
Gene regulation IV Biochemistry 302 March 3,...
15
Gene regulation IV Biochemistry 302 March 3, 2006
Transcript of Gene regulation IV Biochemistry 302 March 3,...

Gene regulation IVBiochemistry 302
March 3, 2006
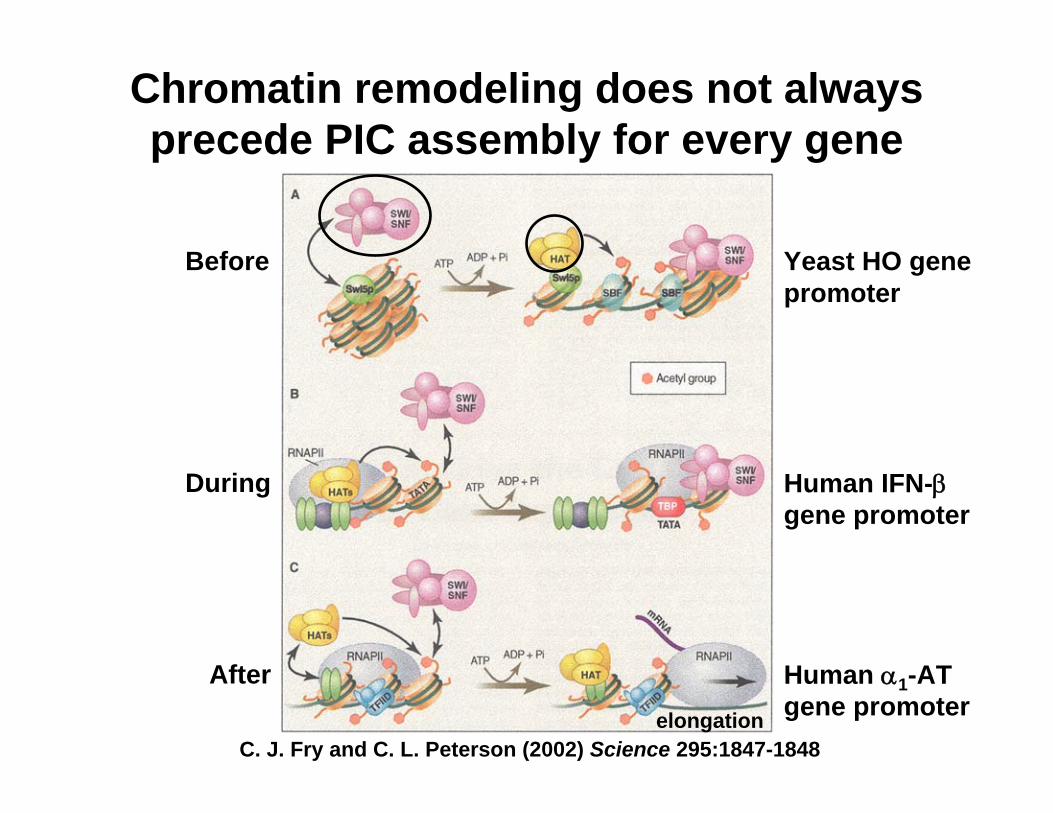
Chromatin remodeling does not always precede PIC assembly for every gene
Before Yeast HO gene promoter
Human IFN-βgene promoter
Human α1-AT gene promoter
During
After
C. J. Fry and C. L. Peterson (2002) Science 295:1847-1848elongation
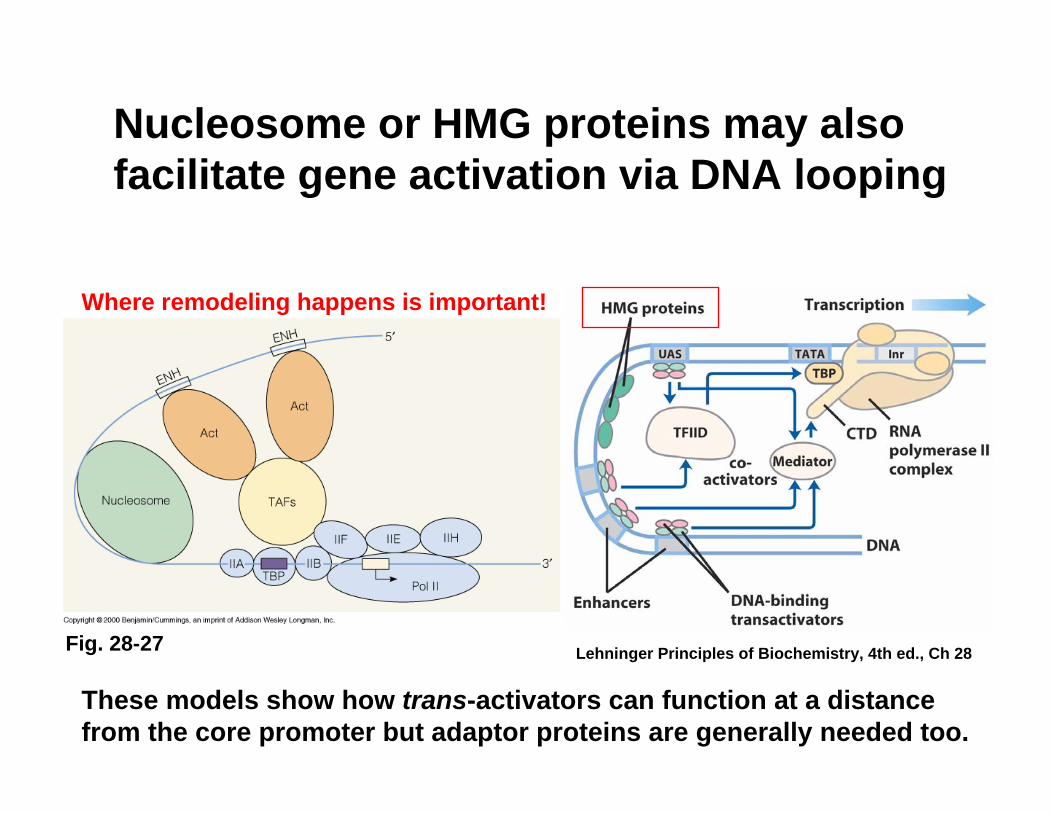
Nucleosome or HMG proteins may also facilitate gene activation via DNA looping
Fig. 28-27
These models show how trans-activators can function at a distance from the core promoter but adaptor proteins are generally needed too.
Lehninger Principles of Biochemistry, 4th ed., Ch 28
Where remodeling happens is important!

Activation machinery: trans-activators TAFs, and other cofactors• Trans-activators (Activators)
– Some bind to cis-elements proximal to the TATA box– Some bind to cis-elements distal to core promoter or
transcription start site (either 5′ or 3′)– Some exhibit cell and tissue-restricted expression.
• TAFs (TBP-associated factors, TBP + TAFs = TFIID)– Adaptor proteins - function via protein-protein interaction– Allow distal DNA-bound activators to interact with GTFs.
• Cofactors (Coactivators), usually large proteins)– Adaptor proteins - function via protein-protein interaction – Serve to link transcriptional activators to each other, to
TAFs, or other components of the GTF complex.– Some possess enzymatic activity (e.g. HATs). – Some exhibit cell and tissue-restricted expression.
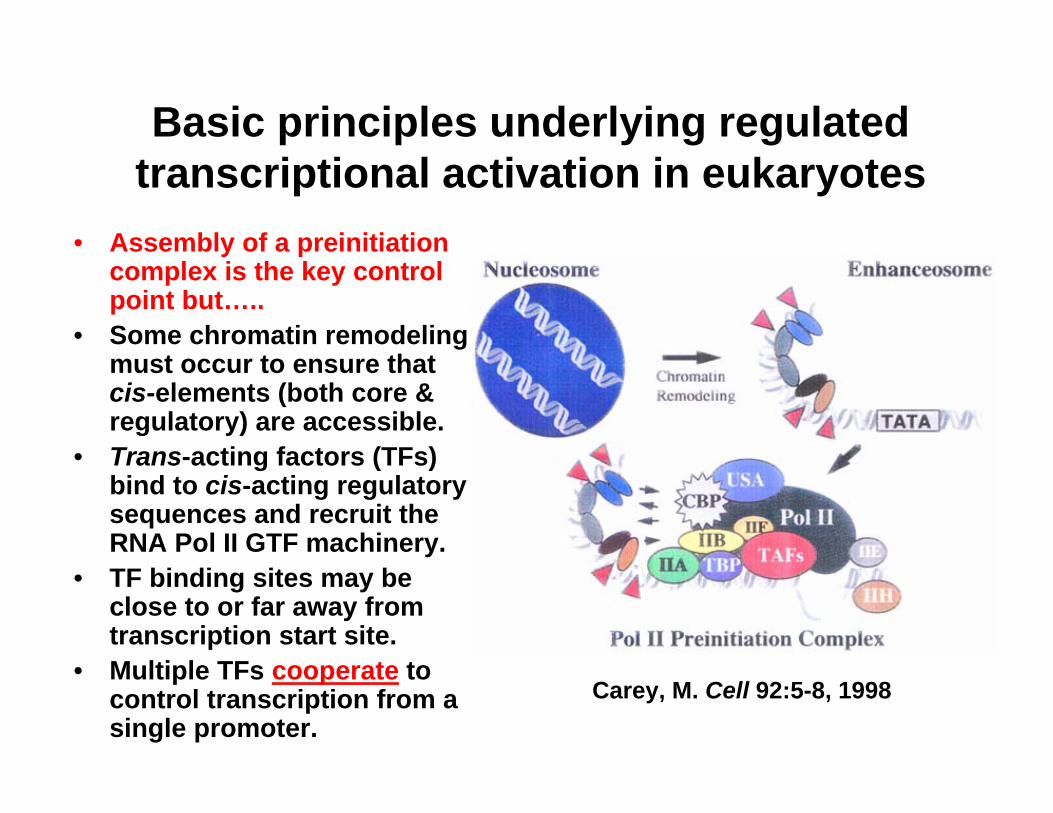
Basic principles underlying regulated transcriptional activation in eukaryotes
• Assembly of a preinitiationcomplex is the key control point but…..
• Some chromatin remodeling must occur to ensure that cis-elements (both core & regulatory) are accessible.
• Trans-acting factors (TFs) bind to cis-acting regulatory sequences and recruit the RNA Pol II GTF machinery.
• TF binding sites may be close to or far away from transcription start site.
• Multiple TFs cooperate to control transcription from a single promoter.
Carey, M. Cell 92:5-8, 1998
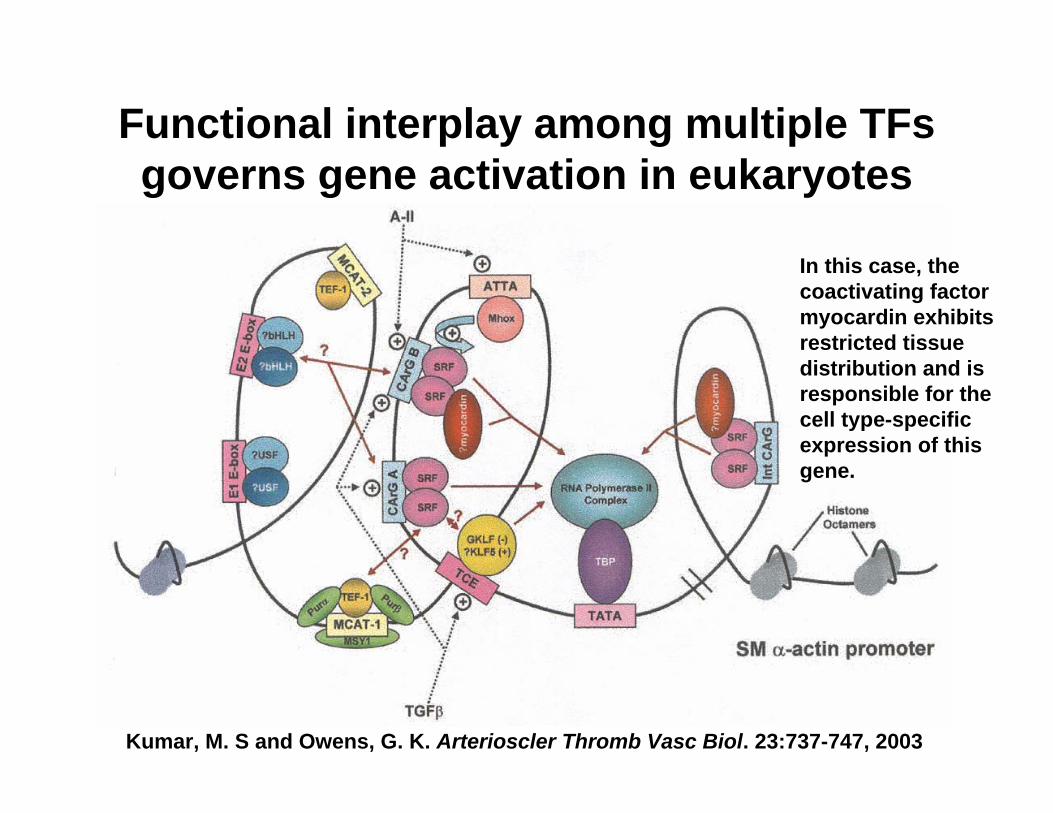
Kumar, M. S and Owens, G. K. Arterioscler Thromb Vasc Biol. 23:737-747, 2003
Functional interplay among multiple TFsgoverns gene activation in eukaryotes
In this case, the coactivating factor myocardin exhibits restricted tissue distribution and is responsible for the cell type-specific expression of this gene.

Paradigm of transcriptional activation: Recruitment of Pol II GTF machinery
• Trans-activators work by interacting with specific cis-elements to recruit the basal transcription machinery to the core promoter.
• Trans-activators also recruit chromatin remodeling factors and other coactivators.
• Direct protein-protein interaction between TF activation domainsand components of the GTF complex are necessary.
– Kd for protein-protein interaction range from ~10−4 to 10−7 M while sequence-specific DNA-binding ~10−8 to 10−10 M.
– Network of contacts among multiple activators & subunits of the RNA Pol II GTF machinery result in transcriptional synergy. Carey, M. Cell 92:5-8, 1998
Parabolic response
Sigmoidal response
K = eΔG/−RT
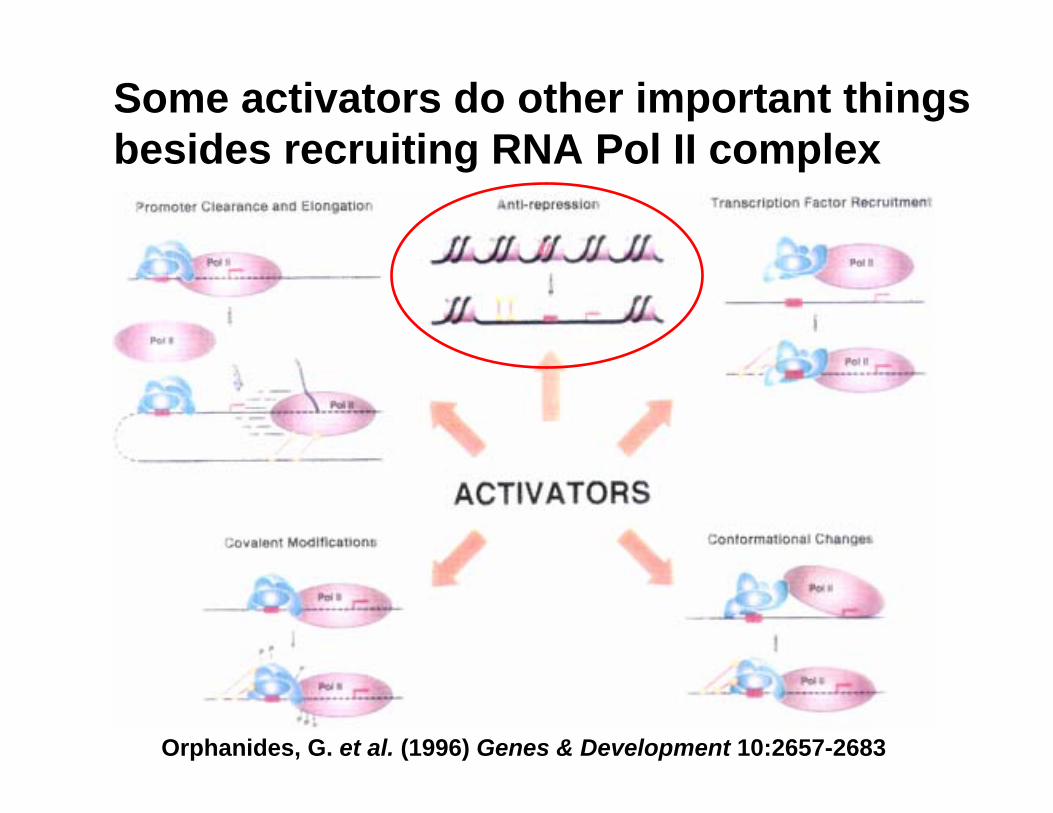
Some activators do other important things besides recruiting RNA Pol II complex
Orphanides, G. et al. (1996) Genes & Development 10:2657-2683
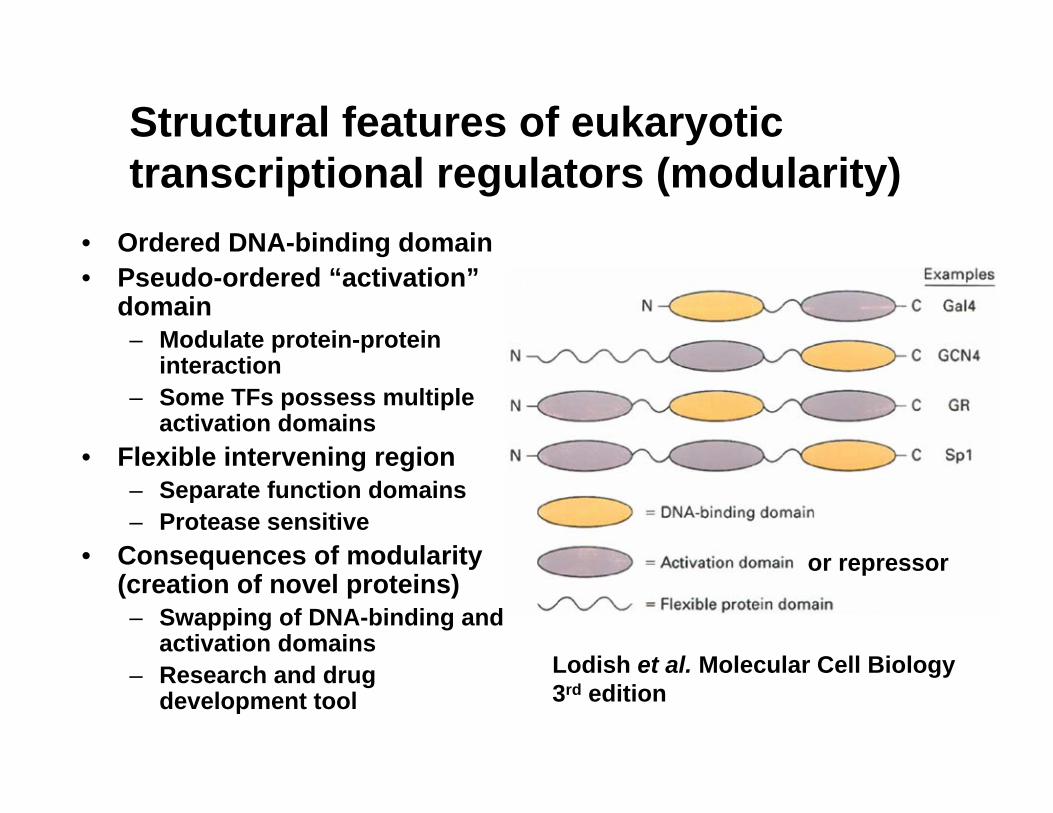
Structural features of eukaryotic transcriptional regulators (modularity)
• Ordered DNA-binding domain• Pseudo-ordered “activation”
domain– Modulate protein-protein
interaction– Some TFs possess multiple
activation domains• Flexible intervening region
– Separate function domains– Protease sensitive
• Consequences of modularity (creation of novel proteins)– Swapping of DNA-binding and
activation domains– Research and drug
development tool
or repressor
Lodish et al. Molecular Cell Biology 3rd edition
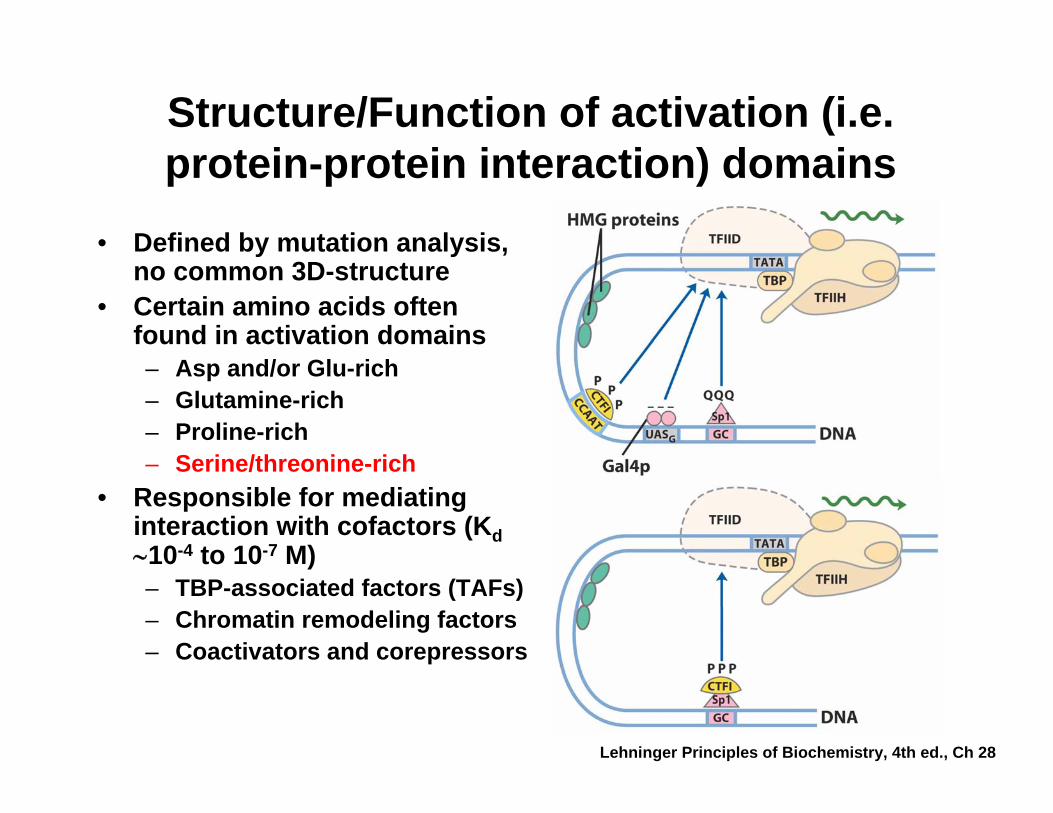
Structure/Function of activation (i.e. protein-protein interaction) domains
• Defined by mutation analysis, no common 3D-structure
• Certain amino acids often found in activation domains– Asp and/or Glu-rich– Glutamine-rich– Proline-rich– Serine/threonine-rich
• Responsible for mediating interaction with cofactors (Kd∼10-4 to 10-7 M)– TBP-associated factors (TAFs)– Chromatin remodeling factors– Coactivators and corepressors
Lehninger Principles of Biochemistry, 4th ed., Ch 28
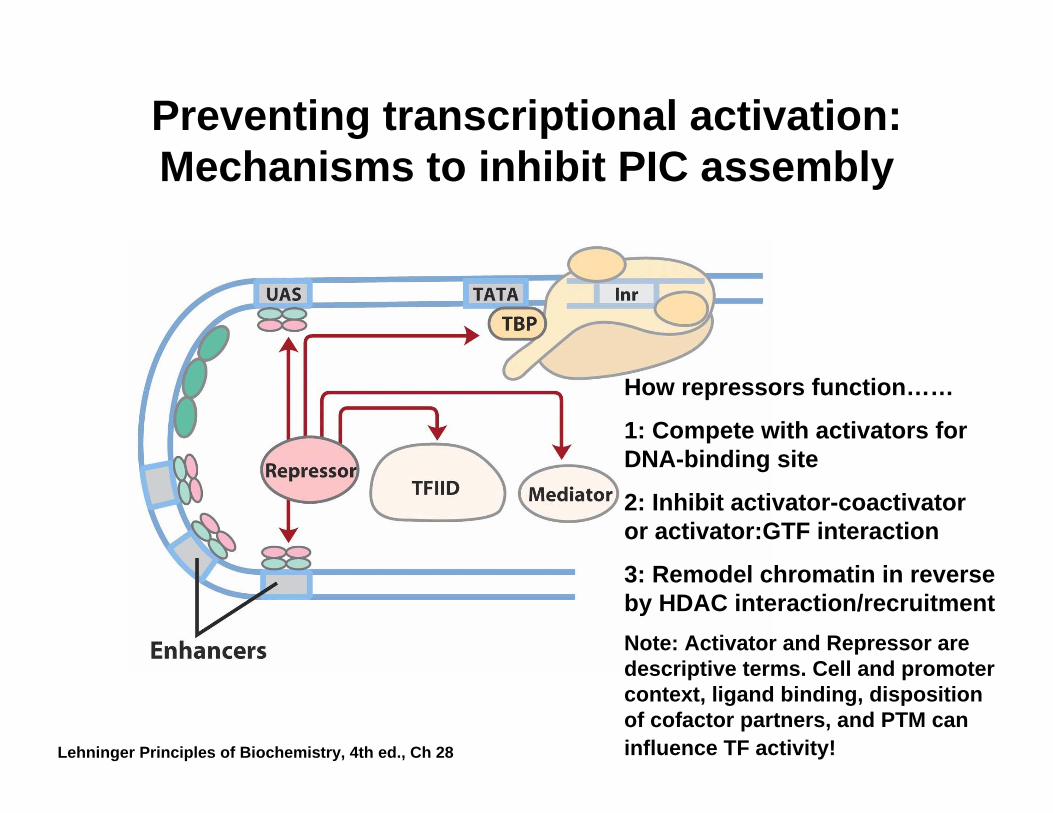
Preventing transcriptional activation: Mechanisms to inhibit PIC assembly
How repressors function……
1: Compete with activators for DNA-binding site
2: Inhibit activator-coactivator or activator:GTF interaction
3: Remodel chromatin in reverse by HDAC interaction/recruitmentNote: Activator and Repressor are descriptive terms. Cell and promoter context, ligand binding, disposition of cofactor partners, and PTM can influence TF activity!Lehninger Principles of Biochemistry, 4th ed., Ch 28
Thermodynamics and chemistry underlying protein-DNA interaction
• General themes− Steric (shape) and electrostatic (charge) compatibility
of protein and DNA− Entropy driven: binding displaces counter ions & water− High affinity (Kd ~10−9 M or less)
• Direct Interactions− Specific recognition of individual base pairs by H-
bonding between amino acids and functional groups− van der Waals contacts to thymine methyl groups − Electrostatic contacts to phosphate oxygen atoms
• Indirect Interactions− Bridging through water molecules− Induced fitting: conformational changes in protein/DNA
complex to promote binding (e.g. TBP)

Zinc finger motif: first described by A. Klug and coworkers in 1986
• Structure inferred based on aasequence analysis of Xenopuslaevis TFIIIA (37 zinc fingers)
• Crystal structure (2.1Å) of the first zinc finger of mouse Zif268 (C2H2, 30 aa) is shown.
– Two antiparallel β strands followed by an α helix, folding stabilized by coordination of a zinc ion (Zn2+) near the base
– Zinc ion is chelated by 2 histidines (from α helix) and 2 cysteines (one from β strand, one from loop)
– Multiple zinc fingers needed for high affinity interaction with DNA (core recognition element: GCG base triplet)
from from PavletichPavletich, N.P. and Sabo, C.O., N.P. and Sabo, C.O.Science Science 252:809252:809--817 (1991) 817 (1991)
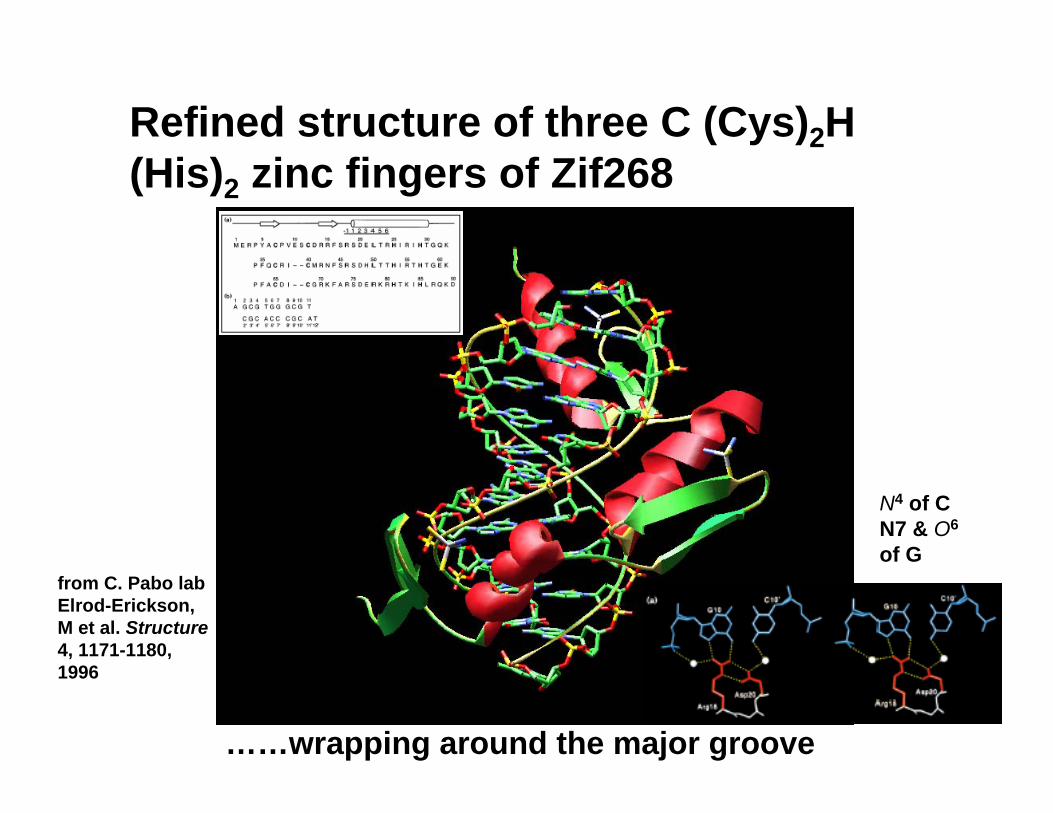
Refined structure of three C (Cys)2H (His)2 zinc fingers of Zif268
……wrapping around the major groove
from C. Pabo lab Elrod-Erickson, M et al. Structure4, 1171-1180, 1996
N4 of C N7 & O6
of G
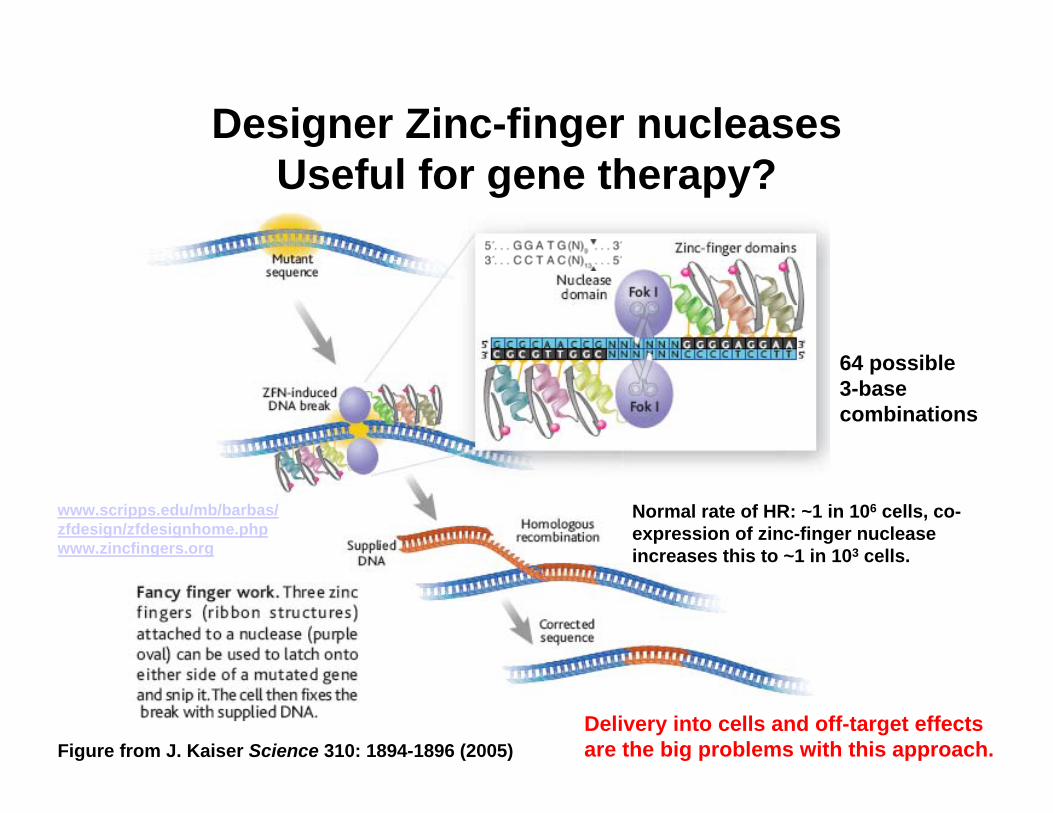
Designer Zinc-finger nucleasesUseful for gene therapy?
64 possible 3-base combinations
Delivery into cells and off-target effects are the big problems with this approach.
Normal rate of HR: ~1 in 106 cells, co-expression of zinc-finger nuclease increases this to ~1 in 103 cells.
Figure from J. Kaiser Science 310: 1894-1896 (2005)
www.scripps.edu/mb/barbas/zfdesign/zfdesignhome.phpwww.zincfingers.org


![AMINO ACIDS [QUALITATIVE TESTS] BCH 302 [PRACTICAL]](https://static.fdocument.org/doc/165x107/56649db35503460f94aa38d5/amino-acids-qualitative-tests-bch-302-practical.jpg)















