
Aντιγραφή Επιδιόρθωση Και Ανασυνδυασμός Του Dna
48
• Aντιγραφή, επιδιόρθωση και ανασυνδυασμός του DNA • DNA Replication, Repair, and Recombination 1
description
dna replication recombinant Dna
Transcript of Aντιγραφή Επιδιόρθωση Και Ανασυνδυασμός Του Dna

• Aντιγραφή, επιδιόρθωση και ανασυνδυασμός του DNA
• DNA Replication, Repair, and Recombination
1
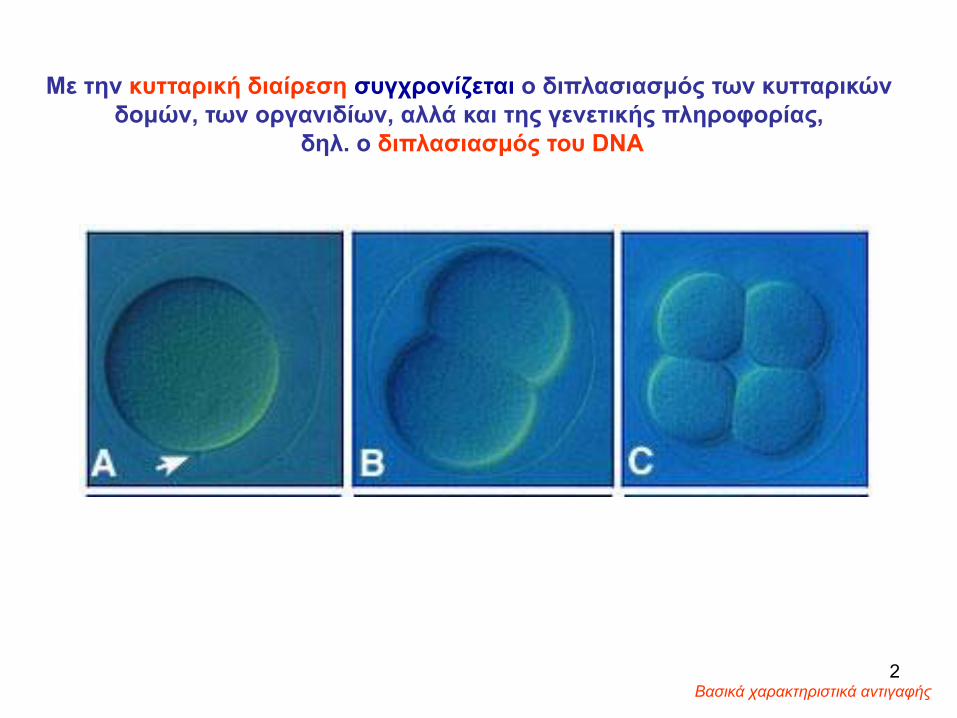
Με την κυτταρική διαίρεση συγχρονίζεται ο διπλασιασμός των κυτταρικών δομών, των οργανιδίων, αλλά και της γενετικής πληροφορίας,
δηλ. ο διπλασιασμός του DNA
Βασικά χαρακτηριστικά αντιγαφής 2

Αντιγραφή του DNADNA Replication
To DNA αντιγράφει επακριβώς τον εαυτό του. Τυχόν λάθη κατά την αντιγραφή στο DNA = Μετάλλαξη(χρωμοσωμική / γενετική μετάλλαξη)
1. Η δυνατότητα ακριβούς αντιγραφής στηρίζεται στο
αυστηρό συμπληρωματικό ζευγάρωμα των βάσεων
(base pairing) C – G, A – T. Ο κάθε κλώνος
χρησιμεύει ως εκμαγείο για τη σύνθεση ενός νέου
συμπληρωματικού κλώνου
Α. Γενικά χαρακτηριστικά της αντιγραφής του DNA
Η δομή του DNA υπέδειξε
το πιθανό μηχανισμό
αυτό-διπλασιασμού του
J. Watson και F. Crick, 19532. ημισυντηρητική 3. DNA pol
3
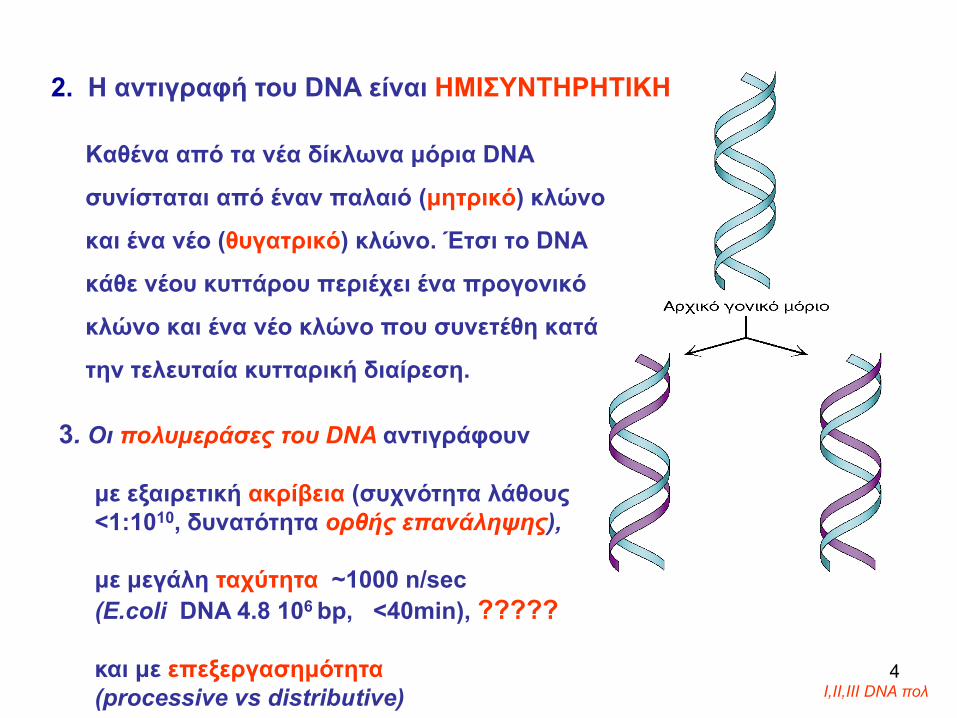
Καθένα από τα νέα δίκλωνα μόρια DNA
συνίσταται από έναν παλαιό (μητρικό) κλώνο
και ένα νέο (θυγατρικό) κλώνο. Έτσι το DNA
κάθε νέου κυττάρου περιέχει ένα προγονικό
κλώνο και ένα νέο κλώνο που συνετέθη κατά
την τελευταία κυτταρική διαίρεση.
2. Η αντιγραφή του DNA είναι ΗΜΙΣΥΝΤΗΡΗΤΙΚΗ
3. Οι πολυμεράσες του DNA αντιγράφουν
με εξαιρετική ακρίβεια (συχνότητα λάθους <1:1010, δυνατότητα ορθής επανάληψης),
με μεγάλη ταχύτητα ~1000 n/sec(E.coli DNA 4.8 106 bp, <40min), ?????
και με επεξεργασημότητα (processive vs distributive) Ι,ΙΙ,ΙΙΙ DNA πολ
4

DNA πολυμεράσες στα βακτήρια
DNA polymerase I IIIII
5’ 3’
3’ 5’
5’ 3’ polymerase
exonuclease(proof reading)
exonuclease
+ + +
+ + +
+ - -
mol/ cell 400 10-20 Why ??
Γονίδιο pol C*pol A pol B
Αριθμός υπομονάδων >1071
Νουκλεοτ. /sec
Επεξεργασημότητα
250-100040 20
>500.00015003-200
Η αντίδραση5
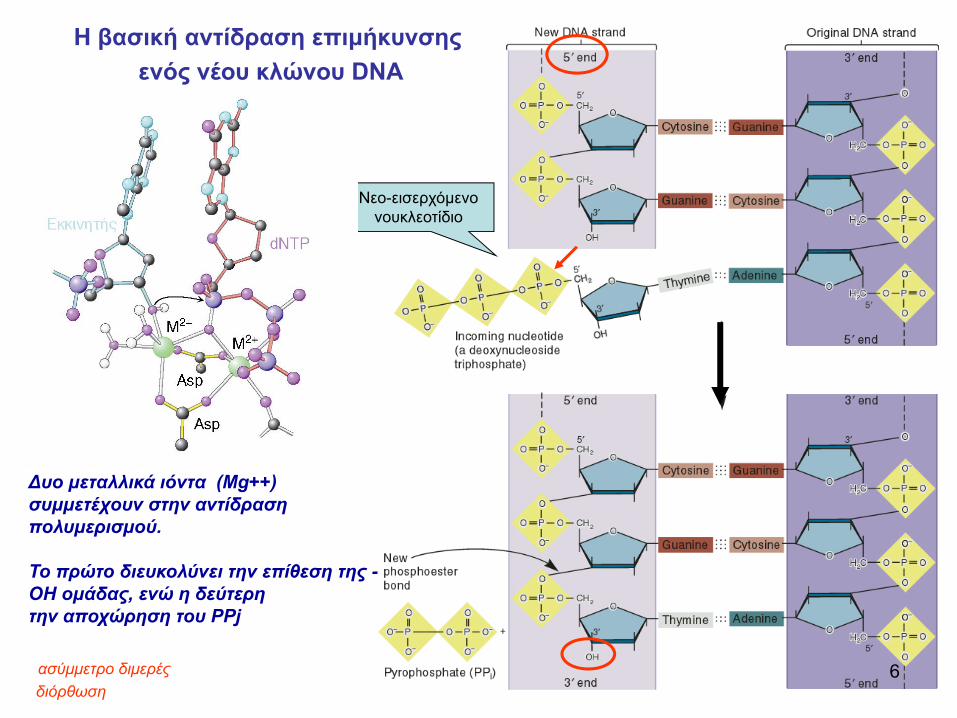
Νεο-εισερχόμενο νουκλεοτίδιο
H βασική αντίδραση επιμήκυνσηςενός νέου κλώνου DNA
διόρθωση
Δυο μεταλλικά ιόντα (Mg++) συμμετέχουν στην αντίδρασηπολυμερισμού.
Το πρώτο διευκολύνει την επίθεση της -ΟΗ ομάδας, ενώ η δεύτερητην αποχώρηση του PPj
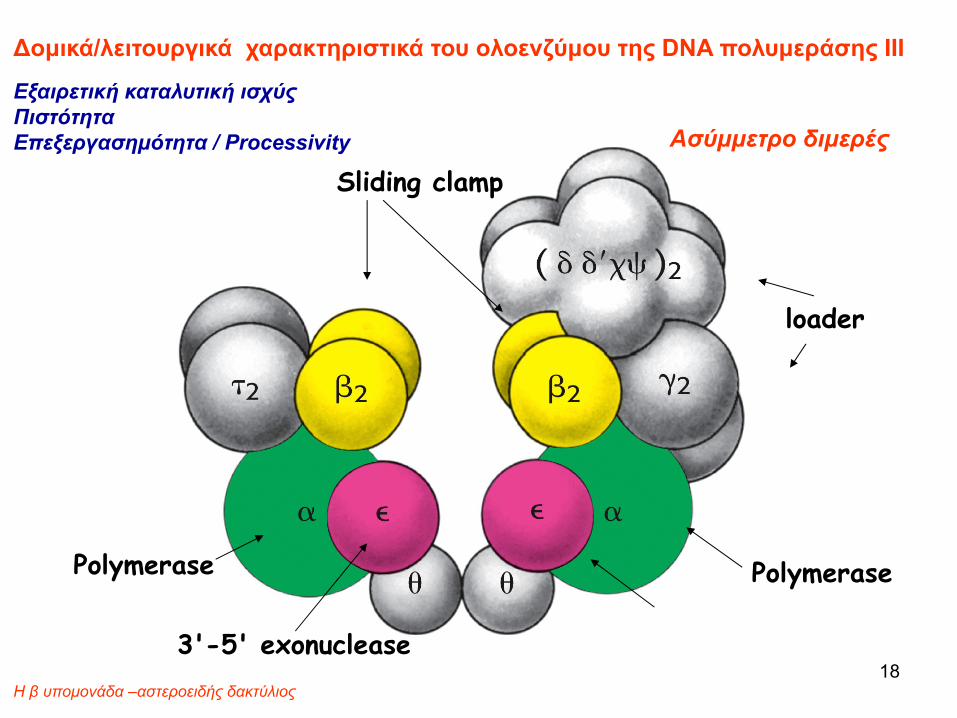
6ασύμμετρο διμερές
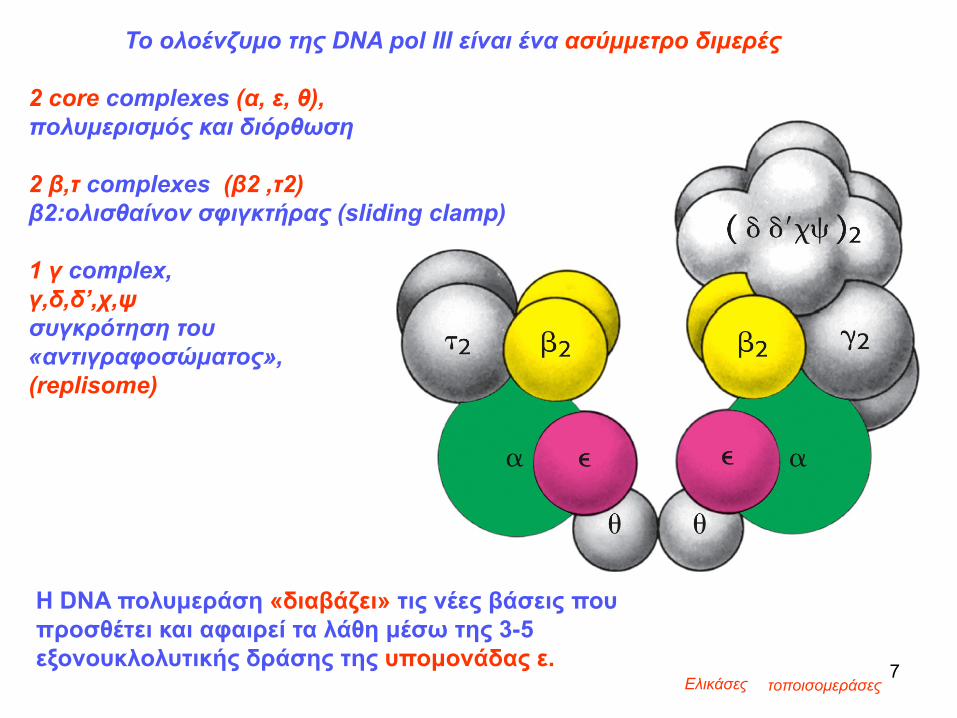
Η DNA πολυμεράση «διαβάζει» τις νέες βάσεις που προσθέτει και αφαιρεί τα λάθη μέσω της 3-5 εξονουκλολυτικής δράσης της υπομονάδας ε.
Ελικάσες τοποισομεράσες7
Το ολοένζυμο της DNA pol III είναι ένα ασύμμετρο διμερές
2 core complexes (α, ε, θ), πολυμερισμός και διόρθωση
2 β,τ complexes (β2 ,τ2) β2:ολισθαίνον σφιγκτήρας (sliding clamp)
1 γ complex, γ,δ,δ’,χ,ψσυγκρότηση του «αντιγραφοσώματος», (replisome)
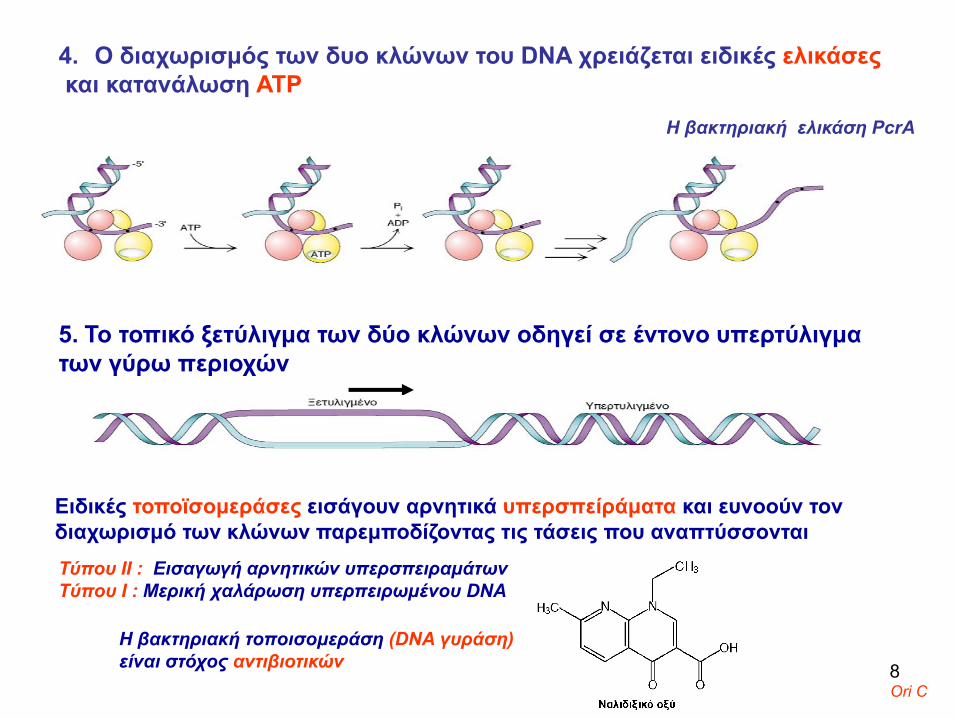
4. Ο διαχωρισμός των δυο κλώνων του DNA χρειάζεται ειδικές ελικάσεςκαι κατανάλωση ΑΤΡ
H βακτηριακή ελικάση PcrA
5. Το τοπικό ξετύλιγμα των δύο κλώνων οδηγεί σε έντονο υπερτύλιγμα των γύρω περιοχών
Ειδικές τοποϊσομεράσες εισάγουν αρνητικά υπερσπείράματα και ευνοούν τον διαχωρισμό των κλώνων παρεμποδίζοντας τις τάσεις που αναπτύσσονται Τύπου ΙΙ : Εισαγωγή αρνητικών υπερσπειραμάτωνTύπου Ι : Μερική χαλάρωση υπερπειρωμένου DNA
Η βακτηριακή τοποισομεράση (DNA γυράση)είναι στόχος αντιβιοτικών
Ori C8

6. Η αντιγραφή του DNA ξεκινά από ένα σημείο και εξελίσσεται προοδευτικά προς τις δυο κατευθύνσεις,
Διχάλα E. coli genome9
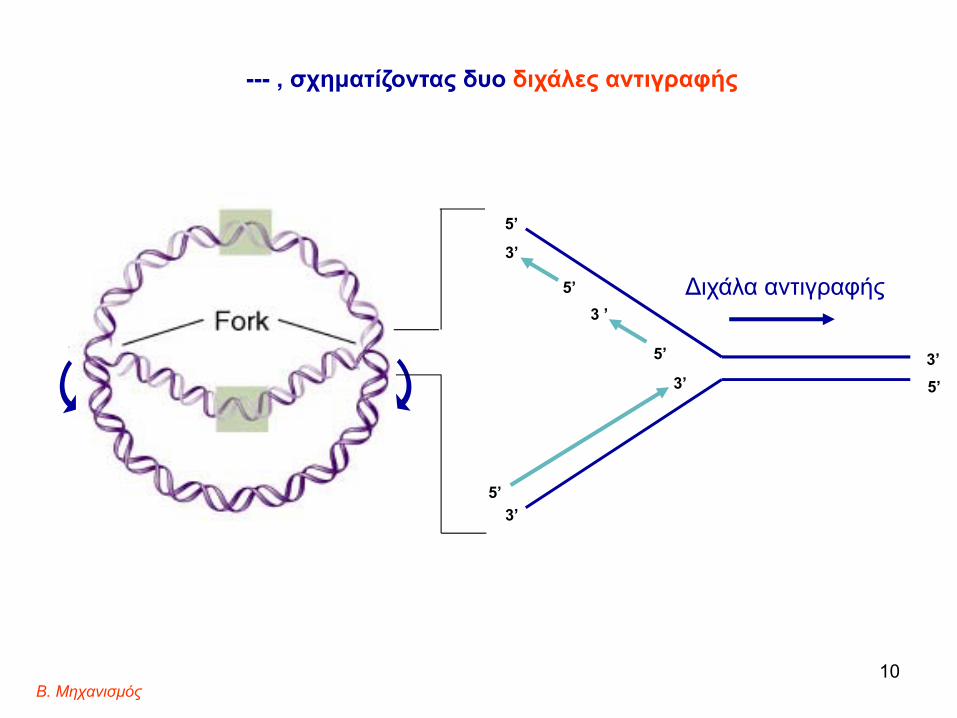
--- , σχηματίζοντας δυο διχάλες αντιγραφής
5’
3 ’
3’
3’
5’3’
5’
3’
5’
5’ Διχάλα αντιγραφής
Β. Μηχανισμός 10

Β. Μηχανισμός της Αντιγραφής του DNA1. Επιλογή του σημείου έναρξης της αντιγραφής στην Ε. Coli
Η έναρξη της αντιγραφής πραγματοποιείται στο oriC (origin of Chromosomal replication). To oriC περιέχει μια ΑΤ περιοχή και 4 επαναλαμβανόμενες αλληλουχίες δέσμευσης της
πρωτείνης DnaA
DnaA-B- SSB11

2. Διαχωρισμός των δυο κλώνων και σχηματισμός συμπλόκου προ-εκκίνησης.
DnaA, DnaB (Ελικάση) και SSB (single strand binding) στις διχάλες αντιγραφής
Διαχωρισμός των δύο κλώνων (Α/Τ)
Δέσμευση τηςελικάσης DnaB
Πρόσδεση της DnaA στο oriC
DnaB
Ο διαχωρισμός των δύο κλώνων επεκτείνεται προς τις δυο κατευθύνσεις σχηματίζοντας 2 διχάλες αντιγραφής
SSB
SSB SSB
SSB
Δέσμευση της SSBστο μονόκλωνο DNA
Σύμπλοκο προ-εκκίνησης
Εναρξη επιμήκυνση primase
12
5’
5’
5’
3’
3’
3’
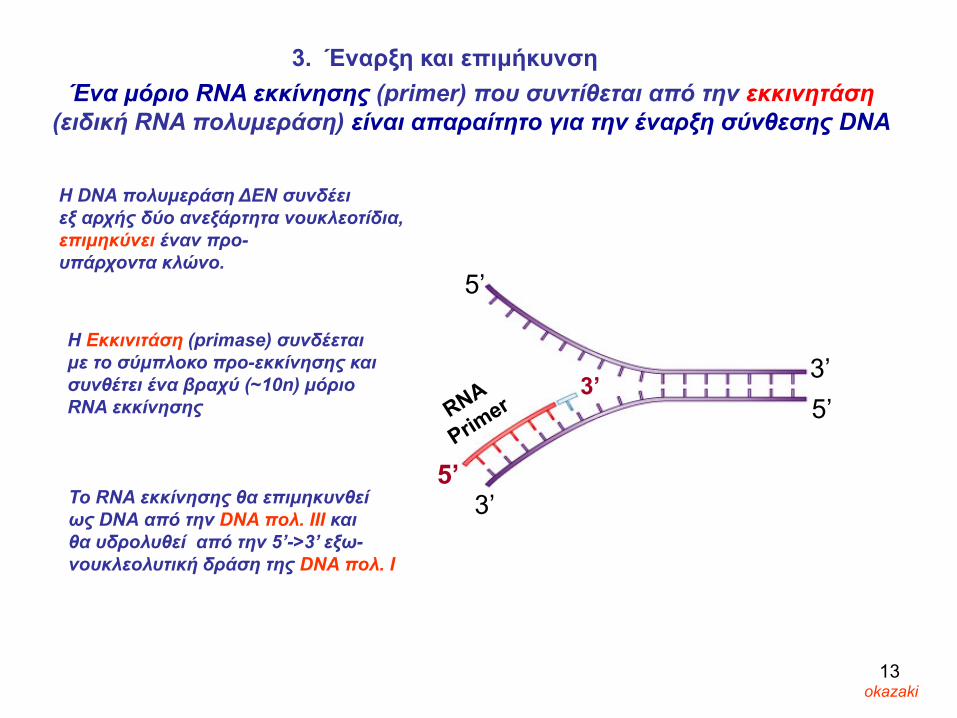
Ένα μόριο RNA εκκίνησης (primer) που συντίθεται από την εκκινητάση(ειδική RNA πολυμεράση) είναι απαραίτητο για την έναρξη σύνθεσης DNA
3. Έναρξη και επιμήκυνση
H DNA πολυμεράση ΔΕΝ συνδέει εξ αρχής δύο ανεξάρτητα νουκλεοτίδια, επιμηκύνει έναν προ-υπάρχοντα κλώνο.
To RNA εκκίνησης θα επιμηκυνθεί ως DNA από την DNA πολ. ΙΙΙ και θα υδρολυθεί από την 5’->3’ εξω-νουκλεολυτική δράση της DNA πολ. Ι
Η Εκκινιτάση (primase) συνδέεται με το σύμπλοκο προ-εκκίνησης και συνθέτει ένα βραχύ (~10n) μόριοRNA εκκίνησης
okazaki13

Ο ένας κλώνος του DNA συντίθεται συνεχώς (προπορευόμενος, leading strand), ενώ ο άλλος (καθυστερών / υπολειπόμενος, lagging strand)
συντίθεται κατά τμήματα (τμήματα Okazaki)
ligase
Μεταξύ των τμημάτων Okazakiαποκαθίστανται οι φωσφοδιεστερικοί δεσμοί από την DNA λιγάση, αφού τo RNA εκκίνησης υδρολυθεί από την 5’->3’ εξω-νουκλεολυτική δράση της DNA πολ. Ι(Nick translation)
5’
3’
3’
5’
3’
5’
3’
5’
Okazaki
5’
3’
3’
5’3’
5’
3’
5’
Διχάλα αντιγραφής
Η DNA polσυνθέτει DNAπάντα με κατεύθυνση 5’ -> 3’
14

Η λιγάση του DNA (ligase) συνδέει το 3’-ΟΗ άκρο ενός μορίου DNA με το 5’–ΡΟ3 άκρο ενός δεύτερου μορίου σε δίκλωνες περιοχές, π.χ στα
τμήματα Okazaki
Μηχανισμός 15

Μηχανισμός αντίδρασης λιγάσης του DNA
1. Ligase (E) + ATP→ PPi +
--OH +DNA-3' O P
OO
O-DNA-3’- 5'-DNA + AMP-OH3.
2. E-AMP + P-5’-DNA→ + AMP--DNA
Ασύμμετρη αλλά συντονισμένη
φωσφαμιδικός δεσμός
HN
Lys
-AMP
P
O
O
O-
5'-DNAO
AMP--DNA
PO
O
O
OH
OΑ
OH
Ligase (E)
E
E
PO
O
O
OH
OΑ
OH
5-5 φωσφοδιεστειρικός δεσμός
P
O
O
O-
5'-DNAOP
O
O
O
OH
OΑ
OH
-AMPE
AMP--DNA
16
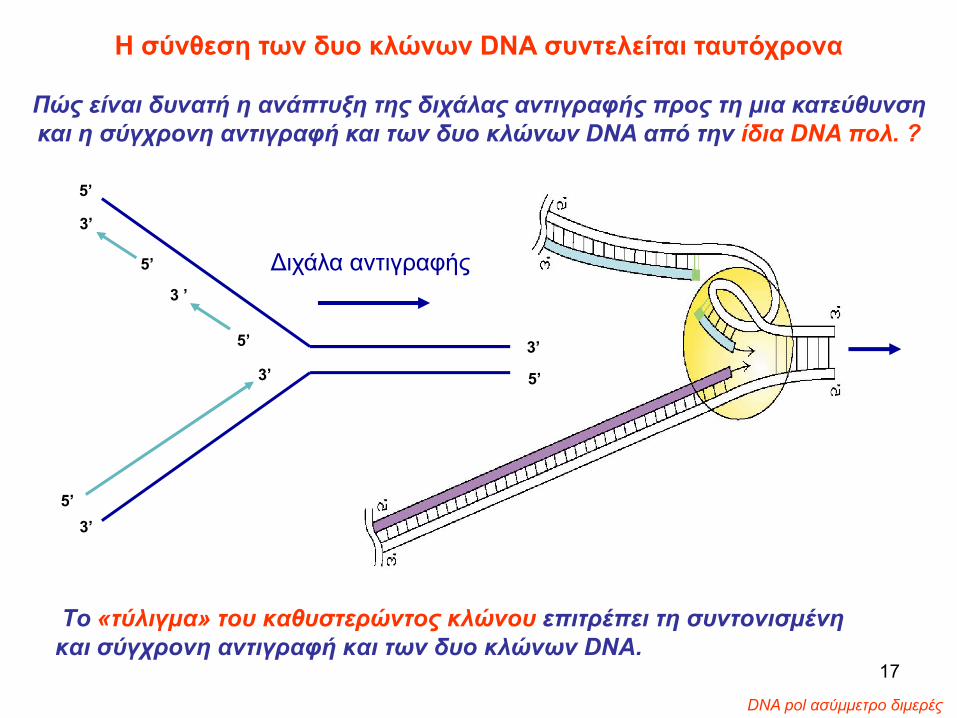
Το «τύλιγμα» του καθυστερώντος κλώνου επιτρέπει τη συντονισμένη και σύγχρονη αντιγραφή και των δυο κλώνων DNA.
5’
3 ’
3’
3’
5’
3’
5’
3’
5’
5’ Διχάλα αντιγραφής
DNA pol ασύμμετρο διμερές
Η σύνθεση των δυο κλώνων DNA συντελείται ταυτόχρονα
Πώς είναι δυνατή η ανάπτυξη της διχάλας αντιγραφής προς τη μια κατεύθυνση και η σύγχρονη αντιγραφή και των δυο κλώνων DNA από την ίδια DNA πολ. ?
17
Polymerase Polymerase
loader
Sliding clamp
3'-5' exonuclease
Δομικά/λειτουργικά χαρακτηριστικά του ολοενζύμου της DNA πολυμεράσης III
Εξαιρετική καταλυτική ισχύςΠιστότητα Επεξεργασημότητα / Processivity Ασύμμετρο διμερές
Η β υπομονάδα –αστεροειδής δακτύλιος 18
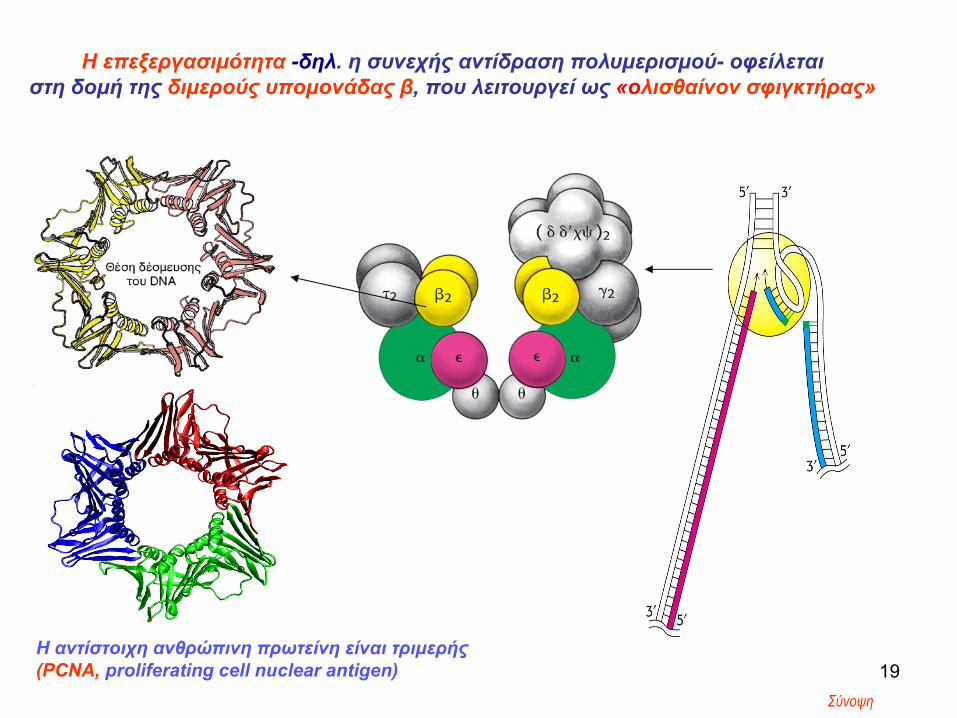
Η επεξεργασιμότητα -δηλ. η συνεχής αντίδραση πολυμερισμού- οφείλεται στη δομή της διμερούς υπομονάδας β, που λειτουργεί ως «ολισθαίνον σφιγκτήρας»
Σύνοψη
19Η αντίστοιχη ανθρώπινη πρωτείνη είναι τριμερής(PCNA, proliferating cell nuclear antigen)
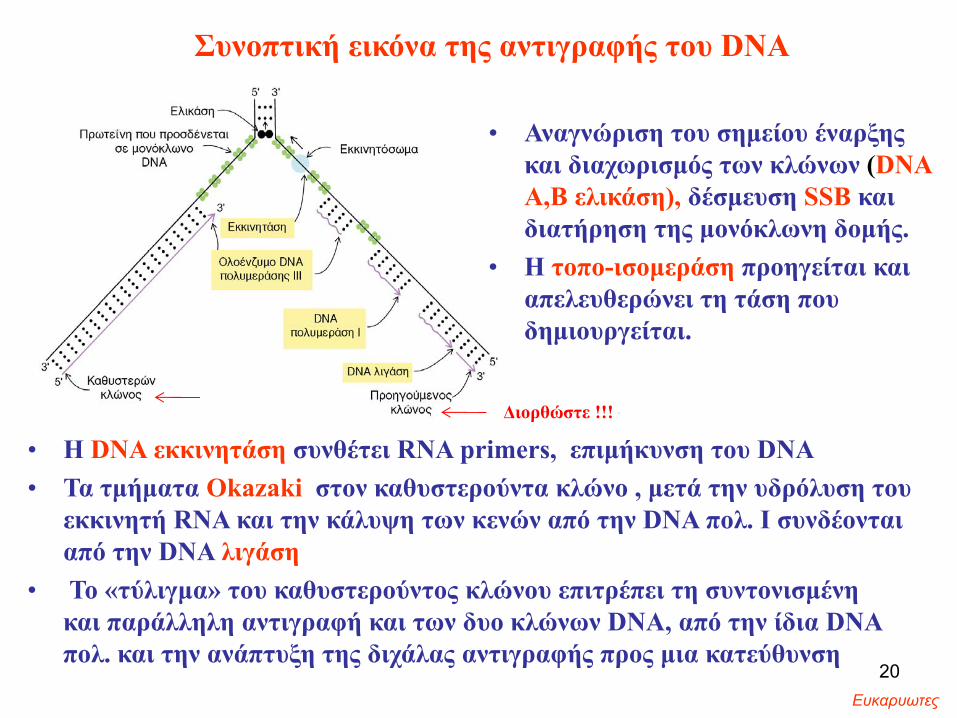
• Η DNA εκκινητάση συνθέτει RNA primers, επιμήκυνση του DNA • Τα τμήματα Okazaki στον καθυστερούντα κλώνο , μετά την υδρόλυση του
εκκινητή RNA και την κάλυψη των κενών από την DNA πολ. Ι συνδέονται από την DNA λιγάση
• Το «τύλιγμα» του καθυστερούντος κλώνου επιτρέπει τη συντονισμένη και παράλληλη αντιγραφή και των δυο κλώνων DNA, από την ίδια DNA πολ. και την ανάπτυξη της διχάλας αντιγραφής προς μια κατεύθυνση
Συνοπτική εικόνα της αντιγραφής του DNA
• Αναγνώριση του σημείου έναρξης και διαχωρισμός των κλώνων (DNAΑ,Β ελικάση), δέσμευση SSB και διατήρηση της μονόκλωνη δομής.
• Η τοπο-ισομεράση προηγείται και απελευθερώνει τη τάση που δημιουργείται.
Διορθώστε !!!
Ευκαρυωτες
20
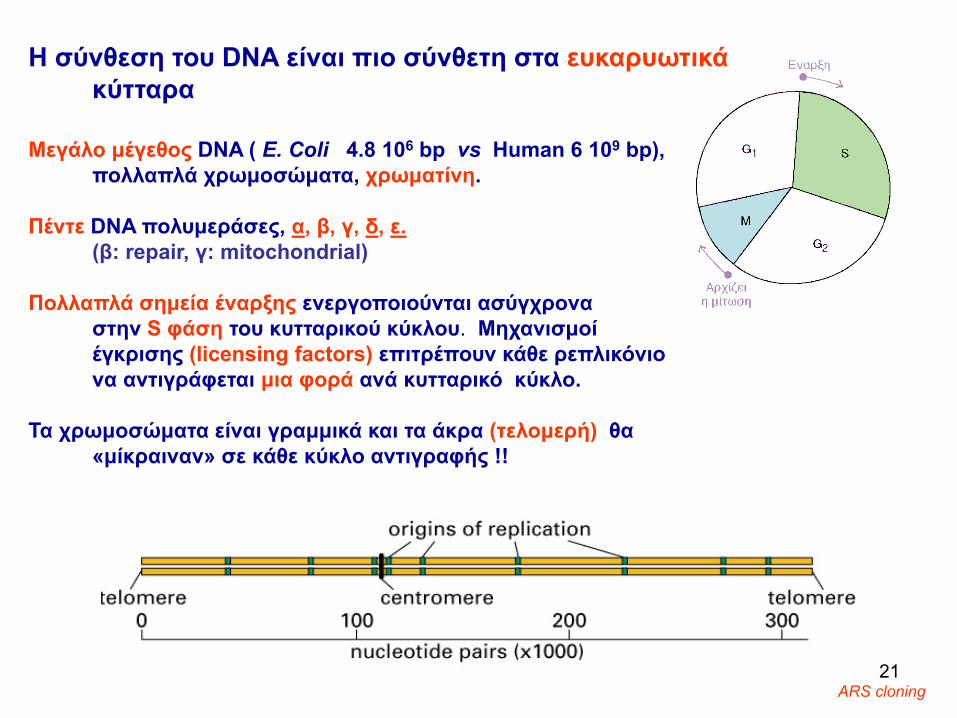
Η σύνθεση του DNA είναι πιο σύνθετη στα ευκαρυωτικάκύτταρα
Μεγάλο μέγεθος DNA ( E. Coli 4.8 106 bp vs Human 6 109 bp), πολλαπλά χρωμοσώματα, χρωματίνη.
Πέντε DNA πολυμεράσες, α, β, γ, δ, ε.(β: repair, γ: mitochondrial)
Πολλαπλά σημεία έναρξης ενεργοποιούνται ασύγχρονα στην S φάση του κυτταρικού κύκλου. Μηχανισμοί έγκρισης (licensing factors) επιτρέπουν κάθε ρεπλικόνιο να αντιγράφεται μια φορά ανά κυτταρικό κύκλο.
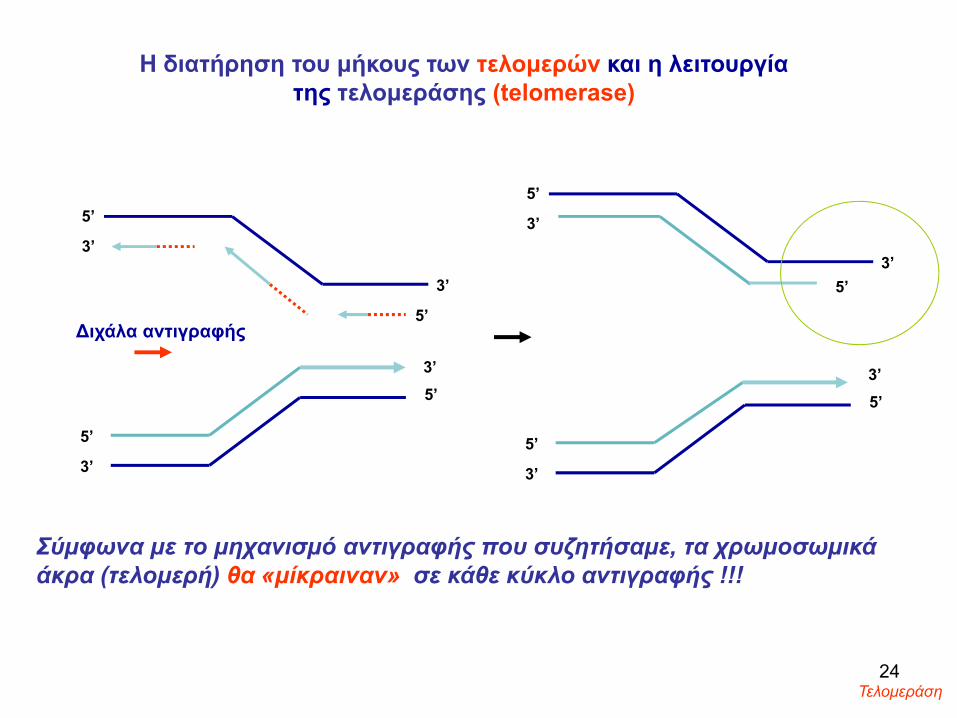
Τα χρωμοσώματα είναι γραμμικά και τα άκρα (τελομερή) θα «μίκραιναν» σε κάθε κύκλο αντιγραφής !!
ARS cloning21
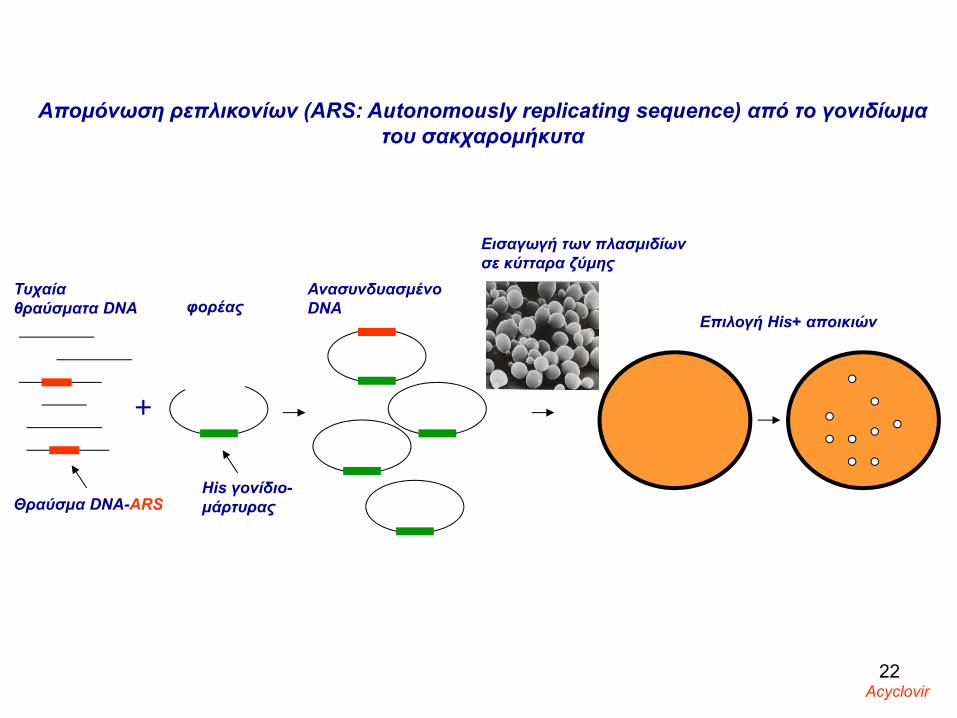
Απομόνωση ρεπλικονίων (ARS: Autonomously replicating sequence) από το γονιδίωμα του σακχαρομήκυτα
Acyclovir
+
Τυχαία θραύσματα DNA
Θραύσμα DNA-ARS
φορέας
His γονίδιο-μάρτυρας
Eισαγωγή των πλασμιδίων σε κύτταρα ζύμης
Επιλογή His+ αποικιών
Ανασυνδυασμένο DNA
22
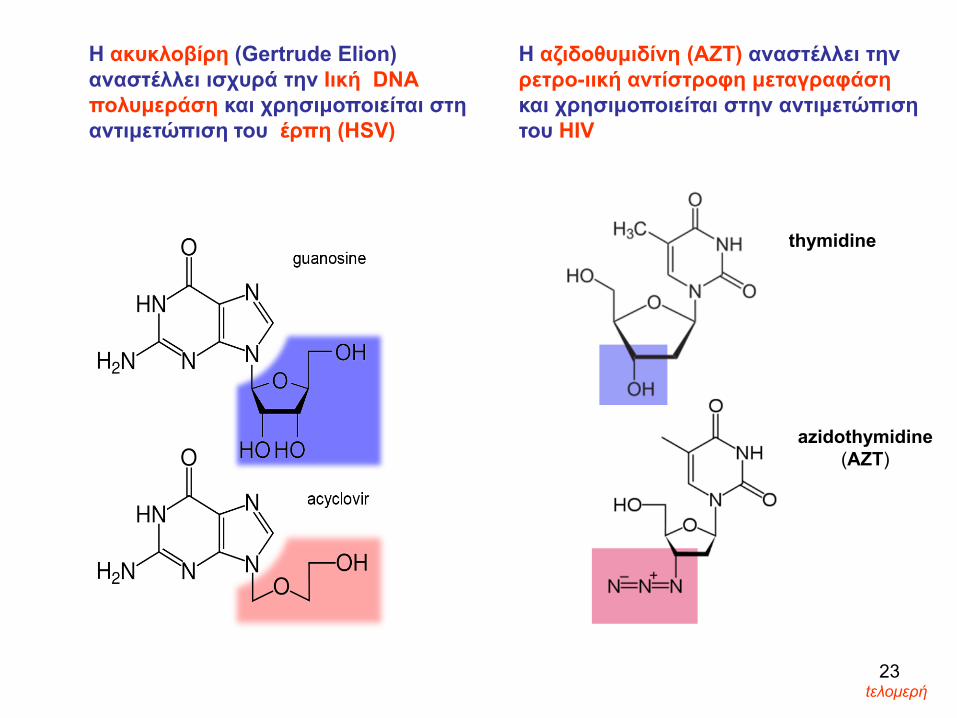
Η ακυκλοβίρη (Gertrude Elion)αναστέλλει ισχυρά την Ιική DNAπολυμεράση και χρησιμοποιείται στη αντιμετώπιση του έρπη (HSV)
tελομερή
azidothymidine(AZT)
thymidine
Η αζιδοθυμιδίνη (ΑΖΤ) αναστέλλει τηνρετρο-ιική αντίστροφη μεταγραφάση και χρησιμοποιείται στην αντιμετώπιση του HIV
23
Η διατήρηση του μήκους των τελομερών και η λειτουργία της τελομεράσης (telomerase)
3’
5’3’
5’
Διχάλα αντιγραφής
3’
5’
5’
3’
3’
5’3’
5’
3’
5’
5’3’
Σύμφωνα με το μηχανισμό αντιγραφής που συζητήσαμε, τα χρωμοσωμικά άκρα (τελομερή) θα «μίκραιναν» σε κάθε κύκλο αντιγραφής !!!
Τελομεράση 24
3’
5’
5’3’
5’
3’
3’
5’
3’
3’
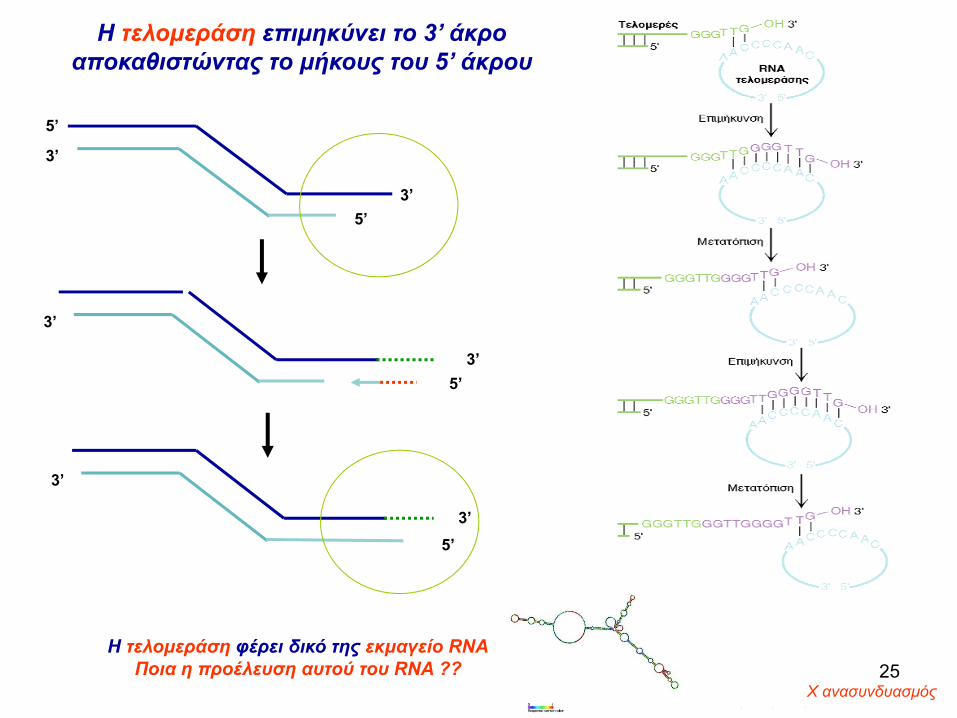
H τελομεράση επιμηκύνει το 3’ άκρο αποκαθιστώντας το μήκους του 5’ άκρου
H τελομεράση φέρει δικό της εκμαγείο RNAΠοια η προέλευση αυτού του RNA ??
Χ ανασυνδυασμός25

Ανασυνδυασμός του DNA (DNA Recombination)H αμοιβαία ανταλλαγή αλληλουχιών μεταξύ δύο μορίων DNA
Oμόλογος ανασυνδυασμός
• Ο ομόλογος ανασυνδυασμός συντελείται κατά τη διάρκεια της σύναψης στη μείωση, του γενικού ανασυνδυασμού στα βακτήρια και κατά την ανταλλαγή γενετικού υλικού στους ιούς
• Προϋποθέτει ομολογία DNA μεταξύ των δυο γονικών μορίων• Ο μοριακός μηχανισμός προτάθηκε από τους Holliday and
Whitehouse (1964).
26
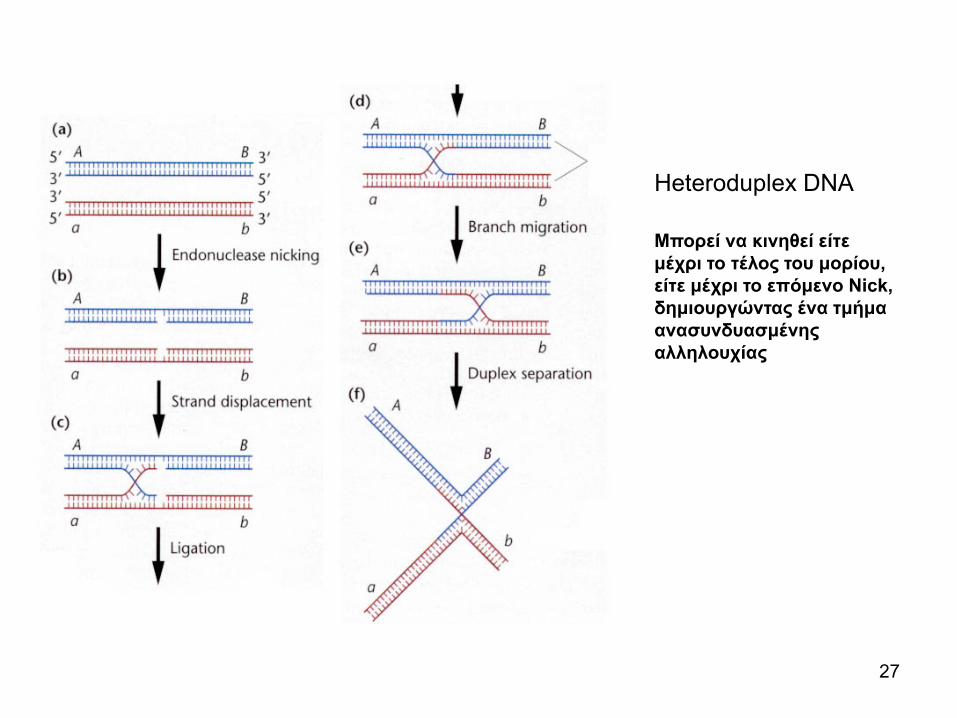
Heteroduplex DNA
Mπορεί να κινηθεί είτε μέχρι το τέλος του μορίου, είτε μέχρι το επόμενο Nick, δημιουργώντας ένα τμήμα ανασυνδυασμένης αλληλουχίας
27
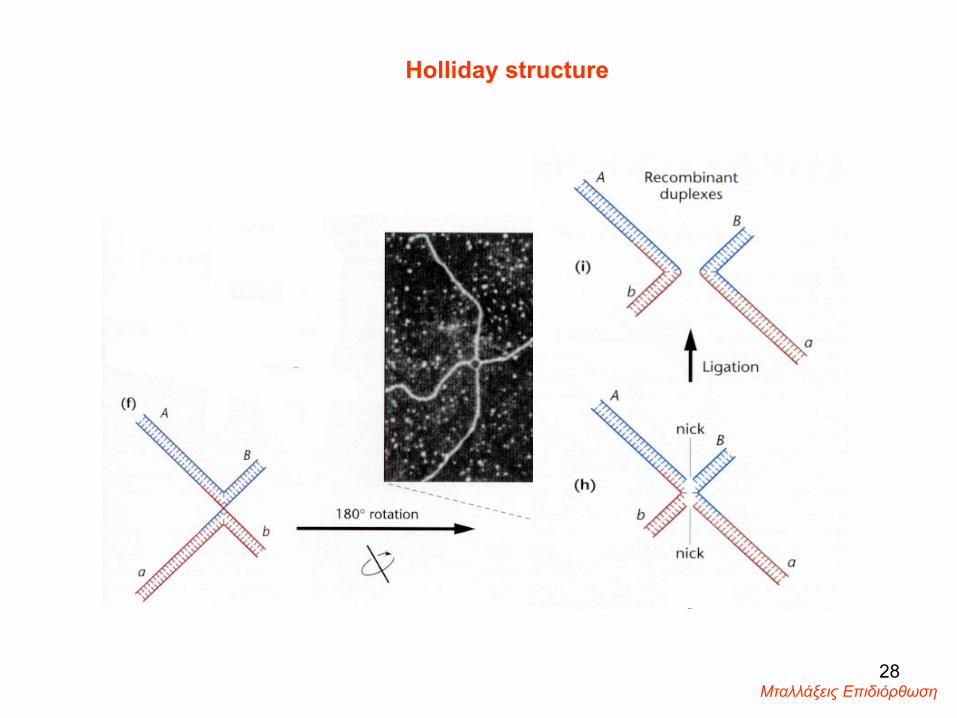
Holliday structure
Μταλλάξεις Επιδιόρθωση 28

Μεταλλάξεις
1. Απάλειψη (deletion) ενός ή περισσοτέρων ζευγών βάσεων
2. Εισδοχή (insertion) ενός ή περισσοτέρων ζευγών βάσεων
3. Αντικατάσταση (substitution) ενός ζεύγους βάσεων από ένα άλλο :Μετάπτωση (transition) Pu->Pu, Py->PyΜεταστροφή (transversion) Pu->Py, Py->Pu
HNO2
Επιδιόρθωση του DNA`Η διεργασία απομάκρυνσης των αταίριαστων βάσεων που:
1. Διαφεύγουν (αν και σπάνια) από τους μηχανισμούς «ορθής επανάληψης» της αντιγραφής του DNA
2. Συμβαίνουν φυσιολογικά ως αυθόρμητες χημικές μεταβολές
3. Επάγονται από περιβαλλοντικούς μεταλλαξογόνους παράγοντες (Χημικά, UV, X-rays)
Oi μεταλλάξεις βραχυπρόθεσμα δεν είναι επιθυμητές !!!
29
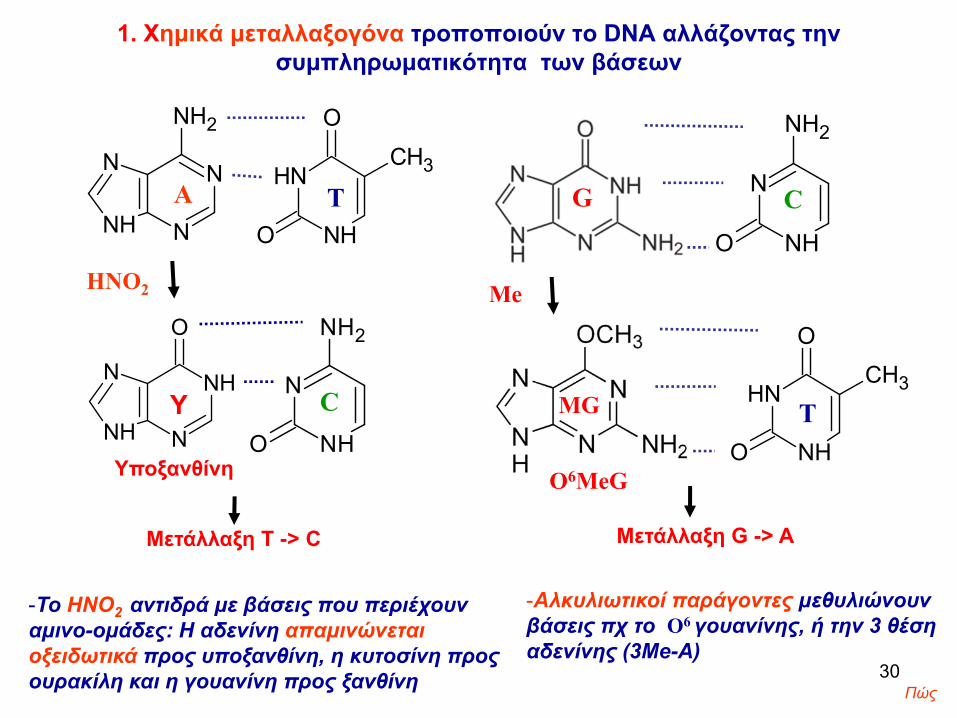
1. Xημικά μεταλλαξογόνα τροποποιούν το DNA αλλάζοντας την συμπληρωματικότητα των βάσεων
N
NH
NH2
ON
NH
NH
N
O
N
N
NH
N
NH2
Α N
NH
NH2
O
C
ΗΝΟ2
C
Υποξανθίνη
Μετάλλαξη Τ -> C
Πώς
-To ΗΝΟ2 αντιδρά με βάσεις που περιέχουν αμινο-ομάδες: H αδενίνη απαμινώνεται οξειδωτικά προς υποξανθίνη, η κυτοσίνη προς ουρακίλη και η γουανίνη προς ξανθίνη
NH
NH
O
O
CH3
Τ
Ο6ΜeG
Μετάλλαξη G -> A
NH
NH
O
O
CH3
Τ
Υ ΜG
G
Μe
-Αλκυλιωτικοί παράγοντες μεθυλιώνουν βάσεις πχ το Ο6 γουανίνης, ή την 3 θέση αδενίνης (3Me-A)
30
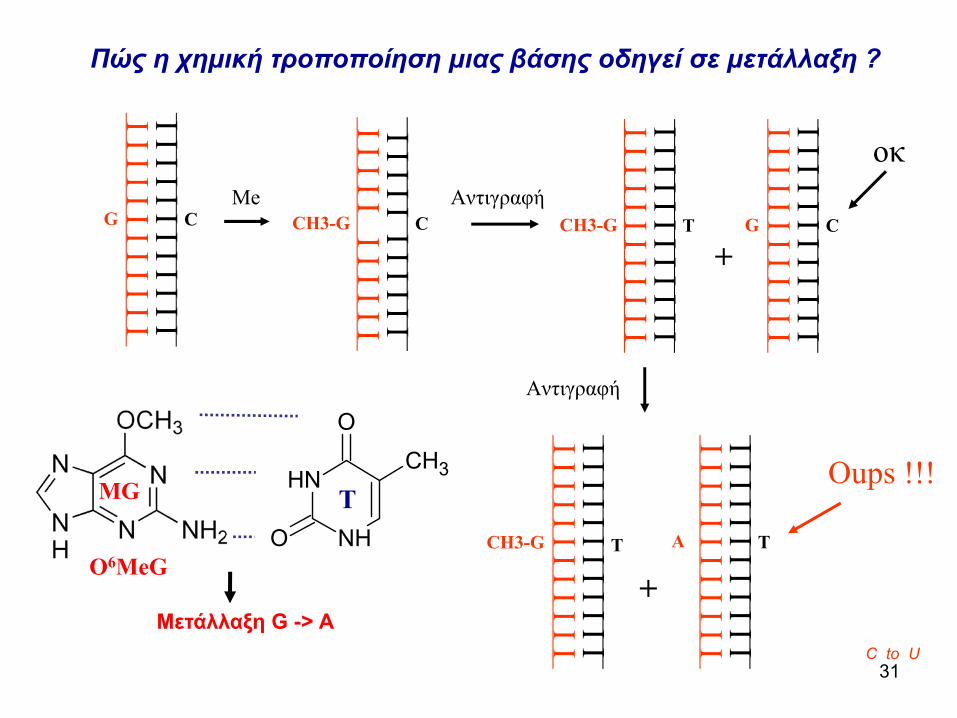
Πώς η χημική τροποποίηση μιας βάσης οδηγεί σε μετάλλαξη ?
I I I I I I I I I I I II I I
I I I
I I I
I I I
CG
I I I I I I I I I I II I I
I I I
I I I
I I
CCH3-GMe Aντιγραφή
I I I I I I I I I I I II I I
I I I
I I I
I I I
ΤCH3-G
I I I I I I I I I I I II I I
I I I
I I I
I I I
CG
I I I I I I I I I I I II I I
I I I
I I I
I I I
ΤCH3-G
I I I I I I I I I I I II I I
I I I
I I I
I I I
ΤΑ
Aντιγραφή
C to U
οκ
Oups !!!
+
+
NH
NH
O
O
CH3
Τ
Ο6ΜeG
Μετάλλαξη G -> A
ΜG
31
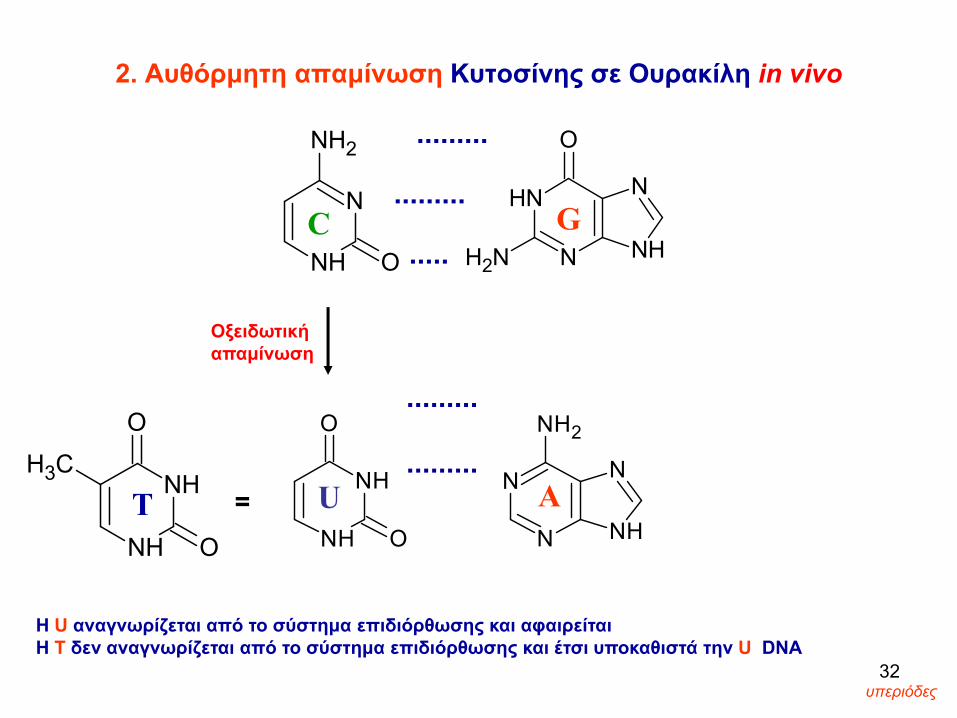
2. Αυθόρμητη απαμίνωση Κυτοσίνης σε Ουρακίλη in vivo
N
NH
NH2
O N
NH
NH
N
NH2
O
NH
NH
O
O N
N
NH
N
NH2
C G
U Α
Oξειδωτική απαμίνωση
NH
NH
O
O
CH3
Τ =
υπεριόδες
Η U αναγνωρίζεται από το σύστημα επιδιόρθωσης και αφαιρείται Η Τ δεν αναγνωρίζεται από το σύστημα επιδιόρθωσης και έτσι υποκαθιστά την U DNA
32
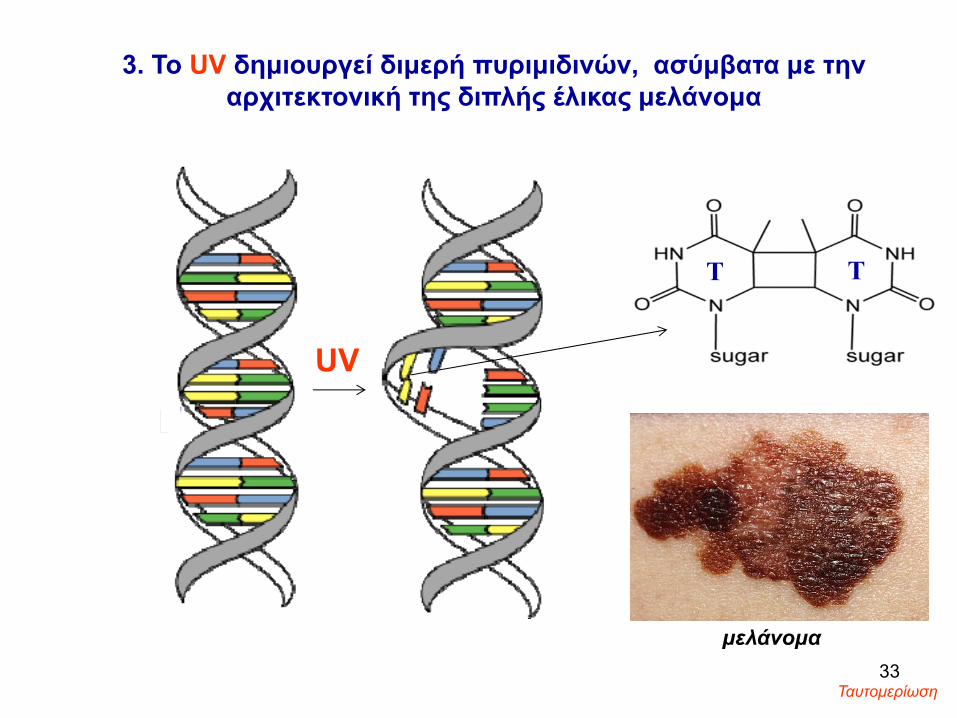
3. Το UV δημιουργεί διμερή πυριμιδινών, ασύμβατα με την αρχιτεκτονική της διπλής έλικας μελάνομα
Ταυτομερίωση
UV
Τ Τ
μελάνομα33
NH
NH
O
O
Br
NH
N
OH
O
Br
N
NH
NH
N
NH2
O
N
N
NH
N
NH2
ΑBrU
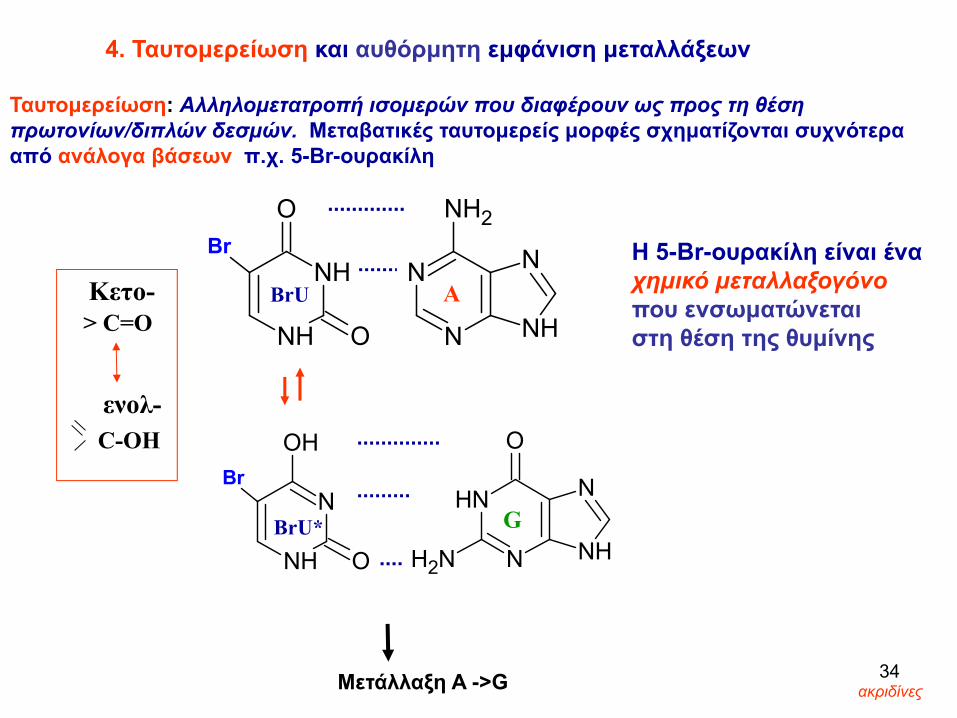
H 5-Br-ουρακίλη είναι ένα χημικό μεταλλαξογόνο που ενσωματώνεται στη θέση της θυμίνης
BrU* G
ακριδίνεςΜετάλλαξη A ->G
C-OH
> C=OΚετο-
ενολ-
34
4. Ταυτομερείωση και αυθόρμητη εμφάνιση μεταλλάξεων
Ταυτομερείωση: Aλληλομετατροπή ισομερών που διαφέρουν ως προς τη θέση πρωτονίων/διπλών δεσμών. Μεταβατικές ταυτομερείς μορφές σχηματίζονται συχνότερα από ανάλογα βάσεων π.χ. 5-Br-ουρακίλη
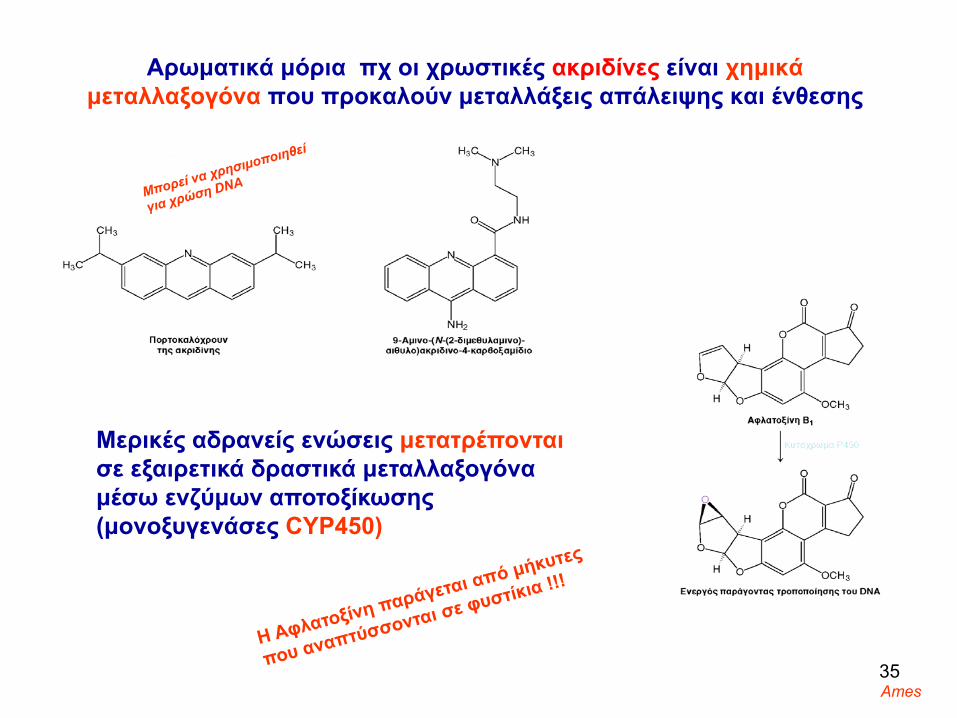
Aρωματικά μόρια πχ οι χρωστικές ακριδίνες είναι χημικά μεταλλαξογόνα που προκαλούν μεταλλάξεις απάλειψης και ένθεσης
35
Μερικές αδρανείς ενώσεις μετατρέπονταισε εξαιρετικά δραστικά μεταλλαξογόναμέσω ενζύμων αποτοξίκωσης (μονοξυγενάσες CYΡ450)
Ames

Η δοκιμασία Ames
Η μεταλλαξογόνος δράση ενός χημικού παράγοντα έλέγχεται με βάση τη συχνότητα πρόκλησης μετάλλάξεων σε βακτήρια, πχ η ανάστροφη μετάλλαξη που επανακαθιστά τα βακτήρια πρωτότροφα για ιστιδίνη
His- His+
Σε ορισμένες περιπτώσεις, η μεταλλαξογόνος δράση ανιχνεύεται μόνο με προ-επώαση του χημικού παράγοντα με ηπατικά εκχυλίσματα
His-
His+
επιδιόρθωση36
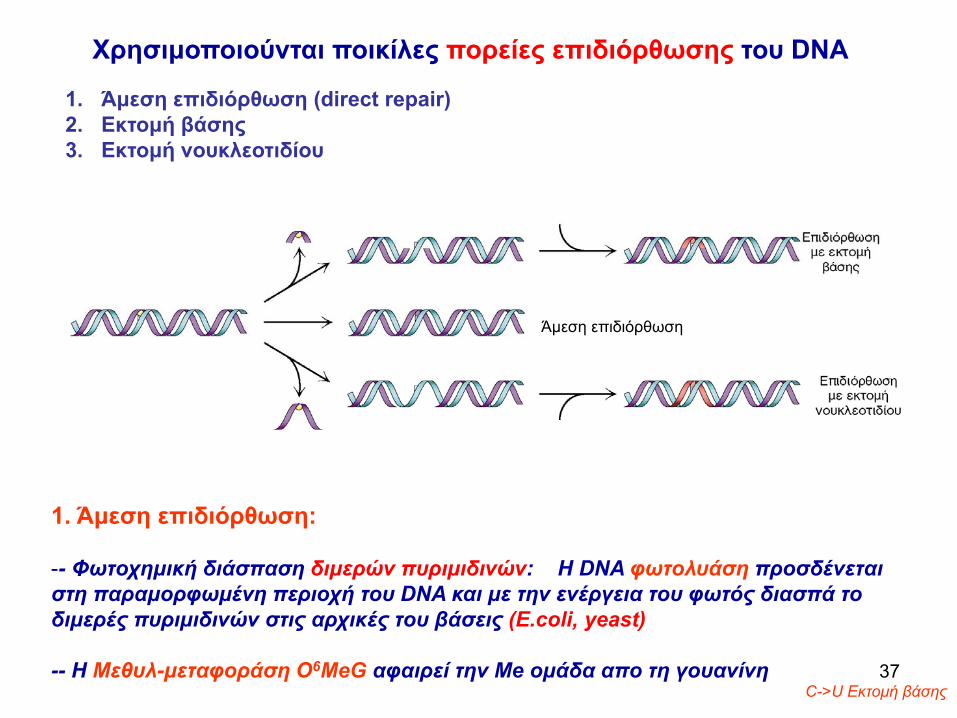
Χρησιμοποιούνται ποικίλες πορείες επιδιόρθωσης του DNA
Άμεση επιδιόρθωση
1. Άμεση επιδιόρθωση:
-- Φωτοχημική διάσπαση διμερών πυριμιδινών: Η DNA φωτολυάση προσδένεται στη παραμορφωμένη περιοχή του DNA και με την ενέργεια του φωτός διασπά το διμερές πυριμιδινών στις αρχικές του βάσεις (E.coli, yeast)
-- Η Μεθυλ-μεταφοράση Ο6MeG αφαιρεί την Μe oμάδα απο τη γουανίνηC->U Εκτομή βάσης
1. Άμεση επιδιόρθωση (direct repair)2. Εκτομή βάσης 3. Εκτομή νουκλεοτιδίου
37
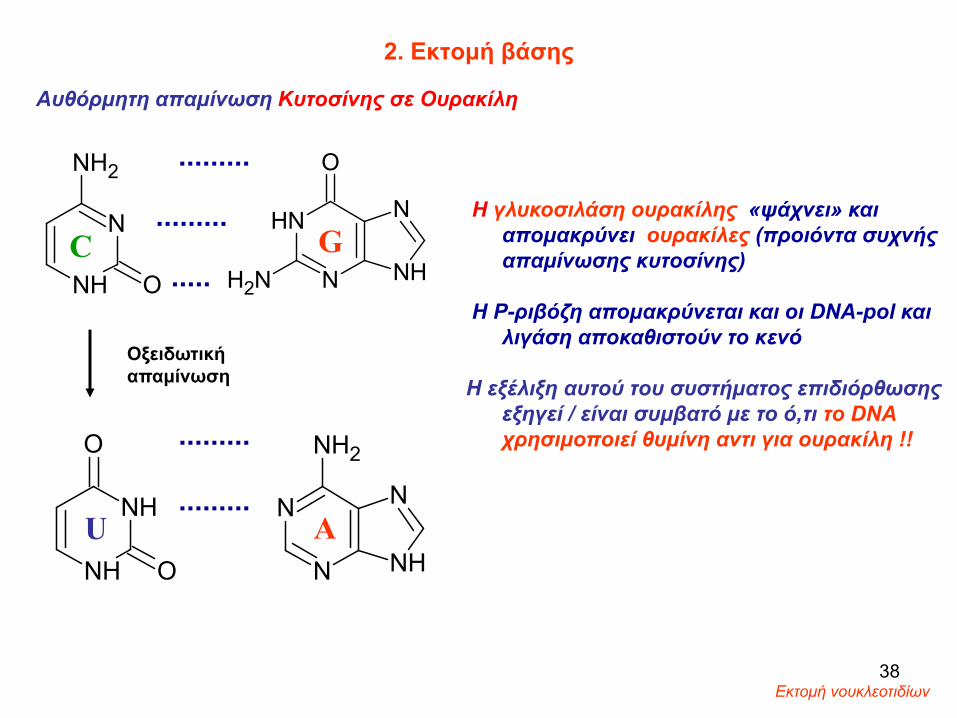
2. Εκτομή βάσης
N
NH
NH2
O N
NH
NH
N
NH2
O
NH
NH
O
O N
N
NH
N
NH2
C G
U Α
Oξειδωτική απαμίνωση
Η γλυκοσιλάση ουρακίλης «ψάχνει» και απομακρύνει ουρακίλες (προιόντα συχνής απαμίνωσης κυτοσίνης)
Η Ρ-ριβόζη απομακρύνεται και οι DNA-pol και λιγάση αποκαθιστούν το κενό
Η εξέλιξη αυτού του συστήματος επιδιόρθωσηςεξηγεί / είναι συμβατό με το ό,τι το DNA χρησιμοποιεί θυμίνη αντι για ουρακίλη !!
Αυθόρμητη απαμίνωση Κυτοσίνης σε Ουρακίλη
Εκτομή νουκλεοτιδίων38
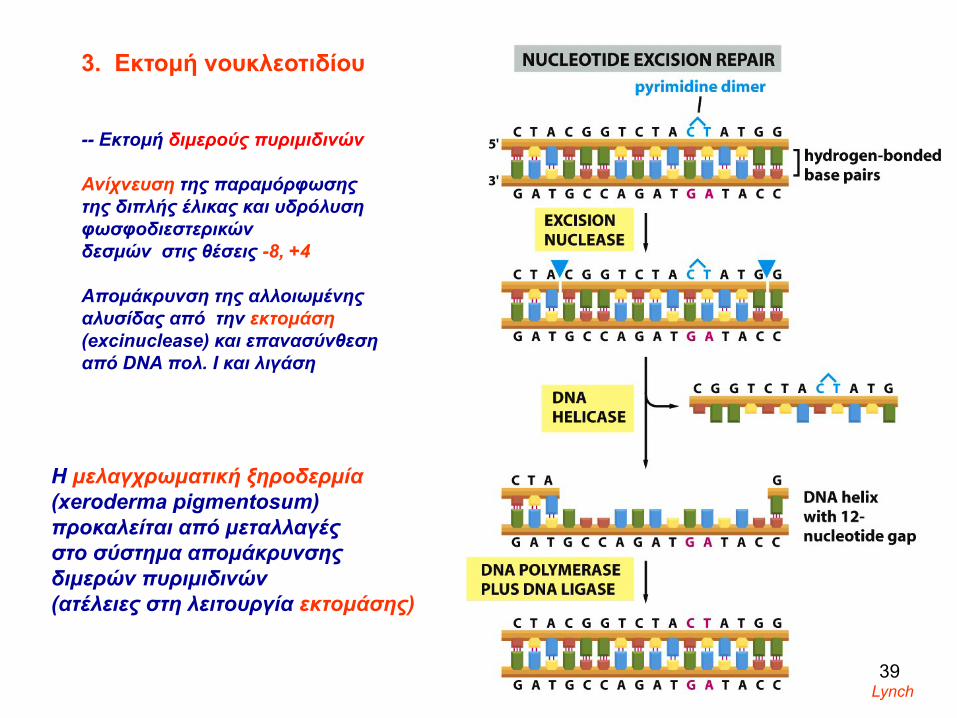
3. Εκτομή νουκλεοτιδίου
-- Εκτομή διμερούς πυριμιδινών
Ανίχνευση της παραμόρφωσης της διπλής έλικας και υδρόλυση φωσφοδιεστερικών δεσμών στις θέσεις -8, +4
Απομάκρυνση της αλλοιωμένης αλυσίδας από την εκτομάση(excinuclease) και επανασύνθεση από DNA πολ. Ι και λιγάση
Lynch
Η μελαγχρωματική ξηροδερμία(xeroderma pigmentosum)προκαλείται από μεταλλαγές στο σύστημα απομάκρυνσης διμερών πυριμιδινών (ατέλειες στη λειτουργία εκτομάσης)
39
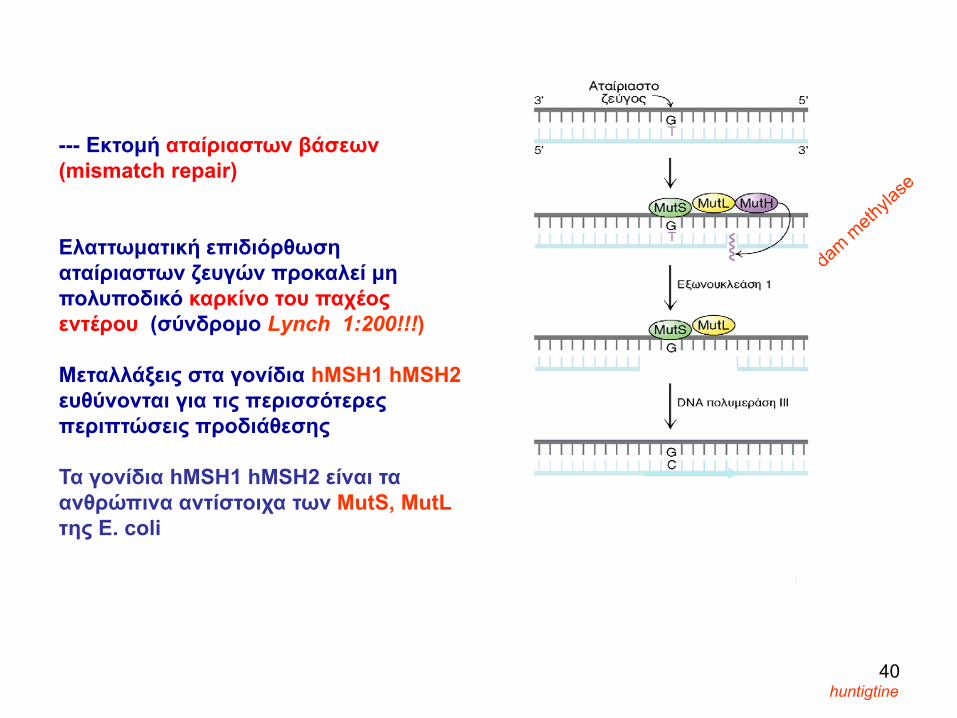
--- Εκτομή αταίριαστων βάσεων (mismatch repair)
Ελαττωματική επιδιόρθωση αταίριαστων ζευγών προκαλεί μη πολυποδικό καρκίνο του παχέος εντέρου (σύνδρομο Lynch 1:200!!!)
Mεταλλάξεις στα γονίδια hMSH1 hMSH2ευθύνονται για τις περισσότερες περιπτώσεις προδιάθεσης
Τα γονίδια hMSH1 hMSH2 είναι τα ανθρώπινα αντίστοιχα των MutS, MutL της E. coli
huntigtine40
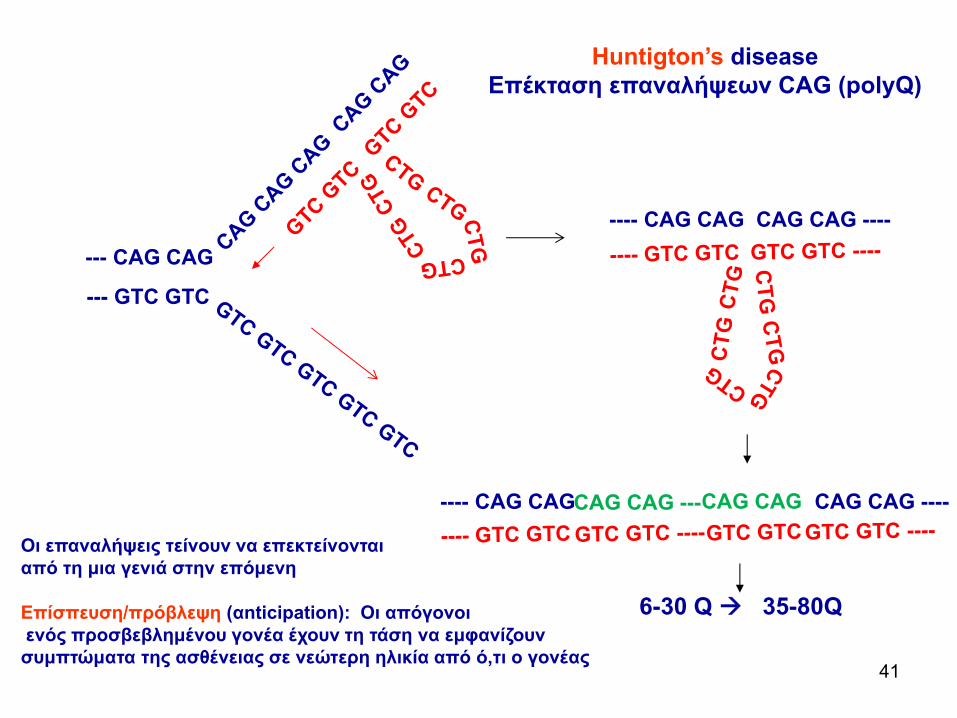
Huntigton’s disease Επέκταση επαναλήψεων CAG (polyQ)
Οι επαναλήψεις τείνουν να επεκτείνονται από τη μια γενιά στην επόμενη
Επίσπευση/πρόβλεψη (αnticipation): Οι απόγονοιενός προσβεβλημένου γονέα έχουν τη τάση να εμφανίζουν συμπτώματα της ασθένειας σε νεώτερη ηλικία από ό,τι ο γονέας
--- CAG CAG
--- GTC GTC
---- CAG CAG CAG CAG ----
---- CAG CAG CAG CAG ----CAG CAGCAG CAG ---
6-30 Q 35-80Q
41

42
Ασκήσεις
Stryer Σελ. 876-8771, 3, 6, 7, 11, 15.
Lehninger Kεφ. 262, 5, 6, 9.
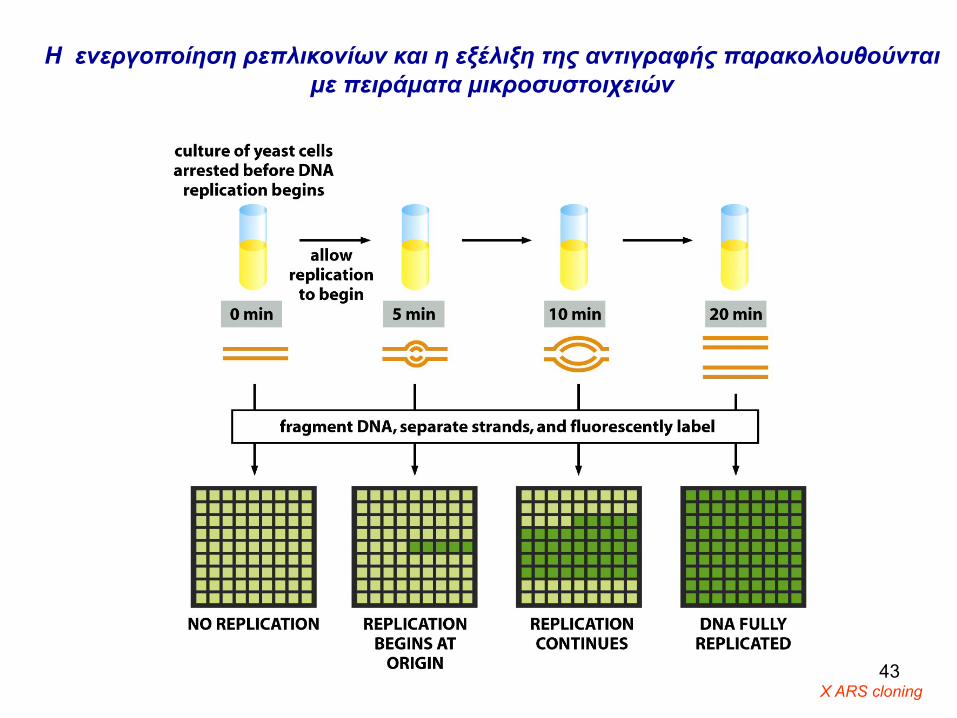
Η ενεργοποίηση ρεπλικονίων και η εξέλιξη της αντιγραφής παρακολουθούνται με πειράματα μικροσυστοιχειών
Χ ARS cloning43
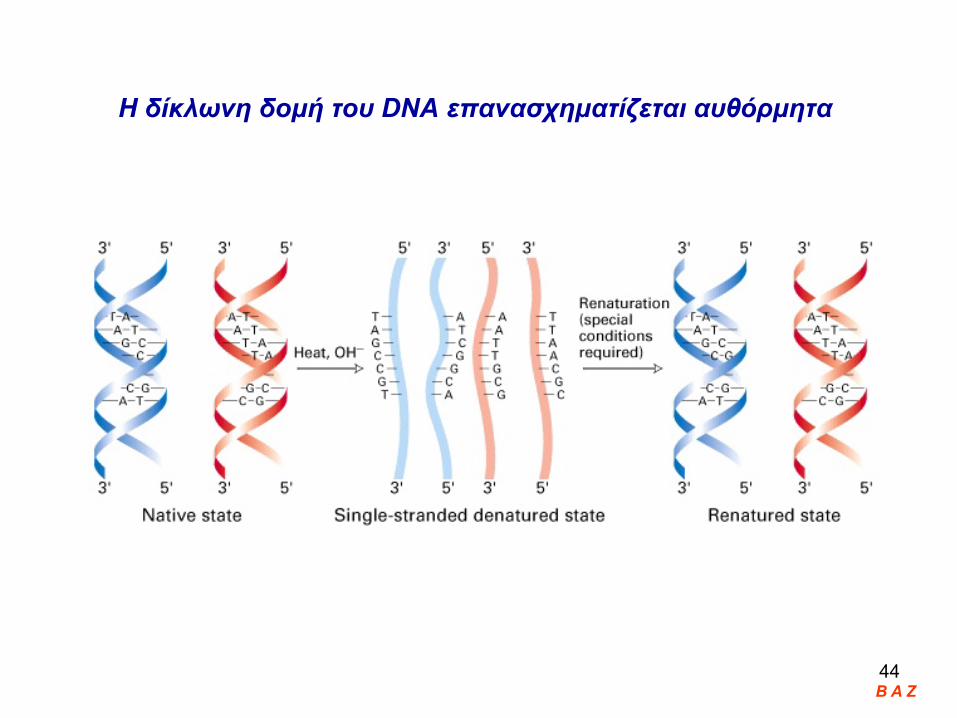
Η δίκλωνη δομή του DNA επανασχηματίζεται αυθόρμητα
Β Α Ζ 44
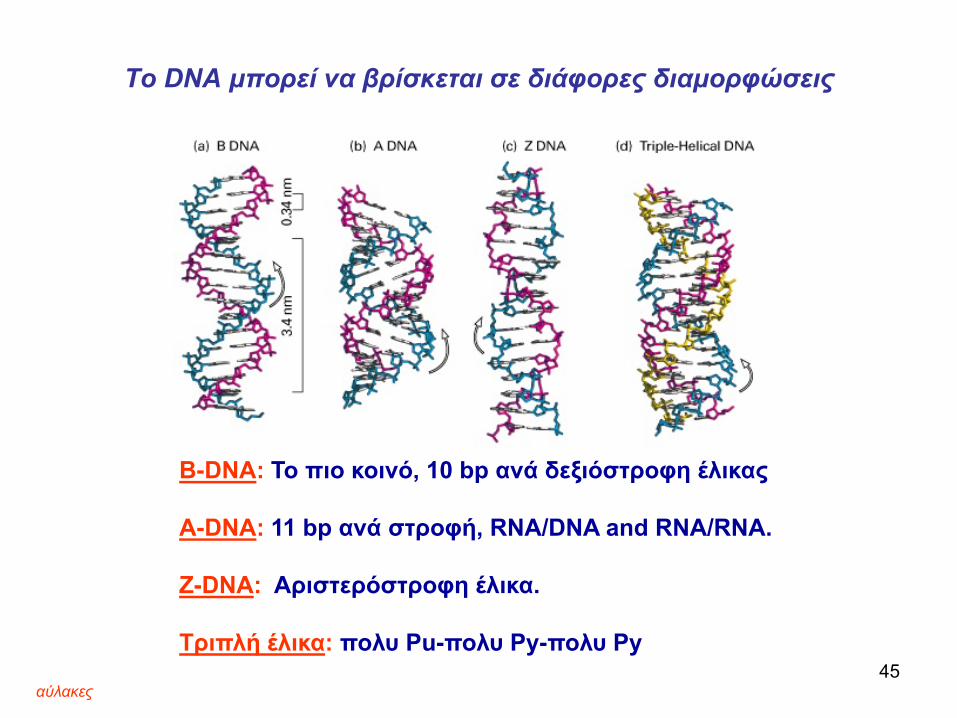
Το DNA μπορεί να βρίσκεται σε διάφορες διαμορφώσεις
B-DNA: Το πιο κοινό, 10 bp ανά δεξιόστροφη έλικας
A-DNA: 11 bp ανά στροφή, RNA/DNA and RNA/RNA.
Z-DNA: Αριστερόστροφη έλικα.
Τριπλή έλικα: πολυ Pu-πολυ Py-πολυ Py
αύλακες45
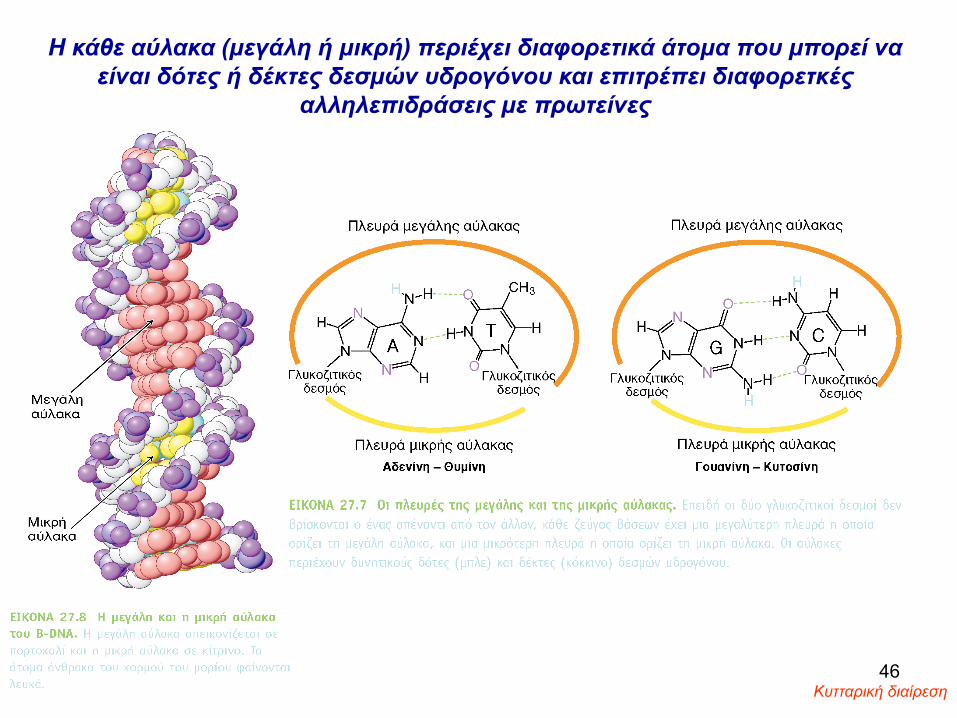
Η κάθε αύλακα (μεγάλη ή μικρή) περιέχει διαφορετικά άτομα που μπορεί να είναι δότες ή δέκτες δεσμών υδρογόνου και επιτρέπει διαφορετκές
αλληλεπιδράσεις με πρωτείνες
Κυτταρική διαίρεση 46
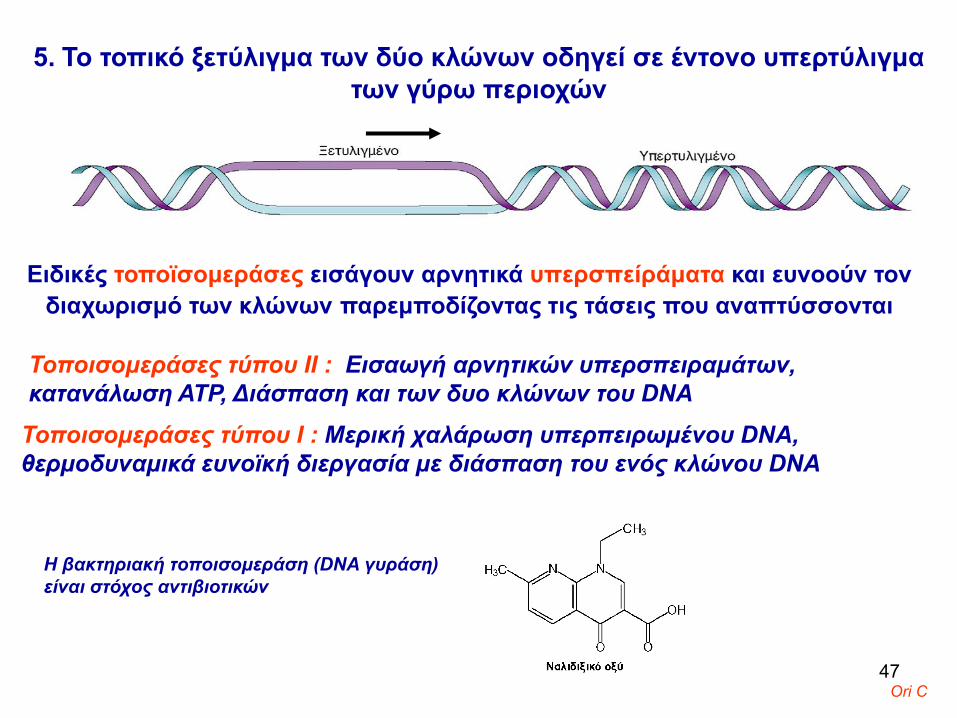
5. Το τοπικό ξετύλιγμα των δύο κλώνων οδηγεί σε έντονο υπερτύλιγμα των γύρω περιοχών
Ειδικές τοποϊσομεράσες εισάγουν αρνητικά υπερσπείράματα και ευνοούν τον διαχωρισμό των κλώνων παρεμποδίζοντας τις τάσεις που αναπτύσσονται
Τοποισομεράσες τύπου ΙΙ : Εισαωγή αρνητικών υπερσπειραμάτων, κατανάλωση ΑΤΡ, Διάσπαση και των δυο κλώνων του DNA
Τοποισομεράσες τύπου Ι : Μερική χαλάρωση υπερπειρωμένου DNA, θερμοδυναμικά ευνοϊκή διεργασία με διάσπαση του ενός κλώνου DNA
Η βακτηριακή τοποισομεράση (DNA γυράση) είναι στόχος αντιβιοτικών
Ori C47
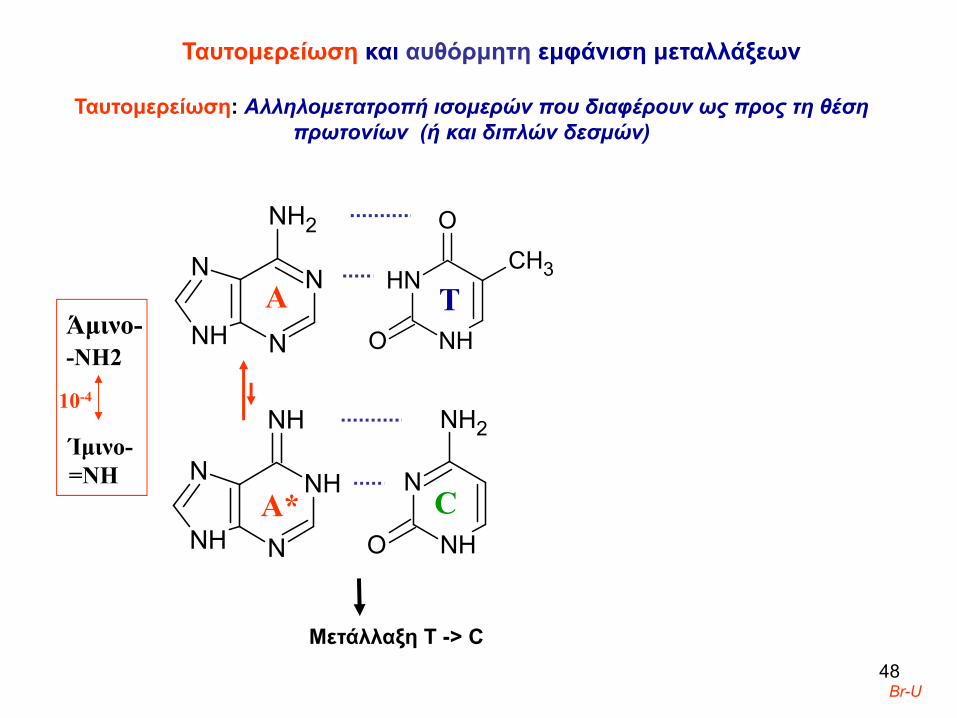
Ταυτομερείωση και αυθόρμητη εμφάνιση μεταλλάξεων
N
N
NH
N
NH2
NH
NH
O
O
CH3
Ταυτομερείωση: Aλληλομετατροπή ισομερών που διαφέρουν ως προς τη θέση πρωτονίων (ή και διπλών δεσμών)
Ίμινο-=ΝΗ
-ΝΗ2Άμινο-
Α Τ
N
NH
NH
N
NH
Α*N
NH
NH2
OC
Μετάλλαξη Τ -> C
Br-U
10-4
48










![Nucleosid * DNA polymerase { ΙΙΙ, Ι } * Nuclease { endonuclease, exonuclease [ 5´,3´ exonuclease]} * DNA ligase * Primase.](https://static.fdocument.org/doc/165x107/56649cab5503460f9496ce53/nucleosid-dna-polymerase-nuclease-endonuclease-exonuclease.jpg)







