
UNIVERSIDADE SÃO JUDAS TADEU PROGRAMA DE PÓS … · PGC-1α Peroxissomo gama coativador 1-alfa...
58
UNIVERSIDADE SÃO JUDAS TADEU PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DO ENVELHECIMENTO ESTUDO MORFOQUANTITATIVO DOS CARDIOMIOCITOS DO ÁTRIO DIREITO DE RATOS WISTAR JOVENS E IDOSOS SÃO PAULO FEVEREIRO/2014
Transcript of UNIVERSIDADE SÃO JUDAS TADEU PROGRAMA DE PÓS … · PGC-1α Peroxissomo gama coativador 1-alfa...

UNIVERSIDADE SÃO JUDAS TADEU
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DO ENVELHECIMENTO
ESTUDO MORFOQUANTITATIVO DOS CARDIOMIOCITOS DO
ÁTRIO DIREITO DE RATOS WISTAR JOVENS E IDOSOS
SÃO PAULO FEVEREIRO/2014

MARCELO SIMONI FERRO
ESTUDO MORFOQUANTITATIVO DOS CARDIOMIOCITOS DO
ÁTRIO DIREITO DE RATOS WISTAR JOVENS E IDOSOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências do Envelhecimento da Universidade São Judas Tadeu como requisito parcial para a obtenção do título de Mestre em Ciências do Envelhecimento. Orientador: Prof. Dr. Romeu Rodrigues de Souza
SÃO PAULO
FEVEREIRO/2014

Ferro, Marcelo Simoni F395e Estereologia dos cardiomiócitos do átrio direto de ratos wistar jovens e
idosos / Marcelo Simoni Ferro. - São Paulo, 2014.
05 f. ; 30 cm.
Orientador: Romeu Rodrigues de Souza.
Dissertação (mestrado) – Universidade São Judas Tadeu,
São Paulo, 2014.
1. Átrio. 2. Envelhecimento. I. Souza, Romeu Rodrigues de. II.
Universidade São Judas Tadeu, Programa de Pós-Graduação Stricto Sensu
em Ciências do Envelhecimento. III. Título
CDD 22 – 155.67
Ficha catalográfica elaborada pela Biblioteca
da Universidade São Judas Tadeu Bibliotecário: Ricardo de Lima - CRB 8/7464

MARCELO SIMONI FERRO
ESTUDO MORFOQUANTITATIVO DOS CARDIOMIÓCITOS DO
ÁTRIO DIREITO DE RATOS WISTAR JOVENS E IDOSOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências do
Envelhecimento da Universidade São Judas Tadeu como requisito parcial para obtenção do título de Mestre em Ciências do Envelhecimento.
BANCA EXAMINADORA
________________________________________________________ Prof. Dr. Romeu Rodrigues de Souza
Orientador
________________________________________________________ Prof. Dr.
________________________________________________________ Prof. Dr.
Data ____/____/____
Resultado ____________

Ao meu amigo e eterno mestre Dr. Laerte Yasushi Kawauchi. Seus ensinamentos e dedicação ao longo dos dez anos de convivência na década de 1990 me fizeram enxergar coisas maravilhosas no mundo do metabolismo, fazendo com que eu me apaixonasse pela vida profissional que tenho hoje. Contudo, com seus ensinamentos e convivência, eu aprendi muito mais do que metabolismo e bioquímica, aprendi com o Laerte a ser uma pessoa humana melhor. A vida nos separou apenas fisicamente. Enquanto fui seu amigo e aluno, seu caráter e postura foram dignos de exemplo que pretendo segui-los até o fim da minha vida. Obrigado por tudo sempre!

AGRADECIMENTOS
Primeiro, agradeço a Deus por ter tido saúde para conseguir terminar mais uma
etapa na minha vida.
A minha esposa Paula Marçal, pela paciência e tolerância comigo nos momentos
turbulentos durante todo esse mestrado. Mais uma vez, você demonstrou ser uma
excelente parceira que eu tive a sorte de encontrar na vida, me compreendendo e
dando incentivo para me tornar um homem melhor a cada dia.
Ao meu Orientador Professor Dr.Romeu Rodrigues, que foi extremamente dedicado
a mim em todos os momentos dessa caminhada. Foram três anos de muito
aprendizado e grandes momentos juntos.
A todos os professores da USJT, por terem me proporcionado, com muita
dedicação, grandes momentos de excelentes aulas que engrandeceram meu
aprendizado.
A minha mãe Célia que, infelizmente, não pode compreender mais as coisas desse
mundo. Contudo, se eu estiver enganado, eu não sou mais aquele menino
considerado com retardo de aprendizado pela coordenadora do ginásio, ok mãe?
Aos meus familiares, meus Pais, Moacyr e Nadia, minha irmã Monica e meu
sobrinho Octávio, por me apoiarem sempre mais nessa trajetória.
A minha sogra, Maria da Penha que, como futura doutora e brilhante professora,
sempre me incentivou a realizar esse mestrado.
Aos amigos, em especial aqueles que sempre me apoiaram e me acompanharam de
perto nesses anos de mestrado. Marcelo Quinn, Renato Freire, Karina Bonalume,
Carina Amorim, João Paulo Almeida, Henrique Costa, Bruno Ferro, Mauricio
Brito, Rodrigo Gimenes, Michely Capobiango, Renan Abraão, Hugo Marçal e Dr.
Hugo Marçal Filho (meu futuro parceiro em consultório). Obrigado de coração!

LISTA DE FIGURAS
Figura 1 – Fotomicrografias de cortes de dois cardiomiócitos do átrio direito de rato Wistar, mostrando grânulos de ANP e BNP no citoplasma (setas). A maior parte dos grânulos está situada junto ao núcleo do cardiomiócito. ......................................................................................... 12
Figura 2 – Fotomicrografias eletrônicas de cortes de cardiomiócitos do átrio direito de ratos Wistar mostrando poro nuclear (seta preta, em a) grânulos de ANP (setas brancas) e BNP (cabeças de setas), mitocôndrias (M), aparelho de Golgi (G, em a), retículo endoplasmático (ER), eucromatina (E) e núcleo (N). ............................. 15
Figura 3 – Fotomicrografias de cortes de cardiomiócitos do AD mostrando em a os componentes do cardiomiócito que foram quantificados e em b o sistema teste dotado de 70 pontos (em vermelho) sobreposto, utilizado para quantificação dos componentes. M – mitocôndrias, G – aparelho de Golgi, RE – retículo endoplasmático, N – núcleo. Seta preta cheia – poro da membrana do núcleo, setas brancas – grânulos de ANP, setas – grânulos de BNP. Barra: 0.2 µm. .................. 32
Figura 4 – Valores da PA sistólica nos dois grupos de ratos estudados: G1 e G2. Valores são média ± desvio padrão (n=10 animais por grupo)........ 33
Figura 5 – Fotomicrografias de corte transversal (a) e longitudinal (b) de cardiomiocitos do átrio direito de rato Wistar, mostrando em a, a medida do diâmetro do cardiomiócito (linha branca vertical) e em b a medida da área do seu núcleo (linha vermelha pontilhada). .................. 33
Figura 6 – Fotomicrografias de cortes transversais de cardiomiócitos do átrio direito de rato Wistar, mostrando em a, a medida do diâmetro de cardiomiócito do G1 e em b do G2 (linhas brancas). Observar maior diâmetro em b do que em a. N – núcleo. ............................................... 34
Figura 7 - Fotomicrografias de cortes transversais de cardiomiócitos do átrio direito de rato Wistar, mostrando em a, a medida da área do núcleo do cardiomiocito do G1 e em b a do G2 (linhas vermelhas pontilhadas). Observar maior área em b do que em a. N – núcleo. ....... 34
Figura 8 – Fotomicrografias eletrônicas de cortes de cardiomiócitos do átrio direito de ratos Wistar dos grupos G1 (a) e G2(b) mostrando os grânulos no citoplasma (setas) junto ao núcleo (N) de cada um dos cardiomiócitos. Barra: 2,1 μm (a); 2,2 μm (b). ........................................ 35
Figura 9 - Fotomicrografias Wistar, dos grupos estudados G1 (a) G2 (b) para mostrar os grânulos de eletrônicas de cortes do átrio direito de ratos ANP (setas pretas), de BNP (setas brancas), o núcleo (N) do cardiomiócito, o aparelho de Golgi (G), o retículo endoplasmático (RE) e as mitocôndrias (M). Observar as diferenças de tamanho e número dos grânulos entre os grupos. Barra: 0,2μm. ............................ 35
Figura 10 – Gráficos representativos do diâmetro do cardiomiócito (µm) (A) e da área de secção do seu núcleo (µm2) (B) nos grupos G1 e G2. Os valores são media ± desvio-padrão. *Significante vs. G1 (P<0,05). ....... 36

Figura 11– Gráficos representativos da Vv [eucromatina] (%) (A) e do número de poros por 10 µm de membrana nuclear (B) nos grupos G1 e G2. Os valores são média ± desvio-padrão. *Significante vs. G1 (P<0,05). ....... 37
Figura 12 - Densidade numérica (número por área) (A) e área dos grânulos de ANP (µm2) (B) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05) ............................................................................................ 38
Figura 13 - Densidade numérica (número por área) (A) e área dos grânulos de BNP (µm2) (B) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05) ............................................................................................ 39
Figura 14 - Densidades de volume (%) do aparelho de Golgi (A), das mitocôndrias (B) e do reticulo endoplasmático (C) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05). Os resultados no aumento da densidade de volume dessas organelas podem estar atrelados ao aumento da expressão do RNA, na produção dos peptídeos natriuréticos que demonstraram queda com o envelhecimento. ..................................................................................... 40
LISTA DE QUADROS
Quadro 1 – Índices Nucleicos.................................................................................... 41
Quadro 2 – Índices Citoplasmáticos .......................................................................... 41
Quadro 3 – Pressão Arterial ...................................................................................... 41
LISTA DE ABREVIATURAS
ANP Peptídeo natriurético atrial
ATP Trifosfato de adenosina
ATPsintase Trifosfato de adenosina sintase
BNP Peptídeo natriurético cerebral (Brian)
DAF-16/FOXO Fatores de transcrição forkhead
DNAmt Ácido desoxirribonucleico mitocondrial
ETS Sistema de transporte de elétrons
I-kβ Proteína inibitória do fator kapa-beta
mtRNAt Ácido ribonucleico tradutor mitocondrial
NAD(P)H Nicotinamida adenina dinucleotídeo (fosfato)
NF-kβ Fator nuclear kapa-beta
NQO1 Quinona oxidoredutase
NRF1 Fator respiratório nuclear 1
OXPHOS Fosforilação oxidativa
PGC-1α Peroxissomo gama coativador 1-alfa
Ran GTPase Proteína Ran dependente de trifosfato de guanosina
RNAm Ácido ribonucleico mensageiro
RNAt Ácido ribonucleico tradutor
TCA Ciclo do ácido tricarboxílico
Vv[eucrom] Densidade de volume da eucromatina
Vv[Golgi] Densidade de volume do aparelho de Golgi
Vv[mitoc] Densidade de volume da mitocôndria
Vv[retíc end] Densidade de volume do retículo endoplasmático

RESUMO
Este trabalho experimental tem o objetivo de comparar as diferenças quantitativas na composição e estrutura dos cardiomiócitos do átrio direito de ratos Wistar com três e 20 meses de idade utilizando a estereologia. Foram estudados 20 ratos (10 animais jovens com três meses de idade e 10 animais idosos com 20 meses de idade) denominados respectivamente de grupos G1 e G2. Fragmentos dos átrios direitos dos animais foram retirados, fixados com solução de glutaraldeído a 2% por 24 horas, processados por técnica ultramicroscópica, incluídos em resina Epon, seccionados e corados pelo tetróxido de ósmio. Foram analisados 10 campos por animal, com aumentos de x7500 (total de campos, 100) e 10 campos com aumentos de x15000 por animal (total de campos, 100), obtidos aleatoriamente, nos quais foram determinados os seguintes parâmetros estereológicos: diâmetro dos cardiomiócitos, área de secção do núcleo, volume relativo da eucromatina (Vv[eucrom]), densidade de poros da membrana nuclear (número de poros por 10 µm de membrana) e volume relativo do complexo de Golgi (Vv[Golgi], das mitocôndrias (Vv[mitoc]) e do retículo endoplasmático (Vv[re]). A densidade numérica e a área dos grânulos de peptídeo natriurético atrial (ANP) e peptídeo natriurético cerebral (BNP), presentes no citoplasma dos cardiomiócitos dos dois grupos foram também determinados. Os valores obtidos foram tabulados e as médias e erro padrão das médias dos grupos, comparadas para avaliar a significância das diferenças entre eles. Os dados foram testados para distribuição normal utilizando o teste de Kolmogorov – Smirnov e o ANOVA de caminho único e o pós-teste t de Student foi utilizado para comparar os grupos, sendo o nível de significância de 0,05. Comparando os animais dos grupos G1 e G2, obtivemos, respectivamente, os seguintes dados : o diâmetro dos cardiomiócitos ao nível do núcleo aumentou de 9,7 ± 0,3 para 11,2 ± 0,2 µm; a área de secção do núcleo aumentou de 10,9 ± 0,78 para 13,8 ± 0,89 µm2; a Vv[eucrom] aumentou de 72,4 ± 0,91 para 78,2 ± 0,89%; o numero de poros por 10 µm de membrana nuclear aumentou de 3,2 ± 0,09 para 5,1 ± 0,1; o Vv[Golgi] aumentou de 1,4 ± 0,1 para 2,0 ± 0,2%; o Vv[mitoc] aumentou de 27,2 ± 1,4 para 30,1 ± 2,9%; o numero de grânulos de ANP por área diminuiu de 29,2 ± 1,8 para 22,1 ± 1,5 e a área desses grânulos diminuiu de 0,130 para 0, 110 µm2. O numero de grânulos de BNP por área diminuiu de 18 ± 2 para 10 ± 3 e a área dos grânulos de BNP diminuiu de 48 ± 5, para 44 ± 3 nm. Essas diferenças foram estatisticamente significantes (p<0,05). Em conclusão: os resultados obtidos indicam que no envelhecimento ocorre hipertrofia dos cardiomiócitos , aumento do diâmetro do núcleo, aumento da densidade de volume da eucromatina, das mitocôndrias, do retículo endoplasmático e complexo de Golgi e redução da síntese de ANP e BNP. Palavras chave: Peptídeo natriurético atrial. Peptídeo natriurético cerebral. Átrio. Envelhecimento. Organelas.

ABSTRACT
This experimental study aimed to compare the quantitative differences in the composition and structure of the right atrial cardiomyocytes of Wistar rats at age of three and 20 months using stereology. 20 rats (10 young animals aged three months, and 10 older animals aged 20 months) called respectively G1 and G2 were studied. Fragments of animal rights atria were removed, fixed with a solution of 2% glutaraldehyde for 24 hours, processed by ultramicroscopic technique, embedded in Epon, sectioned and stained by osmium tetroxide. 10 fields per animal with increases of x7500 (total field of 100) and 10 fields with increases of x15000 per animal (total fields, 100) were randomly analyzed, from which were determined the following stereological parameters: diameter of cardiomyocytes, sectional area of the core, relative volume of euchromatin (Vv[eucrom]), pore density of the nuclear membrane (number of pores per 10 µm of membrane) and relative volume of the Golgi complex (Vv [Golgi], of mitochondria (Vv[mitoc]) and of the endoplasmic reticulum (Vv[re]). The numerical density and area of the granules of atrial natriuretic peptides ( ANP ) and brain natriuretic peptide (BNP) present in the cytoplasm of cardiomyocytes in both groups were also determined. Results were tabulated, and averages and standard errors of the group averages were compared to evaluate the significance of differences. Data were tested for normal distribution using the Kolmogorov-Smirnov test. The ANOVA of single way and post-test t of Student were used to compare the groups, with a significance level of 0.05. Comparing the animals in groups G1 and G2, we obtained, respectively, the following data: cardiomyocyte diameter to the core level increased from 9.7 ± 0.3 to 11.2 ± 0.2 µm; area of core section increased from 10.9 ± 0.78 to 13.8 ± 0.89 μm2 ; Vv[eucrom] increased from 72.4 ± 0.91 to 78.2 ± 0.89%; the number of pores per 10 µm in nuclear membrane increased from 3.2 ± 0.09 to 5.1 ± 0.1; Vv[Golgi] increased from 1.4 ± 0.1 to 2.0 ± 0.2%; Vv[mitoc] increased from 27.2 ± 1.4 to 30.1 ± 2.9%; the number of ANP granules per area decreased by 29.2 ± 1.8 to 22.1 ± 1.5 and the area of these granules decreased from 0.130 to 0.110 μm2. The number of BNP granules per area decreased from 18 ± 2 to 10 ± 3 and the area of the BNP granules decreased from 48 ± 5 to 44 ± 3 nm. These differences were statistically significant (p<0.05). Conclusion: The results indicate that aging results in cardiomyocyte hypertrophy, increased diameter of the core, increased volume density of euchromatin, mitochondria, endoplasmic reticulum and Golgi complex, and reduction of the synthesis of ANP and BNP. Keywords: Atrial natriuretic peptide. Brain natriuretic peptide. Atrium. Aging. Organelles.

SUMÁRIO
INTRODUÇÃO .......................................................................................................... 12
1 REVISÃO DA LITERATURA .................................................................. 14
1.1 ASPECTOS ULTRAESTRUTURAIS E FUNCIONAIS DOS CARDIOMIÓCITOS ATRIAIS ................................................................. 14
1.2 COMPOSIÇÃO E ESTRUTURA DOS CARDIOMIÓCITOS ATRIAIS .... 15
1.2.1 Grânulos de ANP ................................................................................... 15
1.2.2 Grânulos de BNP ................................................................................... 16
1.2.3 Funções do ANP e BNP ......................................................................... 16
1.2.4 Mecanismo de liberação do ANP e BNP ................................................ 17
1.2.5 Importância clinica do ANP/BNP ............................................................ 18
1.2.6 Mitocôndrias ........................................................................................... 19
1.2.7 Biogênese mitocondrial .......................................................................... 20
1.2.8 Função mitocondrial ............................................................................... 20
1.2.9 Efeitos do envelhecimento nas mitocôndrias ......................................... 22
1.2.10 Aparelho de Golgi ................................................................................... 23
1.2.11 Funções do aparelho de Golgi ............................................................... 23
1.2.12 Efeitos do envelhecimento no aparelho de Golgi ................................... 24
1.2.13 Retículo endoplasmático ........................................................................ 24
1.2.14 Função do retículo endoplasmático ........................................................ 25
1.2.15 Efeitos do envelhecimento no retículo endoplasmático .......................... 25
1.2.16 Membrana nuclear e eucromatina .......................................................... 26
1.2.17 Função da membrana nuclear e eucromatina ........................................ 26
1.2.18 Efeitos do envelhecimento na membrana nuclear e eucromatina .......... 27
2 OBJETIVOS ........................................................................................... 29
2.1 OBJETIVO GERAL ................................................................................ 29
2.2 OBJETIVOS ESPECÍFICOS .................................................................. 29
3 MATERIAIS E MÉTODOS ...................................................................... 30
3.1 ANIMAIS E COLETA DE MATERIAL ..................................................... 30
3.2 SACRIFÍCIO DOS ANIMAIS .................................................................. 30
3.3 QUANTIFICAÇÃO DOS COMPONENTES DOS CARDIOMIÓCITOS ... 31
3.4 ANÁLISE ESTATÍSTICA ........................................................................ 32
4 RESULTADOS ....................................................................................... 33
4.1 PRESSÃO ARTERIAL E PESO DOS ANIMAIS ..................................... 33
4.2 MORFOLOGIA DOS CARDIOMIÓCITOS .............................................. 34

4.3 ANÁLISE QUANTITATIVA ..................................................................... 36
4.3.1 Índices nucleicos .................................................................................... 36
4.3.2 Índices citoplasmáticos .......................................................................... 38
DISCUSSÃO ............................................................................................................. 42
CONCLUSÃO ............................................................................................................ 49
REFERÊNCIAS ......................................................................................................... 50

12
INTRODUÇÃO
As mudanças estruturais dos cardiomiócitos dos ventrículos cardíacos
decorrentes do envelhecimento estão bem estabelecidas na literatura (SARMA et al,
2013). Entretanto, as mudanças do envelhecimento nos cardiomiócitos dos átrios
ainda não estão totalmente esclarecidas.
Os cardiomiócitos dos átrios possuem um aparelho secretor constituído por
estruturas nucleares e citoplasmáticas, responsáveis pela produção dos peptídeos
natriuréticos (PENG et al, 2013). Os peptídeos ficam armazenados no citoplasma,
sob a forma de grânulos eletrondensos, alguns contendo o Peptídeo Natriurético
Atrial (ANP) e outros, o Peptídeo Natriuretico Cerebral (BNP) (CAVALLINI et al,
1994) (Fig. 1).
Figura 1 – Fotomicrografias de cortes de dois cardiomiócitos do átrio direito de rato Wistar, mostrando grânulos de ANP e BNP no citoplasma (setas). A maior parte dos grânulos está situada junto ao núcleo do cardiomiócito.
Os peptídeos natriurético ANP e BNP são sintetizados a partir de uma
proteína denominada fator de transcrição kapa βeta (NF-kβ), a qual regula a
expressão gênica do ANP e BNP (GLEZER et al 2000). Segundo os mesmos
autores, o fator NF-kβ, quando não estimulado, encontra-se no citoplasma ligado a
uma proteína inibitória (I-kβ) impedindo sua translocação para o núcleo, sendo
necessária a fosforilação e degradação do I-kβ para que ocorra a translocação e
ativação da RNA polimerase.
Tanto o ANP como o BNP são hormônios que lançados no sangue, atuam
nos túbulos renais produzindo redução da pressão arterial, pelo aumento da
secreção de sódio e água e, consequente queda da volemia (CALIARI E TAFURI,

13
1993; MCGRATH et al, 2005; CLERICO et al, 2006; CHARLOUX et al, 2007; Pan,
2008).
Em ratos idosos, ocorre diminuição significante do conteúdo de ANP nos
átrios em relação a ratos jovens (WU et al, 2008; BEGG et al, 2012). Mifune et al
(1991) observaram redução do número e diâmetro dos grânulos de ANP em átrio
direito de gerbil idoso em relação ao jovem. Em outro estudo, Pollack et al (1997)
observaram redução da secreção de ANP em ratos idosos. Entretanto, na literatura
consultada, não foram encontradas informações mostrando a composição e
estrutura dos cardiomiócitos na idade avançada. Tais dados são importantes como
base para estudos da resposta dos cardiomiócitos a estímulos experimentais em
ratos jovens e idosos ou quando submetidos a estímulos como exercício ou ação de
substâncias químicas, por exemplo.
A estereologia é um método de morfologia quantitativa que permite avaliar as
alterações do envelhecimento nos cardiomiócitos (KAJSTURA et al, 1996). As
alterações estruturais nos cardiomiócitos podem ser avaliadas pela análise
quantitativa relativa de seus componentes nucleares e citoplasmáticos utilizando
tanto a estereologia como a medida dos diâmetros de seus componentes
(MAKSIMOV et al, 2004). Existe concordância na literatura que métodos
morfológicos quantitativos, tais como a estereologia, são eficientes para determinar
alterações do envelhecimento em cardiomiócitos (ÁGUILA et al, 1998).
O presente trabalho experimental tem o objetivo de comparar as diferenças
quantitativas na composição e ultraestrutura dos cardiomiócitos em duas fases
extremas da vida do rato Wistar: três e 20 meses de idade. Estas idades no rato
correspondem, nos seres humanos, ao adulto jovem e ao idoso (GERRITY e CLIFF,
1972).

14
1 REVISÃO DA LITERATURA
Estudos sobre alterações estruturais do envelhecimento nos cardiomiócitos
ventriculares vêm sendo realizados desde longo tempo. Sabe-se que com o
avançar da idade ocorre hipertrofia dos cardiomiócitos ventriculares (ZHANG et al,
2007). Acredita-se que a causa desse fenômeno seja a redução do seu número com
o envelhecimento, o que levaria ao aumento da espessura do miocárdio, visando a
manter a força de ejeção (Yang et al, 2012). Esta é uma alteração normal do
envelhecimento. Entretanto, outros fatores podem interferir nesse processo, durante
o envelhecimento, como, por exemplo, a presença de hipertensão arterial (ZHAO, et
al 2010) que, associada à idade (CANNONE et al, 2011), poderia aumentar ainda
mais a hipertrofia dos cardiomiócitos.
Em relação aos cardiomiócitos atriais, as alterações decorrentes do
envelhecimento ainda não estão totalmente esclarecidas na literatura. A estereologia
é um método morfológico quantitativo que permite analisar com precisão alterações
nos cardiomiócitos em diferentes idades (KAJSTURA et al, 1996).
1.1 ASPECTOS ULTRAESTRUTURAIS E FUNCIONAIS DOS CARDIOMIÓCITOS
ATRIAIS
Os cardiomiócitos atriais possuem um núcleo central, organelas
citoplasmáticas bem evidentes, como o aparelho de Golgi, mitocôndrias e retículo
endoplasmático, além da presença de numerosos grânulos de tamanhos variados,
contendo os denominados peptídeos natriuréticos atriais (ANP e BNP) (Fig. 2). Estes
componentes nucleares e citoplasmáticos estão todos envolvidos na produção e
liberação do ANP e BNP.

15
Figura 2 – Fotomicrografias eletrônicas de cortes de cardiomiócitos do átrio direito de ratos Wistar mostrando poro nuclear (seta preta, em a) grânulos de ANP (setas brancas) e BNP (cabeças de setas), mitocôndrias (M), aparelho de Golgi (G, em a), retículo endoplasmático (ER), eucromatina (E) e núcleo (N).
Por esse motivo, os componentes nucleares e citoplasmáticos dos
cardiomiócitos, especialmente a cromatina nuclear, os poros da membrana nuclear,
o aparelho de Golgi, mitocôndrias e retículo endoplasmático foram denominados por
alguns autores de aparelho secretor dos cardiomiócitos (MAKSIMOV et al, 2004),
devido ao fato de que a produção e secreção do ANP e BNP depende desses
componentes.
A maior parte do ANP e BNP encontram-se armazenados no citoplasma dos
cardiomiócitos atriais sob a forma de grânulos de tamanhos variados em associação
com os componentes citoplasmáticos dos cardiomiócitos (CAVALLINI et al, 1994;
FREDERICO, 2010; KLAIBER, et al, 2011) (Fig. 2). Os grânulos estão geralmente
situados próximo ao núcleo, sendo que uns poucos estão espalhados pelo
citoplasma (TOYOSHIMA et al, 1996). A distribuição quantitativa de grânulos difere
entre os átrios. Gama et al (2007) mostraram presença de maior número desses
grânulos nos cardiomiócitos do átrio direito do que no esquerdo.
1.2 COMPOSIÇÃO E ESTRUTURA DOS CARDIOMIÓCITOS ATRIAIS
1.2.1 Grânulos de ANP
Os grânulos de ANP são esféricos, regulares e apresentam-se ao
microscópio eletrônico, nos cortes, como círculos contendo um limite nítido,
envolvendo um halo em torno de uma área mais escura, contendo o ANP (Figura 2).

16
Skepper et al (1988) analisaram as dimensões dos grânulos atriais em ratos e
verificaram que eles medem de 100 a 250 nanômetros de diâmetro com variações,
em diferentes espécies.
Os grânulos de ANP foram descritos pela primeira vez por Bruno Kisch em
1956 (SILVA at al, 2003) em cobaias. Posteriormente, quando se constatou que
esses grânulos continham uma substância química que atuava como fator
natriurético, a substância passou a ser denominada de Peptídeo Natriurético Atrial
(PALADE et al, 1961). Para verificar a função do ANP dos grânulos, De Bold et al
(1981) prepararam extrato de átrios e injetaram em peritônio de ratos. Observaram
aumento na diurese, excreção renal de sódio, diminuição da pressão arterial e
aumento do hematócrito nos animais que receberam o extrato atrial. A partir dessa
descoberta, numerosos trabalhos têm sido realizados sobre os cardiomiócitos atriais
e sobre os grânulos de ANP em várias situações.
1.2.2 Grânulos de BNP
Os grânulos de BNP, também esféricos, não possuem halo e são geralmente
menores, apresentando frequentemente espículas na sua superfície (DICSTEIN,
1998 e DICSTEIN, 2001) (Fig. 2). Esses grânulos de BNP foram descobertos
recentemente, sendo também conhecidos como peptídeo natriurético cerebral
(reconhecido pela sigla BNP, de Brain Natriuretic Peptide). Este peptídeo é assim
denominado porque foi identificado, inicialmente, no cérebro de macacos, sendo
que, em humanos, é produzido, principalmente, pelos átrios e ventrículos cardíacos,
em resposta a uma sobrecarga de pressão ou volume. Assim como o ANP, o BNP
promove diurese e vasodilatação (DICSTEIN, 1998; MAEDA, 1998 e DICSTEIN,
2001, CANNONE et al, 2011). Além de ser produzido também no cérebro, ele difere
do ANP, pelo fato de ter uma vida curta, de apenas 22 min, o que sugere que ele
seja um hormônio de emergência do coração, refletindo o momento atual de
sobrecarga ventricular (VILLACORTA et al, 2002).
1.2.3 Funções do ANP e BNP
Ambos os peptídeos, tanto o ANP como o BNP, têm demonstrado uma ação
decisiva nos mecanismos de controle da pressão arterial (ZHAO, PANSY E NAVAR

17
2010), pois quando lançados na corrente sanguínea, atuam nos túbulos renais e
produzem redução da pressão arterial, pelo aumento da secreção de sódio e água, e
consequente queda da volemia (CALIARI e TAFURI, 1993; CLERICO, et al, 2006;
PAN, 2008).
A partir da descoberta desses hormônios, vários estudos foram realizados.
Assim, por exemplo, para demonstrar as ações do ANP, Schiebinger & Greening,
(1992) mostraram que a retirada bilateral dos átrios eliminava a liberação destes
peptídeos e bloqueava a excreção de água e sódio com retenção de líquidos nos
tecidos corporais, devido ao aumento do volume plasmático. Para mostrar a
importância desse hormônio, Miglioranza et al (2013) demostraram que o ANP atua
também na monitoração de tratamento dos pacientes com insuficiência cardíaca,
sugerindo-se que o peptídeo poderia servir como parâmetro terapêutico, propondo o
retorno do peptídeo aos níveis séricos basais como objetivo final do tratamento, e
não mais apenas na melhora dos sintomas. Woodman et al (2008) também
relataram que o ANP promove efeitos benéficos, não somente no sistema
cardiovascular, como também possui atividades antioxidantes na disfunção
endotelial em ratos diabéticos. Apesar de como esse mecanismo funciona ainda
estar obscuro, os mesmos autores demonstraram que o ANP melhora o estresse
oxidativo vascular em conjunto com a função endotelial, independente de quaisquer
efeitos sobre os níveis de glicose no plasma. Cannone et al (2011) relataram que, o
ANP tem ainda outras ações importantes além do aumento da diurese, dentre as
quais a vasodilatação, a supressão do sistema renina-angiotensina-aldosterona e a
inibição da hipertrofia dos cardiomiócitos. Para efeitos elucidativos sobre a função
fisiológica do ANP, Wu et al (2008) observaram que peixes adaptados a nadar tanto
em água salgada como em água doce, como por exemplo o salmão, tinham
concentrações significativamente maiores de ANP para manter a homeostase e a
regulação do sódio quando esses peixes migram da água doce para a água salgada.
1.2.4 Mecanismo de liberação do ANP e BNP
Segundo Soualmia et al (2007), o ANP produzido pelos átrios é secretado na
corrente sanguínea em resposta a vários estímulos, dentre eles os do sistema
nervoso autônomo (SNA) e do sistema renina-angiotensina. O conteúdo dos
grânulos é eliminado para o meio extracelular por exocitose. A sinalização para

18
liberação do ANP e BNP ocorre por dois mecanismos: encurtamento da musculatura
dos átrios (CHO et al, 1991; SEUL et al, 1992) e aumento da volemia, com dilatação
das paredes das cavidades atriais (SKEPPER et al, 1989; VOEUX, 2002). Estes
autores observaram maior quantidade de grânulos em animais submetidos à
expansão volumétrica dos átrios quando comparado com os animais não
submetidos a essa condição. Uma variedade de estímulos fisiológicos e
deflagradores pode promover o estiramento dos átrios e a liberação desses
peptídeos na corrente sanguínea. Entre esses são citados: exercícios físicos
aeróbicos, hipóxia, isquemia e aumento do estresse (FLATO et al, 2009).
Em relação ao estresse, de acordo com Flato et al (2009), na ocorrência de
estresse o cortisol e a adrenalina são liberados na corrente sanguínea, provocando
aumento da pressão arterial. Isso ocorre porque o eixo hipotálamo-pituitária-adrenal
é sensivelmente alterado em condições de estresse crônico, elevando não só o
cortisol, mas também a arginina vasopressina (AVP) e as catecolaminas,
provocando aumento da vasoconstrição arterial e consequentemente elevação da
pressão arterial (ESPINER et al 2002). Segundo Viana (1991) e Shaikh et al (2013),
esses fenômenos elevariam a pressão intra-atrial pela elevação do volume atrial;
outros fatores que podem elevar essa pressão são: agentes constritores, imersão
em água, taquicardia atrial e dieta rica em sais (VIANA, 1991, SHAIKH et al, 2013).
Apesar da liberação do ANP estar principalmente relacionada à ação dos
hormônios, como cortisol, AVP, catecolaminas e renina-angiotensina-aldosterona,
Schiebinger e Greening (1992) e Turkovsh et al (2013) demonstraram que a
norepinefrina, endotelina e vasopresina também atuam como mediadores
responsáveis pela liberação do ANP. A sua liberação é mais eficaz durante a
contração dos átrios por estímulo extrínseco, quando são ativadas ambas as
cavidades atriais (SKEPPER et al, 1988; FENZL et al, 2013).
1.2.5 Importância clinica do ANP/BNP
Após a descoberta das ações fisiológicas do ANP, este passou a ser utilizado
clinicamente, como terapia, em pacientes com doenças cardiovasculares
(McGRATH et al, 2005). Vários trabalhos experimentais passaram a ser realizados
demonstrando as aplicações do ANP. Myoshi et al (2001) observaram, em pacientes
internados com hipertensão arterial, que o uso de ANP intravenoso reduzia a

19
pressão sanguínea sistólica, pré-carga e pós-carga, e melhorava o desempenho do
ventrículo esquerdo. Ehara et al (2013) compararam o efeito do ANP associado à
nitroglicerina na remodelação do ventrículo esquerdo em pacientes com Infarto
Agudo do Miocárdio (IAM). Nesse estudo foi administrado ANP e nitroglicerina, e foi
observado, nos sujeitos que receberam ANP, que o tratamento surtiu efeito
significativo na remodelação do ventrículo esquerdo.
Outros pesquisadores realizaram trabalhos com infusão intravenosa contínua
de ANP em pacientes portadores de fibrilação atrial e insuficiência cardíaca
congestiva e observaram melhora nas primeiras 24 e 48 horas da fibrilação atrial,
melhora na resistência vascular periférica, pressão arterial sistólica, além de
decréscimo significativo na resistência vascular pulmonar, acréscimo na excreção de
diurese e sódio, decréscimo significativo de creatinina com melhora da
hemodinâmica (McGRATH et al, 2005). Graffe et al (2012) demonstraram que esse
processo ocorre pelo aumento de uma proteína denominada aquaporina 2,
responsável pelo transporte de água através da membrana nos ductos coletores dos
rins, como se fossem canais de água. Quando há aumento significativo de ANP na
corrente sanguínea, essa proteína se desloca para o citoplasma, diminuindo a
reabsorção de água nos ductos renais, aumentando a diurese, melhorando assim os
níveis dos marcadores relatados acima (BEATZ et al 2012).
No estudo de Sward et al (2001), foi infundido ANP intravenoso contínuo em
pacientes com disfunção renal aguda, sendo observada uma resposta significativa
na hemodinâmica nas primeiras 48 horas, aumento da filtração glomerular e fluxo
renal, diminuição da resistência vascular renal com aumento na fração de filtração.
Em outro estudo, Abassi et al (2013) demonstraram que a infusão de ANP em ratos
promoveu um aumento da taxa da excreção urinária, aumento da excreção de sódio
e aumento na taxa da filtração glomerular em ratos induzidos a nefropatias, quando
comparados ao grupo controle.
1.2.6 Mitocôndrias
A mitocôndria é um tipo de organela que contém características bem distintas
quando comparado a outras organelas. É a única que possui dupla membrana, uma
interna outra externa, e carrega seu próprio genoma (SAFDAR et al 2011). Acredita-
se que essa organela originou-se de bactérias primitivas pelo próprio processo

20
evolutivo (NELSON e COX 2005). Anatomicamente, os mesmos autores relataram
que a membrana interna de uma única mitocôndria do fígado pode ter mais de
10.000 conjuntos de sistemas de transferência de elétrons (cadeia respiratória) e de
moléculas de ATP sintase distribuídas sobre a superfície da membrana. Contudo, a
mitocôndria do músculo cardíaco apresenta três vezes mais conjuntos de sistemas
de transferência de elétrons e uma área de membrana interna muito maior quando
comparado às mitocôndrias hepáticas pela alta demanda de energia necessária
nesses tipos de tecidos.
1.2.7 Biogênese mitocondrial
A biogênese mitocondrial é um processo complexo que requer a expressão
coordenada de aproximadamente 1500 proteínas codificadas entre os genomas
nuclear e mitocondrial (SAFDAR et al, 2011). Jeong et al (2012) afirmaram que a
regulação da biogênese das mitocôndrias ocorre através do peroxissomo gama
coativador 1-alfa (PGC-1α) em conjunto com o fator respiratório nuclear1 (NRF1),
dois fatores de transcrição atrelados a síntese de ácidos ribonucleicos mensageiros
(RNAm) de codificação das proteínas mitocondrial. O NRF1 inicia a transcrição
responsável pela biogênese das mitocôndrias, enquanto que o PGC-1α estabiliza a
rede da regulação dos genes. Os mesmos autores ainda afirmam que a biogênese
mitocondrial é um processo complexo envolvendo várias proteínas e genes
nucleares, evidenciando a necessidade de mais estudos para elucidar melhor esse
processo. Contudo, Vilardo et al (2012) relataram que os RNA de transferência
(RNAt), presentes nos ribossomos, são os adaptadores essenciais na decodificação
do RNAm na biogênese celular, demontrando que modificações no RNAt
mitocondrial (mtRNAt), embora em pequeno número, parece ser fundamental para
alcançar o correto enrolamento dessa organela.
1.2.8 Função mitocondrial
Tecidos que exigem alto processo oxidativo pela grande demanda de energia,
como a musculatura esquelética e a musculatura cardíaca, dependem das
mitocôndrias para um perfeito funcionamento. A capacidade oxidativa do tecido
muscular e a preservação das mitocôndrias dependem do PGC-1α, considerado um

21
regulador máster do metabolismo energético (RUIZ et al, 2012). As mitocôndrias
utilizam o oxigênio e demais substratos para a maior parte da produção de trifosfato
de adenosina (ATP) no interior das células, que são essenciais para numerosos
processos fisiológicos, dentre eles produção de energia, potencial redox, modulação
do cálcio e diversas vias metabólicas (CALI, OTTOLINI e BRINI, 2012; PETERSON,
JOHANSEN e RAVUSSI, 2012). Estas organelas também participam de outros
processos celulares, incluindo sinais de tradução, regulação do ciclo celular,
estresse oxidativo, termogênese e apoptose (PETERSON, JOHANSEN e RAVUSSI,
2012).
A mitocôndria é responsável por 95% da produção de ATP no interior das
células, e muito numerosa em tecidos que exigem grandes quantidades de energia,
como o coração, músculo esquelético e neurônios (VADAVALKAR et al 2012).
Substratos como glicose, ácidos graxos e aminoácidos são convertidos em energia
no interior das mitocôndrias no ciclo do ácido tricarboxilico (TCA) pelo sistema de
transporte de elétrons (ETS) para a produção de ATP. Esse processo é chamado de
fosforilação oxidativa (OXPHOS), sendo fundamental para diversos processos
celulares (PETERSON, JOHANSEN E RAVUSSIN, 2012). Dorn (2013) relatou, em
seu trabalho, que, por causa da grande quantidade de ATP ( g dia) para
alimentar a contração do músculo cardíaco, o coração tem a maior densidade de
mitocôndrias quando comparado a qualquer outro órgão.
Além de sua função central na produção de ATP, as mitocôndrias também
estão envolvidas no processo de morte celular, a apoptose (PETERSON,
JOHANSEN e RAVUSSIN, 2012). A apoptose é definida como um modo
extremamente sincronizado de morte celular, sendo caracterizada por características
morfológicas distintas, incluindo a condensação de cromatina e fragmentação
nuclear. A importância da sinalização tem sido reconhecida na regulação da célula
durante o funcionamento normal e na doença (RASUL et al 2013). Os mesmos
autores relataram que duas vias principais estão envolvidas para iniciar a apoptose,
a intrínseca ou mitocondrial, e a extrínseca, causada por processos quimicos, ou
radioativos que interagem com um receptor específico na membrana plamática. A
primeira consequência do inicio do processo da apoptose é o aumento da
permeabilidade da membrana mitocondrial externa, permitindo a fuga do citocromo
c, ativando uma enzima proteolítica, denominada de caspase, responsável pela
degradação das proteínas no citoplasma (NELSON e COX 2005). Os mesmos

22
autores ainda relataram que esse é um caso expressivo de uma proteína
executando duas funções muito diferentes entre si.
1.2.9 Efeitos do envelhecimento nas mitocôndrias
No envelhecimento em humanos, ratos e macacos, anormalidades nas
mitocôndrias e a mutagênese no ácido desoxirribonucleico mitocondrial (DNAmt)
estão atreladas à degeneração de multissistemas, intolerância ao estresse e
diminuição de energia (SAFDAR et al, 2011). Recentemente, foi mencionada a ação
de uma enzima denominada nicotinamida adenina dinucleotideo (fosfato) [NAD(P)H]:
Quinona oxidoredutase (NQO1), ou NAD(P)H:NQO1, a qual estaria diretamente
envolvida no processo de envelhecimento celular mitocondrial do músculo cardíaco
(LEE et al, 2012). Segundo os autores, essa enzima modula a relação NAD+/NADH,
uma das causas conhecidas do envelhecimento celular e doenças relacionadas ao
envelhecimento, que estariam diretamente atreladas às mitocondrias e na produção
de energia. A função dessa enzima é diminuida em tecidos envelhecidos, podendo
ser minimizada com a indução de restrição calórica na dieta.
Nas mitocôndrias dos cardiomiócitos, Peterson, Johansen e Ravussin (2012)
observaram declínio nas funções bioquímicas e bioenergéticas no processo de
envelhecimento. Os mesmos autores relataram que as mitocôndrias disfuncionais
desempenham um papel chave na redução da função muscular, evidenciando que
mudanças bioquímicas e bioenergéticas, citadas acima, nas mitocôndrias, não
refletem somente na diminuição das funções do tecido muscular, mas também estão
associadas a determinadas patologias, dentre elas resistência a insulina,
inflamações sistêmicas e infiltração de gorduras no músculo (PETERSON,
JOHANNSEN e RAVUSSIN, 2012).
Quando o número ou a atividade mitocondrial é insuficiente, o corpo humano
entra rapidamente em fadiga pela deficiência de ATP. Por esse motivo, o controle da
quantidade e das funções das mitocôndrias é extremamente importante na busca da
otimização do balanço energético dos tecidos (JEONGETAl, 2012). De acordo com
Cali, Ottoni e Brini (2012), quando o número, volume, forma e distribuição das
mitocôndrias, no interior das células são bem controlados, as células entram
facilmente em homeostase.

23
1.2.10 Aparelho de Golgi
Séculos atrás, o anatomista Italiano chamado Camilo Golgi descrevia uma
nova organela que, posteriormente, levaria o seu nome, o aparelho de Golgi ou
complexo de Golgi (ALONSO, TOMÁS e MENARGUES, 2013). O aparelho de Golgi
é constituído por um numero variável de vesículas esféricas de diversos tamanhos
que variam a sua localização dependendo do tecido (LADINSKY et al 1999).
Segundo Alonso, Tomas e Menargues (2013), as primeiras imagens ultraestruturais
obtidas a partir de cortes ultrafinos mostrou a complexidade excepcional dessa
organela e, consequentemente, através da microscopia eletrônica e a estereologia,
foi possível utilizar dados para informações sobre o aparelho de Golgi, o qual é
composto de cisternas achatadas rodeadas por túbulos e vesículas. Os mesmos
autores ainda relataram que o número de cisternas depende de cada organismo e
seu lúmen é geralmente bastante estreito (10-20 nm), o que permite a interação das
enzimas de glicosilação presentes nas suas membranas. Ispolatov e Musch (2013),
corroborando com os trabalhos acima, ainda demonstraram que, em mamífero, o
aparelho de Golgi é composto por seis a oito cisternas não idênticas na sua
morfologia.
1.2.11 Funções do aparelho de Golgi
O aparelho de Golgi apresenta múltiplas funções, dentre as mais importantes
o enderaçamento das moléculas e proteínas síntetizadas nas células,
encaminhando-as para as vesículas de secreção (WARD et al 2001). Alonso, Tomas
e Menargues (2013) descreveram duas funções principais do aparelho de Golgi. A
primeira é a modificação pós-tradução de proteínas e lípidos provenientes do
retículo endoplasmático, principalmente a sua glicosilação. A segunda é a
concentração, a embalagem e a exportação destes produtos modificados para o
destino final, dentro ou fora da célula. Assim, o aparelho de Golgi é, ao mesmo
tempo, uma fábrica de glicanos eficiente e um ponto de logística na realização
destas funções, sendo uma matriz surpreendentemente de complexo de membranas
equipados com máquinas precisas.
Apesar do grande volume de tráfego de entrada e de saída, que é capaz de
manter a sua arquitetura e, embora também seja suficientemente flexível para

24
desmontar e remontar sob certas condições, tais como a mitose, em células
neuroendócrinas, pró-hormônios como o ANP são frequentemente glicosilados e
proteoliticamente processados antes de ser classificados em grânulos secretores
pelo próprio aparelho de Golgi (VÁZQUEZ-MARTÍNEZ et al, 2012).
1.2.12 Efeitos do envelhecimento no aparelho de Golgi
No aparelho de Golgi, Kiosses e Kalnins (1993) observaram alterações
degenerativas significativas no envelhecimento em células endoteliais da parede da
aorta. Nakagomi et al (2008) demonstraram fragmentação no aparelho de Golgi em
células nervosas com o envelhecimento. Outros autores, como Ispolatov e Musch
(2013), relataram perda de proteínas nas cisternas no aparelho de Golgi, que
especifica diminuição na fusão das vesículas à medida que essas organelas
envelhecem. Os mesmos autores demonstraram analiticamente que a concentração
de proteínas que regulam a fusão dessas organelas decai exponencialmente com o
número de cisterna. O mecanismo de como isso funciona ainda é obscuro, relataram
os autores.
1.2.13 Retículo endoplasmático
Nas células eucariontes, o retículo endoplasmático se encontra no citoplasma
próximo ao núcleo. Em sua morfologia, apresenta uma rede de vesículas achatadas,
vesículas esféricas e túbulos que se intercomunicam formando um sistema contínuo.
Esses elementos possuem uma parede formada por unidade de membrana que
delimita cavidades, as cisternas, emergentes do envelope nuclear, no qual
constituem um sistema de túneis, de forma muito variável presentes no citoplasma.
O retículo endoplasmático contém dois domínios distintos, o liso e o rugoso,
que apresenta ribossomos localizados na sua membrana (AMARILIO et al, 2005).
Os mesmos autores ainda afirmam que o retículo endoplasmático se conecta com
outras organelas, tais como o aparelho de Golgi, mitocôndrias e membrana
plasmática para diversos processos metabólicos. Segundo Elgaard e Helenius
(2003), o lúmen do retículo endoplasmático é extracitosólico, ou seja,
topologicamente equivalente ao meio extracelular, sendo, por conseguinte, diferente
do humor citosólico.

25
1.2.14 Função do retículo endoplasmático
O retículo endoplasmático tem várias funções diferenciadas, dentre elas a
translocação de proteínas através da membrana do próprio retículo, a integração de
proteínas dentro da membrana, modificação das proteínas dentro do lúmen, síntese
de fosfolipídios e esteroides no lado citosólico da membrana, armazenamento de
íons de cálcio no lúmen e regulação na liberação do cálcio para o citosol (VOELTZ
et al, 2002). Outros autores, como Helgaard e Helenius (2003), também relataram
que o retículo endoplasmático fornece um ambiente otimizador para o dobramento
de proteínas e sua maturação.
Embora a função do retículo endoplasmático tenha sido extensivamente
estudada, o mecanismo pelo qual essa organela mantém a sua estrutura
característica in vivo permanece largamente desconhecida. Estudos de levedura e
células de mamífero têm mostrado que o tamanho e/ou estrutura do retículo
endoplasmático é extremamente sensível a determinadas condições de estresse
celular (AMARILIO et al, 2004).
1.2.15 Efeitos do envelhecimento no retículo endoplasmático
O estresse do retículo endoplasmático, sob quaisquer condições adversas,
pode levar à diminuição da síntese de proteínas e, consequentemente, ao aumento
da expressão das chaperonas moleculares, as quais promovem o dobramento
adequado das proteínas na recuperação das células (CHEN et al, 2013). Autores
como Cook et al (2012) relataram que o retículo endoplasmático tem fator crucial na
homeostase do cálcio intracelular, e que, no envelhecimento do tecido cardíaco,
esse processo fica extremamente comprometido. Os mesmos autores também
afirmaram que a manutenção dos níveis intracelulares de cálcio é fundamental para
a função das células e sua sobrevivência. Segundo Janczewski e Lakkata (2010), as
concentrações de cálcio dentro das células tendem a desregular-se, em
consequência do envelhecimento, comprometendo a homeostase. Os autores
afirmam também que esse desequilíbrio desencadeia complicações fisiológicas
significativas, como a função sistólica e diastólica e, consequentemente, arritmias.
Esse processo é determinado pela estrutura e característica de permeabilidade da
membrana do próprio retículo, que sofre alterações no processo de envelhecimento.

26
1.2.16 Membrana nuclear e eucromatina
A membrana nuclear de células eucarióticas é rica em proteínas que são
necessárias para a estrutura nuclear, a organização cromossomica, a reparação do
DNA e no controle transcricional (KATTA et al, 2013). O DNA do núcleo é altamente
compactado e organizado, de modo a permitir ser ajustado dentro do limite do
próprio núcleo (RODRIGUES e BJERLIN 2013). Compreender o papel da estrutura
da cromatina e dinâmica na regulação das funções nucleares, incluindo a transcrição
e replicação, é um grande desafio da atual pesquisa em genômica (JULIENNE et al,
2013).
A eucromatina é caracterizada por uma densidade elevada de genes e é
geralmente mais acessível para a ativação da transcrição em comparação com a
heterocromatina. A heterocromatina, por outro lado, é caracterizada por atividade de
transcrição baixa e de sequências de DNA repetitivas (RODRIGUES e BJERLIN
2013). A replicação do DNA em células humanas requer a progressão paralela ao
longo do genoma de milhares de mecanismos de replicação. O conhecimento
abrangente de herança genética em diferentes fases de desenvolvimento baseia-se
na elucidação dos mecanismos que regulam a localização e progressão dessas
máquinas por toda a duração da fase de síntese do DNA do ciclo celular (BAKER et
al, 2012).
1.2.17 Função da membrana nuclear e eucromatina
A organização nuclear de organismos eucarióticos é altamente complexa e
dinâmica. Além dos vários componentes estruturais do núcleo, as interações intra e
intercromossômicas, bem como os contatos entre cromatina e componentes
nucleares não é estático, está em constante mudança em resposta a estímulos
ambientais, como a mudança de disponibilidade de nutrientes e processos de
desenvolvimento celular (RODRIGUES e BJERLIN, 2013). Os mesmos autores
relataram que a organização nuclear de eucariontes apresenta uma estrutura
ordenada com conservação evolutiva, abordando diferentes aspectos de como a
organização da cromatina nuclear está associada com a transcrição e replicação
celular.

27
O processo de transcrição depende da membrana nuclear para realização da
saída do RNAm do núcleo para o citoplasma, assim como o transporte de proteínas
dependentes dos poros da membrana nuclear (KATTA et al, 2013). Normalmente,
esses transportes ocorrem através de uma proteína denominada Ran guanosina
trifosfato (Ran GTPase), responsável pela regulação de processos variados, dentre
eles o transporte núcleo-citoplasma, mitose, formação de envelope nuclear e
principalmente a biogênese dos poros da própria membrana nuclear (BIRD et al,
2013). Segundo Seedorf et al (1998), a participação da Ran GTPase no transporte
de proteínas entre o núcleo e o citoplasma é crucial tanto para síntese proteica
quanto para o próprio DNA.
1.2.18 Efeitos do envelhecimento na membrana nuclear e eucromatina
Trabalhos sobre alterações nucleares no envelhecimento são escassos.
Wang et al (2013) encontraram aumento dos marcadores de NF-kβ em núcleos de
células musculares lisas de animais idosos em relação aos jovens. Kiosses e
Kalnins (1993) observaram alterações na cromatina nuclear em células endoteliais
da parede da aorta em coelhos idosos. Segundo Glezer et al (2000), o fator NF-kβ
aumentado no núcleo interage na ativação da RNA polimerase, principalmente em
processos inflamatórios crônicos. Autores como Wood et al (2010) relataram que a
estrutura da cromatina afeta a acessibilidade da transcrição do DNA, seu reparo e
replicação. Segundo os mesmos autores, essas mudanças estruturais observadas
na cromatina ocorrem durante o desenvolvimento, mas pouco se sabe sobre as
alterações durante o envelhecimento.
Autores como Reidel et al (2013) relataram que fatores de transcrição
forkhead (DAF-16/FOXO) estariam diretamente atrelados como mediadores na
extensão da vida, pela remodelação da cromatina dependentes de ATP, que,
segundo os mesmos autores, sofrem mudanças com o estresse e o envelhecimento.
Contudo, como esse mecanismo funciona ainda está longe de ser compreendido.
Reidel et al (2013) vão mais longe quando mencionam que o DAF-16/FOXO abre
uma nova dimensão à forma de como uma alteração de estados da cromatina pode
regular a resposta ao estresse e a longevidade. Qualquer potencial interligado entre
a remodelação da cromatina dependente de ATP e marcas epigenéticas na

28
regulação da vida útil da célula será de suma importância a ser investigado no
futuro, afirmam os autores.

29
2 OBJETIVOS
2.1 OBJETIVO GERAL
O presente trabalho tem como objetivo comparar as diferenças quantitativas
na composição e ultraestrutura dos cardiomiócitos do átrio direito de ratos Wistar em
duas fases da vida desses animais (três e 20 meses de idade), utilizando a
estereologia e métodos imunohistoquimicos.
2.2 OBJETIVOS ESPECÍFICOS
Comparar as diferenças quantitativas dos seguintes elementos dos
cardiomiócitos do átrio direito de ratos Wistar, com três e 20 meses de idade em
relação a (ao):
1- Diâmetro dos cardiomiócitos (em µm);
2- Área de secção do núcleo (em µm2);
3- Densidade de poros da membrana nuclear (número de poros por 10 µm de
membrana nuclear);
4- Densidade numérica dos grânulos de ANP (número por área de 100 µm2);
5- Área dos grânulos de ANP (em µm);
6- Densidade numérica dos grânulos de BNP (número por área de 100 µm2);
7- Área dos grânulos de BNP (em µm);
8- Densidade de volume da eucromatina do núcleo (Vv[eucrom]);
9- Densidade de volume do complexo de Golgi (Vv[Golgi];
10- Densidade de volume das mitocôndrias (Vv[mitoc]),
11- Densidade de volume do retículo endoplasmático (Vv[retíc end];
12- Medida da pressão arterial.

30
3 MATERIAIS E MÉTODOS
3.1 ANIMAIS E COLETA DE MATERIAL
Foram utilizados neste estudo 20 corações de ratos Wistar machos. Dez
animais (Grupo jovem, G1) tinham 3 meses de idade e outros 10 (Grupo idoso, G2)
tinham idade de 20 meses. Os animais foram mantidos em gaiolas coletivas (3 em
cada) em ambiente com temperatura de 23ºc e ciclo claro-escuro controlado de 12
horas. Os ratos tiveram livre acesso à dieta contendo 60% de carboidratos, 30% de
proteínas e 10% de lípides (Nuvilab CR1, Nuvtal nutrientes Ltda, Curitiba, PR) e
receberam água ad libitum. Momentos antes do sacrifício dos animais, os ratos
foram acondicionados em um tubo cilíndrico de acrílico, de maneira adequada para
realização das medidas de pressão arterial sistólica (PA). Para medida da PA, a
cauda dos ratos foi envolvida por manguito de borracha que foi ligado a um
esfigmomanômetro que foi insuflado em intervalos de aproximadamente 50
segundos. Um transdutor de pulso (sensor) capturou os sinais enviados que foram
registrados em computador. Estes procedimentos foram realizados após um período
de adaptação dos animais.O projeto foi submetido e aprovado pelo Comitê de Ética
da Universidade São Judas Tadeu (Protocolo 015/2006).
3.2 SACRIFÍCIO DOS ANIMAIS
Os animais de cada grupo foram eutanasiados, através de decaptação, o
coração foi coletado seccionando-se os vasos da base próxima ao coração e os
átrios direitos foram separados no sulco atrioventricular. De cada átrio, foram feitos
fragmentos com aproximadamente 2 a 3 mm², com lamina nova, dos quais três,
obtidos ao acaso, foram mergulhados em solução fixadora de glutaraldeído a 2,5% e
2% de paraformaldeído, pós-fixados em solução de 1,5 % de tetróxido de ósmio e
emblocados em resina Epon. Posteriormente, de cada bloco foram feitos cortes
semifinos, que foram corados pelo azul de toluidina, os quais foram utilizados para
localizar campos contendo cardiomiócitos em cortes transversais e campos de
cardiomiócitos em cortes longitudinais. Após esse procedimento, os blocos foram
utilizados para fazer cortes ultrafinos que foram corados, coletados em telas e
examinados ao microscópio eletrônico de transmissão (ICB, USP), quando foram

31
capturadas, ao acaso, 10 fotomicrografias, com aumentos de x7500 e dez com
x15000, por animal para exame e realização de estudos qualitativos e quantitativos.
3.3 QUANTIFICAÇÃO DOS COMPONENTES DOS CARDIOMIÓCITOS
Nas fotomicrografias eletrônicas, os cardiomiócitos do átrio direito foram
submetidos a análise quantitativa, utilizando o programa de análise de imagens
computadorizado (Axio Vision, Zeiss) do Laboratório de Estudos Morfoquantitativos
e Imunohistoquímicos da Universidade São Judas Tadeu (LEMI).
O dietro dos cardiomiócitos foi medido em 10 células por animal, seccionadas
transversalmente ao nível do núcleo (Fig. 4a) e a área de secção do núcleo foi
medida em 10 cardiomiócitos por animal, seccionados longitudinalmente ao nível do
núcleo (Fig. 4b).
As densidades de volume ou volumes relativos foram determinadas por
contagem de pontos que tocavam os componentes dos cardiomiócitos de acordo
com a fórmula (Gundersen, 1986; Mayhew, 2008):
Sendo: Vv[Estrutura] = Densidade de volume; P[Estrutura]= Número de pontos sobre a
estrutura em questão e, PT = Número total de pontos do sistema –teste (n=70) (Fig.
5).

32
Figura 3 – Fotomicrografias de cortes de cardiomiócitos do AD mostrando em a os componentes do cardiomiócito que foram quantificados e em b o sistema teste dotado de 70 pontos (em vermelho) sobreposto, utilizado para quantificação dos componentes. M – mitocôndrias, G – aparelho de Golgi, RE – retículo endoplasmático, N – núcleo. Seta preta cheia – poro da membrana do núcleo, setas brancas – grânulos de ANP, setas – grânulos de BNP. Barra: 0.2 µm.
3.4 ANÁLISE ESTATÍSTICA
Os valores obtidos foram tabulados e as médias e erro padrão das médias
dos grupos comparadas para avaliar a significância das diferenças entre eles. Os
dados foram testados para distribuição normal utilizando o teste de Kolmogorov –
Smirnov e o ANOVA de caminho único e o pós-teste t de Student foi utilizado para
comparar os grupos, sendo o nível de significância de 0,05.
33
4 RESULTADOS
4.1 PRESSÃO ARTERIAL E PESO DOS ANIMAIS
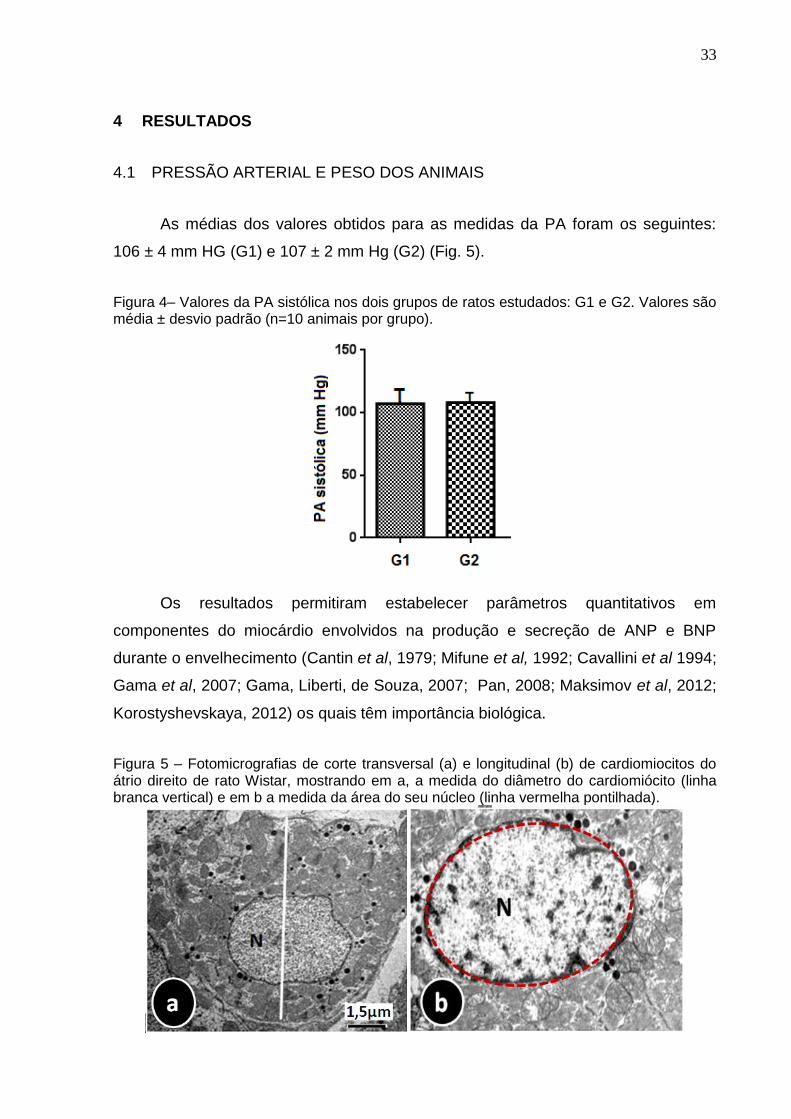
As médias dos valores obtidos para as medidas da PA foram os seguintes:
106 ± 4 mm HG (G1) e 107 ± 2 mm Hg (G2) (Fig. 5).
Figura 4– Valores da PA sistólica nos dois grupos de ratos estudados: G1 e G2. Valores são média ± desvio padrão (n=10 animais por grupo).
Os resultados permitiram estabelecer parâmetros quantitativos em
componentes do miocárdio envolvidos na produção e secreção de ANP e BNP
durante o envelhecimento (Cantin et al, 1979; Mifune et al, 1992; Cavallini et al 1994;
Gama et al, 2007; Gama, Liberti, de Souza, 2007; Pan, 2008; Maksimov et al, 2012;
Korostyshevskaya, 2012) os quais têm importância biológica.
Figura 5 – Fotomicrografias de corte transversal (a) e longitudinal (b) de cardiomiocitos do átrio direito de rato Wistar, mostrando em a, a medida do diâmetro do cardiomiócito (linha branca vertical) e em b a medida da área do seu núcleo (linha vermelha pontilhada).
34
4.2 MORFOLOGIA DOS CARDIOMIÓCITOS
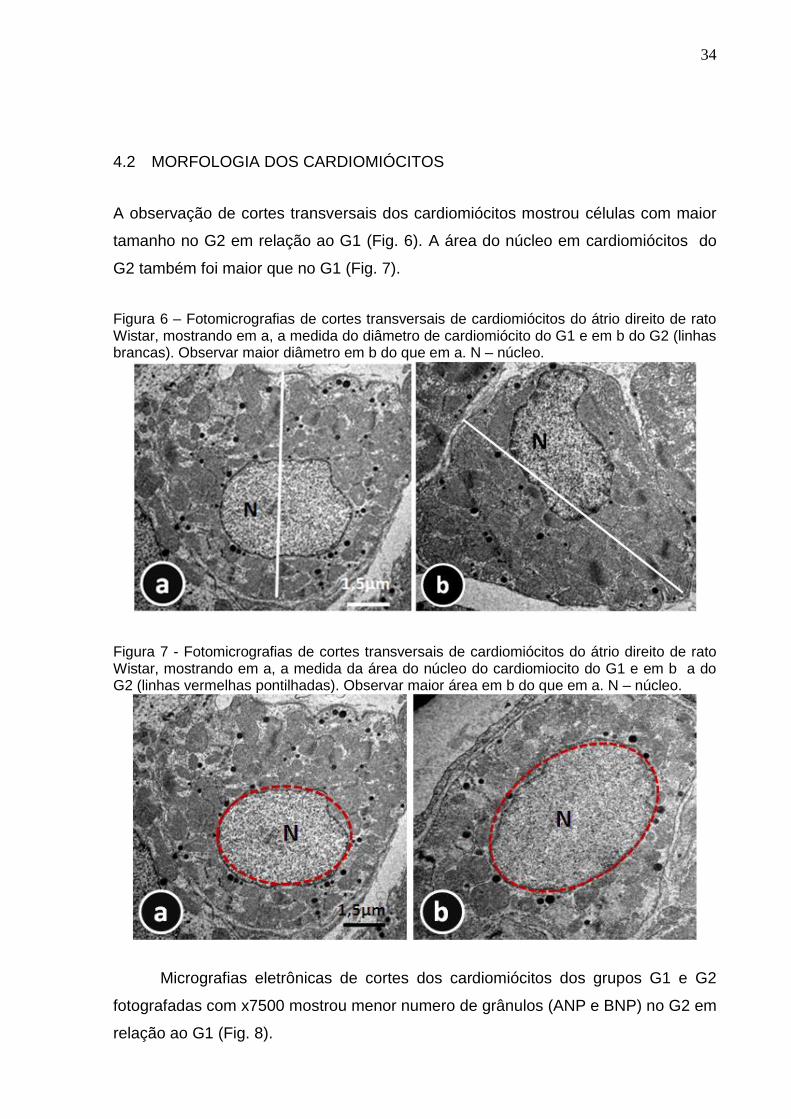
A observação de cortes transversais dos cardiomiócitos mostrou células com maior
tamanho no G2 em relação ao G1 (Fig. 6). A área do núcleo em cardiomiócitos do
G2 também foi maior que no G1 (Fig. 7).
Figura 6 – Fotomicrografias de cortes transversais de cardiomiócitos do átrio direito de rato Wistar, mostrando em a, a medida do diâmetro de cardiomiócito do G1 e em b do G2 (linhas brancas). Observar maior diâmetro em b do que em a. N – núcleo.
Figura 7 - Fotomicrografias de cortes transversais de cardiomiócitos do átrio direito de rato Wistar, mostrando em a, a medida da área do núcleo do cardiomiocito do G1 e em b a do G2 (linhas vermelhas pontilhadas). Observar maior área em b do que em a. N – núcleo.
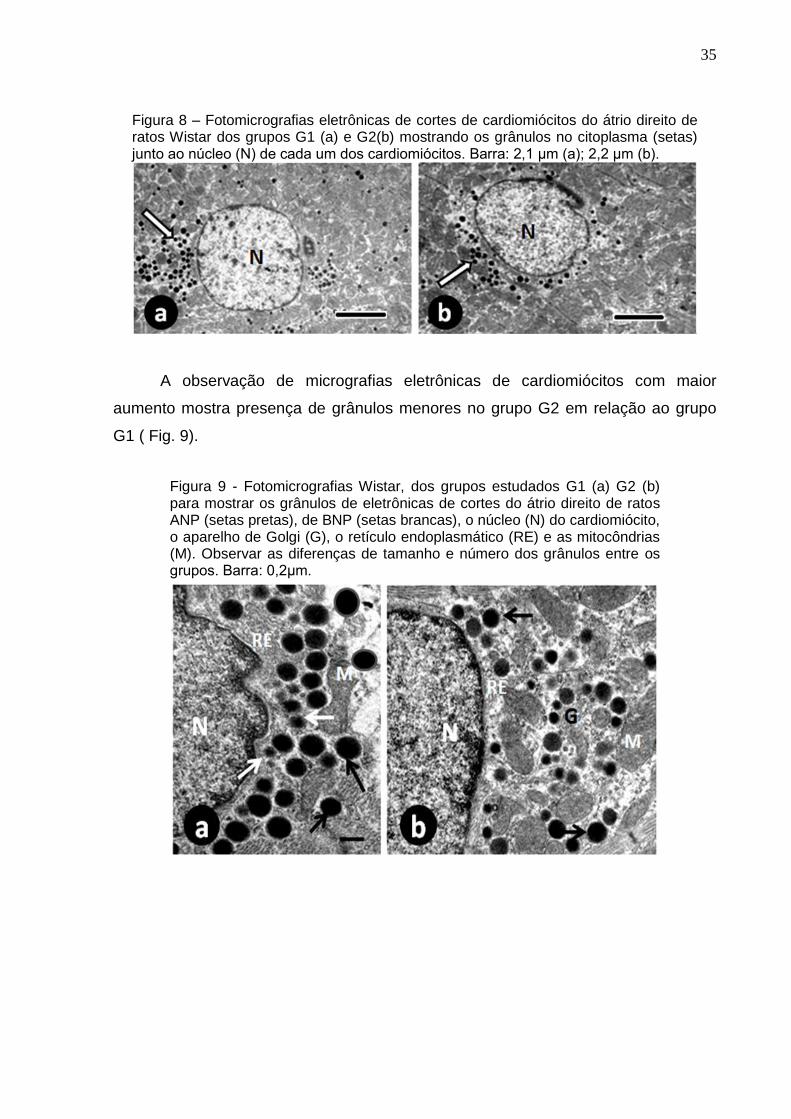
Micrografias eletrônicas de cortes dos cardiomiócitos dos grupos G1 e G2
fotografadas com x7500 mostrou menor numero de grânulos (ANP e BNP) no G2 em
relação ao G1 (Fig. 8).
35
Figura 8 – Fotomicrografias eletrônicas de cortes de cardiomiócitos do átrio direito de ratos Wistar dos grupos G1 (a) e G2(b) mostrando os grânulos no citoplasma (setas) junto ao núcleo (N) de cada um dos cardiomiócitos. Barra: 2,1 μm (a); 2,2 μm (b).
A observação de micrografias eletrônicas de cardiomiócitos com maior
aumento mostra presença de grânulos menores no grupo G2 em relação ao grupo
G1 ( Fig. 9).
Figura 9 - Fotomicrografias Wistar, dos grupos estudados G1 (a) G2 (b) para mostrar os grânulos de eletrônicas de cortes do átrio direito de ratos ANP (setas pretas), de BNP (setas brancas), o núcleo (N) do cardiomiócito, o aparelho de Golgi (G), o retículo endoplasmático (RE) e as mitocôndrias (M). Observar as diferenças de tamanho e número dos grânulos entre os grupos. Barra: 0,2μm.
36
4.3 ANÁLISE QUANTITATIVA
Os resultados quantitativos obtidos mostram diferenças que foram
estatisticamente significantes entre os dois grupos.
4.3.1 Índices nucleicos
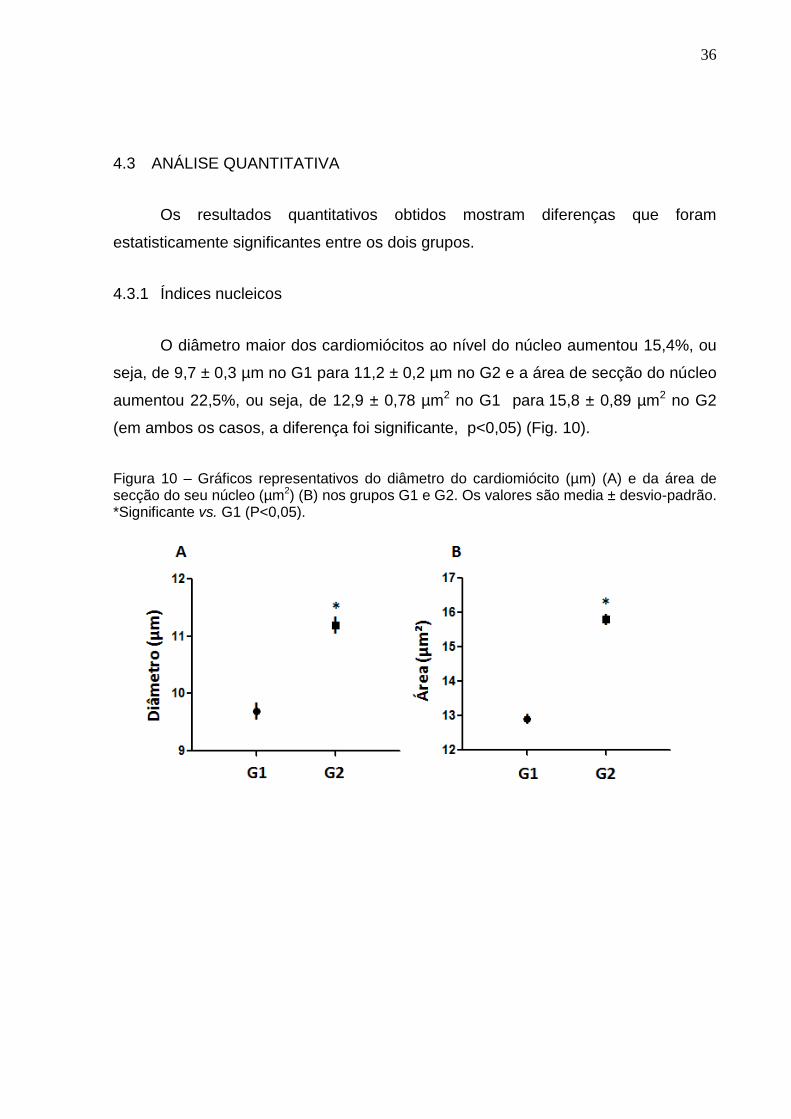
O diâmetro maior dos cardiomiócitos ao nível do núcleo aumentou 15,4%, ou
seja, de 9,7 ± 0,3 µm no G1 para 11,2 ± 0,2 µm no G2 e a área de secção do núcleo
aumentou 22,5%, ou seja, de 12,9 ± 0,78 µm2 no G1 para 15,8 ± 0,89 µm2 no G2
(em ambos os casos, a diferença foi significante, p<0,05) (Fig. 10).
Figura 10 – Gráficos representativos do diâmetro do cardiomiócito (µm) (A) e da área de secção do seu núcleo (µm2) (B) nos grupos G1 e G2. Os valores são media ± desvio-padrão. *Significante vs. G1 (P<0,05).
37
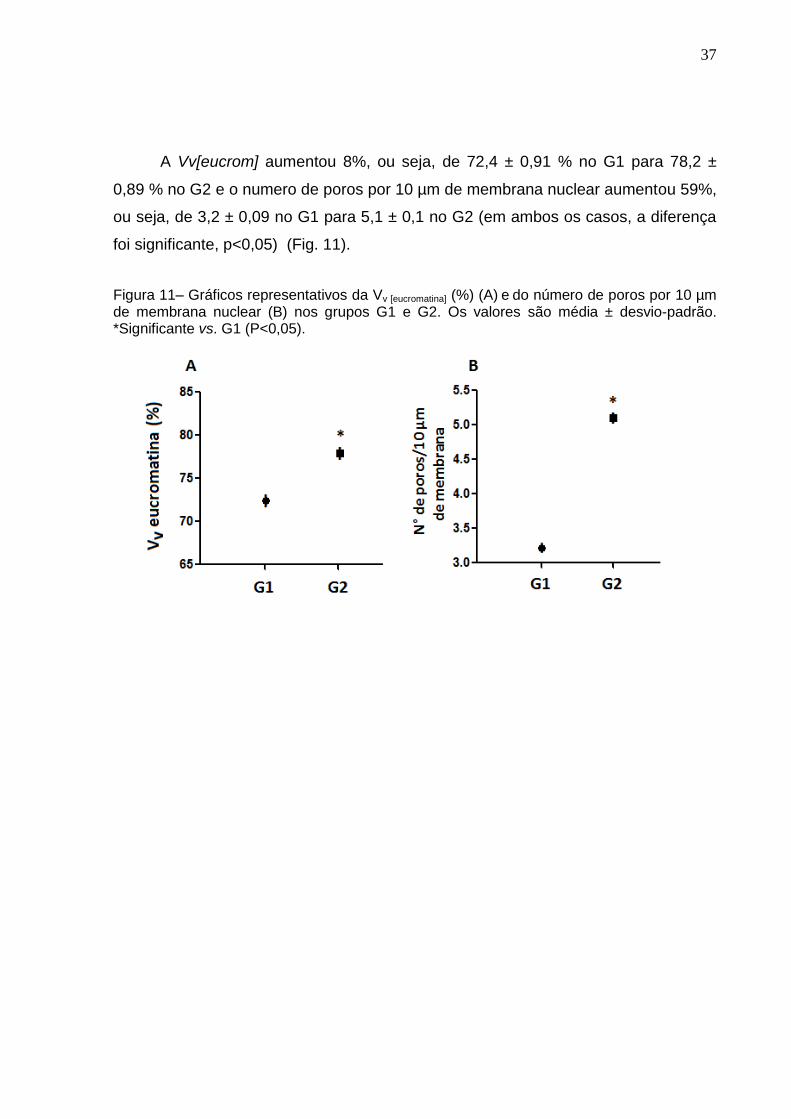
A Vv[eucrom] aumentou 8%, ou seja, de 72,4 ± 0,91 % no G1 para 78,2 ±
0,89 % no G2 e o numero de poros por 10 µm de membrana nuclear aumentou 59%,
ou seja, de 3,2 ± 0,09 no G1 para 5,1 ± 0,1 no G2 (em ambos os casos, a diferença
foi significante, p<0,05) (Fig. 11).
Figura 11– Gráficos representativos da Vv [eucromatina] (%) (A) e do número de poros por 10 µm de membrana nuclear (B) nos grupos G1 e G2. Os valores são média ± desvio-padrão. *Significante vs. G1 (P<0,05).
38
4.3.2 Índices citoplasmáticos
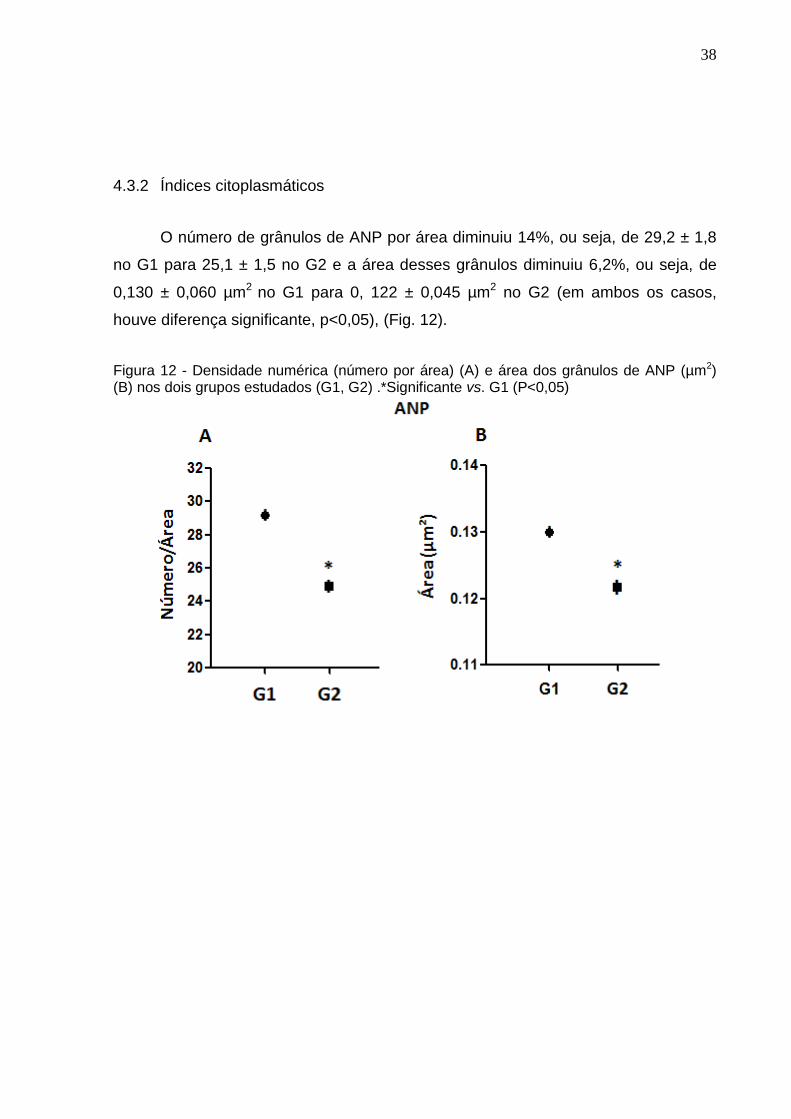
O número de grânulos de ANP por área diminuiu 14%, ou seja, de 29,2 ± 1,8
no G1 para 25,1 ± 1,5 no G2 e a área desses grânulos diminuiu 6,2%, ou seja, de
0,130 ± 0,060 µm2 no G1 para 0, 122 ± 0,045 µm2 no G2 (em ambos os casos,
houve diferença significante, p<0,05), (Fig. 12).
Figura 12 - Densidade numérica (número por área) (A) e área dos grânulos de ANP (µm2) (B) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05)
39
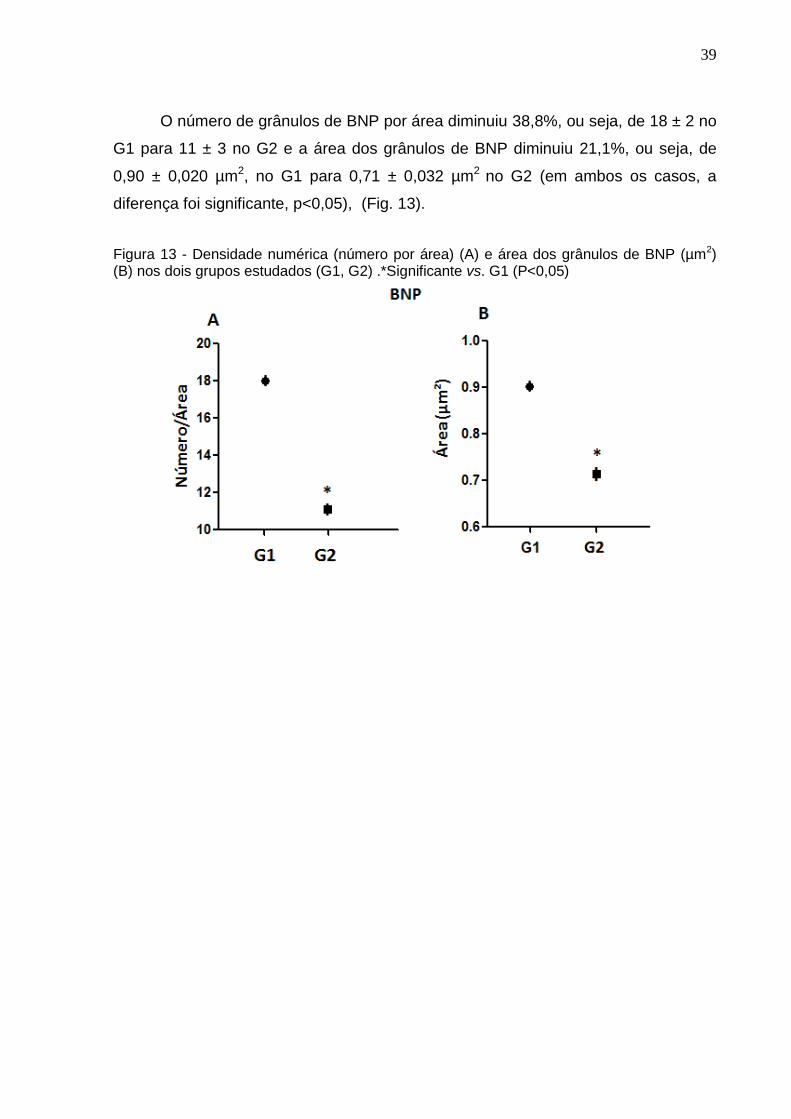
O número de grânulos de BNP por área diminuiu 38,8%, ou seja, de 18 ± 2 no
G1 para 11 ± 3 no G2 e a área dos grânulos de BNP diminuiu 21,1%, ou seja, de
0,90 ± 0,020 µm2, no G1 para 0,71 ± 0,032 µm2 no G2 (em ambos os casos, a
diferença foi significante, p<0,05), (Fig. 13).
Figura 13 - Densidade numérica (número por área) (A) e área dos grânulos de BNP (µm2) (B) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05)
40
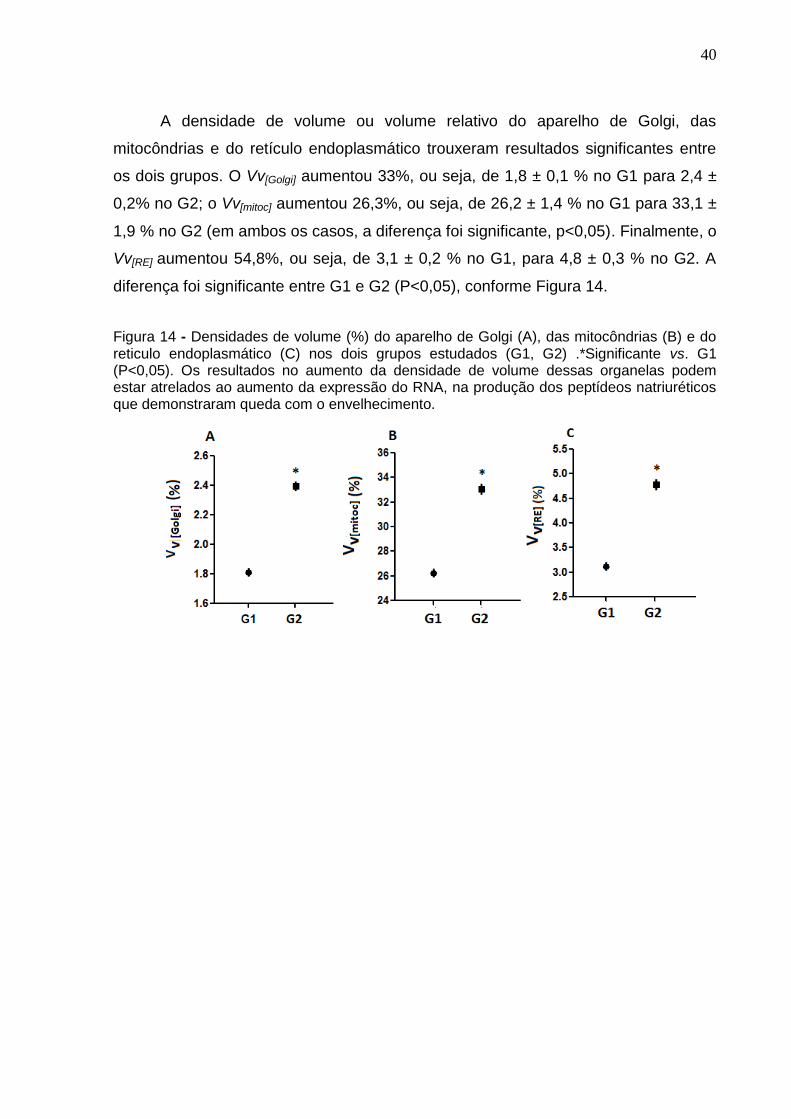
A densidade de volume ou volume relativo do aparelho de Golgi, das
mitocôndrias e do retículo endoplasmático trouxeram resultados significantes entre
os dois grupos. O Vv[Golgi] aumentou 33%, ou seja, de 1,8 ± 0,1 % no G1 para 2,4 ±
0,2% no G2; o Vv[mitoc] aumentou 26,3%, ou seja, de 26,2 ± 1,4 % no G1 para 33,1 ±
1,9 % no G2 (em ambos os casos, a diferença foi significante, p<0,05). Finalmente, o
Vv[RE] aumentou 54,8%, ou seja, de 3,1 ± 0,2 % no G1, para 4,8 ± 0,3 % no G2. A
diferença foi significante entre G1 e G2 (P<0,05), conforme Figura 14.
Figura 14 - Densidades de volume (%) do aparelho de Golgi (A), das mitocôndrias (B) e do reticulo endoplasmático (C) nos dois grupos estudados (G1, G2) .*Significante vs. G1 (P<0,05). Os resultados no aumento da densidade de volume dessas organelas podem estar atrelados ao aumento da expressão do RNA, na produção dos peptídeos natriuréticos que demonstraram queda com o envelhecimento.
41
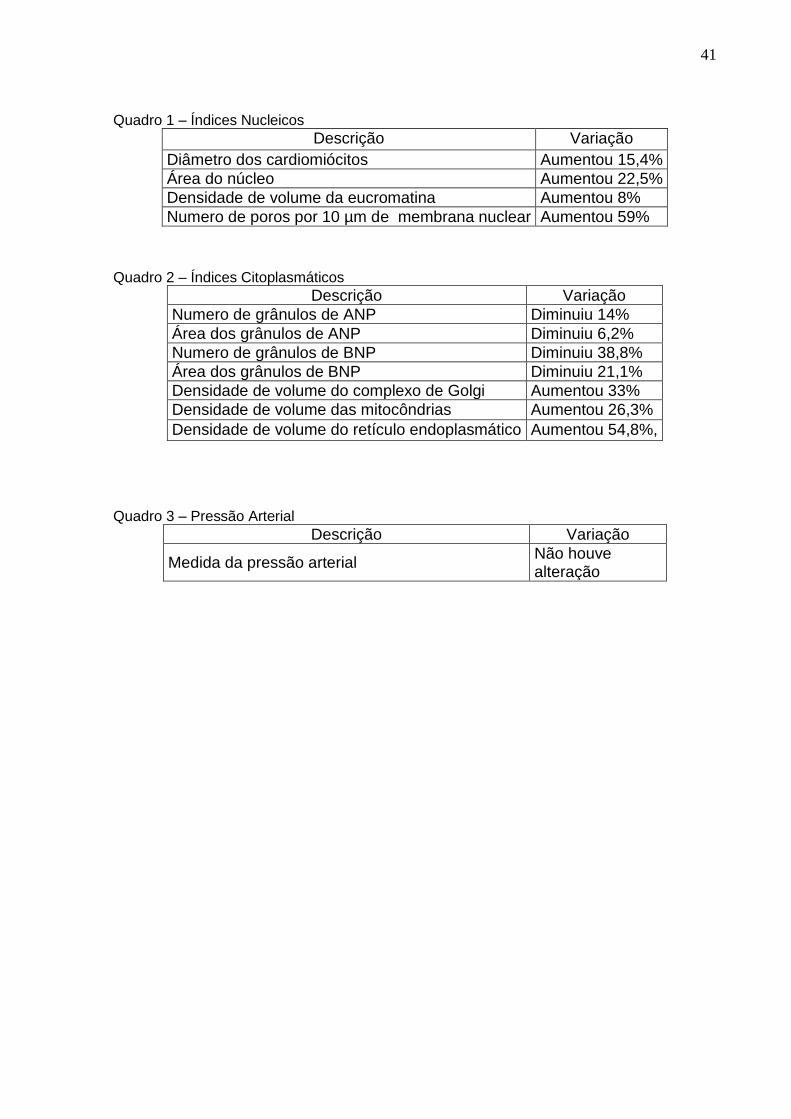
Quadro 1 – Índices Nucleicos
Descrição Variação
Diâmetro dos cardiomiócitos Aumentou 15,4%
Área do núcleo Aumentou 22,5%
Densidade de volume da eucromatina Aumentou 8%
Numero de poros por 10 µm de membrana nuclear Aumentou 59%
Quadro 2 – Índices Citoplasmáticos
Descrição Variação
Numero de grânulos de ANP Diminuiu 14%
Área dos grânulos de ANP Diminuiu 6,2%
Numero de grânulos de BNP Diminuiu 38,8%
Área dos grânulos de BNP Diminuiu 21,1%
Densidade de volume do complexo de Golgi Aumentou 33%
Densidade de volume das mitocôndrias Aumentou 26,3%
Densidade de volume do retículo endoplasmático Aumentou 54,8%,
Quadro 3 – Pressão Arterial
Descrição Variação
Medida da pressão arterial Não houve alteração

42
DISCUSSÃO
As doenças cardíacas ainda são a principal causa de morte no mundo,
matando duas vezes mais que o câncer, chegando a 17,3 milhões de mortes anuais,
segundo dados da Organização Mundial da Saúde (OMS) de 2011. De acordo com
a mesma OMS, a hipertensão arterial tem liderado esse quadro dentre as demais
patologias cardíacas, segundo essa estatística. Por esse motivo, cada vez mais o
controle da pressão arterial se tornou algo comum nos consultórios médicos.
Problemas de estresse crônico, alimentação errada, falta de sono, são só algumas
das causas da elevação da pressão arterial e desequilíbrio na função cardiovascular,
principalmente em idosos. Segundo Cannonne et al (2011), tais fatores podem
interferir com a função dos cardiomiócitos e promover consequências graves no
decorrer do envelhecimento. Sabemos que as patologias como infarto agudo do
miocárdio, derrame e aneurisma podem estar diretamente atreladas ao desequilíbrio
da pressão arterial (ZHAO, et al 2010). Essa tendência é agravar-se ainda mais com
a idade, pois os seres humanos estão cada vez mais submetidos às pressões
sociais, desencadeando aumentos consideráveis nos níveis diários de estresse e
consequentemente alterações cardíacas.
Desta forma, o presente estudo sobre os efeitos do envelhecimento nos
componentes dos cardiomiócitos torna-se necessário para a compreensão das
possíveis alterações sobre o funcionamento dos peptídeos natriurético e as
respectivas organelas referentes à fisiologia desse tecido. Sabemos que os
componentes nuclear e citoplasmático sofrem alterações com a idade em diferentes
tipos de células, ocorrendo seu aumento nas fases mais precoces da vida (MISAKA
et al, 2013). Contudo, há necessidade de mais estudos relacionados ao
envelhecimento desses componentes, principalmente no tecido cardíaco. Essas e
outras razões motivaram o objetivo de avaliar os componentes dos cardiomiócitos,
através a estereologia, para verificar as alterações nesses componentes no
envelhecimento.
De acordo com os resultados deste trabalho, a análise morfoquantitativa dos
cardiomiócitos jovens e idosos do átrio direito demonstrou aumento significante no
seu diâmetro no grupo idoso em relação ao grupo jovem. Este resultado vem
corroborar os resultados do trabalho de Zhang et al (2007), porém realizado no
ventrículo, onde foi demonstrado que, com o avançar da idade, houve hipertrofia nos

43
cardiomiócitos ventriculares. Para Yang et al (2012), a causa desse fenômeno é a
redução do número de células decorrente do próprio envelhecimento,
desencadeando a hipertrofia das células cardíacas remanescentes e aumentando a
espessura do miocárdio para manter sua força de ejeção. Zhao et al (2010)
relataram que a hipertrofia não é desencadeada somente pelo envelhecimento, mas
também pelo aumento da pressão arterial, comum em faixas etárias mais
avançadas.
Ao que parece, o cardiomiócito procura manter a homeostase para não
comprometer a fisiologia de bombeamento sanguíneo para os demais tecidos do
corpo e, com isso, ao envelhecer, seus componentes sofrem alterações
ultraestruturais, tanto nucleares quanto citoplasmáticas, para adaptações fisiológicas
decorrentes do próprio envelhecimento.
Da mesma forma, os resultados do presente trabalho referente à área do
núcleo e a densidade da eucromatina também demonstraram aumentos significante
no grupo idoso, quando comparado ao grupo jovem. Os poros da membrana nuclear
também demonstraram aumento na quantidade de poros no grupo idoso em relação
ao grupo jovem. Trabalhos sobre alterações nucleares no envelhecimento são
escassos. Embora Wang et al (2013) tenham encontrado aumento dos marcadores
de NF-kβ em núcleos de células musculares lisas de animais idosos em relação aos
jovens, não podemos afirmar que o aumento da densidade de volume da
eucromatina estaria diretamente atrelado a esse fato, mas indiretamente sim. No
trabalho de Glezer et al (2000), os autores demonstraram que o fator NF-kβ,
aumentado no núcleo, interage na ativação da RNA polimerase, principalmente em
processos inflamatórios crônicos e no envelhecimento, evidenciando uma maior
atividade da cromatina nesses casos. Contudo, autores como Wood et al (2010)
relataram que a estrutura da cromatina afeta a acessibilidade da transcrição do
DNA, demonstrando que essas mudanças estruturais observadas na cromatina
ocorrem durante o desenvolvimento. Entretanto, pouco se sabe sobre essas
alterações durante o envelhecimento. De acordo com Glezer et al (2000), o NF-kβ
aumentado no núcleo aumenta a expressão da RNA polimerase podendo ser um
fator determinante da densidade da eucromatina avaliada no presente trabalho.
As alterações dos componentes nucleares e citoplasmáticos parece estarem
atreladas e interdependentes no processo de senescência, como podemos
evidenciar pelos resultados deste trabalho em relação à produção de ANP e BNP.

44
Em relação aos componentes citoplasmáticos, o presente trabalho
demonstrou uma diminuição significante tanto nos grânulos de ANP quanto nos
grânulos de BNP no grupo idoso em relação ao grupo jovem, corroborando com os
resultados do trabalho de Wu et al (2008), quando os autores relatam uma
diminuição significante de ANP em ratos idosos em relação aos jovens utilizando a
mesma técnica de estereologia. Entretanto, Cavallini et al (1994) não encontraram
alterações significantes entre as concentrações de ANP em relação aos ratos jovens
e idosos, também utilizando a estereologia. Nenhum dos dois autores mencionou o
BNP. É possível que essas diferenças de resultados sejam devidas à metodologia
utilizada pelos autores.
Da mesma forma que o número de grânulos apresentou alterações entre os
grupos, os resultados do presente estudo mostraram que o diâmetro dos grânulos
de ANP e BNP foi significantemente menor no grupo idoso em relação ao jovem.
Esses resultados concordam com os do trabalho de Miffune et al (1991), que
mostrou diminuição significante no diâmetro dos grânulos de ANP no grupo idoso
quando comparado ao jovem. Contudo, esses autores também nada mencionaram
sobre o BNP.
Apesar de Gama et al (2007) terem demonstrado que a distribuição
quantitativa dos grânulos nos átrios direito e esquerdo é diferente, com maior
presença de grânulos no átrio direito, não há evidências em outros estudos que
corroborem essa afirmativa. Pelo contrário, Cavallini et al (1994) demonstraram que
não há alterações significantes entre a quantidade de ANP nos átrios direito e
esquerdo tanto em ratos jovens, como em adultos e idosos.
Ultimamente, novas evidências apontam para outras funções importantes
para o ANP. Assim por exemplo, o ANP tem chamado a atenção de pesquisadores
com relação ao processo de envelhecimento relacionado aos ritmos biológicos
(CHARLOUX et al, 2007). Afzal (2011) afirmou que níveis plasmáticos muito
elevados de ANP e BNP produzidos pelo miocárdio são consequências diretas de
causas de falência cardíaca.
A importância das pesquisas com o ANP fica demonstrada quando avaliamos
as principais causas de morte em todo o mundo. Cannone et al (2011)
demonstraram, num trabalho de avaliação genética das doenças cardiovasculares,
que a suscetibiliade para doenças cardiometabólicas apoiam o possível papel
protetor dos peptídeos natriuréticos por seus efeitos favoráveis sobre a função

45
metabólica. Chen et al (2011) foram mais longe ao relatarem que os peptídeos
natriuréticos são os novos candidatos a terapias contra doenças cardiorrenais,
demonstrando, em seus resultados, que terapias com peptídeos natriuréticos
constituem um fato no controle de doenças cardíacas.
Apesar de, no presente trabalho, tanto o ANP quanto o BNP apresentarem
diminuição significante tanto no número quanto no diâmetro em idosos, as demais
organelas citoplasmáticas demonstraram o oposto. A densidade de volume do
aparelho de Golgi aumentou de forma significante no grupo idoso em relação ao
grupo jovem no átrio direito. O mesmo ocorreu também nas mitocôndrias e no
retículo endoplasmático, relatado neste trabalho.
No trabalho de Ispolatov e Musch (2013), os autores relataram perda de
proteínas nas cisternas no aparelho de Golgi, que especifica diminuição na fusão
das vesículas à medida que essas organelas envelhecem. Porém, quando os
mesmos autores demonstraram analiticamente que a concentração de proteínas que
regulam a fusão dessas organelas decai exponencialmente com o número de
cisternas, parece que, ao envelhecer, o aparelho de Golgi aumenta o volume de
suas cisternas em decorrência da diminuição das proteínas que regulam sua fusão
e/ou vice-versa. Contudo, não podemos fazer nenhuma afirmação em relação a isso,
mas apenas relatar uma coincidência entre os resultados observados, pois, como os
autores mesmo mencionaram, o mecanismo de como isso funciona ainda é
totalmente obscuro, carecendo de mais estudos para se chegar a alguma conclusão
sobre essa organela.
Nas mitocôndrias, o aumento da densidade de volume por cardiomiócito
observado no presente trabalho pode corroborar os resultados dos estudos de
Preston et al (2008) e Peterson, Johansen e Ravussin (2012). Os primeiros autores
relataram uma diminuição na função energética nessas organelas em ratos idosos,
principalmente na codificação dos genes da cadeia respiratória, diminuição da
atividade das enzimas ATP sintase, NADH desidrogenase e capacidade diminuída
na utilização do oxigênio na síntese de ATP quando comparados a ratos jovens. Os
segundos evidenciaram um aumento na apoptose em tecidos cardiacos. Ambos os
autores ainda relataram um aumento do número de mitocondrias nos cardiomiócitos
nos ratos idosos em relação aos jovens. Esse fato os levou a crer que uma
compensação pela deficiencia energética na diminuição da produção de ATP pode
ter provocado uma forma adaptativa na manutenção das funções do músculo

46
cardíaco. Acreditamos que essas adaptações fisiológicas possam também ter
ocorrido neste presente trabalho, quando a densidade de volume das mitocôndrias
apresentou um resultado maior no grupo idoso do que no grupo jovem.
Como o músculo cardíaco necessita de grandes quantidades de energia, as
mitocôndrias velhas perdem a capacidade de gerar grandes quantidades de ATP
(LEE et al, 2012), ficando evidente a necessidade de aumento do número dessas
organelas por compensação para o correto funcionamento fisiológico do
cardiomiócito, uma vez que, os ratos utilizados nos dois grupos eram saudáveis.
Segundo Jeong et al (2012), quando o número ou a atividade mitocondrial é
insuficiente, o corpo humano entra rapidamente em fadiga pela deficiência do
próprio ATP. Por outro lado, de acordo com Cali et al (2012), quando o número,
volume, forma e distribuição das mitocôndrias, no interior das células, é bem
controlado, as células conseguem entrar facilmente em homeostase. As adaptações
mitocondriais podem estar atreladas a uma cascata bioquímica que se inicia desde o
núcleo até a completa biogênese da própria organela (SAFDAR et al, 2011).
Quando Reidel et al (2013) relataram sobre os fatores de transcrição DAF-
16/FOXO estarem diretamente atrelados como mediadores na extensão da vida,
pela remodelação da cromatina dependentes de ATP, parece que estamos diante de
uma possível adaptação referente ao aumento da densidade de volume tanto da
eucromatina como das mitocôndrias, apresentado não só neste presente trabalho
mas também nos trabalhos de Preston et al (2008), sobre mitocôndrias e Glezer et al
(2000), sobre fator de transcrição NF-kβ e cromatina. Apesar de Reidel et al (2013)
relatarem que a forma de como este mecanismo funciona ainda está longe de ser
compreendida, os autores mencionaram que este fator de transcrição abre uma
nova dimensão à forma de como uma alteração de estado da cromatina pode
regular a resposta ao estresse e à longevidade, pois qualquer remodelação da
cromatina é dependente de ATP. Os autores ainda acreditam que uma deficiência
na produção de ATP, produzido pelas mitocôndrias, poderia provocar alterações na
densidade da própria cromatina.
Assim como as demais organelas citoplasmáticas dos cardiomiócitos no
presente estudo demonstraram aumento na densidade de volume, o retículo
endoplasmático também apresentou aumento na sua densidade de volume no grupo
idoso quando comparado ao grupo jovem. Assim sendo, Tanjore et al (2013)
relataram que a capacidade do retículo endoplasmático em se manter sob condições

47
de estresse diminui com o envelhecimento, o que pode contribuir para que a
incidência dos distúrbios fibróticos cresçam com a idade. Entretanto, Babusikowa et
al (2012) relataram que, embora a atividade da enzima cálcio-ATPase diminuisse
com o avançar da idade, não foram observadas diferenças nas quantidades relativas
de proteínas relacionadas ao retículo endoplasmático. Por outro lado, a acumulação
significativa de dano oxidativo em proteína ocorreu com o envelhecimento. Os
resultados deste estudo fizeram os autores relatarem que a alteração relacionada à
idade na atividade da enzima cálcio-ATPase no coração de rato não é uma
consequência da diminuição dos níveis de proteína dessa organela, mas pode surgir
a partir de modificações oxidativas de proteínas do próprio retículo endoplasmático.
Danos oxidativos celulares causados por espécies reativas de oxigênio poderiam
contribuir para alternâncias relacionadas à idade no relaxamento do miocárdio.
Ainda segundo esses autores, alterações mofológicas no reticulo não foram
observadas em ratos idosos quando comparados aos jovens.
Porém, em outro estudo, autores como Chen et al (2013) relataram que o
estresse do retículo endoplasmático, sob quaisquer condições prejudiciais, inclusive
o envelhecimento, pode levar à diminuição da síntese de proteínas e,
consequentemente, ao aumento da expressão das chaperonas moleculares, as
quais promovem o dobramento adequado das proteínas na recuperação das células,
aumentando suas concentrações no interior dessa organela. Portanto, talvez o
resultado no aumento da densidade de volume dessa organela, observado no
presente trabalho, possa ser um reflexo do aumento das chaperonas no interior do
próprio retículo endoplasmático. Contudo, seriam necessários estudos mais
especificos para verificar como esse mecanismo realmente funciona.
O aumento da densidade de volume das organelas, o aumento do diâmetro
do cardiomiócito e da área do núcleo, parece ser uma adequação na busca da
homeostase desse tipo de tecido. O envelhecimento dos cardiomiócitos pode levar a
um aumento da expressão do RNAm, para adequar as proteínas nesses tecidos
quando estes se encontram envelhecidos. Portanto, como foi demonstrado no
presente trabalho, ao envelhecer, os peptídeos natriuréticos nos cardiomiócitos
diminuem, assim como seu diâmetro. Desta forma, por esses hormônios serem
derivados de proteínas, acreditamos que a carência da adequação das organelas
em sintetizar proteínas faça com que a densidade das mesmas aumente em
consequência disso. Contudo, para confirmar essa possibilidade, seria necessária

48
uma investigação no perfil metabólico dos cardiomiócitos junto a um novo estudo
morfológico utilizando a estereologia. Com isso, poderíamos chegar a um desfecho
mais plausível sobre os resultados que obtivemos no presente estudo, relatando o
porquê das alterações morfológicas nesse tipo de tecido decorrentes do
envelhecimento.

49
CONCLUSÃO
A análise dos resultados apresentados no presente trabalho permite concluir
que:
1- A pressão arterial se manteve inalterada com o envelhecimento.
2- Os cardiomiócitos de ratos idosos, quando examinados pela estereologia,
mostram alterações morfológicas e ultraestrutuais significantes.
3- Os grânulos de ANP e BNP demonstram significante queda, tanto no número
quanto no diâmetro no grupo idoso quando comparado ao jovem.
4- Componentes citoplasmáticos, como aparelho de Golgi, mitocôndrias e retículo
endoplasmático, demonstram aumento significante na densidade de volume no
grupo idoso em relação ao grupo jovem.
5- O diâmetro dos cardiomiócitos, a área do núcleo e a densidade da eucromatina
aumentam significantemente com o envelhecimento.
Portanto, houve uma aumento na hipertrofia dos átrios nos cardiomiócitos com
redução da sua função endócrina.

50
REFERÊNCIAS
ABASSI Z; WEISSMAN I., KARRAM T., GOLTSMAN I., HOFFMAN A., BETTER O.S., WINAVER J. Restoration of renal responsiveness to atrial natriuretic Peptide in experimental nephrotic syndrome by albumin infusion. Am J Nephrol. 2013;38(4):292-9. AGUILA M.B., RODRIGUES-APFEL M.I., MANDARIM-DE-LACERDA C.A. Stereology of the myocardium and blood biochemistry in aged rats fed with a cholesterol-rich and canola oil diet (n-3 fatty acid rich).; Basic Res Cardiol. 1998 Jun;93(3):182-91. AMARILIO R., RAMACHANDRAN S., SABANAY H., LEV S. Differential Regulation of Endoplasmic Reticulum Structure through VAP-Nir Protein Interaction..J Biol Chem 2005 18;280(7):5934-44. BABUŠÍ OVÁ E., LEHOTSKÝ J., DOBROTA D., RAČAY P., KAPLÁN P. Age-associated changes in Ca(2+)-ATPase and oxidative damage in sarcoplasmic reticulum of rat heart. Physiol Res. 2012 Dec 14;61(5):453-60. BAETZ N.W., STAMER W.D., YOOL A.J. Stimulation of aquaporin-mediated fluid transport by cyclic GMP in human retinal pigment epithelium in vitro. Invest Ophthalmol Vis Sci. 2012 Apr 24;53(4):2127-32. BAKER A., AUDIT B., CHEN C., MOINDROT B., LELEU A., GUILBAUD, G. RAPPAILLES A., VAILLANT C., GOLDAR A., MONGELARD F., D'AUBENTON-CARAFA Y., HYRIEN O., THERMES C., ARNEODO A. Replication Fork Polarity Gradients Revealed by Megabase-Sized U-Shaped Replication Timing Domains in Human Cell Lines. PLoS Comput Biol. 2012; 8(4): e1002443. BEGG D.P., SINCLAIR A.J., WEISINGER R.S. Reductions in water and sodium intake by aged male and female rats. Nutr Res.; 2012 Nov;32(11):865-72. BIRD S. L., HEALD R., WEIS K. RanGTP and CLASP1 cooperate to position the mitotic spindle. Mol Biol Cell. 2013; 15; 24(16): 2506–2514. CALIARI M., V., TAFURI W. L.. O coração endócrino. A descoberta do peptídeo natriurético atrial. Arq Bras Cardiol; 1993 61(5): 283-286. CANNONE V., BOERRIGTER G., CATALIOTTI A., COSTELO-BOERRIGTER L. C., OLSON T. M., McKIE P. M., HEUBLEIN D. M., LAHR D. B., BAILEY K. R., AVERNA M.; REDFIELD M. M., RODEHEFF R. J., BOURNETT J. C.. A Genetic Variant of the Atrial Natriuretic Peptide Gene Is Associated With Cardiometabolic Protection in the General Community. J Am Coll Cardiol 2011 vol 58, nº 6. 629-36. CAVALLINI G., CLERICO A., DEL CHICCA M., GORI Z., BERGAMINI E. Changes in endocrine atrial rat cardiocytes during growth and aging: an ultrastructural, morphometric and endocrinological study. Aging. 1994 Jun;6(3):167-74.
http://www.ncbi.nlm.nih.gov/pubmed?term=Ra%C4%8Day%20P%5BAuthor%5D&cauthor=true&cauthor_uid=22881224

51
CHARLOUX A., BRANDENBERGER G., PIQUARD F., GENY B., Desregulation of Pulsative in Aging IV. Pulsative Signaling and Cardiovascular Aging: Functions and Regulation of Natriuretic Peptide Signaling. BP 426, 67091, Dec. (2007). Ageing Res Rev. 2008 Jul;7(3):151-63. CHEN G., GONG M., YAN M., ZHANG X. Sevoflurane induces endoplasmic reticulum stress mediated apoptosis in hippocampal neurons of aging rats. PLoS One 2013;8(2):e57870. CHO K.W., SEUL K.H., KIM S.H., SEUL K.M., KOH G.Y. Atrial pressure, distension, and pacing frequency in ANP secretion in isolated perfused rabbit atria.. Am J Physiol. 1991; 260 (1 Pt 2):R39-46. CLERICO A., ZUCCHELLI G., PILO A., PASSINO C., EMDIN M. Clinical relevance of biological variation: the lesson of brain natriuretic peptide (BNP) and NT-proBNP assay. [email protected] 2006.44(4):366-78. COOK N. L., VIOLA H. M., SHAROV V. S., HOOL L. C., SCHÖNEICH C., DAVIES M. J.Myeloperoxidase-derived oxidants inhibit sarco/endoplasmic reticulum Ca2+-ATPase activity, and perturb Ca2+ homeostasis in human coronary artery endothelial cells. Free Radic Biol Med. 2012, 1; 52(5): 951–961. DE BOLD A.J., BORENSTEIN H.B., VERESS A.T., SONNENBERG H. A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Reprinted from Life Sci. 28:89-94, 1981.. J Am Soc Nephrol. 2001, 12(2):403-9; discussion 403-8, 408-9. DICSTEIN K. Natriuretic peptides in detection of heart failure. Lancet 1998; 35: 3-4. DICSTEIN K. Atrial fibrillation after cardiac surgery. Ann Intern Med. 2001; 135: 1061-73. EHARA S., NAKAMURA Y., MATSUMOTO K., HASEGAWA T., SHIMADA K., TAKAGI M., HANATANI A., IZUMI Y., TERASHIMA M., YOSHIYAMA M. Effects of intravenous atrial natriuretic peptide and nitroglycerin on coronary vasodilation and flow velocity determined using 3 T magnetic resonance imaging in patients with nonischemic heart failure. Heart Vessels. 2013, 28(5):596-605. ELLGAARD L., HELENIUS A. Quality control in the endoplasmic reticulum Molecular cell biology 2003; 4. 181-191. ESPINER E.A., LEIKIS R., FERCH R.D., MACFARLANE M.R., BONKOWSKI J.A., FRAMPTON C.M., RICHARDS A.M. The neuro-cardio-endocrine response to acute subarachnoid haemorrhage. Clin Endocrinol . 2002;56(5):629-35. FEDERICO C. Natriuretic Peptide system and cardiovascular disease.. Heart Views. 2010; 11(1):10-5.
http://www.ncbi.nlm.nih.gov/pubmed?term=de%20Bold%20AJ%5BAuthor%5D&cauthor=true&cauthor_uid=11158233
http://www.ncbi.nlm.nih.gov/pubmed?term=Sonnenberg%20H%5BAuthor%5D&cauthor=true&cauthor_uid=11158233

52
FENZL M., SCHNIZER W., AEBLI N., SCHLEGEL C., VILLIGER B., DISCH A., GREDIG J., ZAUGG T., KREBS J. Release of ANP and fat oxidation in overweight persons during aerobic exercise in water. Int J Sports Med. 2013; 34(9):795-9. FLATO U.A.P., SLHESSARENKO J.R., GUIMARÃES H.P., GOMES R.S., LOPES R.D. Peptídeo natriurético na emergência: quando usar?. Rev Bras Clin Med. 2009; 7: 398-405. GAMA E.F., LIBERTI E.A., DE SOUZA R.R. Effects of pre- and postnatal protein deprivation on atrial natriuretic peptide -(ANP-) granules of the right auricular cardiocytes. An ultrastructural morphometric study. Eur J Nutr. 2007; 46(5):245-50. GERRITY G.R., Cliff W.J. The aortic tunica intima in young and aging rats. Exp Mol Pathol 1972; 16:382:402. GRAFFE C.C., BECH J.N., PEDERSEN E.B. Effect of high and low sodium intake on urinary aquaporin-2 excretion in healthy humans. Am J Physiol Renal Physiol. 2012; 15;302(2):F264-75. ISPOLATOV I., MÜSCH A. A Model for the Self-Organization of Vesicular Flux and Protein Distributions in the Golgi Apparatus. PLoS Comput Biol. 2013 July; 9(7): e1003125. JANCZEWSKI A. M., LAKATTA E. G.Modulation of sarcoplasmic reticulum Ca2+ cycling in systolic and diastolic heart failure associated with aging. Heart Fail Rev. 2010 September; 15(5): 431–445. KAJISTURA J., CHENG W., SARANGARAJAM R. et al. Necrotic and apoptotic myocyte cell death in the aging heart of Fisher 344 hertz. Am J Physiol 1996, 271:H1215-1228. JULIENNE H., ZOUFIR A., AUDITB., ARNEODO A.OUZOUNIS C. A.Human Genome Replication Proceeds through Four Chromatin States. PLoS Comput Biol. 2013; 9(10): e1003233. KIOSSES B.W., KALNINS V.I. Age-related changes in the position of centrosomes in endothelial cells of the rabbit aorta. Exp Gerontol. 1993 Jan-Feb;28(1):69-75. KLAIBER M., DANKWORTH B., KRUSE M., HARTMANN M., NIKOLAEV V.O., YANG R.B., VÖLKER K., GAßNER B., OBERWINKLER H., FEIL R., FREICHEL M., GROSCHNER K., SKRYABIN B.V., FRANTZ S., BIRNBAUMER L., PONGS O., KUHN M. A cardiac pathway of cyclic GMP-independent signaling of guanylyl cyclase A, the receptor for atrial natriuretic peptide. Proc Natl Acad Sci U S A. 2011; 8;108(45):18500-5. LADINSKY M. S., MASTRONARDE D.N., MCINTOSH J. R., HOWELL K. E., STAEHELIN L. A. Golgi Structure in Three Dimensions: Functional Insights from the Normal Rat Kidney Cell. The Journal of Cell Biology; 144, 6, 22, 1999 1135–1149.

53
LAN L., NAKAJIMA S., WEI L., SUN L., HSIEH C.L., SOBOL R.W., BRUCHEZ M., VAN HOUTEN B., YASUI A., LEVINE A.S.Novel method for site-specific induction of oxidative DNA damage reveals differences in recruitment of repair proteins to heterochromatin and euchromatin. Nucleic Acids Res. 2013; 29. [Epub ahead of print] LEE J.S., PARK A.H., LEE S.H., LEE S.H., KIM J.H., YANG S.J., YEOM Y.I., KWAK T.H., LEE D., LEE S.J., LEE C.H., KIM J.M., KIM D. Beta-lapachone, a modulator of NAD metabolism, prevents health declines in aged mice. PLoS One. 2012;7(10):E47122. LIU Q., ZHANG J., XU Y., HUANG Y., WU C. Effect of carvedilol on cardiomyocyte apoptosis in a rat model of myocardial infarction: A role for toll-like receptor 4. Indian J Pharmacol. 2013 Sep;45(5):458-63. MAEDA K., TAKAYOSHI T., WADA A, HISANAGA T., KINOSHITA M. Plasma brain natriuretic peptide as a biochemical marker of high left ventricular end-diastolic pressure in patients with symptomatic left ventricular dysfunction. Am Heart J. 1998; 135: 825-32. MAKSIMOV V. F., KOROSTYSHEVSKAYA I. M., MARKEL' A. L., SHMERLING, M. D. YAKOBSON, G. S. Structural characteristics of cardiomyocytes in the right atrium of NISAG rats. Bull Exp Biol Med. 2004 Jul;138(1):1-4. MARTÍNEZ-ALONSO E., TOMÁS M., e MARTÍNEZ-MENÁRGUEZ J.A. Morpho-Functional Architecture of the Golgi Complex of Neuroendocrine Cells. Front Endocrinol. 2013; 4: 41. McGRATH M.F., DE BOLD M.L., DE BOLD A.J. The endocrine function of the heart. 2005; 16(10):469-77. Epub 2005. MIFUNE H., SUZUKI S., NODA Y., HAYASHI Y., MOCHIZUKI K. Atrial natriuretic peptide (ANP)-granules of auricular cardiocytes in aging Mongolian gerbils. Jikken Dobutsu. 1991 Oct;40(4):573-7. MIGLIORANZA M.H., GARGANI L., SANT'ANNA R.T., ROVER M.M., MARTINS V.M., MANTOVANI A., WEBER C., MORAES M.A., FELDMAN C.J., KALIL R.A., SICARI R., PICANO E., LEIRIA T.L. Lung Ultrasound for the Evaluation of Pulmonary Congestion in Outpatients: A Comparison With Clinical Assessment, Natriuretic Peptides, and Echocardiography. JACC Cardiovasc Imaging. 2013; 26. pii: S1936-878X(13)00610-4. MISAKA T., SUZUKI S., MIYATA M., KOBAYASHI A., SHISHIDO T., ISHIGAMI A., SAITOH S., HIROSE M., KUBOTA I., TAKEISHI Y. Deficiency of senescence marker protein 30 exacerbates angiotensin II-induced cardiac remodelling. Cardiovasc Res. 2013, 1;99(3):461-70. MISAKA T., SUZUKI S., MIYATA M., KOBAYASHI A., ISHIGAMI A., SHISHIDO T., SAITOH S., KUBOTA I., TAKEISHI Y. Senescence marker protein 30 inhibits
http://www.ncbi.nlm.nih.gov/pubmed?term=Markel%27%20AL%5BAuthor%5D&cauthor=true&cauthor_uid=15514738
http://www.ncbi.nlm.nih.gov/pubmed?term=Shmerling%20MD%5BAuthor%5D&cauthor=true&cauthor_uid=15514738
http://www.ncbi.nlm.nih.gov/pubmed?term=Shmerling%20MD%5BAuthor%5D&cauthor=true&cauthor_uid=15514738
http://www.ncbi.nlm.nih.gov/pubmed?term=de%20Bold%20ML%5BAuthor%5D&cauthor=true&cauthor_uid=16269246
http://www.ncbi.nlm.nih.gov/pubmed?term=de%20Bold%20AJ%5BAuthor%5D&cauthor=true&cauthor_uid=16269246

54
angiotensin II-induced cardiac hypertrophy and diastolic dysfunction. Biochem Biophys Res Commun. 2013, 13;439(1):142-7. MIYOSHI H., TAKAYAMA Y., TAMURA T., KITASHIRO S., IZUOKA T., SAITO D., IMURO Y., IWASAKA T. Regional myocardial function at the papillary muscle insertion site. Jpn J Physiol. 2001;51(1):109-14. NAKAGOMI S., BARSOUM M.J., BOSSY-WETZEL E., SÜTTERLIN C., MALHOTRA V., LIPTON S.A. A Golgi fragmentation pathway in neurodegeneration. Neurobiol Dis. 2008; 29(2):221-31. PALADE G.E. Blood capilares of the heart and other organs. Circul. 1961; 24:368-88 PAN SS. Alterations of atrial natriuretic peptide in cardiomyocytes and plasma of rats after different intensity exercise. Scand J Med Sci Sports. 2008; 18(3):346-53. PENG J.Y., XIN H.Y., HAN P., ZHAO H.B., BAI L A.N., CAO B.Y. Identification and gene expression analyses of natriuretic peptide system in the ovary of goat (Capra hircus). Gene. 2013, 25;524(2):105-13. PETERSON C.M., JOHANNSEN D.L., RAVUSSIN E. Skeletal muscle mitochondria and aging: a review. J Aging Res. 2012;2012:194821. POLLACK J.A., SKVORAK J.P., NAZIAN S.J., LANDON C.S., DIETZ J.R. Alterations in atrial natriuretic peptide (ANP) secretion and renal effects in aging. J Gerontol A Biol Sci Med Sci. 1997; 52(4):B196-202. RADEMAKER M.T., RICHARDS A. M. Cardiac natriuretic peptides for cardiac health. Clinical Science 2005; 108, 23–36. RIEDEL C. G., DOWEN R. H., LOURENCO G. F., KIRIENKO N. V., HEIMBUCHER T., WEST J. A., BOWMAN S. K., KINGSTON R. E., DILLIN A., ASARA J. M., RUVKUN G.DAF-16/FOXO employs the chromatin remodeller SWI/SNF to promote stress resistance and longevity. Nat Cell Biol. 2013 May; 15(5): 491–501. RASUL A., KHAN M., ALI M., LI J., LI X. Targeting Apoptosis Pathways in Cancer with Alantolactone and Isoalantolactone. The Scientific World Journal Vol 2013 (2013), Art ID 248532, 9 pages. RODRIGUEZ A., BJERLING P. The links between chromatin spatial organization and biological function. Biochem Soc Trans. 2013; 1; 41(Pt 6): 1634–1639. SAGNELLA G. A. Practical implications of current natriuretic peptide. Journal of Renin-Angiotensin-Aldosterone System 2000 1: 304-315. SARMA S., CARRICK-RANSON G., FUJIMOTO N., ADAMS-HUET B., BHELLA P.S., HASTINGS J.L., SHAFER K.M., SHIBATA S., BOYD K., PALMER D., SZCZEPANIAK E.W., SZCZEPANIAK L.S., LEVINE B.D.T he Effects of Age and Aerobic Fitness on Myocardial Lipid Content. Circ Cardiovasc Imaging. 2013; 6(6):1048-55.
http://www.ncbi.nlm.nih.gov/pubmed?term=Johannsen%20DL%5BAuthor%5D&cauthor=true&cauthor_uid=22888430

55
SCHIEBINGER R.J, GREENING K.M. Interaction between stretch and hormonally stimulated atrial natriuretic peptide secretion. Department of Medicine, Wayne State University, Detroit 48201. Am J Physiol. 1992; 262(1 Pt 2):H78-83. SEEDORF M., DAMELIN M., KAHANA J., TAURA T., SILVER P. A.Interactions between a Nuclear Transporter and a Subset of Nuclear Pore Complex Proteins Depend on Ran GTPase. Mol Cell Biol. 1999 Feb;19(2):1547-57. SEUL K.H, PARK J.K, PARK B.O, KIM S.H, CHO K.W. Effects of acceleration stress on the secretion of atrial natriuretic peptide (ANP) in rats. Physiologist. 1992; 35(1 Suppl):S67-8. SHAIKH Q., AHMED B., AHMED M., MAHAR J.H., AHMAD M., AHMED A., MAJEED F., ALI F.S., KHAN M., KAMAL A.K. Left atrial volumes and associated stroke subtypes. BMC Neurol. 2013; 18;13(1):149. SHORT B., HAAS A., BARR F.A. Golgins and GTPases, giving identity and structure to the Golgi apparatus. Biochim Biophys Acta. 2005; 10;1744(3):383-95. SILVA L.B., FERREIRA C.A., BLACHER C., LEAES P., HADDAD H. [B-type natriuretic peptide and cardiovascular disease]. Arq Bras Cardiol. 2003; 81(5):529-39. SKEPPER J.N., NAVARATNAM V., MARTENSZ N.D. Effects of expansion of blood volume and bilateral vagotomy on specific heart granules and release of atrial natriuretic peptide in the rat. Cell And Tissue Research. 1989; 258(1): 211-218. SKEPPER J.N., WOODWARD J.M., NAVARATNAM V. Immunocytochemical localization of natriuretic peptide sequences in the human right auricle. J Mol Cell Cardiol. 1988; 20(4):343-53. SOUALMIA H., DJERIDANE Y., EURIN J., TOUITOU Y. (2007). Aging and Photoperiod Affect The Daily Rhythm Patterno of Atrial Natriuretic Peptide in The Rat Atrium. j.peptides. 2007; 09.007. 2356-60. SWÄRD K., VALSON F., RICKSTEN S.E. Long-term infusion of atrial natriuretic peptide (ANP) improves renal blood flow and glomerular filtration rate in clinical acute renal failure. Acta Anaesthesiol Scand. 2001; 45(5):536-42. TUROVSKY E.A., TUROVSKAYA M.V., DOLGACHEVA L.P., ZINCHENKO V.P., DYNNIK V.V. Acetylcholine promotes Ca2+ and NO-oscillations in adipocytes implicating Ca2+→NO→cGMP→cADP-ribose→Ca2+ positive feedback loop--modulatory effects of norepinephrine and atrial natriuretic peptide. PLoS One. 2013; 16;8(5):e 63483. TOYOSHIMA Y., SUZUKI S., AWAL M.A. Atrial natriuretic peptide (ANP)-grnules of auricular cardiocytes in dehydrated and rehydrated mice. Exp Anim 1996; 45(2):135-140.
http://www.ncbi.nlm.nih.gov/pubmed?term=Sw%C3%A4rd%20K%5BAuthor%5D&cauthor=true&cauthor_uid=11309000

56
VÁZQUEZ-MARTÍNEZ R., DÍAZ-RUIZ A., ALMABOUADA F., RABANAL-RUIZ Y., GRACIA-NAVARRO F., MALAGÓN M.M. Revisiting the regulated secretory pathway: from frogs to human. Gen Comp Endocrinol. 2012; 1;175(1):1-9. VILLACORTA H., DUARTE A., DUARTE N.M., CARRANO A., MESQUITA E.T., DOHMANN H.J.F., FERREIRA F.E.G. Valor do Peptídeo Natriurético do Tipo B no Diagnóstico de Insuficiência Cardíaca Congestiva em Pacientes Atendidos com Dispnéia na Unidade de Emergência. Arq Bras Cardiol. 2002; 79(6): 564-8. VIANNA P.T.G. Rim e Anestesia. Revista Brasileira de Anestesiologia. 1991; 41(5): 347-356. VOELTZ G. K., ROLLS M.M., RAPOPORT T. A. Structural organization of the endoplasmic reticulum. EMBO 2002 October; 3(10): 944–950. VOEUX P.L. Integração dos Mecanismos Renais para o Controle do Volume Sanguíneo e do Volume Líquido Extra;celular; e Regulação Renal do Potássio, do Cálcio, do Fosfato e do Magnésio. In: GUYTON, A. C.; HALL, J. E. Tratado de Fisiologia Médica. 2002; 8: 312-327. WANG M., ZHANG N., WANG W., WANG B., XU Z., YANG Y. Expression and function of vascular endothelial growth inhibitor in aged porcine bladder detrusor muscle cells. Biogerontology. 2013; 14(5):543-56. WARD T. H., POLISHCHUK R. S., CAPLAN S., HIRSCHBERG K., LIPPINCOTT-SCHWARTZ J. Maintenance of Golgi structure and function depends on the integrity of ER export. JCB 155. 4. 557-570 WOODMAN O. L., MALAKUL W., CAO A. H., XU Q., RITCHIE R. H. Atrial Natriuretic Peptide Prevents Diabetes-Induced Endothelial Dysfunction. Life Sci. 2008; 9;82(15-16):847-54. WU Q., CAI O., CHEN S., WANG W. (2008). Corin: New Insights into the Natriuretic Peptide System. Kidney Int. 2009; 75(2): 142–146.44195. YANG T., CHIAO Y.A., WANG Y., VOORHEES A., HAN H.C., LINDSEY M.L., JIN Y.F. Mathematical modeling of left ventricular dimensional changes in mice during aging. BMC Syst Biol. 2012;6 Suppl 3:S10. ZHANG X.P., VATNER S.F., SHEN Y.T., ROSSI F., TIAN Y., PEPPAS A., RESUELLO R.R., NATIVIDAD F.F., VATNER D.E. Increased apoptosis and myocyte enlargement with decreased cardiac mass; distinctive features of the aging male, but not female, monkey heart. J Mol Cell Cardiol. 2007; 43(4):487-91. ZHAO D., PANSY N., NAVAR L. N. ANP-Mediated Inhibition of Distal Nephron Fractional Sodium Reabsorption in Wild-Type and Mice Overexpressing Natriuretic Peptide Receptor. Am J Physiol Renal Physiol. 2010; 298(1): F103–F108.
http://www.ncbi.nlm.nih.gov/pubmed?term=Almabouada%20F%5BAuthor%5D&cauthor=true&cauthor_uid=21907200
















![Breve revisão sobre Testes de Hipóteses · Não é possível reduzir simultaneamente a probabilidade dos dois tipos de erro: diminuir P[Erro Tipo I] significa reduzir a gama de](https://static.fdocument.org/doc/165x107/5e4aca984ac3ea5f41757324/breve-reviso-sobre-testes-de-hipteses-no-possvel-reduzir-simultaneamente.jpg)

