
Title The inhibition of NLRP3 signaling attenuates liver ...
8
Title The inhibition of NLRP3 signaling attenuates liver injury in an α -galactosylceramide-induced hepatitis model( 本文(Fulltext) ) Author(s) 神戸, 歩 Report No.(Doctoral Degree) 博士(医学) 医博甲第1123号 Issue Date 2020-03-25 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/79317 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
Transcript of Title The inhibition of NLRP3 signaling attenuates liver ...

Title The inhibition of NLRP3 signaling attenuates liver injury in an α-galactosylceramide-induced hepatitis model( 本文(Fulltext) )
Author(s) 神戸, 歩
Report No.(DoctoralDegree) 博士(医学) 医博甲第1123号
Issue Date 2020-03-25
Type 博士論文
Version ETD
URL http://hdl.handle.net/20.500.12099/79317
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。

The inhibition of NLRP3 signaling attenuates liver injury in an a-galactosylceramide-induced hepatitis model
Ayumu Kanbe a, Hiroyasu Ito a, *, Yukari Omori a, Akira Hara b, Mitsuru Seishima a
a Department of Informative Clinical Medicine, Gifu University Graduate School of Medicine, 1-1 Yanagido, Gifu City 501-1194, Japanb Department of Tumor Pathology, Gifu University Graduate School of Medicine, 1-1 Yanagido, Gifu City 501-1194, Japan
a r t i c l e i n f o
Article history:Received 6 June 2017Accepted 12 June 2017Available online 14 June 2017
Keywords:Immune-mediated liver injuryInflammasomesNLRP3Alpha-galactosylceramide
a b s t r a c t
Inflammasomes are involved in innate immune responses. Several NOD-Like receptors (NLRs) participatein the formation of inflammasomes. NACHT, LRR and PYD domains-containing protein 3 (NALP3) belongsto the NLR family and recognizes adenosine triphosphate (ATP), crystals, and Reactive Oxygen Species(ROS). This study examined the effect of inflammasomes on alpha-galactosylceramide (GalCer)-inducedliver injury using NALP3-knockout (KO) mice. GalCer administration induced inflammasome activationand IL-1b-maturation. In NALP3-KO mice treated with GalCer, serum ALT levels were significantlyreduced compared with those in GalCer-treated WT mice. Histological examination revealed decreasednecrosis in NALP3-KO mice compared with WT mice, consistent with ALT levels. Expression of proin-flammatory cytokines (such as IL-6, and TNF-a) and chemokines was also significantly suppressed inNALP3-KO mice. Moreover, flow cytometry analysis revealed fewer infiltrating immune cells in the liversof GalCer-treated NALP3-KO mice. Inportantly, the frequency of MDSCs (CD11bþGr-1int cells), whichsuppress the immune response, was significantly increased in GalCer-treated NALP3-KO mice. Inconclusion, NALP3 inhibition attenuated liver injury in GalCer-induced hepatitis. The inhibition of NALP3signaling coused be a therapeutic target in immune-mediated liver injury.
© 2017 Elsevier Inc. All rights reserved.
1. Introduction
Autoimmune hepatitis (AIH) is an immune-mediated inflam-mation and liver injury of uncertain cause. It is characterized by aninfiltration of immune cells in the liver, an increase in g-globulins,antiantibodies, and HLA associations. Immunosuppressive therapysuch as treatment with corticosteroids is effective for AIH [1].Concanavalin A (Con A) has been widely used to model hostimmune-mediated liver injury [2]. Con A administration in vivoinduces lymphocyte activation and recruitment into the liver. TheCon A-induced hepatitis model has a unique pathogenesis as wellas characteristics of immune-mediated hepatitis in humans,including autoimmune hepatitis and acute viral hepatitis. Previousstudies demonstrated that natural killer T (NKT) cells are involvedin the development of Con A-induced hepatitis model. Deficiency ofNKT cells significantly reduced Con A-induced liver injury [3].Activation of NKT cells by alpha-galactosylceramide (GalCer) alsoinduces liver inflammation and injury [4,5]. Thus, AIH can be
studied in an immune-mediated liver injurymodel using Con A andGalCer.
The innate immune system is activated when pattern recogni-tion receptors (PRRs) detect a pathogen. The principle PRRs familiesare the Toll-like receptors (TLRs), the NOD-Like receptors (NLRs),and the RIG-I-Like receptors (RLRs) [6]. NLRs are intracellularsensors of pathogen-associated molecular patterns (PAMPS), andseveral NLRs are involved in the formation of inflammasomes [7].The formation of inflammasome-related molecules leads to theactivation of caspase-1, resulting in the cleavage of pro-IL-1b andpro-IL-18 into their mature forms. IL-1b and IL-18 are critical for theprogression of inflammation in the microenvironment. Several re-ports have examined the role of inflammasomes in auto-immunediseases. A recent study found that inflammasome activation wasinvolved in the pathogenesis of collagen-induced arthritis [8].NACHT, LRR and PYD domain-containing protein 3 (NALP3) plays acritical role in the development of systemic lupus erythematosus(SLE) [9]. Moreover, many reports indicate that inflammasome ac-tivity is related to auto-immune diseases in humans [10e12].However, it remains unclear whether the activity of inflamma-somes is involved in AIH, which is one of the immune-mediated* Corresponding author.
E-mail address: [email protected] (H. Ito).
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate/ybbrc
http://dx.doi.org/10.1016/j.bbrc.2017.06.0490006-291X/© 2017 Elsevier Inc. All rights reserved.
Biochemical and Biophysical Research Communications 490 (2017) 364e370
diseases.In this study, we used NALP3-KO mice to examined the role of
NALP3, in the progression of GalCer-induced liver injury usingNALP3-KO mice. We demonstrated that NALP3 deficiency reducedthe inflammatory response in the liver and thus liver injury, byincreasing CD11bþ/Gr-1intermediate cells.
2. Materials and methods
2.1. Mice
C57BL/6J (H-2b) wild-type (WT) mice (age, 6e10 weeks;weight; 20e30 g) were obtained from Japan SLC Inc. (Shizuoka,Japan). LRR and PYD domains-containing protein 3 (NALP3)knockout (KO) mice on a C57BL/6J background were obtained fromGenentech, Inc. (South San Francisco, CA). All procedures wereconducted in accordance with the guidelines of the National In-stitutes of Health Guide for the Care and Use of Laboratory Animals,and with the guidelines for the care and use of animals establishedby the Animal Care and Use Committee of Gifu University (Gifu,Japan).
2.2. Reagents
Alpha-galactosylceramide (GalCer) was obtained from Funa-koshi Co. Ltd. (Tokyo, Japan) and stored as a 200 mg/ml stock so-lution in dimethyl sulfoxide (DMSO). Isoliquiritigenin (ISL), a NALP3inhibitor, was purchased from Sigma-Aldrich (St Louis, MO).
2.3. Animal treatment
GalCer was diluted in pyrogen-free saline to obtain the indicateddose just before the intraperitoneal injection of a total volume of200 ml/mouse. Hepatocellular injury was monitored biochemicallythrough measurement serum alanine aminotransferase (ALT) ac-tivity. At appropriate time points, mice were killed by cervicaldislocation and necropsy was performed. Liver tissue samples werefixed in 10% neutral buffered formalin, embedded in paraffin, andsectioned; the sections were then stained with hematoxylin andeosin.
2.4. Measurement of serum transaminase
Hepatocyte damage was assessed at the indicated time pointsafter GalCer injection through measurement of serum ALT activityusing an automated clinical analyzer (BM2250; JEOL Ltd., Tokyo).
2.5. Real-time reverse transcription (RT)ePCR
Total RNA was extracted from mouse liver using ISOGEN II re-agent (Nippon Gene, Tokyo, Japan), and reverse transcribed intocDNA using the High Capacity cDNA Reverse Transcription Kit(Applied Biosystems, Foster City, CA). The resulting cDNA was usedas a template for real-time RT-PCR with primer-robe sets for IL-1b,TNF-a, CCL2, MIP1a, CXCL10, IFN-g, and 18S rRNA, the latter as aninternal control, according to the manufacturer's instructions(TaqMan® Gene Expression Assays and Universal PCR Master mix;Applied Biosystems). Real-time RT-PCR was carried out using theLight-Cycler® 480 system (Roche Diagnostic Systems, Basel,Switzerland).
2.6. Enzyme-linked immunosorbent assay (ELISA)
IFN-gconcentration in culture supernatant and serum fromGalCer treated mice were measured using ELISA. Ninety-six-well
Immulon 2HB plates (Thermo Lab Systems, OH, USA) were coatedwith purified IFN-g-specific antibody and then blocked withphosphate-buffered saline (PBS) containing 1% bovine serum al-bumin (BSA). Culture supernatants or mouse serum were thenadded to the wells of the ELISA plate, and bound cytokine wasdetected by the addition of biotin-labeled secondary IFN-g-specificantibody (all antibodies were obtained from eBioscience), followedby phosphatase-conjugated avidin and a phosphatase-specificcolorimetric substrate (Sigma-Aldrich). Standard curves weregenerated using recombinant cytokines purchased from R&DSystems.
2.7. Western blot analysis
Protein (20 mg) from lysed wound sites was subjected to sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) andtransferred to nitrocellulose membranes. After blocking non-specific reaction with 5% skim milk, the membrane was incubatedwith anti-pro-caspase-1, anti-cleaved caspase1(p20), precursor IL-
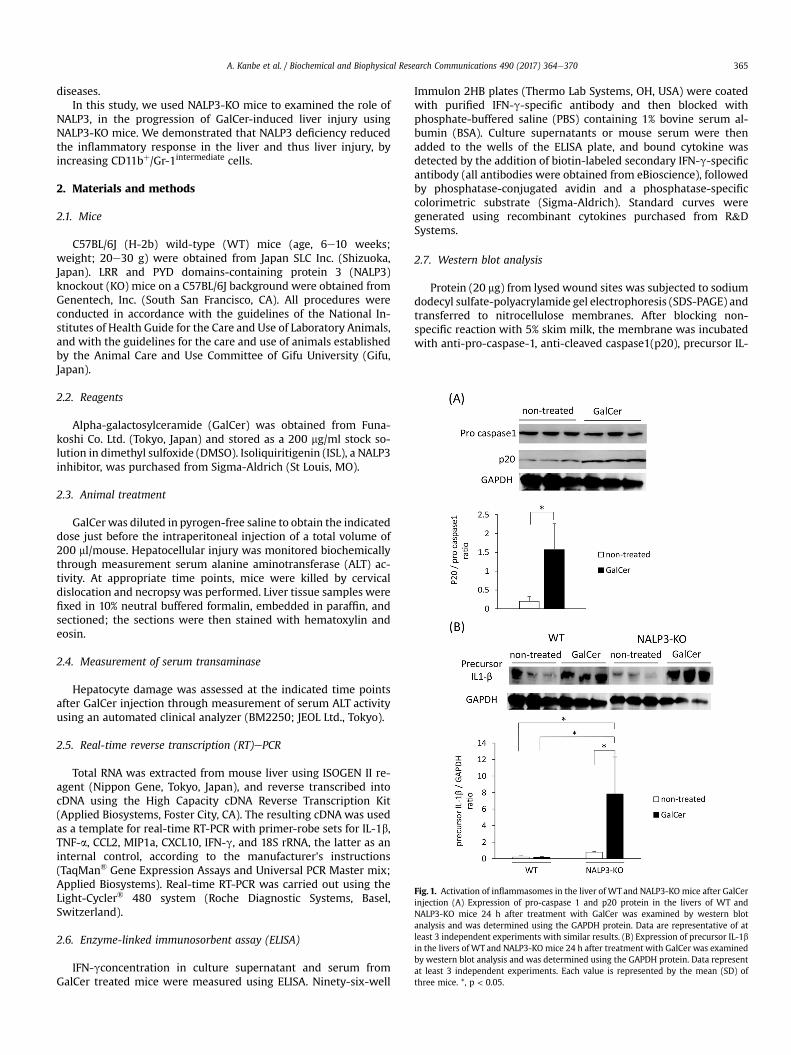
Fig. 1. Activation of inflammasomes in the liver of WT and NALP3-KO mice after GalCerinjection (A) Expression of pro-caspase 1 and p20 protein in the livers of WT andNALP3-KO mice 24 h after treatment with GalCer was examined by western blotanalysis and was determined using the GAPDH protein. Data are representative of atleast 3 independent experiments with similar results. (B) Expression of precursor IL-1bin the livers of WT and NALP3-KO mice 24 h after treatment with GalCer was examinedby western blot analysis and was determined using the GAPDH protein. Data representat least 3 independent experiments. Each value is represented by the mean (SD) ofthree mice. *, p < 0.05.
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370 365
1b. And anti-GAPDH antibodies for 60 min at room temperatureand subsequently incubated with peroxidase-labeled anti-mouseor -rabbit IgG antibody for 60 min at room temperature. Immu-noreactive protein bands were visualized with ECL plus (GEHealthcare UK Ltd., England).
2.8. Hepatic mononuclear cell preparation and flow cytometricanalysis
Hepatic mononuclear cells (MNCs) were isolated and purified aspreviously described [13]. Briefly, the excised liver was cut intosmall pieces with scissors, pressed through a 200-gauge stainlesssteel mesh, and suspended in PBS. Lymphocytes were separatedfrom parenchymal hepatocytes and hepatocyte nuclei by Ficoll-Conray (IBL, Gunma, Japan), and washed twice in ice-cold
medium. Cell viability and cell number were assessed by trypanblue exclusion. For flow cytometry, 2� 105 liver MNCs were stainedusing a standard protocol. The following antibodies were used:fluorescein isothiocyanate (FITC)-labeled rat anti-mouse Gr-1 mAb(clone RB6-8C5; Biosciences) and, phycoerythrin (PE)-labeled ratanti-mouse CD11b mAb (clone M1/70; Biosciences). Samples weremeasured on a FACStar flow cytometer and data analysis was per-formed using CellQuest software (BD Biosciences).
2.9. Statistical analysis
Values are expressed as the mean ± SD. Differences betweenexperimental and control groups were analyzed by the Kruskal-Wallis test followed by Scheffe's test. A value of p < 0.05 wasconsidered statistically significant.
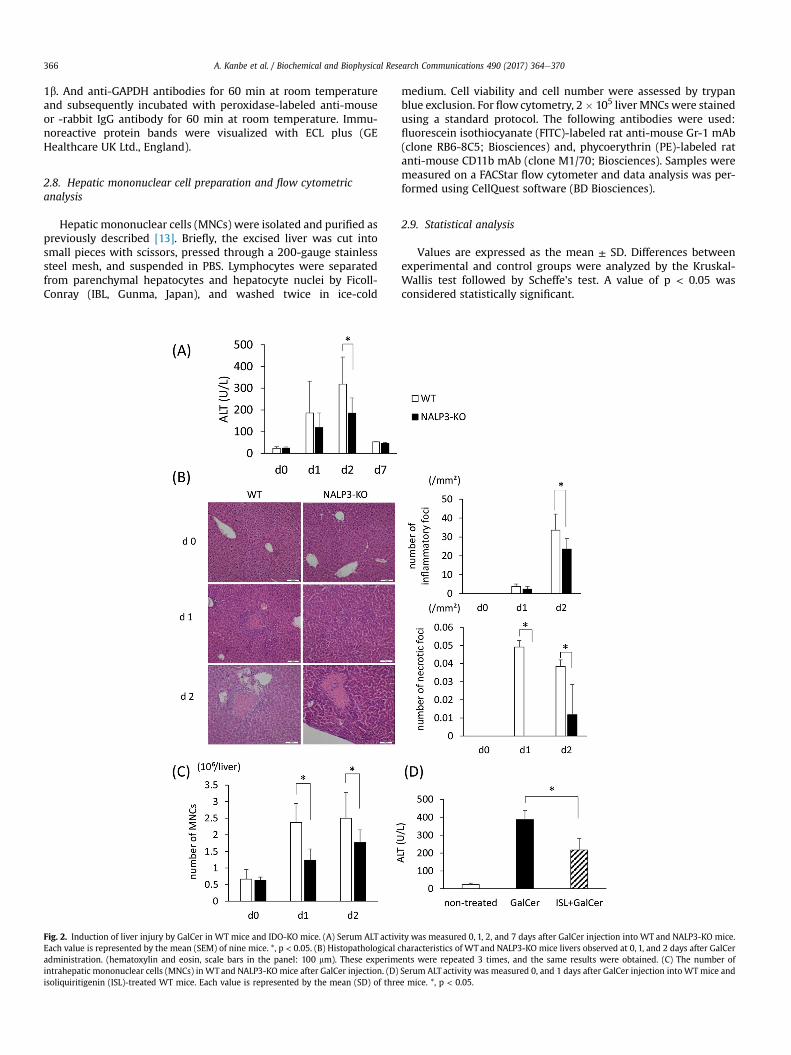
Fig. 2. Induction of liver injury by GalCer in WT mice and IDO-KO mice. (A) Serum ALT activity was measured 0, 1, 2, and 7 days after GalCer injection into WT and NALP3-KO mice.Each value is represented by the mean (SEM) of nine mice. *, p < 0.05. (B) Histopathological characteristics of WT and NALP3-KO mice livers observed at 0, 1, and 2 days after GalCeradministration. (hematoxylin and eosin, scale bars in the panel: 100 mm). These experiments were repeated 3 times, and the same results were obtained. (C) The number ofintrahepatic mononuclear cells (MNCs) in WT and NALP3-KO mice after GalCer injection. (D) Serum ALT activity was measured 0, and 1 days after GalCer injection into WT mice andisoliquiritigenin (ISL)-treated WT mice. Each value is represented by the mean (SD) of three mice. *, p < 0.05.
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370366
3. Results
3.1. Activation of the NALP3 inflammasome in the liver aftersystemic administration of GalCer
We first measured the expression of pro-caspase-1 and cleavedpro-caspase-1 (p20) by western blot to examine inflammasomeactivation in the liver after the administration of GalCer. As shownin Fig. 1A, the expression of p20 in WT mice significantly increasedafter GalCer injection. The activation of caspase-1 cleaves pro- IL-1binto its mature form.Which is then is rapidly secreted from the cells[14]. Thus, we examined the expression of pro-IL-1b significantlyincreased in GalCer treated NALP3-KO mice (Fig. 1B).
3.2. GalCer-induced liver injury in WT mice and NALP3-KO mice
To clarify the role of inflammasome activation in Galcer-inducedliver injury, serum ALT levels were measured after GalCer admin-istration to both WT and NALP3-KO mice. As shown in Fig. 2A anddays after GalCer injection, ALT levels significantly decreased inNALP3-KO mice. Consistent with this finding, histological exami-nation revealed that the number of necrotic foci in NALP3-KO micewere significantly reduced 1 day after GalCer injection (Fig. 2B). Thenumber of inflammatory foci after GalCer injection also decreasedin NALP3-KO mice. Necroinflammatory foci are defined as theinfiltration of inflammatory cells into a region in which more thanthree, hepatocytes are undergoing necrosis, and inflammatory cellfoci are defined as the existence of more than ten inflammatory cellin one paraffin sections of the liver. The data are represented as thenumerical value per area (1 mm2). Additionally, we counted thenumber of intrahepatic MNCs in WT and NALP3-KO mice afterisolating MNCs. The number of MNCs also decreased in NALP3-KOmice after GalCer injection (Fig. 2C). Next, we used ISL, which is apotent inhibitor of NALP3 signaling, to confirm data from the
examination of NALP3-KO mice. Pre-treatment using ISL markedlyreduced serum ALT levels in WT mice in response to GalCer in-jection (Fig. 2D).
3.3. Cytokine and chemokine expression in WT mice and NALP3-KOmice treated with GalCer
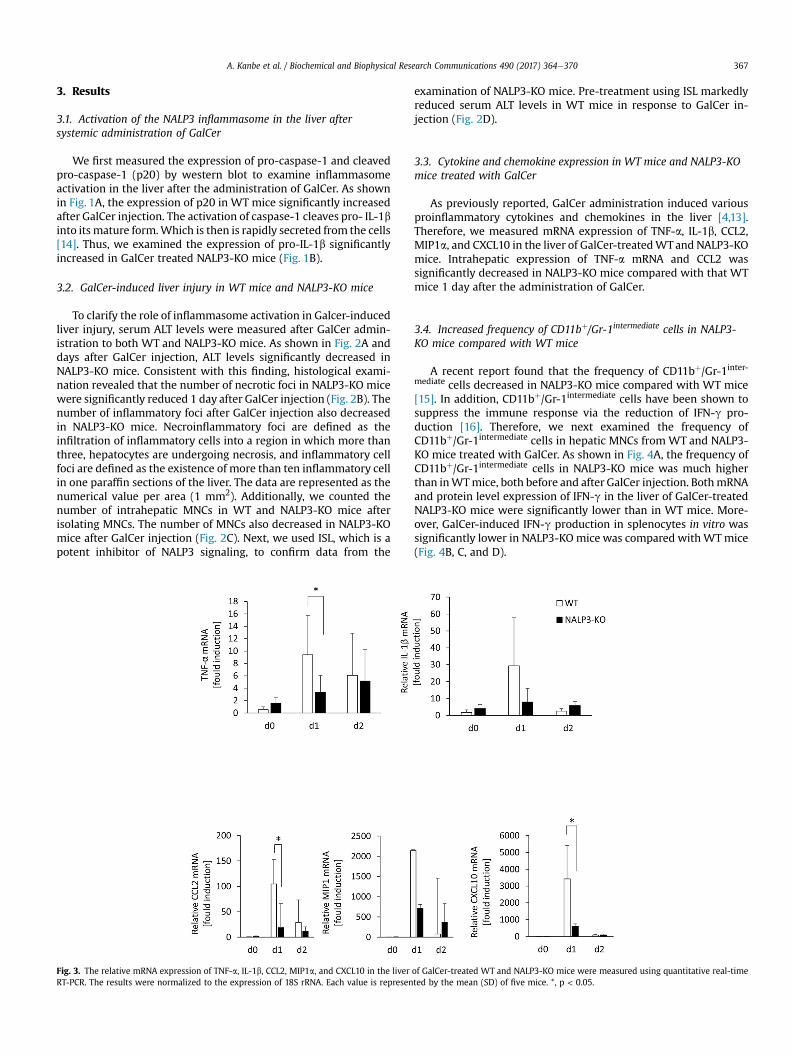
As previously reported, GalCer administration induced variousproinflammatory cytokines and chemokines in the liver [4,13].Therefore, we measured mRNA expression of TNF-a, IL-1b, CCL2,MIP1a, and CXCL10 in the liver of GalCer-treatedWTand NALP3-KOmice. Intrahepatic expression of TNF-a mRNA and CCL2 wassignificantly decreased in NALP3-KO mice compared with that WTmice 1 day after the administration of GalCer.
3.4. Increased frequency of CD11bþ/Gr-1intermediate cells in NALP3-KO mice compared with WT mice
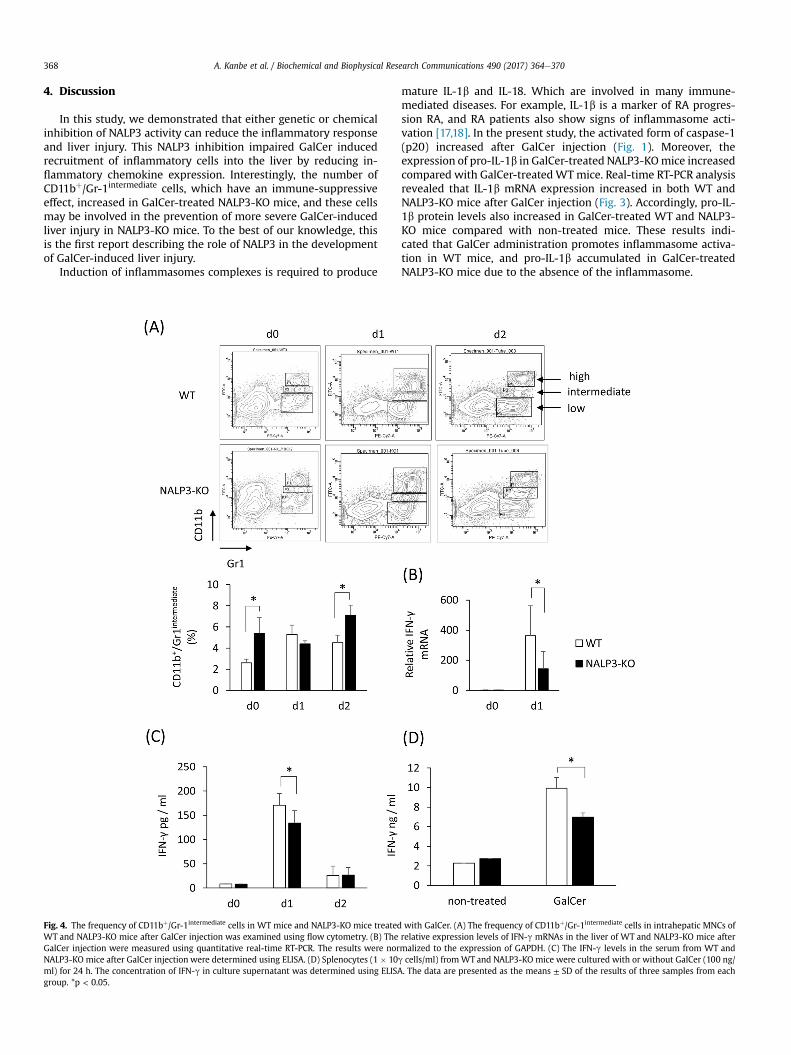
A recent report found that the frequency of CD11bþ/Gr-1inter-mediate cells decreased in NALP3-KO mice compared with WT mice[15]. In addition, CD11bþ/Gr-1intermediate cells have been shown tosuppress the immune response via the reduction of IFN-g pro-duction [16]. Therefore, we next examined the frequency ofCD11bþ/Gr-1intermediate cells in hepatic MNCs from WT and NALP3-KO mice treated with GalCer. As shown in Fig. 4A, the frequency ofCD11bþ/Gr-1intermediate cells in NALP3-KO mice was much higherthan inWTmice, both before and after GalCer injection. BothmRNAand protein level expression of IFN-g in the liver of GalCer-treatedNALP3-KO mice were significantly lower than in WT mice. More-over, GalCer-induced IFN-g production in splenocytes in vitro wassignificantly lower in NALP3-KO mice was compared withWTmice(Fig. 4B, C, and D).
Fig. 3. The relative mRNA expression of TNF-a, IL-1b, CCL2, MIP1a, and CXCL10 in the liver of GalCer-treated WT and NALP3-KO mice were measured using quantitative real-timeRT-PCR. The results were normalized to the expression of 18S rRNA. Each value is represented by the mean (SD) of five mice. *, p < 0.05.
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370 367
4. Discussion
In this study, we demonstrated that either genetic or chemicalinhibition of NALP3 activity can reduce the inflammatory responseand liver injury. This NALP3 inhibition impaired GalCer inducedrecruitment of inflammatory cells into the liver by reducing in-flammatory chemokine expression. Interestingly, the number ofCD11bþ/Gr-1intermediate cells, which have an immune-suppressiveeffect, increased in GalCer-treated NALP3-KO mice, and these cellsmay be involved in the prevention of more severe GalCer-inducedliver injury in NALP3-KO mice. To the best of our knowledge, thisis the first report describing the role of NALP3 in the developmentof GalCer-induced liver injury.
Induction of inflammasomes complexes is required to produce
mature IL-1b and IL-18. Which are involved in many immune-mediated diseases. For example, IL-1b is a marker of RA progres-sion RA, and RA patients also show signs of inflammasome acti-vation [17,18]. In the present study, the activated form of caspase-1(p20) increased after GalCer injection (Fig. 1). Moreover, theexpression of pro-IL-1b in GalCer-treated NALP3-KOmice increasedcompared with GalCer-treatedWTmice. Real-time RT-PCR analysisrevealed that IL-1b mRNA expression increased in both WT andNALP3-KO mice after GalCer injection (Fig. 3). Accordingly, pro-IL-1b protein levels also increased in GalCer-treated WT and NALP3-KO mice compared with non-treated mice. These results indi-cated that GalCer administration promotes inflammasome activa-tion in WT mice, and pro-IL-1b accumulated in GalCer-treatedNALP3-KO mice due to the absence of the inflammasome.
Fig. 4. The frequency of CD11bþ/Gr-1intermediate cells in WT mice and NALP3-KO mice treated with GalCer. (A) The frequency of CD11bþ/Gr-1intermediate cells in intrahepatic MNCs ofWT and NALP3-KO mice after GalCer injection was examined using flow cytometry. (B) The relative expression levels of IFN-g mRNAs in the liver of WT and NALP3-KO mice afterGalCer injection were measured using quantitative real-time RT-PCR. The results were normalized to the expression of GAPDH. (C) The IFN-g levels in the serum from WT andNALP3-KO mice after GalCer injection were determined using ELISA. (D) Splenocytes (1 � 10g cells/ml) from WT and NALP3-KO mice were cultured with or without GalCer (100 ng/ml) for 24 h. The concentration of IFN-g in culture supernatant was determined using ELISA. The data are presented as the means ± SD of the results of three samples from eachgroup. *p < 0.05.
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370368

Inflammasome activation is involved in the production of IL-1band IL-18. Previous studies have demonstrated that expression ofIL-1b expression is increased in the liver and the serum of patientswith hepatitis [19,20], and an IL-18 neutralizing antibody attenu-ated liver injury in a Con A-induced hepatitis model [21]. IL-1b andIL-18 are also involved in the progression of other immune-mediated diseases. An IL-1 receptor antagonist has been shownto be effective in patients with RA [22,23]. Thus, IL-1b and IL-18 playa critical role in immune-mediated diseases, and inflammasomesthat regulate the production and maturation of these cytokines arealso involved in the progression of such diseases. In the presentstudy, GalCer-induced liver injury was significantly attenuated inNALP3-KO mice compared with WT mice (Fig. 2). Histological ex-amination and real-time RT-PCR analysis indicated that NALP3deficiency reduced liver inflammation after GalCer injection.Therefore, the reduction of inflammation may lead to the attenu-ation of GalCer-induced liver injury.
Histological examination, found fewer immune cells infiltratingthe liver in GalCer-treated NALP3-KO mice compared with GalCer-treated WT mice. This reduction was partially related to thedecrease in chemokine expression in NALP3-KO mice injected withGalCer (Fig. 3). The absolute number of intrahepatic MNCs was alsoreduced in NALP3-KO mice compared with WT mice. Interestingly,the frequency of CD11bþ/Gr-1intermediate cells after GalCer treatmentincreased in NALP3-KO mice compared with WT mice. Consistentwith these results, Chow MT et al. [24] demonstrated that the fre-quency of CD11bþ/Gr-1intermediate cells increased in tumor-bearingNLRP3-KO mice compared with WT mice. Myeloid-derived sup-pressor cells (MDSCs) are immunosuppressive and have beenshown to express CD11b and Gr-1 on their cell surfaces. CD11bþ/Gr-1þ cells are divided into two populations: CD11bþ/Gr-1high cells andCD11bþ/Gr-1 intermediate cells. A recent report demonstrated that thenumber of CD11bþ/Gr-1 intermediate cells increased in lethal tuber-culosis infection in mice [16]. In mice infected with Mycobacteriumtuberculosis these CD11bþ/Gr-1 intermediate cells suppressed IFN-gproduction from bone marrow cells and the subsequent activationof immune cells. Thus, the accumulation of CD11bþ/Gr-1 intermediate
cells in the liver may suppress the inflammatory response byreducing IFN-g. Therefore, the accumulation of CD11bþ/Gr-1 inter-
mediate cells in NALP3-KO mice is not a little involved in the atten-uation of GalCer-induced liver injury in NALP3-KO mice. Bycontrast, another study demonstrated that the activation ofinflammasomes impaired immune suppression in MDSCs in GVHDmouse model [25]. Thus, it remains unclear how the inhibition ofNALP3 signaling affects MDSC development, and further studies areneeded.
Although there is no essential therapy for AIH, corticosteroidtreatment is often used and effective for the control of AIH [26].However, corticosteroid treatment has many side effects, andseveral support therapies are required for these side effects. Thus,new treatment strategies for AIH must be developed, and this re-quires identifying and inhibiting the molecules involved in AIHprogression.
In the present study, we revealed the role of NALP3 in GalCer-induced liver injury. The inhibitor of NALP3 significantly reducedliver injury after the administration of GalCer (Fig. 2D). Anddecreased the expression of pro-inflammatory cytokines and che-mokines, leading to the recruitment suppression of immune cellsinto the liver. Moreover, the frequency of CD11bþ/Gr-1 intermediate
MDSCs increased in GalCer-treated NALP3-KOmice. The increase ofMDSCs may be involved in the attenuation of GalCer-induced liverinjury in the absence of NALP3 activation. These observationsindicate that inhibition of NALP3 signaling could be a therapeutictarget in AIH.
Conflict of interest
All the authors declares no conflict of interest.
Acknowledgements
We thank Genentech, Inc. (South San Francisco, CA) forproviding the NALP3-KO mice.
Transparency document
Transparency document related to this article can be foundonline at http://dx.doi.org/10.1016/j.bbrc.2017.06.049.
References
[1] A.J. Czaja, Difficult treatment decisions in autoimmune hepatitis, World J.Gastroenterol. 16 (2010) 934e947.
[2] G. Xia, S. Wu, Y. Zhang, Anti-4-1BB monoclonal antibodies attenuate conca-navalin A-induced immune-mediated liver injury in mice, Exp. Ther. Med. 12(2016) 1263e1268.
[3] Y. Kaneko, M. Harada, T. Kawano, M. Yamashita, Y. Shibata, F. Gejyo, et al.,Augmentation of Valpha14 NKT cell-mediated cytotoxicity by interleukin 4 inan autocrine mechanism resulting in the development of concanavalin A-induced hepatitis, J. Exp. Med. 191 (2000) 105e114.
[4] H. Ito, M. Hoshi, H. Ohtaki, A. Taguchi, K. Ando, T. Ishikawa, et al., Ability of IDOto attenuate liver injury in alpha-galactosylceramide-induced hepatitis model,J. Immunol. 185 (2010) 4554e4560.
[5] H. Ito, N. Koide, F. Hassan, S. Islam, G. Tumurkhuu, I. Mori, et al., Lethalendotoxic shock using alpha-galactosylceramide sensitization as a newexperimental model of septic shock, Lab. Invest. 86 (2006) 254e261.
[6] T. Kawai, S. Akira, Toll-like receptors and their crosstalk with other innatereceptors in infection and immunity, Immunity 34 (2011) 637e650.
[7] H. Guo, J.B. Callaway, J.P. Ting, Inflammasomes: mechanism of action, role indisease, and therapeutics, Nat. Med. 21 (2015) 677e687.
[8] Y. Zhang, Y. Zheng, H. Li, NLRP3 inflammasome plays an important role in thepathogenesis of collagen-induced arthritis, Mediat. Inflamm. 2016 (2016),9656270.
[9] A. Lu, H. Li, J. Niu, S. Wu, G. Xue, X. Yao, et al., Hyperactivation of the NLRP3inflammasome in myeloid cells leads to severe organ damage in experimentallupus, J. Immunol. 198 (2017) 1119e1129.
[10] A. Kastbom, D. Verma, P. Eriksson, T. Skogh, G. Wingren, P. Soderkvist, Geneticvariation in proteins of the cryopyrin inflammasome influences susceptibilityand severity of rheumatoid arthritis (the Swedish TIRA project), Rheumatol.Oxf. 47 (2008) 415e417.
[11] C.A. Yang, S.T. Huang, B.L. Chiang, Sex-dependent differential activation ofNLRP3 and AIM2 inflammasomes in SLE macrophages, Rheumatol. Oxf. 54(2015) 324e331.
[12] R.J. Mathews, J.I. Robinson, M. Battellino, C. Wong, J.C. Taylor, S. Eyre, et al.,Evidence of NLRP3-inflammasome activation in rheumatoid arthritis (RA);genetic variants within the NLRP3-inflammasome complex in relation tosusceptibility to RA and response to anti-TNF treatment, Ann. Rheum. Dis. 73(2014) 1202e1210.
[13] H. Ito, K. Ando, T. Nakayama, M. Taniguchi, T. Ezaki, K. Saito, et al., Role ofValpha 14 NKT cells in the development of impaired liver regenerationin vivo, Hepatology 38 (2003) 1116e1124.
[14] D. Brough, N.J. Rothwell, Caspase-1-dependent processing of pro-interleukin-1beta is cytosolic and precedes cell death, J. Cell Sci. 120 (2007) 772e781.
[15] M.T. Chow, J. Sceneay, C. Paget, C.S. Wong, H. Duret, J. Tschopp, et al., NLRP3suppresses NK cell-mediated responses to carcinogen-induced tumors andmetastases, Cancer Res. 72 (2012) 5721e5732.
[16] E.N. Tsiganov, E.M. Verbina, T.V. Radaeva, V.V. Sosunov, G.A. Kosmiadi,I.Y. Nikitina, et al., Gr-1dimCD11bþ immature myeloid-derived suppressorcells but not neutrophils are markers of lethal tuberculosis infection in mice,J. Immunol. 192 (2014) 4718e4727.
[17] J.F. Camargo, P.A. Correa, J. Castiblanco, J.M. Anaya, Interleukin-1beta poly-morphisms in Colombian patients with autoimmune rheumatic diseases,Genes Immun. 5 (2004) 609e614.
[18] C.A. Yang, B.L. Chiang, Inflammasomes and human autoimmunity: acomprehensive review, J. Autoimmun. 61 (2015) 1e8.
[19] M. Shindo, G.E. Mullin, L. Braun-Elwert, N.V. Bergasa, E.A. Jones, S.P. James,Cytokine mRNA expression in the liver of patients with primary biliarycirrhosis (PBC) and chronic hepatitis B (CHB), Clin. Exp. Immunol. 105 (1996)254e259.
[20] S. Satoh, A.K. Nussler, Z.Z. Liu, A.W. Thomson, Proinflammatory cytokines andendotoxin stimulate ICAM-1 gene expression and secretion by normal humanhepatocytes, Immunology 82 (1994) 571e576.
[21] A. Ikeda, N. Aoki, M. Kido, S. Iwamoto, H. Nishiura, R. Maruoka, et al., Pro-gression of autoimmune hepatitis is mediated by IL-18-producing dendritic
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370 369

cells and hepatic CXCL9 expression in mice, Hepatology 60 (2014) 224e236.[22] S. Cohen, E. Hurd, J. Cush, M. Schiff, M.E. Weinblatt, L.W. Moreland, et al.,
Treatment of rheumatoid arthritis with anakinra, a recombinant humaninterleukin-1 receptor antagonist, in combination with methotrexate: resultsof a twenty-four-week, multicenter, randomized, double-blind, placebo-controlled trial, Arthritis Rheum. 46 (2002) 614e624.
[23] X. Le Loet, D. Nordstrom, M. Rodriguez, A. Rubbert, P. Sarzi-Puttini,J.M. Wouters, et al., Effect of anakinra on functional status in patients withactive rheumatoid arthritis receiving concomitant therapy with traditionaldisease modifying antirheumatic drugs: evidence from the OMEGA trial,
J. Rheumatol. 35 (2008) 1538e1544.[24] M.T. Chow, J. Tschopp, A. Moller, M.J. Smyth, NLRP3 promotes inflammation-
induced skin cancer but is dispensable for asbestos-induced mesothelioma,Immunol. Cell Biol. 90 (2012) 983e986.
[25] B.H. Koehn, P. Apostolova, J.M. Haverkamp, J.S. Miller, V. McCullar, J. Tolar, etal., GVHD-associated, inflammasome-mediated loss of function in adoptivelytransferred myeloid-derived suppressor cells, Blood 126 (2015) 1621e1628.
[26] A.J. Czaja, Diagnosis and management of autoimmune hepatitis: current statusand future directions, Gut Liver 10 (2016) 177e203.
A. Kanbe et al. / Biochemical and Biophysical Research Communications 490 (2017) 364e370370


















