n’avez pas le droit de le modifier, de le transformer ou de l’adapter.
Consulter la licence creative commons complète en français : http://creativecommons.org/licences/by-nc-nd/2.0/fr/
Ces conditions d’utilisation (attribution, pas d’utilisation
commerciale, pas de modification) sont symbolisées par les icônes positionnées en pied de page.

REM
ER
CIEM
EN
TS
Je tiens à remercier en premier lieu le Dr. Jacques Noireaud pour ses nombreux conseils, les connaissances et expériences qu’il m’a transmis, la confiance qu’il m’a accordée, son accessibilité, sa disponibilité, sa bonne humeur et sa gentillesse. Je tiens aussi à remercier le Dr. Ramaroson Andriantsitohaina de m’avoir accueilli au sein de son unité et sans qui l’occasion de découvrir le monde de la recherche ne m’aurait pas été donnée. J’adresse également mes remerciements à Badreddine Lahouel et Edward Milbank pour leurs nombreux conseils et remarques Je tiens enfin à remercier l’ensemble des membres de l’unité Inserm U1063 pour leur aide et leur disponibilité tout au long de ces 2 mois de stage.

Sommaire
INTRODUCTION : ........................................................................................................................ 1
1. Contexte général .................................................................................................................... 1 2. L’athérosclérose ..................................................................................................................... 1 3. Les microparticules ................................................................................................................. 2 4. Le couplage excitation-contraction ........................................................................................ 2 5. Objectif de l’étude .................................................................................................................. 3
MATERIEL ET METHODES ............................................................................................................ 4
1. Culture des macrophages RAW 264.7 et obtention des microparticules
macrophagiques .................................................................................................................................... 4 1.1. Culture cellulaire de la lignée RAW 264.7 ..................................................................................... 4 1.2. Obtention des microparticules ..................................................................................................... 4 2. Isolement des cardiomyocytes ............................................................................................... 4 2.1. Prélèvement des cœurs de souris C57BL/6 adultes ........................................................................ 4 2.2. Montage Langendorff et digestion enzymatique ............................................................................. 4 3. Analyse fonctionnelle : Mesure de la contraction et du transitoire calcique des
cardiomyocytes isolés ........................................................................................................................... 5 3.1. Marquage des cardiomyocytes par la sonde Fura-2AM ................................................................... 5 3.2. Systèmes d’enregistrement ......................................................................................................... 6 4. Analyse moléculaire ................................................................................................................ 6 4.1. Dosage des protéines ................................................................................................................. 6 4.2. Western-Blot ............................................................................................................................. 6 5. Analyses statistiques .............................................................................................................. 7
RESULTATS .................................................................................................................................. 7
1. Le TNFαest présent dans les MPs macrophagiques ................................................................ 7 2. TNF per se induit un effet inotrope négatif ........................................................................... 7 3. Le TNF-α n’a pas d’effet sur les transitoires calciques ........................................................... 8 4. Les MPS induisent un effet inotrope négatif ........................................................................... 9 5. Le SPD-304 inhibe l’effet des MPs macrophagiques ............................................................... 9
DISCUSSION ............................................................................................................................. 10
CONCLUSION............................................................................................................................. 10
REFERENCES BIBLIOGRAPHIQUES ........................................................................................... 11

Liste des abréviations:
Act D : Actinomycine D
Apo B : Apolipoprotéine B
ATP : Adénosine-5’-Triphosphate
BSA : Bovine Serum Albumine (Sérum d’albumine bovine)
Ca2+: Calcium
CICR : Calcium-Induced Calcium Release
DMSO: Diméthyl sulfoxyde
ESM : Erreur standard à la moyenne
HRP : Horseradish peroxydase (Peroxydase de raifort)
I-CAM : Intercellular Adhesion Molecule (Protéine d’adhésion intercellulaire)
ICaL : Canaux calciques potentiel-dépendants de type L (Long Lasting)
IFN-γ: Interféron gamma
IL-1β: Interleukine 1β
IL-8 : Interleukine 8
LDL : Low Density Lipoprotein (Lipoprotéine de basse densité)
M-CSF : Macrophage colony-stimulating Factor
MCP-1: Monocyte Chemoattractant Protein 1 (Protéine chimioattractante pour les monocytes)
MPs : Microparticules
NCX : Echangeur Sodium/Calcium
OMS: Organisation Mondiale de la Santé
oxLDL : Oxidized Low-Density Lipoprotein (Lipoprotéine de basse densité oxydée)
RPE : Résonance Paramagnétique Electronique
RS : Réticulum sarcoplasmique
RYR : Récepteur sensible à la ryanodine
SERCA : Sarco/Endoplasmic Reticulum Ca2+-ATPase
SVF: Sérum de veau foetal
TACE: TNF-α-converting enzyme (Enzyme de conversion du TNF-α)
TBS-Tween : Tris-buffered saline and Tween 20
TNF-α : Tumor necrosis factor alpha (Facteur de nécrose de tumeur alpha)
TNF-R1/2 : Récepteurs au TNF-α de type 1 et 2
V-CAM 1 : Vascular Cell Adhesion Protein 1 (Protéine-1 d’adhésion vasculaire)

mots-clés : Athérosclérose, macrophage, microparticule, TNF-α, cardiomyocyte, souris adulte
Keywords : Atherosclerosis, microparticle, TNF-α, cardiomyocyte, adult mouse.
AB
ST
RA
CT
In case of atherosclerosis, inflammation in the coronary arteries involves the activation of monocytes/ macrophages and the release of microparticles (MPs) in the bloodstream. Thus, these MPs can come close to myocardial cardiomyocytes. Moreover, an important expression of tumor necrosis factor (TNF)-α by macrophages of atherosclerotic plaque was highlighted, which makes possible that these MPs carry this cytokine and thus are able to activate the TNF receptors of type 1 and 2 on the cardiomyocytes. A preliminary study in cardiomyocytes isolated from adult mice, demonstrated that MPs from the macrophage RAW 264.7 have the effect of reducing and slowing the shortening of sarcomeres (negative inotropic effect) without affecting calcium transients (i.e. the release of Ca2+ from the sarcoplasmic reticulum). The objective of this study was to determine whether TNF-α is actually carried by MPs and if so, if he plays a role in the observed effects of MPs on the contractile performance of adult mouse cardiomyocytes. The experimental approach combines a method of functional analysis, i.e. the simultaneous analysis of sarcomeres contraction and calcium transients, and a method of molecular biology (i.e. Western blot) to detect the presence of TNF-α in the MPs. The results show the presence of TNF-α in the MPs from the macrophages and that TNF-α has a negative inotropic effect on the contraction of cardiomyocytes. Moreover, the negative inotropic effect of MPs from macrophages on cardiomyocyte contractility is removed after a pretreatment with SPD-304 (10 μM), a specific and irreversible inhibitor of TNF-α receptors. Thus, the TNF-α pathway could be involved in the effect of MPs from macrophages on contractility of cardiomyocytes. Now, it would be interesting to confirm the involvement of TNF-α carried by these MPs by studying various signaling pathways of TNF-α, such as NO/ cGMP / PKG, phospholipase A2/arachidonic acid and sphingosines pathway.
RÉS
UM
É Dans le cas de l’athérosclérose, l’inflammation au niveau des artères coronaires implique l’activation des
monocytes/macrophages et la libération de microparticules (MPs) dans la circulation sanguine, qui peuvent ainsi venir à proximité des cardiomyocytes myocardique. Par ailleurs, une expression importante du facteur de nécrose tumoral (TNF)-α par les macrophages de la plaque d’athérome a été mise en évidence, ce qui rend fortement probable que les MPs issues de ces derniers puissent porter cette cytokine et activer ainsi les récepteurs TNF de type 1 et 2 portés par les cardiomyocytes. Une étude préliminaire réalisée dans des cardiomyocytes isolés de souris adulte, a permis de montrer que des MPs macrophagiques issues de la lignée RAW 264.7, ont pour effet de diminuer et de ralentir le raccourcissement des sarcomères (effet inotrope négatif), sans affecter les transitoires calciques (i.e. la libération de Ca2+ du réticulum sarcoplasmique). L’objectif de cette étude a donc été de déterminer si le TNF-α est effectivement porté par les MPs et si oui, s’il joue un rôle dans les effets observés des MPs sur la performance contractile des cardiomyocytes murins adultes. L’approche expérimentale utilisée combine une méthode d’étude fonctionnelle, i.e. l’analyse concomitante de la contraction des sarcomères et des transitoires calciques, ainsi qu’une méthode de biologie moléculaire (i.e. western-blot) permettant de détecter la présence éventuelle de TNF-α dans les MPs. Les résultats obtenus montrent tout d’abord la présence de TNF-α dans les MPs macrophagiques et que le TNF-α a un effet inotrope négatif sur la contraction des cardiomyocytes. De plus, l’effet inotrope négatif des MPs macrophagiques sur la
contractilité cardiomyocytaire est supprimé suite à un prétraitement par le SPD-304 (10 M), inhibiteur spécifique et irréversible des récepteurs au TNF-α. La voie du TNF-α pourrait donc bien être impliquée dans l’effet des MPs macrophagiques sur la contractilité des cardiomyocytes. Il serait maintenant intéressant de confirmer l’implication du TNF-α porté par ces MPs en explorant divers voies de signalisation du TNF-α, comme les voies NO/cGMP/PKG, phospholipase A2/acide arachidonique, sphingosines.
Présidence de l'université
40 rue de rennes – BP 73532
49035 Angers cedex
Tél. 02 41 96 23 23 | Fax 02 41 96 23 00

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 1
INTRODUCTION :
1. Contexte général
Depuis ces 40 dernières années, de nombreux
progrès ont été réalisés dans la prévention, le
diagnostic et le traitement des maladies
cardiovasculaires (Braunwald et Bristow, 2000).
Toutefois, malgré ces grandes avancées, les
maladies cardiovasculaires continuent de représenter
aujourd’hui une des principales causes de mortalité
dans le monde (World Health Organization, 2007).
L’Organisation Mondiale de la Santé (OMS) a estimé
que 17 millions de décès en 2008 étaient imputables
aux pathologies cardiovasculaires, soit environ un
tiers de la mortalité mondiale totale (World Health
Organization, 2011). Le diagnostic et la prise en
charge de ces pathologies sont difficiles et ces
dernières ont un impact économique important
(Bloom et al., 2011). Les projections des
épidémiologistes indiquent que les maladies
cardiovasculaires, notamment d’origine ischémique,
devraient rester l’une des principales causes de
mortalité au moins jusqu’à 2030 (Beaglehole et
Bonita, 2008).
2. L’athérosclérose
Il s’agit d’une pathologie cardiovasculaire chronique
très répandue qui s’amplifie avec l’âge. Elle est sous-
jacente à des événements ischémiques coronariens
(infarctus du myocarde) et cérébrovasculaires
(accident vasculaire cérébral). Elle est caractérisée
par le dépôt, dans les artères, d’une plaque
d’athérome essentiellement composée de lipides
riches en cholestérol enveloppée dans une sclérose,
i.e. une gaine fibreuse formée par l’épaississement
de la paroi artérielle (Lusis, 2000). L’OMS l’a définie
comme étant une association variable de
remaniements de l’intima des artères de moyen et de
gros calibre, consistant en une accumulation locale
de lipides, de glucides complexes, de sang et de
produits sanguins, de tissus fibreux et de dépôts
calcaires. Le tout s’accompagne de modifications de
la média. Cette pathologie a longtemps été
considérée uniquement comme un dépôt passif de
lipides, mais aujourd’hui l’athérosclérose est
également définie comme une maladie inflammatoire
chronique entraînée par les lipides, en particulier les
LDLs (« low density lipoproteins ») contenant
l’apolipoprotéine (Apo) B, et les leucocytes (Swirski
et Nahrendorf, 2013). L’athérosclérose peut aussi
toucher des artères de plus petites tailles, comme les
artères coronaires (Libby et al., 2002).
L’accumulation de lipides, et notamment de LDL
oxydés (oxLDL) dans les artères entre l’endothélium
et la lame élastique interne induit une augmentation
de l’expression de certaines molécules d’adhésion
sur la surface endothéliale des vaisseaux, dont la V-
CAM 1 (« Vascular Cell Adhesion Protein 1 »), l’ICAM
(« Intercellular Adhesion Molecule ») et la P-Sélectine
(Poston et Johnson-Tidey, 1996). Ces molécules
permettent l’adhésion de monocytes sanguins
circulants qui d’ordinaire ne se fixent pas
(Blankenberg et al., 2003). Ensuite, sous l’effet de
facteurs chimiotactiques, dont la MCP-1 (« Monocyte
Chemoattractant Protein 1 ») et l’interleukine 8 (IL-8),
les monocytes adhérents pénètrent l’intima à travers
les jonctions inter-endothéliales. Ces facteurs sont
fortement exprimés par les cellules endothéliales de
la plaque d’athérome. Les monocytes se différencient
ensuite en macrophages sous l’action du M-CSF
(« Macrophage colony-stimulating factor ») sécrété
par les cellules endothéliales. Ces derniers expriment

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 2
le récepteur Apo B/E et jouent un rôle « d’éboueurs »
(« scavenger ») en phagocytant les oxLDL accumulés
dans l’espace sous-endothélial. Les macrophages se
chargent donc en cholestérol, ce qui conduit à leur
transformation en cellules dites « spumeuses » qui
s’accumulent. Cette accumulation engendre une
réaction inflammatoire chronique et des lymphocytes
T sont à leur tour recrutés dans l’intima. Les
lymphocytes T libèrent de l’interféron (IFN)-γ et du
TNF (« tumor necrosis factor »)-α/β qui stimulent les
cellules spumeuses qui vont à leur tour libérer des
cytokines pro-inflammatoires et/ou pro-apoptotiques
telles que le TNF-α et l’IL-1β. Ces cytokines
renforcent le caractère pro-inflammatoire de la plaque
d’athérome et ont une action autocrine sur les
macrophages transformés en cellules spumeuses.
Elles les activent et induisent la libération de
microparticules (MPs) (Spagnoli et al., 2007).
3. Les microparticules
Les MPs sont dérivées de fragments de membrane
plasmique issue de la cellule qui les sécrète. Leur
taille varie de 0,1 à 1μm de diamètre. Elles peuvent
se retrouver libérées dans le milieu extracellulaire
suite à une activation cellulaire ou à un phénomène
apoptotique (Biro et al., 2007; Sewify et al., 2013).
Au départ décrites comme des « déchets
cellulaires », « platelet dust » (Wolf, 1967), elles sont
aujourd’hui considérées comme de véritables
vecteurs biologiques. En effet, elles portent aussi
bien des marqueurs membranaires que différents
constitutants du cytoplasme ou du noyau (ADN,
ARNm ou miRNA). Les stimuli à l’origine de la
production de microparticules sont multiples et leur
contenu est très varié (Budaj et al., 2012; Cocucci
et al., 2009; Distler et al., 2005). En théorie, toutes
les cellules peuvent en produire, mais les modèles
les plus utilisés pour leur étude sont ceux qui sont le
plus facilement collectables, c'est-à-dire les cellules
sanguines circulantes telles que les plaquettes, les
lymphocytes et les monocytes. La sécrétion de MPs
est souvent liée à une apoptose déclenchée soit par
un stress oxydatif important, de grandes forces de
cisaillement « shear stress » ou des lésions.
Toutefois, la sécrétion de MPs peut aussi être
déclenchée par une activation de la cellule. C’est par
exemple le cas des plaquettes et des cellules
endothéliales (Jenkins et al., 2013). Une fois
libérées, elles pourront soit intéragir avec différents
ligands portés par leurs cellules cibles et ainsi activer
différentes cascades de signalisation, soit transférer
certains de leurs constituants via des phénomènes
d’internalisation ou de fusion membranaire (Hugel et
al., 2005). Les MPs permettent ainsi à leurs cellules
cibles d’acquérir de nouvelles propriétés et fontions
biologiques. Leur implication a été démontrée dans
de nombreux processus physiopathologiques tels que
la thrombose, les diabètes, l’inflammation,
l’angiogenèse, le processus tumoral et
l’athérosclérose (Freyssinet, 2003 ; Piccin, 2007).
4. Le couplage excitation-
contraction
Le couplage excitation-contraction cardiaque est un
processus qui englobe la stimulation électrique des
cardiomyocytes ventriculaires (i.e. du myocarde) et
ainsi la contraction du cœur permettant d’assurer une
distribution sanguine dans tout l’organisme. Les
cardiomyocytes ventriculaires contiennent des
protéines myofibrillaires longitudinales regroupées en
sarcomères, donnant cet aspect strié à la cellule
cardiaque observée en microscopie optique. Le
raccourcissement simultané de ces sarcomères au
niveau cellulaire entraîne le phénomène de

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 3
contraction myocardique. Le calcium (Ca2+) est le
messager ubiquitaire essentiel pour cette activité
contractile. La propagation d’un potentiel d’action,
initié au niveau sinusal auriculaire cardiaque, le long
du sarcolemme et du système tubulaire transverse
des cardiomyocytes induit une entrée de Ca2+ dans le
cardiomyocyte via l’ouverture des canaux calciques
potentiel-dépendants de type L (Long Lasting) (ICaL).
Cette entrée d’ions Ca2+ va activer des récepteurs-
canaux sensibles à la ryanodine (RYR) qui vont
permettre une sortie massive de Ca2+ du RS, i.e. le
transitoire calcique. Cette sortie de Ca2+ du RS est
amplifiée par un rétrocontrôle positif autocatalytique,
le mécanisme de CICR (« Calcium-induced Calcium
Release ») (Lee et Keener, 2008). La concentration
calcique passe de 10-8 à 10-6 M. Le Ca2+ induit
l’activation des myofibrilles et ainsi le
raccourcissement sarcomérique, i.e. la contraction.
La concentration de Ca2+ cytosolique doit ensuite
diminuer pour permettre la relaxation de l’appareil
contractile. Le Ca2+ est ainsi principalement (chez la
souris 93%) repompé par la pompe ATPasique
SERCA (« Sarco/Endoplasmic Reticulum Ca2+-
ATPase ») du RS, et aussi (7%) expulsé par
l’échangeur Sodium/Calcium NCX et la Ca2+ ATPase
du sarcolemme vers le milieu extracellulaire (Bers,
2002).
5. Objectif de l’étude
Dans le cas de l’athérosclérose, l’inflammation au
niveau des artères coronaires implique l’activation
des monocytes/macrophages et la libération de MPs
dans la circulation sanguine, qui ainsi peuvent venir à
proximité des cardiomyocytes myocardique. Par
ailleurs, une expression importante du TNF-α par les
macrophages de la plaque d’athérome a été mise en
évidence, ce qui rend fortement probable que les
MPs issues de ces derniers puisse porter cette
cytokine (Martinez et al., 2005 ; Tedgui et Mallat,
2006). TNF-α est une protéine impliquée dans
l’inflammation systémique et appartient à un groupe
de cytokines qui stimulent la réaction inflammatoire
aiguë. Il existe deux formes de TNF-α, une forme
transmembranaire et une forme libre (Horiuchi et al.,
2010). Le TNF-α transmembranaire (26 kDa) est un
précurseur de la forme soluble du TNF-α (17 kDa), on
le retrouve exprimé par les macrophages activés et il
peut-être clivé sous l’action d’une « TNF-α-converting
enzyme » (TACE) pour générer la forme soluble du
TNF-α. Cette forme soluble libre est capable de se
lier aux récepteurs TNF de type 1 et 2 (TNF-R1 et
TNF-R2) portés par les cardiomyocytes.
Une étude préliminaire réalisée dans des
cardiomyocytes isolés de souris adulte, a permis de
montrer que des MPs macrophagiques issues de la
lignée RAW 264.7, ont pour effet de diminuer et de
ralentir le raccourcissement des sarcomères (effet
inotrope négatif), sans affecter les transitoires
calciques (Milbank et al., 2014). L’objectif de cette
étude a donc été de déterminer si le TNF-α est
effectivement porté par les MPs et si oui, s’il joue un
rôle dans les effets observés des MPs sur la
performance contractile des cardiomyocytes murins
adultes. Un inhibiteur spécifique et irréversible des
récepteurs aux TNF-α, le SPD-304 (10μM ;
Calbiochem) a été utilisé afin de vérifier cette
implication du TNF-α.

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 4
Matériel et méthodes
1. Culture des macrophages
RAW 264.7 et obtention
des microparticules
macrophagiques
1.1. Culture cellulaire de la lignée RAW 264.7
RAW 264.7 est une lignée de macrophages obtenue
à partir de souris (Mus Musculus) BALB/c (American
Type Culture Collection, Manasses, VA, USA). Ces
cellules ont été par la suite mises en culture dans un
milieu DMEM (Lonza), additionné de 10% de sérum
de veau fœtal (SVF) décomplémenté, de 1% de
Pénicilline/Streptomycine, et de 1% de L-gluthamine.
Les cellules ont été placées dans une atmosphère
humidifiée à 5% de CO2 et à 37°C. Le milieu a été
changé toutes les 48 heures, lorsque les cellules
étaient à 90% de confluence environ. Les
macrophages RAW 264.7 utilisés dans cette étude
pour produire des microparticules l’ont été entre des
repiquages R3 et R10.
1.2. Obtention des microparticules
La lignée cellulaire de macrophages RAW 264.7 a
donc été utilisée pour la production de MPs. Les
cellules ont été réensemencées à une densité de
2.106 cellules/mL puis mises en culture comme décrit
ci-dessus. La production de MPs a été déclenchée
par traitement durant 24 heures avec soit avec de
l’actinomycine D (ActD, 1μg/mL), soit avec de
l’interleukine-1β (IL-1β ; 10ng/mL) seul, soit du TNF-α
(5ng/mL) seul, soit du TNF-α et de l’IL-1β combinés.
Le traitement le plus efficace pour induire la
production de MPs est l’ActD, qui induit l’apoptose
des macrophages. Quatre types de MPs sont ainsi
produites selon le type de stimulation. Un surnageant
a été obtenu par deux centrifugations de 750 g
pendant 15 minutes, et de 1500 g pendant 5 minutes
afin d’éliminer les cellules et les différents débris
cellulaires. Les MPs contenues dans le surnageant
ont été isolées grâce à deux centrifugations
successives pendant 30 minutes à 13 000 g, puis
deux fois 45 min à 13 000 g. Les MPs contenues
dans le surnageant ont été récupérées dans du NaCl
à 0,9 % et conservées à 4°C jusqu’à leur utilisation.
2. Isolement des
cardiomyocytes
2.1. Prélèvement des cœurs de souris C57BL/6 adultes
Les souris utilisées pour l’isolement des
cardiomyocytes ventriculaires sont des mâles de
souche C57BL/6. Elles ont été soumises à un régime
standard et furent utilisées à l’âge de 14 semaines.
Les cardiomyocytes ventriculaires ont été isolés par
dissociation mécanique comme décrit précédemment
(Pinz et al, 2011). Les souris sont soumises à un
prétraitement par de l’héparine (Héparine Choay,
Sanofi Aventis, Paris, 0,05 ml par voie
intrapéritonéale). L’héparine permet d’éviter toute
coagulation sanguine et éventuel infarctus. Après
vingt minutes, les animaux sont sacrifiés dans une
cuve à CO2. Une incision thoracique est ensuite
réalisée, afin d’exposer le cœur.
2.2. Montage Langendorff et digestion enzymatique
Une fois détaché, le cœur est canulé par l’aorte et
placé sur le montage Langendorff. Les artères
coronaires du ventricule gauche sont donc perfusées

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 5
de façon rétrograde par une solution tampon HANKS-
HEPES sans Ca2+ ajouté, dite de lavage, à 37°C
(NaCl : 113 mM, KCl : 4,7 mM, KH2PO4: 0,6 mM,
KHCO3 : 10 mM, Na2HPO4 : 0,6 mM, NaHCO3 : 12
mM, HEPES : 10 mM, MgCl2 :1,2 mM, glucose : 5,5
mM, taurine : 29 mM, pH ajusté à 7,4 avec du NaOH
5N) ce qui permet d’enlever toutes traces sanguines
dans les artères coronaires. Du 2,3-Butanedione
monoxime (BDM, 10 mM) est ajouté dans cette
solution de lavage. Il s’agit d’une substance
cardioplégique qui d’inhibe l’ATPase de la myosine et
empêche rapidement et réversiblement la formation
des ponts entre actine et myosine. Cela permet au
cœur de rester en permanence relaxé et prévient les
dommages dus aux contractions terminales, à
l’ischémie et à la reperfusion (Louch et al, 2011).
Toutefois, l’utilisation de ce milieu très réduit en Ca2+
entraîne une fragilisation du sarcolemme qui peut
induire une accumulation de sodium (Na+). Si bien
que lors de la réintroduction du Ca2+ extracellulaire,
l’échange Na+/Ca2+ est dans le sens d’une entrée de
Ca2+, ce qui va induire une surcharge en Ca2+
intracellulaire dramatique pour la survie de la cellule.
C’est pourquoi la solution de lavage contient aussi de
la taurine (3 mM) qui permet de protèger les cellules
contre ce phénomène appelé « paradoxe calcique ».
Il existe en effet un symport taurine/Na+ permettant
de vider la cellule d’une surcharge de sodium.
Ensuite, une solution de lavage contenant de la
Liberase (Solution commerciale contenant différents
types de collagénases ; Roche, 10 ml/mL) est
perfusée jusqu’à ce qu’une augmentation du débit
d’écoulement soit observée (6-9 min), ce qui
témoigne d’une bonne digestion des connections
intercellulaires. Une fois digéré, le cœur est détaché
par coupure au niveau des oreillettes. Puis, les
cellules sont dissociées par séparation mécanique,
filtrées et mises à sédimenter pendant 10 minutes. La
concentration calcique du milieu est alors augmentée
durant une phase de réplétion en 4 étapes
successives de 5 min jusqu’à atteindre 1 mM.
Pour les études par Western-Blot, les cardiomyocytes
sont ensuite déposés dans des boites de Pétri (35
mm de diamètre) prétraitées avec de la laminine (20
mg/mL) en présence d’une solution de Tyrode (NaCl :
113mM, KCl :4,7mM, MgCl2 : 1,2 mM, Na2HPO4 : 0,6
mM, NaHCO3 : 4,4 mM, HEPES : 20 mM, glucose :
10 mM, taurine : 20 mM, Na pyruvate : 3 mM, CaCl2 :
1,25mM, pH : 7,35) pendant 2-3 heures.
Pour les études fonctionnelles, ce n’est pas la boite
de Pétri qui est prétraitée avec de la laminine (20
mg/mL) mais une lamelle de verre (Corning, 25x25
mm) placée à l’intérieur de la boite de Pétri (35 mm)
en présence de la même solution de Tyrode pendant
2-3 heures.
3. Analyse fonctionnelle :
Mesure de la contraction
et du transitoire calcique
des cardiomyocytes
isolés
3.1. Marquage des cardiomyocytes par la sonde Fura-2AM
Les cardiomyocytes sont chargés ou non avec du
Fura-2AM (ester acétoxyméthylé perméant,
Molecular Probes, 1μM dans du DMSO [diméthyl
sulfoxide]) à température ambiante et obscurité
pendant 20 minutes, puis lavés durant 5 minutes
avec la solution de Tyrode afin de permettre la
désestérification du Fura-2 intracellulaire.

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 6
3.2. Systèmes d’enregistrement
Les raccourcissements des sarcomères et les
transitoires calciques ont été enregistrés
simultanément en imposant une stimulation électrique
de champ (0,5 Hz, 0,1 ms, 7-15 V) à l’aide du
système IonOptix (Myocyte Calcium and Contractility
Recording System, Dublin, Irlande) couplé à un
microscope à fluorescence inversée (Episcopic
fluorescence attachment ef-inv-II ; Inverted
microscope AE30-31). La contraction des sarcomères
est enregistrée à une fréquence de 500 Hz tandis que
les transitoires calciques sont enregistrés à une
fréquence de 250 Hz. Les réponses sont analysées
avec le logiciel IonWizard 6.3 (IonOptix).
4. Analyse moléculaire
Suite aux différentes stimulations, les cardiomyocytes
sont détachés par grattage, puis les protéines en sont
extraites. Pour cela, les cellules sont récupérées
dans des tubes eppendorf et centrifugés à 500g
pendant 5 minutes à 20°C. Le culot est repris dans du
PBS, puis centrifugé à 1500g pendant 5 minutes. Le
surnageant est ensuite éliminé et le culot plongé dans
l’azote liquide. Dans un premier temps, le dosage des
protéines est réalisé selon la méthode de Lowry. Ce
dosage permet d’orienter la manière dont il est
possible d’exploiter l’échantillon par la suite et
notamment de savoir si la quantité de protéines est
suffisante pour une détection par Western-Blot. Le
reste de l’échantillon est ensuite utilisé pour réaliser
des Western-Blot.
4.1. Dosage des protéines
Le dosage des protéines a été réalisé en suivant le
protocole de Lowry. 5 μL d’échantillons sont déposés
en duplicate sur une plaque 96 puits. 25 μL de réactif
A (BioRad), contenant des ions Cu2+, sont ensuite
ajoutés. Les atomes d’azote contenus dans les
liaisons peptidiques des protéines réduisent ces ions
Cu2+ en ions Cu+. Puis, 200 μL d’un réactif B
(BioRad) sont ajoutés. Ce dernier va former un
complexe de couleur bleu avec les ions Cu+. La
plaque est laissée à incuber 10 minutes à l’abri de la
lumière puis l’absorbance est mesurée à 570 nm
grâce à un lecteur de microplaques (Mithras LB 940,
Berthold technologies). La détermination de la
concentration se fait par comparaison avec
l’absorbance mesurée d’une gamme étalon de BSA
(« Bovine serum albumine ») 5%.
4.2. Western-Blot
Les protéines sont séparées sur un gel de poly-
acrylamide (Novex Protein from Life technologies,
NuPAGE 4-12% Bis-Tri Gel ; 1,0 mm X 12 well ou 1,5
mm X 10 well) pendant 90 minutes à 150 V, puis
transférées sur une membrane de nitrocellulose
pendant 90 minutes à 110 V à 4°C. La membrane est
alors colorée au rouge Ponceau puis saturée avec
une solution de sérum d’albumine bovine (BSA) à 5%
pendant 90 minutes à température ambiante afin
d’éliminer les sites de fixation non spécifiques.
Ensuite, la membrane est mise à incuber avec
l’anticorps primaire pendant 90 minutes à
température ambiante, ou à 4°C durant la nuit. Le
lendemain, 3 lavages de 10 minutes au TBS-Tween
(Tris buffered saline and Tween) (20 mM Tris ;
61,5mM NaCl ; 0,1% Tween) sont effectués afin
d’éliminer l’anticorps en excès. L’anticorps
secondaire couplé à la peroxydase est alors mis à
incuber avec la membrane pendant 90 minutes à
température ambiante, puis une autre série de
lavages, 3 fois 10 minutes, avec du TBS-Tween est
effectuée. Les protéines sont révélées par

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 7
chemiluminescence soit avec du luminol (Santa Cruz)
pendant 1 minute, soit grâce au kit de révélation
SuperSignal® West Femto Maximum Sensitivity
Substrate (ThermoScientific, Fischer) pendant 3
minutes. La révélation s’effectue grâce à un système
d’acquisition en chambre noire (ChemiSmart 500,
Vilbert Lourmat) et du logiciel Chemi-Capt 5.0. Les
différents anticorps qui ont été utilisés sont l’anticorps
polyclonal anti-TNF-α (Santa Cruz, 1/500) produit
chez la chèvre, l’anticorps polyclonal anti-α-tubuline
(Santa Cruz, 1/500) produit chez la chèvre et l’anti-
IgG de chèvre conjugué à HRP (Santa Cruz, 1/5000)
produit chez l’âne.
5. Analyses statistiques
Les différentes bandes d’intérêt des Western Blot
sont quantifiées relativement à l’aide du logiciel BIO-
1D. Ces résultats sont exprimés en moyenne ± erreur
standard à la moyenne (ESM) de “n” expériences, i.e.
le nombre de souris étudiées. Les différences entre
les groupes de données ont été comparées à l’aide
d’un Mann-Whitney non paramétrique à l’aide du
logiciel Prism 5.0. Une valeur de P inférieur à 0,05 a
été admise pour que les différences entre les
données soient considérées comme significatives.
Les résultats des analyes fonctionnelles sont
exprimés en moyenne ± ESM de « n » expériences,
i.e. le nombre de cellules étudiées. Les différences
entre les groupes de données ont été comparées sur
le logiciel Prism 5.0 à l’aide du test t (test de Student)
non apparié et par la méthode statistique ANOVA,
suivie d’un test de comparaisons multiples de
Bonferroni, pour les expériences utilisant les MPs.
Une valeur de P inférieure à 0,05 a été admise pour
que les différences entre les données soient
considérées comme significatives.
Résultats
1. Le TNFαest présent dans
les MPs macrophagiques
Le TNF-α a été détecté par Western-blot sur des
extraits protéiques de MPs issues de macrophages
RAW 264.7 traitées avec de l’ActD (figure 1). Sur les
membranes de nitrocellulose, on peut notamment
distinguer les bandes correspondant aux deux formes
de TNF-α, libre (17kDa) et transmembranaire
(26kDa).
Figure 1 : Western-blot montrant la présence de
TNF-α dans les MPs
Expression du TNF-α (n=3)
2. TNF per se induit un
effet inotrope négatif
L’étude de l’effet du TNF-α per se (800 U/mL) sur la
contractilité des cardiomyocytes (figure 2) indique
que le TNF-α a un effet inotrope négatif sur les
cardiomyocytes dès 15 min et induit un
ralentissement des cinétiques de raccourcissement et
de relaxation sarcomérique.

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 8
Figure 2: Effet du TNF-α per se (800 U/mL) sur la
contractilité des cardiomyocytes
A : Amplitude maximale de raccourcissement (% de
variation de la longueur basale de sarcomère)
B : Vitesse maximale de raccourcissement (-dL/dt, en
μm/sec)
C : Vitesse maximale de réallongement (+dL/dt, en
μm/sec)
Les valeurs représentent les moyennes ± ESM; n=24-
28 cellules par groupe, *P<0.05 vs. Contrôle (CTRL).
3. Le TNF-α n’a pas d’effet
sur les transitoires
calciques
L’étude de l’effet du TNF-α (800 U/mL) sur les
transitoires calciques des cardiomyocytes (figure 3)
indique qu’il n’y a pas d’effets significatifs du TNF-α
sur les différents paramètres du transitoire calcique.
Figure 3: Effets contrôles du TNF-α (800 U/mL)
sur les transitoires calciques des cardiomyocytes
A : Pourcentage de variation du rapport de
fluorescence F340/F380
B : Vitesse maximale de sortie de Ca2+ à l’extérieur
du RS (depV)
C : Temps au pic depuis t0 (milliseconde) (peak T)
D : Vitesse maximale de repompage du Ca2+ (retV)
E : Constante de tps de repompage calcique (en
milliseconde) (TAU)
Les valeurs représentent les moyennes ± ESM; n=24-
28 cellules par groupe, *P<0.05 vs. Contrôle (CTRL).
Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 9
4. Les MPS induisent un
effet inotrope négatif
L’étude de l’effet des MPs sur la contractilité des
cardiomyocytes (figure 4) indique que les MPs ont un
effet inotrope négatif rapide sur les cardiomyocytes
car ils induisent une diminution des
raccourcissements sarcomériques.
Figure 4 : Effet des MPs à 1 et 10 μg/ml sur la
contractilité des cardiomyocytes
A : Enregistrement typique des raccourcissements
sarcomérique
B : Enregistrement typique des transitoires calciques
C : histogramme présentant l’effet des MPs à 1 et 10
μg/ml sur le pourcentage de raccourcissement
sarcomérique. Les valeurs représentent les
moyennes ± ESM; n=28-50 cellules par groupe,
*P<0,05 vs. contrôle (CTRL).
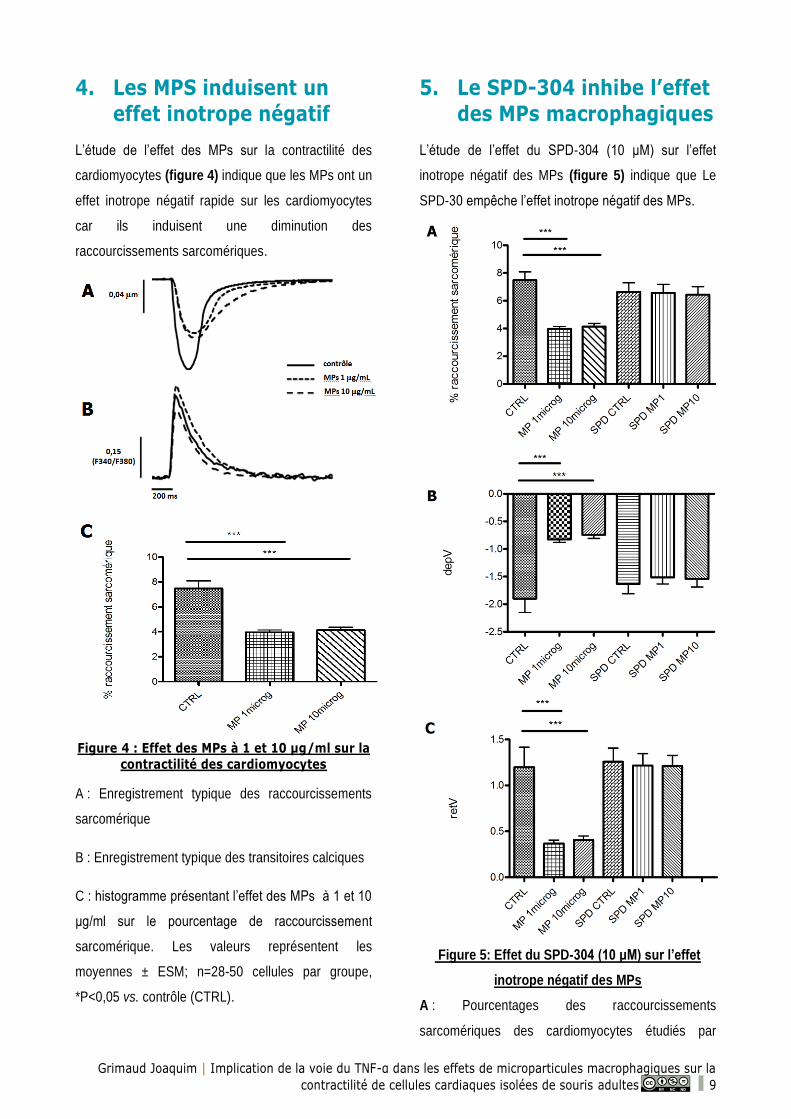
5. Le SPD-304 inhibe l’effet
des MPs macrophagiques
L’étude de l’effet du SPD-304 (10 μM) sur l’effet
inotrope négatif des MPs (figure 5) indique que Le
SPD-30 empêche l’effet inotrope négatif des MPs.
Figure 5: Effet du SPD-304 (10 μM) sur l’effet
inotrope négatif des MPs
A : Pourcentages des raccourcissements
sarcomériques des cardiomyocytes étudiés par

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 10
vidéo-imaging en présence ou non de MPs et de
SPD-304
B : Vitesse maximale de contraction sarcomérique
(depV, μm/sec)
C : Vitesse maximale de relaxation sarcomérique
(retV, μm/sec)
Les valeurs représentent les moyennes ± ESM; n=16-
50 cellules par groupe, *P<0,05 vs. contrôle (CTRL).
Discussion
Lors de cette étude, la présence de TNF-α a bien été
confirmée dans les MPs. De plus le TNF-α a un effet
inotrope négatif sur la contraction, i.e. provoque une
diminution de la contractilité des cardiomyocytes.
Cependant, cette diminution de la contractilité ne
s’accompagne pas d’une modification des transitoires
calciques. Nous avons confirmé que les MPs
induisent un effet inotrope négatif. L’inhibition de la
voie du TNF-α par son inhibiteur SPD-304 rétablie la
contractilité normale. L’ensemble de ces résultats
suggère que le TNF-α est impliqué dans l’effet
inotrope négatif des MPs issues de macrophages sur
la contractilité des cardiomyocytes.
Il existe un autre mécanisme d’intéraction potentiel
entre MPs et cardiomyocytes différent de l’intéraction
ligand-récepteur, l’internalisation. Ce processus
d’internalisation des MPs sur des cardiomyocytes a
déjà été étudié (Milbank et al., 2014). Il a ainsi été
montré que les MPs commencent à être internalisées
seulement après 1 heure au contact d’un
cardiomyocyte, tandis que l’effet inotrope négatif des
MPs est observé seulement après 5 minutes.
L’implication de ce mode d’action est donc écartée.
Le temps de traitement avec les MPs est également
trop court pour entraîner des modifications
d’expression de protéines impliquées dans le
couplage excitation-contraction.
Les résultats indiquent que le TNF-α
transmembranaire est porté par les MPs issues des
macrophages RAW 264.7 traités avec de l’Act D. De
plus, il a été montré que le TNF-α transmembranaire
est capable d’intéragir avec le TNF-R1 et le TNF-R2
(Horiuchi et al., 2010). Ces récepteurs TNF-R1 et
TNF-R2 se retrouvent exprimés à la surface des
cardiomyocytes (Higuchi et al., 2004).
Il n’y a pas eu d’effet observé des MPs sur le
transitoire calcique. Il est néanmoins possible qu’il y
ait un effet et que cet effet ne soit pas détectable
avec la technique utilisée ou que la libération de Ca2+
à partir du RS cache la diminution éventuelle de
l’entrée de Ca2+ par ICaL. Cependant, d’autres
travaux indiquent que le TNF-α aurait plutôt un effet
sur la sensibilité des protéines contractiles au Ca2+
(Goldhaber et al., 1996). En présence de TNF-α, la
sensibilité des protéines contractiles, comme l’actine,
la myosine ou la troponine I au Ca2+ serait diminuée.
Conclusion
La voie du TNF-α pourrait donc bien être impliquée
dans l’effet des MPs macrophagiques sur la
contractilité des cardiomyocytes. Il serait maintenant
intéressant de confirmer l’implication du TNF-α porté
par ces MPs en explorant diverses voies de
signalisation du TNF-α, comme la voie du
NO/cGMP/PKG, la voie phospholipase A2/acide
arachidonique et la voie sphingosines. Différents
inhibiteurs pharmacologiques pourraient être utilisés
comme la L-Nitroarginine (LNA) qui est un inhibiteur
de la « nitric oxide synthase » (NOS), l’ODQ (1H-
[1,2,4]oxadiazolo-[4, 3-a]quinoxalin-1-one) qui est un

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 11
inhibiteur de la guanylate cyclase soluble (GCs), le
KT 5823 qui est un inhibiteur de la protéine kinase G
(PKG), l’AACOCF3 (Arachidonyl trifluoromethyl
ketone) qui est un inhibiteur de la phospholipase-A2
et le SKI-II (Sphingosine kinase inhibitor 2). Un
« cross-talk » entre la voie de la PLA2/AA et la voie
médiée par le TNF-α a par ailleurs été démontré
récemment (Saito et al., 2012).
Références bibliographiques
- Beaglehole R, Bonita R (2008). Global public
health: A scorecard. Lancet 372: 1988-96.
- Bers DM (2002). Cardiac excitation-
contraction coupling. Nature, 415:198-205.
- Biro E, Lok CA, Hack CE, van der Post JA,
Schaap MC, Sturk A, Nieuwland R (2007).
Cell-derived microparticles and complement
activation in preeclampsia versus normal
pregnancy. Placenta, 28: 928-35.
- Blankenberg S, Barbaux S, Tiret L (2003).
Adhesion molecules and atherosclerosis.
Atherosclerosis, 191-203
- Bloom DE, Cafiero ET, Jané-Llopis E,
Abrahams-Gessel S, Bloom LR, Fathima S,
Feigl AB, Gaziano T, Mowafi M, Pandya A,
Prettner K, Rosenberg L, Seligman B, Stein
AZ, Weinstein C (2011). The Global
Economic Burden Of Noncommunicable
Diseases. Geneva: World Economic Forum
- Braunwald E, Bristow MR (2000).
Congestive heart failure: fifty years of
progress. Circulation, 102: IV14-23
- Budaj M, Poljak Z, Duris I, Kasko M, Imrich
R, Kopani M, Maruscakova L, Hulin I (2012).
Microparticles: a component of various
diseases. Pol Arch Med Wewn, 122 Suppl 1:
24-9.
- Cocucci E, Racchetti G, Meldolesi J (2009).
Shedding microvesicles: artefacts no more.
Trends Cell Biol, 19: 43-51.
- Distler JH, Pisetsky DS, Huber LC, Kalden
JR, Gay S, Distler O (2005). Microparticles
as regulators of inflammation: novel players
of cellular crosstalk in the rheumatic
diseases. Arthritis Rheum, 52: 3337-48.
- Freyssinet JM (2003). Cellular
microparticles: what are they bad or good
for? J Thromb Haemost 1: 1655-62.
- Goldhaber JI, Kim KH, Natterson PD,
Lawrence T, Yang P, Weiss JN (1996).
Effects of TNF-alpha on [Ca2+]i and
contractility in isolated adult rabbit ventricular
myocytes. Am J Physiol, 271:H1449-55.
- Higuchi Y, McTiernan CF, Frye CB,
McGowan BS, Chan TO, Feldman AM
(2004). Tumor necrosis factor receptors 1
and 2 differentially regulate survival, cardiac
dysfunction, and remodeling in transgenic
mice with tumor necrosis factor-alpha-
induced cardiomyopathy. Circulation, 109:
1892-7.
- Horiuchi T, Mitoma H, Harashima S,
Tsukamoto H, Shimoda T (2010).
Transmembrane TNF-alpha: structure,
function and interaction with anti-TNF
agents. Rheumatology (Oxford), 49:1215-28.
- Hugel B, Martinez MC, Kunzelmann C,
Freyssinet JM (2005). Membrane
microparticles: two sides of the coin.
Physiology (Bethesda) 20, 22-27.
- Jenkins NT, Padilla J, Boyle LJ, Credeur DP,
Laughlin MH, Fadel PJ (2013). Disturbed
Blood Flow Acutely Induces Activation and
Apoptosis of the Human Vascular
Endothelium. Hypertension, 61:615-21.

Grimaud Joaquim | Implication de la voie du TNF-α dans les effets de microparticules macrophagiques sur la
contractilité de cellules cardiaques isolées de souris adultes 12
- Lee YS, Keener JP (2008). A calcium-
induced calcium release mechanism
mediated by calsequestrin. J Theor Biol, 253:
668-79.
- Libby P, Ridker PM, Maseri A (2002).
Inflammation and Atherosclerosis.
Circulation, 105: 1135-43.
- Louch WE, Sheehan KA, Wolska BM (2011).
Methods in cardiomyocyte isolation, culture,
and gene transfer. J Mol Cell Cardiol 51:
288-98.
- Lusis AJ (2000). Atherosclerosis. Nature
407: 233-41.
- Martínez MC, Tesse A, Zobairi F,
Andriantsitohaina R (2005). Shed membrane
microparticles from circulating and vascular
cells in regulating vascular function. Am J
Physiol Heart Circ Physiol 288: H1004-9.
- Milbank E, Soletti R, Andriantsitohaina R,
Noireaud J (2014). Negative inotropic and
proapototic effects of macrophagic
microparticles in adult murine
cardiomyocytes. Fund Clin Pharmacology
28, 79.
- Piccin A, Murphy WG, Smith OP (2007).
Circulating microparticles: pathophysiology
and clinical implications. Blood Rev 21: 157-
71.
- Pinz I, Zhu M, Mende U, Ingwall JS (2011).
An improved isolation procedure for adult
mouse cardiomyocytes. Cell Biochem
Biophys 61: 93-101.
- Poston RN, Johnson-Tidey RR (1996).
Localized adhesion of monocytes to human
atherosclerotic plaques demonstrated in
vitro: implications for atherogenesis.
American Journal of Pathology, 149: 73-80.
- Saito Y, Watanabe K, Fujioka D, Nakamura
T, Obata JE, Kawabata K, Watanabe Y,
Mishina H, Tamaru S, Kita Y, Shimizu T,
Kugiyama K (2012). Disruption of group IVA
cytosolic phospholipase A(2) attenuates
myocardial ischemia-reperfusion injury partly
through inhibition of TNF-α-mediated
pathway. Am J Physiol Heart Circ Physiol,
302: H2018-30.
- Sewify EM, Sayed D, Abdel Aal RF, Ahmad
HM, Abdou MA (2013). Increased circulating
red cell microparticles (RMP) and platelet
microparticles (PMP) in immune
thrombocytopenic purpura. Thromb Res,
131: e59-63.
- Spagnoli LG, Bonanno E, Sangiorgi G,
Mauriello A (2007). Role of inflammation in
atherosclerosis. The Journal of Nuclear
Medicine, 48: 1800-15.
- Swirski FK, Nahrendorf M (2013). Leukocyte
behavior in atherosclerosis, myocardial
infarction, and heart failure. Science,
339:161-6.
- Tedgui A, Mallat Z (2006). Cytokines in
atherosclerosis: pathogenic and regulatory
pathways. Physiol Rev 86: 515-81.
- Wolf P (1967). The nature and significance
of platelet products in human plasma. Br J
Haematol 13: 269-88.
- World Health Organization (2007).
Prevention of Cardiovascular Disease,
Pocket Guidelines for Assessment and
Management of Cardiovascular Risk.
- World Health Organization (2011). Global
Atlas on cardiovascular disease prevention
and control

ENGAGEMENT
DE NON PLAGIAT Je, soussigné(e) ......................................................................
déclare être pleinement conscient(e) que le plagiat de documents ou d’une partie d’un document publiée sur toutes formes de support, y compris l’internet, constitue une violation des droits d’auteur ainsi qu’une fraude caractérisée. En conséquence, je m’engage à citer toutes les sources que j’ai utilisées pour écrire ce rapport ou mémoire.
signé par l'étudiant(e) le 31 / 05 / 2014
Cet engagement de non plagiat doit être signé et joint
à tous les rapports, dossiers, mémoires.
Présidence de l'université
40 rue de rennes – BP 73532
49035 Angers cedex
Tél. 02 41 96 23 23 | Fax 02 41 96 23 00


















