
Adiponectina e TNF-α no desenvolvimento de Mieloma Múltiplo · prognóstico do mieloma múltiplo...
75
Adiponectina e TNF-α no desenvolvimento de Mieloma Múltiplo Carla Susana Pessegueiro Serra Dissertação de Mestrado em Oncologia Outubro 2010
Transcript of Adiponectina e TNF-α no desenvolvimento de Mieloma Múltiplo · prognóstico do mieloma múltiplo...

Adiponectina e TNF-α no desenvolvimento de Mieloma
Múltiplo
Carla Susana Pessegueiro Serra
Dissertação de Mestrado em Oncologia
Outubro 2010

Carla Susana Pessegueiro Serra
Adiponectina e TNF-α no desenvolvimento de Mieloma Múltiplo
Dissertação de Candidatura ao grau de Mestre em Oncologia submetida ao Instituto de Ciências Biomédicas de Abel Salazar da Universidade do Porto. Orientador – Professor Doutor Rui Medeiros Categoria – Professor Associado Afiliação – Instituto de Ciências Biomédicas Abel Salazar da Universidade do Porto e Instituto Português de Oncologia de Francisco Gentil, EPE - Porto. Co-orientador – Dr. Ricardo Ribeiro Categoria – Investigador Afiliação – Instituto de Ciências Biomédicas Abel Salazar da Universidade do Porto e Instituto Português de Oncologia de Francisco Gentil, EPE - Porto.

II
Agradecimentos
Ao Prof. Dr. Rui Medeiros, meu orientador, pela oportunidade dada
de desenvolver este trabalho, todo o tempo dispensado e,
nomeadamente, todas as sugestões construtivas empreendidas ao
longo do desenvolvimento deste trabalho e a sua revisão.
Ao Dr. Ricardo Ribeiro, meu co-orientador pela ajuda prestada ao
longo de todo o trabalho, disponibilidade, incansável apoio e
paciência. Muito obrigada.
Ao Dr. Carlos Mendes um especial agradecimento por ter tornado
este projecto possível e pela incansável ajuda. Muito obrigada.
Ao Prof. Dr. Carlos Lopes por todo o apoio prestado, pelo seu
carinho e amabilidade.
Ao Dr. Luís Araújo e Dr. Mário Mariz pela possibilidade de ter
tornado este projecto possível e por toda disponibilidade prestada.
Ao Dr. Luís Leite e Dr. Nuno Duarte por toda disponibilidade e ajuda
prestada ao longo do projecto.
Ao meu amigo Leandro por toda a preocupação e ajuda prestada e
pela grande amizade que nos une.
Às colegas do grupo de Oncologia Molecular Cátia, Célia e Virgínia
e às colegas de trabalho Ana Aguiar, Carina, Filomena, Raquel e
Ana Paula pelo apoio e pela sempre prontidão em ajudar.
À minha família sempre presente, em especial à minha mãe e irmã
por todo o apoio, confiança e paciência.

III
Resumo
Alguns estudos têm vindo a demonstrar associação entre obesidade e mieloma
múltiplo. Esta associação parece dever-se à modificação do padrão de produção de
adipocinas em doentes obesos, nomadamente a adiponectina e o factor de necrose
tumoral α (TNF-α), que parecem apresentar um papel no desenvolvimento de vários
tumores sólidos, assim como em doenças do foro hematológico, tais como as leucemias
e o mieloma múltiplo.
Neste estudo, foram avaliados os níveis circulantes de adiponectina e TNF-α no
sangue periférico e a sua concentração na medula óssea, no momento do diagnóstico e
após tratamento, com o intuito de esclarecer a sua associação com a patogénese e
prognóstico do mieloma múltiplo e compreender os mecanismos envolvidos.
Foram utilizadas amostras de sangue periférico e de líquido intersticial medular
(LIM) de 13 indivíduos com mieloma múltiplo (MM) ou plasmocitoma, 8 com gamapatia
monoclonal de significado indeterminado (MGUS) e 12 controlos emparelhados por idade
e género. As amostras foram colhidas entre Janeiro e Junho de 2010, tendo sido
doseados os níveis de adiponectina total (tAd), globular (gAd) e TNF-α no plasma e no
LIM, no diagnóstico e os níveis de adiponectina total (tAd), globular (gAd) no plasma, 1
mês após tratamento. O diagnóstico foi obtido de acordo com os critérios clássicos
morfológicos, clínicos e laboratoriais e a classificação do estádio segundo o sistema
internacional de estadiamento (ISS).
Não se observaram diferenças com significado estatístico entre MM e MGUS para
níveis circulantes de tAd e gAd (p>0,05). Na análise efectuada de acordo com o
estadiamento ISS verificamos ausência de associação com a adiponectina no líquido
intersticial mas tendência para níveis sanguíneos mais baixos de gAd nos indivíduos com
estádios ≥2 (p=0,094), sugestivo de pior prognóstico, assim como para o TNF-α (8,8 ±
4,0; 38,7 ± 9,9 p<0,036). Foi observada correlação, no momento do diagnóstico, entre os
níveis de TNF-α doseado LIM e no plasma de doentes com MM (r = 0,942; p = 0,002).
Também foram presenciadas correlações inversas entre os níveis de TNF-α do LIM com
a concentração de adiponectina globular plasmática, (r = -0,810; p=0,027) e entre o
número de plasmócitos contabilizados morfologicamente em lâminas de medula óssea,
com a adiponectina globular intersticial (r = -0,680; p = 0,031). Considerando estas

IV
correlações é possível que a adiponectina possa indirectamente diminuir o crescimento
dos plasmócitos através do seu efeito supressor na produção de TNF-α e
consequentemente diminuição da viabilidade dos plasmócitos.
Não foi detectada a expressão dos receptores de adiponectina nos plasmócitos ou
nas células de linhagem linfóide, sugerindo uma eventual acção indirecta da
adiponectina, ou alternativamente, esta poderá apenas representar o estado metabólico
associado à obesidade, podendo outras adipocinas possuir maior relevância na sua
acção nos plasmócitos.
Para confirmar as associações encontradas serão necessários estudos com
amostragens mais significativas. Os nossos resultados ajudam a explorar os mecanismos
que poderão envolver estas adipocinas e o mieloma múltiplo.

V
Abstract
Some studies have come to demonstrate the association between obesity and
multiple myeloma. This association appears to be due to a change in the production of
citokines in the state of obesity, particularly adiponectin and TNF-α seem to take part in
the development of some solid tumors and in leukemia.
In this study, were evaluated the adiponectin and TNF-α circulating levels in the
peripheral blood and its concentration in bone marrow, at the diagnosis and after
treatment, in order to clarify its association with the pathogenesis and prognosis of
multiple myeloma and understand the mechanisms involved.
Samples of peripheral blood and medular interstitial liquid of 13 individuals with
myeloma (MM) or plasmacytoma, 8 with monoclonal gammopathy of undetermined
significance (MGUS) and 12 controls paired up for age and gender had been used. These
samples were collected between January and June of 2010, and assayed the levels of
total (tAd) and globular adiponectin (gAd) and TNF-α in the plasma and interstitial liquid of
the bone marrow, at the diagnosis and 1 month after treatment. The diagnosis was made
according to morphologic, clinical and laboratorial traditional criteria and the stage
classification according to international system for staging (ISS).
There were no statistically significant differences between MM and MGUS
circulating levels of tAd and gAd (p> 0.05). In the analysis carried out in accordance with
the ISS stage, we found no association with adiponectin in the interstitial liquid but a trend
to lower blood levels of gAd in individuals with stage ≥2 (p=0,094), suggesting a worse
prognosis, as well as for TNF-α (8,8 ± 4,0; 38,7 ± 9,9 p<0,036). A correlation was
observed at the time of diagnosis between TNF-α levels assayed in the medular interstitial
liquid and in plasma of patients MM (r = 0.942, p = 0.002). We also observe inverse
correlations between levels of TNF-α in bone marrow interstitial fluid and the plasma
concentration of globular adiponectin (r = -0.810, p = 0.027) and between the number of
plasma cells accounted in bone marrow slides, with interstitial globular adiponectin (r = -
0.680, p = 0.031).Considering these correlations is possible that adiponectin may
indirectly reduce the growth of plasmocytes by its suppressive effect on TNF-α production
and subsequent decline of plasmocytes viability.
It was not detected the expression of adiponectin receptors in the plasmocytes or
cells of lymphoid lineage, suggesting a possible indirect action of adiponectin, or

VI
alternatively, it can only represent the metabolic state associated with obesity; other
adipokines may have greater relevance in their action in plasmocytes.
To confirm these associations, studies with more significant number of samples
have to be performed. Our results help to explore the mechanisms that may involve these
adipokines with multiple myeloma.

VII
Lista de abreviaturas
ACC - Carboxilase acetil-CoA
ADN – Ácido desoxiribonúcleico
AMPK - Proteína cinase activada
bFGF - Factor de crescimento dos fibroblastos
BJP - Proteínas de bence-jones
CCL-2 - Quimiocina C-C-2
CXCR4- Receptor de quimiocinas CXC 4
EMP - Plasmocitoma extramedular
FAZ - Sintase de ácidos gordos
gAd ou gAcrp30 - Adiponectina globular
HB EGF - Factor de crescimento epidérmico de ligação à heparina
HDL – Colesterol de alto peso molecular
HGF - Factor de crescimento da heparina
HMW - Adiponectina de alto peso molecular
ICAM-1- Molécula de adesão intercelular
IFN-γ – Interferão γ
Ig – Imunoglobulina
IL-1β – Interleucina 1β
IL-3 – Interleucina 3
IL-6 - Interleucina 6
IMC – Índice de massa corporal
ISS - Sistema internacional de estadiamento
LFA-1 - Antigénio associado ao leucócito
LIM - Líquido intersticial medular
LMA-M1 – Leucemia mieloblástica aguda M1
LMW - Adiponectina de baixo peso molecular
MCP-1- Proteína quimio-atractora de monócitos 1
MGUS - Gamopatia monoclonal de significado indeterminado
MIF - Factor de migração inibitório
MIP-1α - proteína inflamatória macrófagos
MM - Mieloma múltiplo

VIII
MMW – Adiponectina de médio peso molecular
NF-kB - Factor nuclear-kB
PAI-1 - Inibidor do activador do plasminogénio 1
PCR - Proteína C reactiva
PDGF-BB - Factor de crescimento derivado de plaquetas BB
PPAR α - Receptor do activador da proliferação de peroxisomas α
PPAR γ - Receptor do activador da proliferação de peroxisomas γ
RPM – Rotações por minuto
SBP - Plasmocitoma ósseo solitário
tAd ou Acrp30 –Adiponectina total
TBS - tris buffered saline
TNF-α – Factor de necrose tumoral α
TZDs - Tiazidas
VCAM-1 - Molécula de adesão celular vascular
VEGF - Factor de crescimento do endotélio vascular
VF - Visfatina
VLA- 4 - Antigénio muito tardio
WHO - World Health Organization

IX
Índice
Agradecimentos .............................................................................................................. II
Resumo ...........................................................................................................................III
Abstract ........................................................................................................................... V
Lista de abreviaturas .................................................................................................... VII
Índice de figuras ............................................................................................................ XI
Índice de quadros ......................................................................................................... XII
Índice de gráficos ........................................................................................................ XIII
1. Introdução ................................................................................................................... 1
1.1 Neoplasias: Aspectos gerais e biopatologia ............................................................ 2
1.2 Células plasmáticas: Conceito e fisiopatologia ........................................................ 3
1.3 Neoplasias de células plasmáticas: Gamapatias monoclonais ................................ 6
1.3.1 Mieloma Múltiplo: Características gerais e critérios de diagnóstico ................... 9
1.3.2 Plasmocitoma: Características gerais e critérios de diagnóstico ......................10
1.3.3 MGUS: Características gerais e critérios de diagnóstico ..................................11
1.4 Tecido adiposo: Conceito e fisiopatologia...............................................................13
1.5 Adiponectina ..........................................................................................................17
1.5.1 Caracterização molecular e genética ...............................................................17
1.5.2 Receptores ADIPOR1 e ADIPOR2 ..................................................................18
1.5.3 Caracterização fisiológica ................................................................................19
1.6 Adiponectina e as neoplasias .................................................................................22
1.7 Obesidade, adiponectina - mieloma múltiplo / MGUS/plasmocitoma ......................25
2. Objectivos ..................................................................................................................26
3. Material e Métodos .....................................................................................................28
3.1 População ..............................................................................................................29
3.2 Processamento de amostras ..................................................................................30

X
3.3 Análise estatística ..................................................................................................31
4. Resultados .................................................................................................................32
5. Discussão ...................................................................................................................40
6. Conclusão e perspectivas futuras ............................................................................47
7. Referências ................................................................................................................50

XI
Índice de figuras
Figura 1: Processo de captação dos precursores das células plasmáticas do sangue
periférico para a medula óssea (Harousseau and Moreau, 2009). .................................... 4
Figura 2: Interacções entre as células plasmáticas neoplásicas e o micro ambiente da
medula óssea (Harousseau and Moreau, 2009).. ............................................................. 5
Figura 3: Principais proteínas encontradas em cada banda eletroforética (Silva, 2008). .. 6
Figura 4: Mecanismos oncogénicos envolvidos na diferenciação das células plasmáticas.
Adaptado de:(Kyle and Rajkumar, 2004). ......................................................................... 7
Figura 5: Patogénese e progressão de gamapatia monoclonal de significado
indeterminado para mieloma múltiplo. Adaptado de:(Rajkumar, 2005). ...........................12
Figura 6: Tecido adiposo: componentes celulares, moléculas sintetizadas e seu
envolvimento com a inflamação (Tilg and Moschen, 2008). .............................................15
Figura 7: Factores contribuintes para o aumento do risco da obesidade associada a
neoplasias. Adaptado de: (Barb et al., 2007). ..................................................................16
Figura 8: Estrutura e formas oligoméricas da adiponectina total (Kadowaki and
Yamauchi, 2005). .............................................................................................................17
Figura 9: Estrutura dos receptores de adiponectina (Kadowaki and Yamauchi, 2005). ...18
Figura 10: Mecanismos de expressão da adiponectina pela activação dos agonistas do
receptor do activador da proliferação de peroxisomas (Tilg and Moschen, 2008). ...........21
Figura 11: Possíveis mecanismos moleculares implicados na regulação do crescimento
das células tumorais pela proteína kinase activada por 5´-AMP. Adaptado de (Kelesidis et
al., 2006). .........................................................................................................................24
Figura 12: Imunocitoquímica para os anticorpos ADIPOR1 (a) e ADIPOR2 (b) em
esfregaço de medula óssea de doentes com mieloma múltiplo. ......................................39

XII
Índice de quadros
Quadro 1: Sistema internacional de estadiamento (ISS). Adaptado de: (Silva, 2009). ..... 8
Quadro 2: Classificação internacional de alteração de peso de acordo com o Índice de
Massa Corporal. Adaptado de (OMS, 2004). ...................................................................14
Quadro 3: Características clínicas e analíticas dos doentes com mieloma múltiplo e
gamapatia monoclonal de significado indeterminado .......................................................33
Quadro 4: Associação entre a obesidade e os casos de mieloma múltiplo, gamapatia
monoclonal de significado indeterminado ........................................................................34
Quadro 5: Concentração no sangue periférico de adiponectina total, globular e TNF-α em
cada um dos grupos no momento de diagnóstico ............................................................34
Quadro 6: Concentração de adiponectina total, globular e TNF-α nos casos e nos
controlos no momento do diagnóstico no LIM ..................................................................35
Quadro 7: Concentração de adiponectina total, globular e TNF-α nos casos de mieloma
múltiplo em sangue periférico e LIM no momento de diagnóstico, de acordo com o
estadiamento ISS ............................................................................................................36
Quadro 8: Comparação dos níveis de adiponectina total e globular, nos casos de
mieloma múltiplo, em sangue periférico no momento do diagnóstico e após 1 mês de
tratamento .......................................................................................................................37

XIII
Índice de gráficos
Gráfico 1: Correlação entre adiponectina total (a), globular (b) e TNF-α (c) doseados no
LIM e no plasma de doentes com mieloma múltiplo, no momento de diagnóstico ...........38

1. Introdução

2
1. Introdução
1.1 Neoplasias: Aspectos gerais e biopatologia
Nos países desenvolvidos, as doenças neoplásicas são consideradas a segunda
causa de morte sendo ainda responsáveis pela elevada taxa de morbilidade. As
estimativas da Organização Mundial de Saúde até 2030 sugerem aumentos na incidência
e mortalidade (OMS, 2008).
Actualmente, parece estar estabelecido que o diagnóstico tardio das neoplasias
se encontra associado a uma maior mortalidade. Assim, a identificação de grupos de
risco e o estabelecimento de novos biomarcadores são fundamentais para o diagnóstico
precoce e orientação terapêutica.
As neoplasias são doenças complexas, sabendo-se que na sua etiopatogenia
estão envolvidos factores genéticos e ambientais. As neoplasias malignas caracterizam-
se pela divisão celular descontrolada que escapam ao controle dos factores que,
normalmente, regulam a sua proliferação e maturação. A progressão tumoral é, na
maioria dos casos, lenta e envolve a acumulação de mutações em proto-oncogenes,
genes supressores tumorais, genes associados à regulação da apoptose ou genes
envolvidos na reparação do ADN (Lengauer et al., 1998).
Segundo (Hanahan and Weinberg, 2000), as células tumorais apresentam seis
características essenciais que permitem o seu desenvolvimento e progressão: auto-
suficiência em factores de crescimento, insensibilidade a sinais inibidores de crescimento,
potencial replicativo ilimitado, fuga à apoptose, capacidade de invasão tecidular e a
metastização. Ao longo dos últimos anos tem-se vindo a acumular evidências sobre o
papel do micro ambiente na carcinogénese, estando estabelecida a interacção entre
células tumorais e células estromais envolventes (Rajala and Scherer, 2003; Finak et al.,
2008).

3
1.2 Células plasmáticas: Conceito e fisiopatologia
As células plasmáticas, também denominadas de plasmócitos, são linfócitos B
totalmente diferenciados que desempenham funções específicas de produção e secreção
de imunoglobulinas com o objectivo de combater e eliminar os agentes causadores de
infecção, como as bactérias ou os vírus (Radbruch et al., 2006; Alexander et al., 2007).
As células precursoras das células plasmáticas, tal como todas as células do
sistema hematopoiético, têm uma origem comum na medula óssea, a partir de uma stem
cell pluripotente (figura 1). No entanto, durante a fase terminal de diferenciação
desencadeada nos nódulos linfáticos poderão ocorrer erros genéticos a nível das células
precursoras, os plasmablastos (Harmening, 1997; Greipp et al., 2005; Radbruch et al.,
2006).
Os plasmablastos mutados gerados são transportados através do sangue
periférico para a medula óssea, onde serão produzidos clones. O mecanismo sob o qual
estas células precursoras são deslocadas para a medula óssea ainda se encontra por
clarificar. Contudo, foi proposto que a ligação da proteína quimio-atractora de monócitos
1 (MCP-1), expressa em células estromais, ao receptor de quimiocinas CXC 4 (CXCR4),
expresso nos plasmablastos parece desempenhar um papel importante na migração e
afinidade medular. O microambiente da medula óssea parece estar organizado para
capturar precursores das células tumorais circulantes, oferecendo-lhes condições
favoráveis para a sua diferenciação e expansão clonal (Caligaris-Cappio et al., 1985).
Assim, a MCP-1 activa os plasmablastos CXCR4 positivos, estimulando a sua adesão ao
endotélio através de moléculas de adesão pelo antigénio associado ao leucócito (LFA-1)
e pelo antigénio muito tardio (VLA-4). Desta forma, os plasmablastos conseguem migrar
para a medula óssea e assim ficarem expostos às células estromais (Radbruch et al.,
2006; Harousseau and Moreau, 2009) (figura 2).
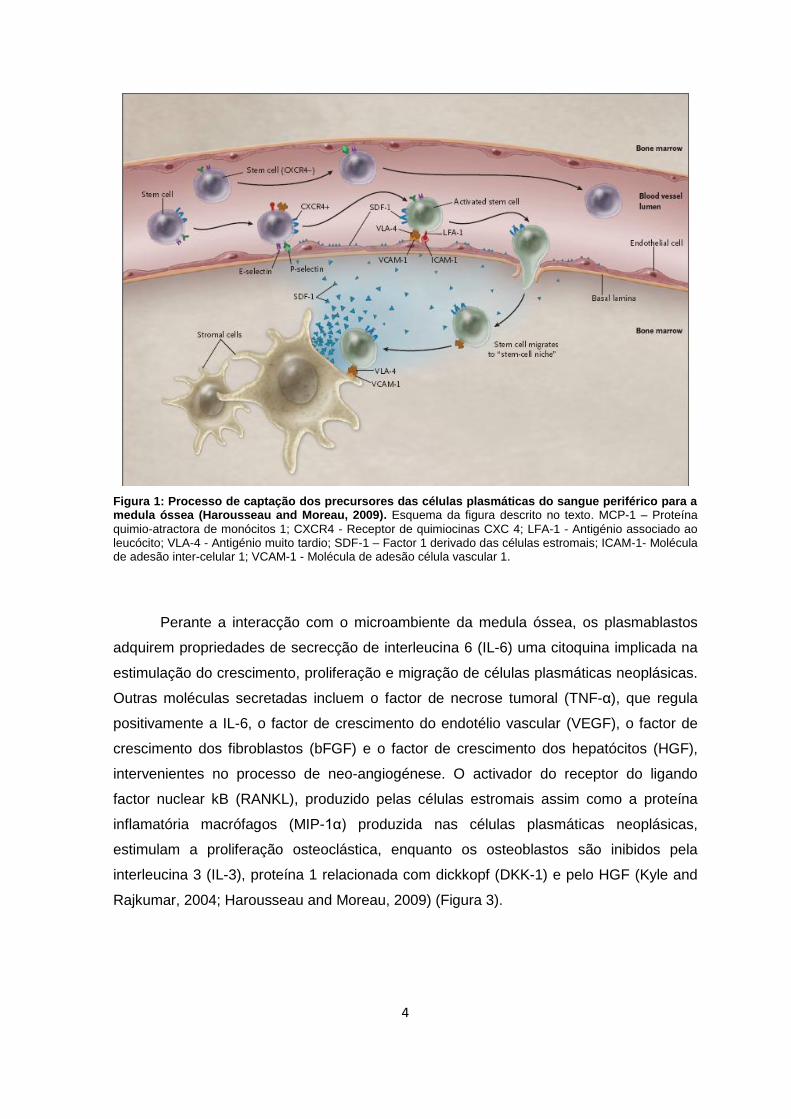
4
Figura 1: Processo de captação dos precursores das células plasmáticas do sangue periférico para a medula óssea (Harousseau and Moreau, 2009). Esquema da figura descrito no texto. MCP-1 – Proteína
quimio-atractora de monócitos 1; CXCR4 - Receptor de quimiocinas CXC 4; LFA-1 - Antigénio associado ao leucócito; VLA-4 - Antigénio muito tardio; SDF-1 – Factor 1 derivado das células estromais; ICAM-1- Molécula de adesão inter-celular 1; VCAM-1 - Molécula de adesão célula vascular 1.
Perante a interacção com o microambiente da medula óssea, os plasmablastos
adquirem propriedades de secrecção de interleucina 6 (IL-6) uma citoquina implicada na
estimulação do crescimento, proliferação e migração de células plasmáticas neoplásicas.
Outras moléculas secretadas incluem o factor de necrose tumoral (TNF-α), que regula
positivamente a IL-6, o factor de crescimento do endotélio vascular (VEGF), o factor de
crescimento dos fibroblastos (bFGF) e o factor de crescimento dos hepatócitos (HGF),
intervenientes no processo de neo-angiogénese. O activador do receptor do ligando
factor nuclear kB (RANKL), produzido pelas células estromais assim como a proteína
inflamatória macrófagos (MIP-1α) produzida nas células plasmáticas neoplásicas,
estimulam a proliferação osteoclástica, enquanto os osteoblastos são inibidos pela
interleucina 3 (IL-3), proteína 1 relacionada com dickkopf (DKK-1) e pelo HGF (Kyle and
Rajkumar, 2004; Harousseau and Moreau, 2009) (Figura 3).
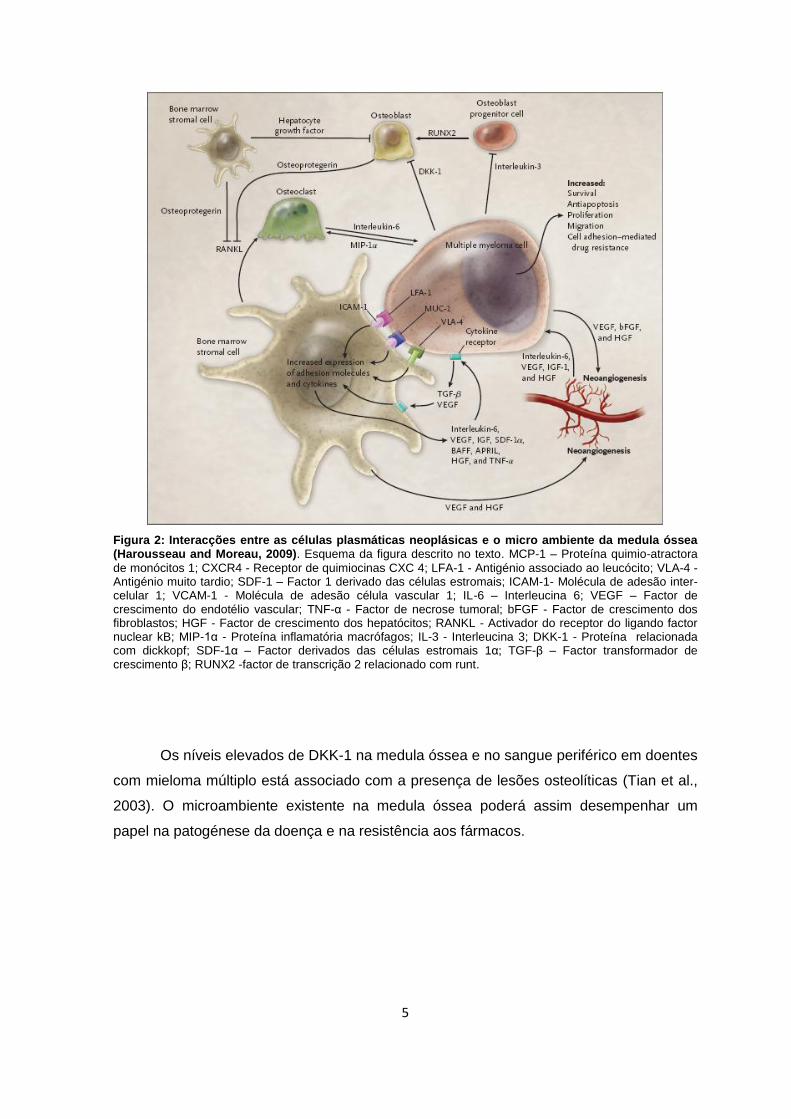
5
Figura 2: Interacções entre as células plasmáticas neoplásicas e o micro ambiente da medula óssea (Harousseau and Moreau, 2009). Esquema da figura descrito no texto. MCP-1 – Proteína quimio-atractora
de monócitos 1; CXCR4 - Receptor de quimiocinas CXC 4; LFA-1 - Antigénio associado ao leucócito; VLA-4 - Antigénio muito tardio; SDF-1 – Factor 1 derivado das células estromais; ICAM-1- Molécula de adesão inter-celular 1; VCAM-1 - Molécula de adesão célula vascular 1; IL-6 – Interleucina 6; VEGF – Factor de crescimento do endotélio vascular; TNF-α - Factor de necrose tumoral; bFGF - Factor de crescimento dos fibroblastos; HGF - Factor de crescimento dos hepatócitos; RANKL - Activador do receptor do ligando factor nuclear kB; MIP-1α - Proteína inflamatória macrófagos; IL-3 - Interleucina 3; DKK-1 - Proteína relacionada com dickkopf; SDF-1α – Factor derivados das células estromais 1α; TGF-β – Factor transformador de crescimento β; RUNX2 -factor de transcrição 2 relacionado com runt.
Os níveis elevados de DKK-1 na medula óssea e no sangue periférico em doentes
com mieloma múltiplo está associado com a presença de lesões osteolíticas (Tian et al.,
2003). O microambiente existente na medula óssea poderá assim desempenhar um
papel na patogénese da doença e na resistência aos fármacos.

6
1.3 Neoplasias de células plasmáticas: Gamapatias monoclonais
As gamapatias definem-se pelo aumento sérico de imunoglobulinas (Igs) e podem
ser de dois tipos, policlonais ou monoclonais. As gamapatias policlonais são
caracterizadas por um aumento homogéneo e difuso da fracção gama da electroforese
das proteínas séricas, representando a produção de vários tipos de imunoglobulinas
pelos vários clones de plasmócitos. Encontram-se associadas a um grupo heterogéneo
de situações não malignas, fruto de diversos processos inflamatórios ou reactivos, como
infecções virais, alergias e infecções crónica (Turgeon, 1999; Dispenzieri et al., 2001;
Kyle et al., 2002).
As gamapatias monoclonais resultam da produção excessiva de um tipo
específico de imunoglobulina ou de um fragmento por um único clone plasmocitário
potencialmente neoplásico (Attaelmannan and Levinson, 2000; Dispenzieri et al., 2001;
Silva, 2008). A imunoglobulina monoclonal, também designada de componente M,
proteína M ou paraproteína, é reconhecida na electroforese sérica ou urinária, por uma
banda de migração restrita na fracção gama (pico monoclonal) (Attaelmannan and
Levinson, 2000; Faria, 2007). A imunoglobulina é constituída por duas cadeias
polipéptidicas pesadas da mesma classe e subclasse (IgG, IgA, IgM, IgD e IgE) e por
duas cadeias polipeptidicas leves do mesmo tipo lambda (λ) ou kappa (k)(Attaelmannan
and Levinson, 2000; Faria, 2007) (figura 4). Quando a banda representada pelas
imunoglobulinas monoclonais de cadeias leves é identificada na urina, estas são
denominadas de proteínas de Bence-Jones (BJP).
Figura 3: Principais proteínas encontradas em cada banda eletroforética (Silva, 2008).

7
O mieloma múltiplo, o plasmocitoma e as gamapatias monoclonais de significado
indeterminado (MGUS) são alguns dos modelos de gamapatias monoclonais
provenientes de alterações das células plasmáticas. O seu diagnóstico requer o
estabelecimento de vários critérios bem definidos que têm por base as manifestações
clínicas e os estudos laboratoriais para verificação de possíveis alterações orgânicas.
Este inclui a detecção e quantificação do componente monoclonal, o exame da medula
óssea para verificação da infiltração plasmocitária e o estudo genético (Silva, 2008).
Durante a replicação celular e diferenciação dos linfócitos B maduros em células
plasmáticas poderão ocorrer erros genéticos, mais especificamente a nível dos
plasmablastos. Assim, a pesquisa de alterações cromossómicas é importante, pois
influenciam o prognóstico da doença. As variações mais comuns ocorrem no
cromossoma 14q32.3 originando alterações a nível do locus da cadeia pesada da
imunoglobulina (IgH). A hiperploidia encontra-se associada a melhor prognóstico,
enquanto que a deleção do cromossoma 17p13, verificado em 10% dos indivíduos, e as
translocações dos cromossomas 14q32.3 (50%), 4p16.3 (15%), o 16q23 (5%), 11q13,
6p21 e 20q11 encontram-se associadas a pior prognóstico da doença. Uma grande
variedade de outras alterações genéticas pode ocorrer em fases mais avançadas da
doença, como mutações em genes da família do oncogene RAS e do gene supressor
tumoral P53, as translocações do gene MYC e as metilações da P16 (Kyle and Rajkumar,
2004; Martinez, 2007; Harousseau and Moreau, 2009) (Figura 5).
Ocorrências oncogénicas nas células plasmáticas
Figura 4: Mecanismos oncogénicos envolvidos na diferenciação das células plasmáticas. Adaptado de:(Kyle and Rajkumar, 2004). Esquema da figura descrito no texto. IgH - cadeia pesada da imunoglobulina; MYC – factor de transcrição nuclear; RAS- proteína transdutora de sinal; NF-kB - Factor nuclear-kB; P53 – Gene supressor tumoral.
Translocação IgH 14q32 (4p26, 11q13, 16q23, 20q11, 12p13… Delecção 13 Trissomia 3, 5, 7, 9, 11, 15, 19, 21
N-Ras, K-Ras Desregulação MYC Mutação P53 Mutação da via activada do NF-kB

8
Por outro lado, as alterações verificadas no microambiente da medula óssea
também poderão exercer uma influência bem marcada neste tipo de patologias. Podemos
incluir o efeito pró-angiogénico das moléculas segregadas pelos plasmócitos neoplásicos,
a supressão imunológica e a acção da IL-6 e do VEGF.
As manifestações clínicas neste tipo de alterações das células plasmáticas podem
variar desde assintomática até estados avançados dolorosos e incapacitantes devido ao
envolvimento ósseo. Os sinais e sintomas manifestados mais frequentemente incluem a
perda de peso, cansaço, osteólise, insuficiência renal, supressão da hematopoiese, maior
risco de infecções, hipercalcémia, hiperviscosidade do sangue e amiloidose (Bataille et
al., 1984; Attaelmannan and Levinson, 2000; Kyle et al., 2002; Alexander et al., 2007;
Hungria, 2007; Grethlein, 2009; Harousseau and Moreau, 2009; Silva, 2009; Barbara J.
Bain, 2010).
O sistema de estadiamento clínico desenvolvido por Durie & Salmon para
estabelece o diagnóstico de gamapatias baseia-se na correlação de vários factores
(hemoglobina, cálcio sérico, componente monoclonal e creatinina) com a massa tumoral
(Greipp et al., 2005). No entanto, evidências recentes têm vindo a mostrar que este
sistema é incapaz de se relacionar adequadamente com a sobrevivência global e com o
tempo livre de doença (Zaidi and Vesole, 2001; Greipp et al., 2005). Desta forma, tornou-
se necessário o desenvolvimento de novos parâmetros que permitissem uma melhor
correlação clínica e uma estratificação de subgrupos com diferentes valores preditivos e
prognóstico. A combinação da β2 – microglobulina com a albumina sérica resultou num
sistema de estadiamento simples e confiável, possibilitando uma divisão em três estádios
clínicos, denominado sistema internacional de estadiamento (ISS) (quadro 1) (Zaidi and
Vesole, 2001; Greipp et al., 2005; Silva, 2009).
Quadro 1 - Sistema internacional de estadiamento (ISS). Adaptado de: (Silva, 2009).
Estádio I β2 microglobulina sérica <3,5mg/gL
Albumina sérica >3,5g/gL
Estádio II β2 microglobulina sérica <3,5mg/gL e albumina sérica <3,5g/gL
ou
β2 microglobulina sérica >3,5mg/gL e albumina sérica >3,5g/dL
Estádio III β2 microglobulina sérica >5,5mg/dL

9
1.3.1 Mieloma Múltiplo: Características gerais e critérios de diagnóstico
O mieloma múltiplo é uma neoplasia que representa cerca de 1% da totalidade
das neoplasias malignas e cerca de 10% das neoplasias hematológicas. É mais
frequente em indivíduos do sexo masculino com idades superiores a 65 anos e é mais
incidente na raça negra (Landgren et al., 2006; Alexander et al., 2007; Harousseau and
Moreau, 2009). Dados epidemiológicos referem igualmente que apenas 2% dos
indivíduos com idades inferiores a 40 anos poderão desenvolver este tipo de patologia
(Alexander et al., 2007; Lynch et al., 2008; Silva, 2009). Actualmente, tem-se vindo a
verificar um aumento na incidência de MM, reflexo de um maior conhecimento da história
natural da patologia e da sua patogénese, de um avanço dos recursos laboratoriais de
diagnóstico e do aumento da esperança média de vida (Silva, 2009). O MM possui mau
prognóstico, na medida em que somente um terço dos pacientes sobrevive 5 ou mais
anos e apenas 16% sobrevivem até aos 10 anos. Desta forma, a identificação de factores
de risco e de biomarcadores poderão ter impacto na morbilidade e mortalidade desta
patologia (Larsson and Wolk, 2007; Weiss et al., 2009). Estudos ocupacionais assinalam
um aumento do risco em indivíduos expostos a pesticidas, produtos químicos, radiações
ionizantes, asbestos e solventes específicos, como o benzeno (Lope et al., 2008). Para
além da exposição crónica a agentes poluentes, outros factores poderão estar implicados
na etiologia do MM, como a estimulação imunológica crónica, história familiar de MM e a
co-existência de doenças auto-imunes (Larsson and Wolk, 2007).
O MM é uma neoplasia progressiva e incurável caracterizada pela proliferação
desregulada e clonal na medula óssea, de células plasmáticas neoplásicas com
características morfológicas e funcionais semelhantes às células plasmáticas normais
(Turgeon, 1999; Grethlein, 2009; Barbara J. Bain, 2010). Desta forma, podem apresentar
características morfológicas tipicamente normais como o núcleo excêntrico, cromatina
grosseira e aparelho de Golgi vísivel, ou por outro lado podem apresentar modificações
moderadas ou displásicas. As alterações citológicas verificadas frequentemente incluem
a assincronia de maturação do citoplasma e do núcleo (citoplasma maduro e cromatina
nuclear difusa com nucléolos proeminentes), pleomorfismo celular e presença de corpos
de Russell (Turgeon, 1999; Beutler, 2001; Grethlein, 2009; Barbara J. Bain, 2010).
A proteína M mais frequentemente secretada é a IgG (60%), seguida da IgA (20%), IgM,
IgE ou IgD (2%), biclonal (8%) e 15 a 20% apenas produzem imunoglobulinas de cadeias

10
leves. Cerca de 7% de pacientes não produz nenhum tipo de imunoglobulinas, a nível
sérico e urinário, denominando-se a patologia por mieloma não secretor.
A imunofenotipagem completa dos plasmócitos tumorais, demonstra grande
heterogeneidade na expressão de marcadores associados à linhagem B e à
diferenciação celular. Os plasmócitos normais expressam CD19+, CD38+, CD138+,
CD45+ e são negativos para CD56-. O plasmócitos neoplásicos, na sua maioria
expressam CD56+ (62% - 75% dos casos), CD138+, CD38+, CD20+, CD28+, CD33+,
CD117+ em 28% dos casos podendo apresentar positividade fraca ou negativa para
CD45+/-. As moléculas de adesão, principalmente a CD56 e o CD86, vão permitir a sua
ligação a locais metastáticos extra-medulares quando se encontram distantes do
microambiente medular, daí que a expressão destes anticorpos se encontre relacionada
com maior agressividade e com pior prognóstico (Epstein, 1992; Omede et al., 1993;
Teoh and Anderson, 1997; Robillard et al., 2003; Lin et al., 2004; Matsui et al., 2004;
Falcão, 2007; Barbara J. Bain, 2010).
Os critérios para diagnóstico do MM envolvem a presença de mais de 10% de
células plasmáticas no aspirado de medula óssea; presença de uma proteína monoclonal
no soro e / ou urina >3g/dL, com excepção dos pacientes com MM não secretor;
evidência de alterações orgânicas atribuídas à proliferação das células plasmáticas como
a hipercalcémia (cálcio ≥11.5mg/100mL), insuficiência renal (creatinina sérica >1.75
g/dL); anemia normocrómica, normocítica com valores de hemoglobina de <10g /dL;
lesões ósseas, incluindo lesões líticas, osteoporose ou fracturas e outros como a
hiperviscosidase sanguínea e infecções bacterianas recorrentes, superiores a 2 episódios
por ano (Kyle et al., 2002; Hungria, 2007; Kyle and Rajkumar, 2009; Barbara J. Bain,
2010).
1.3.2 Plasmocitoma: Características gerais e critérios de diagnóstico
O plasmocitoma é a terminologia utilizada para a neoplasia caracterizada pela
invasão de plasmócitos neoplásicos localizados numa determinada área sem evidência

11
de disseminação proliferativa sistémica (Kumar et al., 2003). Os plasmocitomas são
responsáveis por 5 a 10% do total das neoplasias de células plasmáticas e podem ser
classificados numa lesão óssea única (plasmocitoma ósseo solitário, SBP) ou numa lesão
de tecidos moles (plasmocitoma extramedular). Os plasmocitomas não tratados poderão
evoluir para MM em cerca de 2 a 3 anos (Kumar et al., 2003).
Os critérios para diagnóstico de ambos os tipos de plasmocitoma incluem a
existência de uma área isolada de destruição óssea devido a um clone de plasmócitos
(SBP) ou a presença de células plasmáticas monoclonais no tecido de biópsia, infiltração
da medula óssea por plasmócitos inferior a 10%, ausência de anemia, hipercalcémia ou
insuficiência renal e baixa concentração de proteína M sérica (IgG <3g/dL; IgA <2g/dL)
e/ou urinária (λ ou k <1g/24H) (Dimopoulos et al., 2000; Faria, 2007; Fanning, 2008).
A indução da angiogénese é uma característica das células plasmáticas
neoplásicas, que poderá contribuir para a progressão do plasmocitoma para MM, na
medida em que permitirá a invasão da medula óssea pelos plasmócitos neoplásicos e
consequentemente, o crescimento tumoral (Kumar et al., 2003; Rajkumar, 2005).
1.3.3 MGUS: Características gerais e critérios de diagnóstico
A MGUS é a gamapatia mais frequente, presente em aproximadamente 3% da
população acima dos 50 anos, apresentando uma prevalência aumentada com a
progressão da idade (1.7% em indivíduos entre os 50 e os 59 anos e superior a 5% em
indivíduos a partir dos 70 anos) (Faria, 2007).
Os factores que contribuem para o desenvolvimento da MGUS, assim como os
que conduzem à sua evolução para MM ainda se encontram por elucidar. No entanto,
estudos referem que indivíduos com evidência de MGUS apresentam um risco mais
acentuado de progressão para MM (Cesana et al., 2002; Rajkumar, 2005; Landgren et
al., 2006; Lynch et al., 2008; Weiss et al., 2009). O risco estimado de progressão de
MGUS para MM é de aproximadamente de 19%, no intervalo de tempo de 2 a 19 anos
(Grethlein, 2009).
A MGUS é caracterizada por apresentar ausência de sintomatologia clínica,
infiltração da medula óssea por plasmócitos inferior a 10%, pela similaridade evidente nas

12
alterações cromossómicas referenciadas em doentes com MM (figura 19), pela presença
sérica e /ou urinária de IgG com concentração inferior a 3g/dL ou IgA <2g/dL e k ou λ
<1g/24h, pela inexistência de lesões ósseas, anemia ou falência renal. A proteína M mais
frequente é a IgG (55.5%), seguida da IgM (20%), IgA (10%), biclonal (8%), cadeias leves
(6%) e IgD (<0.5%) (Rajkumar, 2005; Faria, 2007; Lynch et al., 2008; Landgren et al.,
2009).
Os indivíduos com MGUS não são tratados, no entanto é necessário efectuar um
follow-up das suas condições pré-malignas, devido à possível evolução para MM. O risco
de progressão é afectado pela concentração sérica da imunoglobulina monoclonal, pelo
aumento de plasmócitos neoplásicos na medula óssea, pelo aumento da angiogénese e
pelo comprometimento orgânico que caracteriza o MM (figura 6). No entanto, a razão
pela qual o MGUS evolui para MM é desconhecida, continuando esta a ser uma
importante questão ainda por resolver (Rajkumar, 2005; Landgren et al., 2009; Weiss et
al., 2009).
Figura 5: Patogénese e progressão de gamapatia monoclonal de significado indeterminado para mieloma múltiplo. Adaptado de:(Rajkumar, 2005). Esquema da figura descrito no texto. RANKL - Activador
do receptor do ligando factor nuclear kB; MIP-1α - Proteína inflamatória macrófagos; IL-6 - Interleucina 6; VEGF – Factor de crescimento do endotélio vascular;N-Ras - Proteína transdutora de sinal; K-Ras - Proteína transdutora de sinal; MGUS – Gamapatia monoclonal de significado indeterminado.

13
Vários autores têm referenciado a obesidade como factor de risco ao
desenvolvimento de vários tumores sólidos, incluindo o do cólon, mama, endométrio, rim
e esófago (Barb et al., 2007). O excesso de peso e a obesidade também parecem estar
associados com o desenvolvimento de MM, no entanto as evidências epidemiológicas
são ainda reduzidas devido ao escasso número de estudos (Lauta, 2003; Birmann et al.,
2007; Larsson and Wolk, 2007). Este poderá ser um dado para a patogénese do MM,
com consequências no desenvolvimento de estratégias preventivas da doença e
abordagem terapêutica.
1.4 Tecido adiposo: Conceito e fisiopatologia
Actualmente, os avanços nas ciências biomédicas têm vindo a demonstrar uma
mudança de conceito relativamente às funções dos tecidos e órgãos do organismo
humano. O tecido adiposo representa um típico exemplo desta mudança. Para além da
sua clássica função de armazenamento de energia, representa o maior órgão endócrino
do organismo, produtor de hormonas e outras moléculas com impacto na regulação do
metabolismo energético, sensibilidade à insulina, imunidade, inflamação e metabolismo
ósseo (Haluzik et al., 2004; Dietze-Schroeder et al., 2005). O tecido adiposo é constituído
por adipócitos e por uma fracção vascular estromal, na qual estão incluídos os pré-
adipócitos, macrófagos, as células mesenquimais, endoteliais e fibroblastos. No tecido
adiposo são produzidas várias citocinas, (TNF-α, IL-6 e IL-8), hormonas (adiponectina,
leptina e resistina), factores de crescimento (VEGF, bFGF) e mediadores do processo de
coagulação (inibidor do activador do plasminogénio 1 - PAI-1), que representam um papel
relevante no desenvolvimento da obesidade, na proliferação celular e no interface entre o
sistema imunológico e metabólico (Fruhbeck et al., 2001; Onuma et al., 2003; Tilg and
Moschen, 2006).
A obesidade é definida pela alteração nutricional mais frequente nos países
desenvolvidos e a sua prevalência tem vindo a aumentar significativamente (OMS, 2006).
14
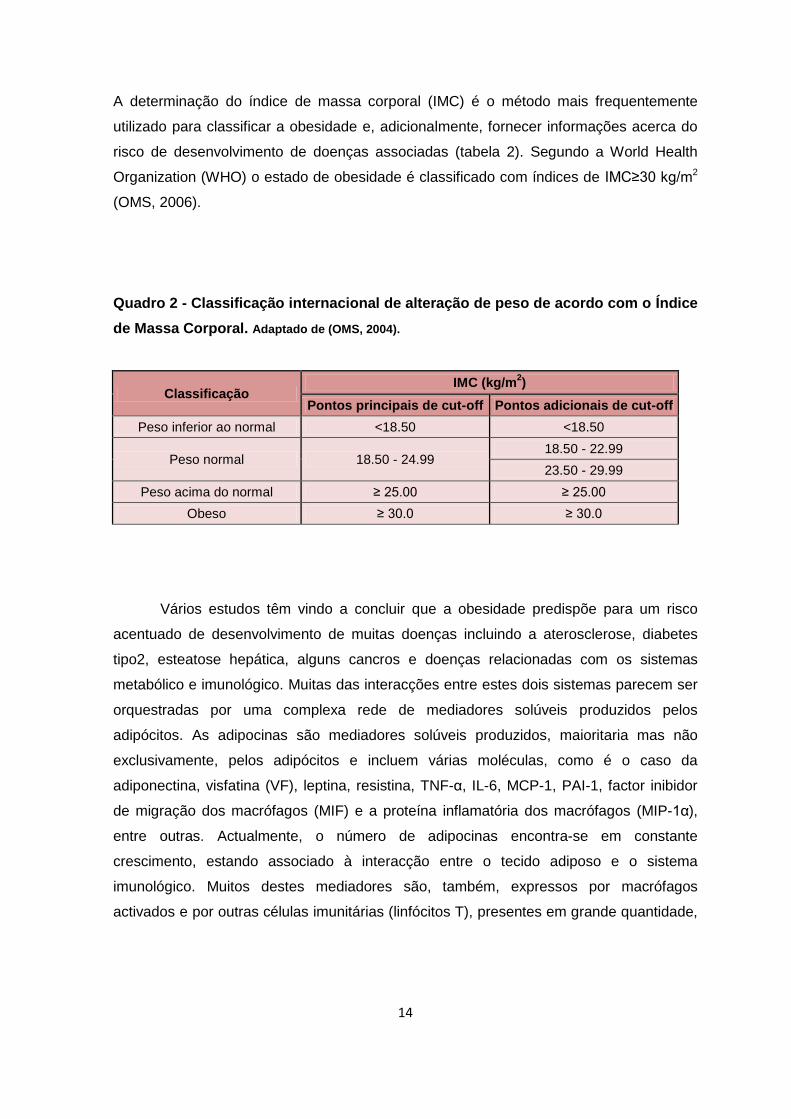
A determinação do índice de massa corporal (IMC) é o método mais frequentemente
utilizado para classificar a obesidade e, adicionalmente, fornecer informações acerca do
risco de desenvolvimento de doenças associadas (tabela 2). Segundo a World Health
Organization (WHO) o estado de obesidade é classificado com índices de IMC≥30 kg/m2
(OMS, 2006).
Quadro 2 - Classificação internacional de alteração de peso de acordo com o Índice
de Massa Corporal. Adaptado de (OMS, 2004).
Classificação IMC (kg/m
2)
Pontos principais de cut-off Pontos adicionais de cut-off
Peso inferior ao normal <18.50 <18.50
Peso normal 18.50 - 24.99 18.50 - 22.99
23.50 - 29.99
Peso acima do normal ≥ 25.00 ≥ 25.00
Obeso ≥ 30.0 ≥ 30.0
Vários estudos têm vindo a concluir que a obesidade predispõe para um risco
acentuado de desenvolvimento de muitas doenças incluindo a aterosclerose, diabetes
tipo2, esteatose hepática, alguns cancros e doenças relacionadas com os sistemas
metabólico e imunológico. Muitas das interacções entre estes dois sistemas parecem ser
orquestradas por uma complexa rede de mediadores solúveis produzidos pelos
adipócitos. As adipocinas são mediadores solúveis produzidos, maioritaria mas não
exclusivamente, pelos adipócitos e incluem várias moléculas, como é o caso da
adiponectina, visfatina (VF), leptina, resistina, TNF-α, IL-6, MCP-1, PAI-1, factor inibidor
de migração dos macrófagos (MIF) e a proteína inflamatória dos macrófagos (MIP-1α),
entre outras. Actualmente, o número de adipocinas encontra-se em constante
crescimento, estando associado à interacção entre o tecido adiposo e o sistema
imunológico. Muitos destes mediadores são, também, expressos por macrófagos
activados e por outras células imunitárias (linfócitos T), presentes em grande quantidade,

15
no tecido adiposo de indivíduos obesos, representando, desta forma uma fonte adicional
e desequilibrada da sua produção.
Assim, em situações de obesidade verifica-se uma indução da produção do TNF-
α, IL-6, PBEF/vistafina, leptina e IL-1β, infiltração de macrófagos e linfócitos T no tecido
adiposo, supressão da expressão da adiponectina, aumento sérico de produtos de fase
aguda (proteína C reactiva - PCR), que no seu conjunto vão contribuir para o
desenvolvimento de inflamação local e sistémica (Figura 7).
Figura 6: Tecido adiposo: componentes celulares, moléculas sintetizadas e seu envolvimento com a
inflamação (Tilg and Moschen, 2008). Esquema da figura descrito no texto. MIP-1α - Proteína inflamatória
macrófagos; IL-6 - Interleucina 6; TNF-α - Factor de necrose tumoral; MCP-1 – Proteína quimio-atractora de
monócitos 1; MIP-1α - Proteína inflamatória macrófagos; IL-1β - Interleucina 1β; MIF - factor inibidor de
migração dos macrófagos.
O MCP-1, o MIF e o MIP-1α foram recentemente identificados como potenciais
factores de infiltração dos macrófagos no tecido adiposo. Uma vez presentes e activos no
tecido adiposo, os macrófagos em conjunto com os adipócitos, podem perpetuar um ciclo
com feed-back positivo no recrutamento de macrófagos e produção de citoquinas pró-
inflamatórias (Okamoto et al., 2006; Tilg and Moschen, 2006, 2008).

16
Os macrófagos representam a principal fonte de produção de TNF-α no tecido
adiposo. O TNF-α tem um efeito supressor da produção de adiponectina nos adipócitos
apresentando-se como um dos mecanismos para níveis mais baixos deste mediador anti-
inflamatório nos indivíduos obesos. A diminuição da produção de adiponectina pode
contribuir para o desenvolvimento de tumores, devido à interacção com as vias da
insulina, com a inflamação e com a angiogénese (Barb et al., 2007). (Figura 8)
Figura 7: Factores contribuintes para o aumento do risco da obesidade associada a neoplasias. Adaptado de: (Barb et al., 2007). Esquema da figura descrito no texto. FFA –Ácidos gordos livres; IGFBPs –
Proteínas de ligação ao factor de crescimento insulínico; IGF – Factor crescimento insulínico 1; E2 – Estradiol; T – Testosterona; 17βHSD – desidrogenase hidroxiesteróide 17
Em situações patológicas de obesidade, o desequilíbrio na concentração destas
adipocinas, poder-se-á reflectir no desenvolvimento de patologias malignas, como o
cancro do endométrio, mama, próstata e aparelho digestivo (Kelesidis et al., 2006; Barb
et al., 2007). Mais recentemente, estudos demonstraram o relacionamento da obesidade
com patologias hematológicas, como a leucemia mieloblástica aguda e o mieloma
múltiplo (Petridou et al., 2006; Dalamaga et al., 2009).
O tecido adiposo é um órgão endócrino complexo, que se encontra envolvido, em
importantes processos biológicos do organismo humano, nomeadamente através de
adipocinas produzidaspelo mesmo e segregadas para o sangue e interstícios. Este
estudo incidirá sobre, uma das principais adipocinas anti-inflamatórias produzida pelos
adipócitos, a adiponectina.

17
1.5 Adiponectina
1.5.1 Caracterização molecular e genética
A adiponectina, também conhecida por GBP-28 (Nakano et al., 1996) apM1
(Maeda et al., 1996) adipoQ (Hu et al., 1996) ou Acrp30 (Scherer et al., 1995), é uma
proteína produzida nos adipócitos. É codificada pelo gene apM1, localizado no
cromossoma 3q27, (Avcu et al., 2006; Kelesidis et al., 2006; Barb et al., 2007; Waki and
Tontonoz, 2007) (figura 9).
Actualmente, são conhecidas várias formas de adiponectina circulantes no
plasma, a total (fAd) e a globular (gAd), sendo a fAd a mais predominante em circulação,
com concentrações plasmáticas de aproximadamente 10µg/mL.
A fAd constituída por 244 aminoácidos, apresentando uma região N-terminal com
uma grande afinidade estrutural para o colagénio VIII, X e complemento C1q e uma
região C-terminal globular que origina diferentes formas multiméricas (Barb et al., 2007;
Guerre-Millo, 2008). Pode apresentar-se sob três isoformas estáveis e distintas, como
trímero (adiponectina de baixo peso molecular - LMW), hexâmero (médio peso molecular
- MMW) ou como adiponectina de alto peso molecular (HMW), encontrando-se esta
última, presente em maior concentração em circulação (Kadowaki and Yamauchi, 2005)
(Figura 9).
A gAd resulta da clivagem proteolítica da adiponectina HMW, consistindo numa
molécula constituída por 110 aminoácidos (Kadowaki and Yamauchi, 2005; Barb et al.,
2007).
Figura 8: Estrutura e formas oligoméricas da adiponectina total (Kadowaki and Yamauchi, 2005).

18
1.5.2 Receptores ADIPOR1 e ADIPOR2
A adiponectina é produzida principalmente nos adipócitos. Vários autores
verificaram a expressão dos seus receptores em outras células e tecidos, nomeadamente
nos osteoblastos (Berner et al., 2004) e no tecido músculo-esquelético (Delaigle et al.,
2004), hepatócitos (Berg et al., 2001), medula óssea (Yokota et al., 2002; Luo et al.,
2006), em miócitos cardíacos (Ding et al., 2007) e em células do sangue periférico,
nomeadamente nos neutrófilos (Crawford et al., 2010). Encontram-se descritos dois tipos
de receptores de adiponectina, o ADIPOR1 e o ADIPOR2, com distribuições e afinidades
específicas para as diferentes formas de adiponectina circulante. Ambos são proteínas
integrais de membrana constituídas por sete domínios transmembranares, uma
terminação interna N-terminal e uma externa C-terminal (Figura 10).
Figura 9: Estrutura dos receptores de adiponectina (Kadowaki and Yamauchi, 2005). Esquema da figura
descrito no texto.
O ADIPOR1 apresenta uma maior afinidade para a gAd comparativamente com as
várias isoformas de fAd. Está abundantemente expresso nos tecidos do músculo-
esquelético e em células endoteliais. O ADIPOR2 apresenta similar afinidade para todas
as formas de adiponectina e encontra-se, predominantemente, nos hepatócitos
(Yamauchi et al., 2003; Fang and Sweeney, 2006; Okamoto et al., 2006; Tilg and
Moschen, 2006; Guerre-Millo, 2008).
Vários estudos reportaram que a caderina T, um receptor celular de superfície
envolvido na interacção célula-célula mediada por cálcio e na sinalização de células
endoteliais e do músculo liso, parece funcionar como receptor para as isoformas MMW e
HMW de adiponectina, existindo compatibilidade para a gAd e LMW. Vários autores

19
sugeriram que a caderina-T, devido à ausência do domínio intracelular, actua como co-
receptor de um receptor de sinalização não identificado. (Tilg and Moschen, 2006; Barb et
al., 2007; Waki and Tontonoz, 2007).
Também em linhas de células tumorais e tumores foi demonstrada a presença de
ADIPORs na próstata, mama, endométrio, fígado, cólon e em neuroblastoma (Wang et
al., 2005b; Bub et al., 2006; Dieudonne et al., 2006; Barb et al., 2007), sugerindo efeitos
directos da adiponectina mediado pelos seus receptores neste tipo de células.
1.5.3 Caracterização fisiológica
A adiponectina possui um papel importante nos metabolismos lipídico e da glicose
e no desenvolvimento e progressão de patologias relacionadas com a obesidade, embora
o seu processo de envolvimento não esteja ainda totalmente esclarecido, sabendo-se
que a eficácia biológica da adiponectina depende da disponibilidade sérica das suas
isoformas e da capacidade funcional dos seus receptores.
Estudos prévios revelaram a intervenção da adiponectina nos mecanismos
reguladores de ácidos gordos livres (oxidação de ácidos gordos) e na glicogenólise.
(Guerre-Millo, 2008).
A adiponectina apresenta alguns efeitos biológicos nas células do sistema
imunológico, na medida em que está implicada na indução da apoptose dos monócitos,
na estimulação da via da proteína quinase activada por 5´-AMP (AMPK), na inibição do
crescimento dos precursores dos macrófagos e na supressão da actividade fagocítica
dos macrófagos. Também é responsável pela supressão da produção de IL-6, IL-8 e
induz a síntese de IL-10 e do receptor antagonista da IL-1 (IL-1RA).
A adiponectina globular, através da ligação ao receptor ADIPOR1, suprime a
activação do factor nuclear-kB (NF-kB) (Arkan et al., 2005; Tilg and Moschen, 2006),
implicado na produção de TNF-α e IFN-γ (Tilg and Moschen, 2006; Barb et al., 2007;
Waki and Tontonoz, 2007).
A adiponectina é um importante mediador na regulação da sensibilidade à insulina
e na supressão da inflamação. Vários estudos indicam que a estimulação dos receptores
ADIPOR1 e ADIPOR2 desencadeiam, nos tecidos do músculo-esquelético e no fígado, a
activação da AMPK e do receptor do activador da proliferação de peroxissomas α e γ
(PPAR α e γ), responsáveis pela oxidação de ácidos gordos e metabolismo da glicose
(Kern et al., 2003; Okamoto et al., 2006; Kanazawa et al., 2007; Guerre-Millo, 2008).

20
A AMPK actua como um sensor na conservação energética da célula e apresenta
um papel fundamental no controlo do balanço energético sistémico através regulação da
ingestão alimentar, peso corporal e homeostase da glicose e dos lípidos. Para além da
adiponectina, pode ser activada por outras moléculas a leptina, IL-6 ou catecolaminas.
Uma vez activada, exerce efeitos directos em enzimas específicas que são a chave
reguladora das proteínas e ácidos gordos. (Tilg and Moschen, 2006)
Nos tecidos do músculo-esquelético a adiponectina globular e HMW activam a
AMPK conduzindo à estimulação da fosforilação da carboxilase acetil-CoA (ACC),
oxidação dos ácidos gordos e consumo de glicose. No fígado, apenas a fAd activa a
AMPK, reduzindo moléculas envolvidas na gluconeogénese e promovendo a fosforilação
do ACC e a oxidação dos ácidos gordos.
O PPAR α e γ são membros dos receptores hormonais nucleares da super-família
de ligandos dependentes de factores de transcrição, responsáveis pela regulação do
metabolismo energético e pela homeostase (Ding et al., 2007).
Vários autores referem que a adiponectina é responsável, via activação de PPAR-
α, pela combustão dos ácidos gordos e consumo de energia que predispõe para o
decréscimo do conteúdo de triglicerídeos no fígado e nos tecidos do músculo-esquelético
e, consequentemente, para a indução da sensibilidade à insulina. Os níveis de
adiponectina circulante são regulados positivamente pela actividade dos agonistas do
PPAR-γ, como é o caso das tiazidas (TZDs) (figura 11) (Kern et al., 2003; Rajala and
Scherer, 2003; Kadowaki and Yamauchi, 2005; Tilg and Moschen, 2008).
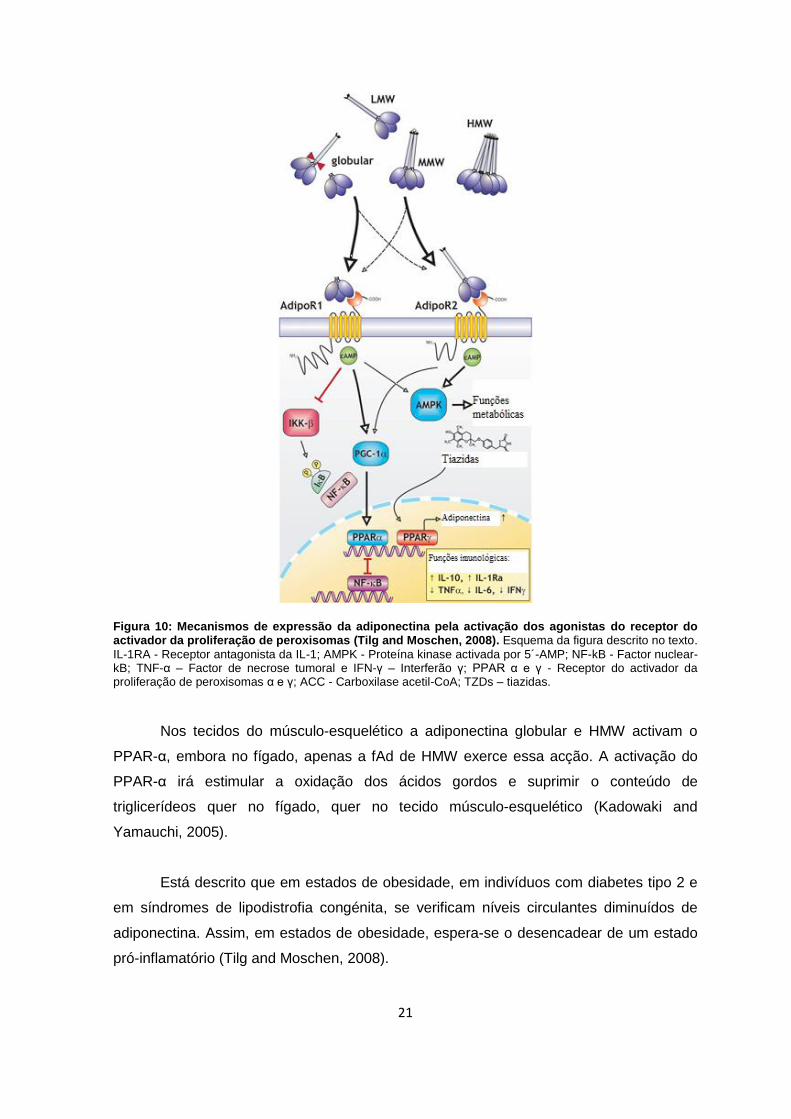
21
Figura 10: Mecanismos de expressão da adiponectina pela activação dos agonistas do receptor do activador da proliferação de peroxisomas (Tilg and Moschen, 2008). Esquema da figura descrito no texto.
IL-1RA - Receptor antagonista da IL-1; AMPK - Proteína kinase activada por 5´-AMP; NF-kB - Factor nuclear-kB; TNF-α – Factor de necrose tumoral e IFN-γ – Interferão γ; PPAR α e γ - Receptor do activador da proliferação de peroxisomas α e γ; ACC - Carboxilase acetil-CoA; TZDs – tiazidas.
Nos tecidos do músculo-esquelético a adiponectina globular e HMW activam o
PPAR-α, embora no fígado, apenas a fAd de HMW exerce essa acção. A activação do
PPAR-α irá estimular a oxidação dos ácidos gordos e suprimir o conteúdo de
triglicerídeos quer no fígado, quer no tecido músculo-esquelético (Kadowaki and
Yamauchi, 2005).
Está descrito que em estados de obesidade, em indivíduos com diabetes tipo 2 e
em síndromes de lipodistrofia congénita, se verificam níveis circulantes diminuídos de
adiponectina. Assim, em estados de obesidade, espera-se o desencadear de um estado
pró-inflamatório (Tilg and Moschen, 2008).

22
Nos últimos anos verifica-se uma evidência crescente de associação entre a
resistência à insulina e a inflamação, possivelmente mediada pelo TNF-α. Com o
aumento do tecido adiposo várias adipocinas induzem a activação e infiltração de
macrófagos. Estes, quando activados, segregam citoquinas pró-inflamatórias que podem
alterar a sensibilidade à insulina e estimular a infiltração dos monócitos e macrófagos no
tecido adiposo. Os pré-adipócitos também podem segregar quimiocinas, mediante
estimulação pelo TNF-α, podendo contribuir para a infiltração de macrófagos. Estes sinais
de amplificação podem comprometer a sinalização da insulina e, eventualmente, causar
uma resistência sistémica à insulina (Xu et al., 2003).
Adicionalmente, também foi descrito o relacionamento da adiponectina com o
processo de angiogénese, na qual esta adipocitoquina actuará como um inibidor directo,
actuando em factores de crescimento que regulam a proliferação celular da parede
vascular das células. Por outro lado, a adiponectina tem a capacidade de activar
directamente um grupo de caspases (3, 8 e 9) que se encontram envolvidas na apoptose
celular das células endoteliais, implicando de uma forma indirecta, a inibição celular do
crescimento tumoral pelo decréscimo da neovascularização (Brakenhielm et al., 2004;
Wang et al., 2005b; Kelesidis et al., 2006; Molica et al., 2009).
1.6 Adiponectina e as neoplasias
Vários estudos epidemiológicos revelaram que a obesidade representa um factor
de risco, no desenvolvimento de neoplasias. Um estudo prospectivo com
aproximadamente 1 milhão de indivíduos adultos nos EUA durante um período de 16
anos verificou associação entre o IMC e o risco de morte relativo a várias neoplasias
(Calle et al., 2003).
Os órgãos ricos em tecido adiposo, como a mama e a medula óssea, poderão ser
locais predisponentes ao desenvolvimento de neoplasias. Contudo, a oncogénese poderá
não estar forçosamente associada à massa gorda corporal, mas sim influenciada

23
maioritariamente, pelos perfis de secreção fisiopatológicos induzidos pelos adipócitos nos
microambientes oncogénicos (Rajala and Scherer, 2003).
Desta forma, a adiponectina possui um papel de protecção relativamente aos
mecanismos de iniciação e desenvolvimento tumoral celular. Assim, relativamente à sua
acção directa nas células, um estudo demonstrou, in vitro, que a adiponectina suprime o
crescimento das linhas celulares mielomonocíticas, induzindo a apoptose das células
progenitoras mielomonocíticas, modula a expressão dos genes relacionados com a
apoptose nas células LMA-M1 e regula negativamente a expressão do gene BCL-2
(Yokota et al., 2000). Apresenta ainda uma relação indirecta com o desenvolvimento de
cancro através da interacção com vários factores de crescimento, incluindo o factor de
crescimento derivado de plaquetas BB (PDGF-BB), o factor de crescimento epidérmico
de ligação à heparina (HB EGF), o VEGF e o FGF, responsáveis pelo seu envolvimento
na regulação da angiogénese, aterogénese, carcinogénese e inflamação. A adiponectina
influencia a biodisponibilidade destes factores de crescimento, regulando assim, a
fisiopatologia desses processos com efeitos anti-ateroscleróticos, anti-angiogénicos e
anti-proliferativos (Wang et al., 2005b; Kelesidis et al., 2006; Molica et al., 2009).
Uma das principais moléculas intervenientes na sinalização celular da
adiponectina é a AMPK que apresenta um papel particularmente importante no
mecanismo de regulação do crescimento das células tumorais. Assim, a AMPK activada
pela adiponectina poderá exercer efeitos directos, incluindo a inibição da sintase de
ácidos gordos (FAS) e a activação do TSC2, um supressor tumoral, que regula
negativamente a síntese de proteínas pela inibição da proteína mTOR. Estas vias
moleculares representam alguns dos mecanismos pelos quais a adiponectina ao
estimular a AMPK, inibe o crescimento e/ou a sobrevivência das células tumorais
(Kelesidis et al., 2006) (Figura 12).
A correlação negativa entre os níveis circulantes de adiponectina, a obesidade e o
conhecimento das suas funções de inibição da proliferação celular e da indução da
apoptose celular, sugerem plausibilidade biológica para um papel inibidor do
desenvolvimento tumoral e confirmam as evidências previamente estabelecidas relativas
à associação entre obesidade e as neoplasias.

24
Figura 11: Possíveis mecanismos moleculares implicados na regulação do crescimento das células tumorais pela proteína kinase activada por 5´-AMP. Adaptado de (Kelesidis et al., 2006). Esquema da
figura descrito no texto. AMPK - Proteína kinase activada por 5´-AMP; PI-3K – fosfatidilinositol-3-kinase; AKT – Proteína B kinase; FAS - sintase de ácidos gordos; mTOR – proteína de alvo da rapamicina
Vários estudos de caso-controlo mostraram que os níveis circulantes de
adiponectina se encontram inversamente correlacionados com o cancro na mama
(Korner et al., 2007; Tworoger et al., 2007), endométrio (Soliman, 2005; Cust et al., 2007),
próstata (Michalakis et al., 2007), colo-rectal (Wei et al., 2005), gástrico (Ishikawa et al.,
2005), renal (Spyridopoulos et al., 2007), leucemia mieloblástica aguda (Petridou et al.,
2006), leucemia linfoblástica crónica / síndrome mielodisplásico (Avcu et al., 2006) e
mieloma múltiplo (Dalamaga et al., 2009; Reseland et al., 2009).
Outros estudos mais recentes investigaram a possível acção da adiponectina no
processo de hematopoiese na medula óssea. O microambiente da medula óssea é
constituído por vários elementos, dentre os quais se destacam citoquinas, componentes
da matriz extracelular, adipócitos e vários tipos de células do estroma, como é o caso dos
fibroblastos, das células endoteliais e das células fagocíticas mononucleares. Desta
forma, como a formação celular hematopoiética se encontra dependente do equilíbrio
existente entre estes componentes e tendo em conta as acções que a adiponectina
exerce em alguns destes elementos, esta poderá ter um papel na regulação da
hematopoiese (Yokota et al., 2002; Iversen and Wiig, 2005; Petridou et al., 2006;
Crawford et al., 2010).

25
1.7 Obesidade, adiponectina - mieloma múltiplo / MGUS/plasmocitoma
Estudos anteriores investigaram a provável implicação da obesidade no risco de
desenvolvimento de MM, embora os potenciais mecanismos pelos quais esta relação é
estabelecida ainda permaneçam por elucidar (Birmann et al., 2007; Larsson and Wolk,
2007). Foi sugerido que o aumento do número total de plasmócitos em risco de sofrer
transformação maligna e a contribuição de citoquinas envolvidas na proliferação e
sobrevivência dos plasmócitos podem explicar o risco de MM em obesos (Lauta, 2003). A
IL-6 tem sido descrita como a citoquina que parece ser também influente na patogénese
do MM e na proliferação anormal dos plasmócitos (Lauta, 2003). A produção e a
actividade de IL-6 são reguladas por vários mecanismos e pode estar relacionada com
níveis de adiponectina observados em doentes com MM, possivelmente decorrente da
sobreprodução de IL-6 e TNF-α na medula óssea.
Alguns autores sugeriram uma associação entre a obesidade e o mieloma
múltiplo(Dalamaga et al., 2009; Reseland et al., 2009). Estes estudaram os níveis de
adiponectina, leptina e resistina em doentes com MM. Em ambos os estudos foi
observada associação inversa entre os níveis séricos de adiponectina e o risco para
desenvolvimento de MM: no trabalho de Reseland et al. (2009) foi avaliada ainda a
concentração de adiponectina no momento da recidiva de MM, não se tendo verificado
qualquer relação com o prognóstico.
Permanece por estabelecer e confirmar a associação da adiponectina com o MM,
nomeadamente qual a isoforma envolvida e o seu envolvimento como marcador de
prognóstico.

2. Objectivos

27
2. Objectivos
Objectivos gerais
Estudar a influência da adiponectina e do TNF-α no desenvolvimento de mieloma
múltiplo.
Avaliar a influência do tratamento na concentração de adiponectina, em doentes com
mieloma múltiplo.
Objectivos específicos
Dosear a adiponectina total e globular no sangue periférico e no líquido intersticial
medular em doentes com mieloma múltiplo e em controlos.
Avaliar os níveis de adiponectina antes e após tratamento.
Dosear os níveis de TNF-α no sangue periférico e líquido intersticial medular.
Avaliar a expressão dos receptores da adiponectina nos plasmócitos de doentes com MM
no momento do diagnóstico.

3. Material e Métodos

29
3. Material e métodos
3.1 População
Este estudo do tipo caso-controlo incluiu amostras de sangue periférico e medula
óssea de 24 indivíduos, 14 do género feminino e 10 do género masculino. Participaram
neste estudo doentes com diagnóstico de MM e plasmocitoma (n = 13; idade = 64,3 ±
10,7), os quais foram englobados no mesmo grupo, devido às características de
malignidade e tratamento; e MGUS (n = 8; idade = 62,9 ± 13,6), incidentes no Serviço de
Onco-Hematologia do Instituto Português de Oncologia de Francisco Gentil, EPE – Porto,
entre Janeiro e Junho de 2010.
O grupo controlo inclui 12 indivíduos, 7 homens e 5 mulheres (idade = 62,2 ± 8,0)
sem qualquer tipo de patologia neoplásica, recrutados a partir do Banco de Dadores de
Sangue do IPO Porto e emparelhados com os doentes de acordo com o grupo etário.
Nenhum dos controlos desenvolveu MM, MGUS ou plasmocitoma durante o estudo.
O diagnóstico foi obtido de acordo com os critérios clássicos morfológicos, clínicos e
laboratoriais em todos os estádios da doença (I, II e III), segundo o sistema de
internacional de estadiamento (ISS).
Os doentes com MM foram divididos à data do diagnóstico em dois grupos de
tratamento. Um grupo com os doentes elegíveis para auto-transplante de medula óssea
(idade inferior ou igual a 65 anos) tratados com talidomida e dexametasona (tratamento
A). No outro grupo encontram-se os doentes não elegíveis para transplante (idade
superior a 65 anos), tratados com melfalam, talidomida e prednisolona (tratamento B). O
tratamento A consiste em ciclos de talidomida e dexametasona até à data do transplante.
A talidomida é administrada contínuamente (200mg/dia) enquanto que a dexametasona é
administrada durante 7 dias, a cada 28 dias. O tratamento B consiste me ciclos de 28
dias, até ao máximo de 12 ciclos (aproximadamente 1 ano). São administrados
100mg/dia de talidomida, enquanto o melfalam e a prednisolona são administrados
durante 7 dias, a cada 28 dias.
O projecto de investigação, assim como o acesso aos processos clínicos foi
avaliado e aprovado pela Comissão de Ética do Instituto. Apenas foram incluídos no
estudo os doentes que aceitaram participar e assinaram o consentimento informado.

30
3.2 Processamento de amostras
As amostras de medula óssea e sangue periférico foram obtidas,
respectivamente, por punção medular da crista ilíaca e venopunção do antebraço, na
fase de diagnóstico da patologia. Após 1 mês de tratamento, foram novamente colhidas
amostras de sangue periférico. Em todos os casos apenas foram utilizadas amostras
biológicas remanescentes dos procedimentos de diagnóstico, nunca o colocando em
causa. Todas as amostras foram centrifugadas a 2000 rpm durante 10 minutos à
temperatura ambiente. O plasma e o líquido intersticial medular recolhido foi armazenado
a -20ºC.
As lâminas de aspirado de medula óssea, obtidas através punção medular, foram
fixadas em acetona refrigerada durante 5 minutos e após secagem foram armazenadas a
-20ºC.
O processo de colheita e armazenamento das amostras e em todos os indivíduos
foi realizado obedecendo criteriosamente ao mesmo protocolo estabelecido.
Quantificação de adiponectina, TNF-α
As concentrações de adiponectina total e globular no plasma e no líquido
intersticial foram determinadas sempre em duplicado através de ELISA. Para detecção da
gAd utilizou-se um kit específico: Human Globular Adiponectin gAcrp30 (Alpha Diagnostic
International, EUA). As amostras foram diluídas numa concentração 1:10.
A detecção de tAd foi realizada através do kit AviBion Human Adiponectin Acrp30
(Orgenium Laboratories, Finlândia) no analisador Dynex DS2 system. As amostras foram
diluídas 1:1000.
A concentração de TNF-α foi determinada no plasma e no líquido intersticial
medular em duplicado para cada amostra através de radioimunoensaio (RIA). Foi usado
o kit TNF-α-IRMA (Diasource ImmunoAssays S.A.) no analisador LKB 1271 RIAGAMA.

31
Imunocitoquímica dos receptores de adiponectina
A pesquisa de receptores de adiponectina, ADIPOR1 e ADIPOR2 foi efectuada
nas lâminas de medula óssea preparadas aquando da punção medular de diagnóstico.
Após descongelação das lâminas em acetona (-20 C) e lavagem em tris buffered saline
(TBS) foi efectuado bloqueio da Peroxidase endógena. Os anticorpos primários ADIPOR1
(cat.nº H-001-23) e ADIPOR2 (cat.nº H-001-44) (Phoenix Pharmaceuticals, Inc.) foram
incubados durante 1h à temperatura ambiente (diluição 1:750 e 1:220, respectivamente),
foi utilizado para detecção da reacção o sistema Power Vision + Poly – anti Ms/Rb/Rt IgG
biotin-free (Immunologic BV). Após incubação com o HRP durante 30 minutos à
temperatura ambiente, foi realizada revelação com DAB e coloração com hematoxilina.
O critério de positividade foi reconhecido pela coloração castanha na membrana
citoplasmática.
3.3 Análise estatística
Os resultados relativos às variáveis categóricas foram realizados através da
frequência (percentagem), tendo sido utilizado o teste de qui-quadrado de Pearson para
avaliar diferenças entre proporções, ou quando adequado o teste exacto de Fisher.
No que respeita ao estudo das variáveis contínuas, foram efectuados os testes de
normalidade Kolmogorov-Smirnov e Shapiro-Wilk, verificando-se a existência de variáveis
não-paraméticas (IMC, TNF-α no momento do diagnóstico, LDH, pico monoclonal e o
número de plasmócitos). Foram usadas como medidas de tendência central a média ±
erro padrão da média e a avaliação de diferenças entre grupos, no momento do
diagnóstico através t-Student medidas independentes e Mann Whitney U, de acordo com
o tipo de variável paramétrica e e não-paramétrica, respectivamente. A comparação da
idade e IMC entre os grupos MM, MGUS e controlos foi efectuada através da ANOVA
factorial de medidas independentes. Para comparação das médias dos mesmos grupos
ao longo do tempo foi usado o teste de t-Student medidas emparelhadas. O grau de
associação entre as variáveis contínuas foi testado através do teste de correlação de
Pearson (r). Foi considerado com significado estatístico o valor de p<0,05. A análise
estatística foi efectuada através do software SPSS 16.0.

4. Resultados

33
4. Resultados
O quadro 3 evidencia as características demográficas e clínicas dos indivíduos
participantes neste estudo. Não se verificaram diferenças significativas a nível de idade
(p=0,879) e de IMC (p=0,973), entre os grupos MM, MGUS e controlos. Observou-se que,
segundo a classificação clínica de obesidade, os valores médios de IMC em cada grupo,
MM, MGUS e controlos se classificam como excesso de peso (28,0 ± 4,8; 28,2 ± 4,8 e
27,8 ± 3,2, respectivamente).
Relativamente às características clínicas dos MM verificou-se que 53,9% se
encontravam no estádio III correspondente ao estadiamento ISS (Quadro 3). A
imunoglobulina mais frequente é a IgG, representada em 69,3% e 75% dos casos de MM
e MGUS, respectivamente (Quadro 3).
As sintomatologias mais relevantes apresentadas pelos doentes no momento do
diagnóstico foram: 57,1% com sintomatologia álgica óssea (MM, 69,2%; MGUS, 37,5%),
23,8% com caquexia/anorexia (MM, 30,8%; MGUS, 12,8%) e 23,8% não apresentavam
sintomatologia no momento de diagnóstico.
Quadro 3 - Características clínicas e analíticas dos doentes com mieloma múltiplo e
gamapatia monoclonal de significado indeterminado
Casos
MM (n=13) MGUS (n=8) p*
Estadiamento (ISS)a
I 6 (46,2 %) 8 (100%)
II 3 (23,1 %) 0 (0%)
III 4 (30,8 %) 0 (0%)
Proteína monoclonala
IgG 9 (69,3%) 6 (75%)
IgA 1 (7,6%) 2 (25%)
Cadeias leves 3 (23,1%) 0 (0%)
Número de plasmócitos na
medula óssea por
contagem morfológicab
38,1 ± 7,7 1,6 ± 0,6 0,001
Pico monoclonalb 24,0 ± 6,7 10,9 ± 1,5 0,310
MM - Mieloma múltiplo. MGUS - Gamapatia monoclonal de significado indeterminado. a n (%);
b média ± erro
padrão. * - Teste Mann Whitney U.
34
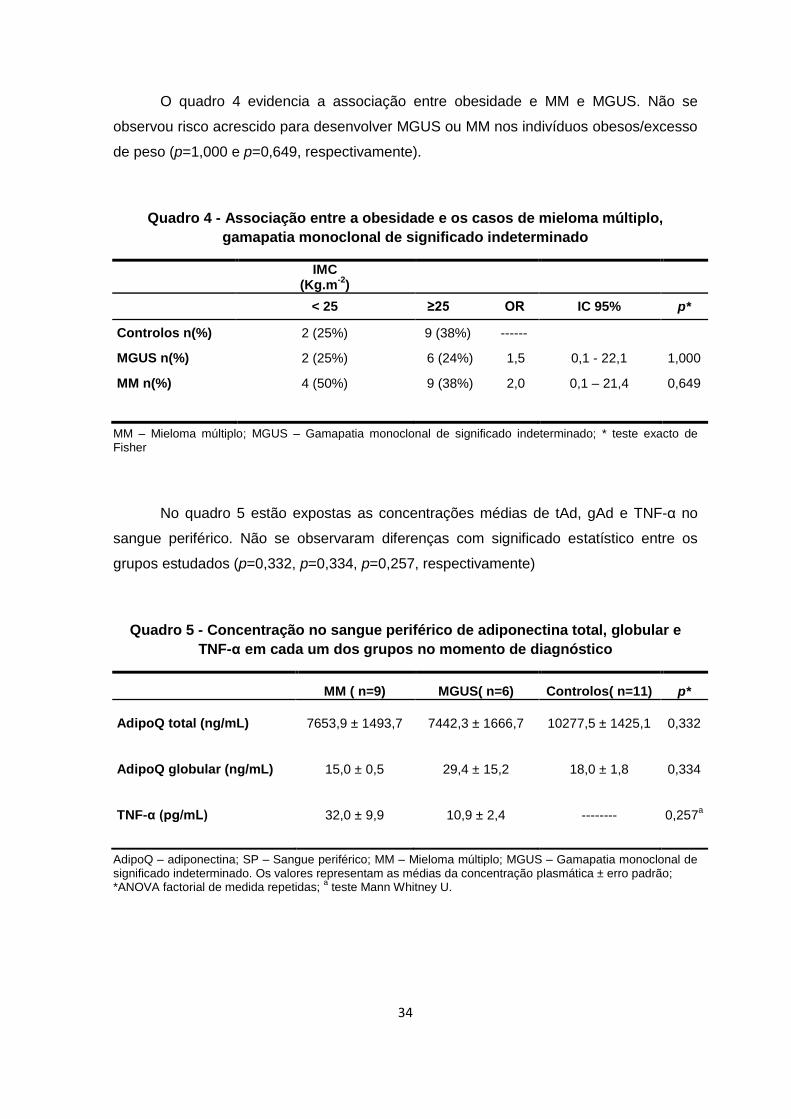
O quadro 4 evidencia a associação entre obesidade e MM e MGUS. Não se
observou risco acrescido para desenvolver MGUS ou MM nos indivíduos obesos/excesso
de peso (p=1,000 e p=0,649, respectivamente).
Quadro 4 - Associação entre a obesidade e os casos de mieloma múltiplo,
gamapatia monoclonal de significado indeterminado
IMC
(Kg.m-2
)
< 25 ≥25 OR IC 95% p*
Controlos n(%) 2 (25%) 9 (38%) ------
MGUS n(%) 2 (25%) 6 (24%) 1,5 0,1 - 22,1 1,000
MM n(%) 4 (50%) 9 (38%) 2,0 0,1 – 21,4 0,649
MM – Mieloma múltiplo; MGUS – Gamapatia monoclonal de significado indeterminado; * teste exacto de Fisher
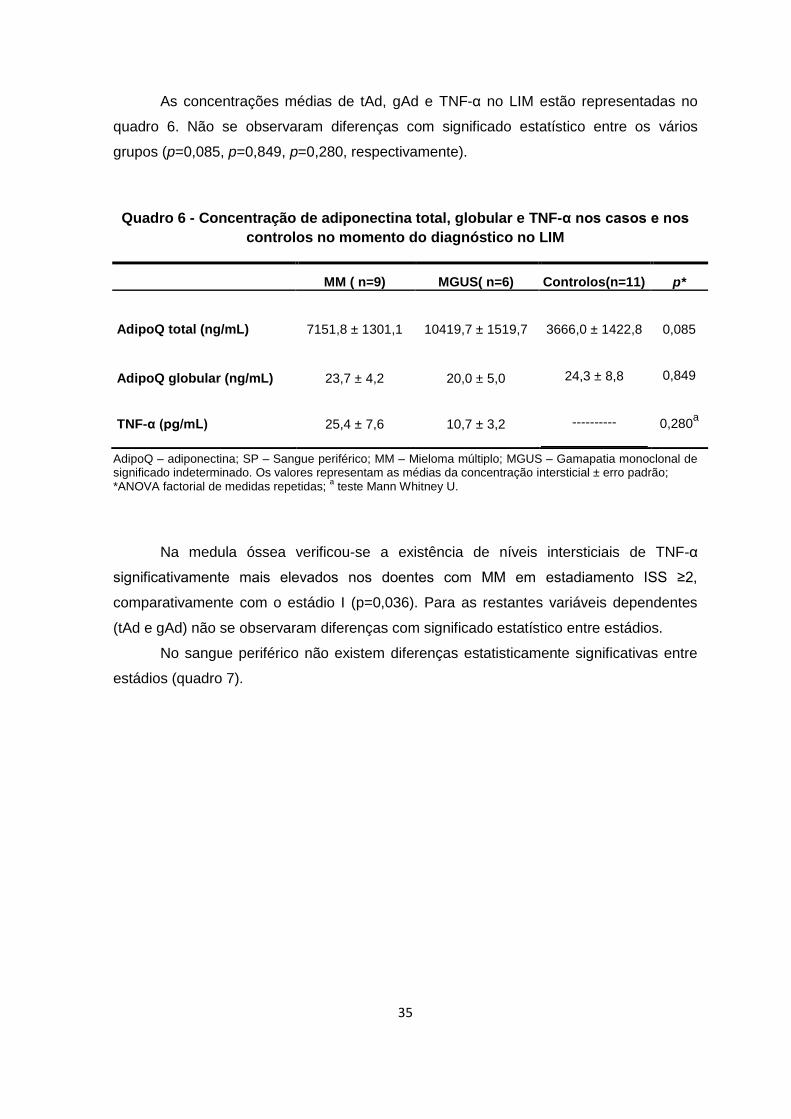
No quadro 5 estão expostas as concentrações médias de tAd, gAd e TNF-α no
sangue periférico. Não se observaram diferenças com significado estatístico entre os
grupos estudados (p=0,332, p=0,334, p=0,257, respectivamente)
Quadro 5 - Concentração no sangue periférico de adiponectina total, globular e
TNF-α em cada um dos grupos no momento de diagnóstico
MM ( n=9) MGUS( n=6) Controlos( n=11) p*
AdipoQ total (ng/mL)
7653,9 ± 1493,7
7442,3 ± 1666,7
10277,5 ± 1425,1
0,332
AdipoQ globular (ng/mL) TNF-α (pg/mL)
15,0 ± 0,5
32,0 ± 9,9
29,4 ± 15,2
10,9 ± 2,4
18,0 ± 1,8
--------
0,334
0,257a
AdipoQ – adiponectina; SP – Sangue periférico; MM – Mieloma múltiplo; MGUS – Gamapatia monoclonal de significado indeterminado. Os valores representam as médias da concentração plasmática ± erro padrão; *ANOVA factorial de medida repetidas;
a teste Mann Whitney U.
35
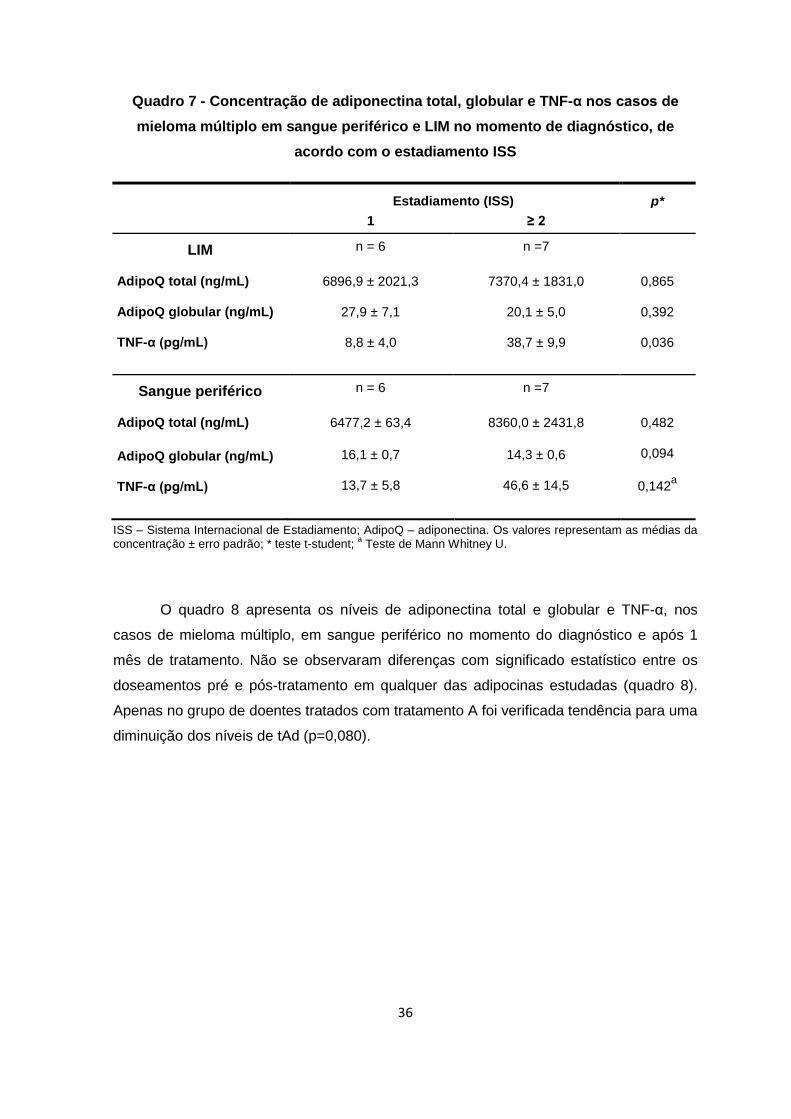
As concentrações médias de tAd, gAd e TNF-α no LIM estão representadas no
quadro 6. Não se observaram diferenças com significado estatístico entre os vários
grupos (p=0,085, p=0,849, p=0,280, respectivamente).
Quadro 6 - Concentração de adiponectina total, globular e TNF-α nos casos e nos
controlos no momento do diagnóstico no LIM
MM ( n=9) MGUS( n=6) Controlos(n=11) p*
AdipoQ total (ng/mL)
7151,8 ± 1301,1
10419,7 ± 1519,7
3666,0 ± 1422,8
0,085
AdipoQ globular (ng/mL) TNF-α (pg/mL)
23,7 ± 4,2
25,4 ± 7,6
20,0 ± 5,0
10,7 ± 3,2
24,3 ± 8,8
----------
0,849
0,280a
AdipoQ – adiponectina; SP – Sangue periférico; MM – Mieloma múltiplo; MGUS – Gamapatia monoclonal de significado indeterminado. Os valores representam as médias da concentração intersticial ± erro padrão; *ANOVA factorial de medidas repetidas;
a teste Mann Whitney U.
Na medula óssea verificou-se a existência de níveis intersticiais de TNF-α
significativamente mais elevados nos doentes com MM em estadiamento ISS ≥2,
comparativamente com o estádio I (p=0,036). Para as restantes variáveis dependentes
(tAd e gAd) não se observaram diferenças com significado estatístico entre estádios.
No sangue periférico não existem diferenças estatisticamente significativas entre
estádios (quadro 7).
36
Quadro 7 - Concentração de adiponectina total, globular e TNF-α nos casos de
mieloma múltiplo em sangue periférico e LIM no momento de diagnóstico, de
acordo com o estadiamento ISS
Estadiamento (ISS) p*
1 ≥ 2
LIM n = 6 n =7
AdipoQ total (ng/mL)
6896,9 ± 2021,3 7370,4 ± 1831,0 0,865
AdipoQ globular (ng/mL) TNF-α (pg/mL)
27,9 ± 7,1
8,8 ± 4,0
20,1 ± 5,0
38,7 ± 9,9
0,392
0,036
Sangue periférico n = 6 n =7
AdipoQ total (ng/mL)
6477,2 ± 63,4 8360,0 ± 2431,8 0,482
AdipoQ globular (ng/mL) TNF-α (pg/mL)
16,1 ± 0,7
13,7 ± 5,8
14,3 ± 0,6
46,6 ± 14,5
0,094
0,142a
ISS – Sistema Internacional de Estadiamento; AdipoQ – adiponectina. Os valores representam as médias da concentração ± erro padrão; * teste t-student;
a Teste de Mann Whitney U.
O quadro 8 apresenta os níveis de adiponectina total e globular e TNF-α, nos
casos de mieloma múltiplo, em sangue periférico no momento do diagnóstico e após 1
mês de tratamento. Não se observaram diferenças com significado estatístico entre os
doseamentos pré e pós-tratamento em qualquer das adipocinas estudadas (quadro 8).
Apenas no grupo de doentes tratados com tratamento A foi verificada tendência para uma
diminuição dos níveis de tAd (p=0,080).
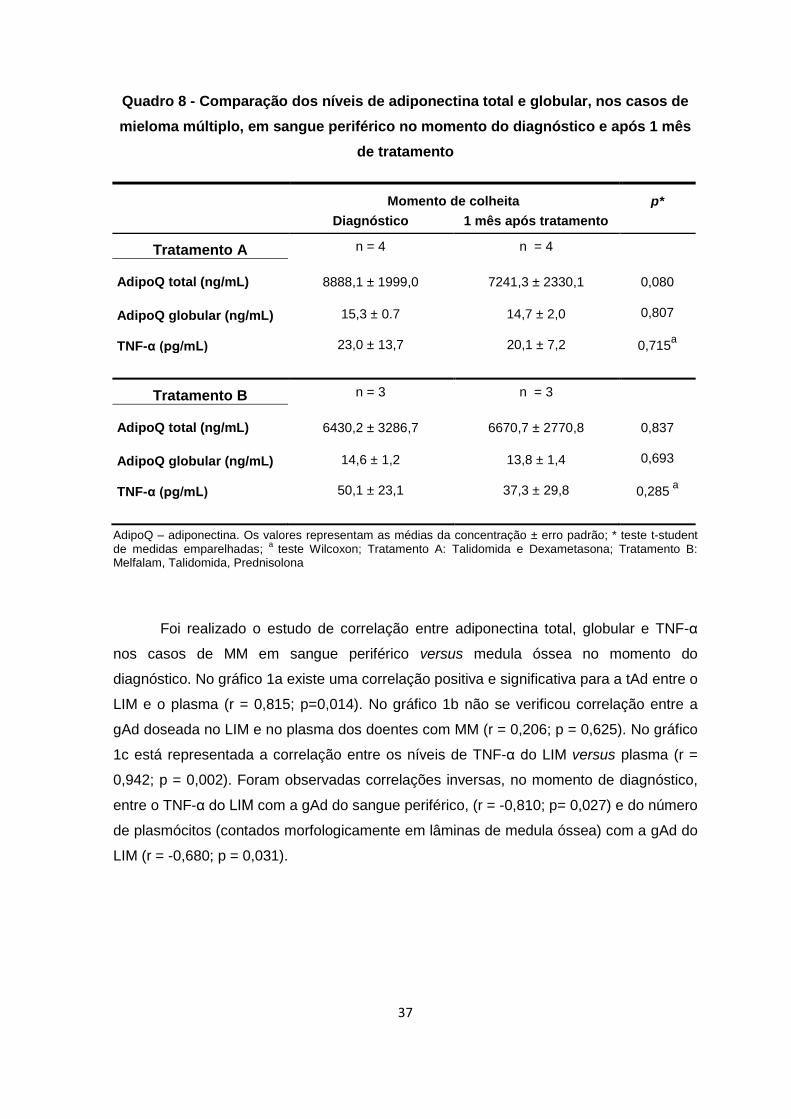
37
Quadro 8 - Comparação dos níveis de adiponectina total e globular, nos casos de
mieloma múltiplo, em sangue periférico no momento do diagnóstico e após 1 mês
de tratamento
Momento de colheita p*
Diagnóstico 1 mês após tratamento
Tratamento A n = 4 n = 4
AdipoQ total (ng/mL)
8888,1 ± 1999,0 7241,3 ± 2330,1 0,080
AdipoQ globular (ng/mL) TNF-α (pg/mL)
15,3 ± 0.7
23,0 ± 13,7
14,7 ± 2,0
20,1 ± 7,2
0,807
0,715a
Tratamento B n = 3 n = 3
AdipoQ total (ng/mL)
6430,2 ± 3286,7 6670,7 ± 2770,8 0,837
AdipoQ globular (ng/mL) TNF-α (pg/mL)
14,6 ± 1,2
50,1 ± 23,1
13,8 ± 1,4
37,3 ± 29,8
0,693
0,285 a
AdipoQ – adiponectina. Os valores representam as médias da concentração ± erro padrão; * teste t-student de medidas emparelhadas;
a teste Wilcoxon; Tratamento A: Talidomida e Dexametasona; Tratamento B:
Melfalam, Talidomida, Prednisolona
Foi realizado o estudo de correlação entre adiponectina total, globular e TNF-α
nos casos de MM em sangue periférico versus medula óssea no momento do
diagnóstico. No gráfico 1a existe uma correlação positiva e significativa para a tAd entre o
LIM e o plasma (r = 0,815; p=0,014). No gráfico 1b não se verificou correlação entre a
gAd doseada no LIM e no plasma dos doentes com MM (r = 0,206; p = 0,625). No gráfico
1c está representada a correlação entre os níveis de TNF-α do LIM versus plasma (r =
0,942; p = 0,002). Foram observadas correlações inversas, no momento de diagnóstico,
entre o TNF-α do LIM com a gAd do sangue periférico, (r = -0,810; p= 0,027) e do número
de plasmócitos (contados morfologicamente em lâminas de medula óssea) com a gAd do
LIM (r = -0,680; p = 0,031).
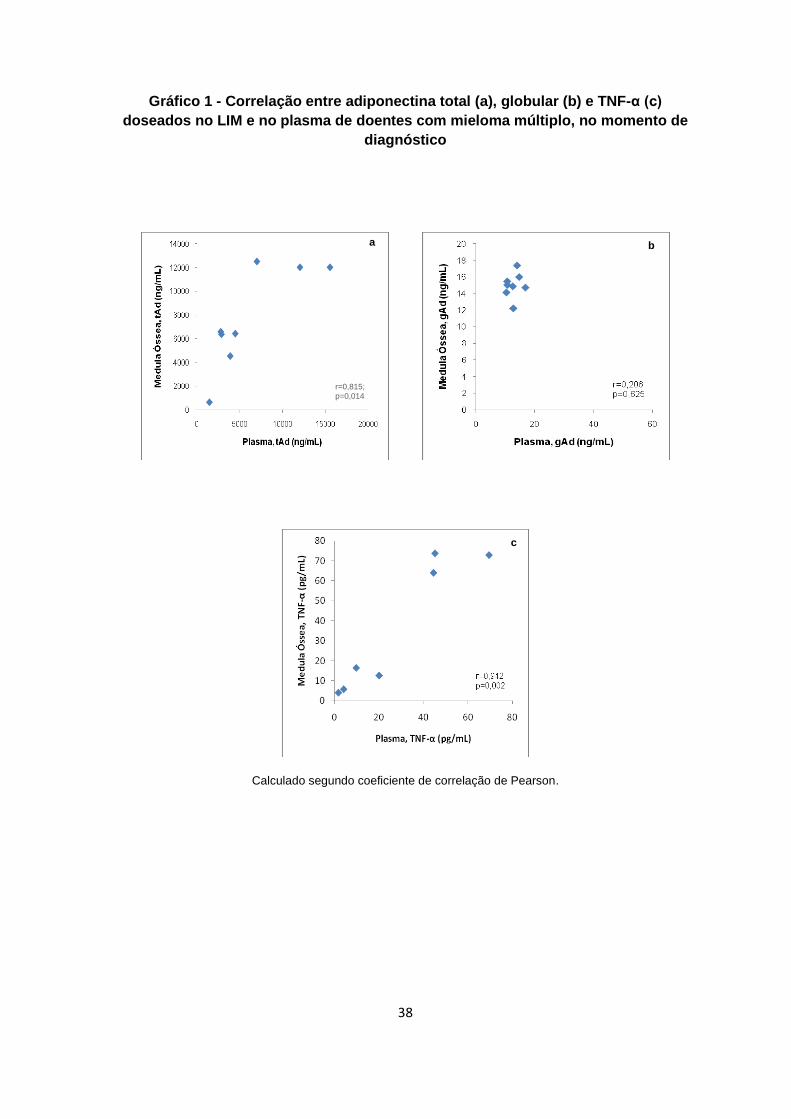
38
Gráfico 1 - Correlação entre adiponectina total (a), globular (b) e TNF-α (c)
doseados no LIM e no plasma de doentes com mieloma múltiplo, no momento de
diagnóstico
Calculado segundo coeficiente de correlação de Pearson.
r=0,815;
p=0,014
a b
c
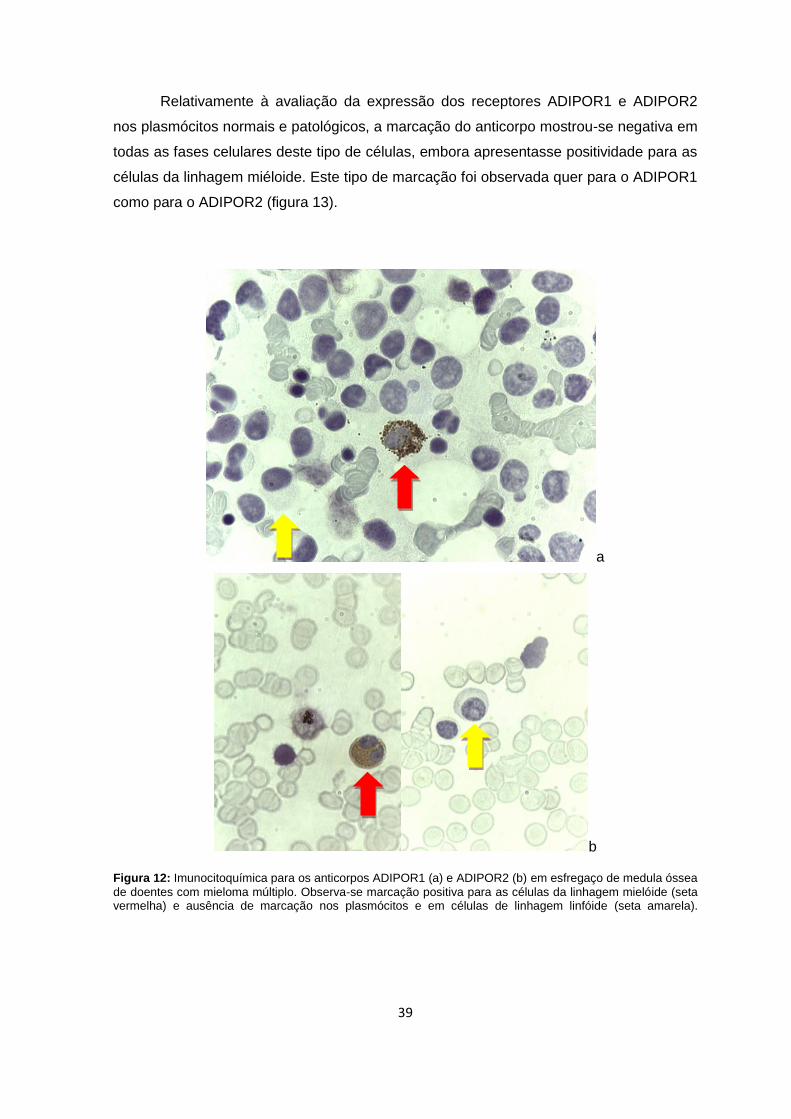
39
Relativamente à avaliação da expressão dos receptores ADIPOR1 e ADIPOR2
nos plasmócitos normais e patológicos, a marcação do anticorpo mostrou-se negativa em
todas as fases celulares deste tipo de células, embora apresentasse positividade para as
células da linhagem miéloide. Este tipo de marcação foi observada quer para o ADIPOR1
como para o ADIPOR2 (figura 13).
a
b
Figura 12: Imunocitoquímica para os anticorpos ADIPOR1 (a) e ADIPOR2 (b) em esfregaço de medula óssea
de doentes com mieloma múltiplo. Observa-se marcação positiva para as células da linhagem mielóide (seta vermelha) e ausência de marcação nos plasmócitos e em células de linhagem linfóide (seta amarela).

5. Discussão

41
5. Discussão
Actualmente, considera-se que o tecido adiposo é um órgão endócrino activo.
Apesar da sua capacidade de produção de várias proteínas, entre as quais a
adiponectina, esta parece desempenhar um importante papel no metabolismo da glicose
e dos lípidos, assim como no desenvolvimento de neoplasias. A associação entre
obesidade e doenças hemato-oncológicas tem sido objectivo de estudo nos últimos anos.
Os estudos epidemiológicos sugerem uma associação entre ser obeso e risco acrescido
de desenvolver síndrome mielodisplásicos, leucemias e mieloma múltiplo (Avcu et al.,
2006; Petridou et al., 2006; Barb et al., 2007; Dalamaga et al., 2009; Reseland et al.,
2009). No que diz respeito ao MM, alguns trabalhos, incluindo meta-análises,
demonstraram que indivíduos obesos têm maior predisposição para desenvolver MM
(Birmann et al., 2007; Larsson and Wolk, 2007; Crawford et al., 2010). No presente
estudo, não observamos uma sobre-representação de indivíduos com excesso de
peso/obesos no grupo de doentes MM (p=0,649), em contraste com os resultados
previamente descritos na literatura. A ausência de associação observada no nosso
estudo poderá dever-se ao reduzido número de indivíduos incluídos no trabalho, que
resulta na diminuição do poder estatístico. No entanto, verificamos que não existiam
diferenças significativas de IMC entre os vários grupos estudados, conferindo maior
homogeneidade inter-grupos e reduzindo a influência desta co-variável nos níveis
circulantes de adiponectina.
A baixa concentração circulante de adiponectina tem sido descrita como factor de
risco para o desenvolvimento de patologias neoplásicas, daí que o estudo da sua
fisiopatologia e mecanismos de actuação adicionarão informação sobre a associação
entre obesidade e cancro. O IMC e a percentagem de gordura estão inversamente
correlacionados com os níveis circulantes de adiponectina (Barb et al., 2007; Birmann et
al., 2007; Larsson and Wolk, 2007).
Desta forma, foi já referido que o doseamento dos níveis de adiponectina
circulante poderá vir a ser usado como um biomarcador no rastreio do cancro e em
doenças relacionadas com a obesidade (Kelesidis et al., 2006). Estudos prévios
mostraram a existência de correlação inversa entre a concentração plasmática de
adiponectina e o desenvolvimento de várias patologias malignas, como o cancro da
mama (Korner et al., 2007; Tworoger et al., 2007), do endométrio (Soliman, 2005; Cust et

42
al., 2007), da próstata (Michalakis et al., 2007), colo-rectal (Wei et al., 2005), gástrico
(Ishikawa et al., 2005) e renal (Spyridopoulos et al., 2007). Relativamente ao risco para
neoplasias hemato-oncológicas, alguns trabalhos também suportam o papel protector da
adiponectina (Avcu et al., 2006; Petridou et al., 2006; Dalamaga et al., 2009; Reseland et
al., 2009).
Dalamaga et al. (2009) e Reseland et al. (2009) observaram concentrações
significativamente mais baixas de adiponectina total nos doentes com MM. Não foram
observadas diferenças com significado estatístico nos níveis circulantes de tAd entre os
MM e os controlos no presente estudo. Assim, os mesmos resultados sugerem que a tAd
não parece discriminar entre MM e controlos, e portanto, não deverá estar envolvida na
fisiopatologia do MM. Nos trabalhos caso-controlo de Dalamaga et al. (2009) foram
incluídos 73 casos de MM e 73 controlos, enquanto no de Reseland et al. (2009)
participaram 23 casos de MM e 23 controlos. No nosso estudo participaram apenas 13
casos de MM e 12 controlos, portanto, limitando a extrapolação de resultados e a
consistência da associação. Contudo, no estudo de Dalamaga et al. (2009) os indivíduos
do grupo MM tinham IMC significativamente mais elevado do que os controlos, não
existindo informação do IMC no estudo de Reseland et al. (2009).
Actualmente, sabe-se que a adiponectina circulante é composta por várias
isoformas com percentagens relativas, que poderão eventualmente variar de acordo com
condições fisiológicas distintas (Kelesidis et al., 2006). Doseamos a gAd e a fAd, embora
também na gAd não tenham sido observadas diferenças entre MM e controlos.
Considerando que a gAd possui maior potência de activação ADIPOR1 (Alexander et al.,
2008; Reseland et al., 2009) tínhamos colocado a hipótese de que a gAd poderia estar
diminuída nos MM quando comparados com os controlos. Adicionamos ao já descrito
doseamento de adiponectina no sangue periférico, o doseamento de tAd e gAd no LIM
dos doentes com MM, com estado pré-neoplásico de MGUS e em 3 controlos. Não se
observaram diferenças com significado estatístico entre MM e MGUS para a tAd e gAd
(p>0,05).
A classificação ISS encontra-se associada com o prognóstico do MM (Kyle and
Rajkumar, 2009). Na análise efectuada de acordo com o estadiamento ISS verificamos
ausência de associação com a adiponectina no LIM mas tendência para níveis
sanguíneos mais baixos de gAd nos indivíduos com estádios ≥2 (p=0,094), portanto
indicador de pior prognóstico. Assim, a gAd em circulação através do seu
efeito/interacção periférica com a β2-microbulina e albumina, poderá reflectir uma relação
fisiológica com estes parâmetros de estadiamento e não com um efeito endócrino que

43
corresponde a uma acção protectora da gAd de MM mais agressivo. Também Dalamaga
et al. (2009) obtiveram resultados sobreponíveis, sugerindo a ausência de valor
prognóstico da adiponectina no MM.
Neste estudo verificamos que a concentração plasmática de gAd se manteve
semelhante no diagnóstico e 1 mês após o tratamento (p>0,05). Relativamente à tAd
observou-se uma tendência para os seus valores circulantes diminuírem após tratamento
com talidomida e dexametasona (tratamento A) (p=0,080), mas não após o tratamento B
(melfalam, talidomida e prednisolona). Reseland et al. (2009) não observaram variações
nos níveis circulantes de tAd em doentes com MM tratados com tratamento A. Contudo, o
número reduzido de doentes com doseamento pré e pós-tratamento no nosso estudo e a
existência de tratamento prévio no grupo submetido ao tratamento A no estudo de
Reseland et al. (2009) poderão explicar a ausência de concordância. Os resultados
apresentados sugerem, na nossa perspectiva, um eventual impacto directo do tratamento
A nos níveis circulantes de tAd, talvez devido ao efeito dos fármacos talidomida e
dexametasona nos adipócitos ou com o efeito deste tratamento no microambiente
medular, uma vez que estes fármacos influenciam as células das linhagens mielóide e
imuno-inflamatórias que interagem com os adipócitos. No entanto não existe literatura
que aborde esta possível associação, daí que seria uma das formas de futuros estudos
no que diz respeito à interacção do tratamento na diminuição dos níveis de adiponectina.
A diminuição da produção de adiponectina pode directa ou indirectamente,
através da resistência à insulina e da promoção da inflamação contribuir largamente para
o aumento do risco de desenvolvimento tumoral. Em situações de obesidade, a
resistência à insulina é o produto da diminuição da adiponectina mas também funciona
como indutor de outras citoquinas pró-inflamatórias, como o TNF-α. Neste estudo foram
avaliados os níveis de TNF-α no sangue periférico e no LIM. Mediante os resultados
obtidos verifica-se, a ausência de diferenças com significado estatístico entre os grupos
de MGUS e MM para os níveis circulantes e para a concentração na medula óssea.
Relativamente aos níveis de TNF-α circulantes antes e após tratamento, verificamos
ausência de diferenças com significado estatístico. É possível que o tempo de tratamento
tenha sido insuficiente para que influenciasse os níveis de TNF-α.
Estudos prévios referem que o TNF-α apresenta efeitos diversos e opostos na
patogénese e no crescimento das células plasmáticas tumorais (Davies et al., 2000;
Jurisic and Colovic, 2002). Assim, sabe-se que o TNF-α desempenha funções

44
importantes na diferenciação e transformação dos linfócitos B no micro ambiente da
medula óssea, pela complexa comunicação com as células estromais. No entanto, foi
demonstrado in vitro que depois de adquirir receptores apropriados pode induzir a
apoptose em células plasmáticas neoplásicas (Davies et al., 2000; Jurisic and Colovic,
2002). Neste estudo, observamos que o TNF-α poderá vir a ser um marcador tumoral
com significado prognóstico para doentes com MM, isto porque em estádios mais
avançados da doença apresenta concentrações medulares significativamente mais
elevadas. Estudos anteriores revelam que altas concentrações circulantes de TNF-α se
encontram associadas a pior prognóstico (Jourdan et al., 1999; Balkwill, 2006). Assim, o
facto de o TNF-α apresentar um efeito indutor do crescimento de células plasmáticas
tumorais (Jourdan et al., 1999; Kelesidis et al., 2006) e também o facto de nesta patologia
existir uma grande quantidade de células estromais que contribuem para a
sobreprodução desta citoquina (Jurisic and Colovic, 2002), pode justificar a elevada
concentração de TNF-α. Outros estudos parecem suportar os nossos resultados, já que
revelaram que o efeito estimulatório do microambiente tumoral aumenta a expressão de
vários genes relacionados com citoquinas inflamatórias, incluindo o TNF-α (Wang et al.,
2005a).
Sabe-se que a adiponectina sofre clivagens e processamentos pós-tradução,
através da acção de enzimas como a elastase dos leucócitos (Kadowaki and Yamauchi,
2005; Waki et al., 2005) para produzir as diferentes isoformas, incluindo a gAd.
Observamos correlação entre a concentração de adiponectina total do LIM e os níveis
plasmáticos (r = 0,815; p=0,014). No entanto, não se observou correlação significativa
entre a gAd da medula óssea e a gAd plasmática. Estes resultados poderão sugerir que
existem mecanismos de processamento distintos da adiponectina na medula óssea. O
presente trabalho sugere que o ambiente da medula óssea poderá influenciar de
diferente modo o processamento da gAd. Outros trabalhos demonstraram previamente
que a adiponectina é produzida na medula óssea por adipócitos e que existe em níveis
mais elevados comparativamente com os níveis plasmáticos (Berner et al., 2004; Iversen
and Wiig, 2005). O nosso estudo evidencia que também em MM o ambiente medular
mantém a produção de adiponectina.
Alguns estudos demonstraram, previamente, que os níveis de TNF-α em doentes
com MM se encontram elevados na medula óssea, na medida em que as células
plasmáticas neoplásicas segregam significantes quantidades desta citocina (Davies et al.,

45
2000). Além disso, o TNF-α promove a proliferação da linhagem das células plasmáticas
neoplásicas e o aumento desta citocina a nível plasmático (Jourdan et al., 1999).
Observámos correlação entre o TNF-α doseado no LIM e no plasma de doentes com MM
(r = 0,942; p = 0,002). Outro estudo referencia que a grande produção de TNF-α pelas
células do estroma da medula óssea provavelmente contribuem para o aumento dos
valores plasmáticos, suportando que estes dados indicam a significância da
determinação desta citoquina em relação ao número de células plasmáticas neoplásicas
existentes. (Jurisic and Colovic, 2002)
Existe ausência de concordância entre os estudos anteriores relativamente à
associação, entre adiponectina total e TNF-α, uma vez que alguns afirmam que a
adiponectina inibe a produção de TNF-α (Kern et al., 2003; Bernstein et al., 2006; Gumbs,
2008; Dalamaga et al., 2009), enquanto que outros referem que a relação inversa entre
adiponectina e TNF-α tem origem em situações de resistência à insulina e da
consequente promoção da inflamação (Wang et al., 2005a; Barb et al., 2007). Neste
estudo observamos correlação inversa, no momento do diagnóstico, entre os níveis de
TNF-α do LIM com a concentração de adiponectina globular plasmática, (r = -0,810;
p=0,027) sugerindo que o mecanismo de acção destas citocinas possa passar pela sua
interacção, o que está de acordo com evidências de estudos anteriores, adicionando
informação relativamente ao efeito especificamente da gAd nos níveis de TNF-α na
medula óssea.
Vários estudos indicam que a adiponectina tem efeitos nas células mesenquimais
do microambiente da medula óssea parecendo desempenhar um papel importante na
regulação da maturação deste tipo de células (Berner et al., 2004). Os adipócitos, as
células do estroma, os osteoblastos e os miócitos derivam das células mesenquimais
(Yokota et al., 2002) suportando eventual influência da concentração de adiponectina na
diferenciação selectiva das linhagens celulares. Outro estudo revela que a adiponectina
detectada na medula óssea, pode inibir clones isolados de progenitores de macrófagos,
mas não inibe a mielopoiese (Yokota et al., 2002). No entanto, a linfopoiese B é inibida
por esta citocina através de mecanismos de acção nas células do estroma. Estes
resultados estão de acordo com a correlação encontrada no nosso estudo, no qual se
observou correlação inversa entre o número de plasmócitos contabilizados
morfologicamente em lâminas de medula óssea, com a adiponectina globular intersticial

46
(r = -0,680; p = 0,031). Alternativamente, considerando a correlação inversa da
adiponectina com o TNF-α, é possível que a adiponectina possa indirectamente diminuir
o crescimento dos plasmócitos através do seu efeito supressor na produção de TNF-α e
consequentemente diminuição da viabilidade dos plasmócitos.
Não existem estudos de expressão dos receptores de adiponectina em tumores
líquidos. No entanto, vários trabalhos demonstram a presença dos ADIPOR1 e ADIPOR2
em vários tecidos normais e tumorais (Berner et al., 2004; Miller et al., 2009). No presente
estudo não se observou marcação para ambos os receptores em plasmócitos normais ou
patológicos para qualquer fase celular. Deste modo, concluímos que a eventual
associação entre níveis circulantes de adiponectina e MM descritos na literatura
(Dalamaga et al., 2009; Reseland et al., 2009) se devem a uma possível acção indirecta
da adiponectina na biologia dos plasmócitos ou dos linfócitos (Crawford et al., 2010). As
células da linhagem mielóide estavam marcadas com ADIPOR1 e ADIPOR2, suportando
uma acção fisiológica da adiponectina neste tipo de células. Existem outros estudos que
argumentam a adiponectina como factor protector em determinadas leucemias da série
mielóide, resta-nos saber se estas células têm capacidade de produção desta citocina, ou
será devida à existente em circulação (Yokota et al., 2002; Iversen and Wiig, 2005;
Petridou et al., 2006; Crawford et al., 2010).

6. Conclusão e perspectivas futuras

48
6. Conclusão e perspectivas futuras
Os resultados do presente estudo sugerem que a adiponectina e o TNF-α não
influenciam o desenvolvimento de mieloma múltiplo, na medida em que quer a nível
plasmático quer a nível do LIM os níveis de gAd, fAd e TNF-α não se encontram
alterados. No entanto, os resultados indicam uma correlação entre os níveis de TNF-α
plasmático e o LIM. Além disso, apontam para a associação com o estádio e prognóstico
do mieloma múltiplo, pois verificamos que em estádios mais avançados da doença
existem níveis aumentados na medula óssea. Estudos futuros deverão confirmar o
doseamento de TNF-α plasmático (menos invasivo) como biomarcador de prognóstico no
mieloma múltiplo. Também se constatou a correlação inversa entre o TNF-α do líquido
intersticial e a gAd plasmática, reflectindo um eventual efeito protector indirecto da gAd
para estádios mais avançados da doença (ISS≥2), devido à sua relação com o TNF-α.
Cumulativamente, a gAd está inversamente associada com o número de plasmócitos, o
que suporta a existência de um mecanismo indirecto para a acção da adiponectina nos
MM, pois não encontramos expressão dos receptores ADIPOR1 E ADIPOR2 nas células
da série linfóide.
Após o trabalho desenvolvido neste projecto e atendendo ao facto de que os
nossos resultados suportam algumas hipóteses mas efectivamente levantam muitas
questões, consideramos como perspectivas e trabalhos futuros o seguinte:
- Incrementar o número de doentes e controlos, contribuindo para aumentar o
poder estatístico das análises.
- Estudar interacção da IL-6 e outras citoquinas com a adiponectina, nas células
plasmáticas dos MM.
- Estudar a possível presença de receptores da adiponectina nas linhagens
celulares das patologias hematológicas mielóides, no sentido de encontrar possíveis
associações entre a produção de adiponectina e o desenvolvimento e prognóstico dessas
patologias.

49
- Estudar a influência da talidomida e da dexametasona in vitro nos adipócitos e
na produção de adiponectina.
- Estudar a relação entre o TNF-α e os plasmócitos.
- Correlacionar os resultados do presente estudo com mediadores de prognóstico
(sobrevivência global, tempo até progressão/recidiva), avaliando o seu valor preditivo de
resposta ao tratamento e valor prognóstico.

50
7. Referências
Alexander, D.D., Mink, P.J., Adami, H.O., Cole, P., Mandel, J.S., Oken, M.M., and
Trichopoulos, D. (2007). Multiple myeloma: a review of the epidemiologic literature. Int J
Cancer 120 Suppl 12, 40-61.
Alexander, S.P., Mathie, A., and Peters, J.A. (2008). Guide to Receptors and Channels
(GRAC), 3rd edition. Br J Pharmacol 153 Suppl 2, S1-209.
Arkan, M.C., Hevener, A.L., Greten, F.R., Maeda, S., Li, Z.W., Long, J.M., Wynshaw-
Boris, A., Poli, G., Olefsky, J., and Karin, M. (2005). IKK-beta links inflammation to
obesity-induced insulin resistance. Nat Med 11, 191-198.
Attaelmannan, M., and Levinson, S.S. (2000). Understanding and identifying monoclonal
gammopathies. Clin Chem 46, 1230-1238.
Avcu, F., Ural, A.U., Yilmaz, M.I., Bingol, N., Nevruz, O., and Caglar, K. (2006).
Association of plasma adiponectin concentrations with chronic lymphocytic leukemia and
myeloproliferative diseases. Int J Hematol 83, 254-258.
Balkwill, F. (2006). TNF-alpha in promotion and progression of cancer. Cancer Metastasis
Rev 25, 409-416.
Barb, D., Williams, C.J., Neuwirth, A.K., and Mantzoros, C.S. (2007). Adiponectin in
relation to malignancies: a review of existing basic research and clinical evidence. Am J
Clin Nutr 86, s858-866.
Barbara J. Bain, D.M.C., Bridget S. Wilkins (2010). Bone Marrow Pathology. In Wiley-
blackwell, pp. 640.
Bataille, R., Grenier, J., and Sany, J. (1984). Beta-2-microglobulin in myeloma: optimal
use for staging, prognosis, and treatment--a prospective study of 160 patients. Blood 63,
468-476.

51
Berg, A.H., Combs, T.P., Du, X., Brownlee, M., and Scherer, P.E. (2001). The adipocyte-
secreted protein Acrp30 enhances hepatic insulin action. Nat Med 7, 947-953.
Berner, H.S., Lyngstadaas, S.P., Spahr, A., Monjo, M., Thommesen, L., Drevon, C.A.,
Syversen, U., and Reseland, J.E. (2004). Adiponectin and its receptors are expressed in
bone-forming cells. Bone 35, 842-849.
Bernstein, L.E., Berry, J., Kim, S., Canavan, B., and Grinspoon, S.K. (2006). Effects of
etanercept in patients with the metabolic syndrome. Arch Intern Med 166, 902-908.
Beutler, E.L., M.A.; Coller, B.S.; Kipss, T.J.; Seligsohn U. (2001). Williams Hematology, M.
Hill, ed., pp. 1943
Birmann, B.M., Giovannucci, E., Rosner, B., Anderson, K.C., and Colditz, G.A. (2007).
Body mass index, physical activity, and risk of multiple myeloma. Cancer Epidemiol
Biomarkers Prev 16, 1474-1478.
Brakenhielm, E., Veitonmaki, N., Cao, R., Kihara, S., Matsuzawa, Y., Zhivotovsky, B.,
Funahashi, T., and Cao, Y. (2004). Adiponectin-induced antiangiogenesis and antitumor
activity involve caspase-mediated endothelial cell apoptosis. Proc Natl Acad Sci U S A
101, 2476-2481.
Bub, J.D., Miyazaki, T., and Iwamoto, Y. (2006). Adiponectin as a growth inhibitor in
prostate cancer cells. Biochem Biophys Res Commun 340, 1158-1166.
Caligaris-Cappio, F., Bergui, L., Tesio, L., Pizzolo, G., Malavasi, F., Chilosi, M., Campana,
D., van Camp, B., and Janossy, G. (1985). Identification of malignant plasma cell
precursors in the bone marrow of multiple myeloma. J Clin Invest 76, 1243-1251.
Calle, E.E., Rodriguez, C., Walker-Thurmond, K., and Thun, M.J. (2003). Overweight,
obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N Engl
J Med 348, 1625-1638.
Cesana, C., Klersy, C., Barbarano, L., Nosari, A.M., Crugnola, M., Pungolino, E.,
Gargantini, L., Granata, S., Valentini, M., and Morra, E. (2002). Prognostic factors for

52
malignant transformation in monoclonal gammopathy of undetermined significance and
smoldering multiple myeloma. J Clin Oncol 20, 1625-1634.
Crawford, L.J., Peake, R., Price, S., Morris, T.C., and Irvine, A.E. (2010). Adiponectin is
produced by lymphocytes and is a negative regulator of granulopoiesis. J Leukoc Biol.
Cust, A.E., Kaaks, R., Friedenreich, C., Bonnet, F., Laville, M., Lukanova, A., Rinaldi, S.,
Dossus, L., Slimani, N., Lundin, E., et al. (2007). Plasma adiponectin levels and
endometrial cancer risk in pre- and postmenopausal women. J Clin Endocrinol Metab 92,
255-263.
Dalamaga, M., Karmaniolas, K., Panagiotou, A., Hsi, A., Chamberland, J., Dimas, C.,
Lekka, A., and Mantzoros, C.S. (2009). Low circulating adiponectin and resistin, but not
leptin, levels are associated with multiple myeloma risk: a case-control study. Cancer
Causes Control 20, 193-199.
Davies, F.E., Rollinson, S.J., Rawstron, A.C., Roman, E., Richards, S., Drayson, M.,
Child, J.A., and Morgan, G.J. (2000). High-producer haplotypes of tumor necrosis factor
alpha and lymphotoxin alpha are associated with an increased risk of myeloma and have
an improved progression-free survival after treatment. J Clin Oncol 18, 2843-2851.
Delaigle, A.M., Jonas, J.C., Bauche, I.B., Cornu, O., and Brichard, S.M. (2004). Induction
of adiponectin in skeletal muscle by inflammatory cytokines: in vivo and in vitro studies.
Endocrinology 145, 5589-5597.
Dietze-Schroeder, D., Sell, H., Uhlig, M., Koenen, M., and Eckel, J. (2005). Autocrine
action of adiponectin on human fat cells prevents the release of insulin resistance-
inducing factors. Diabetes 54, 2003-2011.
Dieudonne, M.N., Bussiere, M., Dos Santos, E., Leneveu, M.C., Giudicelli, Y., and
Pecquery, R. (2006). Adiponectin mediates antiproliferative and apoptotic responses in
human MCF7 breast cancer cells. Biochem Biophys Res Commun 345, 271-279.
Dimopoulos, M.A., Moulopoulos, L.A., Maniatis, A., and Alexanian, R. (2000). Solitary
plasmacytoma of bone and asymptomatic multiple myeloma. Blood 96, 2037-2044.

53
Ding, G., Qin, Q., He, N., Francis-David, S.C., Hou, J., Liu, J., Ricks, E., and Yang, Q.
(2007). Adiponectin and its receptors are expressed in adult ventricular cardiomyocytes
and upregulated by activation of peroxisome proliferator-activated receptor gamma. J Mol
Cell Cardiol 43, 73-84.
Dispenzieri, A., Gertz, M.A., Therneau, T.M., and Kyle, R.A. (2001). Retrospective cohort
study of 148 patients with polyclonal gammopathy. Mayo Clin Proc 76, 476-487.
Epstein, J. (1992). Myeloma phenotype: clues to disease origin and manifestation.
Hematol Oncol Clin North Am 6, 249-256.
Falcão (2007). O valor da imunofenotipagem para o diagnóstico do Mieloma Múltiplo e na
avaliação da doença residual mínima. Revista brasileira hematologia hemoterapia 1, 3-9.
Fang, X., and Sweeney, G. (2006). Mechanisms regulating energy metabolism by
adiponectin in obesity and diabetes. Biochem Soc Trans 34, 798-801.
Fanning (2008). http://emedicine.medscape.com/article/207233-overview.
Faria, J.A. (2007). Gamopatias monoclonais – critérios diagnósticos e diagnósticos
diferenciais. Revista brasileira hematologia hemoterapia, 17-22.
Finak, G., Bertos, N., Pepin, F., Sadekova, S., Souleimanova, M., Zhao, H., Chen, H.,
Omeroglu, G., Meterissian, S., Omeroglu, A., et al. (2008). Stromal gene expression
predicts clinical outcome in breast cancer. Nat Med 14, 518-527.
Fruhbeck, G., Gomez-Ambrosi, J., Muruzabal, F.J., and Burrell, M.A. (2001). The
adipocyte: a model for integration of endocrine and metabolic signaling in energy
metabolism regulation. Am J Physiol Endocrinol Metab 280, E827-847.
Greipp, P.R., San Miguel, J., Durie, B.G., Crowley, J.J., Barlogie, B., Blade, J.,
Boccadoro, M., Child, J.A., Avet-Loiseau, H., Kyle, R.A., et al. (2005). International
staging system for multiple myeloma. J Clin Oncol 23, 3412-3420.
Grethlein, S.J. (2009). http://emedicine.medscape.com/article/204369-overview.

54
Guerre-Millo, M. (2008). Adiponectin: an update. Diabetes Metab 34, 12-18.
Gumbs, A.A. (2008). Obesity, pancreatitis, and pancreatic cancer. Obes Surg 18, 1183-
1187.
Haluzik, M., Parizkova, J., and Haluzik, M.M. (2004). Adiponectin and its role in the
obesity-induced insulin resistance and related complications. Physiol Res 53, 123-129.
Hanahan, D., and Weinberg, R.A. (2000). The hallmarks of cancer. Cell 100, 57-70.
Harmening, D.M. (1997). Clinical Hematology and Fundamentals of Hemostasis, F.A.D.
Company, ed. (Philadelphia), pp. 717.
Harousseau, J.L., and Moreau, P. (2009). Autologous hematopoietic stem-cell
transplantation for multiple myeloma. N Engl J Med 360, 2645-2654.
Hu, E., Liang, P., and Spiegelman, B.M. (1996). AdipoQ is a novel adipose-specific gene
dysregulated in obesity. J Biol Chem 271, 10697-10703.
Hungria, A.B. (2007). Doença óssea em Mieloma Múltiplo. Revista brasileira hematologia
hemoterapia 1, 60-66.
Ishikawa, M., Kitayama, J., Kazama, S., Hiramatsu, T., Hatano, K., and Nagawa, H.
(2005). Plasma adiponectin and gastric cancer. Clin Cancer Res 11, 466-472.
Iversen, P.O., and Wiig, H. (2005). Tumor necrosis factor alpha and adiponectin in bone
marrow interstitial fluid from patients with acute myeloid leukemia inhibit normal
hematopoiesis. Clin Cancer Res 11, 6793-6799.
Jourdan, M., Tarte, K., Legouffe, E., Brochier, J., Rossi, J.F., and Klein, B. (1999). Tumor
necrosis factor is a survival and proliferation factor for human myeloma cells. Eur Cytokine
Netw 10, 65-70.

55
Jurisic, V., and Colovic, M. (2002). Correlation of sera TNF-alpha with percentage of bone
marrow plasma cells, LDH, beta2-microglobulin, and clinical stage in multiple myeloma.
Med Oncol 19, 133-139.
Kadowaki, T., and Yamauchi, T. (2005). Adiponectin and adiponectin receptors. Endocr
Rev 26, 439-451.
Kanazawa, I., Yamaguchi, T., Yano, S., Yamauchi, M., Yamamoto, M., and Sugimoto, T.
(2007). Adiponectin and AMP kinase activator stimulate proliferation, differentiation, and
mineralization of osteoblastic MC3T3-E1 cells. BMC Cell Biol 8, 51.
Kelesidis, I., Kelesidis, T., and Mantzoros, C.S. (2006). Adiponectin and cancer: a
systematic review. Br J Cancer 94, 1221-1225.
Kern, P.A., Di Gregorio, G.B., Lu, T., Rassouli, N., and Ranganathan, G. (2003).
Adiponectin expression from human adipose tissue: relation to obesity, insulin resistance,
and tumor necrosis factor-alpha expression. Diabetes 52, 1779-1785.
Korner, A., Pazaitou-Panayiotou, K., Kelesidis, T., Kelesidis, I., Williams, C.J., Kaprara,
A., Bullen, J., Neuwirth, A., Tseleni, S., Mitsiades, N., et al. (2007). Total and high-
molecular-weight adiponectin in breast cancer: in vitro and in vivo studies. J Clin
Endocrinol Metab 92, 1041-1048.
Kumar, S., Fonseca, R., Dispenzieri, A., Lacy, M.Q., Lust, J.A., Wellik, L., Witzig, T.E.,
Gertz, M.A., Kyle, R.A., Greipp, P.R., et al. (2003). Prognostic value of angiogenesis in
solitary bone plasmacytoma. Blood 101, 1715-1717.
Kyle, R.A., and Rajkumar, S.V. (2004). Multiple myeloma. N Engl J Med 351, 1860-1873.
Kyle, R.A., and Rajkumar, S.V. (2009). Criteria for diagnosis, staging, risk stratification
and response assessment of multiple myeloma. Leukemia 23, 3-9.
Kyle, R.A., Therneau, T.M., Rajkumar, S.V., Offord, J.R., Larson, D.R., Plevak, M.F., and
Melton, L.J., 3rd (2002). A long-term study of prognosis in monoclonal gammopathy of
undetermined significance. N Engl J Med 346, 564-569.

56
Landgren, O., Gridley, G., Turesson, I., Caporaso, N.E., Goldin, L.R., Baris, D., Fears,
T.R., Hoover, R.N., and Linet, M.S. (2006). Risk of monoclonal gammopathy of
undetermined significance (MGUS) and subsequent multiple myeloma among African
American and white veterans in the United States. Blood 107, 904-906.
Landgren, O., Kyle, R.A., Pfeiffer, R.M., Katzmann, J.A., Caporaso, N.E., Hayes, R.B.,
Dispenzieri, A., Kumar, S., Clark, R.J., Baris, D., et al. (2009). Monoclonal gammopathy of
undetermined significance (MGUS) consistently precedes multiple myeloma: a
prospective study. Blood 113, 5412-5417.
Larsson, S.C., and Wolk, A. (2007). Body mass index and risk of multiple myeloma: a
meta-analysis. Int J Cancer 121, 2512-2516.
Lauta, V.M. (2003). A review of the cytokine network in multiple myeloma: diagnostic,
prognostic, and therapeutic implications. Cancer 97, 2440-2452.
Lengauer, C., Kinzler, K.W., and Vogelstein, B. (1998). Genetic instabilities in human
cancers. Nature 396, 643-649.
Lin, P., Owens, R., Tricot, G., and Wilson, C.S. (2004). Flow cytometric
immunophenotypic analysis of 306 cases of multiple myeloma. Am J Clin Pathol 121, 482-
488.
Lope, V., Perez-Gomez, B., Aragones, N., Lopez-Abente, G., Gustavsson, P., Plato, N.,
Zock, J.P., and Pollan, M. (2008). Occupation, exposure to chemicals, sensitizing agents,
and risk of multiple myeloma in Sweden. Cancer Epidemiol Biomarkers Prev 17, 3123-
3127.
Luo, X.H., Guo, L.J., Xie, H., Yuan, L.Q., Wu, X.P., Zhou, H.D., and Liao, E.Y. (2006).
Adiponectin stimulates RANKL and inhibits OPG expression in human osteoblasts
through the MAPK signaling pathway. J Bone Miner Res 21, 1648-1656.
Lynch, H.T., Ferrara, K., Barlogie, B., Coleman, E.A., Lynch, J.F., Weisenburger, D.,
Sanger, W., Watson, P., Nipper, H., Witt, V., et al. (2008). Familial myeloma. N Engl J
Med 359, 152-157.

57
Maeda, K., Okubo, K., Shimomura, I., Funahashi, T., Matsuzawa, Y., and Matsubara, K.
(1996). cDNA cloning and expression of a novel adipose specific collagen-like factor,
apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun 221,
286-289.
Martinez (2007). Factores prognósticos no Mieloma Múltiplo. Revista brasileira
hematologia hemoterapia 1, 27-30.
Matsui, W., Huff, C.A., Wang, Q., Malehorn, M.T., Barber, J., Tanhehco, Y., Smith, B.D.,
Civin, C.I., and Jones, R.J. (2004). Characterization of clonogenic multiple myeloma cells.
Blood 103, 2332-2336.
Michalakis, K., Williams, C.J., Mitsiades, N., Blakeman, J., Balafouta-Tselenis, S.,
Giannopoulos, A., and Mantzoros, C.S. (2007). Serum adiponectin concentrations and
tissue expression of adiponectin receptors are reduced in patients with prostate cancer: a
case control study. Cancer Epidemiol Biomarkers Prev 16, 308-313.
Miller, M., Cho, J.Y., Pham, A., Ramsdell, J., and Broide, D.H. (2009). Adiponectin and
functional adiponectin receptor 1 are expressed by airway epithelial cells in chronic
obstructive pulmonary disease. J Immunol 182, 684-691.
Molica, S., Digiesi, G., Vacca, A., Mirabelli, R., Todoerti, K., Battaglia, C., Morabito, F.,
Neri, A., and Ribatti, D. (2009). Does adiponectin act as an antiangiogenic factor in B-cell
chronic lymphocytic leukemia? Adv Hematol 2009, 287974.
Nakano, Y., Tobe, T., Choi-Miura, N.H., Mazda, T., and Tomita, M. (1996). Isolation and
characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J
Biochem 120, 803-812.
Okamoto, Y., Kihara, S., Funahashi, T., Matsuzawa, Y., and Libby, P. (2006). Adiponectin:
a key adipocytokine in metabolic syndrome. Clin Sci (Lond) 110, 267-278.
Omede, P., Boccadoro, M., Fusaro, A., Gallone, G., and Pileri, A. (1993). Multiple
myeloma: 'early' plasma cell phenotype identifies patients with aggressive biological and
clinical characteristics. Br J Haematol 85, 504-513.

58
OMS (2006). http://who.int/mediacentre/factssheets/fs311/en/index.html. In Organização
Mundial de Saúde.
OMS (2008). http://who.int/features/qa/15/en/index.html. In Organização mundial de
saúde.
Onuma, M., Bub, J.D., Rummel, T.L., and Iwamoto, Y. (2003). Prostate cancer cell-
adipocyte interaction: leptin mediates androgen-independent prostate cancer cell
proliferation through c-Jun NH2-terminal kinase. J Biol Chem 278, 42660-42667.
Petridou, E., Mantzoros, C.S., Dessypris, N., Dikalioti, S.K., and Trichopoulos, D. (2006).
Adiponectin in relation to childhood myeloblastic leukaemia. Br J Cancer 94, 156-160.
Radbruch, A., Muehlinghaus, G., Luger, E.O., Inamine, A., Smith, K.G., Dorner, T., and
Hiepe, F. (2006). Competence and competition: the challenge of becoming a long-lived
plasma cell. Nat Rev Immunol 6, 741-750.
Rajala, M.W., and Scherer, P.E. (2003). Minireview: The adipocyte--at the crossroads of
energy homeostasis, inflammation, and atherosclerosis. Endocrinology 144, 3765-3773.
Rajkumar, S.V. (2005). MGUS and smoldering multiple myeloma: update on
pathogenesis, natural history, and management. Hematology Am Soc Hematol Educ
Program, 340-345.
Reseland, J.E., Reppe, S., Olstad, O.K., Hjorth-Hansen, H., Brenne, A.T., Syversen, U.,
Waage, A., and Iversen, P.O. (2009). Abnormal adipokine levels and leptin-induced
changes in gene expression profiles in multiple myeloma. Eur J Haematol 83, 460-470.
Robillard, N., Avet-Loiseau, H., Garand, R., Moreau, P., Pineau, D., Rapp, M.J.,
Harousseau, J.L., and Bataille, R. (2003). CD20 is associated with a small mature plasma
cell morphology and t(11;14) in multiple myeloma. Blood 102, 1070-1071.
Scherer, P.E., Williams, S., Fogliano, M., Baldini, G., and Lodish, H.F. (1995). A novel
serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 270,
26746-26749.

59
Silva, P.e. (2008). Eletroforese de proteínas séricas: interpretação e correlação clínica.
Revista Médica de Minas Gerais 18(12): 116-122.
Silva, P.e. (2009). Mieloma múltiplo: características clínicas e laboratoriais ao diagnóstico
e estudo prognóstico. Revista brasileira hematologia hemoterapia 2, 63-68.
Soliman, e.a. (2005). Association Between Adiponectin, Insulin Resistance, and
Endometrial Cancer. American Cancer Society, 2376-2381.
Spyridopoulos, T.N., Petridou, E.T., Skalkidou, A., Dessypris, N., Chrousos, G.P., and
Mantzoros, C.S. (2007). Low adiponectin levels are associated with renal cell carcinoma:
a case-control study. Int J Cancer 120, 1573-1578.
Teoh, G., and Anderson, K.C. (1997). Interaction of tumor and host cells with adhesion
and extracellular matrix molecules in the development of multiple myeloma. Hematol
Oncol Clin North Am 11, 27-42.
Tian, E., Zhan, F., Walker, R., Rasmussen, E., Ma, Y., Barlogie, B., and Shaughnessy,
J.D., Jr. (2003). The role of the Wnt-signaling antagonist DKK1 in the development of
osteolytic lesions in multiple myeloma. N Engl J Med 349, 2483-2494.
Tilg, H., and Moschen, A.R. (2006). Adipocytokines: mediators linking adipose tissue,
inflammation and immunity. Nat Rev Immunol 6, 772-783.
Tilg, H., and Moschen, A.R. (2008). Role of adiponectin and PBEF/visfatin as regulators of
inflammation: involvement in obesity-associated diseases. Clin Sci (Lond) 114, 275-288.
Turgeon, M.L. (1999). Clinical Hematology Theory and Procedures, L.W.a. Wilkins, ed.
(Boston), pp. 467.
Tworoger, S.S., Eliassen, A.H., Kelesidis, T., Colditz, G.A., Willett, W.C., Mantzoros, C.S.,
and Hankinson, S.E. (2007). Plasma adiponectin concentrations and risk of incident
breast cancer. J Clin Endocrinol Metab 92, 1510-1516.

60
Waki, H., and Tontonoz, P. (2007). Endocrine functions of adipose tissue. Annu Rev
Pathol 2, 31-56.
Waki, H., Yamauchi, T., Kamon, J., Kita, S., Ito, Y., Hada, Y., Uchida, S., Tsuchida, A.,
Takekawa, S., and Kadowaki, T. (2005). Generation of globular fragment of adiponectin
by leukocyte elastase secreted by monocytic cell line THP-1. Endocrinology 146, 790-796.
Wang, B., Jenkins, J.R., and Trayhurn, P. (2005a). Expression and secretion of
inflammation-related adipokines by human adipocytes differentiated in culture: integrated
response to TNF-alpha. Am J Physiol Endocrinol Metab 288, E731-740.
Wang, Y., Lam, K.S., Xu, J.Y., Lu, G., Xu, L.Y., Cooper, G.J., and Xu, A. (2005b).
Adiponectin inhibits cell proliferation by interacting with several growth factors in an
oligomerization-dependent manner. J Biol Chem 280, 18341-18347.
Wei, E.K., Giovannucci, E., Fuchs, C.S., Willett, W.C., and Mantzoros, C.S. (2005). Low
plasma adiponectin levels and risk of colorectal cancer in men: a prospective study. J Natl
Cancer Inst 97, 1688-1694.
Weiss, B.M., Abadie, J., Verma, P., Howard, R.S., and Kuehl, W.M. (2009). A monoclonal
gammopathy precedes multiple myeloma in most patients. Blood 113, 5418-5422.
Xu, H., Barnes, G.T., Yang, Q., Tan, G., Yang, D., Chou, C.J., Sole, J., Nichols, A., Ross,
J.S., Tartaglia, L.A., et al. (2003). Chronic inflammation in fat plays a crucial role in the
development of obesity-related insulin resistance. J Clin Invest 112, 1821-1830.
Yamauchi, T., Kamon, J., Ito, Y., Tsuchida, A., Yokomizo, T., Kita, S., Sugiyama, T.,
Miyagishi, M., Hara, K., Tsunoda, M., et al. (2003). Cloning of adiponectin receptors that
mediate antidiabetic metabolic effects. Nature 423, 762-769.
Yokota, T., Meka, C.S., Medina, K.L., Igarashi, H., Comp, P.C., Takahashi, M., Nishida,
M., Oritani, K., Miyagawa, J., Funahashi, T., et al. (2002). Paracrine regulation of fat cell
formation in bone marrow cultures via adiponectin and prostaglandins. J Clin Invest 109,
1303-1310.

61
Yokota, T., Oritani, K., Takahashi, I., Ishikawa, J., Matsuyama, A., Ouchi, N., Kihara, S.,
Funahashi, T., Tenner, A.J., Tomiyama, Y., et al. (2000). Adiponectin, a new member of
the family of soluble defense collagens, negatively regulates the growth of
myelomonocytic progenitors and the functions of macrophages. Blood 96, 1723-1732.
Zaidi, A.A., and Vesole, D.H. (2001). Multiple myeloma: an old disease with new hope for
the future. CA Cancer J Clin 51, 273-285; quiz 286-279.











![Considera um triângulo [ABC], rectângulo em B e cujos ... · PDF filequadrado do maior, obtenho sempre um número que não é múltiplo de dois.” ... de o cubo escolhido ter só](https://static.fdocument.org/doc/165x107/5a76a81b7f8b9aa3688d5e78/considera-um-triangulo-abc-rectangulo-em-b-e-cujos-quadrado-do-maior.jpg)
