
N-terminal acetylation and other functions of Nα-acetyltransferases
8
Biol. Chem., Vol. 393, pp. 291–298, April 2012 • Copyright © by Walter de Gruyter • Berlin • Boston. DOI 10.1515/hsz-2011-0228 Minireview N-terminal acetylation and other functions of N α -acetyltransferases Jolien Hollebeke 1,2 , Petra Van Damme 1,2 and Kris Gevaert 1,2, * 1 Department of Medical Protein Research, Flanders Interuniversity Institute for Biotechnology (VIB), Ghent University, A. Baertsoenkaai 3, B-9000 Ghent, Belgium 2 Department of Biochemistry, Ghent University, B-9000 Ghent, Belgium * Corresponding author e-mail: [email protected] Abstract Protein N-terminal acetylation by N α -acetyltransferases (NATs) is an omnipresent protein modification that affects a large number of proteins. The exact biological role of N-terminal acetylation has, however, remained enigmatic for the overall majority of affected proteins, and only for a rather small number of proteins, N-terminal acetylation was linked to various protein features including stability, local- ization, and interactions. This minireview tries to summarize the recent progress made in understanding the functionality of N-terminal protein acetylation and also focuses on nonca- nonical functions of the NATs subunits. Keywords: α-N-acetylation; N α -acetyltransferases; N-terminal acetylation of proteins. General introduction on N-terminal acetylation Cotranslational modification of the N-terminus of eukaryotic proteins occurs very frequently. For instance, about 85% of all human proteins are acetylated at their N α -terminus, whereas in yeast, this number totals to about 68% (Van Damme et al., 2011c). Besides eukaryotic proteins, it has been shown that several proteins from prokaryotes and archaebacteria are also N-terminally acetylated. However, this is usually posttransla- tional as is the case for N-terminal acetylation of proteins in chloroplasts (Bradshaw et al., 1998; Polevoda and Sherman, 2003; Zybailov et al., 2008). Different enzyme complexes are responsible for the acetylation of nascent protein N-termini, a modification which thus far was found to be irreversible. Each of these enzyme complexes has its own substrate speci- ficity; however, substrate redundancy among these enzymes has been shown. In total, six different N α -acetyltransferases (NATs) have been reported in humans (Table 1 ). NatA recognizes and acetylates N-termini from which the initiator methionine was removed by methionine aminopeptidases and then start with small amino acids such as Ser, Thr, Ala, Val, or Gly. When the initiator methionine is followed by Asp, Glu, or Gln, it can be acetylated by NatB. NatC acetylates initiator methionines followed by hydrophobic amino acids (Leu, Ile, Tyr, or Phe), NatD acetylates the N-termini of the histones H2A and H4, and NatE acetylates proteins where the initiator methionine is followed by Leu, Ala, Lys, or Met (Polevoda et al., 2009; Hole et al., 2011). Finally, the recently discov- ered NatF, which is specific for higher eukaryotes, acetylates N-termini mainly starting with Met followed by Lys, Leu, Ile, Trp, or Phe (Van Damme et al., 2011c). Over the years, different techniques have been used to study protein N-terminal acetylation. Early on, protein 2D-gel electrophoresis was used (Polevoda et al., 1999) to identify NAT substrates as the addition of an acetyl group on protein N-termini lowers the isoelectric point of the affected protein and thus alters electrophoretic mobility of that protein in the gel. More recently, gel-free proteomics techniques were devel- oped that directly focus on isolated N-terminal peptides rather than on proteins. N α -acetylated N-termini can be isolated using so-called positive or negative selection methods. Strong cation exchange (SCX) at low pH is an example of a posi- tive selection method in which N α -acetylated N-termini are enriched because of their reduced positive charge compared to internal peptides that hold a primary α-amine. Negative selec- tion methods deplete non-N α -acetylated peptides from N α - acetylated ones. Amine-reactive resins, affinity-tag directed beads, and N-terminal combined fractional diagonal chroma- tography (COFRADIC) are all examples of negative selection procedures. A recent overview of such proteomics technolo- gies was presented by Van Damme et al. (see Van Damme et al., 2011a, and references therein). In general, all these tech- niques helped to identify the specificity of the different NATs as well as their in vivo substrate repertoires. As such, much is currently known about the specificity of cotranslational NAT-mediated N-terminal acetylation, and it has been spec- ulated that this modification might affect several cellular processes. However, for the multitude of proteins targeted by N-terminal acetylation, the real physiological function of this modification remains to be determined. Here we present a general view on what is currently known about the biological implications for N-terminal acetylation, together with a summary of the functions of NAT subunits (in)dependent of their NAT activity, which might help to elucidate some of the observed phenotypes when knocking down/out (a) specific NAT(s). Brought to you by | Universitaetsbibliothek der LMU Muenchen Authenticated | 129.187.254.47 Download Date | 6/18/14 11:01 PM
Transcript of N-terminal acetylation and other functions of Nα-acetyltransferases

Biol. Chem., Vol. 393, pp. 291–298, April 2012 • Copyright © by Walter de Gruyter • Berlin • Boston. DOI 10.1515/hsz-2011-0228
Minireview
N-terminal acetylation and other functions of N α -acetyltransferases
Jolien Hollebeke 1,2 , Petra Van Damme 1,2 and Kris Gevaert 1,2, *
1 Department of Medical Protein Research , Flanders Interuniversity Institute for Biotechnology (VIB), Ghent University, A. Baertsoenkaai 3, B-9000 Ghent , Belgium 2 Department of Biochemistry , Ghent University, B-9000 Ghent , Belgium
* Corresponding authore-mail: [email protected]
Abstract
Protein N-terminal acetylation by N α -acetyltransferases (NATs) is an omnipresent protein modifi cation that affects a large number of proteins. The exact biological role of N-terminal acetylation has, however, remained enigmatic for the overall majority of affected proteins, and only for a rather small number of proteins, N-terminal acetylation was linked to various protein features including stability, local-ization, and interactions. This minireview tries to summarize the recent progress made in understanding the functionality of N-terminal protein acetylation and also focuses on nonca-nonical functions of the NATs subunits.
Keywords: α -N-acetylation; N α -acetyltransferases; N-terminal acetylation of proteins.
General introduction on N-terminal acetylation
Cotranslational modifi cation of the N-terminus of eukaryotic proteins occurs very frequently. For instance, about 85 % of all human proteins are acetylated at their N α -terminus, whereas in yeast, this number totals to about 68 % (Van Damme et al. , 2011c ). Besides eukaryotic proteins, it has been shown that several proteins from prokaryotes and archaebacteria are also N-terminally acetylated. However, this is usually posttransla-tional as is the case for N-terminal acetylation of proteins in chloroplasts (Bradshaw et al. , 1998 ; Polevoda and Sherman , 2003 ; Zybailov et al. , 2008 ). Different enzyme complexes are responsible for the acetylation of nascent protein N-termini, a modifi cation which thus far was found to be irreversible. Each of these enzyme complexes has its own substrate speci-fi city; however, substrate redundancy among these enzymes has been shown. In total, six different N α -acetyltransferases (NATs) have been reported in humans (Table 1 ). NatA
recognizes and acetylates N-termini from which the initiator methionine was removed by methionine aminopeptidases and then start with small amino acids such as Ser, Thr, Ala, Val, or Gly. When the initiator methionine is followed by Asp, Glu, or Gln, it can be acetylated by NatB. NatC acetylates initiator methionines followed by hydrophobic amino acids (Leu, Ile, Tyr, or Phe), NatD acetylates the N-termini of the histones H2A and H4, and NatE acetylates proteins where the initiator methionine is followed by Leu, Ala, Lys, or Met (Polevoda et al. , 2009 ; Hole et al. , 2011 ). Finally, the recently discov-ered NatF, which is specifi c for higher eukaryotes, acetylates N-termini mainly starting with Met followed by Lys, Leu, Ile, Trp, or Phe (Van Damme et al. , 2011c ).
Over the years, different techniques have been used to study protein N-terminal acetylation. Early on, protein 2D-gel electrophoresis was used (Polevoda et al. , 1999 ) to identify NAT substrates as the addition of an acetyl group on protein N-termini lowers the isoelectric point of the affected protein and thus alters electrophoretic mobility of that protein in the gel. More recently, gel-free proteomics techniques were devel-oped that directly focus on isolated N-terminal peptides rather than on proteins. N α -acetylated N-termini can be isolated using so-called positive or negative selection methods. Strong cation exchange (SCX) at low pH is an example of a posi-tive selection method in which Nα-acetylated N-termini are enriched because of their reduced positive charge compared to internal peptides that hold a primary α -amine. Negative selec-tion methods deplete non-N α -acetylated peptides from N α -acetylated ones. Amine-reactive resins, affi nity-tag directed beads, and N-terminal combined fractional diagonal chroma-tography (COFRADIC) are all examples of negative selection procedures. A recent overview of such proteomics technolo-gies was presented by Van Damme et al. (see Van Damme et al. , 2011a , and references therein). In general, all these tech-niques helped to identify the specifi city of the different NATs as well as their in vivo substrate repertoires. As such, much is currently known about the specifi city of cotranslational NAT-mediated N-terminal acetylation, and it has been spec-ulated that this modifi cation might affect several cellular processes. However, for the multitude of proteins targeted by N-terminal acetylation, the real physiological function of this modifi cation remains to be determined.
Here we present a general view on what is currently known about the biological implications for N-terminal acetylation, together with a summary of the functions of NAT subunits (in)dependent of their NAT activity, which might help to elucidate some of the observed phenotypes when knocking down/out (a) specifi c NAT(s).
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

292 J. Hollebeke et al.
N-terminal acetylation can affect protein
stability
The N-end rule links the half life of a protein to its N-terminal amino acid, with eight amino acids being primary destabiliz-ing (Ile, Leu, Phe, Tyr, Trp, Arg, His, and Lys) and a further four (Asn, Gln, Asp, and Glu) that need to be modifi ed (by deamidation or arginylation) before they can be recognized by ubiquitin ligases. Asp and Glu need to be arginylated before they can be recognized by ubiquitin ligases, making them sec-ondary destabilizing residues, while Asn and Gln are tertiary destabilizing residues because they fi rst need to be converted to the secondary destabilizing residues Asp and Glu, respec-tively, by deamidation prior to arginylation.
For many years, it was believed that N-terminal acetyla-tion protects proteins from being degraded (Persson et al. , 1985 ), likely because N α -acetylated N-termini were supposed to block (N-terminal) ubiquitination and subsequent protein degradation (Hershko et al. , 1984 ). An example of a protein following this rule is the tumor suppressor protein p19 ARF (Kuo et al. , 2004 ) (Figure 1 A). However, many studies con-tradict each other, for instance, by showing that N α -acetylated proteins are degraded at the same rate as proteins with a non-blocked N-terminus (Brown , 1979 ), whereas for the α -mel-anocyte-stimulating hormone, it was shown that N-terminal acetylation increases its stability (Guo et al. , 2004 ), and in the algae species Chlamydomonas reinhardtii , it was found that unstable chloroplast proteins are rarely N-terminally acety-lated (Bienvenut et al. , 2011 ).
The group of Varshavsky showed that for some yeast proteins, N-terminal acetylation creates specifi c N-terminal proteins structures, N α -degrons, which are recognized by the ubiquitin ligase Doa10, marking these proteins with ubiquitin, which fi nally leads to their degradation (Hwang et al. , 2010 ) (Figure 1 B). In fact, this was shown to be the case for eight N-terminally acetylated yeast proteins. Given this link between N-terminal acetylation and ubiquitination, it is now believed that such a protein degradation targeting strategy might serve as a sort of quality control mechanism; when a protein is being translated, N α -acetylated and rapidly
folded, interacts with a chaperone, or becomes part of a com-plex, its recognition by Doa10 and its further degradation are prevented. Hence, this also implies that several other factors, besides the presence or absence of an N α -acetyl group, deter-mine the fate of a protein.
Protein localization can be infl uenced by
N-terminal acetylation
Besides infl uencing protein stability, evidence is accumulat-ing that N-terminal acetylation can steer the localization of proteins. Arl3p and Arl8p are two Arf-like GTPases, which differ from other ADP ribosylation factor (Arf) family mem-bers and Arf-like GTPases in that they lack a Gly at posi-tion 2 required for N-myristoylation and membrane targeting (Behnia et al. , 2004 ; Hofmann and Munro , 2006 ; Starheim et al. , 2009 ). Instead, Arl3p and Arl8p contain a Phe (yeast Arl3p), Tyr (human Arl3p), Leu (human Arl8a), or Ile (human Arl8b) at this position, making all of these N-termini substrates of NatC. In fact, Arl3p requires its N α -acetyl group for its targeting to the Golgi membrane by its interaction with the Golgi membrane-residing protein Sys1p (Figure 2 A). When Arl3p ’ s Phe or Tyr is replaced by an Ala (representa-tive of a NatA-type substrate) or Glu (NatB substrate), Arl3p no longer localized to the Golgi, hinting to the fact that Arl3p needs its natural N-terminal structure for proper localization. N-terminal acetylation of Arl8p was also shown to facilitate membrane targeting to lysosomes, but this did not determine organellar specifi city.
Proteins that are destined for the secretory pathway typi-cally contain an N-terminal signal sequence, which targets them to the endoplasmatic reticulum (ER). Two different pathways lead to protein translocation via the ER: a post-translational, Sec62-dependent pathway and a cotransla-tional, signal recognition particle (SRP)-dependent pathway (Figure 2 B and C). Forte et al. recently showed that there is a marked difference in amino acid usage at a protein ’ s second position when comparing cytosolic proteins and proteins containing signal sequences (Forte et al. , 2011 ); cytosolic proteins tend to hold small or acidic residues, while proteins containing signal sequences preferably hold bulky or basic residues. Furthermore, this amino acid bias is conserved during evolution. Forte and coworkers further demonstrated that N-terminal acetylation prevents translo-cation to the ER by creating protein mutants in which an additional residue between the initiator methionine and the natural residue at the protein ’ s second position was added to allow for N-terminal acetylation. For regular cotransla-tional ER translocation, protein N-terminal acetylation is prevented because SRP and NATs are thought to compete for binding to the ribosome. It is currently, however, not fully understood how exactly N-terminal acetylation blocks protein translocation, but the difference in charge distribu-tion at the N-terminus appears not to be the major determi-nant. Instead, the authors reason that N-terminal acetylation rather marks proteins to be folded and thereby to remain in the cytoplasm.
Table 1 The composition and substrate specifi city of the N α -acetyltransferases.
Nat Subunits Substrates
NatA Naa10p (Ard1p) Ser, Ala, Gly, Thr, Val, CysNaa15p (Nat1p)
NatB Naa20p (Nat3p) Met-Glu-, Met-Asp-, Met-Asn-, Met-Gln-Naa25p (Mdm20p)
NatC Naa30p (Mak3p) Met-Leu-, Met-Ile-, Met-Trp-, Met-Phe-Naa35p (Mak10p)
Naa38p (Mak31p)NatD Naa40p (Nat4p) Ser-Gly-Gly-, Ser-Gly-Arg-NatE Naa50p (Nat5p) Met-Leu-, Met-Ala-, Met-Lys-,
Met-Met-NatF Naa60p Met-Lys-, Met-Leu-, Met-Ile-,
Met-Trp-, Met-Phe-
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

Functions of N α -acetyltransferases 293
Metabolism and apoptosis linked to N-terminal
acetylation
Protein knockdown experiments have linked protein N-terminal acetylation to cell cycle regulation and apoptosis.
Arnesen and coworkers showed that knockdown of the NatA or the NatC complex led to the induction of p53-dependent apoptosis and argued that certain anti-apoptotic proteins were less or no longer functional because of reduced protein N-terminal acetylation (Gromyko et al. , 2010 ). In contrast,
Figure 1 N-terminal acetylation may signal protein degradation. N-terminal acetylation of proteins can have different effects on protein stability. (A) Proteins can be N-terminally ubiquitinated and subse-quently degraded. N-terminal acetylation can prevent N-terminal ubiquitination and thus increase protein stability. (B) The initiator methionine of a protein can be N α -acetylated, but when the second amino acid (green) is small, this allows the removal of the initiator methionine by a methionine aminopeptidase and subsequent N-terminal acetylation. N-terminal acetylation may promote degradation as a secondary destabiliz-ing residue gets recognized by the ubiquitin ligase Doa10 leading to proteasomal degradation of the ubiquitinated NAT substrate ( a ), or this N α -acetylated protein can be folded and remain stable in the cell ( b ).
Figure 2 N-terminal acetylation can steer protein localization. (A) In the case of Arl3p, N-terminal acetylation is needed to direct this protein to the Golgi protein Sys1p. (B) Proteins with a bulky or acidic residue (red) at the second position in their signal sequence are not N α -acetylated. When fully translated, they interact with chaperones and are directed toward the Sec61 channel of the ER membrane. (C) The cotranslational pathway of protein translocation. The SRP protein binds to the ribosome on the same site as the NATs and thereby prevents N-terminal acetylation. SRP recognizes the signal sequence and targets the nascent polypeptide to the Sec61 channel via the SRP receptor.
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

294 J. Hollebeke et al.
Yi et al. ( 2011 ) showed that N-terminal acetylation induces apoptosis. They used subtiligase to enzymatically biotinylate nonblocked protein N-termini with NatA or NatB specifi c-ity and further quantifi ed changes in N-terminal acetylation. In this way, it was shown that the N-terminus of caspase-2 gets acetylated by NatA and that this modifi cation is neces-sary for its interaction with the adaptor protein RIP associ-ated Ich-1/Ced-3 homologous protein with a death domain (RAIDD), resulting in activation of caspase-2 and ultimately to induction of apoptosis. They further reasoned that the cel-lular metabolic status could have an effect on N-terminal acetylation given that acetyl-coenzyme A (CoA) is the major cofactor of the NATs. This indeed seemed to be the case as they could demonstrate that changing the expression of B-cell lymphoma – extra large (Bcl-xL), an antiapoptotic Bcl-2 family member known to affect cellular metabolism, caused a change in the degree of N-terminal acetylation of certain NatA or NatB substrates (caspases 2, 3, and 9). The proposed mechanism would thus be that Bcl-xL lowers the concentration of acetyl-CoA, thereby lowering N-terminal acetylation of particular caspases and concomitantly inhibit-ing apoptosis.
These opposing effects on apoptosis might be explained by the fact that NATs and their subunits (see further) can have different effects on (different) proteins in various pathways. This means that even a slight difference in the knockdown effi ciency of a protein might lead to different phenotypes as is the case in the examples discussed above.
The impact of N-terminal acetylation on protein
synthesis
The majority of cytosolic proteins of higher eukaryotes are acetylated at their N-terminus. Ribosomal proteins, for instance, are also known to be N α -acetylated, but the impact of this modifi cation on their function is unclear. Kamita et al. ( 2011 ) used yeast NAT-deletion strains and 2D-gel electrophoresis to study N-terminal acetylation of ribo-somal proteins. Ribosomes function in protein synthesis; hence, it stands to reason that N-terminal acetylation of ribosomal proteins might have an effect on protein syn-thesis. Kamita and coworkers indeed showed a decrease of 27 % and 23 % in the protein synthesis rate of NatA and NatB deletion strains, respectively. The underlying cause is not fully understood; however, in the NatB deletion strain, a defect in ribosome assembly was noticed, whereas in the NatA deletion strain, a reduction of translation fi delity was observed.
Protein-protein interactions steered by
N-terminal acetylation
Various proteins need an acetylated N-terminus in order to interact with other proteins. As discussed above, this is the case for caspase-2 and its adaptor protein RAIDD (Yi et al. , 2011 ), for Arl3p and its interaction with Sys1p (Behnia et
al. , 2004 ), and for several ribosomal subunits (Kamita et al. , 2011 ).
Another example where N-terminal acetylation is involved in protein-protein interaction is the interaction between actin, myosin, and tropomyosin. It has been shown that the NatB-mediated N-terminal acetylation of tropomyosin is necessary to strengthen the end-to-end contacts between tropomyo-sin dimers (Coulton et al. , 2010 ). Interestingly, N-terminal acetylation of tropomyosin is dispensable for actin binding, but increases the integrity of actin-tropomyosin fi laments, thereby stabilizing the position of tropomyosin polymer on fi lamentous actin.
Actin itself is also acetylated at its N-terminus after its N α -acetylated initiator methionine (of class I actins like β -actin) is removed. For class II actins such as α -actin, the initiator methionine is removed followed by the removal of the N α -acetylated cysteine at the second position. Subsequent posttranslational N-terminal acetylation in vivo is likely performed in part by Naa10 and/or NatA (Van Damme et al. , 2011b ), and this acetylation was found to strengthen actin ’ s interaction with myosin (Abe et al. , 2000 ).
Alternative functions of NAT subunits
NATs consist of two or more subunits. Reducing the expres-sion of some of these subunits leads to different phenotypes. Different subunits of a given NAT complex sometimes also possess differential localization patterns. Among others, such observations may contribute to the fact that NAT subunits may hold functions independent of their actual acetyltrans-ferase function. Some recent examples of this are described in the following sections and illustrated in Figure 3 .
Noncanonical acetylation by Naa10
Naa10 or arrest defective-1 (Ard1) is the best-studied NAT subunit as well as the fi rst identifi ed NAT in yeast (Lee et al. , 1988 ; Mullen et al. , 1989 ). Of particular note is that just recently, a Ser-to-Pro mutation at position 37 of Naa10 was linked to a lethal human X-linked disorder, making it the fi rst mutation in the NAT machineries linked to a human dis-order. In fact, this mutation occurs in an evolutionary con-served domain and has been shown to have an effect on the N-terminal acetylation activity of Naa10 (Rope et al. , 2011 ). What makes Naa10 further interesting is that, besides pos-sessing N-terminal acetylation activity, Naa10 has also been reported to N ε -acetylate substrates, a feature that points to an alternative function of Naa10 apart from its canonical N-terminal acetylation functionality.
Different variants of Naa10 are found in mammals; e.g., in mouse, three splice variants have been described. One of these was shown to N ε -acetylate HIF-1α leading to its deg-radation (Jeong et al. , 2002 ). In human cells, however, data on Naa10-dependent N ε -acetylation of HIF-1 α are confl ict-ing and rather point to the fact that HIF-1 α is probably not N ε -acetylated by Naa10, although both proteins
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM
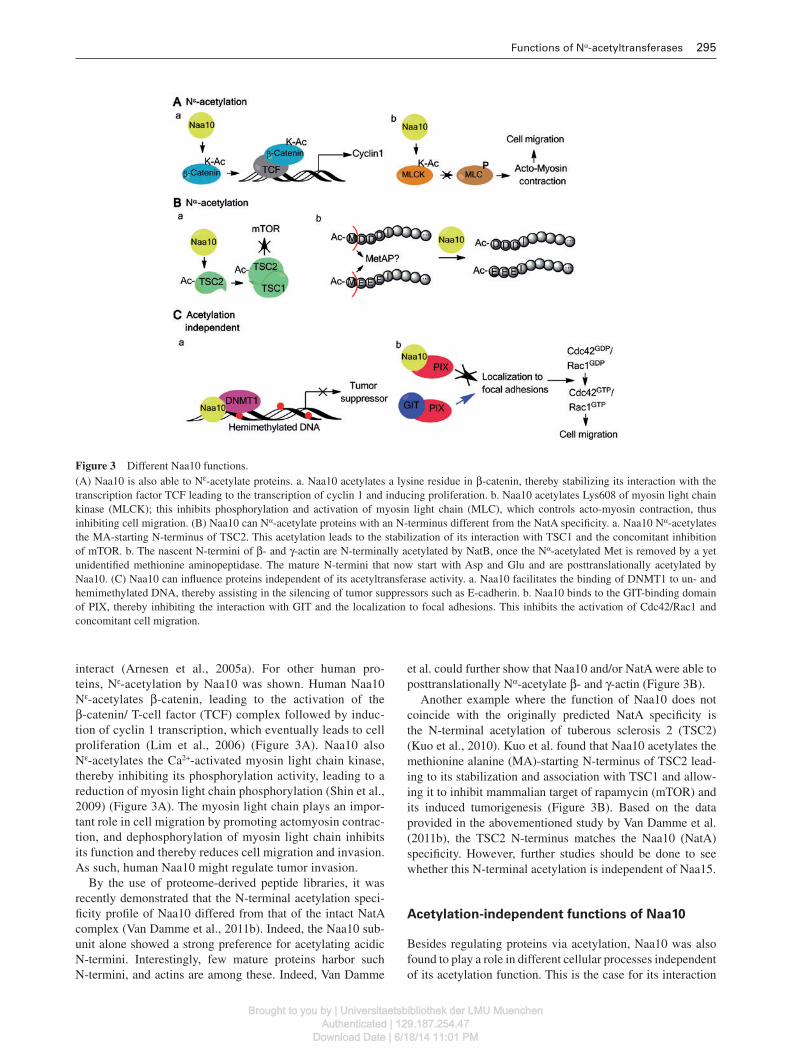
Functions of N α -acetyltransferases 295
Figure 3 Different Naa10 functions. (A) Naa10 is also able to N ε -acetylate proteins. a . Naa10 acetylates a lysine residue in β -catenin, thereby stabilizing its interaction with the transcription factor TCF leading to the transcription of cyclin 1 and inducing proliferation. b . Naa10 acetylates Lys608 of myosin light chain kinase (MLCK); this inhibits phosphorylation and activation of myosin light chain (MLC), which controls acto-myosin contraction, thus inhibiting cell migration. (B) Naa10 can N α -acetylate proteins with an N-terminus different from the NatA specifi city. a. Naa10 N α -acetylates the MA-starting N-terminus of TSC2. This acetylation leads to the stabilization of its interaction with TSC1 and the concomitant inhibition of mTOR. b. The nascent N-termini of β - and γ -actin are N-terminally acetylated by NatB, once the N α -acetylated Met is removed by a yet unidentifi ed methionine aminopeptidase. The mature N-termini that now start with Asp and Glu and are posttranslationally acetylated by Naa10. (C) Naa10 can infl uence proteins independent of its acetyltransferase activity. a . Naa10 facilitates the binding of DNMT1 to un- and hemimethylated DNA, thereby assisting in the silencing of tumor suppressors such as E-cadherin. b . Naa10 binds to the GIT-binding domain of PIX, thereby inhibiting the interaction with GIT and the localization to focal adhesions. This inhibits the activation of Cdc42/Rac1 and concomitant cell migration.
interact (Arnesen et al. , 2005a ). For other human pro-teins, N ε -acetylation by Naa10 was shown. Human Naa10 N ε -acetylates β -catenin, leading to the activation of the β -catenin/ T-cell factor (TCF) complex followed by induc-tion of cyclin 1 transcription, which eventually leads to cell proliferation (Lim et al. , 2006 ) (Figure 3 A). Naa10 also N ε -acetylates the Ca 2 + -activated myosin light chain kinase, thereby inhibiting its phosphorylation activity, leading to a reduction of myosin light chain phosphorylation (Shin et al. , 2009 ) (Figure 3 A). The myosin light chain plays an impor-tant role in cell migration by promoting actomyosin contrac-tion, and dephosphorylation of myosin light chain inhibits its function and thereby reduces cell migration and invasion. As such, human Naa10 might regulate tumor invasion.
By the use of proteome-derived peptide libraries, it was recently demonstrated that the N-terminal acetylation speci-fi city profi le of Naa10 differed from that of the intact NatA complex (Van Damme et al. , 2011b ). Indeed, the Naa10 sub-unit alone showed a strong preference for acetylating acidic N-termini. Interestingly, few mature proteins harbor such N-termini, and actins are among these. Indeed, Van Damme
et al. could further show that Naa10 and/or NatA were able to posttranslationally N α -acetylate β - and γ -actin (Figure 3 B).
Another example where the function of Naa10 does not coincide with the originally predicted NatA specifi city is the N-terminal acetylation of tuberous sclerosis 2 (TSC2) (Kuo et al. , 2010 ). Kuo et al. found that Naa10 acetylates the methionine alanine (MA)-starting N-terminus of TSC2 lead-ing to its stabilization and association with TSC1 and allow-ing it to inhibit mammalian target of rapamycin (mTOR) and its induced tumorigenesis (Figure 3 B). Based on the data provided in the abovementioned study by Van Damme et al. (2011b) , the TSC2 N-terminus matches the Naa10 (NatA) specifi city. However, further studies should be done to see whether this N-terminal acetylation is independent of Naa15.
Acetylation-independent functions of Naa10
Besides regulating proteins via acetylation, Naa10 was also found to play a role in different cellular processes independent of its acetylation function. This is the case for its interaction
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

296 J. Hollebeke et al.
with DNA-methyltransferase 1 (DNMT1) (Figure 3 C). Lee and coworkers reasoned that Naa10 could be an oncoprotein and proved this in different types of lung cancer (Lee et al. , 2010 ). To unravel the mechanism by which Naa10 contrib-utes to tumorigenesis, a yeast 2-hybrid screen was conducted to identify interaction partners, and one of these partners was DNMT1. Although they suggested that Naa10 N α -acetylated DNMT1, acetylation was not found to play a role in the inter-action between both proteins. Furthermore, it must be noted that DNMT1 holds a proline at the second position, which was shown to prevent N-terminal acetylation (Goetze et al. , 2009 ). Lee et al. demonstrated that Naa10 is able to bind unmethylated and hemimethylated DNA, and because of this ability, Naa10 strengthens the association between DNMT1 and its substrate DNA (in particular for de novo methylation). In this respect, it was shown that Naa10 regulates methyla-tion of the promoter region of E-cadherin, thereby reducing it expression. This, in turn, explains why Naa10 is considered as an oncoprotein as it is involved in the silencing of tumor suppressor genes (such as E-cadherin and the large tumor suppressor, homolog 2 protein).
As discussed above, Naa10 is able to inhibit cell migration by N ε -acetylation of the myosin light chain kinase (Shin et al. , 2009 ). Hua et al. ( 2011 ) demonstrate that Naa10 is able to inhibit cell migration and metastasis via an acetylation-inde-pendent manner. In their model, Naa10 binds to the G-protein coupled receptor kinase-interacting protein (GIT) binding domain of the p21-activated kinase (PAK) interacting exchange factor (PIX) via its acetyltransferase domain, preventing the binding of GIT to PIX and consequently inhibiting its local-ization to focal adhesions where it normally activates Rac1/Cdc42 through its guanine exchange factor (GEF) activity (Figure 3 C). Rac1 and Cdc42 are two Rho GTPases that con-trol actin dynamics and play important roles in cell migration. By preventing their activation, cell motility is consequently decreased, thus turning Naa10 into a tumor suppressor.
Possible alternative functions for Naa15
Not much is known about alternative functions of Naa15 and especially not whether these would be independent of Naa10. It has, for instance, been shown that Naa15 is overexpressed in thyroid neoplasm (Arnesen et al. , 2005b ). Interestingly, the levels of its expected partner in the NatA complex, Naa10, only correlated to those of Naa15 in tumors in which Naa15 expres-sion was reduced compared to non-neoplastic tissues. Other papers indicated that Naa15 may play a role in the cytotoxic activity of 5-fl uorouracil as upon its administration, expres-sion of Naa15 is downregulated, while Naa10 and Naa50 expression is upregulated hinting toward a function of Naa15 independent from the NatA complex (Takubo et al. , 2009 ).
Noncanonical functions of NatB and NatC
For NatB, it was shown that Naa25 is localized exclusively to the cytoplasm, while Naa20 displays a dual localization pat-tern, in the cytoplasm and the nucleus, pointing to a possible
alternative function of Naa20 (Starheim et al. , 2008 ). Another indication of an alternative function is the observation of dif-ferent phenotypes when the different subunits are knocked down. Knocking down Naa20 leads to an increase in p21 lev-els, while knockdown of Naa25 decreases p21 levels. Further research is necessary to pinpoint to the exact underlying mechanism causing this difference.
Multiple lines of evidence also hint to alternative functions for the individual subunits of NatC (Naa30, Naa35, and Naa38). Both Naa30 (Mak3) and Naa35 (Mak10) localize exclusively to the cytoplasm, while Naa38 (Mak31) is also present in the nucleus, possibly because of its small size that enables it to pass through the nuclear pore complex (Starheim et al. , 2009 ). Furthermore, Naa38 belongs to the Sm and Sm-like family of proteins, which associate with RNA and are often involved in RNA processing, hinting to an alike alternative function of Naa38. In Arabidopsis thaliana , it was also shown in that dele-tion of AtNaa35 did not display the same phenotype compared to the AtNaa30 knockout, again pointing to possible additional functions of Naa30 (Pesaresi et al. , 2003 ).
Conclusions
Much is still unknown and unclear about the functional impli-cation of protein N-terminal acetylation and the functions of the different NAT (subunit)s in general. Nevertheless, the NATs and their evoked modifi cations might govern many dif-ferent cellular processes. This hypothesis is supported by the fact that the NAT subunits themselves may hold individual functions that are not directly related to their NAT activity. Many of such reported functions actually depend on the cell type and conditions studied, meaning that results obtained in one cell type or tissue cannot be generalized to all other cell types. For example, in the case of Naa10 (NatA), Gromyko et al . showed that knockdown of NatA in HeLa cells led to the induction of p53-dependent apoptosis, while Yi and cowork-ers demonstrated that knockdown of NatA in HeLa cells inhibited apoptosis. In this respect, it is also not clear whether Naa10 acts as an oncoprotein or rather as a tumor suppressor, as both functions were put forward when making use of differ-ent model systems. Because Naa10 is involved in a multitude of pathways, it is maybe not that surprising that a knockdown of NatA gives such different phenotypes. This is the reason why, to our opinion, it is important to perform research on the functions of the NAT subunits individually. In this way, the phenotypes that are not the direct consequence of the loss of N-terminal acetylation due to the loss of a NAT can be segregated from those that are the consequence of the loss of the function of a NAT subunit independent of N-terminal acetylation. Future research should enable a further under-standing of the precise function of this common, yet incom-pletely understood, protein modifi cation.
Acknowledgements
J.H. is supported by a PhD grant of the Institute for the Promotion of Innovation through Science and Technology in Flanders
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

Functions of N α -acetyltransferases 297
(IWT-Vlaanderen). P.V.D. is a Postdoctoral Fellow of the Research Foundation-Flanders (FWO-Vlaanderen). The authors further acknowledge support of research grants from the Fund for Scientifi c Research-Flanders (Belgium) (project number G.0042.07 and G.0440.10) and the Inter University Attraction Poles (IUAP06).
References
Abe, A., Saeki, K., Yasunaga, T., and Wakabayashi, T. (2000). Acetylation at the N-terminus of actin strengthens weak inter-action between actin and myosin. Biochem. Biophys. Res. Commun. 268 , 14 – 19.
Arnesen, T., Gromyko, D., Horvli, D., Fluge, O., Lillehaug, O., and Varhaug, J.E. (2005a). Expression of N-acetyl transferase human and human Arrest defective 1 proteins in thyroid neoplasms. Thyroid 15 , 1131 – 1136.
Arnesen, T., Kong, X., Evjenth, R., Gromyko, D., Varhaug, J.E., Lin, Z., Sang, N., Caro, J., and Lillehaug, J.R. (2005b). Interaction between HIF-1 alpha (ODD) and hARD1 does not induce acetylation and destabilization of HIF-1 alpha. FEBS Lett. 579 , 6428 – 6432.
Behnia, R., Panic, B., Whyte, J.R., and Munro, S. (2004). Targeting of the Arf-like GTPase Arl3p to the Golgi requires N-terminal acetylation and the membrane protein Sys1p. Nat. Cell Biol. 6 , 405 – 413.
Bienvenut, W.V., Espagne, C., Martinez, A., Majeran, W., Valot, B., Zivy, M., Vallon, O., Adam, Z., Meinnel T., and Giglione, C. (2011). Dynamics of post-translational modifi cations and protein stability in the stroma of Chlamydomonas reinhardtii chloro-plasts. Proteomics 11 , 1734 – 1750.
Bradshaw, R.A., Brickey, W.W., and Walker, K.W. (1998). N-terminal processing: the methionine aminopeptidase and N α -acetyl trans-ferase families. Trends Biochem. Sci. 23 , 263 – 267.
Brown, J.L. (1979). A comparison of the turnover of alpha-N-acetylated and nonacetylated mouse L-cell proteins. J. Biol. Chem. 254 , 1447 – 1449.
Coulton, A.T., East, D.A., Galinska-Rakoczy, A., Lehman, W., and Mulvihill, D.P. (2010). The recruitment of acetylated and unacetylated tropomyosin to distinct actin polymers permits the discrete regulation of specifi c myosins in fi ssion yeast. J. Cell Sci. 123 , 3235 – 3243.
Forte, G.M., Pool, M.R., and Stirling, C.J. (2011). N-terminal acety-lation inhibits protein targeting to the endoplasmic reticulum. PLoS Biol. 9 , e1001073.
Goetze, S., Qeli, E., Mosimann, C., Staes, A., Gerrits, B., Roschitzki, B., Mohanty, S., Niederer, E.M., Laczko, E., Timmerman, E., et al. (2009). Identifi cation and functional characterization of N-terminally acetylated proteins in Drosophila melanogaster . PLoS Biol. 7 , e1000236.
Gromyko, D., Arnesen, T., Ryningen, A., Varhaug, J.E., and Lillehaug, J.R. (2010). Depletion of the human Nalpha-terminal acetyltransferase A induces p53-dependent apoptosis and p53-independent growth inhibition. Int. J. Cancer 127 , 2777 – 2789.
Guo, L., M ü nzberg, H., Stuart, R.C., Nillni, E.A., and Bj ø rb æ k, C. (2004). N-acetylation of hypothalamic α -melanocyte-stimulating hormone and regulation by leptin. Proc. Natl. Acad. Sci. USA 101 , 11797 – 11802.
Hershko, A., Heller, H., Eytan, E., Kaklij, G., and Rose, I.A. (1984). Role of the alpha-amino group of protein in ubiquitin-mediated protein breakdown. Proc. Natl. Acad. Sci. USA 81 , 7021 – 7025.
Hofmann, I. and Munro, S. (2006). An N-terminally acetylated Arf-like GTPase is localised to lysosomes and affects their motility. J. Cell Sci. 119 , 1494 – 1503.
Hole, K., Van Damme, P., Dalva, M., Aksnes, H., Glomnes, N., Varhaug, J.E., Lillehaug, J.R., Gevaert, K., and Arnesen, T. (2011). The human N-alpha-acetyltransferase 40 (hNaa40p/hNatD) is conserved from yeast and N-terminally acetylates his-tones H2A and H4. PLoS One 6 , e24713.
Hua, K.-T., Tan, C.-T., Johansson, G., Lee, J.-M., Yang, P.-W., Lu, H.-Y., Chen, C.-K., Su, J.-L., Chen, P. B., Wu, Y.-L., et al. (2011). N-alpha-acetyltransferase 10 protein suppresses cancer cell metastasis by binding PIX proteins and inhibiting Cdc42/Rac1 activity. Cancer Cell 19 , 218 – 231.
Hwang, C.-S., Shemorry, A., and Varshavsky, A. (2010). N-terminal acetylation of cellular proteins creates specifi c degradation signals. Science 327 , 973 – 977.
Jeong, J.-W., Bae, M.-K., Ahn, M.-Y., Kim, S.-H., Sohn, T.-K., Bae, M.-H., Yoo, M.-A., Song, E.J., Lee, K.-J., and Kim, K.-W. (2002). Regulation and destabilization of HIF-1 α by ARD1-mediated acetylation. Cell 111 , 709 – 720.
Kamita, M., Kimura, Y., Ino, Y., Kamp, R.M., Polevoda, B., Sherman, F., and Hirano, H. (2011). N[ α ]-Acetylation of yeast ribosomal proteins and its effect on protein synthesis. J. Proteomics 74 , 431 – 441.
Kuo, M.-L., den Besten, W., Bertwistle, D., Roussel, M.F., and Sherr, C.J. (2004). N-terminal polyubiquitination and degradation of the Arf tumor suppressor. Genes Dev. 18 , 1862 – 1874.
Kuo, H.-P., Lee, D.-F., Chen, C.-T., Liu, M., Chou, C.-K., Lee, H.-J., Du, Y., Xie, X., Wei, Y., Xia, W., et al. (2010). ARD1 stabiliza-tion of TSC2 suppresses tumorigenesis through the mTOR sig-naling pathway. Sci. Signal. 3 , ra9.
Lee, F.J., Lin, L.W., and Smith, J.A. (1988). Purifi cation and charac-terization of an N α -acetyltransferase from Saccharomyces cer-evisiae . J. Biol. Chem. 263 , 14948 – 14955.
Lee, C.-F., Ou, D.S.C., Lee, S.-B., Chang, L.-H., Lin, R.-K., Li, Y.-S., Upadhyay, A.K., Cheng, X., Wang, Y.-C., Hsu, H.-S., et al. (2010). hNaa10p contributes to tumorigenesis by facilitat-ing DNMT1-mediated tumor suppressor gene silencing. J. Clin. Invest. 120 , 2920 – 2930.
Lim, J.H., Park, J.W., and Chun, Y.S. (2006). Human arrest defective 1 acetylates and activates beta-catenin, promoting lung cancer cell proliferation. Cancer Res. 66 , 10677 – 10682.
Mullen, J.R., Kayne, P.S., Moerschell, R.P., Tsunasawa, S., Gribskov, M., Colavito-Shepanski, M., Grunstein, M., Sherman, F., and Sternglanz, R. (1989). Identifi cation and characterization of genes and mutants for an N-terminal acetyltransferase from yeast. EMBO J. 8 , 2067 – 2075.
Persson, B., Flinta, C., von Heijne, G., and J ö rnvall, H. (1985). Structures of N-terminally acetylated proteins. Eur. J. Biochem. 152 , 523 – 527.
Pesaresi, P., Gardner, N.A., Masiero, S., Dietzmann, A., Eichacker, L., Wickner, R., Salamini, F., and Leister, D. (2003). Cytoplasmic N-terminal protein acetylation is required for effi cient photosyn-thesis in Arabidopsis . Plant Cell 15 , 1817 – 1832.
Polevoda, B. and Sherman, F. (2003). N-terminal acetyltransferases and sequence requirements for N-terminal acetylation of eukary-otic proteins. J. Mol. Biol. 325 , 595 – 622.
Polevoda, B., Norbeck, J., Takakura, H., Blomberg, A., and Sherman, F. (1999). Identifi cation and specifi cities of N-terminal acetyl-transferases from Saccharomyces cerevisiae . EMBO J. 18 , 6155 – 6168.
Polevoda, B., Arnesen, T., and Sherman, F. (2009). A synopsis of eukaryotic Nalpha-terminal acetyltransferases: nomenclature, subunits and substrates. BMC Proc. 3 (Suppl 6), S2.
Rope, A.F., Wang, K., Evjenth, R., Xing, J., Johnston, J.J., Swensen, J.J., Johnson, W.E., Moore, B., Huff, C.D., Bird, L.M., et al. (2011). Using VAAST to identify an X-linked disorder resulting
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM

298 J. Hollebeke et al.
in lethality in male infants due to N-terminal acetyltransferase defi ciency. Am. J. Hum. Genet. 89 , 28 – 43.
Shin, D.H., Chun, Y.S., Lee, K.H., Shin, H.W., and Park, J.W. (2009). Arrest defective-1 controls tumor cell behavior by acetylating myosin light chain kinase. PLoS One 4 , e7451.
Starheim, K.K., Arnesen, T., Gromyko, D., Ryningen, A., Varhaug, J.E., and Lillehaug, J.R. (2008). Identifi cation of the human N(alpha)-acetyltransferase complex B (hNatB): a complex important for cell-cycle progression. Biochem. J. 415 , 325 – 331.
Starheim, K.K., Gromyko, D., Evjenth, R., Ryningen, A., Varhaug, J.E., Lillehaug, J.R., and Arnesen, T. (2009). Knockdown of human N alpha-terminal acetyltransferase complex C leads to p53-dependent apoptosis and aberrant human Arl8b localization. Mol. Cell. Biol. 29 , 3569 – 3581.
Takubo, K., Tsuchiya, H., Kurimasa, A., Arnesen, T., Ryoke, K., and Shiota, G. (2009). Involvement of N-acetyltransferase human in the cytotoxic activity of 5-fl uorouracil. Anticancer Drugs 20 , 668 – 675.
Van Damme, P., Arnesen, T., and Gevaert, K. (2011a). Protein alpha-N-acetylation studied by N-terminomics. FEBS J. 278 , 3822 – 3834.
Van Damme, P., Evjenth, R., Foyn, H., Demeyer, K., De Bock, P.-J., Lillehaug, J.R., Vandekerckhove, J., Arnesen, T., and Gevaert, K. (2011b). Proteome-derived peptide libraries allow detailed analy-sis of the substrate specifi cities of N α -acetyltransferases and point to hNaa10p as the post-translational actin N α -acetyltransferase. Mol. Cell. Proteomics 10 , M110 004580.
Van Damme, P., Hole, K., Pimenta-Marques, A., Helsens, K., Vandekerckhove, J., Martinho, R.G., Gevaert, K., and Arnesen, T. (2011c). NatF contributes to an evolutionary shift in protein N-terminal acetylation and is important for normal chromosome segregation. PLoS Genet. 7 , e1002169.
Yi, C.H., Pan, H., Seebacher, J., Jang, I.-H., Hyberts, S.G., Heffron, G.J., Vander Heiden, M.G., Yang, R., Li, F., Locasale, J.W., et al. (2011). Metabolic regulation of protein N-alpha-acetylation by Bcl-xL promotes cell survival. Cell 146 , 607 – 620.
Zybailov, B., Rutschow, H., Friso, G., Rudella, A., Emanuelsson, O., Sun, Q., and van Wijk, K.J. (2008). Sorting signals, N-terminal modifi cations and abundance of the chloroplast proteome. PLoS One 3 , e1994.
Received October 10, 2011; accepted December 19, 2011
Brought to you by | Universitaetsbibliothek der LMU MuenchenAuthenticated | 129.187.254.47
Download Date | 6/18/14 11:01 PM


















