Granzyme B-dependent proteolysis acts as a switch to
26
Granzyme B-dependent proteolysis acts as a switch to enhance the pro-inflammatory activity of IL-1α Inna S. Afonina 1 , Graham A. Tynan 2 , Susan E. Logue 1 , Sean P. Cullen 1 , Michael Bots 3 , Alexander U. Lüthi 1 , Emer P. Reeves 4 , Noel G. McElvaney 4 , Jan P. Medema 3 , Ed C. Lavelle 2 , and Seamus J. Martin 1,5 1 Molecular Cell Biology Laboratory, Dept. of Genetics, The Smurfit Institute, Trinity College, Dublin 2, Ireland 2 Adjuvant Research Group, School of Biochemistry and Immunology, Trinity College, Dublin 2, Ireland. 3 LEXOR (Lab of Exp. Oncology and Radiobiology), Center for Experimental Molecular Medicine, Academic Medical Center (AMC), Amsterdam, The Netherlands. 4 Respiratory Research Division, Department of Medicine, Education and Research Centre, Royal College of Surgeons in Ireland, Beaumont Hospital, Dublin 9, Ireland. Summary Granzyme B is a cytotoxic lymphocyte-derived protease that plays a central role in promoting apoptosis of virus-infected target cells, through direct proteolysis and activation of constituents of the cell death machinery. However, previous studies have also implicated granzymes A and B in the production of pro-inflammatory cytokines, via a mechanism that remains undefined. Here we show that IL-1α is a substrate for granzyme B and that proteolysis potently enhanced the biological activity of this cytokine in vitro as well as in vivo. Consistent with this, compared with full-length IL-1α, granzyme B-processed IL-1α exhibited more potent activity as an immunoadjuvant in vivo. Furthermore, proteolysis of IL-1α within the same region, by proteases such as calpain and elastase, was also found to enhance its biological potency. Thus, IL-1α processing by multiple immune-related proteases, including granzyme B, acts as a switch to enhance the pro-inflammatory properties of this cytokine. Keywords IL-1α; granzyme B; inflammation; proteolysis; cell death; apoptosis Introduction Granzyme (Gzm) B is a protease typically found in the cytotoxic granules of Natural Killer (NK) and cytotoxic T cells (CTLs) and, upon delivery into the cytoplasm of virus-infected target cells, promotes apoptosis to limit virus replication and dissemination (reviewed in Afonina et al., 2010). Gzm B promotes apoptosis through proteolysis of the BH3-only protein Bid, as well as via proteolytic processing and activation of caspase-3 (Afonina et al., 2010). However, accumulating evidence suggests that granzymes A and B may also influence the production of pro-inflammatory cytokines, although the molecular targets of these proteases in inflammatory contexts remain to be defined (Sower et al., 1996a,b; Metkar et al., 2008; Froelich et al., 2009). Interestingly, a number of non-cytotoxic cell 5 Correspondence: Seamus J. Martin, Ph.D. [email protected]. The authors have no conflicting financial interests to declare. Europe PMC Funders Group Author Manuscript Mol Cell. Author manuscript; available in PMC 2012 April 04. Published in final edited form as: Mol Cell. 2011 October 21; 44(2): 265–278. doi:10.1016/j.molcel.2011.07.037. Europe PMC Funders Author Manuscripts Europe PMC Funders Author Manuscripts
Transcript of Granzyme B-dependent proteolysis acts as a switch to

Granzyme B-dependent proteolysis acts as a switch to enhancethe pro-inflammatory activity of IL-1α
Inna S. Afonina1, Graham A. Tynan2, Susan E. Logue1, Sean P. Cullen1, Michael Bots3,Alexander U. Lüthi1, Emer P. Reeves4, Noel G. McElvaney4, Jan P. Medema3, Ed C.Lavelle2, and Seamus J. Martin1,5
1Molecular Cell Biology Laboratory, Dept. of Genetics, The Smurfit Institute, Trinity College,Dublin 2, Ireland 2Adjuvant Research Group, School of Biochemistry and Immunology, TrinityCollege, Dublin 2, Ireland. 3LEXOR (Lab of Exp. Oncology and Radiobiology), Center forExperimental Molecular Medicine, Academic Medical Center (AMC), Amsterdam, TheNetherlands. 4Respiratory Research Division, Department of Medicine, Education and ResearchCentre, Royal College of Surgeons in Ireland, Beaumont Hospital, Dublin 9, Ireland.
SummaryGranzyme B is a cytotoxic lymphocyte-derived protease that plays a central role in promotingapoptosis of virus-infected target cells, through direct proteolysis and activation of constituents ofthe cell death machinery. However, previous studies have also implicated granzymes A and B inthe production of pro-inflammatory cytokines, via a mechanism that remains undefined. Here weshow that IL-1α is a substrate for granzyme B and that proteolysis potently enhanced thebiological activity of this cytokine in vitro as well as in vivo. Consistent with this, compared withfull-length IL-1α, granzyme B-processed IL-1α exhibited more potent activity as animmunoadjuvant in vivo. Furthermore, proteolysis of IL-1α within the same region, by proteasessuch as calpain and elastase, was also found to enhance its biological potency. Thus, IL-1αprocessing by multiple immune-related proteases, including granzyme B, acts as a switch toenhance the pro-inflammatory properties of this cytokine.
KeywordsIL-1α; granzyme B; inflammation; proteolysis; cell death; apoptosis
IntroductionGranzyme (Gzm) B is a protease typically found in the cytotoxic granules of Natural Killer(NK) and cytotoxic T cells (CTLs) and, upon delivery into the cytoplasm of virus-infectedtarget cells, promotes apoptosis to limit virus replication and dissemination (reviewed inAfonina et al., 2010). Gzm B promotes apoptosis through proteolysis of the BH3-onlyprotein Bid, as well as via proteolytic processing and activation of caspase-3 (Afonina et al.,2010). However, accumulating evidence suggests that granzymes A and B may alsoinfluence the production of pro-inflammatory cytokines, although the molecular targets ofthese proteases in inflammatory contexts remain to be defined (Sower et al., 1996a,b;Metkar et al., 2008; Froelich et al., 2009). Interestingly, a number of non-cytotoxic cell
5Correspondence: Seamus J. Martin, Ph.D. [email protected].
The authors have no conflicting financial interests to declare.
Europe PMC Funders GroupAuthor ManuscriptMol Cell. Author manuscript; available in PMC 2012 April 04.
Published in final edited form as:Mol Cell. 2011 October 21; 44(2): 265–278. doi:10.1016/j.molcel.2011.07.037.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

types, such as mast cells and basophils, have also been found to secrete Gzm B uponactivation (Tschopp et al., 2006; Strik et al., 2007). Furthermore, Gzm B is found at elevatedlevels in serum from Rheumatoid arthritis (RA) patients and in other inflammatoryconditions associated with elevated levels of IL-1 (Lauw et al., 2000; Tak et al., 1999; Kimet al., 2007). For these reasons, we explored whether Gzm B may contribute to theprocessing and activation of IL-1.
IL-1 is a cytokine with diverse biological activities and is produced in the early stages ofinfection or sterile injury where it plays an important role in instigating immune responses(Dinarello, 1996; Kono et al., 2010). Although IL-1 is produced in two different forms(IL-1α and IL-1β), encoded by distinct genes, these cytokines bind to the same receptor andshare similar biological activities (Dinarello, 1996). Despite their relatedness in structureand biological activity, IL-1α and IL-1β are post-translationally processed in a verydifferent manner. IL-1α is expressed as a ~31 kDa polypeptide and in this form is capable ofbinding to the IL-1 receptor and is biologically active (Mosley et al., 1987a,b). IL-1β is alsoexpressed as a ~31 kDa polypeptide, but in marked contrast to IL-1α, full length IL-1β isincapable of binding to the IL-1 receptor and consequently, exhibits little biological activity(Mosley et al., 1987a,b; Hazuda et al., 1990). IL-1β acquires biological activity upon limitedinternal processing by the aspartic acid-specific protease, caspase-1 (Thornberry et al, 1992).Caspase-1-dependent processing of IL-1β at Asp116 unlocks the biological activity of thiscytokine. Gzm A has also been found to cleave IL-1β in vitro, however the functionalconsequences of this event remain unclear (Irmler et al., 1995).
IL-1α is typically released from cells as a result of injury or death and circulating IL-1αlevels are persistently elevated in inflammatory conditions such as RA. Although full-lengthIL-1α displays biological activity, this cytokine is susceptible to proteolysis by certainintracellular proteases. For example, the calcium-activated protease, calpain-1, promotesrestricted proteolysis of IL-1α to produce a ~17 kDa protein, although this reportedly haslittle effect on the biological activity of this cytokine (Kobayashi et al., 1990; Carruth et al.,1991). Therefore, it remains unclear what role calpain-mediated processing of IL-1α serves.As a full-length protein, IL-1α is typically cell-associated, and a portion of this cell-associated form may be present at the plasma membrane (Kurt-Jones et al., 1985).Proteolytic processing of IL-1α may remove a nuclear localization signal or permit sheddingof membrane-associated IL-1α and increase the likelihood of secretion (Watanabe andKobayashi, 1994; Matsuhima et al., 1986). Thus, proteolysis of IL-1α may convert thiscytokine from a cell-associated form to a soluble molecule that could exert more systemiceffects. An additional possibility is that proteolysis of IL-1α may modulate the biologicalpotency of this cytokine, although this issue has not been explored in any detail.
Here we report that human IL-1α, but not IL-1β, is a substrate for the CTL/NK cellprotease, Gzm B, and that processing of this cytokine by the latter protease enhanced itsbiological activity several-fold. A similar increase in IL-1α activity was also seen uponprocessing of IL-1α by other proteases, such as calpain-1, elastase and mast cell chymase.Thus, Gzm B and other proteases that are produced during inflammatory responses maypromote inflammation, at least in part, through conversion of pro-IL-1α to its mature form.
ResultsIL-1α is a substrate for granzyme B
Because recent studies have suggested that CTL/NK granzymes may have extracellular pro-inflammatory roles, we set out to explore whether Gzm B was capable of processing any ofthe major pro-inflammatory cytokines. To address this issue, we screened a panel of in vitrotranscribed and translated members of the interleukin family (IL-1 to IL-17) for
Afonina et al. Page 2
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

susceptibility to proteolysis by Gzm B, as well as a range of other aspartic acid-specificproteases as controls (data not shown). All proteases were confirmed to be active by virtueof their ability to cleave synthetic tetrapeptide substrates (data not shown) or control proteinssuch as Bid (Figure 1A). The outcome of this screen identified full length IL-1α as asubstrate for Gzm B but not for any of the other proteases tested. As Figure 1A shows,whereas caspases-1, -3, -4, -5, and – 7 failed to cleave IL-1α, Gzm B produced a majorcleavage product at ~17 kDa. In contrast, Gzm B did not cleave IL-1β, whereas caspase-1did so, as expected (Figure 1A). To confirm the specificity of IL-1α proteolysis by Gzm B,we titrated this protease over a range of concentrations from 12.5 to 100 nM. IL-1β failed tobe cleaved by Gzm B at any of these concentrations, whereas proteolysis of IL-1α, as wellas the known Gzm B substrate, Bid, was readily observed (Figure 1B). Importantly, becausehuman and murine GzmB have been shown to have divergent substrate preferences (Cullenet al., 2007), we also tested whether murine IL-1α was a substrate for Gzm B and found thatboth human and mouse Gzm B cleaved murine IL-1α within the nanomolar range (FigureS1A and S1B).
IL-1α is cleaved by granzyme B at Asp103To map the Gzm B cleavage site within IL-1α, we expressed and purified the full-lengthcytokine from E. coli and incubated recombinant IL-1α with Gzm B to promote proteolysis.Cleavage products were then analysed by mass spectrometry, which suggested thatproteolysis occurred between amino acids 80 and 112 (Figure 1C). Because Gzm B has analmost absolute preference for proteolysis after Asp residues and typically cleaves withinIXXD/S motifs, this suggested that IL-1α was cleaved after Asp103, within the motifIAND103, which is conserved among mammals (Figure 1D). To confirm this as the site ofproteolysis, we generated a point mutation Asp>Ala103 in IL-1α and tested thesusceptibility of this mutant to proteolysis by Gzm B. As Figure 1E shows, this mutant fullyresisted proteolysis, as did a deletion mutant of IL-1α lacking the N-terminal 103 aminoacids (Figure 1F), confirming that Gzm B cleaves IL-1α at Asp103.
Granzyme B-mediated proteolysis of IL-1α potentiates biological activityPrevious studies have shown that calpain-1 can process IL-1α at Phe118 (Kobayashi et al.,1990), however the significance of this cleavage event for IL-1α bioactivity has not beendetermined. Indeed, it is widely assumed that proteolytic processing of IL-1α within its N-terminal region has little effect on the biological activity of this cytokine. For these reasons,we wondered whether Gzm B-mediated processing of IL-1α could impact upon itsbiological activity. To explore this issue, we measured the ability of IL-1α to stimulateproduction of IL-6 and IL-8 from HeLa cells and primary HUVECs, as both cell types areknown to respond to this cytokine (Bertelsen and Sanfridson, 2007; Rhim et al., 2008).
To mimic the Gzm B cleavage product of IL-1α, we generated a truncated form of thiscytokine missing the N-terminal 103 amino acids (IL-1α104-271). Using this form of IL-1αwe confirmed that both HeLa cells and primary HUVECs were highly responsive to thiscytokine (Figure 2A), with both cell types producing IL-6 and IL-8 in response toIL-1α104-271. HUVECs also produced GM-CSF under the same conditions (Figure 2A).These observations confirmed that truncation of IL-1α after Asp103 produced a biologicallyactive cytokine. However, this type of artificial truncation does not necessarily faithfullyreproduce the natural cleaved form of full length IL-1α, as proteolysis of a protein does notnecessarily disassociate the resulting cleavage products.
To compare the effects of Gzm B-mediated proteolysis of IL-1α with the full-lengthcytokine, we incubated bacterially expressed IL-1α precursor in the presence or absence ofGzm B, followed by assessment of biological activity as above. Surprisingly, proteolytic
Afonina et al. Page 3
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

processing of IL-1α by Gzm B increased the activity of this cytokine several-fold (Figure2B). Similar observations were also made in time course experiments, once again indicatingthat Gzm B-mediated proteolysis dramatically increased the bioactivity of IL-1α (Figure2C).
Importantly, we ruled out the possibility that co-purifying microbial contaminants in theGzm B preparations contributed to the effects seen. First, we expressed and purified acatalytically inactive Gzm B mutant (Gzm BSA) in the same yeast system that was used togenerate wild type Gzm B (Figure S2A-C). This mutant failed to cleave IL-1α and alsofailed to enhance the activity of IL-1α on HeLa or HUVECs (Figure S2C and S2D). Second,neither wild type nor mutant forms of Gzm B induced cytokines when added directly toHeLa cells in the absence of IL-1α (Figure S2E). Third, whereas HeLa cells are veryresponsive to IL-1α, these cells fail to produce IL-6 or IL-8 in response to a wide panel ofmicrobial components (i.e. pathogen-associated molecular patterns) such aslipopolysaccharide that would represent possible contaminants (Figure S2F).
Granzyme B does not directly influence inflammatory cytokine production by HeLa orHUVECs
To further rule out the possibility that residual Gzm B activity within the cleaved IL-1αpreparations was responsible for the increased pro-inflammatory effects seen, we neutralizedthis granzyme with the serpin protease inhibitor PI-9, confirming that no residual proteaseactivity remained after the incubation period with IL-1α (Figure 3A and B). Addition ofPI-9 after Gzm B-mediated proteolysis of IL-1α had no effect on the increased potency ofthe Gzm B-treated IL-1α preparations (Figure 3C), demonstrating that Gzm B was notdirectly acting upon the cells used to assay IL-1α activity. Similar observations were alsomade with HUVECs (data not shown).
We also directly compared the potency of full length IL-1α with the artificially-truncatedIL-1α104-271 protein (which mimics the Gzm B-cleaved form of this cytokine) and onceagain observed considerable differences in activity between these forms of IL-1α (Figure3D). Furthermore, incubation of truncated IL-1α104-271 with Gzm B did not have any effecton the activity of this cytokine, as expected (Figure 3E). Similarly, incubation of the Gzm B-non-cleavable form of IL-1α (IL-1αD103A) with this granzyme, also failed to enhance theactivity of this protein (data not shown). Collectively, these data provide strong evidencethat proteolysis of IL-1α by Gzm B enhances the biological activity of this protein up to 10-fold.
Proteolysis of IL-1α by calpain, as well as other proteases, enhances the bioactivity of thiscytokine
As mentioned earlier, calpain-1 has been shown to cleave IL-1α at Phe118 (Kobayashi etal., 1990), close to the Gzm B cleavage site at Asp103 (Figure 1D). Therefore, we wonderedwhether calpain-mediated proteolysis of IL-1α also enhanced the activity of this cytokine, asprevious studies have failed to address this issue. We first compared the relative efficiencyof IL-1α processing by calpain-1 versus Gzm B. As Figure 4A illustrates, calpain cleavedIL-1α very efficiently at nanomolar concentrations, producing a cleavage pattern distinctfrom that of Gzm B. Furthermore, calpain-1 also cleaved the Gzm B-resistant IL-1αD103A
mutant (Figure 4A), confirming that proteolysis by these proteases occurs at different siteswithin IL-1α.
We then explored the effects of calpain-1-mediated proteolysis on IL-1α activity using theHeLa and HUVEC assays as before. As Figure 4B illustrates, similar to our earlierobservations with Gzm B, proteolysis of IL-1α by calpain-1 also enhanced the potency of
Afonina et al. Page 4
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

this cytokine to a similar degree. Thus, proteolysis of IL-1α, by either Gzm B or calpain,greatly enhanced the biological activity of IL-1α, which suggests that restricted proteolysisof IL-1α within its N-terminus has a role in regulating the potency of this cytokine.
Elastase and mast cell chymase have been previously implicated as IL-1β-processingproteases (Black et al., 1988; Hazuda et al., 1990; Mizutani et al., 1991), but it is unclearwhether IL-1α is also processed by these enzymes. Therefore, we explored the effects ofelastase and mast cell chymase on IL-1α and found that both proteases did indeed cleavethis cytokine, albeit producing different processing patterns (Figure 4C and E). Elastasegenerated an IL-1α cleavage product very similar in mobility to the Gzm B cleavageproduct of IL-1α (Figure 4C), consistent with proteolysis at a candidate site at Ala101(Figure 4G). Mast cell chymase produced an IL-1α cleavage product running at an almostidentical mobility to the product produced by calpain cleavage (Figure 4E), consistent withcleavage at a candidate site at Phe116 (Figure 4G). In agreement with this, both enzymesalso enhanced the potency of IL-1α to an extent almost identical to Gzm B (Figure 4D andF). Thus, proteolysis of IL-1α within its N-terminus, which can be effected by multiple pro-inflammatory proteases, acts in a switch-like manner to potentiate the biological activity ofthis cytokine.
Intracellular and extracellular proteolysis of IL-1α during NK-mediated killingSeveral non-cytotoxic cell types, such as plasmacytoid dendritic cells, mast cells,keratinocytes and basophils have been reported to express Gzm B, but not perforin, uponactivation (Strik et al., 2007; Hernandez-Pigeon et al., 2007; Tschopp et al., 2006). This ispuzzling, as gene knockout studies have shown that perforin is essential for the cytotoxiceffects of granzymes (Kagi et al., 1994; Lowin et al., 1994). However, other studies havesuggested that granzymes may have extracellular roles as pro-inflammatory moleculesunrelated to their role in cell killing (Sower et al., 1996a,b; Metkar et al., 2008).Furthermore, several studies have also reported that Gzm B-expressing cells can activelysecrete this protease into the extracellular space (Spaeny-Dekking et al., 1998; Prakash et al.,2009). This suggests that Gzm B might have extracellular functions, independent of its roleas a pro-apoptotic protease, in certain settings.
To explore this possibility, we investigated IL-1α processing as a consequence of Gzm Bsecretion into the extracellular space. First, we actively permeabilized the NK cell line, YT,using the pore-forming protein, streptolysin O, or through freeze thaw-induced cell lysis topromote granzyme release. This NK line does not express all of the granzymes but doesexpress Gzm B, Gzm H and Gzm M in addition to perforin (Sedelies et al., 2004). As Figure5A shows, active permeabilization of YT cells resulted in robust IL-1α proteolysis, with theresulting cleavage fragment running at an identical mobility to that produced throughproteolysis of IL-1α by recombinant Gzm B. YT cells were also incubated in the presenceor absence of HeLa target cells to induce secretion of granzymes, followed by assessment ofthe ability of culture supernatants derived from these assays to cleave full length IL-1α. AsFigure 5B shows, proteolysis of IL-1α was readily detected using supernatants derived fromNK cells over a range of effector to target cell ratios. A time course analysis revealed thatproteolysis was already detectable within 1h of incubation of IL-1α with YT cellsupernatants, whereas incubation with HeLa cell supernatants alone failed to cleave thiscytokine (Figure 5C). Because it was formally possible that proteases other than Gzm B,released from YT cells, might be responsible for the proteolysis seen, we also exploredwhether the Gzm B-specific inhibitory serpin, PI-9, could inhibit IL-1α proteolysis withinYT-derived cell supernatants. These experiments confirmed that proteolysis of IL-1α in theextracellular space was Gzm B-dependent as this could be completely inhibited throughaddition of PI-9 to the reactions (Figure 5D). Furthermore, the Gzm B non-cleavableIL-1αD103A point mutant was completely resistant to proteolysis by YT-derived cell
Afonina et al. Page 5
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

supernatants, further confirming that the proteolysis seen was Gzm B-dependent (Figure5E).
We also explored the ability of YT cells to cleave cell-associated IL-1α during NK killing.As Figure 5F shows, intracellular IL-1α was also cleaved efficiently during target cellapoptosis. Similar results were also found using a different NK cell line, NK-92, as effectorsin similar cell killing assays (Figure 5G). Collectively, these data demonstrate thatendogenous Gzm B is capable of cleaving IL-1α, either intracellularly during NK-mediatedkilling, or as a consequence of release of both proteins into the extracellular space.
IL-1α cleaving activity in inflammatory patientsTo explore whether IL-1α also underwent processing during inflammation, we obtainedbronchoalveolar lavage fluids (BALF) from patients with persistent inflammatory lungconditions (cystic fibrosis, chronic obstructive pulmonary disease, bronchiectasis) as well ashealthy donors. Previous studies have shown that such fluids contain high levels ofinflammatory proteases, particularly elastase, due to invasion of the lungs by neutrophils andother cells of the innate immune system (McElvaney et al., 1991; Reeves et al., 2010). AsFigure 5H illustrates, IL-1α underwent rapid proteolysis to fragments very similar to theGzm B/elastase/calpain-cleaved forms of this cytokine in BALF from several cystic fibrosisand bronchiectasis patients, whereas little processing was seen in controls. Thus, proteasescapable of processing IL-1α to its mature form are secreted into the extracellular spaceunder inflammatory conditions in vivo.
Granzyme B-cleaved IL-1α exhibits enhanced activity in vivoWe next explored whether the Gzm B-cleaved form of IL-1α also exhibited greaterbioactivity in vivo. To address this question, we compared the adjuvant activity of full-length versus Gzm B-cleaved IL-1α in an antigen-driven model, using ovalbumin (OVA).Balb/c mice were immunized with OVA, either alone, or in combination with full-length orGzm B-cleaved IL-1α, followed by boosting with the same combinations two weeks later.One week after boosting, OVA-specific antibodies were measured in the serum as well asOVA-specific T-cell responses in the peritoneal lavage and spleens of these mice. As Figure6A and Figure S3 shows, both forms of IL-1α significantly boosted antibody responses toOVA, with the cleaved form of this cytokine exhibiting considerably greater potency in thisregard. Furthermore, OVA-specific IL-4, IL-5, and IL-10 production from peritoneal lavagecell preparations was also significantly greater in mice that received the Gzm B-cleavedform of IL-1α (Figure 6B). Polyclonal stimulation with anti-CD3 also produced more robustcytokine responses from mice that had received cleaved IL-1α (Figure 6C). A similar trendtowards more robust OVA-specific responses was also observed in splenocytes from thesame mice, but this was less statistically significant than in peritoneal exudates (Figure 6D).Collectively, these data lend further support to our in vitro observations that Gzm B-dependent proteolysis of IL-1α acts to enhance the pro-inflammatory effects of thiscytokine.
To again rule out the possibility that a contaminant within the Gzm B preparations couldcontribute to the enhanced IL-1α activity seen in vivo, we used the catalytically inactivemutant form of this protease (Gzm BSA; Figure S2A-C), to explore whether this mutantcould enhance IL-1α activity in vivo. As Figure 7A shows, pre-incubation with Gzm BSA
failed to potentiate IL-1α activity under conditions where incubation with wild type Gzm Bclearly did so.
We also asked whether the IL-1α mutant (D103A) incapable of undergoing processing byGzm B (Figure 1E) possessed reduced activity in vivo, when compared to wild type IL-1α.
Afonina et al. Page 6
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

As Figure 7B illustrates, IL-1α-dependent enhancement of OVA-specific antibodyresponses were consistently less pronounced with IL-1αD103A than with wild type IL-1α,suggesting that endogenous Gzm B contributes to the potency of IL-1α in vivo.
Finally, we also explored IL-1α-dependent enhancement of OVA-specific antibodyresponses in wild type versus gzmB-/- mice. Although it is important to note that severalproteases can process IL-1α to a more active form, as we have shown here, we werenonetheless interested to explore whether the absence of Gzm B had a significant impact onIL-1α activity in vivo. As Figure 7C and 7D show, we found that IL-1α-dependentenhancement of OVA-specific antibodies was greater in wild type animals than that seen ingzmB-/- mice. Note however that baseline antibody responses to OVA alone were higher ingzmB-/- animals for reasons that are unclear. Nonetheless, the enhancement effect of IL-1αon OVA-specific responses was significantly higher in wild type animals than in knockouts,again suggesting that endogenous Gzm B contributes to IL-1α processing in vivo.
Collectively, these data reveal a hitherto unappreciated consequence of IL-1α processing tothe mature ~17 kDa cytokine. The N-terminal IL-1α pro-peptide appears to function as asensor for multiple pro-inflammatory proteases, proteolysis of which switches the activity ofthis cytokine from a basal to a hyperactive state.
DISCUSSIONHere we report that IL-1α is a substrate for the CTL/NK protease Gzm B and thatproteolysis of this cytokine by the latter protease very significantly increased its biologicalpotency. These data suggest that Gzm B may enhance inflammatory responses in certainsituations through proteolytic processing of IL-1α. We have further shown that proteolysisof IL-1α by calpain-1, elastase and mast cell chymase, at sites close to but distinct from theGzm B processing site, also potentiated the activity of this cytokine. This suggests thatproteolysis of IL-1α within its N-terminus, irrespective of the protease involved, acts as aswitch to increase the biological potency of this cytokine. Thus, contrary to current thinking,limited proteolysis of IL-1α increases biological activity and serves as a further level ofregulation on this cytokine.
Proteolysis of IL-1α may induce a conformational change in this protein that increases itsaffinity for the IL-1 receptor complex, thereby increasing biological potency. Alternatively,the IL-1α N-terminus may partly occlude the receptor binding domain, removal of whichnow permits a more stable interaction with the IL-1R complex. This possibility is consistentwith a previous study that utilized sensitivity to proteinase K-mediated proteolysis toexplore whether IL-1α underwent a conformational change as a consequence of removal ofits N-terminus (Hazuda et al., 1991). Although the latter study did not explore the functionaleffects of IL-1α proteolysis in any detail, it did suggest that proteolysis of this cytokineinitiated a profound conformational change in the molecule from a proteinase K-sensitive toa proteinase K-insensitive state (Hazuda et al., 1991). This change is most likely reflected inan altered conformation that has increased IL-1 receptor affinity.
There are a number of scenarios where Gzm B may encounter IL-1α. First, in a classicalCTL/NK killing reaction, where the target cell expresses IL-1α, this cytokine may becomeprocessed within the target cell and exacerbate pro-inflammatory responses if released fromdying cells prior to clearance of the latter by phagocytes. In this context, it is relevant to notethat Bid was substantially more sensitive to Gzm B-mediated proteolysis than IL-1α. Thus,although the concentration of Gzm B achieved in a CTL/NK killing reaction is unknown, atlow nanomolar concentrations of Gzm B robust proteolysis of Bid and apoptosis might
Afonina et al. Page 7
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

occur without significant proteolysis of IL-1α, with proteolysis of the latter becoming moresignificant upon exposure to higher granzyme concentrations.
Second, there are now a number of studies suggesting that NK cells and CTLs, as well asother cell types, may actively secrete granzymes into the extracellular space (Spaeny-Dekking et al., 1998; Prakash et al., 2009). In this scenario, where injury or infection hasresulted in necrosis and release of IL-1α, Gzm B released from CTL/NK cells—or indeedone of the growing number of non-cytotoxic cell types that have been found to express thisgranzyme—could process this cytokine and enhance its potency. In this context it isnoteworthy to mention that several studies have detected circulating Gzm B at elevatedlevels in rheumatoid arthritis patients and in patients with melioidosis (Spaeny-Dekking etal., 1998; Tak et al., 1999; Lauw et al., 2000). Furthermore, Gzm B expression and secretionhas been repeatedly detected in non-cytotoxic, pro-inflammatory immune cells such asmacrophages, plasmacytoid dendritic cells, activated mast cells, basophils, B lymphocytes,macrophages, keratinocytes, platelets, human articular chondrocytes, and breast carcinomacells (reviewed in Afonina et al., 2010), further suggesting that this granzyme may havealternative, non-cytotoxic functions. Proteases such as elastase and mast cell chymase areabundantly released via degranulation of neutrophils and mast cells during inflammatoryreactions, providing a ready means by which these proteases may encounter IL-1α releasedthrough cell damage. Indeed, several recent studies have shown that IL-1α is a majorinitiator of inflammatory responses to necrotic cells (Chen et al., 2007; Kono and Rock,2008; Kono et al., 2010).
A recent study found that whereas IL-1α was readily released from cells undergoingnecrosis, this cytokine was retained within the nucleus of cells undergoing apoptosis (Cohenet al., 2010). Retention of IL-1α within the nucleus appears to be facilitated by a nuclearlocalization signal within its N-terminus, the region that is separated from the moleculethrough Gzm B-dependent proteolysis. Thus, in addition to increasing biological potencythrough enhancing IL-1α binding to its receptor, proteolysis of IL-1α by proteases such asGzm B may also facilitate its release by overriding the nuclear retention mechanisms thatappear to operate during some forms of cell death.
In summary, in addition to acting as a direct instigator of cell death upon delivery into targetcells, Gzm B may also serve in a hitherto unsuspected capacity as an amplifier ofinflammatory responses through restricted proteolysis of IL-1α. We have also shown thatproteolysis of IL-1α by several other proteases, each of which cleave this cytokine atdistinct sites within its N-terminus, enhanced the biological potency of this cytokine. IL-1αprocessing to the mature ~ 17 kDa protein acts as a switch to enhance the pro-inflammatoryproperties of this cytokine, from a basal level of activity to a hyperactive one. Thus, incommon with IL-1β, the biological activity of IL-1α is also regulated through restrictedproteolysis.
MATERIALS AND METHODSReagents
Recombinant caspase-1, -3, -4, -5, -7 were expressed and purified from bacteria as describedpreviously (Luthi et al., 2009). Recombinant poly-histidine-tagged human PI-9 wasexpressed and purified from bacteria according to standard procedures. Recombinant wildtype Gzm B (both human and murine) and the catalytically inactive mutant, Gzm BSA, wereexpressed and purified from Pichia pastoris yeast as described previously (Cullen et al.,2007). cDNA clones encoding full-length human and murine IL-1α were obtained fromOrigene and subcloned into pET45b. Recombinant IL-1α and derivatives were expressed inE. coli and purified as described in supplemental experimental procedures. Human calpain-1
Afonina et al. Page 8
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

and human mast cell chymase were purchased from Calbiochem, human elastase waspurchased from Serva. Polyclonal anti-IL-1α antibody was from AbD Serotec, goat anti-rabbit HRPO secondary antibody was from Jackson ImmunoResearch. Ac-IETD-AFC waspurchased from Bachem (UK). Lipopolysaccharides (LPS), Mannan, Lipoteichoic acid(LTA), Muramyl dipeptide (MDP) were purchased from Sigma (Ireland) Ltd., Pam3CSK4,Flagellin, CpG were purchased from Invivogen (France). Unless otherwise stated, all otherreagents were purchased from Sigma (Ireland) Ltd.
Cell cultureHeLa cells were cultured in RPMI media, supplemented with 5% fetal calf serum, YT cellsin RPMI supplemented with 10% serum and HUVEC (PromoCell, Germany) in endothelialcell growth media with added growth supplement (PromoCell, Germany). Cells werecultured at 37°C in humidified atmosphere with 5% CO2.
Cell-killing assaysHeLa cells were transiently transfected with IL-1α expression plasmid using GeneJuicetransfection reagent (Merck, Ireland). For NK-mediated killing, HeLa cells were incubatedwith YT or NK-92 cells at 20:1 and 10:1 effector to target ratios, respectively. NK cellswere removed from the target cell monolayer after 3-5 h and HeLa cells were furtherincubated for total of 24 h. Cell lysates were generated at 2 × 107 cells/ml and analysed bywestern blotting.
AnimalsBalb/c mice were purchased from Harlan (UK). Animal experiments were in accordancewith the regulations of the Trinity College Dublin ethics committee and the Irish Departmentof Health. Granzyme B null mice, on the C57BL/6 background, were bred at AcademicMedical Center (AMC), Amsterdam, The Netherlands.
Determination of cytokine and antibody concentrationsCytokines were determined by ELISA as described previously (Luthi et al., 2009). Pairedantibodies for mouse IL-4, IL-5, IL-10 and IFN-γ were purchased from BD PharMingen(UK), human IL-6, IL-8 and GM-CSF DuoSets were purchased from R&D Systems (UK).Serum immunoglobulin titres were determined as described previously (Lavelle et al.,2001).
IL-1α bioactivity assayHeLa cells were plated at a density of 4 × 104 cells per well and primary HUVECs wereplated at 2 × 104 cells per well on 24-well plates. 24 h later cells were incubated with theindicated concentrations of recombinant IL-1α, or recombinant annexin V (as a controlprotein), and culture supernatants were collected at the specified time-points for quantitativecytokine analysis by ELISA.
Bronchoalveolar lavage fluid analysisBronchoalveolar lavage fluid samples from control individuals or patients with cysticfibrosis (CF), chronic obstructive pulmonary disease (COPD) or bronchiectasis werecollected as described previously (Reeves et al., 2010). Informed patient consent wasobtained and ethical approval was granted by the Beaumont Hospital Institutional ReviewBoard.
Afonina et al. Page 9
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWork in the Martin laboratory is supported by SRC and PI grants from Science Foundation Ireland (07/SRC/B1144and 08/IN.1/B2031) and The Wellcome Trust UK (082749). The Lavelle laboratory is supported by ScienceFoundation Ireland grant 07/SRC/B1144. I.S.A. was a Health Research Board of Ireland scholar. S.J.M is a ScienceFoundation Ireland Principal Investigator.
ReferencesAfonina I, Cullen SP, Martin SJ. Cytotoxic and non cytotoxic roles of the CTL/NK proteases
granzyme B. Immunological Reviews. 2010; 235:105–116. [PubMed: 20536558]
Bertelsen M, Sanfridson A. TAB1 modulates IL-1alpha mediated cytokine secretion but is dispensablefor TAK1 activation. Cell Signal. 2007; 19:646–57. [PubMed: 17052891]
Black RA, Kronheim SR, Cantrell M, Deeley MC, March CJ, Prickett KS, Wignall J, Conlon PJ,Cosman D, Hopp TP, et al. Generation of biologically active interleukin-1 beta by proteolyticcleavage of the inactive precursor. J. Biol. Chem. 1988; 263:9437–42. [PubMed: 3288634]
Carruth LM, Demczuk S, Mizel SB. Involvement of a calpain-like protease in the processing of themurine interleukin 1 alpha precursor. J. Biol. Chem. 1991; 266:12162–7. [PubMed: 2061304]
Chen CJ, Kono H, Golenbock D, Reed G, Akira S, Rock KL. Identification of a key pathway requiredfor the sterile inflammatory response triggered by dying cells. Nat Med. 2007; 13:851–6. [PubMed:17572686]
Choy JC, McDonald PC, Suarez AC, Hung VH, Wilson JE, McManus BM, Granville DJ. Granzyme Bin atherosclerosis and transplant vascular disease: association with cell death and atheroscleroticdisease severity. Mod Pathol. 2003; 16:460–70. [PubMed: 12748253]
Cohen I, Rider P, Carmi Y, Braiman A, Dotan S, White MR, Voronov E, Martin MU, Dinarello CA,Apte RN. Differential release of chromatin-bound IL-1alpha discriminates between necrotic andapoptotic cell death by the ability to induce sterile inflammation. Proc Natl Acad Sci U S A. 2010;107:2574–9. [PubMed: 20133797]
Conlon PJ, Grabstein KH, Alpert A, Prickett KS, Hopp TP, Gillis S. Localization of humanmononuclear cell interleukin 1. J. Immunol. 1987; 139:98–102. [PubMed: 3495602]
Cullen SP, Adrain C, Luthi AU, Duriez PJ, Martin SJ. Human and Murine Granzyme B ExhibitDivergent Substrate Preferences. The Journal of Cell Biology. 2007; 176:435–444. [PubMed:17283187]
Dinarello CA. Biologic basis for interleukin-1 in disease. Blood. 1996; 87:2095–147. [PubMed:8630372]
Eigenbrod T, Park JH, Harder J, Iwakura Y, Núñez G. Cutting edge: critical role for mesothelial cellsin necrosis-induced inflammation through the recognition of IL-1 alpha released from dying cells.J. Immunol. 2008; 181:8194–8. [PubMed: 19050234]
Froelich CJ, Pardo J, Simon MM. Granule-associated serine proteases: granzymes might not just bekiller proteases. Trends Immunol. 2009; 30:117–23. [PubMed: 19217825]
Hazuda DJ, Strickler J, Simon P, Young PR. Structure-function mapping of interleukin 1 precursors.Cleavage leads to a conformational change in the mature protein. J. Biol. Chem. 1991; 266:7081–6. [PubMed: 2016316]
Hernandez-Pigeon H, Jean C, Charruyer A, Haure MJ, Baudouin C, Charveron M, Quillet-Mary A,Laurent G. UVA induces granzyme B in human keratinocytes through MIF: implication inextracellular matrix remodeling. J Biol Chem. 2007; 282:8157–64. [PubMed: 17224449]
Irmler M, Hertig S, MacDonald HR, Sadoul R, Becherer JD, Proudfoot A, Solari R, Tschopp J.Granzyme A is an interleukin 1 beta-converting enzyme. J Exp Med. 1995; 181:1917–22.[PubMed: 7722467]
Afonina et al. Page 10
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Kagi D, Ledermann B, Burki K, Seiler P, Odermatt B, Olsen KJ, et al. Cytotoxicity mediated by Tcells and natural killer cells is greatly impaired in perforin-deficient mice. Nature. 1994; 369:31–37. [PubMed: 8164737]
Kim WJ, Kim H, Suk K, Lee WH. Macrophages express granzyme B in the lesion areas ofatherosclerosis and rheumatoid arthritis. Immunol. Lett. 2007; 111:57–65. [PubMed: 17604848]
Kobayashi Y, Yamamoto K, Saido T, Kawasaki H, Oppenheim JJ, Matsushima K. Identification ofcalcium-activated neutral protease as a processing enzyme of human interleukin 1 alpha. Proc NatlAcad Sci U S A. 1990; 87:5548–52. [PubMed: 2115174]
Kono H, Karmarkar D, Iwakura Y, Rock KL. Identification of the cellular sensor that stimulates theinflammatory response to sterile cell death. J Immunol. 2010; 184:4470–8. [PubMed: 20220089]
Kono H, Rock KL. How dying cells alert the immune system to danger. Nat. Rev. Immunol. 2008;8:279–89. [PubMed: 18340345]
Kurt-Jones EA, Beller DI, Mizel SB, Unanue ER. Identification of a membrane-associated interleukin1 in macrophages. Proc. Natl. Acad. Sci. U S A. 1985; 82:1204–8. [PubMed: 3919388]
Lauw FN, Simpson AJ, Hack CE, Prins JM, Wolbink AM, van Deventer SJ, Chaowagul W, White NJ,van Der Poll T. Soluble granzymes are released during human endotoxemia and in patients withsevere infection due to gram-negative bacteria. J. Infect. Dis. 2000; 182:206–13. [PubMed:10882599]
Lavelle EC, Grant G, Pusztai A, Pfüller U, O’Hagan DT. The identification of plant lectins withmucosal adjuvant activity. Immunology. 2001; 102:77–86. [PubMed: 11168640]
Lowin B, Beermann F, Schmidt A, Tschopp J. A null mutation in the perforin gene impairs cytolytic Tlymphocyte- and natural killer cell-mediated cytotoxicity. Proc. Natl. Acad. Sci. USA. 1994;91:11571–11575. [PubMed: 7972104]
Lüthi AU, Cullen SP, McNeela EA, Duriez PJ, Afonina IS, Sheridan C, Brumatti G, Taylor RC,Kersse K, Vandenabeele P, Lavelle EC, Martin SJ. Suppression of IL-33 Bioactivity throughProteolysis by Apoptotic Caspases. Immunity. 2009; 31:84–98. [PubMed: 19559631]
Matsushima K, Taguchi M, Kovacs EJ, Young HA, Oppenheim JJ. Intracellular localization of humanmonocyte associated interleukin 1 (IL-1) activity and release of biologically active IL-1 frommonocytes by trypsin and plasmin. J Immunol. 1986; 136:2883–91. [PubMed: 2420874]
McElvaney NG, Hubbard RC, Birrer P, Chernick MS, Caplan DB, Frank MM, Crystal RG. Aerosolalpha 1-antitrypsin treatment for cystic fibrosis. Lancet. 1991; 337:392–394. [PubMed: 1671425]
Metkar SS, Menaa C, Pardo J, Wang B, Wallich R, Freudenberg M, Kim S, Raja SM, Shi L, SimonMM, Froelich CJ. Human and mouse granzyme A induce a proinflammatory cytokine response.Immunity. 2008; 29:720–33. [PubMed: 18951048]
Mizutani H, Schechter N, Lazarus G, Black RA, Kupper TS. Rapid and specific conversion ofprecursor interleukin 1 beta (IL-1 beta) to an active IL-1 species by human mast cell chymase. JExp Med. 1991; 174:821–5. [PubMed: 1919436]
Mosley B, Urdal DL, Prickett KS, Larsen A, Cosman D, Conlon PJ, Gillis S, Dower SK. Theinterleukin-1 receptor binds the human interleukin-1 alpha precursor but not the interleukin-1 betaprecursor. J. Biol. Chem. 1987a; 262:2941–4. [PubMed: 2950091]
Mosley B, Dower SK, Gillis S, Cosman D. Determination of the minimum polypeptide lengths of thefunctionally active sites of human interleukins 1 alpha and 1 beta. Proc. Natl. Acad. Sci. U S A.1987b; 84:4572–6. [PubMed: 2955410]
Prakash MD, Bird CH, Bird PI. Active and zymogen forms of granzyme B are constitutively releasedfrom cytotoxic lymphocytes in the absence of target cell engagement. Immunol. Cell Biol. 2009;87:249–54. [PubMed: 19079360]
Reeves EP, Williamson M, Byrne B, Bergin DA, Smith SG, Greally P, O’Kennedy R, O’Neill SJ,McElvaney NG. IL-8 dictates glycosaminoglycan binding and stability of IL-18 in cystic fibrosis.J Immunol. 2010; 184:1642–52. [PubMed: 20026745]
Rhim JH, Kim SA, Lee JE, Kim DJ, Chung HK, Shin KJ, Chung J. Cancer cell-derived IL-1alphainduces IL-8 release in endothelial cells. J. Cancer Res. Clin. Oncol. 2008; 134:45–50. [PubMed:17622559]
Afonina et al. Page 11
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Sedelies KA, Sayers TJ, Edwards KM, Chen W, Pellicci DG, Godfrey DI, Trapani JA. Discordantregulation of granzyme H and granzyme B expression in human lymphocytes. J Biol Chem. 2004;279:26581–26587. [PubMed: 15069086]
Sower LE, Froelich CJ, Allegretto N, Rose PM, Hanna WD, Klimpel GR. Extracellular activities ofhuman granzyme A. Monocyte activation by granzyme A versus alpha-thrombin. J Immunol.1996a; 156:2585–90. [PubMed: 8786323]
Sower LE, Klimpel GR, Hanna W, Froelich CJ. Extracellular activities of human granzymes. I.Granzyme A induces IL6 and IL8 production in fibroblast and epithelial cell lines. Cell Immunol.1996b; 10:159–63.
Spaeny-Dekking EH, Hanna WL, Wolbink AM, Wever PC, Kummer JA, Swaak AJ, et al.Extracellular granzymes A and B in humans: detection of native species during CTL responses invitro and in vivo. J. Immunol. 1998; 160:3610–6. [PubMed: 9531325]
Strik MC, de Koning PJ, Kleijmeer MJ, Bladergroen BA, Wolbink AM, Griffith JM, Wouters D,Fukuoka Y, Schwartz LB, Hack CE, van Ham SM, Kummer JA. Human mast cells produce andrelease the cytotoxic lymphocyte associated protease granzyme B upon activation. Mol. Immunol.2007; 44:3462–72. [PubMed: 17485116]
Tak PP, Spaeny-Dekking L, Kraan MC, Breedveld FC, Froelich CJ, Hack CE. The levels of solublegranzyme A and B are elevated in plasma and synovial fluid of patients with rheumatoid arthritis(RA). Clin. Exp. Immunol. 1999; 116:366–70. [PubMed: 10337032]
Thornberry NA, Bull HG, Calaycay JR, Chapman KT, Howard AD, Kostura MJ, Miller DK,Molineaux SM, Weidner JR, Aunins J, et al. A novel heterodimeric cysteine protease is requiredfor interleukin-1 beta processing in monocytes. Nature. 1992; 356:768–74. [PubMed: 1574116]
Tschopp CM, Spiegl N, Didichenko S, Lutmann W, Julius P, Virchow JC, Hack CE, Dahinden CA.Granzyme B, a novel mediator of allergic inflammation: its induction and release in bloodbasophils and human asthma. Blood. 2006; 108:2290–9. [PubMed: 16794249]
Watanabe N, Kobayashi Y. Selective release of a processed form of interleukin 1 alpha. Cytokine.1994; 6:597–601. [PubMed: 7893968]
Afonina et al. Page 12
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
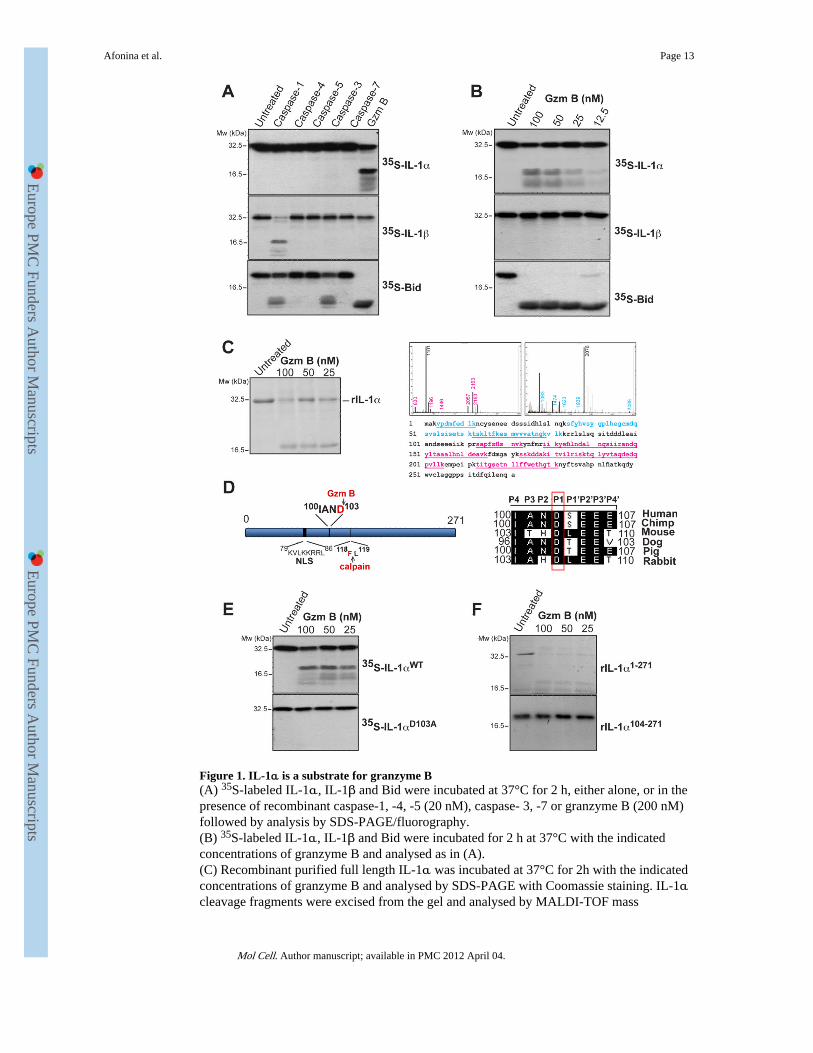
Figure 1. IL-1α is a substrate for granzyme B(A) 35S-labeled IL-1α, IL-1β and Bid were incubated at 37°C for 2 h, either alone, or in thepresence of recombinant caspase-1, -4, -5 (20 nM), caspase- 3, -7 or granzyme B (200 nM)followed by analysis by SDS-PAGE/fluorography.(B) 35S-labeled IL-1α, IL-1β and Bid were incubated for 2 h at 37°C with the indicatedconcentrations of granzyme B and analysed as in (A).(C) Recombinant purified full length IL-1α was incubated at 37°C for 2h with the indicatedconcentrations of granzyme B and analysed by SDS-PAGE with Coomassie staining. IL-1αcleavage fragments were excised from the gel and analysed by MALDI-TOF mass
Afonina et al. Page 13
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

spectrometry. Mass spectra of cleavage fragments are indicated, with coverage of the twocleavage fragments underlined in blue and red, respectively.(D) Schematic representation of IL-1α indicating nuclear localisation signal and granzymeB and calpain-1 cleavage sites. A sequence alignment of the putative granzyme B cleavagesite in IL-1α from a number of mammals is shown to the right. The P1 Asp residue isindicated.(E) 35S-labeled IL-1αWT and IL-1αD103A mutant were incubated at 37°C for 2h with theindicated concentrations of granzyme B followed by analysis by SDS-PAGE/fluorography.(F) Recombinant IL-1αFL and IL-1α104-271 were incubated for 2 h at 37°C with theindicated concentrations of granzyme B, then analysed by SDS-PAGE and Coomassiestaining. All data shown are representative of at least three independent experiments.
Afonina et al. Page 14
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
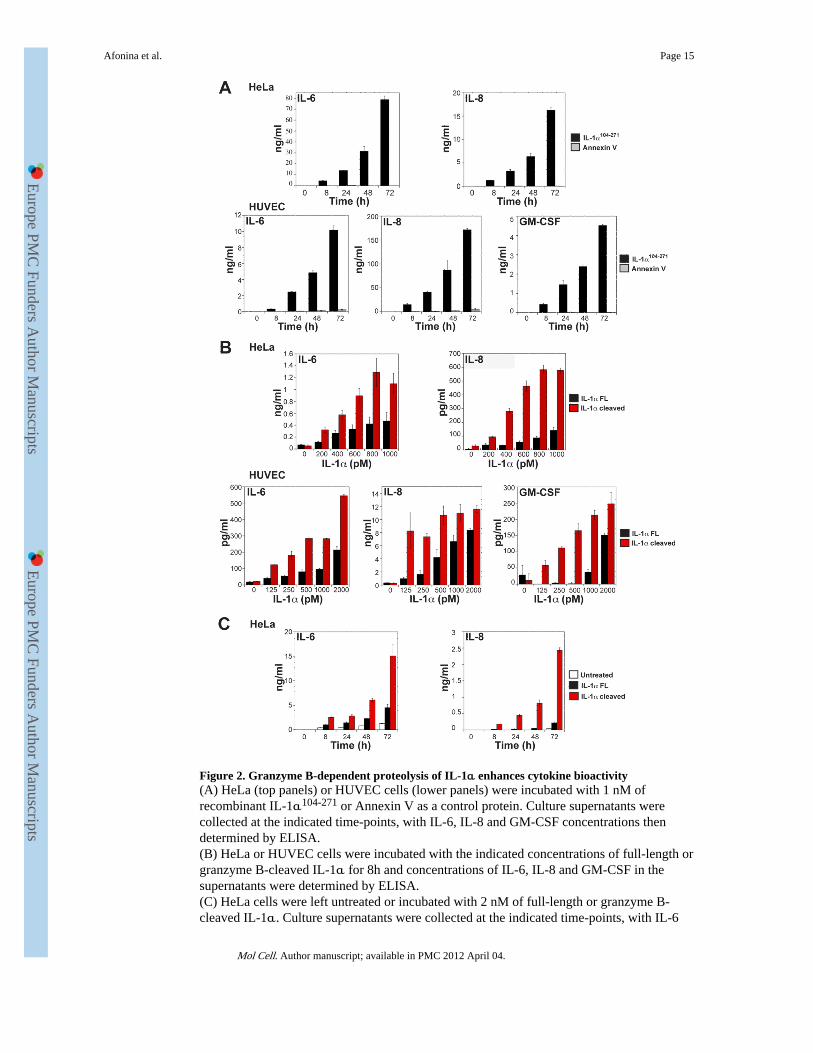
Figure 2. Granzyme B-dependent proteolysis of IL-1α enhances cytokine bioactivity(A) HeLa (top panels) or HUVEC cells (lower panels) were incubated with 1 nM ofrecombinant IL-1α104-271 or Annexin V as a control protein. Culture supernatants werecollected at the indicated time-points, with IL-6, IL-8 and GM-CSF concentrations thendetermined by ELISA.(B) HeLa or HUVEC cells were incubated with the indicated concentrations of full-length orgranzyme B-cleaved IL-1α for 8h and concentrations of IL-6, IL-8 and GM-CSF in thesupernatants were determined by ELISA.(C) HeLa cells were left untreated or incubated with 2 nM of full-length or granzyme B-cleaved IL-1α. Culture supernatants were collected at the indicated time-points, with IL-6
Afonina et al. Page 15
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

and IL-8 concentrations subsequently determined by ELISA. All results are representative ofat least three independent experiments. Error bars represent the mean ± the SEM of triplicateexperiments.
Afonina et al. Page 16
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
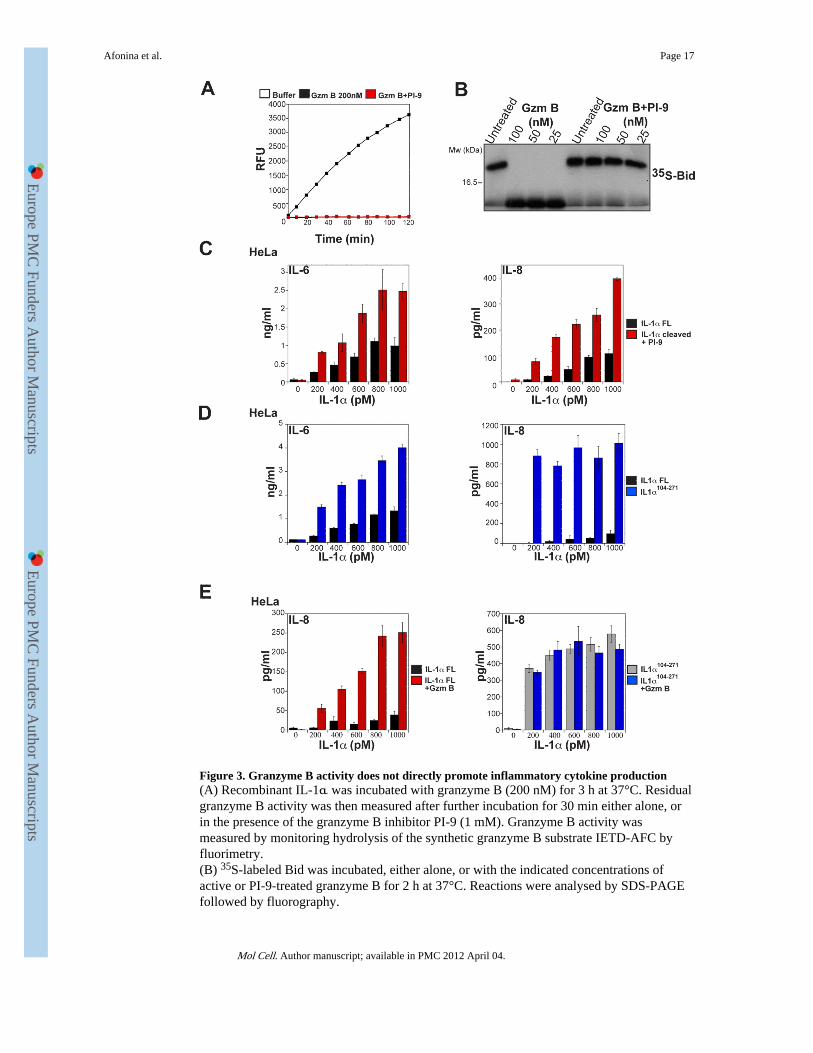
Figure 3. Granzyme B activity does not directly promote inflammatory cytokine production(A) Recombinant IL-1α was incubated with granzyme B (200 nM) for 3 h at 37°C. Residualgranzyme B activity was then measured after further incubation for 30 min either alone, orin the presence of the granzyme B inhibitor PI-9 (1 mM). Granzyme B activity wasmeasured by monitoring hydrolysis of the synthetic granzyme B substrate IETD-AFC byfluorimetry.(B) 35S-labeled Bid was incubated, either alone, or with the indicated concentrations ofactive or PI-9-treated granzyme B for 2 h at 37°C. Reactions were analysed by SDS-PAGEfollowed by fluorography.
Afonina et al. Page 17
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(C) HeLa cells were incubated for 8 h with the indicated concentrations of full-length orgranzyme B-cleaved IL-1α, where residual granzyme B activity after proteolysis of IL-1αwas inhibited by addition of saturating amounts of PI-9. IL-6 and IL-8 levels in culturesupernatants were determined by ELISA.(D) HeLa cells were incubated for 8 h with the indicated concentrations of IL-1αFL orIL-1α104-271. IL-6 and IL-8 levels were determined by ELISA.(E) HeLa cells were incubated for 8 h with the indicated concentrations of IL-1αFL,granzyme B-treated IL-1αFL, IL-1α104-271, or granzyme B-treated IL-1α104-271, asindicated. IL-6 and IL-8 levels were determined by ELISA.Results are representative of at least three independent experiments. Error bars represent themean ±SEM of determinations from three independent experiments.
Afonina et al. Page 18
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
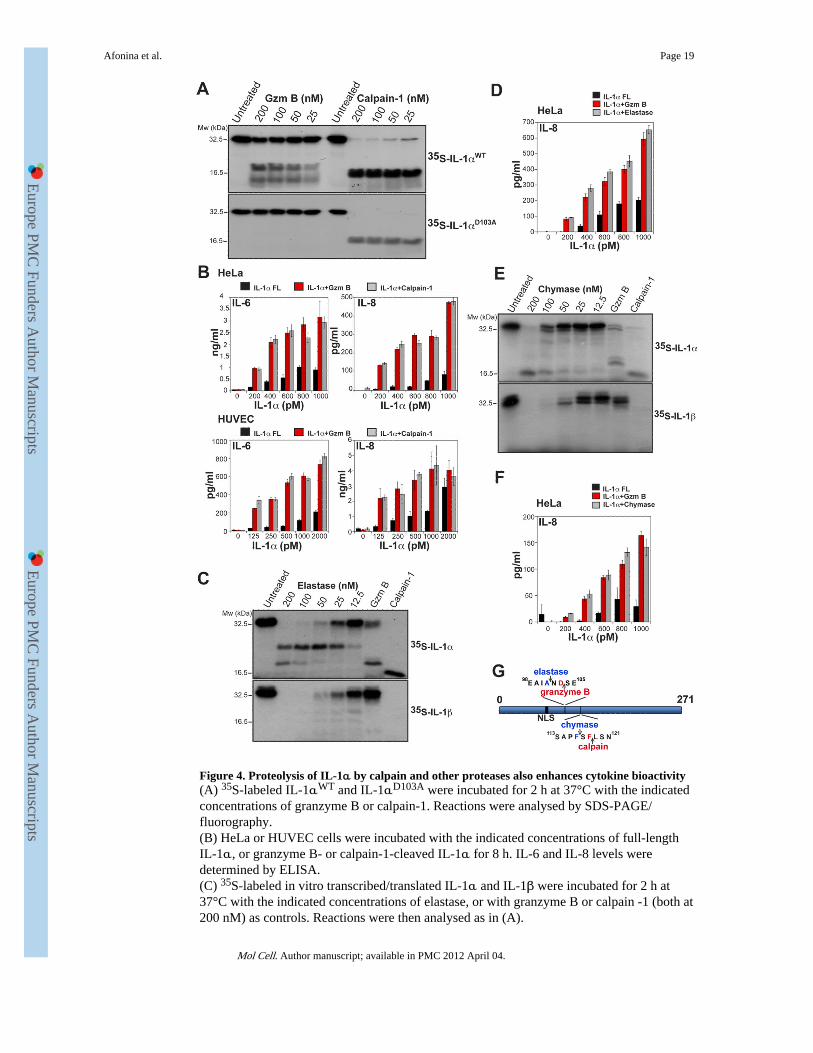
Figure 4. Proteolysis of IL-1α by calpain and other proteases also enhances cytokine bioactivity(A) 35S-labeled IL-1αWT and IL-1αD103A were incubated for 2 h at 37°C with the indicatedconcentrations of granzyme B or calpain-1. Reactions were analysed by SDS-PAGE/fluorography.(B) HeLa or HUVEC cells were incubated with the indicated concentrations of full-lengthIL-1α, or granzyme B- or calpain-1-cleaved IL-1α for 8 h. IL-6 and IL-8 levels weredetermined by ELISA.(C) 35S-labeled in vitro transcribed/translated IL-1α and IL-1β were incubated for 2 h at37°C with the indicated concentrations of elastase, or with granzyme B or calpain -1 (both at200 nM) as controls. Reactions were then analysed as in (A).
Afonina et al. Page 19
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(D) HeLa cells were incubated for 8 h with the indicated concentrations of full-length IL-1α,or IL-1α cleaved with either granzyme B, or elastase, as indicated. IL-6 and IL-8 levelswere then determined by ELISA.(E) 35S-labeled in vitro transcribed/translated IL-1α and IL-1β were incubated for 2 h at37°C with the indicated concentrations of mast cell chymase, or with granzyme B orcalpain-1 (both at 200 nM) as controls. Reactions were then analysed as in (A).(F) HeLa cells were incubated for 8 h with the indicated concentrations of full-lengthIL-1α, or IL-1α cleaved with either granzyme B, or chymase, as indicated. IL-6 and IL-8levels were then determined by ELISA.All results are representative of at least three independent experiments. Error bars representthe mean ±SEM of determinations from three experiments.(G) Schematic of IL-1α indicating sites of granzyme B- and calpain-mediated proteolysisand predicted sites of chymase- and elastase-mediated proteolysis.
Afonina et al. Page 20
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 5. IL-1α is cleaved, either extracellularly or intracellularly, by endogenous granzyme B(A) YT cells were either left untreated or were permeabilized via 2 cycles of freeze-thaw(Fr-Th), or addition of 10 μg/ml of streptolysin O (SLO) for 90 min. Recombinant IL-1αwas added to the supernatants which were further incubated for 2 h at 37°C and analysed forIL-1α processing by immunoblotting.(B) YT cells were incubated for 18 h with HeLa target cells at the indicated effector:targetratios in the presence of recombinant IL-1α. Culture supernatants were analysed byimmunoblot for IL-1α processing. Serum-derived transferrin served as a loading control.
Afonina et al. Page 21
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(C) HeLa cells were incubated with recombinant extracellular IL-1α for the indicated times,in the presence or absence of YT cells, as indicated. Supernatants were then analysed forIL-1α processing by immunoblot.(D) YT cells were incubated for 6 h with HeLa target cells at an E:T ratio of 10:1 in thepresence of recombinant IL-1α and PI-9, as indicated. Supernatants were then analysed forIL-1α processing by immunoblot. Serum-derived transferrin served as a loading control.(E) YT cells were incubated for 6 h with HeLa target cells at an E:T ratio of 10:1 in thepresence of either recombinant IL-1αWT or IL-1αD103A mutant, as indicated. Supernatantswere then analysed for IL-1α processing by immunoblot. Serum-derived transferrin servedas a loading control.(F, G) HeLa cells were transfected with IL-1α expression plasmid. 24h later cells wereexposed to YT cells (F) or NK-92 cells (G) cells at an E:T ratio of 20:1 or 5:1, respectively.NK cells were removed 3-5 h later and HeLa cells were further incubated for the total of 24h, after which cell lysates were generated and immunoblotted for the indicated proteins. Celldeath was assessed by annexin V/propidium iodide binding, measured by flow cytometry.All data shown are representative of at least three independent experiments.(H) Recombinant purified full length IL-1α was incubated at 37°C for 20 min with 10 μl ofBALF samples from control patients or patients with cystic fibrosis, bronchiectasis orchronic obstructive pulmonary disease (COPD). IL-1α processing was analysed byimmunoblot. As controls, full-length IL-1α and IL-1α104-271 were expressed in HEK293Tcells and were included to facilitate size comparison of cleavage products.
Afonina et al. Page 22
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
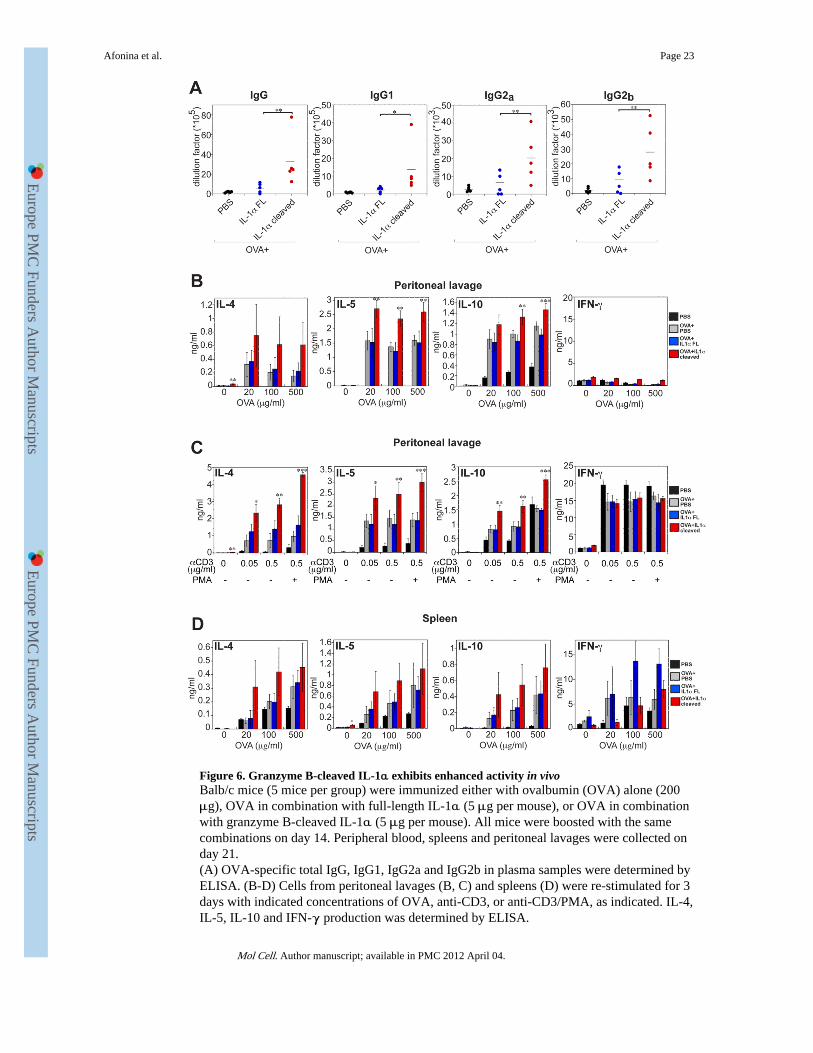
Figure 6. Granzyme B-cleaved IL-1α exhibits enhanced activity in vivoBalb/c mice (5 mice per group) were immunized either with ovalbumin (OVA) alone (200μg), OVA in combination with full-length IL-1α (5 μg per mouse), or OVA in combinationwith granzyme B-cleaved IL-1α (5 μg per mouse). All mice were boosted with the samecombinations on day 14. Peripheral blood, spleens and peritoneal lavages were collected onday 21.(A) OVA-specific total IgG, IgG1, IgG2a and IgG2b in plasma samples were determined byELISA. (B-D) Cells from peritoneal lavages (B, C) and spleens (D) were re-stimulated for 3days with indicated concentrations of OVA, anti-CD3, or anti-CD3/PMA, as indicated. IL-4,IL-5, IL-10 and IFN-γ production was determined by ELISA.
Afonina et al. Page 23
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
All measurements were taken in triplicate. Error bars represent the mean ± SEM from eachgroup of five mice. Significance levels, ***= p<.001, **= p<.05, *= p<.01, by students t-test.
Afonina et al. Page 24
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
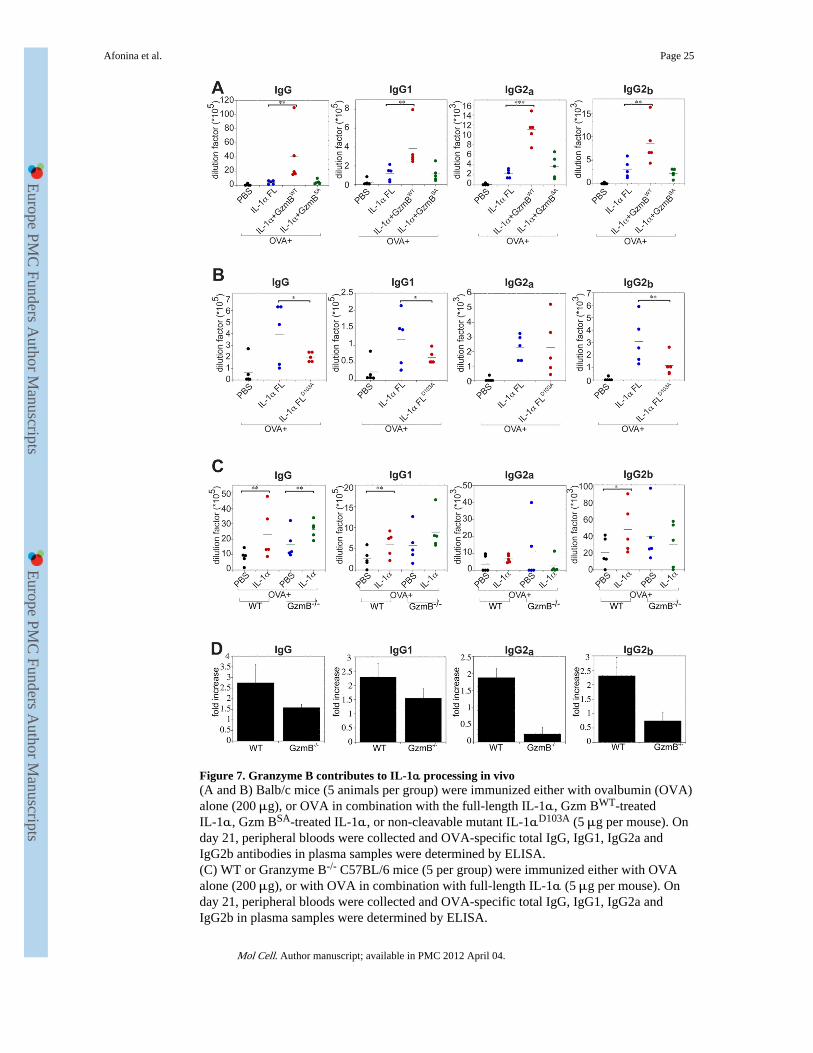
Figure 7. Granzyme B contributes to IL-1α processing in vivo(A and B) Balb/c mice (5 animals per group) were immunized either with ovalbumin (OVA)alone (200 μg), or OVA in combination with the full-length IL-1α, Gzm BWT-treatedIL-1α, Gzm BSA-treated IL-1α, or non-cleavable mutant IL-1αD103A (5 μg per mouse). Onday 21, peripheral bloods were collected and OVA-specific total IgG, IgG1, IgG2a andIgG2b antibodies in plasma samples were determined by ELISA.(C) WT or Granzyme B-/- C57BL/6 mice (5 per group) were immunized either with OVAalone (200 μg), or with OVA in combination with full-length IL-1α (5 μg per mouse). Onday 21, peripheral bloods were collected and OVA-specific total IgG, IgG1, IgG2a andIgG2b in plasma samples were determined by ELISA.
Afonina et al. Page 25
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(D) Fold increase in antibody production in WT versus Granzyme B-/- mice treated withOVA plus IL-1α, compared with OVA alone. The analysis shown is based on the datapresented in (C). Error bars represent the mean ± SEM from each group of five mice.Significance levels, ***= p<.001, **= p<.05, *= p<.01, by students t-test.
Afonina et al. Page 26
Mol Cell. Author manuscript; available in PMC 2012 April 04.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts