Boundary Layer - Thicknesses
5
Boundary Layer - Thicknesses Baumgart, J. and Freidrich, B.M., 2014, “Swimming across scales”, Nature Physics, Vol. 10, pp. 711 – 712. Gazzola, M., Argentina, M., and Mahadevan, L., 2014, “Scaling macroscopic aquatic locomotion”, Nature Physics, Vol. 10, pp. 758 – 761. Swimming #, Sw = ωAL/ν a b Laminar regime Amphibians Mammals Birds Fish Reptiles Turbulent regime Larvae Re ∼ Sw Re ∼ Sw 4/3 Re St St ∼ Re −1/4 St = 0.3 Sw Re 10 1 10 –1 1 10 8 10 7 10 6 10 5 10 4 10 3 10 2 10 1 10 9 10 8 10 7 10 6 10 5 10 4 10 3 10 2 10 1 1 10 9 10 8 10 7 10 6 10 5 10 4 10 3 10 2 10 1 1 L U = 2πf = tail beat frequency A ω Bolus of liquid
Transcript of Boundary Layer - Thicknesses

Boundary Layer - Thicknesses
Baumgart, J. and Freidrich, B.M., 2014, “Swimming across scales”, Nature Physics, Vol. 10, pp. 711 – 712.
Gazzola, M., Argentina, M., and Mahadevan, L., 2014, “Scaling macroscopic aquatic locomotion”, Nature Physics,
Vol. 10, pp. 758 – 761.
Swimming #, Sw = ωAL/ν
LETTERS NATURE PHYSICS DOI: 10.1038/NPHYS3078
a
b
Laminar regime
Amphibians
Mammals
Birds
Fish
Reptiles
Turbulent regime
Larvae
Re ∼ Sw
Re ∼ Sw
4/3
ReSt
St ∼ Re−1/4St = 0.3
Sw
Re
101
10–1
1
108107106105104103102101
109
108
107
106
105
104
103
102
101
11091081071061051041031021011
Figure 2 | Scaling aquatic locomotion: measurements. a, Data fromamphibians, larvae, fish, marine birds and mammals show that the scaledspeed of the organism Re=UL/⌫ varies with the scaled frequency of theoscillatory propulsor Sw=!AL/⌫ according to equations (1) and (2) overeight decades. Data fit for the laminar regime yields Re=0.03Sw1.31 withR2 =0.95, and for the turbulent regime yields Re=0.4Sw1.02 withR2 =0.99. b, The Strouhal number St= fA/U, with f =!/2⇡ , dependsweakly on Reynolds number St⇠Re�1/4 for Sw< 104 (blue) and isindependent for Sw> 104 (red), consistent with our scaling relationshipsand earlier observations30.
Because aquatic organisms live in water, testing the dependenceof our scaling relationships on viscosity requires manipulatingthe environment. Although this has been done on occasion27
and is consistent with our scaling relations (SupplementaryInformation), numerical simulations of the Navier–Stokesequations coupled to the motion of a swimming body allow us totest our power laws directly by varying Sw via the viscosity ⌫ only(Supplementary Information). In Fig. 3, we show the results fortwo-dimensional anguilliform swimmers28,29. The data from ournumerical experiments straddle both sides of the crossover from thelaminar to the turbulent regime and are in quantitative agreementwith ourminimal scaling theory, and our simple estimate for Recritical.To further challenge our theoretical scaling relationships, in Fig. 3,we plot the results of three-dimensional simulations performed byvarious groups using di�erent numerical techniques19,22,24,28; theyalso collapse onto the same power laws (details in SupplementaryInformation). The agreementwith both two- and three-dimensionalnumerical simulations, which are not a�ected by environmentaland behavioural vagaries, gives us further confidence inour theory.
Traditionally, most studies of locomotion use the Strouhalnumber St = !A/U , a variable borrowed from engineering, to
Sw = 200
Sw = 600
Sw = 2,000
Sw = 20,000
Re ∼ Sw
Re ∼ Sw
4/3Laminar
regime
104a
b
103
102
102 103 104 105101
Turbulent regime
Re
Sw
Figure 3 | Scaling aquatic locomotion: simulations. a, Two- andthree-dimensional direct numerical simulations of swimming creaturesconfirm equations (1) and (2). Circles correspond to two-dimensionalsimulations, while squares correspond to three-dimensional simulations(details about sources and numerical techniques can be found in theSupplementary Information). In the case of two-dimensional simulations, adata fit for the laminar regime yields Re=0.04Sw4/3 with R2 =0.99, andfor the turbulent regime yields Re=0.43Sw with R2 =0.99. Remarkably,three-dimensional simulations performed by various groups19,22,24,28 andwith di�erent numerical techniques (Supplementary Information) confirmour scaling relations (Re=0.02Sw4/3 with R2 = 1.00, and Re=0.26Sw withR2 =0.99). b, For several Sw we display the vorticity fields (red—positive,blue—negative) generated by a two-dimensional anguilliform swimmerinitially located on the rightmost side of the figure.
characterize the underlying dynamics. Although this is reasonablefor many engineering applications such as vortex shedding,vibration and so on, in a biological context it is worth emphasizingthat St confounds input A–! and output U variables, capturesonly one length scale by assuming A⇠ L, and does not accountfor varying fluid environments characterized by ⌫. For biologicallocomotion, Sw is a more natural variable as it captures the
760 NATURE PHYSICS | VOL 10 | OCTOBER 2014 | www.nature.com/naturephysics
© 2014 Macmillan Publishers Limited. All rights reserved
NATURE PHYSICS DOI: 10.1038/NPHYS3078 LETTERS
L
U
Re (10n)
= 2πf = tail beat frequency
L2
A 2
A
n = 1 432
a
b
c
5 87 96
ω
Thrust 2 A2 L
Skin drag ( L)1/2 U3/2
A/L
ρ
ρ ν
Pressure drag U2 L ρ
ρ
Bolus of liquid
ω
ω
Figure 1 | Aquatic swimming. a, The organisms considered here (Supplementary Information) span eight orders of magnitude in Reynolds number andencompass larvae (from mayfly to zebrafish), fish (from goldfish, to stingrays and sharks), amphibians (tadpoles), reptiles (alligators), marine birds(penguins) and large mammals (from manatees and dolphins to belugas and blue whales). Blue fish sketch by Margherita Gazzola. b, Swimmer of length Lis propelled forward with velocity U by pushing a bolus of water14,20,24 through body undulations characterized by tail beat amplitude A and frequency !.c, Thrust and drag forces on a swimmer. Thrust is the reaction force associated with accelerating (A!2) the mass of liquid per unit depth ⇢L2 weighted bythe local angle A/L (therefore ⇢LA may be understood as the mass of liquid channelled downstream). For laminar boundary layers, the drag is dominatedby viscous shear (skin drag), whereas for turbulent boundary layers, the drag is dominated by pressure (pressure drag).
As most species when swimming at high speeds maintain anapproximately constant value of the specific tail beat amplitudeA/L(refs 8,11), relation (2) reduces toU/L⇠ f , providing a mechanisticbasis for Bainbridge’s empirical relation.
In Fig. 2a, we plot all data from over 1,000 di�erentmeasurements compiled from a variety of sources (SupplementaryInformation) in terms of Re and Sw, for fish (from zebrafishlarvae to stingrays and sharks), amphibians (tadpoles), reptiles(alligators), marine birds (penguins) and large mammals (frommanatees and dolphins to belugas and blue whales). The organismsvaried in size from 0.001 to 30m, while their propulsion frequencyvaried from 0.25 to 100Hz. The dimensionless numbers we useto scale the data provides a natural division of aquatic organismsby size, with fish larvae at the bottom left, followed by smallamphibians, fish, birds, reptiles, and large marine mammals at thetop right. We see that the data, which span nearly eight orders ofmagnitude in the Reynolds number, are in agreement with ourpredictions, and show a natural crossover from the laminar powerlaw (1) to the turbulent power law (2) at a Reynolds number ofapproximately Re' 3, 000. To understand this, we note that theskin friction starts to be dominated by the pressure drag when
the thickness of the laminar boundary layer is comparable to halfthe oscillation amplitude. Therefore, a minimal estimate for thecritical Reynolds number Recritical associated with the laminar–turbulent transition is given by the relation � ' A/2. For a flatplate26 � = 5
p⌫L/U and given a typical value of A/L= 0.2, we
obtain Recritical ' (10L/A)2 = 2,500, which is in agreement withexperimental data.
Naturally, some organisms do not hew exactly to our scalingrelationships. Indeed, sirenians (manatees) slightly fall below theline, whereas anuran tadpoles lie slightly above it (SupplementaryInformation). We ascribe these di�erences to intermittent modesof locomotion involving a combination of acceleration, steadyswimming and coasting that these species often use. Other reasonsfor the deviations could be related to di�erent gaits in which part orthe entire body is used, as in carangiform or anguilliform motion.Moreover, morphological variations associated with the body, tailand fins may play a role by directly a�ecting the hydrodynamicprofile, or indirectly bymodifying the gaits. However, the agreementwith our minimal scaling arguments suggests that the role ofthese specifics is secondary, given the variety of shapes and gaitsencompassed in our experimental data set.
NATURE PHYSICS | VOL 10 | OCTOBER 2014 | www.nature.com/naturephysics 759
© 2014 Macmillan Publishers Limited. All rights reserved

Boundary Layer - Thicknesses
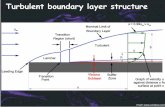
Boundary Layer Thickness Definitions 1. (99%) boundary layer thickness, d:
2. displacement thickness, dD or d*:
y
u
d
u=0.99U
U
y
u
U y
u dD
U mass flow deficit
mass flow deficit
effective displaced boundary

Boundary Layer - Thicknesses
Example: Consider the mass flow rate between two parallel plates in which a boundary layer has formed: Determine the mass flow rate through the channel in terms of the displacement thickness.
H/2
H/2
core velocity, U’
boundary layer velocity, u(y)
free stream velocity, U

Boundary Layer - Thicknesses
3. momentum thickness, dM or Q:
y
u
U y
u dM
U momentum deficit
( )ò¥=
=
-=y
y
dyuUu0
rmomentum deficit
MU dr 2=
effective displaced boundary

Boundary Layer - Thicknesses
Example: Consider the boundary layer flow over a flat plate.
Determine the drag acting on the plate in terms of the momentum thickness.
y
x
U
d
U
u
x
top of CV follows a streamline so that there is no flow out from the top of the CV
h
D


















