
SUPPLEMENTARY INFORMATION with E34 and G35 of ubiquitin in the structure of the RNF4...
21
WWW.NATURE.COM/NATURE | 1 SUPPLEMENTARY INFORMATION doi:10.1038/nature11376 Supplementary Table 1 Data collection and refinement statistics Data collection Space group P2 1 2 1 2 1 Cell dimensions a, b, c (Å) 50.5, 65.1, 189.7 α, β, γ (°) 90, 90, 90 Resolution (Å) 99 – 2.21 (2.26 – 2.21)* R merge 0.033 (0.172) I/σI 44 (7) Completeness (%) 99 (92) Redundancy 10.6 (3.5) Refinement Resolution (Å) 95 – 2.21 (2.26 – 2.21) No. reflections 30,468 R work / R free 20.8 (26.2) / 24.2 (31.0) No. atoms Protein 4607 Zn 2+ 4 Water 137 B-factors Protein 49 Zn 2+ 33 Water 41 R.m.s deviations Bond lengths (Å) 0.008 Bond angles (º) 1.2 *Highest resolution shell is shown in parenthesis. One crystal was used to collect the data set.
Transcript of SUPPLEMENTARY INFORMATION with E34 and G35 of ubiquitin in the structure of the RNF4...

W W W. N A T U R E . C O M / N A T U R E | 1
SUPPLEMENTARY INFORMATIONdoi:10.1038/nature11376SUPPLEMENTARY TABLE
Supplementary Table 1 Data collection and refinement statistics Data collection Space group P212121 Cell dimensions a, b, c (Å) 50.5, 65.1, 189.7 α, β, γ (°) 90, 90, 90 Resolution (Å) 99 – 2.21 (2.26 – 2.21)* Rmerge 0.033 (0.172) I/σI 44 (7) Completeness (%) 99 (92) Redundancy 10.6 (3.5) Refinement Resolution (Å) 95 – 2.21 (2.26 – 2.21) No. reflections 30,468 Rwork / Rfree 20.8 (26.2) / 24.2 (31.0) No. atoms Protein 4607 Zn2+ 4 Water 137 B-factors Protein 49 Zn2+ 33 Water 41 R.m.s deviations Bond lengths (Å) 0.008 Bond angles (º) 1.2
*Highest resolution shell is shown in parenthesis. One crystal was used to collect the data set.
SUPPLEMENTARY INFORMATION
2 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
350
0
50
100
150
200
250
300
5040 60 70 80 90 100 110
Abs
orba
nce
at 2
80 n
m (m
AU
)
Elution volume (ml)
Ubiquitin
UbcH5A(S22R/C85K)–Ub126 -
63 -
35 -25 -17 -12 -
7.5 -
kDa
UbcH5A(S22R/C85K)–Ub
Ubiquitin
Load
Fractions
a
b c
26
126 -63 -
35 -25 -
17 -12 -
7.5 -
kDa
Time (h):
Conjugationreaction
2280 Time (h): 0
Cleavagewith TEV
18
UbcH5A–HisUbUbcH5A–Ub
UbiquitinHisUbiquitin
Ni-NTAchromatography
Load
FTLoad
UbcH5A–HisUbUbcH5A(S22R/C85K)
HisUbiquitin
Ni-NTAchromatography
FT Elution
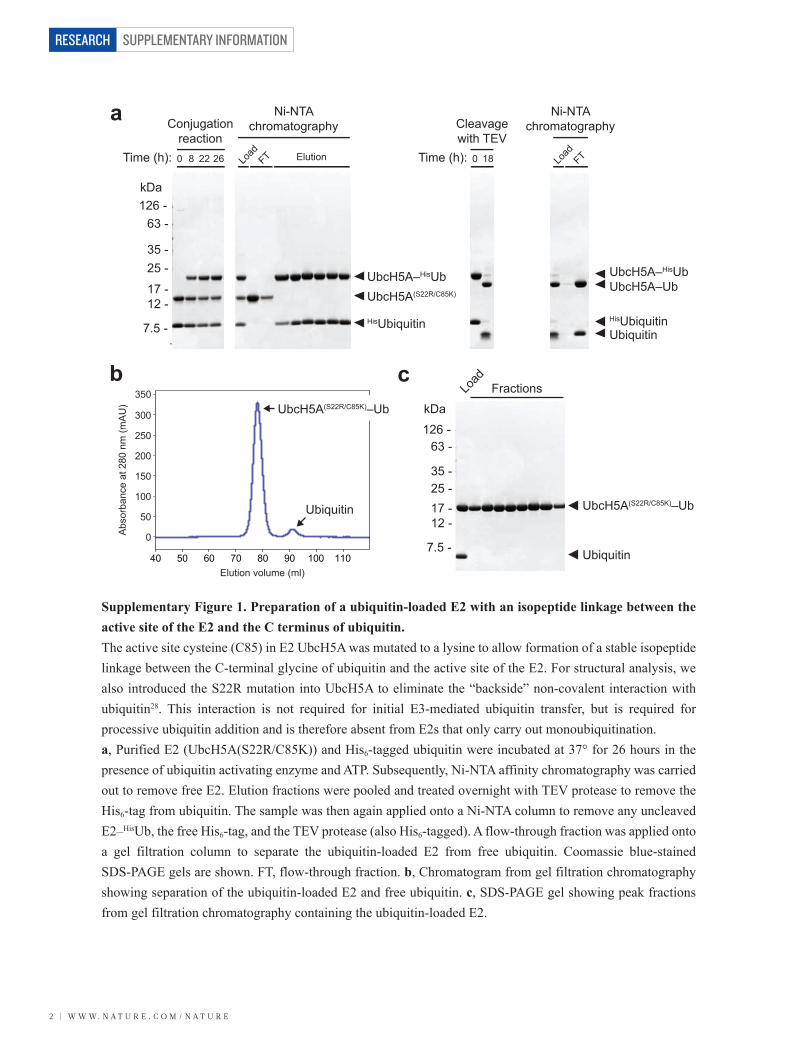
Supplementary Figure 1. Preparation of a ubiquitin-loaded E2 with an isopeptide linkage between the active site of the E2 and the C terminus of ubiquitin. The active site cysteine (C85) in E2 UbcH5A was mutated to a lysine to allow formation of a stable isopeptide linkage between the C-terminal glycine of ubiquitin and the active site of the E2. For structural analysis, we also introduced the S22R mutation into UbcH5A to eliminate the “backside” non-covalent interaction with ubiquitin28. This interaction is not required for initial E3-mediated ubiquitin transfer, but is required for processive ubiquitin addition and is therefore absent from E2s that only carry out monoubiquitination. a, Purified E2 (UbcH5A(S22R/C85K)) and His6-tagged ubiquitin were incubated at 37° for 26 hours in the presence of ubiquitin activating enzyme and ATP. Subsequently, Ni-NTA affinity chromatography was carried out to remove free E2. Elution fractions were pooled and treated overnight with TEV protease to remove the His6-tag from ubiquitin. The sample was then again applied onto a Ni-NTA column to remove any uncleaved E2–HisUb, the free His6-tag, and the TEV protease (also His6-tagged). A flow-through fraction was applied onto a gel filtration column to separate the ubiquitin-loaded E2 from free ubiquitin. Coomassie blue-stained SDS-PAGE gels are shown. FT, flow-through fraction. b, Chromatogram from gel filtration chromatography showing separation of the ubiquitin-loaded E2 and free ubiquitin. c, SDS-PAGE gel showing peak fractions from gel filtration chromatography containing the ubiquitin-loaded E2.
SUPPLEMENTARY FIGURES
W W W. N A T U R E . C O M / N A T U R E | 3
SUPPLEMENTARY INFORMATION RESEARCH
_
y₁₇
I
y₁₆
a₂
Y
y₁₅
b₃
H
y₁₄
b₄
P
y₁₃
b₅
N
y₁₂
b₆
I
y₁₁
b₇
N
y₁₀
b₈
S
y₉
b₉
N
y₈
b₁₀
G
y₇
b₁₁
S
y₆
b₁₂
I
y₅
b₁₃
gl
K
y₄
b₁₄
I
y₃
b₁₅
D
y₂
b₁₆
I
y₁
b₁₇
I R _
y₁175.119
a₂249.1598
y₂-NH₃271.1765
y₂288.203
y₃-NH₃384.2605
y₃401.2871
b₃414.2136
y₄-NH₃499.2875
b₄511.2663
y₄516.314
b₅625.3093
y₅629.3981 b₆-NH₃
721.3668
b₆738.3933
b₁₃²⁺776.897
y₁₃²⁺778.9414
b₇852.4363
y₆-NH₃854.5094
y₆871.536
b₈-H₂O921.4577
b₈939.4683
y₇984.62
b₁₇²⁺1004.037
b₉1053.511
y₈1071.652
b₁₀1110.533
y₉1128.674
b₁₁-H₂O1179.554
b₁₁1197.565
y₁₀-NH₃1225.69
y₁₀1242.716
b₁₂-H₂O1292.638
b₁₂1310.649
y₁₁-NH₃1312.722
y₁₁1329.748
y₁₂-NH₃1426.765
y₁₂1443.791
y₁₃-NH₃1539.849
b₁₃1552.787
y₁₃1556.875
y₁₄-NH₃1653.892
b₁₄1665.871
y₁₄1670.918
y₁₅-NH₃1750.945
b₁₅-NH₃1763.871
y₁₅1767.971
b₁₅1780.898
b₁₆1893.982
y₁₆1905.03
b₁₇2007.066
y₁₇-NH₃2051.067
y₁₇2068.093
020
4060
8010
012
0
Rel
ativ
e Ab
unda
nce
01
23
4
Intensity [10e7]
200 400 600 800 1000 1200 1400 1600 1800 2000 2200
m/z [amu]
UbcH5A(S22R/C85K)
UbcH5A(S22R/C85K)–Ub
12.5-
17.0-
25.0-kDa
-Ub +Ub
UbcH5A(S22R/C85K)
0
5E+09
1E+10
1.5E+10
2E+10
GG-K85 LRGG-K85
Pep
tide
inte
nsity
-Ub+Ub
a
c
d
b
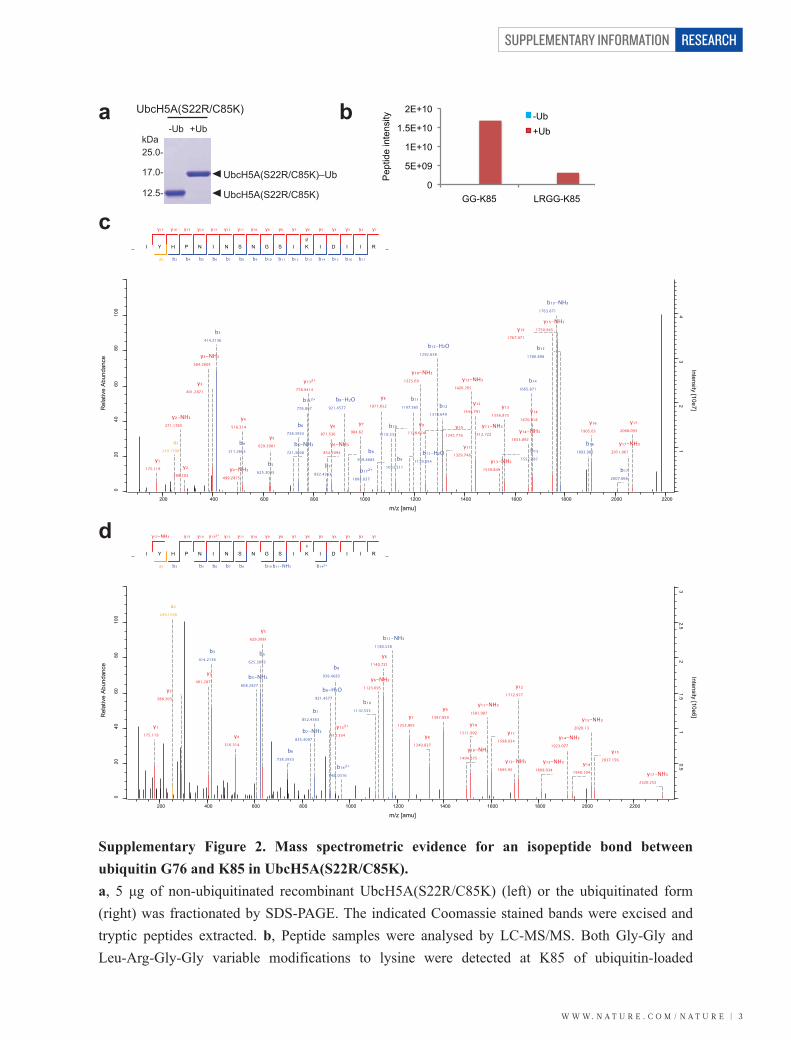
Supplementary Figure 2. Mass spectrometric evidence for an isopeptide bond between ubiquitin G76 and K85 in UbcH5A(S22R/C85K). a, 5 μg of non-ubiquitinated recombinant UbcH5A(S22R/C85K) (left) or the ubiquitinated form (right) was fractionated by SDS-PAGE. The indicated Coomassie stained bands were excised and tryptic peptides extracted. b, Peptide samples were analysed by LC-MS/MS. Both Gly-Gly and Leu-Arg-Gly-Gly variable modifications to lysine were detected at K85 of ubiquitin-loaded
_
y₁₇-NH₃
I
a₂
Y
y₁₅
b₃
H
y₁₄
P
y₁₃²⁺
b₅
N
y₁₂
b₆
I
y₁₁
b₇
N
y₁₀
b₈
S
y₉
N
y₈
b₁₀
G
y₇
b₁₁-NH₃
S
y₆
I
y₅
lr
K
y₄
b₁₄²⁺
I
y₃
D
y₂
I
y₁
I R _
y₁175.119
a₂249.1598
y₂288.203
y₃401.2871
b₃414.2136
y₄516.314
b₅-NH₃608.2827
b₅625.3093
y₅629.3981
b₆738.3933
b₇-NH₃835.4097
b₇852.4363
y₁₃²⁺913.534
b₈-H₂O921.4577
b₈939.4683
b₁₄²⁺968.0316
b₁₀1110.533
y₆-NH₃1123.695
y₆1140.721
b₁₁-NH₃1180.538
y₇1253.805
y₈1340.837
y₉1397.859
y₁₀-NH₃1494.875
y₁₀1511.902
y₁₁-NH₃1581.907
y₁₁1598.934
y₁₂-NH₃1695.95
y₁₂1712.977
y₁₃-NH₃1809.034
y₁₄-NH₃1923.077
y₁₄1940.104
y₁₅-NH₃2020.13
y₁₅2037.156
y₁₇-NH₃2320.252
020
4060
8010
012
0
Rel
ativ
e Ab
unda
nce
00.5
11.5
22.5
3
Intensity [10e6]
200 400 600 800 1000 1200 1400 1600 1800 2000 2200
m/z [amu]
SUPPLEMENTARY INFORMATION
4 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
WB: α-UbcH5A
Inpu
t (4%
)M
BP
RN
F4
RN
F4(M
140A
/R18
1A)
C85K
Inpu
t (4%
)M
BP
RN
F4
RN
F4( M
140A
/R18
1A)
C85SUbcH5A:
17 -12 -
kDa
63 -
45 -
kDa
MBP
MBP–RNF4
Coomassie blue-stainedSDS-PAGE gel
UbcH5A–Ub
WB: α-UbcH5A
C85K S22R/C85K
Inpu
t (15
%)
MB
P
C85SUbcH5A:
E2 siteY193A
RING1–RING2linear fusion
E2 siteY193AIn
put (
15%
)M
BP
RING1–RING2linear fusion
–– – ––– – –
––– –
–
UbcH5A–Ub
MBP–RING1–RING2MBP
17 -12 -
45 -63 -
–– – ––– – –
––– –
–
Inpu
t (15
%)
MB
P
RING1–RING2linear fusion
–– – ––– – –
––– –
–
Mutations in:
RIN
G1–R
ING
2
Coomassie blue-stained SDS-PAGE gels
kDa
17 -
25 -
45 -
128 -
8 -
kDa
UbcH5A(C85K)–Ub
RNF4
RNF4:Time (h):
– +0 5 24 48 72 10
015
00 5 24 48 72 10
015
0
a
cd
b
UbcH5A(S22R/C85K). c, MS/MS spectrum of the peptide 73-IYHPNINSNGSIKLDILR-90 containing K85 showing the Gly-Gly adduct characteristic of ubiquitination. The peptide was confidently identified with a high score (326.53) and low mass error (-0.33965). d, MS/MS spectrum of the peptide 73-IYHPNINSNGSIKLDILR-90 containing K85 with the Leu-Arg-Gly-Gly adduct characteristic of ubiquitination with a single missed tryptic cleavage at the C terminus. Again high score (176.23) and low mass error (0.3127 ppm) are evident. Together, these identify an isopeptide linkage between G76 of ubiquitin and K85 of the UbcH5A mutant.
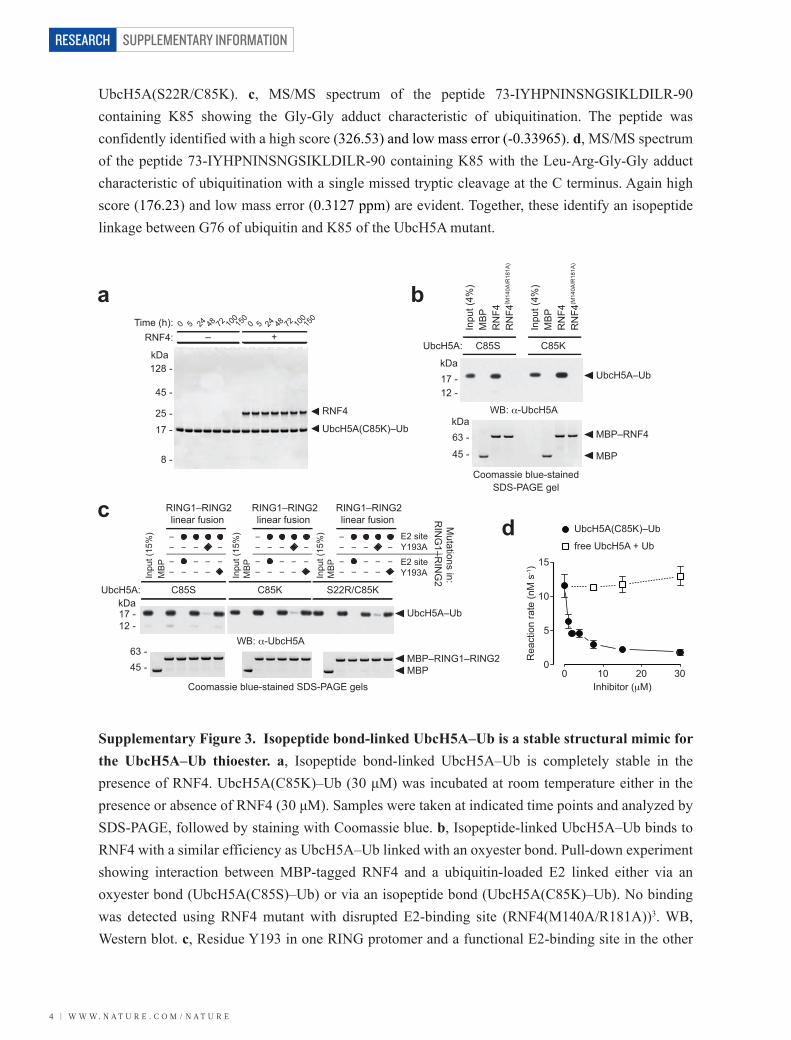
Supplementary Figure 3. Isopeptide bond-linked UbcH5A–Ub is a stable structural mimic for the UbcH5A–Ub thioester. a, Isopeptide bond-linked UbcH5A–Ub is completely stable in the presence of RNF4. UbcH5A(C85K)–Ub (30 μM) was incubated at room temperature either in the presence or absence of RNF4 (30 μM). Samples were taken at indicated time points and analyzed by SDS-PAGE, followed by staining with Coomassie blue. b, Isopeptide-linked UbcH5A–Ub binds to RNF4 with a similar efficiency as UbcH5A–Ub linked with an oxyester bond. Pull-down experiment showing interaction between MBP-tagged RNF4 and a ubiquitin-loaded E2 linked either via an oxyester bond (UbcH5A(C85S)–Ub) or via an isopeptide bond (UbcH5A(C85K)–Ub). No binding was detected using RNF4 mutant with disrupted E2-binding site (RNF4(M140A/R181A))3. WB, Western blot. c, Residue Y193 in one RING protomer and a functional E2-binding site in the other
0 10 20 30
UbcH5A(C85K)–Ub
free UbcH5A + Ub
Inhibitor (μM)
Rea
ctio
n ra
te (n
M s
-1)
0
5
10
15
W W W. N A T U R E . C O M / N A T U R E | 5
SUPPLEMENTARY INFORMATION RESEARCH
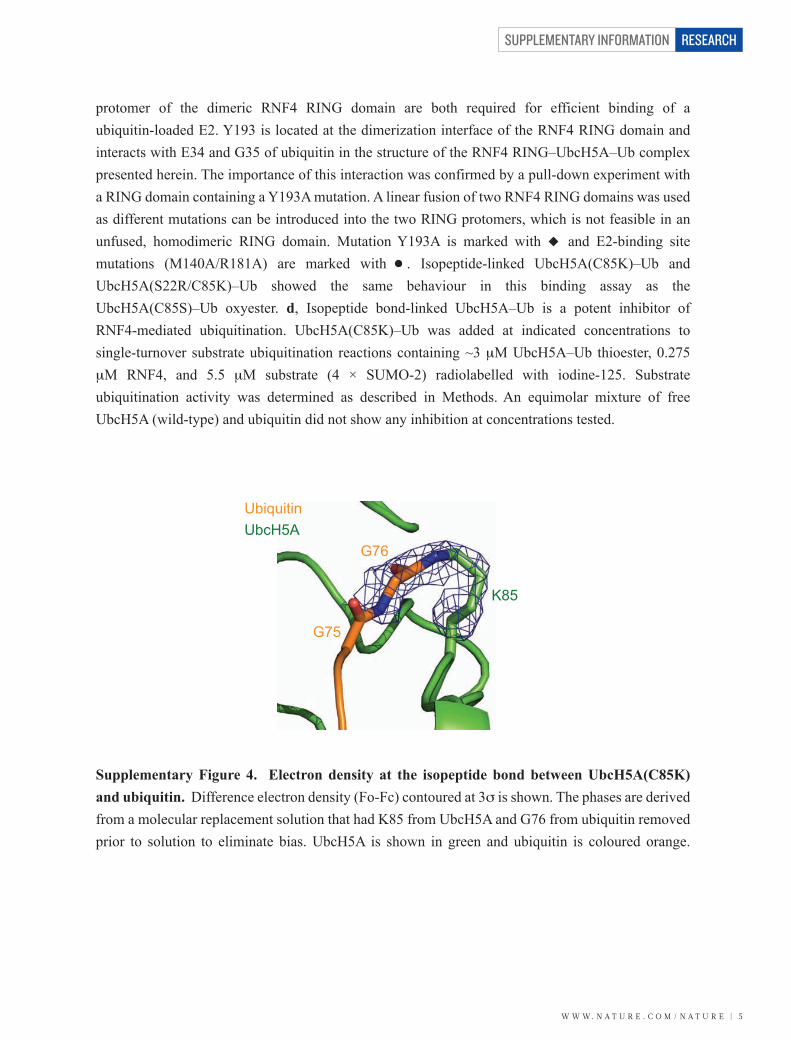
Supplementary Figure 4. Electron density at the isopeptide bond between UbcH5A(C85K) and ubiquitin. Difference electron density (Fo-Fc) contoured at 3σ is shown. The phases are derived from a molecular replacement solution that had K85 from UbcH5A and G76 from ubiquitin removed prior to solution to eliminate bias. UbcH5A is shown in green and ubiquitin is coloured orange.
G75
G76
UbiquitinUbcH5A
K85
protomer of the dimeric RNF4 RING domain are both required for efficient binding of a ubiquitin-loaded E2. Y193 is located at the dimerization interface of the RNF4 RING domain and interacts with E34 and G35 of ubiquitin in the structure of the RNF4 RING–UbcH5A–Ub complex presented herein. The importance of this interaction was confirmed by a pull-down experiment with a RING domain containing a Y193A mutation. A linear fusion of two RNF4 RING domains was used as different mutations can be introduced into the two RING protomers, which is not feasible in an unfused, homodimeric RING domain. Mutation Y193A is marked with and E2-binding site mutations (M140A/R181A) are marked with . Isopeptide-linked UbcH5A(C85K)–Ub and UbcH5A(S22R/C85K)–Ub showed the same behaviour in this binding assay as the UbcH5A(C85S)–Ub oxyester. d, Isopeptide bond-linked UbcH5A–Ub is a potent inhibitor of RNF4-mediated ubiquitination. UbcH5A(C85K)–Ub was added at indicated concentrations to single-turnover substrate ubiquitination reactions containing ~3 μM UbcH5A–Ub thioester, 0.275 μM RNF4, and 5.5 μM substrate (4 × SUMO-2) radiolabelled with iodine-125. Substrate ubiquitination activity was determined as described in Methods. An equimolar mixture of free UbcH5A (wild-type) and ubiquitin did not show any inhibition at concentrations tested.
SUPPLEMENTARY INFORMATION
6 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
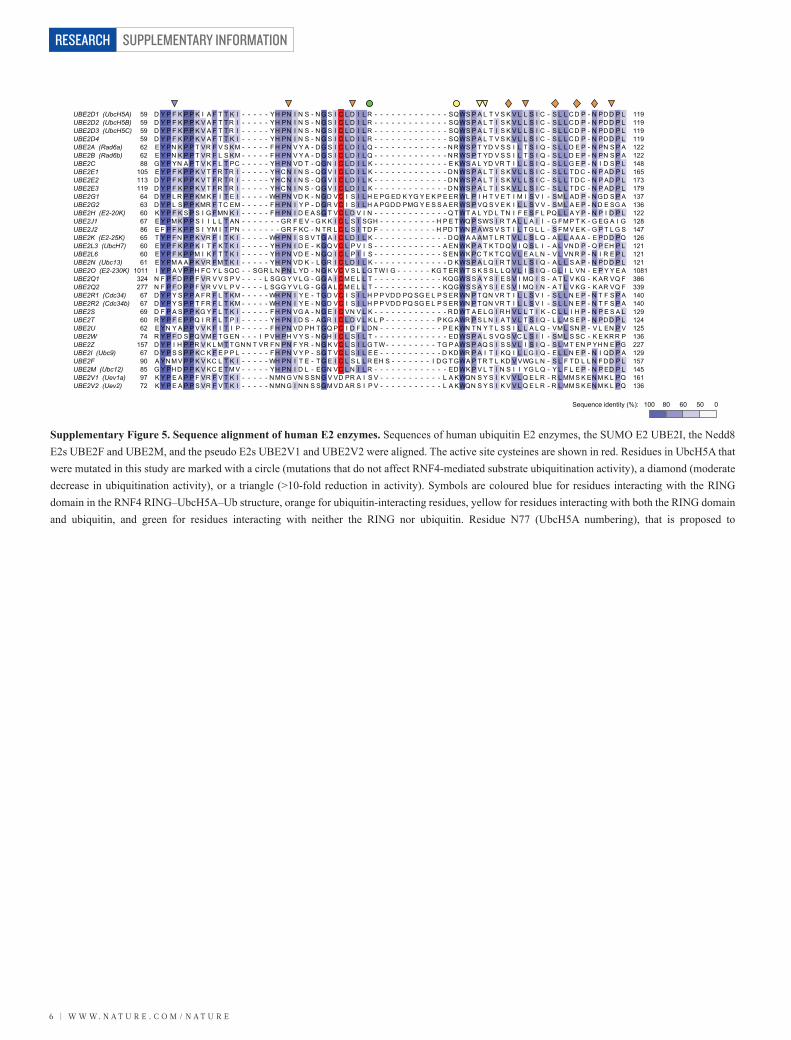
UBE2D1 (UbcH5A)UBE2D2 (UbcH5B)UBE2D3 (UbcH5C)UBE2D4UBE2A (Rad6a)UBE2B (Rad6b)UBE2CUBE2E1UBE2E2UBE2E3UBE2G1UBE2G2UBE2H (E2-20K)UBE2J1UBE2J2UBE2K (E2-25K)UBE2L3 (UbcH7)UBE2L6UBE2N (Ubc13)UBE2O (E2-230K)UBE2Q1UBE2Q2UBE2R1 (Cdc34)UBE2R2 (Cdc34b)UBE2SUBE2TUBE2UUBE2WUBE2ZUBE2I (Ubc9)UBE2FUBE2M (Ubc12)UBE2V1 (Uev1a)UBE2V2 (Uev2)
59595959626288
105113119646360678665606061
1011324277676769606274
1576790859772
1191191191191221221481651731791371361221281471261211211211081386339140140129124125136227129157145161136
D Y P F K P P K I A F T T K I - - - - - YH PN I N S - N G S I C L D I L R - - - - - - - - - - - - - SQWS P A L T V S K V L L S I C - S L L C D P - N PD D P LD Y P F K P P K V A F T T R I - - - - - YH PN I N S - N G S I C L D I L R - - - - - - - - - - - - - SQWS P A L T I S K V L L S I C - S L L C D P - N PD D P LD Y P F K P P K V A F T T R I - - - - - YH PN I N S - N G S I C L D I L R - - - - - - - - - - - - - SQWS P A L T I S K V L L S I C - S L L C D P - N PD D P LD Y P F K P P K V A F T T K I - - - - - YH PN I N S - N G S I C L D I L R - - - - - - - - - - - - - SQWS P A L T V S K V L L S I C - S L L C D P - N PD D P LE Y PN K P P T VR F V S KM - - - - - F H PN V Y A - D G S I C L D I L Q - - - - - - - - - - - - - N R WS P T YD V S S I L T S I Q - S L L D E P - N PN S P AE Y PN K P P T VR F L S KM - - - - - F H PN V Y A - D G S I C L D I L Q - - - - - - - - - - - - - N R WS P T YD V S S I L T S I Q - S L L D E P - N PN S P AG Y P YN A P T V K F L T PC - - - - - YH PN VD T - QGN I C L D I L K - - - - - - - - - - - - - E KWS A L YD VR T I L L S I Q - S L L G E P - N I D S P LE Y P F K P P K V T F R T R I - - - - - YH C N I N S - QG V I C L D I L K - - - - - - - - - - - - - D N WS P A L T I S K V L L S I C - S L L TD C - N P AD P LD Y P F K P P K V T F R T R I - - - - - YH C N I N S - QG V I C L D I L K - - - - - - - - - - - - - D N WS P A L T I S K V L L S I C - S L L TD C - N P AD P LD Y P F K P P K V T F R T R I - - - - - YH C N I N S - QG V I C L D I L K - - - - - - - - - - - - - D N WS P A L T I S K V L L S I C - S L L TD C - N P AD P LD Y P L R P P KM K F I T E I - - - - - WH PN VD K - N GD VC I S I L H E PG ED K YG Y E K P E ER WL P I H T V E T I M I S V I - SM L AD P - N GD S P AD Y P L S P P KMR F T C EM - - - - - F H PN I Y P - D GR VC I S I L H A PGD D PMG Y E S S A ER WS P VQ S V E K I L L S V V - SM L A E P - N D E SG AK Y P F K S P S I G F MN K I - - - - - F H PN I D E A SG T VC L D V I N - - - - - - - - - - - - - Q TWT A L YD L TN I F E S F L PQ L L A Y P - N P I D P LE Y PM K P P S I I L L T AN - - - - - - - GR F E V - G K K I C L S I SGH - - - - - - - - - - H P E TWQ P SWS I R T A L L A I I - G FM P T K - G EG A I GE F P F K P P S I YM I T PN - - - - - - - GR F KC - N TR L C L S I TD F - - - - - - - - - - H PD TWN P AWS V S T I L TG L L - S FM V E K - G P T L G ST Y P F N P P K VR F I T K I - - - - - WH PN I S S V T G A I C L D I L K - - - - - - - - - - - - - D QWA A AM T L R T V L L S L Q - A L L A A A - E PD D PQE Y P F K P P K I T F K T K I - - - - - YH PN I D E - KGQ VC L P V I S - - - - - - - - - - - - A EN WK P A T K TD Q V I Q S L I - A L VN D P - Q P EH P LE Y P F K P PM I K F T T K I - - - - - YH PN VD E - N GQ I C L P I I S - - - - - - - - - - - - S EN WK PC T K TC Q V L E A L N - V L VN R P - N I R E P LE Y PM A A P K VR F M T K I - - - - - YH PN VD K - L GR I C L D I L K - - - - - - - - - - - - - D KWS P A L Q I R T V L L S I Q - A L L S A P - N PD D P LI Y P A V P PH F C Y L SQC - - SGR L N PN L YD - N G K VC V S L L G TW I G - - - - - - KG T ER WT S K S S L L Q V L I S I Q - G L I L VN - E P Y Y E AN F P F D P P F VR V V S P V - - - - L SGG Y V L G - GG A I C M E L L T - - - - - - - - - - - - KQGWS S A Y S I E S V I MQ I S - A T L V KG - K AR VQ FN F P F D P P F VR V V L P V - - - - L SGG Y V L G - GG A L C M E L L T - - - - - - - - - - - - KQGWS S A Y S I E S V I MQ I N - A T L V KG - K AR VQ FD Y P Y S P P A F R F L T KM - - - - - WH PN I Y E - T GD VC I S I L H P P VD D PQ SG E L P S ER WN P TQN VR T I L L S V I - S L L N E P - N T F S P AD Y P Y S P P T F R F L T KM - - - - - WH PN I Y E - N GD VC I S I L H P P VD D PQ SG E L P S ER WN P TQN VR T I L L S V I - S L L N E P - N T F S P AD F P A S P P KG Y F L T K I - - - - - F H PN VG A - N G E I C VN V L K - - - - - - - - - - - - - R D WT A E L G I R H V L L T I K - C L L I H P - N P E S A LR Y P F E P PQ I R F L T P I - - - - - YH PN I D S - AGR I C L D V L K L P - - - - - - - - - P KG AWR P S L N I A T V L T S I Q - L L M S E P - N PD D P LE YN Y A P P V V K F I T I P - - - - - F H PN VD PH T GQ PC I D F L D N - - - - - - - - - - - P E KWN TN Y T L S S I L L A L Q - VM L SN P - V L EN P VR Y P F D S PQ VM F T G EN - - - I P VH PH V Y S - N GH I C L S I L T - - - - - - - - - - - - - ED WS P A L S VQ S VC L S I I - SM L S SC - K E KR R PD Y P I H P PR V K L M T T GN N T VR F N PN F YR - N G K VC L S I L G TW - - - - - - - - - TG P AWS P AQ S I S S V L I S I Q - S L M T EN P YH N E PGD Y P S S P P KC K F E P P L - - - - - F H PN V Y P - SG T VC L S I L E E - - - - - - - - - - - D KD WR P A I T I KQ I L L G I Q - E L L N E P - N I QD P AA YN M V P P K V KC L T K I - - - - - WH PN I T E - T G E I C L S L L R EH S - - - - - - - I D G TGWA P TR T L KD V VWG L N - S L F TD L L N FD D P LG Y PH D P P K V KC E T M V - - - - - YH PN I D L - EGN VC L N I L R - - - - - - - - - - - - - ED WK P V L T I N S I I YG L Q - Y L F L E P - N P ED P LK Y P E A P P F VR F V T K I - - - - - N MN G VN S SN G V VD PR A I S V - - - - - - - - - - - L A KWQN S Y S I K V V L Q E L R - R L MM S K EN M K L PQK Y P E A P P S VR F V T K I - - - - - N MN G I N N S SGM VD AR S I P V - - - - - - - - - - - L A KWQN S Y S I K V V L Q E L R - R L MM S K EN M K L PQ
Supplementary Figure 5. Sequence alignment of human E2 enzymes. Sequences of human ubiquitin E2 enzymes, the SUMO E2 UBE2I, the Nedd8 E2s UBE2F and UBE2M, and the pseudo E2s UBE2V1 and UBE2V2 were aligned. The active site cysteines are shown in red. Residues in UbcH5A that were mutated in this study are marked with a circle (mutations that do not affect RNF4-mediated substrate ubiquitination activity), a diamond (moderate decrease in ubiquitination activity), or a triangle (>10-fold reduction in activity). Symbols are coloured blue for residues interacting with the RING domain in the RNF4 RING–UbcH5A–Ub structure, orange for ubiquitin-interacting residues, yellow for residues interacting with both the RING domain and ubiquitin, and green for residues interacting with neither the RING nor ubiquitin. Residue N77 (UbcH5A numbering), that is proposed to
100Sequence identity (%): 80 60 50 0

W W W. N A T U R E . C O M / N A T U R E | 7
SUPPLEMENTARY INFORMATION RESEARCH
stabilize the tetrahedral intermediate during the nucleophilic attack on the activated thioester bond, is highly conserved in active E2 enzymes. Residue D87 in UbcH5A hydogen bonds to the main chain of ubiquitin R74 in the RNF4 RING–UbcH5A–Ub structure. Mutation D87A abolished activity, indicating that this interaction is critical for locking the C-terminal tail of ubiquitin in the E2 active site groove. Aspartate or serine is found in this position in most of the E2 enzymes that were shown to be active with RING-type E3 ligases. Serine can be engaged in a similar hydrogen bonding interaction as aspartate. This was indeed observed in the SUMO–RanGAP1–Ubc9–RanBP2 complex23,25 (see Supplementary Fig. 17c). D117, a residue proposed to position and activate the incoming substrate lysine, is often aspartate or serine in other E2s. It was shown that D127S Ubc9 (corresponding to D117 in UbcH5A) retains activity, but a decrease in catalytic efficiency was observed21, suggesting that E2s with a serine in this position might be less efficient in activating the incoming substrate lysine. In the RING–UbcH5A–Ub complex, ubiquitin is folded back onto the E2, contacting the α2 helix of UbcH5A. This interaction is centered on L104 in UbcH5A and I44 in ubiquitin. A hydrophobic residue is found in the position of UbcH5A L104 in most of the E2 enzymes that were shown to be active with RING E3s.
Supplementary Figure 6. The active site groove shows higher conservation than other surfaces in E2 enzymes. Conserved residues present in active human E2 enzymes (Supplementary Fig. 5) were mapped onto the surface of UbcH5A in the RNF4 RING–UbcH5A–Ub complex. Identical residues are indicated in red, highly conserved residues in pink and conserved residues in green. Ubiquitin and the RNF4 RING domain are shown in ribbon representation, with ubiquitin coloured orange and the two protomers in the dimeric RING domain coloured cyan and blue. Conserved residues surround the E2 active site groove, the region that makes extensive contacts with the C-terminal tail of ubiquitin in the RNF4 RING–UbcH5A–Ub structure. A patch of conserved residues is also located at the RING interaction interface.
UbcH5A
Ubiquitin
RNF4 RING
RNF4 RING

SUPPLEMENTARY INFORMATION
8 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
UbcH5A
RING
RING
Ubiquitin
UbcH5A
RING
RINGK4
M1
P61
E9
R5
F62
L97P95
S94
Q92
M140
D141
I138
R181
P137F162
T179
P178
C139
C163
C158 C180
C177H160
A96
a b
Supplementary Figure 7. Interaction interface between the RING domain of RNF4 and UbcH5A. a, Overall view of the complex of the dimeric RNF4 RING domain bound to ubiquitin-loaded UbcH5A (top panel) and the same complex with ubiquitin removed to show the interaction interface between the RING domain and UbcH5A (bottom panel). The two monomers in the dimeric RING domain are coloured cyan and blue, UbcH5A is coloured green, and ubiquitin is orange. Zinc ions are shown as grey spheres. b, Detailed view of the interaction interface between one monomer of the RNF4 RING (blue) and UbcH5A (green). Residues at the interface and zinc-coordinating residues in the RING domain are shown as sticks, hydrogen bonds are indicated as black dashes. In UbcH5A, the interaction interface is formed by the α1 helix (residues K4 to D12) and by two loops spanning residues P61 to K63 and Q92 to T98, respectively. The latter loop contains an SPA motif (S94, P95 and A96) that is conserved in many E2 enzymes. UbcH5A residues P61 and F62 together with the SPA motif insert into a shallow hydrophobic groove on the surface of the RING domain delineated by residues P137 and I138 from the first zinc-binding loop, F162, the α2 helix residues (D169 to N173), and P178 from the loop chelating the second zinc ion.
α1
α2
α1
α2
W W W. N A T U R E . C O M / N A T U R E | 9
SUPPLEMENTARY INFORMATION RESEARCH
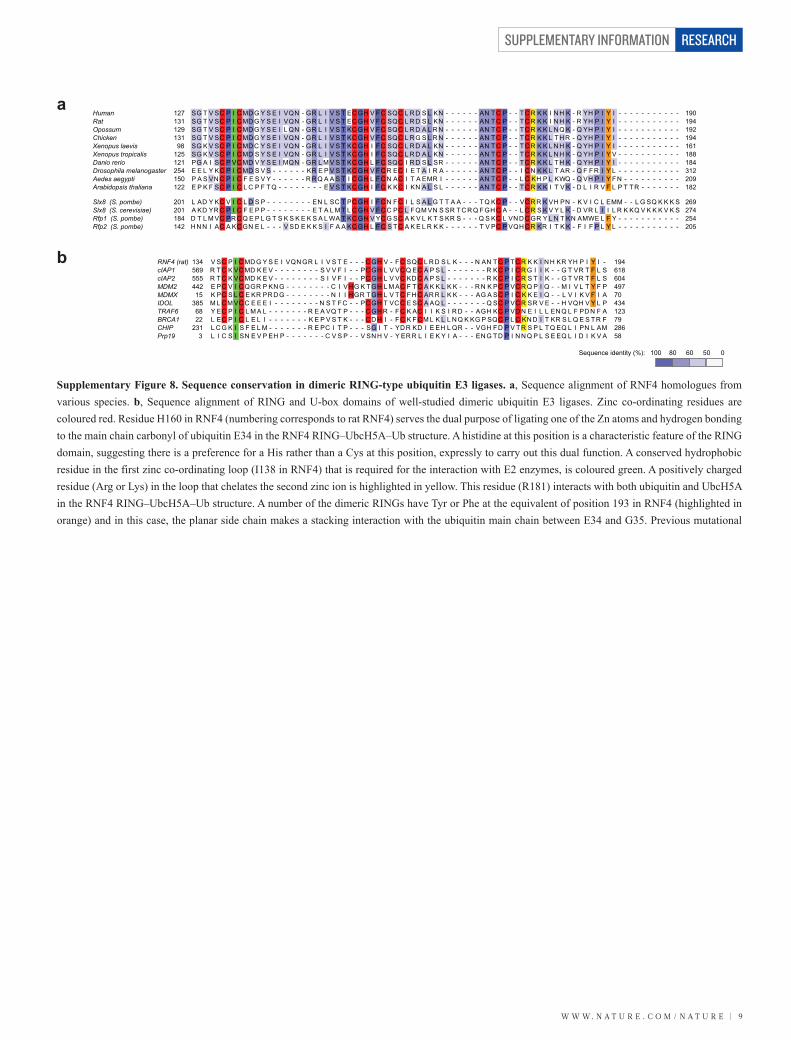
HumanRatOpossumChickenXenopus laevisXenopus tropicalisDanio rerioDrosophila melanogasterAedes aegyptiArabidopsis thaliana
Slx8 (S. pombe)Slx8 (S. cerevisiae)Rfp1 (S. pombe)Rfp2 (S. pombe)
12713112913198
125121254150122
201201184142
190194192194161188184312209182
269274254205
SG T V SC P I C MD G Y S E I VQN - GR L I V S T EC GH V F C SQC L R D S L KN - - - - - - AN TC P - - TC R K K I N H K - R YH P I Y I - - - - - - - - - - -SG T V SC P I C MD G Y S E I VQN - GR L I V S T EC GH V F C SQC L R D S L KN - - - - - - AN TC P - - TC R K K I N H K - R YH P I Y I - - - - - - - - - - -SG T V SC P I C MD G Y S E I L QN - GR L I V S T KC GH V F C SQC L R D A L R N - - - - - - AN TC P - - TC R K K L N Q K - Q YH P I Y I - - - - - - - - - - -SG T V SC P I C MD G Y S E I VQN - GR L I V S T KC GH V F C SQC L R G S L R N - - - - - - AN TC P - - TC R K K L TH R - Q YH P I Y I - - - - - - - - - - -SG K V SC P I C MD C Y S E I VQN - GR L I V S T KC GH I FC SQC L R D A L KN - - - - - - AN TC P - - TC R K K L N H K - Q YH P I Y I - - - - - - - - - - -SG K V SC P I C MD S Y S E I VQN - GR L I V S T KC GH I FC SQC L R D A L KN - - - - - - AN TC P - - TC R K K L N H K - Q YH P I Y V - - - - - - - - - - -PG A I SC P VC MD V Y S E I MQN - GR L M V S T KC GH L F C SQC I R D S L SR - - - - - - AN TC P - - TC R K K L TH K - Q YH P I Y I - - - - - - - - - - -E E L Y KC P I C MD S V S - - - - - - KR E P V S T KC GH V F C R EC I E T A I R A - - - - - - AN TC P - - I C N K K L T AR - Q F FR I Y L - - - - - - - - - - -P A S VN C P I C F E S V Y - - - - - - R R Q A A S T I C GH L F C N AC I T A EMR I - - - - - - AN TC P - - L C KH P L KWQ - Q VH P I Y FN - - - - - - - - - -E P K F SC P I C L C P F T Q - - - - - - - - E V S T KC GH I FC K KC I KN A L S L - - - - - - AN TC P - - TC R K K I T V K - D L I R V F L P T TR - - - - - - -
L AD Y KC V I C L D S P - - - - - - - - EN L SC T PC GH I FC N FC I L S A L G T T A A - - - TQ KC P - - VC R R K VH PN - K V I C L EMM - - L G SQ K K K SA KD YR C P I C F E P P - - - - - - - - E T A L M T L C GH V F C C PC L FQM VN S SR TC R Q FGH C A - - L C R S K V Y L K - D VR L I I L R K KQ V K K K V K SD T L M VC PR C Q E P L G T S K S K E K S A L WA T KC GH V YC G SC A K V L K T S KR S - - - Q S KC L VN D C GR Y L N T KN AMWE L F Y - - - - - - - - - - -H N N I AC A KC GN E L - - - V SD E K K S I F A A KC GH L FC S TC A K E L R K K - - - - - - T V PC P VQH C R KR I T K K - F I F P L Y L - - - - - - - - - - -
Supplementary Figure 8. Sequence conservation in dimeric RING-type ubiquitin E3 ligases. a, Sequence alignment of RNF4 homologues from various species. b, Sequence alignment of RING and U-box domains of well-studied dimeric ubiquitin E3 ligases. Zinc co-ordinating residues are coloured red. Residue H160 in RNF4 (numbering corresponds to rat RNF4) serves the dual purpose of ligating one of the Zn atoms and hydrogen bonding to the main chain carbonyl of ubiquitin E34 in the RNF4 RING–UbcH5A–Ub structure. A histidine at this position is a characteristic feature of the RING domain, suggesting there is a preference for a His rather than a Cys at this position, expressly to carry out this dual function. A conserved hydrophobic residue in the first zinc co-ordinating loop (I138 in RNF4) that is required for the interaction with E2 enzymes, is coloured green. A positively charged residue (Arg or Lys) in the loop that chelates the second zinc ion is highlighted in yellow. This residue (R181) interacts with both ubiquitin and UbcH5A in the RNF4 RING–UbcH5A–Ub structure. A number of the dimeric RINGs have Tyr or Phe at the equivalent of position 193 in RNF4 (highlighted in orange) and in this case, the planar side chain makes a stacking interaction with the ubiquitin main chain between E34 and G35. Previous mutational
100Sequence identity (%): 80 60 50 0
RNF4 (rat)cIAP1cIAP2MDM2MDMXIDOLTRAF6BRCA1CHIPPrp19
13456955544215
3856822
2313
194618604497704341237928658
V SC P I C MD G Y S E I VQN GR L I V S T E - - - C GH V - F C SQC L R D S L K - - - N AN TC P TC R K K I N H KR YH P I Y I -R T C K VC MD K E V - - - - - - - - S V V F I - - PC GH L V VC Q EC A P S L - - - - - - - R KC P I C R G I I K - - G T VR T F L SR T C K VC MD K E V - - - - - - - - S I V F I - - PC GH L V VC KD C A P S L - - - - - - - R KC P I C R S T I K - - G T VR T F L SE PC V I C QGR P KN G - - - - - - - - C I VH G K TGH L M AC F TC A K K L K K - - - R N K PC P VC R Q P I Q - - M I V L T Y F PK PC S L C E KR PR D G - - - - - - - - N I I H GR TGH L V TC FH C AR R L K K - - - AG A SC P I C K K E I Q - - L V I K V F I AM L C M VC C E E E I - - - - - - - - N S T FC - - PC GH T VC C E SC A AQ L - - - - - - - Q SC P VC R SR V E - - H VQH V Y L PY EC P I C L M A L - - - - - - - R E A VQ T P - - - C GH R - F C K AC I I K S I R D - - AGH KC P VD N E I L L EN Q L F PD N F AL EC P I C L E L I - - - - - - - K E P V S T K - - - C D H I - FC K F C M L K L L N Q K KG P SQC P L C KN D I T KR S L Q E S TR FL C G K I S F E L M - - - - - - - R E PC I T P - - - SG I T - YD R KD I E EH L QR - - VGH FD P V TR S P L TQ EQ L I PN L AML I C S I SN E V P EH P - - - - - - - C V S P - - V SN H V - Y ER R L I E K Y I A - - - EN G TD P I N N Q P L S E EQ L I D I K V A
a
b
SUPPLEMENTARY INFORMATION
1 0 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
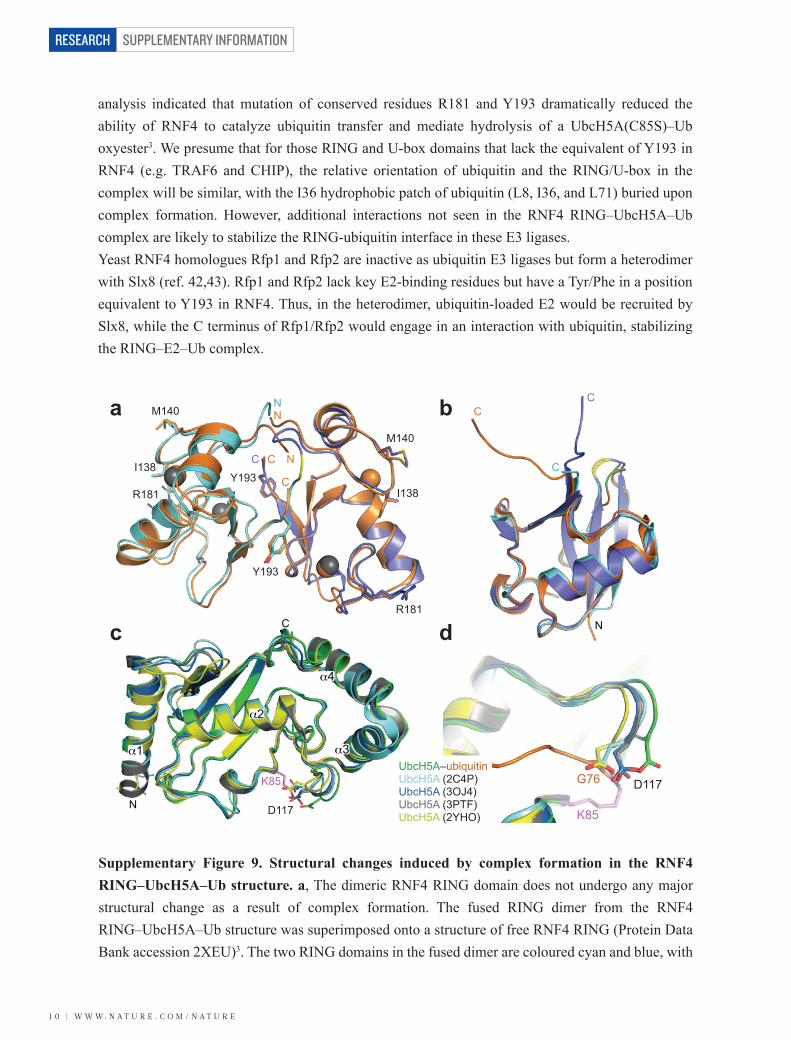
analysis indicated that mutation of conserved residues R181 and Y193 dramatically reduced the ability of RNF4 to catalyze ubiquitin transfer and mediate hydrolysis of a UbcH5A(C85S)–Ub oxyester3. We presume that for those RING and U-box domains that lack the equivalent of Y193 in RNF4 (e.g. TRAF6 and CHIP), the relative orientation of ubiquitin and the RING/U-box in the complex will be similar, with the I36 hydrophobic patch of ubiquitin (L8, I36, and L71) buried upon complex formation. However, additional interactions not seen in the RNF4 RING–UbcH5A–Ub complex are likely to stabilize the RING-ubiquitin interface in these E3 ligases.Yeast RNF4 homologues Rfp1 and Rfp2 are inactive as ubiquitin E3 ligases but form a heterodimer with Slx8 (ref. 42,43). Rfp1 and Rfp2 lack key E2-binding residues but have a Tyr/Phe in a position equivalent to Y193 in RNF4. Thus, in the heterodimer, ubiquitin-loaded E2 would be recruited by Slx8, while the C terminus of Rfp1/Rfp2 would engage in an interaction with ubiquitin, stabilizing the RING–E2–Ub complex.
a
c d
b
α2
α3
α4
α1
D117
K85 D117
K85
G76UbcH5A–ubiquitinUbcH5A (2C4P)UbcH5A (3OJ4)
UbcH5A (2YHO)UbcH5A (3PTF)
M140
I138
R181
M140
I138
R181
Y193
Y193
NN
NCC
C
N
C
CC
C
N
Supplementary Figure 9. Structural changes induced by complex formation in the RNF4 RING–UbcH5A–Ub structure. a, The dimeric RNF4 RING domain does not undergo any major structural change as a result of complex formation. The fused RING dimer from the RNF4 RING–UbcH5A–Ub structure was superimposed onto a structure of free RNF4 RING (Protein Data Bank accession 2XEU)3. The two RING domains in the fused dimer are coloured cyan and blue, with

W W W. N A T U R E . C O M / N A T U R E | 1 1
SUPPLEMENTARY INFORMATION RESEARCH
the linker region (a single glycine residue) coloured yellow. The uncomplexed RNF4 RING is shown in orange. Key residues involved in interactions with E2 (I138 and M140), ubiquitin (Y193), and both E2 and ubiquitin (R181) are shown as sticks. Zinc ions are shown as spheres and N- and C-termini are labelled with N and C, respectively. b, Superposition of ubiquitin from the RNF4 RING–UbcH5A–Ub complex (orange) with free ubiquitin structures 1UBQ32 (blue) and 3ONS44 (residues 1-72, cyan). Residue L8 is highlighted in green (complexed ubiquitin) or yellow (structures of free ubiquitin). The loop at L8 moves over 4 Å to contact both UbcH5A and the RING in the RNF4 RING–UbcH5A–Ub complex. c, Superposition of UbcH5A from the present structure (green) with structures of free UbcH5A (Protein Data Bank accession 2C4P, cyan) and UbcH5A in a variety of non-covalent complexes: UbcH5A bound to the IDOL RING domain14 (Protein Data Bank accession 2YHO, yellow), UbcH5A in complex with ubiquitin12 (Protein Data Bank accession 3PTF, grey), and UbcH5A–ubiquitin–A20 zinc finger 4 complex13 (Protein Data Bank accession 3OJ4, blue). In the RING–UbcH5A–Ub complex, the α1 helix of UbcH5A and the loop which connects to the β1 strand adjust their position to avoid clashing with K6, G47 and H68 of ubiquitin. This loop, however, varies in other structures of the E2, suggesting it is a mobile region. The D117 loop moves as a consequence of isopeptide bond formation between the C terminus of ubiquitin and K85 in UbcH5A (shown in sticks representation and coloured violet). The unconjugated structures all have the side chain of D117 (shown as sticks) pointing towards C85 in a position that would clash with the isopeptide (thioester) bond. d, Detailed view of the D117 loop in the UbcH5A structures shown in c. The isopeptide bond that connects the C-terminal tail of ubiquitin (orange) with UbcH5A residue K85 (violet) in the RNF4 RING–UbcH5A–Ub structure, is shown as sticks.
SUPPLEMENTARY INFORMATION
1 2 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
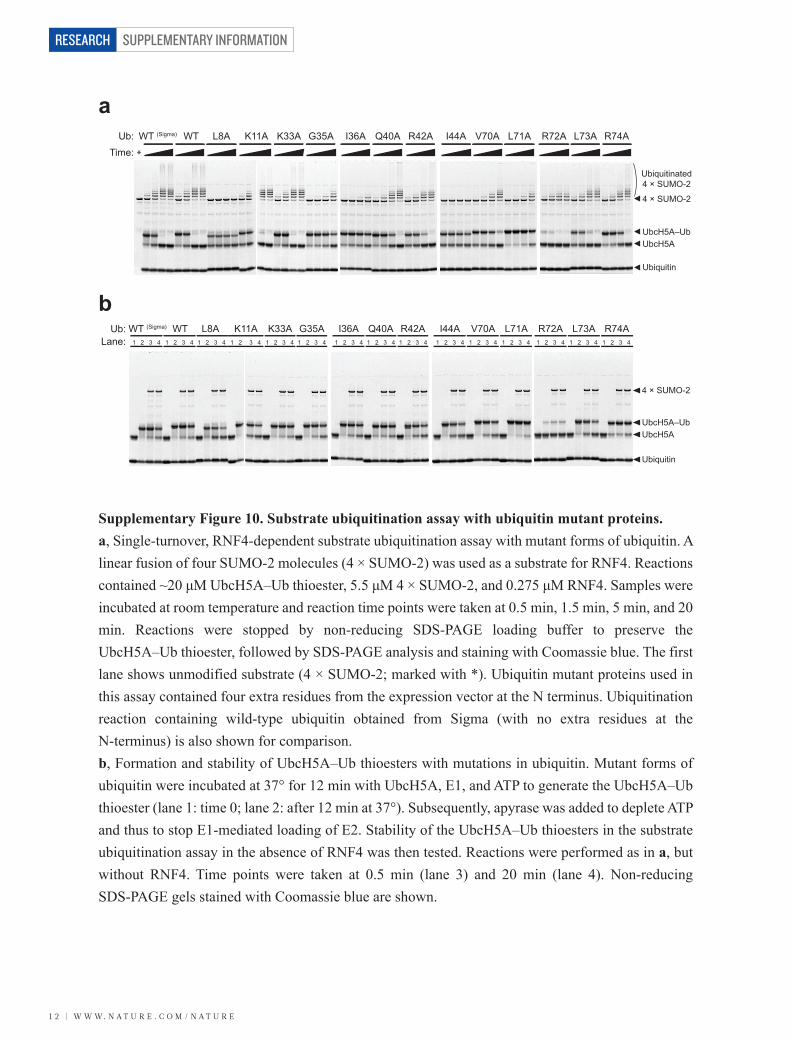
Ub:Time:
WT (Sigma) WT L8A K11A K33A G35A I36A Q40A R42A I44A V70A L71A R72A R74AL73A
UbcH5A UbcH5A–Ub
Ubiquitin
4 × SUMO-2
Ubiquitinated4 × SUMO-2
UbcH5A UbcH5A–Ub
Ubiquitin
4 × SUMO-2
I36A Q40A R42A1 2 3 4 1 2 3 4 1 2 3 4 1
I44A V70A L71A2 3 4 1 2 3 4 1 2 3 4
R72A R74AL73A1 2 3 4 1 2 3 4 1 2 3 4
Ub:Lane: 1
WT (Sigma) WT L8A K11A K33A G35A2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 41 2 3 4
a
b
Supplementary Figure 10. Substrate ubiquitination assay with ubiquitin mutant proteins.a, Single-turnover, RNF4-dependent substrate ubiquitination assay with mutant forms of ubiquitin. A linear fusion of four SUMO-2 molecules (4 × SUMO-2) was used as a substrate for RNF4. Reactions contained ~20 μM UbcH5A–Ub thioester, 5.5 μM 4 × SUMO-2, and 0.275 μM RNF4. Samples were incubated at room temperature and reaction time points were taken at 0.5 min, 1.5 min, 5 min, and 20 min. Reactions were stopped by non-reducing SDS-PAGE loading buffer to preserve the UbcH5A–Ub thioester, followed by SDS-PAGE analysis and staining with Coomassie blue. The first lane shows unmodified substrate (4 × SUMO-2; marked with *). Ubiquitin mutant proteins used in this assay contained four extra residues from the expression vector at the N terminus. Ubiquitination reaction containing wild-type ubiquitin obtained from Sigma (with no extra residues at the N-terminus) is also shown for comparison. b, Formation and stability of UbcH5A–Ub thioesters with mutations in ubiquitin. Mutant forms of ubiquitin were incubated at 37° for 12 min with UbcH5A, E1, and ATP to generate the UbcH5A–Ub thioester (lane 1: time 0; lane 2: after 12 min at 37°). Subsequently, apyrase was added to deplete ATP and thus to stop E1-mediated loading of E2. Stability of the UbcH5A–Ub thioesters in the substrate ubiquitination assay in the absence of RNF4 was then tested. Reactions were performed as in a, but without RNF4. Time points were taken at 0.5 min (lane 3) and 20 min (lane 4). Non-reducing SDS-PAGE gels stained with Coomassie blue are shown.
*
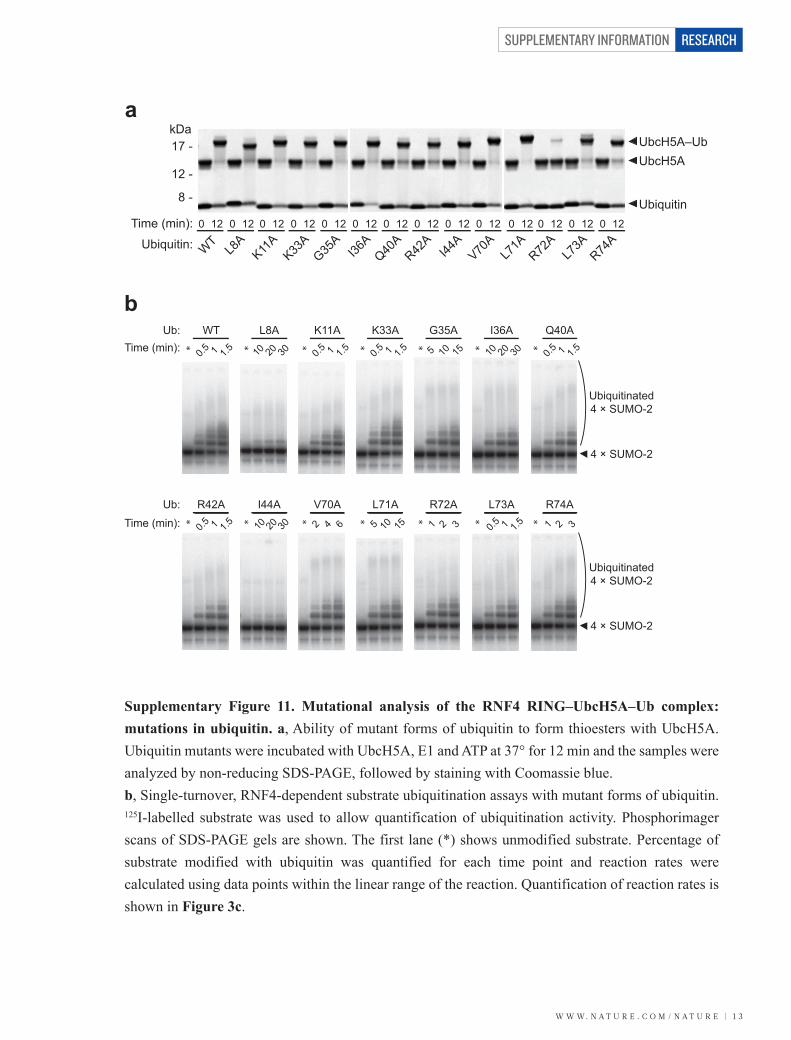
W W W. N A T U R E . C O M / N A T U R E | 1 3
SUPPLEMENTARY INFORMATION RESEARCH
Supplementary Figure 11. Mutational analysis of the RNF4 RING–UbcH5A–Ub complex: mutations in ubiquitin. a, Ability of mutant forms of ubiquitin to form thioesters with UbcH5A. Ubiquitin mutants were incubated with UbcH5A, E1 and ATP at 37° for 12 min and the samples were analyzed by non-reducing SDS-PAGE, followed by staining with Coomassie blue. b, Single-turnover, RNF4-dependent substrate ubiquitination assays with mutant forms of ubiquitin. 125I-labelled substrate was used to allow quantification of ubiquitination activity. Phosphorimager scans of SDS-PAGE gels are shown. The first lane (*) shows unmodified substrate. Percentage of substrate modified with ubiquitin was quantified for each time point and reaction rates were calculated using data points within the linear range of the reaction. Quantification of reaction rates is shown in Figure 3c.
UbcH5A–Ub UbcH5A
Ubiquitin
Ubiquitin:
120
WT
120
L8A
Time (min): 120
K11A
120
K33A
120
G35A
120
I36A
120
Q40A
120
R42A
120
I44A
120
V70A
120
L71A
120
R72A
120
L73A
120
R74A
17 -kDa
12 -
8 -
a
bUb:
Time (min): 0.5 1 1.5
WT
0.5 1 1.5 0.5 1 1.5
K11A
0.5 1 1.5
K33A
10 20 30
L8A
10 20 30
I36A Q40A
5 10 15
G35A
4 × SUMO-2
Ubiquitinated4 × SUMO-2
* * * * * * *
Ub:
Time (min): 0.5 1 1.5
R42A
0.5 1 1.5
L73A
10 20 30
I44A
2 4 6
V70A
5 10 15
L71A
1 2 3
R72A
1 2 3
R74A
4 × SUMO-2
Ubiquitinated4 × SUMO-2
* * * * * * *

SUPPLEMENTARY INFORMATION
1 4 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
a
b
UbcH5A:Time:
WT N77A D87A R90A Q92A L97A K101A D112A N114A D117A
UbcH5A UbcH5A–Ub
Ubiquitin
4 × SUMO-2
Ubiquitinated4 × SUMO-2
UbcH5A:Lane:
WT N77A
UbcH5A UbcH5A–Ub
Ubiquitin
4 × SUMO-2
1 2 3 4 1 2 3 4 1
D87A R90A Q92A2 3 4 1 2 3 4 1 2 3 4
L97A K101A D112A1 2 3 4 1 2 3 4 1 2 3 4
N114A D117A1 2 3 4 1 2 3 4
Supplementary Figure 12. Substrate ubiquitination assay with UbcH5A mutant proteins.a, Single-turnover, RNF4-dependent substrate ubiquitination assay with mutant forms of UbcH5A. Reactions were performed as described in Supplementary Figure 10a. Reactions were stopped by non-reducing SDS-PAGE loading buffer, followed by SDS-PAGE analysis. Coomassie blue-stained gels are shown.b, Formation and stability of UbcH5A–Ub thioesters with mutations in UbcH5A. The experiment was performed as described in Supplementary Figure 10b. Lanes 1 and 2: formation of UbcH5A–Ub thioesters. Mutant forms of UbcH5A were incubated with ubiquitin, E1, and ATP to generate the UbcH5A–Ub thioester (lane 1: time 0; lane 2: after 12 min at 37°). Lanes 3 and 4: stability of the UbcH5A–Ub thioesters in the substrate ubiquitination assay in the absence of RNF4. Time points were taken at 0.5 min (lane 3) and 20 min (lane 4). Non-reducing SDS-PAGE gels stained with Coomassie blue are shown.
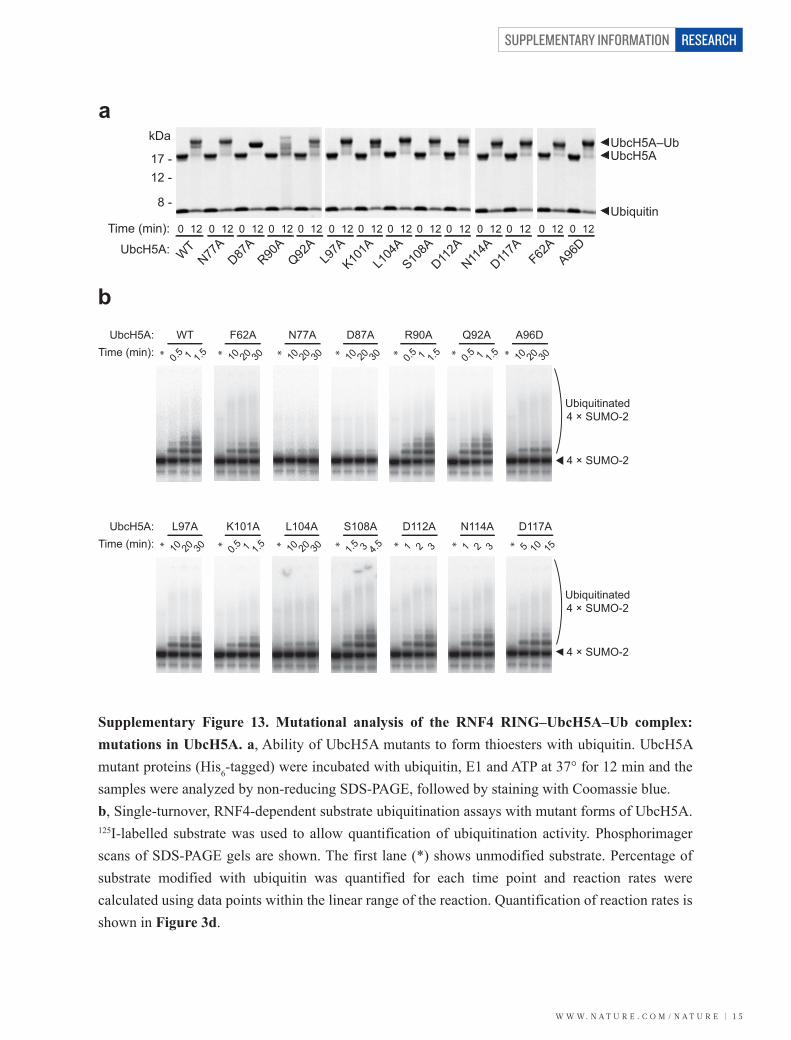
W W W. N A T U R E . C O M / N A T U R E | 1 5
SUPPLEMENTARY INFORMATION RESEARCH
Supplementary Figure 13. Mutational analysis of the RNF4 RING–UbcH5A–Ub complex: mutations in UbcH5A. a, Ability of UbcH5A mutants to form thioesters with ubiquitin. UbcH5A mutant proteins (His6-tagged) were incubated with ubiquitin, E1 and ATP at 37° for 12 min and the samples were analyzed by non-reducing SDS-PAGE, followed by staining with Coomassie blue. b, Single-turnover, RNF4-dependent substrate ubiquitination assays with mutant forms of UbcH5A. 125I-labelled substrate was used to allow quantification of ubiquitination activity. Phosphorimager scans of SDS-PAGE gels are shown. The first lane (*) shows unmodified substrate. Percentage of substrate modified with ubiquitin was quantified for each time point and reaction rates were calculated using data points within the linear range of the reaction. Quantification of reaction rates is shown in Figure 3d.
a
b
UbcH5A:
120
WT
120
N77A
Time (min): 120
D87A
120
R90A
120
Q92A
120
L97A
120
K101A
120
L104
A120
S108A
120
D112A
120
N114A
120
D117A
120
F62A
120
A96D
UbcH5A–Ub UbcH5A
Ubiquitin
17 -
kDa
12 -
8 -
UbcH5A:Time (min): 1.5 3 4.5
S108A
*10 20 30
L104A
* 1 2 3
D112A
* 5 10 15
D117A
*10 20 30
L97A
* 0.5 1 1.5
K101A
*
4 × SUMO-2
Ubiquitinated4 × SUMO-2
1 2 3
N114A
*
UbcH5A:Time (min): 0.5 1 1.5
WT
* 0.5 1 1.5
R90A
* 0.5 1 1.5
Q92A
*10 20 30
N77A
*10 20 30
F62A
* 10 20 30
A96D
*10 20 30
D87A
*
4 × SUMO-2
Ubiquitinated4 × SUMO-2

SUPPLEMENTARY INFORMATION
1 6 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
126 -
35 -25 -17 -12 -
8 -
kDa
126 -
35 -25 -17 -12 -
8 -
kDa
UbcH5A(C85S) UbcH5A–Ub
Ubiquitin
RNF4
E1
UbcH5A(C85S) UbcH5A–Ub
Ubiquitin
RNF4
E1
WT+– – – –
L8A+–
K33A+
G35A+
I36A+
Ub:RNF4:Time:
R42A+–
I44A+–
L71A+– –
R74A+
Ub:RNF4:Time:
Supplementary Figure 14. UbcH5A–Ub oxyester hydrolysis assay with mutant forms of ubiquitin. To generate UbcH5A(C85S)–Ub oxyesters, UbcH5A(C85S) was incubated with ubiquitin (wild-type or mutant) and E1 in the presence of ATP, followed by addition of apyrase to deplete ATP and thus to stop the E1-mediated charging reaction. This material was then incubated at room temperature either in the presence or absence of RNF4. Time points were taken as follows: reactions without RNF4: 2.5 min and 180 min; reactions with RNF4: 2.5 min, 30 min, 60 min, 90 min, and 180 min. Reactions were stopped with SDS-PAGE loading buffer and analyzed by SDS-PAGE. Coomassie blue-stained gels are shown. Quantification of reaction rates is shown in Figure 3e.

W W W. N A T U R E . C O M / N A T U R E | 1 7
SUPPLEMENTARY INFORMATION RESEARCH
UbcH5A(C85S) UbcH5A–Ub RNF4
E1
UbcH5A(C85S) UbcH5A–Ub
Ubiquitin
Ubiquitin
RNF4
E1
WT+–
N77A+–
D87A+–
UbcH5A(C85S):RNF4:Time:
A96D+–
L97A+–
K101A+–
L104A+–
S108A+–
N114A+–
D117A+–
UbcH5A(C85S):RNF4:Time:
126 -kDa
126 -
35 -25 -17 -12 -
8 -
kDa
Supplementary Figure 15. UbcH5A–Ub oxyester hydrolysis assay with mutant forms of UbcH5A. His6-tagged UbcH5A(C85S) mutants were loaded with ubiquitin in E1-mediated reaction, followed by depletion of ATP. The yield of the charging reaction was low for UbcH5A(N77A/C85S) and UbcH5A(C85S/L104A). The UbcH5A(C85S)–Ub oxyesters were then incubated at room temperature either in the presence or absence of RNF4. Time points were taken as follows: reactions without RNF4: 2.5 min and 180 min; reactions with RNF4: 2.5 min, 15 min, 30 min, 60 min, and 180 min. Reactions were stopped with SDS-PAGE loading buffer and analyzed by SDS-PAGE. Coomassie blue-stained gels are shown. Quantification of reaction rates is shown in Figure 3f.
35 -25 -17 -12 -
8 -

SUPPLEMENTARY INFORMATION
1 8 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
UbiquitinUbiquitin
UbcH5AUbc1
G76
G76
C85N77
D117
C88N79
D120
Supplementary Figure 16. The C-terminal tail of ubiquitin adopts a different conformation in the model of free Ubc1–Ub thioester compared to UbcH5A–Ub bound to the RNF4 RING. a, Ubc1 in the NMR model of the Ubc1–Ub thioester15 (Protein Data Bank accession 1FXT) was superimposed onto UbcH5A in the model of the UbcH5A–Ub thioester bound to the RNF4 RING described herein. Ubc1 and ubiquitin in the Ubc1–Ub thioester are coloured grey and bluish grey, respectively. UbcH5A and ubiquitin from the RNF4 RING–UbcH5A–Ub complex are coloured green and orange, respectively. The RNF4 RING domain is not shown. b, Detailed view of the E2 active sites. The active site cysteines of the E2s and the C-terminal G76 of ubiquitin are shown in sticks representation. Proposed catalytic residues N77 and D117 in UbcH5A and the corresponding residues N79 and D120 in Ubc1 are also shown as sticks. Colour scheme is as in a.
a b
UbcH5A–ubiquitin
Ubc1–ubiquitin (1FXT)
W W W. N A T U R E . C O M / N A T U R E | 1 9
SUPPLEMENTARY INFORMATION RESEARCH
N124N114
N124N114
D117 D117
Y87 Y87
L119 L119A129 A129
G76 G76G97 G97
G96 G96G75 G75
R74 R74
L73 L73
UbiquitinUbcH5A
SUMO-1RanGAP1
Ubc9UbiquitinUbcH5A
SUMO-1RanGAP1
Ubc9 D127 D127
N77
N85 N85
N77
R72 R72
L71 L71
E93
T95 T95
E93
Q92 Q92
K524 K524
C93
S95 S95
C93
K85 K85
D87 D87Q94 Q94
Ubc9UbcH5A
SUMO-1
RanBP2Ubiquitin
RNF4 RING
RNF4 RING
RanGAP1
Ubc9UbcH5A
SUMO-1
RanBP2
Ubiquitin
RNF4 RING
RNF4 RINGRanGAP1
a
b
c
Supplementary Figure 17 (continued)
150°

SUPPLEMENTARY INFORMATION
2 0 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
UbcH5A
Ubiquitin
RNF4 RING
RNF4 RING RanGAP1
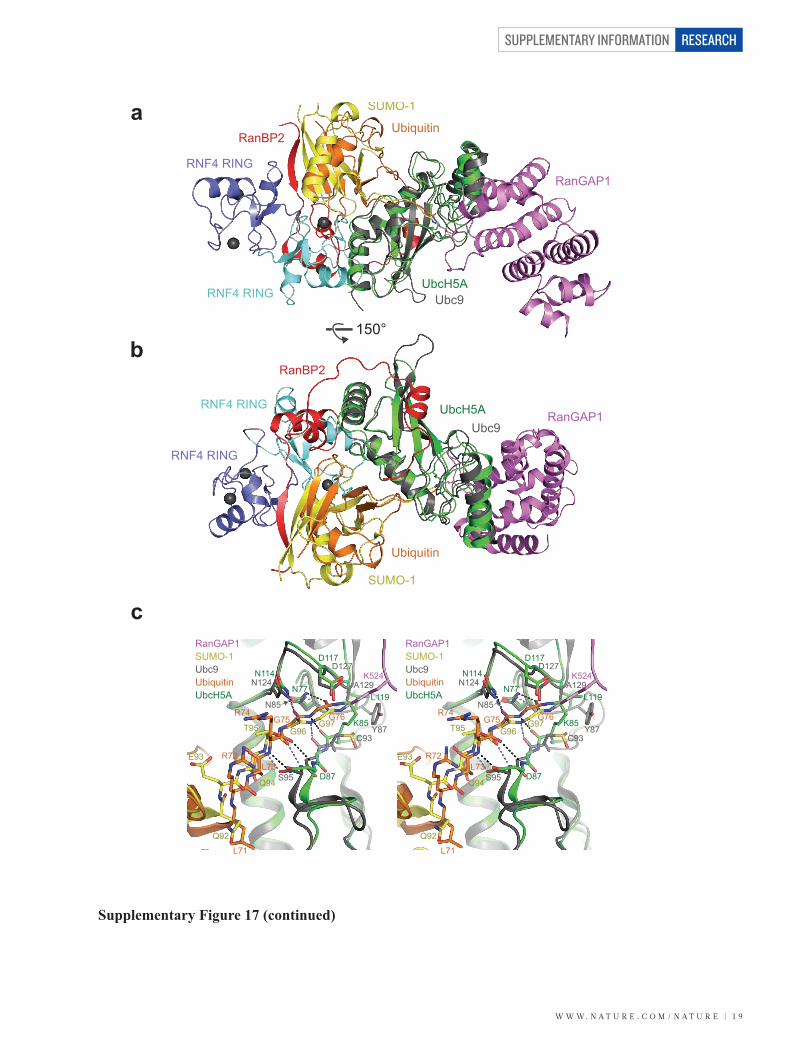
Supplementary Figure 17. Structural comparison of the RNF4 RING–UbcH5A–ubiquitin complex with a complex of SUMO-modified RanGAP1 bound to Ubc9 and the SUMO E3 ligase RanBP2. a, Ubc9 in the SUMO–RanGAP1–Ubc9–RanBP2 complex25 (Protein Data Bank accession 3UIP) was superimposed onto UbcH5A in the RNF4 RING–UbcH5A(S22R/C85K)–Ub complex. The two protomers of the dimeric RNF4 RING domain are coloured cyan and blue, UbcH5A is green, ubiquitin is orange, SUMO-1 is yellow, RanGAP1 is violet, Ubc9 is grey, and RanBP2 is red. Zinc ions are shown as grey spheres. b, As in a, but the structures are rotated by 150° as indicated. c, Stereo view of the active sites of the E2s. Colour scheme is as in a. Hydrogen bonds are indicated by dashed lines coloured either black (RNF4 RING–UbcH5A–Ub complex) or blue (SUMO–RanGAP1–Ubc9–RanBP2 complex). d, Model for a RING–substrate–UbcH5A–Ub thioester complex. The SUMO–RanGAP1–Ubc9–RanBP2 structure provides a model for how a substrate might interact with the RING-activated UbcH5A–Ub thioester. To generate this model, UbcH5A in our model of the RING–UbcH5A–Ub thioester complex was superimposed onto Ubc9 in the SUMO–RanGAP1–Ubc9– RanBP2 complex. RanGAP1 was then moved by 1.9 Å to stop all interpenetration with UbcH5A. In this position, the side chain of K524 in RanGAP1 is poised to attack the activated thioester bond in the UbcH5A–Ub thioester. Proteins are coloured as in a. e, Detailed view of the thioester bond with the incoming substrate lysine. In our model of a hypothetical substrate (RanGAP1) approaching the activated thioester bond, D117 in UbcH5A positions and activates the incoming substrate lysine for a nucleophilic attack. The resulting tetrahedral intermediate would be stabilized by a hydrogen bond with the side chain of N77. An intricate network of hydrogen bonding interactions (grey dashes) between the C-terminal tail of ubiquitin and the active site groove of UbcH5A lock the thioester bond in the activated configuration.
UbcH5AUbiquitin
RanGAP1
d
e
G76G75R74
L73C85
D117N114
D87
L86
K524
N77

W W W. N A T U R E . C O M / N A T U R E | 2 1
SUPPLEMENTARY INFORMATION RESEARCH
SUPPLEMENTARY REFERENCES 42 Kosoy, A., Calonge, T. M., Outwin, E. A. & O'Connell, M. J. Fission yeast Rnf4 homologs
are required for DNA repair. J. Biol. Chem. 282, 20388–20394 (2007).
43 Sun, H., Leverson, J. D. & Hunter, T. Conserved function of RNF4 family proteins in
eukaryotes: targeting a ubiquitin ligase to SUMOylated proteins. EMBO J. 26, 4102–4112
(2007).
44 Huang, K. Y., Amodeo, G. A., Tong, L. & McDermott, A. The structure of human ubiquitin
in 2-methyl-2,4-pentanediol: a new conformational switch. Protein Sci. 20, 630-639 (2011).










![A colorimetric method for α-glucosidase activity assay … · reversibly bind diols with high affinity to form cyclic esters [23]. Herein, based on these findings, a ...](https://static.fdocument.org/doc/165x107/5b696db67f8b9a24488e21b4/a-colorimetric-method-for-glucosidase-activity-assay-reversibly-bind-diols.jpg)







