Tumor Evasion of the Immune System by Converting CD4 CD25 T ...
11
of February 15, 2018. This information is current as β Tumor-Derived TGF- T Regulatory Cells: Role of + CD25 + T Cells into CD4 - CD25 + Converting CD4 Tumor Evasion of the Immune System by Beverly A. Teicher and Chung Lee Irwin Park, Ximing Yang, Qiang Zhang, Scott Lonning, Victoria C. Liu, Larry Y. Wong, Thomas Jang, Ali H. Shah, http://www.jimmunol.org/content/178/5/2883 doi: 10.4049/jimmunol.178.5.2883 2007; 178:2883-2892; ; J Immunol References http://www.jimmunol.org/content/178/5/2883.full#ref-list-1 , 30 of which you can access for free at: cites 54 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2007 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 15, 2018 http://www.jimmunol.org/ Downloaded from by guest on February 15, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Tumor Evasion of the Immune System by Converting CD4 CD25 T ...

of February 15, 2018.This information is current as
βTumor-Derived TGF- T Regulatory Cells: Role of+CD25
+ T Cells into CD4−CD25+Converting CD4Tumor Evasion of the Immune System by
Beverly A. Teicher and Chung LeeIrwin Park, Ximing Yang, Qiang Zhang, Scott Lonning, Victoria C. Liu, Larry Y. Wong, Thomas Jang, Ali H. Shah,
http://www.jimmunol.org/content/178/5/2883doi: 10.4049/jimmunol.178.5.2883
2007; 178:2883-2892; ;J Immunol
Referenceshttp://www.jimmunol.org/content/178/5/2883.full#ref-list-1
, 30 of which you can access for free at: cites 54 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 15, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Tumor Evasion of the Immune System by ConvertingCD4�CD25� T Cells into CD4�CD25� T Regulatory Cells:Role of Tumor-Derived TGF-�
Victoria C. Liu,* Larry Y. Wong,* Thomas Jang,* Ali H. Shah,* Irwin Park,* Ximing Yang,*Qiang Zhang,* Scott Lonning,† Beverly A. Teicher,† and Chung Lee1*
CD4�CD25� T regulatory (Treg) cells were initially described for their ability to suppress autoimmune diseases in animal models.An emerging interest is the potential role of Treg cells in cancer development and progression because they have been shown tosuppress antitumor immunity. In this study, CD4�CD25� T cells cultured in conditioned medium (CM) derived from tumor cells,RENCA or TRAMP-C2, possess similar characteristics as those of naturally occurring Treg cells, including expression of Foxp3,a crucial transcription factor of Treg cells, production of low levels of IL-2, high levels of IL-10 and TGF-�, and the ability tosuppress CD4�CD25� T cell proliferation. Further investigation revealed a critical role of tumor-derived TGF-� in convertingCD4�CD25� T cells into Treg cells because a neutralizing Ab against TGF-�, 1D11, completely abrogated the induction of Treg
cells. CM from a nontumorigenic cell line, NRP-152, or irradiated tumor cells did not convert CD4�CD25� T cells to Treg cellsbecause they produce low levels of TGF-� in CM. Finally, we observed a reduced tumor burden in animals receiving 1D11. Thereduction in tumor burden correlated with a decrease in tumor-derived TGF-�. Treatment of 1D11 also reduced the conversionof CD4� T cells into Treg cells and subsequent Treg cell-mediated suppression of antitumor immunity. In summary, we havedemonstrated that tumor cells directly convert CD4�CD25� T cells to Treg cells through production of high levels of TGF-�,suggesting a possible mechanism through which tumor cells evade the immune system. The Journal of Immunology, 2007, 178:2883–2892.
A key aspect of tumor immunology is immune surveil-lance in the tumor-bearing host. It has been almost halfa century since the concept of immune surveillance was
first proposed by Burnet (1). This hypothesis has been debated andtested over the years. Despite the ability to generate immune re-activity against tumor Ags, the immune surveillance program canbe overpowered by tumors with eventual tumor progression (2–5).Tumor-infiltrating lymphocytes (TILs)2 were identified in both hu-man and murine tumors and were shown to recognize and rejecttumor cells in immunotherapy (4, 6). Despite their ability to infil-trate into tumor sites, the presence of TILs, however, is not alwaysbeneficial. Although a better prognosis in patients with elevatedTILs has been observed in several studies, the mere presence ofTILs is insufficient to reject tumors (7–9). Rather, the type of in-filtrating cell is the essential factor in the rejection of the tumor(10–15). Numerous studies have shown that CD4� Th cells andCD8� CTL are beneficial in antitumor immunity, whereas thepresence of CD4�CD25� T regulatory (Treg) cells is detrimentalbecause the presence of Treg cells is associated with reduced sur-vival (10–15).
CD4�CD25� Treg cells are a unique lineage of the T cell pop-ulation that regulates the immune system by inhibiting immunecell proliferation. In the 1970s, it was first proposed by Gershonand Kondo (53, 54) that T lymphocytes could function as helpercells as well as regulatory cells to suppress the immune response.In addition to suppressing CD4� T cells and NK cells, Treg cellssuppress activation and proliferation of CD8� T cells (19). Treg
cells inhibit target cells through a wide range of cytokines. Besidesexpressing TGF-�, Treg cells produce IL-10, and express CTLA-4,glucocorticoid-induced TNFR, and transcription factor Foxp3(20–22). It is well established that Treg cells are a major player inthe regulation of the immune system to prevent autoimmunity bysuppressing harmful autoreactive immune cells (16 –18, 23, 24).Recently, an emerging interest has been focused on the associ-ation between Treg cells and cancer development. These cellshave been shown to suppress antitumor immunity, and deple-tion of Treg cells in vivo enhances antitumor immunity and fa-cilitates tumor rejection (25–28).
There are at least two subsets of Treg cells. One subset, alsoknown as naturally occurring Treg cells, develops during the nor-mal process of T cell maturation in the thymus. The other subsetdevelops as a consequence of activation of mature T cells underparticular conditions in the periphery (28, 29). The negative reg-ulatory role of Treg cells has been amply demonstrated, suggestingthat the presence of Treg cells in the tumor microenvironmentcould indeed promote, instead of suppressing, tumor progressionby inhibiting antitumor immunity (13–15, 30). However, the originof these Treg cells in the tumor sites remains elusive. It is unknownwhether they are naturally occurring Treg cells that are recruited tothe tumor sites or whether they arrive at tumor sites originally asCD4� Th cells that are later converted to Treg cells in the tumormicroenvironment. Because the presence of Treg cells in the tumor
*Department of Urology, Northwestern University Medical School, Chicago, IL 60611;and †Genzyme, Framingham, MA 01701
Received for publication June 15, 2006. Accepted for publication December 21, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Chung Lee, Department ofUrology, Tarry 16-733, Northwestern University Medical School, 303 East ChicagoAvenue, Chicago, IL 60611. E-mail address: [email protected] Abbreviations used in this paper: TIL, tumor-infiltrating lymphocyte; CM, condi-tioned medium; DC, dendritic cell; Treg, T regulatory.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

sites is associated with poor prognosis in patients, identification ofthe origin of these cells will provide valuable information in treat-ment of cancer. In this study, we observed the tumor conversion ofnaive CD4�CD25� T cells into CD4�CD25�Foxp3� Treg cellsthrough the production of high levels of TGF-�. Furthermore, neu-tralization of TGF-� abrogated this conversion both in vitro andin vivo.
Materials and MethodsMice and cell lines
Male C57BL/6 and BALB/c mice (6 – 8 wk of age; Harlan Sprague-Dawley) were maintained in pathogen-free facilities at the Center forComparative Medicine at Northwestern University Feinberg School ofMedicine in accordance with established guidelines of the Animal Care andUse Committee of Northwestern University. The mouse prostate tumorTRAMP-C2 cells were cultured in RPMI 1640 medium supplemented with10% FBS and 1% penicillin/streptomycin (Invitrogen Life Technologies).The mouse renal cell carcinoma RENCA cells were cultured in DMEMsupplemented with 10% FBS and 1% penicillin/streptomycin. The NRP-152 cells are nontumorigenic rat prostate epithelium cells and were cul-tured in DMEM supplemented with 10% FBS, 1% penicillin/streptomycin,5 �g/ml insulin, and 0.1 �M dexamethasone. All cells were kept at37°C/5% CO2.
Isolation of CD4�CD25� and CD4�CD25� T cells
For studies with CM, CD4�CD25� and CD4�CD25� T cells from adultmouse spleens were isolated using a mouse CD4�CD25� T RegulatoryCell Isolation Kit, according to manufacturer’s instructions (Miltenyi Bio-tec). For coculturing studies, T cells were stained with rat anti-mouse CD4and CD25 mAbs (BD Pharmingen) after coculturing with TRAMP-C2 orNRP-152 cells for 7 days and sorted on a FACS sorter into CD4�CD25�
and CD4�CD25� T cell populations. All purified T cells had a purity of�90%, as determined by FACS.
Coculture of TRAMP-C2 or NRP-152 cells with CD4�CD25�
T cells
CD4�CD25� T cells (4 � 106) were isolated from the spleen of adultC57BL/6 mice (Miltenyi Biotec) and cultured with TRAMP-C2 or NRP-152 cells (2 � 106) in the presence of 1 �g/ml rat anti-mouse anti-CD3mAb (BD Pharmingen) and 3000 rad-irradiated, T cell-depleted spleno-cytes (1 � 107) as APCs for 7 days with replacing of TRAMP-C2 orNRP-152 cells every 2 days. T cells cultured in T cell medium with anti-CD3 Ab and APCs were used as a control. After the end of the 7-dayculture, cells were sorted on a FACS sorter according to the cell surfaceCD25 expression into CD4�CD25� and CD4�CD25� T cell populations.To determine the rate of conversion, isolated CD4�CD25� T cells werecocultured with TRAMP-C2, as described above, for 5 days. T cells werecollected on each day and stained for CD4 and CD25 expression levelswith Abs against CD4 and CD25 (BD Pharmingen) and analyzed by FACS.The effect of number of TRAMP-C2 cells on conversion was determinedby coculturing TRAMP-C2 cells and isolated CD4�CD25� T cells at dif-ferent ratios (0:1, 0.25:1, 0.5:1, 0.75:1, and 1:1) for 5 days. At the endpoint, T cells were collected and stained for cell surface expression levelsof CD4 and CD25 (BD Pharmingen) and analyzed by FACS.
Culture of CD4�CD25� T cells with conditioned medium (CM)
Isolated CD4�CD25� T cells (1 � 106/well in 24-well plate; MiltenyiBiotec) were cultured either in complete T cell medium (RPMI 1640 sup-plemented with 10% FBS, 1% penicillin/streptomycin, 10 mM HEPESbuffer, 10 mM nonessential amino acids, 1 mM sodium pyruvate, 4 mML-glutamine, and 0.05 mM 2-ME) or in CM collected from a 3-day cultureof RENCA, TRAMP-C2, NRP-152, or 20,000 rad-irradiated TRAMP-C2cells in the presence of 1 �g/ml plate-bound anti-CD3 and 2 �g/ml solubleanti-CD28 Abs (BD Pharmingen) for 5 days. T cells cultured in T cellmedium were used as a control. In some cases, a neutralizing Ab againstTGF-� 1D11 (100 �g/ml; Genzyme) was added to CM to block TGF-�signaling. To determine the concentration of CM on conversion ofCD4�CD25� to CD4�CD25� T cells, 0.01, 0.1, 0.2, 0.5, and 0.9 ml ofTRAMP-C2 CM were added to T cell medium in a final volume of 1 ml,and isolated CD4�CD25� T cells were cultured in diluted CM for 5 days,as described above. Five days later, T cells were collected and stained forcell surface CD4 and CD25 expression levels (BD Pharmingen) and ana-lyzed by FACS.
Foxp3 RT-PCR
To detect Foxp3 expression, total RNA was extracted from T cells culturedunder various conditions using a QiaEasy RNA Isolation Kit (Qiagen). Atwo-step RT-PCR was performed by first reverse transcribing RNA tocDNA using a First-Strand DNA Synthesis Kit (Invitrogen Life Technol-ogies), and then PCR was conducted using Foxp3-specific primers, as de-scribed previously (22).
Intracellular detection of Foxp3
To detect Foxp3 protein levels, T cells were stained using a Foxp3-specificmAb, according to the manufacturer’s instructions (eBioscience). Briefly,cells were fixed and permeablized in Fix/Perm Buffer and incubated overnightin the dark at 4°C. The next day, an Fc block (BD Pharmingen) was used toblock the nonspecific binding before adding Fopx3 Ab, and cells wereincubated in the dark at 4°C for 30 min. A rat IgG2a FITC Ab was used asan isotype control. Cells were then washed and analyzed by FACS.
ELISA
Isolated CD4�CD25� T cells were cultured with or without TRAMP-C2CM. Five days later, they were collected, washed extensively, plated at adensity of 1 � 105/well in a U-bottom 96-well plate, and restimulated with1 �g/ml plate-bound anti-CD3 and 2 �g/ml soluble anti-CD28 Abs (BDPharmingen) for 48 h (for IL-2 and IL-10 assays) or 72 h (TGF-� assay).ELISA were performed to measure the cytokine levels in the cell-freesupernatant, according to manufacturers’ instructions (IL-2, IL-10 ELISA,BioSource International; TGF-� ELISA, R&D Systems). For TGF-�ELISA, serum-free medium was used. To measure TGF-� production inCM of various cell lines, all cells were plated at a density of 1 � 106/T-25flask and cultured in serum-free medium for 48 h. Cell-free supernatantwas collected and TGF-� production was measured using a TGF-� ELISAkit, according to the manufacturer’s instructions (R&D Systems). To detectTGF-� production in CM from irradiated TRAMP-C2 cells, TRAMP-C2cells were irradiated (20,000 rad) and cultured in serum-free T cell medium(lacking 10% FBS) for 48 h, and CM was collected for TGF-� ELISA.
T cell suppression assay
T cells (5 � 104) from various culture conditions were cultured withfreshly isolated CD4�CD25� T cells (5 � 104 or 1.5 � 104) from spleensof wild-type mice in the presence of 0.5 �g/ml anti-CD3 Ab (BD Pharm-ingen) and 3000 rad-irradiated, T cell-depleted splenocytes (5 � 104) for72 h in a 96-well U-bottom plate. During the last 6–18 h, 1 �Ci/well[3H]thymidine (Amersham Biosciences) was added to the culture. Cellswere harvested and counted in a scintillation counter. For the titrationassay, CD4�CD25� T cells were cultured with TRAMP-C2 CM for 5days. Five days later, T cells were collected and cultured at various con-centrations with CD4�CD25� T cells (5 � 104) for suppression assays.For the in vivo study, 1.5 � 104/well CD4�CD25� T cells isolated fromlung tissues were used.
Immunofluorescence staining of mouse lung tissues
C57BL/6 mice were injected i.v. via tail vein with 1 � 106 TRAMP-C2cells. Thirty days later, animals were sacrificed and lung tissue was fixedin formalin and cut into 4-�m sections for immunofluorescence stain-ing. CD4 FITC, CD25 PE (1/100 dilution; BD Pharmingen), and Foxp3(eBioscience) were used for the histological staining. For TGF-� staining,a 1D11 Ab was used as the primary Ab (1/100 dilution; Genzyme) anda goat anti-mouse Texas Red Ab was used as the secondary Ab (1/400dilution; Santa Cruz Biotechnology). Nuclei were counterstained withHoechst 33342 dye (Sigma-Aldrich). Sections were viewed under aconfocal microscope.
Tumor induction and treatment with 1D11
BALB/c mice were injected with 1 � 106 RENCA cells i.v. via the tailvein. Five mice received a neutralizing Ab against TGF-� 1D11; five micereceived control IgG Ab 13C4 (Genzyme) at a loading dose of 100 mg/kgi.p. at the day of the injection; and five mice received no treatment. Theexperimental and control groups received 1D11 and 13C4, respectively,every 3 days at a dose of 50 mg/kg over a course of a 30-day experiment.Mice were sacrificed at day 30, and lung tissues were fixed with formalinand cut into 4-�m sections for H&E and immunofluorescence staining.Tumor burden was evaluated by counting numbers of tumor metastases ofH&E-stained lung tissue sections of each animal.
2884 TUMOR-DERIVED TGF-� CONVERTS CD4�CD25� T CELLS TO Treg CELLS
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

CD4�CD25� T cell isolation from lung tissues
BALB/c mice were challenged with RENCA cells and treated with orwithout 1D11 or 13C4, as described above. At the end of the experiment,lung tissues from each animal were digested with 150 U/ml collagenase(Sigma-Aldrich) for 1 h at 37°C. After digestion, lung tissues were cut intosmall pieces, and single-cell suspensions were obtained and filteredthrough 70- and 40-�m cell strainers (VWR). Lymphocytes were separatedusing Percoll (Amersham Biosciences). Briefly, cells were pooled fromanimals in the same group and resuspended in 8 ml of 40% Percoll andloaded onto 3 ml of 70% Percoll in a 15-ml centrifuge tube. Tubes werespun at 2500 rpm for 20 min at room temperature, and the interface (lym-phocytes) was removed using a 3-ml syringe with a 20-gauge needle. Cellswere washed three times with PBS to remove the Percoll. CD4�CD25� Tcells were isolated from enriched lung lymphocytes using MACS (MiltenyiBiotec), according to the manufacturer’s instructions. The purity of isolatedcell populations was �90%, as determined by FACS.
Statistical analysis
Mean � SD are given. For comparison of groups, the one-way ANOVAwith posttest comparison was performed, and a p � 0.05 was consideredsignificant.
ResultsCD4�CD25� and Foxp3� T cells are present within tumormasses
Since Woo et al. (31) observed increased Treg cells in peripheralblood in patients with non-small cell lung tumor than the normaldonors in 2001, there have been numerous studies that reported ahigh frequency of Treg cells in peripheral blood in various humancancers (13, 15, 32, 33). A recent study by Curiel et al. (11) re-ported the presence of CD4�CD25�, Foxp3�CD3� T cells in hu-man ovarian carcinoma. In an attempt to explore this phenomenonand potential connections between tumors and Treg cells, we useda mouse prostate cancer cell line TRAMP-C2 in this study. Toexamine whether CD4�CD25� and Foxp3� T cells are alsopresent in TRAMP-C2 tumors, lung tissues from mice receivingTRAMP-C2 i.v. were used for immunofluorescence studies.CD4�CD25� T cells were detected in the metastatic tumor sitesin lung tissues from these mice (Fig. 1A). More importantly,Foxp3�CD25� T cells were also detected at the tumor sites (Fig.1B). Because Foxp3 is exclusively expressed by Treg cells (22), thepositive staining of Foxp3 at tumor sites suggests these cells areTreg cells. It has been shown in other studies that the presence ofsuch CD4�CD25�Foxp3� Treg cells in tumor sites is associatedwith suppression of antitumor immunity; however, the origin ofthese cells remains elusive. In the following studies, we exploredthe connections between tumor cells and Treg cells and the possibleorigin of such Treg cells in the tumor mass.
T cells cocultured with TRAMP-C2 express Foxp3
Treg cells are developed in thymus or induced and differentiated inthe periphery, such as in the tumor microenvironment (34–37). Ithas been shown that tumor-derived cytokines, such as IL-10,TGF-�, and vascular endothelial growth factor, suppress dendriticcell (DC) maturation and function. As a result, these dysfunctionalDCs induce differentiation of Treg cells in the tumor site (38, 39).However, it is unknown whether tumor cells can directly induceTreg cell differentiation. To investigate the direct relationship be-tween tumors and Treg cells, TRAMP-C2 cells were used in acoculture with CD4�CD25� T cells isolated from C57BL/6 mice.After a 7-day coculture, T cells were sorted on a FACS sorteraccording to the cell surface CD25 expression into CD4�CD25�
and CD4�CD25� populations. Because Foxp3 is a crucial tran-scriptional factor and is exclusively expressed in Treg cells, Foxp3expression in these cells was examined (22). Total RNA was ex-tracted from individual T cell populations, and RT-PCR was per-formed using Foxp3-specific primers. Foxp3 expression was ob-served in control CD4�CD25� Treg cells (Ctrl 25�) as well as inTRAMP-C2-cocultured CD25� T cells (Fig. 2A, top panel). Incontrast, Foxp3 expression was absent in control CD4�CD25� Tcells (Ctrl 25�) or TRAMP-C2-cocultured CD25� T cells (Fig.2A, top panel). In addition, T cells cultured alone did not expressFoxp3 (data not shown). Interestingly, CD4�CD25� T cells cocul-tured with nontumorigenic rat prostate epithelium NRP-152 cellsdid not express Foxp3 in either sorted CD25� or CD25� T cells(Fig. 2A, top panel).
Next, the kinetics of conversion was examined. To study the rateof conversion, TRMAP-C2 cells were cocultured with purifiedCD4�CD25� T cells as above for 0–5 days, and T cells werecollected on each day and analyzed by FACS for CD25 expressionlevels. As shown in Fig. 2B, top panel, the percentage ofCD4�CD25� T cells increased in a time-dependent manner. Inaddition, the ratio of conversion was studied by coculturingTRAMP-C2 and CD4�CD25� T cells at 0:1, 0.1:1, 0.25:1, 0.5:1,0.75:1, and 1:1 for 5 days. At the end of the 5-day culture, T cellswere collected and cell surface CD25 expression was analyzed byFACS. The percentage of CD4�CD25� T cells increased in aratio-dependent manner (Fig. 2B, bottom panel). These results sug-gest that coculture of purified CD4�CD25� T cells with tumorcells TRAMP-C2, but not nontumorigenic NRP-152 cells, inducesCD4�CD25�Foxp3� T cells. Moreover, these results provide ev-idence that tumor cells can indeed directly convert CD4�CD25�
T cells into Treg cells.
FIGURE 1. CD4�CD25� and Foxp3� T cells presentwithin the tumor mass. TRAMP-C2 cells (1 � 106)were injected into C57BL/6 mice i.v. via tail vein. Micewere sacrificed at day 30, lung tissues were fixed informalin and cut into 4-�m sections, and slides wereused to stain for CD4, CD25 (A), and Foxp3 (B) ex-pression and viewed under a confocal microscope. Arepresentative section is shown here. A, CD4�CD25� Tcells in the tumor mass. Red color indicates CD25� Tcells; green, CD4� T cells; and double-positive T cellsare shown as yellow. B, CD25�Foxp3� T cells in thetumor mass. Foxp3� T cells are shown in green. Tumornuclei were counterstained with Hoechst 33342 dye andare shown as blue. Scale bars, 20 �m.
2885The Journal of Immunology
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
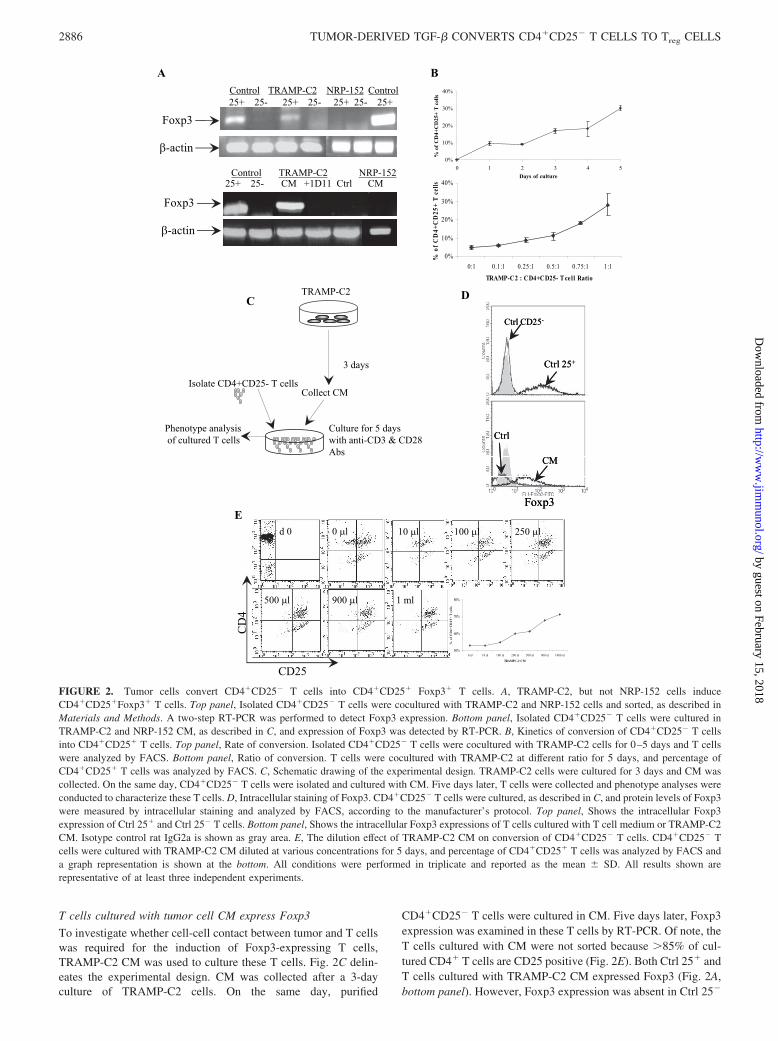
T cells cultured with tumor cell CM express Foxp3
To investigate whether cell-cell contact between tumor and T cellswas required for the induction of Foxp3-expressing T cells,TRAMP-C2 CM was used to culture these T cells. Fig. 2C delin-eates the experimental design. CM was collected after a 3-dayculture of TRAMP-C2 cells. On the same day, purified
CD4�CD25� T cells were cultured in CM. Five days later, Foxp3expression was examined in these T cells by RT-PCR. Of note, theT cells cultured with CM were not sorted because �85% of cul-tured CD4� T cells are CD25 positive (Fig. 2E). Both Ctrl 25� andT cells cultured with TRAMP-C2 CM expressed Foxp3 (Fig. 2A,bottom panel). However, Foxp3 expression was absent in Ctrl 25�
TRAMP-C2
3 days
Collect CMIsolate CD4+CD25- T cells
Culture for 5 dayswith anti-CD3 & CD28Abs
Phenotype analysisof cultured T cells
A B
50%
60%
70%
80%
0 ul 10 ul 100 ul 200 ul 500 ul 900 ul 1000 ul
TRAMPC-2 CM
sllecT
+52D
C+ 4D
Cfo%
4D
C
CD25
0 µl
1 ml900 µl500 µl
250 µl100 µl10 µld 0
0%
10%
20%
30%
40%
0:1 0.1:1 0.25:1 0.5:1 0.75:1 1:1
TRAMP-C2 : CD4+CD25- T cell Ratio
sllecT
+52D
C+ 4D
Cf o%
0%
10%
20%
30%
40%
0 1 2 3 4 5Days of culture
sllecT
+52D
C +4D
Cfo%
C D
E
Foxp3
β-actin
ControlCM25+ 25- CM +1D11 Ctrl
TRAMP-C2 NRP-152
25+ 25- 25+ 25- 25+ 25- 25+Control TRAMP-C2 NRP-152 Control
Foxp3
β-actin
Ctrl CD25-
Ctrl 25+
CM
Ctrl
Foxp3
Ctrl CD25-
Ctrl 25+
Ctrl CD25-
Ctrl 25+
CM
Ctrl
CM
Ctrl
Foxp3
FIGURE 2. Tumor cells convert CD4�CD25� T cells into CD4�CD25� Foxp3� T cells. A, TRAMP-C2, but not NRP-152 cells induceCD4�CD25�Foxp3� T cells. Top panel, Isolated CD4�CD25� T cells were cocultured with TRAMP-C2 and NRP-152 cells and sorted, as described inMaterials and Methods. A two-step RT-PCR was performed to detect Foxp3 expression. Bottom panel, Isolated CD4�CD25� T cells were cultured inTRAMP-C2 and NRP-152 CM, as described in C, and expression of Foxp3 was detected by RT-PCR. B, Kinetics of conversion of CD4�CD25� T cellsinto CD4�CD25� T cells. Top panel, Rate of conversion. Isolated CD4�CD25� T cells were cocultured with TRAMP-C2 cells for 0–5 days and T cellswere analyzed by FACS. Bottom panel, Ratio of conversion. T cells were cocultured with TRAMP-C2 at different ratio for 5 days, and percentage ofCD4�CD25� T cells was analyzed by FACS. C, Schematic drawing of the experimental design. TRAMP-C2 cells were cultured for 3 days and CM wascollected. On the same day, CD4�CD25� T cells were isolated and cultured with CM. Five days later, T cells were collected and phenotype analyses wereconducted to characterize these T cells. D, Intracellular staining of Foxp3. CD4�CD25� T cells were cultured, as described in C, and protein levels of Foxp3were measured by intracellular staining and analyzed by FACS, according to the manufacturer’s protocol. Top panel, Shows the intracellular Foxp3expression of Ctrl 25� and Ctrl 25� T cells. Bottom panel, Shows the intracellular Foxp3 expressions of T cells cultured with T cell medium or TRAMP-C2CM. Isotype control rat IgG2a is shown as gray area. E, The dilution effect of TRAMP-C2 CM on conversion of CD4�CD25� T cells. CD4�CD25� Tcells were cultured with TRAMP-C2 CM diluted at various concentrations for 5 days, and percentage of CD4�CD25� T cells was analyzed by FACS anda graph representation is shown at the bottom. All conditions were performed in triplicate and reported as the mean � SD. All results shown arerepresentative of at least three independent experiments.
2886 TUMOR-DERIVED TGF-� CONVERTS CD4�CD25� T CELLS TO Treg CELLS
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
or CD4�CD25� T cells cultured in T cell medium (Ctrl) (Fig. 2A,bottom panel). Again, CD4�CD25� T cells cultured with NRP-152 CM did not express Foxp3 (Fig. 2A, bottom panel). In addi-tion, protein expression of Foxp3 was evaluated by intracellularstaining using an Ab against Foxp3 and analyzed by FACS (Fig.2D). In agreement with RT-PCR results, FACS analysis demon-strated increased protein expression levels of Foxp3 in T cellscultured in TRAMP-C2 CM (CM) as the Ctrl 25�; however,CD4�CD25� T cells cultured with T cell medium (Ctrl) did notexpress Foxp3 (Fig. 2D, bottom panel).
To study the dilution effect of CM on the conversion, 0.01, 0.1,0.25, 0.5, and 0.9 ml of TRAMP-C2 CM were diluted in T cellmedium in a final volume of 1 ml. Purified CD4�CD25� T cellswere cultured in the various diluted CM for 5 days and analyzed byFACS to examine CD25 expression levels. Fig. 2E shows a rep-resentative FACS profile of the conversion. The starting purifiedCD4�CD25� T cell population is shown as day 0. The graph onthe bottom represents the percentage of CD4�CD25� T cells cul-tured in various diluted CM. The results suggest the conversion ofCD4�CD25� T cells into CD4�CD25� T cells is in a CM con-centration-dependent manner. Taken together, these results indi-cate that cell-cell contact is not required because TRAMP-C2 CMwas sufficient to generate Foxp3-expressing T cells, and this con-version is in a CM concentration-dependent manner. In addition,because Foxp3 is exclusively expressed in Treg cells, results fromthis study suggest that TRAMP-C2 cell CM, but not NRP-152 CM,
can generate Treg cells from CD4�CD25� T cells. All of the fol-lowing experiments were conducted with CM.
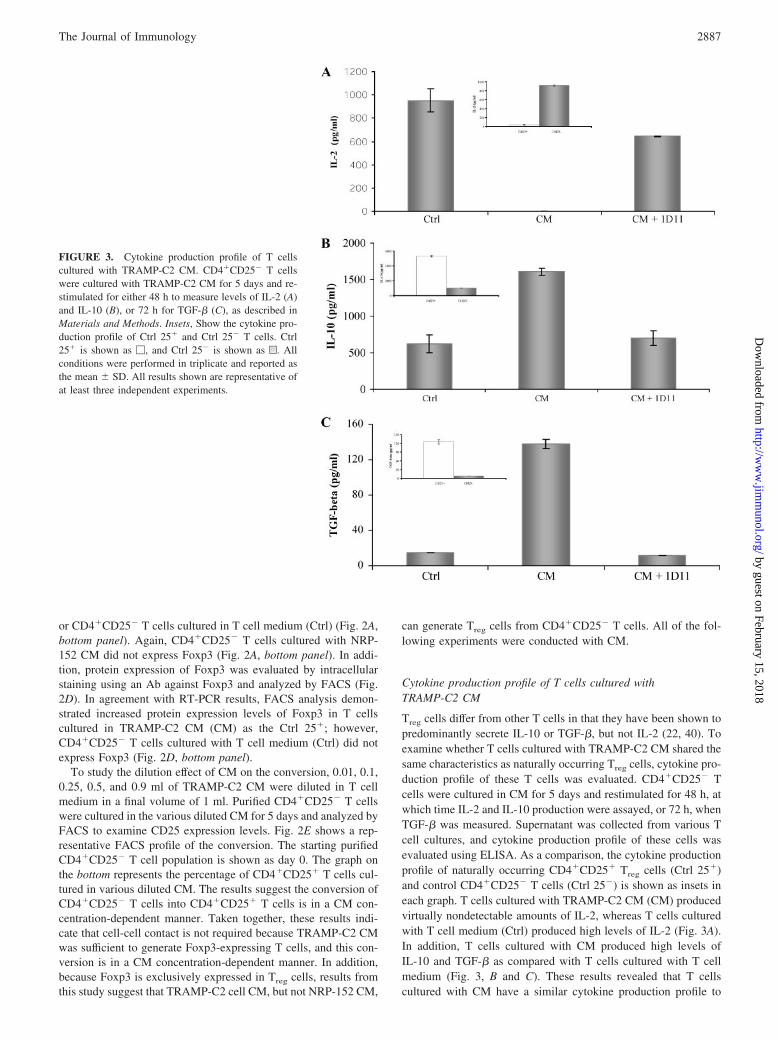
Cytokine production profile of T cells cultured withTRAMP-C2 CM
Treg cells differ from other T cells in that they have been shown topredominantly secrete IL-10 or TGF-�, but not IL-2 (22, 40). Toexamine whether T cells cultured with TRAMP-C2 CM shared thesame characteristics as naturally occurring Treg cells, cytokine pro-duction profile of these T cells was evaluated. CD4�CD25� Tcells were cultured in CM for 5 days and restimulated for 48 h, atwhich time IL-2 and IL-10 production were assayed, or 72 h, whenTGF-� was measured. Supernatant was collected from various Tcell cultures, and cytokine production profile of these cells wasevaluated using ELISA. As a comparison, the cytokine productionprofile of naturally occurring CD4�CD25� Treg cells (Ctrl 25�)and control CD4�CD25� T cells (Ctrl 25�) is shown as insets ineach graph. T cells cultured with TRAMP-C2 CM (CM) producedvirtually nondetectable amounts of IL-2, whereas T cells culturedwith T cell medium (Ctrl) produced high levels of IL-2 (Fig. 3A).In addition, T cells cultured with CM produced high levels ofIL-10 and TGF-� as compared with T cells cultured with T cellmedium (Fig. 3, B and C). These results revealed that T cellscultured with CM have a similar cytokine production profile to
FIGURE 3. Cytokine production profile of T cellscultured with TRAMP-C2 CM. CD4�CD25� T cellswere cultured with TRAMP-C2 CM for 5 days and re-stimulated for either 48 h to measure levels of IL-2 (A)and IL-10 (B), or 72 h for TGF-� (C), as described inMaterials and Methods. Insets, Show the cytokine pro-duction profile of Ctrl 25� and Ctrl 25� T cells. Ctrl25� is shown as �, and Ctrl 25� is shown as u. Allconditions were performed in triplicate and reported asthe mean � SD. All results shown are representative ofat least three independent experiments.
2887The Journal of Immunology
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
naturally occurring Treg cells, and these similarities provided evi-dence to further support the notion that tumor cells convertCD4�CD25� T cells to Treg cells (22, 40).
T cells cultured with tumor cell CM suppress CD4�CD25�
T cell proliferation
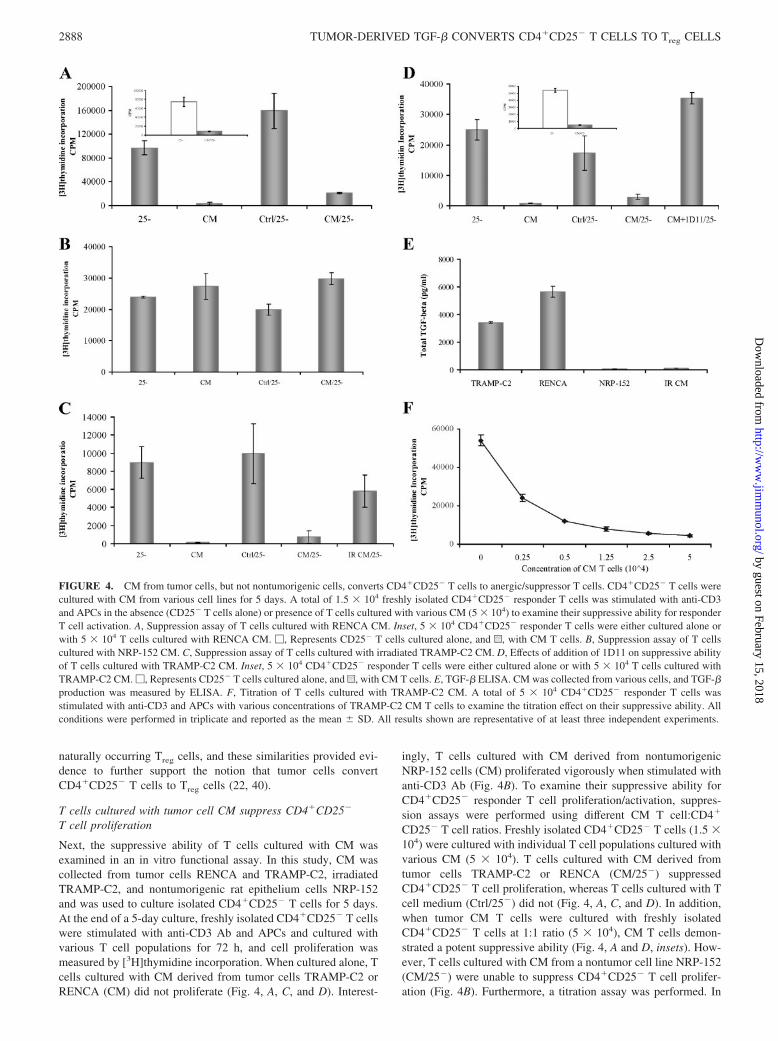
Next, the suppressive ability of T cells cultured with CM wasexamined in an in vitro functional assay. In this study, CM wascollected from tumor cells RENCA and TRAMP-C2, irradiatedTRAMP-C2, and nontumorigenic rat epithelium cells NRP-152and was used to culture isolated CD4�CD25� T cells for 5 days.At the end of a 5-day culture, freshly isolated CD4�CD25� T cellswere stimulated with anti-CD3 Ab and APCs and cultured withvarious T cell populations for 72 h, and cell proliferation wasmeasured by [3H]thymidine incorporation. When cultured alone, Tcells cultured with CM derived from tumor cells TRAMP-C2 orRENCA (CM) did not proliferate (Fig. 4, A, C, and D). Interest-
ingly, T cells cultured with CM derived from nontumorigenicNRP-152 cells (CM) proliferated vigorously when stimulated withanti-CD3 Ab (Fig. 4B). To examine their suppressive ability forCD4�CD25� responder T cell proliferation/activation, suppres-sion assays were performed using different CM T cell:CD4�
CD25� T cell ratios. Freshly isolated CD4�CD25� T cells (1.5 �104) were cultured with individual T cell populations cultured withvarious CM (5 � 104). T cells cultured with CM derived fromtumor cells TRAMP-C2 or RENCA (CM/25�) suppressedCD4�CD25� T cell proliferation, whereas T cells cultured with Tcell medium (Ctrl/25�) did not (Fig. 4, A, C, and D). In addition,when tumor CM T cells were cultured with freshly isolatedCD4�CD25� T cells at 1:1 ratio (5 � 104), CM T cells demon-strated a potent suppressive ability (Fig. 4, A and D, insets). How-ever, T cells cultured with CM from a nontumor cell line NRP-152(CM/25�) were unable to suppress CD4�CD25� T cell prolifer-ation (Fig. 4B). Furthermore, a titration assay was performed. In
FIGURE 4. CM from tumor cells, but not nontumorigenic cells, converts CD4�CD25� T cells to anergic/suppressor T cells. CD4�CD25� T cells werecultured with CM from various cell lines for 5 days. A total of 1.5 � 104 freshly isolated CD4�CD25� responder T cells was stimulated with anti-CD3and APCs in the absence (CD25� T cells alone) or presence of T cells cultured with various CM (5 � 104) to examine their suppressive ability for responderT cell activation. A, Suppression assay of T cells cultured with RENCA CM. Inset, 5 � 104 CD4�CD25� responder T cells were either cultured alone orwith 5 � 104 T cells cultured with RENCA CM. �, Represents CD25� T cells cultured alone, and u, with CM T cells. B, Suppression assay of T cellscultured with NRP-152 CM. C, Suppression assay of T cells cultured with irradiated TRAMP-C2 CM. D, Effects of addition of 1D11 on suppressive abilityof T cells cultured with TRAMP-C2 CM. Inset, 5 � 104 CD4�CD25� responder T cells were either cultured alone or with 5 � 104 T cells cultured withTRAMP-C2 CM. �, Represents CD25� T cells cultured alone, and u, with CM T cells. E, TGF-� ELISA. CM was collected from various cells, and TGF-�production was measured by ELISA. F, Titration of T cells cultured with TRAMP-C2 CM. A total of 5 � 104 CD4�CD25� responder T cells wasstimulated with anti-CD3 and APCs with various concentrations of TRAMP-C2 CM T cells to examine the titration effect on their suppressive ability. Allconditions were performed in triplicate and reported as the mean � SD. All results shown are representative of at least three independent experiments.
2888 TUMOR-DERIVED TGF-� CONVERTS CD4�CD25� T CELLS TO Treg CELLS
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
this study, 5 � 104 freshly isolated CD4�CD25� T cells werestimulated with anti-CD3 Ab and cultured with different numbersof TRAMP-C2 CM-cultured T cells ranging from 0 to 5 � 104.CM T cells showed a potent suppressive ability of CD4�CD25� Tcell proliferation in a dose-dependent manner (Fig. 4F). These re-sults strongly suggest that tumor cells, but not nontumorigeniccells, can indeed directly convert CD4�CD25� T cells into Treg
cells because these T cells demonstrated a potent suppressive abil-ity in the in vitro suppression assay.
The role of tumor cell-derived TGF-� in convertingCD4�CD25� T cells to Treg cells
Many types of tumor cells produce high levels of TGF-�, andexogenously added TGF-� can convert CD4�CD25� T cells intoTreg cells (35, 41–43). Consequently, levels of TGF-� present inthe CM from RENCA, TRAMP-C2, irradiated TRAMP-C2, and
NPR-152 cells were examined using a TGF-� ELISA. The resultsindicated that tumor cells produced high levels of TGF-�, whereas thenontumorigenic NRP-152 and irradiated TRAMP-C2 cells had sig-nificantly lower TGF-� production (Fig. 4E). To elucidate the mech-anism of Treg cell conversion and the potential role of tumor-derivedTGF-�, a neutralizing Ab against TGF-� 1D11 was used. This Abrecognizes TGF-�1, 2, and 3 in human and mouse and neutralizesbiological activities of TGF-� (44). When 1D11 was added toTRAMP-C2 CM, T cells cultured with CM did not express Fopx3(Fig. 2A, bottom panel). In addition, T cells cultured with NRP-152CM did not express Foxp3 because NRP-152 CM had little TGF-�expression (Fig. 4E). Furthermore, neutralization of TGF-� com-pletely changed the profile of cytokine production of these T cells, andcells now expressed high levels of IL-2 and low levels of IL-10 andTGF-� (Fig. 3). More importantly, in the suppression assay, additionof 1D11 abrogated the ability of TRAMP-C2 CM-cultured T cells to
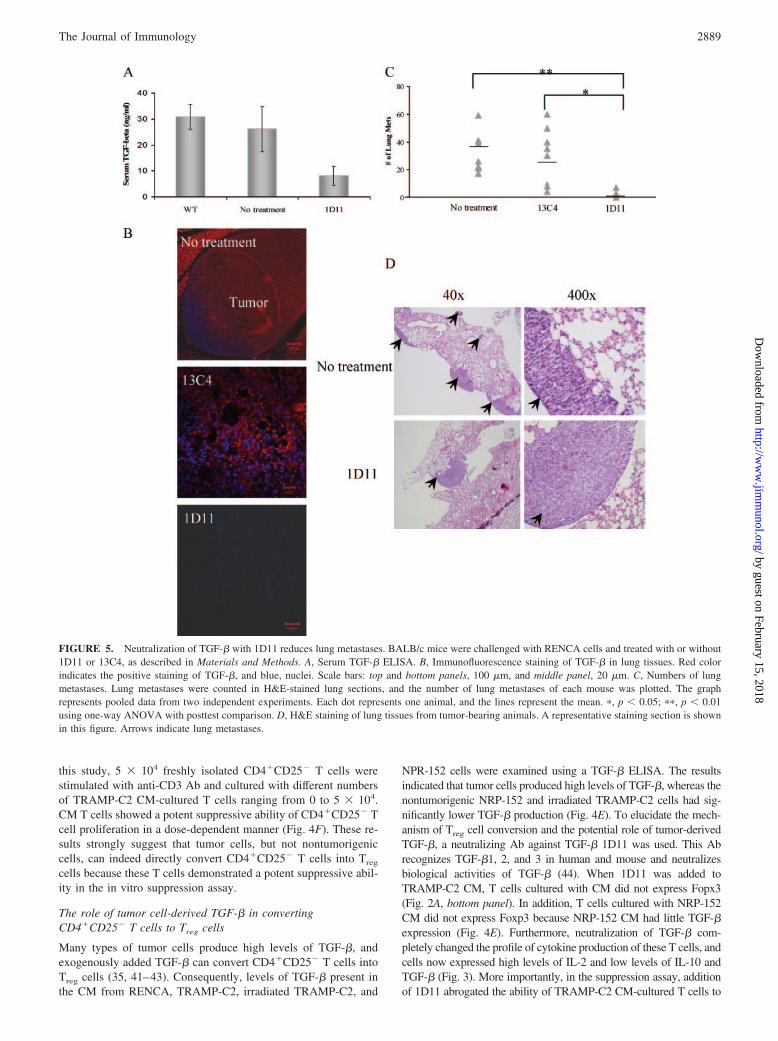
FIGURE 5. Neutralization of TGF-� with 1D11 reduces lung metastases. BALB/c mice were challenged with RENCA cells and treated with or without1D11 or 13C4, as described in Materials and Methods. A, Serum TGF-� ELISA. B, Immunofluorescence staining of TGF-� in lung tissues. Red colorindicates the positive staining of TGF-�, and blue, nuclei. Scale bars: top and bottom panels, 100 �m, and middle panel, 20 �m. C, Numbers of lungmetastases. Lung metastases were counted in H&E-stained lung sections, and the number of lung metastases of each mouse was plotted. The graphrepresents pooled data from two independent experiments. Each dot represents one animal, and the lines represent the mean. �, p � 0.05; ��, p � 0.01using one-way ANOVA with posttest comparison. D, H&E staining of lung tissues from tumor-bearing animals. A representative staining section is shownin this figure. Arrows indicate lung metastases.
2889The Journal of Immunology
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
suppress CD4�CD25� T cell proliferation (Fig. 4D). Finally,CD4�CD25� T cells were cultured with irradiated TRAMP-C2 CM,because TGF-� production was significantly reduced in irradiatedCM (Fig. 4E). As shown in Fig. 4C, the resulting T cells were unableto suppress CD4�CD25� T cell proliferation. Taken together, theseresults have revealed a critical role of tumor-derived TGF-� in con-version of CD4�CD25� T cells into Treg cells. Noticeably, nontu-morigenic NRP-152 cells produce low levels of TGF-� (Fig. 4E) (45),and T cells cultured with CM derived from these cells were unable tosuppress T cell proliferation (Fig. 4B). Collectively, these studiesdemonstrate that tumor cells convert CD4�CD25� T cells into Treg
cells through the production of high levels of TGF-�, whereas non-tumorigenic cells NRP-152 and irradiated TRAMP-C2 were unable toconvert such cells due to the low levels of TGF-� production.
Neutralization of TGF-� reduces the tumor burden in mice
To support the above in vitro observations, an in vivo study wasconducted using the mouse renal cell carcinoma RENCA becausethey produce high levels of TGF-� (Fig. 4E). BALB/c mice wereinjected with 1 � 106 RENCA cells i.v. via the tail vein. On theday of injection, five mice received 1D11 and five mice receivedcontrol Ab13C4 at a loading dose of 100 mg/kg. These mice con-tinued receiving 1D11 or 13C4 at a dose of 50 mg/kg every 3 daysfor a course of 30 days. At the end of the experiment, all animalswere sacrificed, TGF-� levels were measured in serum and lungtissues, and tumor burden was evaluated in all animals. TGF-�ELISA revealed that serum levels of TGF-� were significantlyreduced in 1D11-treated animals, whereas serum from untreatedand 13C4-treated animals contained levels of TGF-� that werecomparable to those of tumor-free animals (Fig. 5A and data notshown). In addition, TGF-� expression in lung tissues was eval-uated. Lung sections from animals were stained with an anti-TGF-� Ab. Fig. 5B shows the TGF-� expression profile of theseanimals. In tumor-bearing mice receiving 13C4 or no treatment,TGF-� was ubiquitously expressed in tumor and lung tissues,whereas no TGF-� expression was detected in lung tissues of micereceiving 1D11. Analysis of tumor burden revealed that animalsreceiving 1D11 had substantially fewer tumor metastases in thelung tissues as compared with those receiving 13C4 or no treat-ment (Fig. 5, C and D). These results demonstrate the efficacy of1D11 in neutralizing TGF-� in treated animals, and such treatmentreduces the tumor burden.
Neutralization of TGF-� abrogates the conversion of Treg cellsin the tumor microenvironment
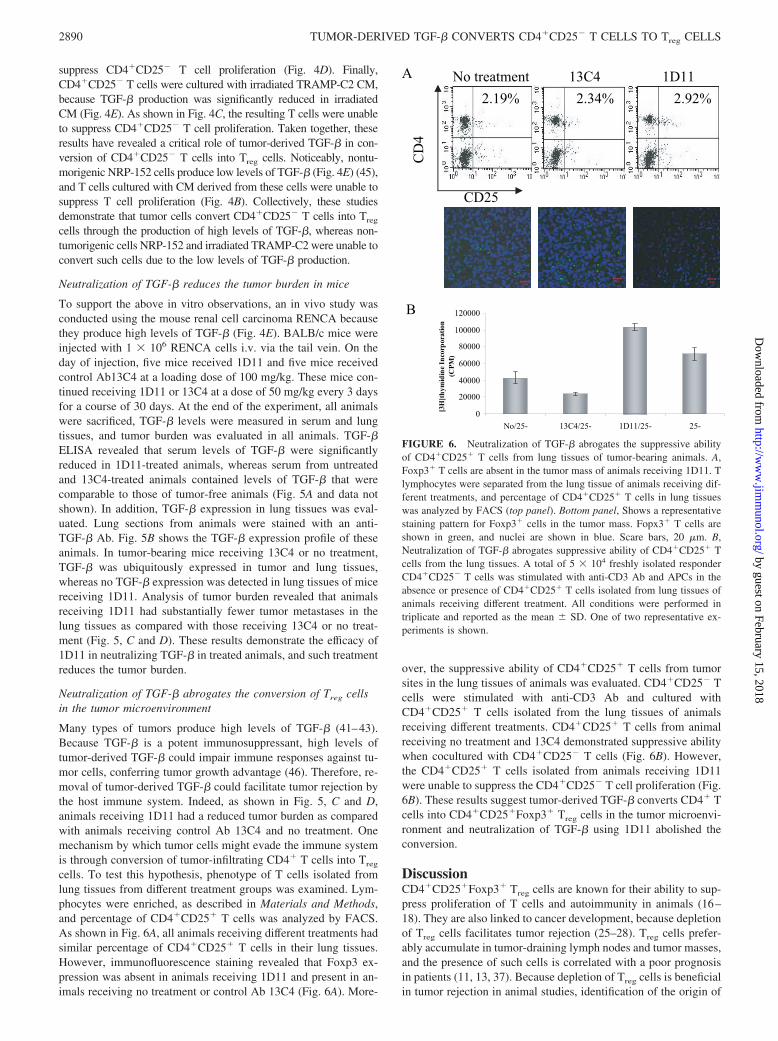
Many types of tumors produce high levels of TGF-� (41–43).Because TGF-� is a potent immunosuppressant, high levels oftumor-derived TGF-� could impair immune responses against tu-mor cells, conferring tumor growth advantage (46). Therefore, re-moval of tumor-derived TGF-� could facilitate tumor rejection bythe host immune system. Indeed, as shown in Fig. 5, C and D,animals receiving 1D11 had a reduced tumor burden as comparedwith animals receiving control Ab 13C4 and no treatment. Onemechanism by which tumor cells might evade the immune systemis through conversion of tumor-infiltrating CD4� T cells into Treg
cells. To test this hypothesis, phenotype of T cells isolated fromlung tissues from different treatment groups was examined. Lym-phocytes were enriched, as described in Materials and Methods,and percentage of CD4�CD25� T cells was analyzed by FACS.As shown in Fig. 6A, all animals receiving different treatments hadsimilar percentage of CD4�CD25� T cells in their lung tissues.However, immunofluorescence staining revealed that Foxp3 ex-pression was absent in animals receiving 1D11 and present in an-imals receiving no treatment or control Ab 13C4 (Fig. 6A). More-
over, the suppressive ability of CD4�CD25� T cells from tumorsites in the lung tissues of animals was evaluated. CD4�CD25� Tcells were stimulated with anti-CD3 Ab and cultured withCD4�CD25� T cells isolated from the lung tissues of animalsreceiving different treatments. CD4�CD25� T cells from animalreceiving no treatment and 13C4 demonstrated suppressive abilitywhen cocultured with CD4�CD25� T cells (Fig. 6B). However,the CD4�CD25� T cells isolated from animals receiving 1D11were unable to suppress the CD4�CD25� T cell proliferation (Fig.6B). These results suggest tumor-derived TGF-� converts CD4� Tcells into CD4�CD25�Foxp3� Treg cells in the tumor microenvi-ronment and neutralization of TGF-� using 1D11 abolished theconversion.
DiscussionCD4�CD25�Foxp3� Treg cells are known for their ability to sup-press proliferation of T cells and autoimmunity in animals (16–18). They are also linked to cancer development, because depletionof Treg cells facilitates tumor rejection (25–28). Treg cells prefer-ably accumulate in tumor-draining lymph nodes and tumor masses,and the presence of such cells is correlated with a poor prognosisin patients (11, 13, 37). Because depletion of Treg cells is beneficialin tumor rejection in animal studies, identification of the origin of
B
A
0
20000
40000
60000
80000
100000
120000
No/25- 13C4/25- 1D11/25- 25-
noitaroprocnIenidimyht]
H3[)
M PC(
No treatment 13C4 1D11
4D
C
CD25
2.19% 2.34% 2.92%
FIGURE 6. Neutralization of TGF-� abrogates the suppressive abilityof CD4�CD25� T cells from lung tissues of tumor-bearing animals. A,Foxp3� T cells are absent in the tumor mass of animals receiving 1D11. Tlymphocytes were separated from the lung tissue of animals receiving dif-ferent treatments, and percentage of CD4�CD25� T cells in lung tissueswas analyzed by FACS (top panel). Bottom panel, Shows a representativestaining pattern for Foxp3� cells in the tumor mass. Fopx3� T cells areshown in green, and nuclei are shown in blue. Scare bars, 20 �m. B,Neutralization of TGF-� abrogates suppressive ability of CD4�CD25� Tcells from the lung tissues. A total of 5 � 104 freshly isolated responderCD4�CD25� T cells was stimulated with anti-CD3 Ab and APCs in theabsence or presence of CD4�CD25� T cells isolated from lung tissues ofanimals receiving different treatment. All conditions were performed intriplicate and reported as the mean � SD. One of two representative ex-periments is shown.
2890 TUMOR-DERIVED TGF-� CONVERTS CD4�CD25� T CELLS TO Treg CELLS
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Treg cells in tumor microenvironment will provide valuable infor-mation to successfully eliminate Treg cells, and thus the suppres-sion of antitumor immunity by these cells (27, 47, 48). To date, theorigin of Treg cells in the tumor microenvironment still remainselusive; however, recent evidence has suggested at least four pos-sible sources of these Treg cells (39). First is trafficking. Tumorcells produce chemokines such as CCL22 that specifically recruitsTreg cells to tumor sites (11). Second is differentiation throughcontacting with tumor-induced immature DCs (38). Third is ex-pansion of Treg cells through DC stimulation (49). Finally, Treg
cells can also be converted from normal CD4� T cells in tumor-bearing animals (37). It appears that there are two distinct types ofTreg cells in the tumor microenvironment. One is naturally occur-ring Treg cells that are either recruited to the tumor sites or stim-ulated to expand, and the other is through conversion of normalCD4� T cells into Treg cells. Although the phenotype of these twotypes of Treg cells is virtually indistinguishable (50), the origin ofthese Treg cells differs, as described above. TGF-� plays a criticalrole in extrathymic generation/conversion of Treg cells eitherthrough direct or indirect mechanisms. The direct conversion ofTreg cells by tumor-derived TGF-� is demonstrated in this study,whereas the indirect mechanism involves induction of immatureDCs by tumor-derived TGF-�, which in turn increase Treg cellconversion (38). Because neither direct expansion of naturally oc-curring Treg cells by DCs nor trafficking of such cells to the tumormicroenvironment requires TGF-� (11, 49), and because a directtreatment with TGF-� induces Treg cell conversion and neutral-ization of TGF-� reduced Treg cells in the tumor microenviron-ment (Fig. 6), it is logical to reason that CD4�CD25�Foxp3� Treg
cells observed in the tumor masses, at least some of them, areconverted by tumor-produced TGF-�. In addition, they possesssuppressive ability of immune cells, thus conferring tumors’ ad-vantage to evade the immune system.
T cells obtained from the in vitro conversion studies are bona fideTreg cells in terms of Foxp3 expression, cytokine production (lowIL-2, high IL-10, and TGF-�), and suppression of CD4�CD25� Tcells (Figs. 2–4). Additional studies revealed a critical role of TGF-�in the conversion because tumor cells (TRAMP-C2 and RENCA)produce high levels of this cytokine, and neutralization of TGF-�completely abrogated the conversion of CD4�CD25� T cells to Treg
cells (Figs. 2–4). The importance of TGF-� in conversion was furtherdemonstrated by the studies of CD4�CD25� T cells cultured in CMderived from a nontumorigenic cell line NRP-152 and irradiatedTRAMP-C2, because these cells lack all the characteristics of Treg
cells due to the absence of TGF-� in CM (Figs. 2–4).Tumor cells are a rich source of TGF-� in the tumor microen-
vironment, and tumor-derived TGF-� becomes a source that in-fluences conversion of normal CD4� T cells into Treg cells. In-deed, studies by Kretschmer et al. (51) have demonstrated thataddition of TGF-� enhances the conversion of naive CD4� T cellsinto Treg cells when stimulated by a minute Ag dose with subop-timal DC activation. The present study suggests that Treg cells inthe tumor sites, at least some of them, are converted from normalCD4� T cells by tumor-derived TGF-�. We thus propose thattumor cells evade the immune system by conversion of Treg cellsthrough production of high levels of TGF-�. When tumor-infil-trating CD4� T cells are exposed to TGF-�-rich environment intumor sites, CD4�CD25�Foxp3� Treg cell conversion occurs. Asa result, the converted Treg cells suppress antitumor immunity,conferring tumor cell advantages to evade the immune system.Indeed, a significant reduction in lung metastases was observed intumor-bearing animals treated with a neutralizing Ab againstTGF-� 1D11 (Fig. 5, C and D). This reduction in tumor burden, atleast in part, was due to the absence of CD4�CD25�Foxp3� Treg
cells in the tumor microenvironment of these animals (Fig. 6A).The absence of Foxp3� Treg cells is most likely due to the lack ofconversion of normal CD4� T cells because of the absence ofTGF-� in these animals. Moreover, it is of interest to point out thatTGF-� produced by host tissues also could play a role in Treg cellconversion, as demonstrated by Thomas and Massague (52) thatblockage of host-produced TGF-� aids tumor rejection. In theirstudies, mice rejected tumor cells engineered to produce a solubleform of TGF-� type II receptor that systemically neutralizesTGF-�, whereas mice injected with tumor cells producing minimallevels of TGF-� eventually succumbed to tumor burden, suggest-ing host-produced TGF-� could also play an important role intumor rejection and as an alternative source of TGF-� in Treg cellconversion.
The present and other recent studies have revealed varioussources of Treg cells present in the tumor microenvironment. How-ever, the possibility still remains that Treg cells observed in thetumor mass in the present study, at least some of them, were re-cruited by tumor-derived CCL22, as demonstrated by Curiel et al.(11). It is likely that the population of Treg cells present in thetumor microenvironment consists of a heterogeneous mixture ofnaturally occurring Treg cells and converted Treg cells. Cytokinesproduced by tumor cells, such as CCL22, IL-10, and TGF-�, canwork in concert to aid tumors in evading the immune system,either by recruitment of naturally occurring Treg cells to tumorsites or through direct or indirect conversion of normal CD4� Tcells to Treg cells (11, 37–39). Furthermore, it has been shown thatfunctional CD4�CD25� Treg cells can be expanded by DCs (49).Finally, Treg cell conversion could also occur independent ofTGF-�, as shown in the study by Kretschmer et al. (51) that T cellswith Ag specificity can be efficiently induced to Treg cells withminute amount of Ag stimulation along with suboptimal DC acti-vation. However, addition of TGF-� further enhances such con-version. It appears that extrathymic generation/conversion of Treg
cells could be Ag specific or nonspecific; in either case, TGF-�plays an essential role in this conversion. It is therefore importantto identify the source of Treg cells in the tumor mass to provide abetter understanding and more efficient measurement in treatmentof existing tumors, as depletion of CD25� T cells will not preventthe conversion of normal CD4� T cells into Treg cells by tumorcells. Combination of CD25� T cell depletion and blockage ofTGF-� should further improve the outcome of treatment of exist-ing tumors. However, because TGF-� is a very potent immuno-suppressant, long-term systematic blockage of TGF-� may resultin autoimmunity in patients; therefore, it is critical to find the del-icate balance between tumor immunity and autoimmunity that ismaintained by Treg cells in the treatment of tumors.
In summary, this study provides evidence that tumor cells candirectly convert normal CD4� T cells into CD4�CD25�Foxp3�
Treg cells through production of TGF-�, thus providing a possiblemechanism whereby tumors evade the host immune surveillanceprogram through conversion of Treg cells.
AcknowledgmentsWe thank Dr. Mary Paniagua and Jeffrey Nelson for assistance with flowcytometry, Dr. Adam Kohm for assistance with T cell proliferation assay,and Dr. Teng-Leong Chew for assistance with microscopy.
DisclosuresThe authors have no financial conflict of interest.
References1. Burnet, F. M. 1970. The concept of immunological surveillance. Prog. Exp. Tu-
mor Res. 13: 1–27.
2891The Journal of Immunology
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

2. Yee, C., J. A. Thompson, D. Byrd, S. R. Riddell, P. Roche, E. Celis, andP. D. Greenberg. 2002. Adoptive T cell therapy using antigen-specific CD8� Tcell clones for the treatment of patients with metastatic melanoma: in vivo per-sistence, migration, and antitumor effect of transferred T cells. Proc. Natl. Acad.Sci. USA 99: 16168–16173.
3. Rosenberg, S. A. 2001. Progress in the development of immunotherapy for thetreatment of patients with cancer. J. Intern. Med. 250: 462–475.
4. Rosenberg, S. A., J. R. Yannelli, J. C. Yang, S. L. Topalian, D. J.Schwartzentruber, J. S. Weber, D. R. Parkinson, C. A. Seipp, J. H. Einhorn, andD. E. White. 1994. Treatment of patients with metastatic melanoma with autol-ogous tumor-infiltrating lymphocytes and interleukin 2. J. Natl. Cancer Inst. 86:1159–1166.
5. Boon, T., and P. van der Bruggen. 1996. Human tumor antigens recognized by Tlymphocytes. J. Exp. Med. 183: 725–729.
6. Dudley, M. E., J. R. Wunderlich, P. F. Robbins, J. C. Yang, P. Hwu,D. J. Schwartzentruber, S. L. Topalian, R. Sherry, N. P. Restifo, A. M. Hubicki,et al. 2002. Cancer regression and autoimmunity in patients after clonal repopu-lation with antitumor lymphocytes. Science 298: 850–854.
7. Zhang, L., J. R. Conejo-Garcia, D. Katsaros, P. A. Gimotty, M. Massobrio,G. Regnani, A. Makrigiannakis, H. Gray, K. Schlienger, M. N. Liebman, et al.2003. Intratumoral T cells, recurrence, and survival in epithelial ovarian cancer.N. Engl. J. Med. 348: 203–213.
8. Clemente, C. G., M. C. Mihm, Jr., R. Bufalino, S. Zurrida, P. Collini, andN. Cascinelli. 1996. Prognostic value of tumor infiltrating lymphocytes in thevertical growth phase of primary cutaneous melanoma. Cancer 77: 1303–1310.
9. Clark, W. H., Jr., D. E. Elder, D. Guerry IV, L. E. Braitman, B. J. Trock,D. Schultz, M. Synnestvedt, and A. C. Halpern. 1989. Model predicting survivalin stage I melanoma based on tumor progression. J. Natl. Cancer Inst. 81:1893–1904.
10. Nakano, O., M. Sato, Y. Naito, K. Suzuki, S. Orikasa, M. Aizawa, Y. Suzuki,I. Shintaku, H. Nagura, and H. Ohtani. 2001. Proliferative activity of intratumoralCD8� T-lymphocytes as a prognostic factor in human renal cell carcinoma: clin-icopathologic demonstration of antitumor immunity. Cancer Res. 61: 5132–5136.
11. Curiel, T. J., G. Coukos, L. Zou, X. Alvarez, P. Cheng, P. Mottram, M.Evdemon-Hogan, J. R. Conejo-Garcia, L. Zhang, M. Burow, et al. 2004. Specificrecruitment of regulatory T cells in ovarian carcinoma fosters immune privilegeand predicts reduced survival. Nat. Med. 10: 942–949.
12. Schumacher, K., W. Haensch, C. Roefzaad, and P.M. Schlag. 2001. Prognosticsignificance of activated CD8� T cell infiltrations within esophageal carcinomas.Cancer Res. 61: 3932–3936.
13. Sasada, T., M. Kimura, Y. Yoshida, M. Kanai, and A. Takabayashi. 2003.CD4�CD25� regulatory T cells in patients with gastrointestinal malignancies:possible involvement of regulatory T cells in disease progression. Cancer 98:1089–1099.
14. Marshall, N. A., L. E. Christie, L. R. Munro, D. J. Culligan, P. W. Johnston,R. N. Barker, and M. A. Vickers. 2004. Immunosuppressive regulatory T cells areabundant in the reactive lymphocytes of Hodgkin lymphoma. Blood 103:1755–1762.
15. Liyanage, U. K., T. T. Moore, H. G. Joo, Y. Tanaka, V. Herrmann, G. Doherty,J. A. Drebin, S. M. Strasberg, T. J. Eberlein, P. S. Goedegebuure, andD. C. Linehan. 2002. Prevalence of regulatory T cells is increased in peripheralblood and tumor microenvironment of patients with pancreas or breast adeno-carcinoma. J. Immunol. 169: 2756–2761.
16. Suri-Payer, E., and H. Cantor. 2001. Differential cytokine requirements forregulation of autoimmune gastritis and colitis by CD4�CD25� T cells.J. Autoimmun. 16: 115–123.
17. Seddon, B., and D. Mason. 1999. Regulatory T cells in the control of autoim-munity: the essential role of transforming growth factor � and interleukin 4 in theprevention of autoimmune thyroiditis in rats by peripheral CD4�CD45RC� cellsand CD4�CD8� thymocytes. J. Exp. Med. 189: 279–288.
18. Asseman, C., S. Mauze, M. W. Leach, R. L. Coffman, and F. Powrie. 1999. Anessential role for interleukin 10 in the function of regulatory T cells that inhibitintestinal inflammation. J. Exp. Med. 190: 995–1004.
19. Murakami, M., A. Sakamoto, J. Bender, J. Kappler, and P. Marrack. 2002.CD25�CD4� T cells contribute to the control of memory CD8� T cells. Proc.Natl. Acad. Sci. USA 99: 8832–8837.
20. Shimizu, J., S. Yamazaki, T. Takahashi, Y. Ishida, and S. Sakaguchi. 2002. Stim-ulation of CD25�CD4� regulatory T cells through GITR breaks immunologicalself-tolerance. Nat. Immunol. 3: 135–142.
21. Read, S., V. Malmstrom, and F. Powrie. 2000. Cytotoxic T lymphocyte-associ-ated antigen 4 plays an essential role in the function of CD25�CD4� regulatorycells that control intestinal inflammation. J. Exp. Med. 192: 295–302.
22. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell de-velopment by the transcription factor Foxp3. Science 299: 1057–1061.
23. Asano, M., M. Toda, N. Sakaguchi, and S. Sakaguchi. 1996. Autoimmune diseaseas a consequence of developmental abnormality of a T cell subpopulation. J. Exp.Med. 184: 387–396.
24. Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh, and M. Toda. 1995. Immuno-logic self-tolerance maintained by activated T cells expressing IL-2 receptor�-chains (CD25): breakdown of a single mechanism of self-tolerance causesvarious autoimmune diseases. J. Immunol. 155: 1151–1164.
25. Shimizu, J., S. Yamazaki, and S. Sakaguchi. 1999. Induction of tumor immunityby removing CD25�CD4� T cells: a common basis between tumor immunityand autoimmunity. J. Immunol. 163: 5211–5218.
26. Golgher, D., E. Jones, F. Powrie, T. Elliott, and A. Gallimore. 2002. Depletion ofCD25� regulatory cells uncovers immune responses to shared murine tumorrejection antigens. Eur. J. Immunol. 32: 3267–3275.
27. Tanaka, H., J. Tanaka, J. Kjaergaard, and S. Shu. 2002. Depletion of CD4�
CD25� regulatory cells augments the generation of specific immune T cells intumor-draining lymph nodes. J. Immunother. 25: 207–217.
28. Akbar, A. N., L. S. Taams, M. Salmon, and M. Vukmanovic-Stejic. 2003. Theperipheral generation of CD4� CD25� regulatory T cells. Immunology 109:319–325.
29. Bluestone, J. A., and A. K. Abbas. 2003. Natural versus adaptive regulatory Tcells. Nat. Rev. Immunol. 3: 253–257.
30. Woo, E. Y., H. Yeh, C. S. Chu, K. Schlienger, R. G. Carroll, J. L. Riley,L. R. Kaiser, and C. H. June. 2002. Cutting edge: regulatory T cells from lungcancer patients directly inhibit autologous T cell proliferation. J. Immunol. 168:4272–4276.
31. Woo, E. Y., C. S. Chu, T. J. Goletz, K. Schlienger, H. Yeh, G. Coukos,S. C. Rubin, L. R. Kaiser, and C. H. June. 2001. Regulatory CD4�CD25� T cellsin tumors from patients with early-stage non-small cell lung cancer and late-stageovarian cancer. Cancer Res. 61: 4766–4772.
32. Ormandy, L. A., T. Hillemann, H. Wedemeyer, M. P. Manns, T. F. Greten, andF. Korangy. 2005. Increased populations of regulatory T cells in peripheral bloodof patients with hepatocellular carcinoma. Cancer Res. 65: 2457–2464.
33. Wolf, A. M., D. Wolf, M. Steurer, G. Gastl, E. Gunsilius, and B. Grubeck-Loebenstein. 2003. Increase of regulatory T cells in the peripheral blood of cancerpatients. Clin. Cancer Res. 9: 606–612.
34. Itoh, M., T. Takahashi, N. Sakaguchi, Y. Kuniyasu, J. Shimizu, F. Otsuka, andS. Sakaguchi. 1999. Thymus and autoimmunity: production of CD25�CD4�
naturally anergic and suppressive T cells as a key function of the thymus inmaintaining immunologic self-tolerance. J. Immunol. 162: 5317–5326.
35. Chen, W., W. Jin, N. Hardegen, K. J. Lei, L. Li, N. Marinos, G. McGrady, andS. M. Wahl. 2003. Conversion of peripheral CD4�CD25� naive T cells toCD4�CD25� regulatory T cells by TGF-� induction of transcription factorFoxp3. J. Exp. Med. 198: 1875–1886.
36. Zheng, S. G., J. D. Gray, K. Ohtsuka, S. Yamagiwa, and D. A. Horwitz. 2002.Generation ex vivo of TGF-�-producing regulatory T cells from CD4�CD25�
precursors. J. Immunol. 169: 4183–4189.37. Valzasina, B., S. Piconese, C. Guiducci, and M. P. Colombo. 2006. Tumor-
induced expansion of regulatory T cells by conversion of CD4�CD25� lympho-cytes is thymus and proliferation independent. Cancer Res. 66: 4488–4495.
38. Ghiringhelli, F., P. E. Puig, S. Roux, A. Parcellier, E. Schmitt, E. Solary,G. Kroemer, F. Martin, B. Chauffert, and L. Zitvogel. 2005. Tumor cells convertimmature myeloid dendritic cells into TGF-�-secreting cells inducingCD4�CD25� regulatory T cell proliferation. J. Exp. Med. 202: 919–929.
39. Zou, W. 2006. Regulatory T cells, tumor immunity and immunotherapy. Nat.Rev. Immunol. 6: 295–307.
40. Nakamura, K., A. Kitani, and W. Strober. 2001. Cell contact-dependent immu-nosuppression by CD4�CD25� regulatory T cells is mediated by cell surface-bound transforming growth factor �. J. Exp. Med. 194: 629–644.
41. Chen, T. C., D. R. Hinton, V. W. Yong, and F. M. Hofman. 1997. TGF-B2 andsoluble p55 TNFR modulate VCAM-1 expression in glioma cells and brain de-rived endothelial cells. J. Neuroimmunol. 73: 155–161.
42. Kong, F. M., M. K. Washington, R. L. Jirtle, and M. S. Anscher. 1996. Plasmatransforming growth factor-�1 reflects disease status in patients with lung cancerafter radiotherapy: a possible tumor marker. Lung Cancer 16: 47–59.
43. Xu, J., A. Ahmad, J. F. Jones, R. Dolcetti, E. Vaccher, U. Prasad, and J. Menezes.2000. Elevated serum transforming growth factor �1 levels in Epstein-Barr virus-associated diseases and their correlation with virus-specific immunoglobulin A(IgA) and IgM. J. Virol. 74: 2443–2446.
44. Dasch, J. R., D. R. Pace, W. Waegell, D. Inenaga, and L. Ellingsworth. 1989.Monoclonal antibodies recognizing transforming growth factor-�: bioactivityneutralization and transforming growth factor �2 affinity purification. J. Immunol.142: 1536–1541.
45. Danielpour, D. 1999. Transdifferentiation of NRP-152 rat prostatic basal epithe-lial cells toward a luminal phenotype: regulation by glucocorticoid, insulin-likegrowth factor-I and transforming growth factor-�. J. Cell Sci. 112: 169–179.
46. Letterio, J. J., and A. B. Roberts. 1998. Regulation of immune responses byTGF-�. Annu. Rev. Immunol. 16: 137–161.
47. Steitz, J., J. Bruck, J. Lenz, J. Knop, and T. Tuting. 2001. Depletion of CD25�
CD4� T cells and treatment with tyrosinase-related protein 2-transduced den-dritic cells enhance the interferon �-induced, CD8� T-cell-dependent immunedefense of B16 melanoma. Cancer Res. 61: 8643–8646.
48. Jones, E., M. Dahm-Vicker, A. K. Simon, A. Green, F. Powrie, V. Cerundolo,and A. Gallimore. 2002. Depletion of CD25� regulatory cells results in suppres-sion of melanoma growth and induction of autoreactivity in mice. Cancer Immun.2: 1.
49. Yamazaki, S., T. Iyoda, K. Tarbell, K. Olson, K. Velinzon, K. Inaba, andR. M. Steinman. 2003. Direct expansion of functional CD25�CD4� regulatory Tcells by antigen-processing dendritic cells. J. Exp. Med. 198: 235–247.
50. Wing, K., Z. Fehervari, and S. Sakaguchi. 2006. Emerging possibilities in thedevelopment and function of regulatory T cells. Int. Immunol. 18: 991–1000.
51. Kretschmer, K., I. Apostolou, D. Hawiger, K. Khazaie, M. C. Nussenzweig, andH. von Boehmer. 2005. Inducing and expanding regulatory T cell populations byforeign antigen. Nat. Immunol. 6: 1219–1227.
52. Thomas, D. A., and J. Massague. 2005. TGF-� directly targets cytotoxic T cellfunctions during tumor evasion of immune surveillance. Cancer Cells 8:369–380.
53. Gershon, R. K., and K. Kondo. 1970. Cell interactions in the induction of toler-ance: the role of thymic lymphocytes. Immunology 18: 723–737.
54. Gershon, R. K., and K. Kondo. 1971. Infectious immunological tolerance. Im-munology 21: 903–914.
2892 TUMOR-DERIVED TGF-� CONVERTS CD4�CD25� T CELLS TO Treg CELLS
by guest on February 15, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
![élyi[1])toBracewell)[2,3] · Technical)Notes) Converting)Erdélyi[1])toBracewell)[2,3]! Erdélyidefinedhis!Hankel!transform!of!order!v!as![1]:! ( )( )1 2 0 gy f xJ xy xy dx(;) ()υ](https://static.fdocument.org/doc/165x107/5edc9e48ad6a402d66675c69/lyi1tobracewell23-technicalnotes-convertingerdlyi1tobracewell23.jpg)