
Interferon β-secreting mesenchymal stem cells combined with minocycline attenuate experimental...
8
Interferon β-secreting mesenchymal stem cells combined with minocycline attenuate experimental autoimmune encephalomyelitis Yun Hou a,1 , Chung Heon Ryu b,1 , Jin Ae Jun a , Seong Muk Kim b , Chang Hyun Jeong a , Sin-Soo Jeun a,c, ⁎ a Department of Biomedical Science, College of Medicine, The Catholic University of Korea, Seoul, Republic of Korea b Postech-Catholic Biomedical Engineering Institute, Seoul St. Mary's Hospital, The Catholic University of Korea, Seoul, Republic of Korea c Department of Neurosurgery, Seoul St. Mary's Hospital, The Catholic University of Korea, Seoul, Republic of Korea abstract article info Article history: Received 13 February 2014 Received in revised form 21 May 2014 Accepted 2 June 2014 Available online xxxx Keywords: IFN-β-secreting mesenchymal stem cells Minocycline Experimental autoimmune encephalomyelitis Multiple sclerosis Blood–spinal cord barrier disruption Tight junctions We previously demonstrated that interferon β (IFN-β)-secreting mesenchymal stem cells (MSCs-IFN-β) strongly reduced the clinical severity of experimental autoimmune encephalomyelitis (EAE), compared with MSCs alone. Recently, minocycline ameliorates the clinical severity of multiple sclerosis (MS). Herein, we evaluated the effects of a combined treatment of MSCs-IFN-β and minocycline on EAE mice. The combined treatment significantly alleviated the clinical severity mainly by maintaining the integrity of blood–spinal cord barrier, in a manner likely involving inhibition of microvascular disruption, matrix metalloproteinases, neuroinflammation, and enhancement of immunomodulatory effects. Therefore, this combined treatment has the potential to improve the functional recovery of patients with MS. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Multiple sclerosis (MS) is the most common demyelinating disease of the central nervous system (CNS). The pathological characteristics of MS include blood–brain barrier (BBB) disruption, infiltration of inflammatory cells into the CNS, demyelination, and lesions in the spinal cord white matter. Interferon-beta (IFN-β), methylprednisolone, minocycline, and atorvastatin have all been demonstrated to reduce the clinical severity of either MS or experimental autoimmune enceph- alomyelitis (EAE), while it has been established that combined treatments have been gaining momentum compared with monotherapy for treating MS or EAE (Giuliani et al., 2005; Luccarini et al., 2008; Chen et al., 2010). Minocycline is a semi-synthetic tetracycline analog suitable for MS treatment. It has been shown to delay disease progression and reduce disease severity by inhibiting matrix metalloproteinase (MMP) activities, modulating the immune response, inhibiting inflammatory influx, and reducing edema in EAE animals and patients with MS (Brundula et al., 2002; Popovic et al., 2002; Yong et al., 2004). However, minocycline is associated with some adverse effects, including systemic lupus erythematosus, serum sickness, and CNS toxicity at high doses (Elkayam et al., 1999; Sturkenboom et al., 1999; Hollborn et al., 2010). To overcome these obstacles associated with minocycline-based regi- mens, we recently demonstrated that a treatment combining a low dose of minocycline with mesenchymal stem cells exerted synergistic therapeutic effects on EAE mice (Hou et al., 2013). Interferon-β has been approved as one of the most promising treatments for MS. However, the development of effective IFN-β- based therapies is hindered by many obstacles, including insufficient duration, a short half-life in vivo, inaccessibility to the CNS, and adverse effects such as irritation at the injection site. The use of IFN-β-secreting mesenchymal stem cells (MSCs-IFN-β) has been proposed to overcome many of these obstacles. In a previous work, MSCs-IFN-β were demon- strated to strongly reduce the extent of inflammatory cell infiltration, alleviate demyelination, and promote a shift in the cytokine response from Th1 to Th2, compared with MSCs in EAE treatment (Ryu et al., 2013). Therefore, we hypothesized that a combined treatment with a low dose of minocycline MSCs-IFN-β may provide better therapeutic effects compared with each treatment individually. The blood–spinal cord barrier (BSCB) is a barrier similar to the BBB; disruption of the BSCB is a primary and continuous pathological characteristic of MS. During neuroinflammation, cytokines produced by pathogenic T-cells, activated astrocytes, and resident spinal cord microglia mediate upregulation of adhesion molecules in the BSCB (Bartanusz et al., 2011). These lead to BSCB activation and its subsequent breakdown, resulting in an influx of immune cells into the CNS Journal of Neuroimmunology xxx (2014) xxx–xxx ⁎ Correspondence to: S.-S. Jeun, Department of Biomedical Science, Department of Neurosurgery, Seoul St. Mary's Hospital, College of Medicine, Catholic University of Korea, 505 Banpo-dong, Socho-gu, Seoul 137-701, Republic of Korea. Tel.: +82 2 2258 7535; fax: +82 2 3482 1853. E-mail address: [email protected] (S.-S. Jeun). 1 These authors contributed equally to this work. JNI-475920; No of Pages 8 http://dx.doi.org/10.1016/j.jneuroim.2014.06.001 0165-5728/© 2014 Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Journal of Neuroimmunology journal homepage: www.elsevier.com/locate/jneuroim Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchymal stem cells combined with minocycline attenuate experimental autoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.org/10.1016/j.jneuroim.2014.06.001
Transcript of Interferon β-secreting mesenchymal stem cells combined with minocycline attenuate experimental...

Journal of Neuroimmunology xxx (2014) xxx–xxx
JNI-475920; No of Pages 8
Contents lists available at ScienceDirect
Journal of Neuroimmunology
j ourna l homepage: www.e lsev ie r .com/ locate / jneuro im
Interferon β-secreting mesenchymal stem cells combined withminocycline attenuate experimental autoimmune encephalomyelitis
Yun Hou a,1, Chung Heon Ryu b,1, Jin Ae Jun a, Seong Muk Kim b, Chang Hyun Jeong a, Sin-Soo Jeun a,c,⁎a Department of Biomedical Science, College of Medicine, The Catholic University of Korea, Seoul, Republic of Koreab Postech-Catholic Biomedical Engineering Institute, Seoul St. Mary's Hospital, The Catholic University of Korea, Seoul, Republic of Koreac Department of Neurosurgery, Seoul St. Mary's Hospital, The Catholic University of Korea, Seoul, Republic of Korea
⁎ Correspondence to: S.-S. Jeun, Department of BiomNeurosurgery, Seoul St. Mary's Hospital, College of MeKorea, 505 Banpo-dong, Socho-gu, Seoul 137-701, Repu7535; fax: +82 2 3482 1853.
E-mail address: [email protected] (S.-S. Jeun).1 These authors contributed equally to this work.
http://dx.doi.org/10.1016/j.jneuroim.2014.06.0010165-5728/© 2014 Elsevier B.V. All rights reserved.
Please cite this article as: Hou, Y., et al., Inteautoimmune encephalomyelitis, J. Neuroimm
a b s t r a c t
a r t i c l e i n f oArticle history:Received 13 February 2014Received in revised form 21 May 2014Accepted 2 June 2014Available online xxxx
Keywords:IFN-β-secreting mesenchymal stem cellsMinocyclineExperimental autoimmune encephalomyelitisMultiple sclerosisBlood–spinal cord barrier disruptionTight junctions
Wepreviously demonstrated that interferonβ (IFN-β)-secretingmesenchymal stem cells (MSCs-IFN-β) stronglyreduced the clinical severity of experimental autoimmune encephalomyelitis (EAE), comparedwith MSCs alone.Recently,minocycline ameliorates the clinical severity ofmultiple sclerosis (MS). Herein,we evaluated the effectsof a combined treatment of MSCs-IFN-β and minocycline on EAE mice. The combined treatment significantlyalleviated the clinical severitymainly bymaintaining the integrity of blood–spinal cord barrier, in amanner likelyinvolving inhibition ofmicrovascular disruption, matrix metalloproteinases, neuroinflammation, and enhancementof immunomodulatory effects. Therefore, this combined treatment has the potential to improve the functionalrecovery of patients with MS.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Multiple sclerosis (MS) is the most common demyelinating diseaseof the central nervous system (CNS). The pathological characteristicsof MS include blood–brain barrier (BBB) disruption, infiltration ofinflammatory cells into theCNS, demyelination, and lesions in the spinalcord white matter. Interferon-beta (IFN-β), methylprednisolone,minocycline, and atorvastatin have all been demonstrated to reducethe clinical severity of either MS or experimental autoimmune enceph-alomyelitis (EAE), while it has been established that combined treatmentshave been gaining momentum compared with monotherapy for treatingMS or EAE (Giuliani et al., 2005; Luccarini et al., 2008; Chen et al., 2010).
Minocycline is a semi-synthetic tetracycline analog suitable for MStreatment. It has been shown to delay disease progression and reducedisease severity by inhibitingmatrixmetalloproteinase (MMP) activities,modulating the immune response, inhibiting inflammatory influx, andreducing edema in EAE animals and patients with MS (Brundula et al.,2002; Popovic et al., 2002; Yong et al., 2004). However, minocycline isassociated with some adverse effects, including systemic lupus
edical Science, Department ofdicine, Catholic University ofblic of Korea. Tel.: +82 2 2258
rferon β-secreting mesenchunol. (2014), http://dx.doi.o
erythematosus, serum sickness, and CNS toxicity at high doses(Elkayam et al., 1999; Sturkenboom et al., 1999; Hollborn et al., 2010).To overcome these obstacles associated with minocycline-based regi-mens, we recently demonstrated that a treatment combining a lowdose of minocycline with mesenchymal stem cells exerted synergistictherapeutic effects on EAE mice (Hou et al., 2013).
Interferon-β has been approved as one of the most promisingtreatments for MS. However, the development of effective IFN-β-based therapies is hindered by many obstacles, including insufficientduration, a short half-life in vivo, inaccessibility to the CNS, and adverseeffects such as irritation at the injection site. The use of IFN-β-secretingmesenchymal stem cells (MSCs-IFN-β) has been proposed to overcomemany of these obstacles. In a previous work, MSCs-IFN-β were demon-strated to strongly reduce the extent of inflammatory cell infiltration,alleviate demyelination, and promote a shift in the cytokine responsefrom Th1 to Th2, compared with MSCs in EAE treatment (Ryu et al.,2013). Therefore, we hypothesized that a combined treatment with alow dose of minocycline MSCs-IFN-β may provide better therapeuticeffects compared with each treatment individually.
The blood–spinal cord barrier (BSCB) is a barrier similar to the BBB;disruption of the BSCB is a primary and continuous pathologicalcharacteristic of MS. During neuroinflammation, cytokines producedby pathogenic T-cells, activated astrocytes, and resident spinal cordmicroglia mediate upregulation of adhesion molecules in the BSCB(Bartanusz et al., 2011). These lead to BSCB activation and its subsequentbreakdown, resulting in an influx of immune cells into the CNS
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001

2 Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
(Schellenberg et al., 2007; Ransohoff and Engelhardt, 2012). Theactivated BSCB also secretes chemokines and cytokines, therebyadditionally stimulating both proliferation and recruitment of inflamma-tory cells into the CNS (Bartanusz et al., 2011). This vicious cycle couldpotentially be disrupted by targeting the mechanisms regulating BSCBfunction and integrity; consistent with this hypothesis, inhibition of BBBdisruption has been shown to attenuate the severity of EAE (Adzemovicet al., 2013). Therefore, further improvements in MS therapeutics may beachieved by inhibiting this activation of the BSCB.
In the present study, we investigated the effects of combinedtreatment with minocycline and MSCs-IFN-β on EAE mice. Ourresults demonstrate that the combined treatment strongly reducedthe severity of neurological disease, especially by inhibition of BSCBdisruption. The data presented here suggest that this combinedtreatment has the potential to improve the functional recovery ofpatients with MS.
2. Materials and methods
2.1. Cell culture and reagents
Human bone marrow-derived mesenchymal stem cells (CatholicMASTER Cells) were obtained from the Catholic Institute of Cell Therapy(CIC, Seoul, Korea). Cells were thawed and cultured according to themanufacturer's instructions (Jeong et al., 2014). Minocycline waspurchased from Sigma-Aldrich (St. Louis, MO, USA). Evans blue dyewas purchased from Sigma.
2.2. Adenovirus infection to drive IFN-β expression
A recombinant adenoviral vector encodingmouse IFN-β (Ad-IFN-β)gene was constructed using the AdEasy vector system (QuantumBiotechnologies, Carlsbad, CA, USA) according to the manufacturer'sinstructions. Briefly, adenoviruses were allowed to bind to the 4HP4peptide in Opti-MEM (Gibco, Invitrogen, Carlsbad, CA, USA) for 30 minat room temperature (RT). Next, hBM-MSCs were incubated withpremixed virus-4HP4 complexes (multiplicity of infection = 50)for 6 h at 37 °C, washed twice with PBS, and incubated in maintenancemedium.
2.3. EAE induction and treatment
All animal protocols were approved by the Institutional Animal Careand Use Committee of the Medical College of Catholic University. EAEwas induced in C57BL/6 mice (12-week-old, female) by immunizationwith the MOG35-55 peptide (Hooke Labs, Lawrence, MA, USA)according to the manufacturer's instructions. Briefly, mice wereinjected subcutaneously with the MOG35-55 peptide,which is emul-sified in complete Freund's adjuvant (CFA) containing Mycobacteri-um tuberculosis. At 2 and 24 h post MOG35-55 injection, micereceived an intraperitoneal pertussis toxin injection. Paralysis asclinical evidence of EAEwas assessed daily starting on day 7 post immu-nization. Clinically, animals were scored as follows: 0, no clinicalsigns; 1, limp tail; 2, partial hind leg paralysis; 3, complete hind legparalysis; 4, complete hind leg paralysis and partial front leg paraly-sis; and 5, moribund or dead. Forty mice were randomly divided into4 groups: PBS (n = 10); minocycline (10 mg/kg, intraperitoneal in-jection, n = 10); MSCs-IFN-β (1.0 × 106 cells in 100 μl PBS for eachmouse, intravenous injection, n = 10); and a combination ofminocycline and MSCs-IFN-β (n= 10). All treatments were initiatedon day 13 post immunization, a timepoint at which all mice mani-fested clinical signs of EAE. Minocycline was administered intraper-itoneally for 37 consecutive days until sacrifice.
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
2.4. Assessment of BSCB integrity by extravasation of Evans blue (EB)
Evans blue dye (2% in saline, 4 ml/kg) was given intravenously toEAE mice between days 26–28 post immunization. Four hours afterEvans blue injection, animals were perfused with PBS to remove theintravascular dye. After decapitation, entire spinal cords were removedandhomogenized in 200 μl N,N-dimethyl formamide (Sigma) for 24h at55 °C and centrifuged at 10,000 ×g for 30 min. The absorbances at620 nm of the supernatants were then measured using a spectropho-tometer (Molecular Devices, Sunnyvale, CA, USA).
2.5. Immunoblot analysis of tight junction proteins
Fresh spinal cords were collected from EAE mice between days26–28 post immunization. Tissues were homogenized in T-PER TissueProtein Extraction Reagent (Thermo, Pittsburgh, PA, USA). The proteinsin the resultant lysates were resolved by sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE) on 10% gels. Proteinswere then transferred to nitrocellulose membranes whichwere probedwith primary antibodies against β-actin (Sigma) or occludin (ChemiconInternational Temecula, CA, USA), followed by horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology, Dallas, TX,USA). Immunoreactive bands were detected using Amersham ECLdetection reagents (GE Healthcare Life Sciences, Pittsburgh, PA, USA).
2.6. Determination of matrix metalloproteinase (MMP) activation bygelatin zymography
To detect the expression levels of MMP-2 and MMP-9 in the freshspinal cords, gelatin zymography was performed using NovexZymogram Gels (Life Technologies, Carlsbad, CM, USA) according tothe manufacturer's instructions. Briefly, tissues were homogenized inT-PER Tissue Protein Extraction Reagent (Thermo). After centrifugation,supernatants were harvested, mixed with Tris-Glycine SDS samplebuffer, and resolved on 8% SDS gels. Gelswere renaturated and incubatedin incubation buffer for 18 h and then developed with 0.5% CoomassieBlue R-250 (Sigma).
2.7. Immunofluorescence stains
To detect the neuroinflammation, mice were sacrificed on day 28post immunization for immunofluorescence staining. Lumbar spinalcord sections were incubated at 4 °C overnight with the followingantibodies: monoclonal mouse anti-glial fibrillary acidic protein(GFAP; Millipore, Temecula, CA, USA), polyclonal rabbit anti-ionizedcalcium-binding adaptor molecule 1 (Iba1; Wako Pure Chemical Indus-tries, Osaka, Japan), or monoclonal mouse anti-CD4 (BD BiosciencesPharmingen, CA, USA). Immunoreactive proteins were visualized withanti-rabbit and anti-mouse Cy3-conjugated secondary antibodies(Jackson ImmunoResearch, West Grove, PA, USA). Immunoreactivityspecificity was confirmed by control reactions, in which primary orsecondary antibodies were omitted. Cell nuclei were counterstainedwith 4-6-diamidino-2-phenylindole (DAPI; Roche, Penzberg,Germany) for 15min. All imageswere acquired using a LSM700 confocalmicroscope (Carl Zeiss, Oberkochen, Germany).
2.8. Quantification of serum cytokines by enzyme-linked immunosorbentassays (ELISAs)
Serum was obtained from all animals in each treatment groupbetween days 26–28 post-immunization. ELISAs were performedusing a Quantikine Immunoassay Kit from R&D Systems (Madison, WI,USA). Briefly, sera were incubated in precoated 96-well plates for 2 hat RT. After three washes, plates were incubated with conjugatedantibody for 2 h at RT and incubated in substrate solution for 30 min.
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001
3Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
Finally, stop solution was added, and the optical density of each well at450 nmwas determined using amicroplate reader (Molecular Devices).
2.9. Quantification and statistical analysis
Quantification was performed by an examiner blinded to thetreatment status of each animal. Six to 8 sections from each of fivetransverse lumbar spinal cords collected from each group were qualita-tively analyzed. H&E and LFB stains were viewed with a Slide Scannerfor Digital Pathology (SCN400, Leica, Wetzlar, Germany) under a ×200objective lens. Immunofluorescence was viewed with a confocal micro-scope under a ×200 objective lens. All images were measured withMetaMorph software, version 7.5 (Molecular Devices). The averagenumber of infiltrated inflammatory cells, positive immunostained cellnumber or fluorescence intensity was presented as the cell number orfluorescence intensity in the lesion sites under each photographedobjective magnification. Lesion size was determined by quantitativehistological analysis of the LFB-counterstained spinal cord sections. Thelesion size was presented as lesion area under photographed objectivemagnification. The tissue content of values of EBwas presented as inten-sity offluorescence. TheMMPdensitiesweremeasuredwithMetaMorphsoftware, version 7.5 (Molecular Devices). All statistical comparisonsbetween the groups were examined using one-way analysis of variance(ANOVA) with post-hoc Bonferroni corrections. p values b0.05 wereconsidered statistically significant.
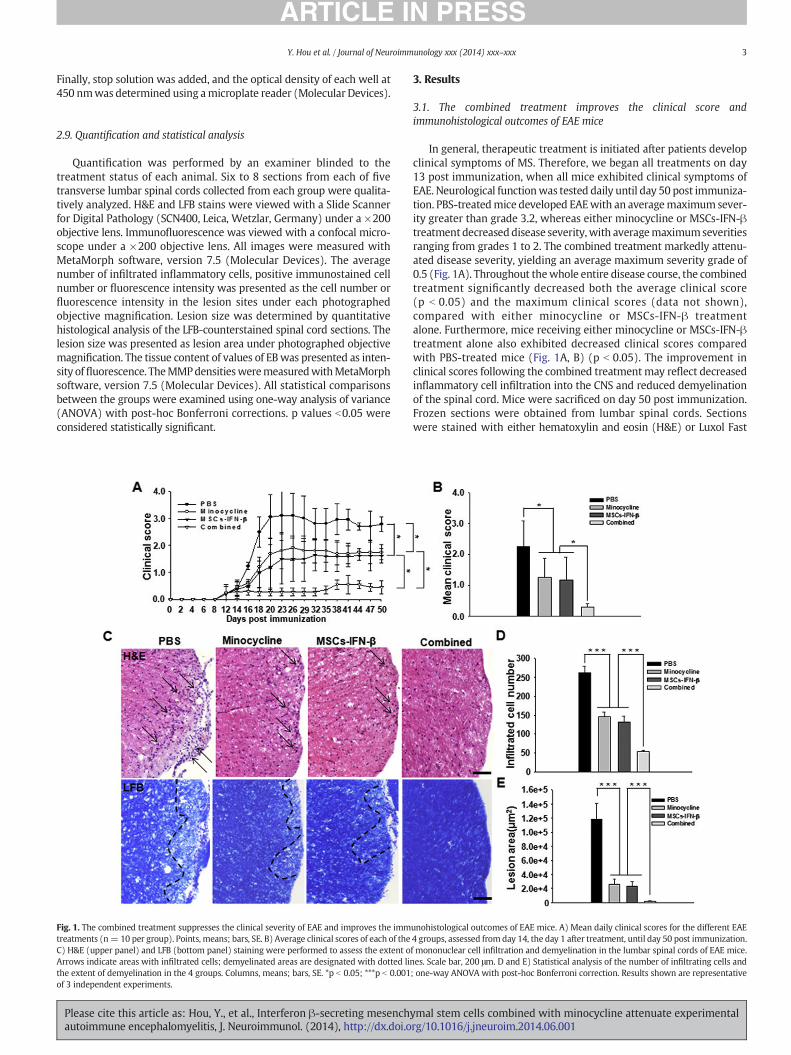
Fig. 1. The combined treatment suppresses the clinical severity of EAE and improves the immtreatments (n= 10 per group). Points, means; bars, SE. B) Average clinical scores of each of theC) H&E (upper panel) and LFB (bottom panel) staining were performed to assess the extent oArrows indicate areas with infiltrated cells; demyelinated areas are designated with dotted linthe extent of demyelination in the 4 groups. Columns, means; bars, SE. *p b 0.05; ***p b 0.001of 3 independent experiments.
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
3. Results
3.1. The combined treatment improves the clinical score andimmunohistological outcomes of EAE mice
In general, therapeutic treatment is initiated after patients developclinical symptoms of MS. Therefore, we began all treatments on day13 post immunization, when all mice exhibited clinical symptoms ofEAE. Neurological functionwas tested daily until day 50 post immuniza-tion. PBS-treatedmice developed EAEwith an averagemaximum sever-ity greater than grade 3.2, whereas either minocycline or MSCs-IFN-βtreatment decreaseddisease severity,with averagemaximumseveritiesranging from grades 1 to 2. The combined treatment markedly attenu-ated disease severity, yielding an average maximum severity grade of0.5 (Fig. 1A). Throughout thewhole entire disease course, the combinedtreatment significantly decreased both the average clinical score(p b 0.05) and the maximum clinical scores (data not shown),compared with either minocycline or MSCs-IFN-β treatmentalone. Furthermore, mice receiving either minocycline or MSCs-IFN-βtreatment alone also exhibited decreased clinical scores comparedwith PBS-treated mice (Fig. 1A, B) (p b 0.05). The improvement inclinical scores following the combined treatmentmay reflect decreasedinflammatory cell infiltration into the CNS and reduced demyelinationof the spinal cord. Mice were sacrificed on day 50 post immunization.Frozen sections were obtained from lumbar spinal cords. Sectionswere stained with either hematoxylin and eosin (H&E) or Luxol Fast
unohistological outcomes of EAE mice. A) Mean daily clinical scores for the different EAE4 groups, assessed fromday 14, the day 1 after treatment, until day 50 post immunization.f mononuclear cell infiltration and demyelination in the lumbar spinal cords of EAE mice.es. Scale bar, 200 μm. D and E) Statistical analysis of the number of infiltrating cells and; one-way ANOVA with post-hoc Bonferroni correction. Results shown are representative
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001
4 Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
Blue (LFB). H&E staining was used to detect infiltrated inflammatorymononuclear cells, which were particularly apparent in the white matterof the spinal cord. The amount of inflammatory cellular infiltrate was sig-nificantly decreased by the combined treatment compared with eitherminocycline orMSCs-IFN-β treatment alone (p b 0.001).Mice receiving ei-ther minocycline or MSCs-IFN-β treatment alone also exhibited a reducedamount of inflammatory cellular infiltrates compared with PBS-treatedmice (p b 0.001) (Fig. 1C, D). To investigate the extent of demyelination,LFB stainingwas performed. In accordancewith the cellular infiltration re-sults, the combined treatment significantly reduced the extent of demye-lination in the white matter of the spinal cord compared with eitherminocycline orMSCs-IFN-β treatment alone (p b 0.001).Mice receiving ei-ther the minocycline or the MSCs-IFN-β treatment also exhibited reduceddemyelination comparedwith PBS-treatedmice (p b 0.001) (Fig. 1C, E). Toensure that the minocycline in the combined treatment did not affect theMSCs used in the treatment, we investigated the effects of minocycline(10 μM) on the viability of MSCs-IFN-β and their ability to secrete IFN-βin vitro. This concentration ofminocyclinewas chosen based on a previousstudy (Hou et al., 2013). Minocycline did not affect the viability of MSCs-IFN-β, nor did it affect the ability of MSCs expressing mouse IFN-β to se-crete IFN-β (Fig. S1).
3.2. The combined treatment reduces neuroinflammation in the spinalcords of EAE mice
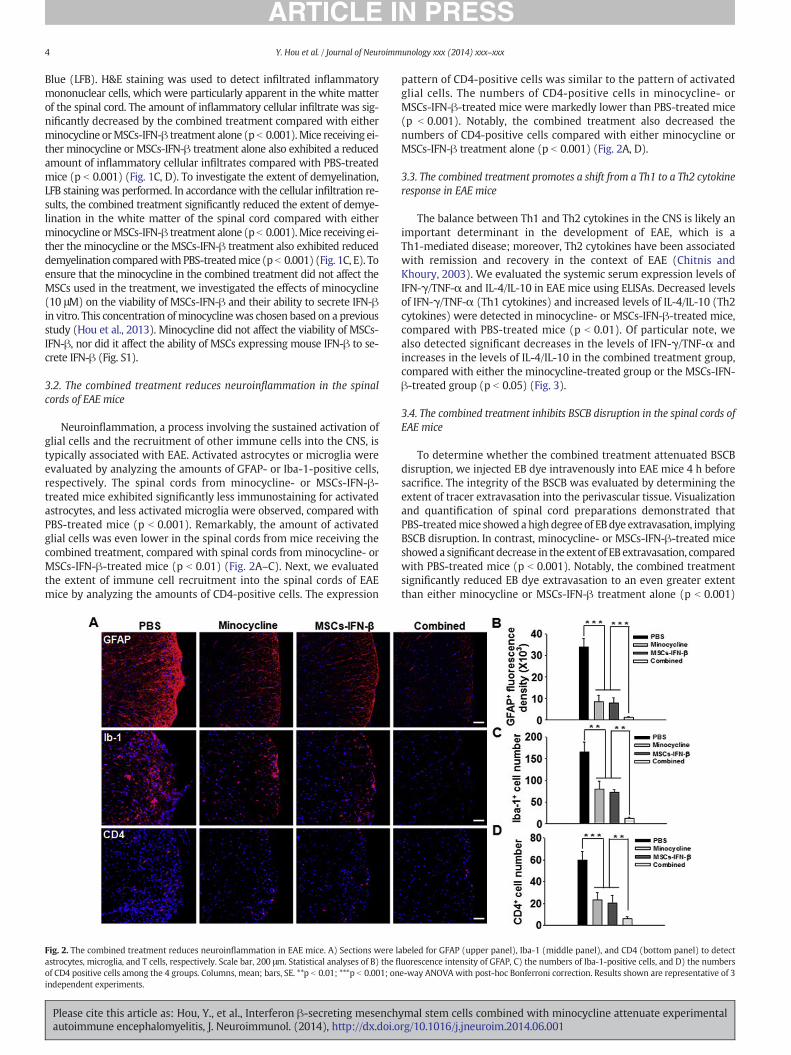
Neuroinflammation, a process involving the sustained activation ofglial cells and the recruitment of other immune cells into the CNS, istypically associated with EAE. Activated astrocytes or microglia wereevaluated by analyzing the amounts of GFAP- or Iba-1-positive cells,respectively. The spinal cords from minocycline- or MSCs-IFN-β-treated mice exhibited significantly less immunostaining for activatedastrocytes, and less activated microglia were observed, compared withPBS-treated mice (p b 0.001). Remarkably, the amount of activatedglial cells was even lower in the spinal cords from mice receiving thecombined treatment, compared with spinal cords from minocycline- orMSCs-IFN-β-treated mice (p b 0.01) (Fig. 2A–C). Next, we evaluatedthe extent of immune cell recruitment into the spinal cords of EAEmice by analyzing the amounts of CD4-positive cells. The expression
Fig. 2. The combined treatment reduces neuroinflammation in EAE mice. A) Sections were lastrocytes, microglia, and T cells, respectively. Scale bar, 200 μm. Statistical analyses of B) the fl
of CD4 positive cells among the 4 groups. Columns, mean; bars, SE. **p b 0.01; ***p b 0.001; onindependent experiments.
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
pattern of CD4-positive cells was similar to the pattern of activatedglial cells. The numbers of CD4-positive cells in minocycline- orMSCs-IFN-β-treated mice were markedly lower than PBS-treated mice(p b 0.001). Notably, the combined treatment also decreased thenumbers of CD4-positive cells compared with either minocycline orMSCs-IFN-β treatment alone (p b 0.001) (Fig. 2A, D).
3.3. The combined treatment promotes a shift from a Th1 to a Th2 cytokineresponse in EAE mice
The balance between Th1 and Th2 cytokines in the CNS is likely animportant determinant in the development of EAE, which is aTh1-mediated disease; moreover, Th2 cytokines have been associatedwith remission and recovery in the context of EAE (Chitnis andKhoury, 2003). We evaluated the systemic serum expression levels ofIFN-γ/TNF-α and IL-4/IL-10 in EAE mice using ELISAs. Decreased levelsof IFN-γ/TNF-α (Th1 cytokines) and increased levels of IL-4/IL-10 (Th2cytokines) were detected in minocycline- or MSCs-IFN-β-treated mice,compared with PBS-treated mice (p b 0.01). Of particular note, wealso detected significant decreases in the levels of IFN-γ/TNF-α andincreases in the levels of IL-4/IL-10 in the combined treatment group,compared with either the minocycline-treated group or the MSCs-IFN-β-treated group (p b 0.05) (Fig. 3).
3.4. The combined treatment inhibits BSCB disruption in the spinal cords ofEAE mice
To determine whether the combined treatment attenuated BSCBdisruption, we injected EB dye intravenously into EAE mice 4 h beforesacrifice. The integrity of the BSCB was evaluated by determining theextent of tracer extravasation into the perivascular tissue. Visualizationand quantification of spinal cord preparations demonstrated thatPBS-treatedmice showed ahighdegree of EBdye extravasation, implyingBSCB disruption. In contrast, minocycline- or MSCs-IFN-β-treated miceshowed a significant decrease in the extent of EB extravasation, comparedwith PBS-treated mice (p b 0.001). Notably, the combined treatmentsignificantly reduced EB dye extravasation to an even greater extentthan either minocycline or MSCs-IFN-β treatment alone (p b 0.001)
abeled for GFAP (upper panel), Iba-1 (middle panel), and CD4 (bottom panel) to detectuorescence intensity of GFAP, C) the numbers of Iba-1-positive cells, and D) the numberse-way ANOVA with post-hoc Bonferroni correction. Results shown are representative of 3
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001
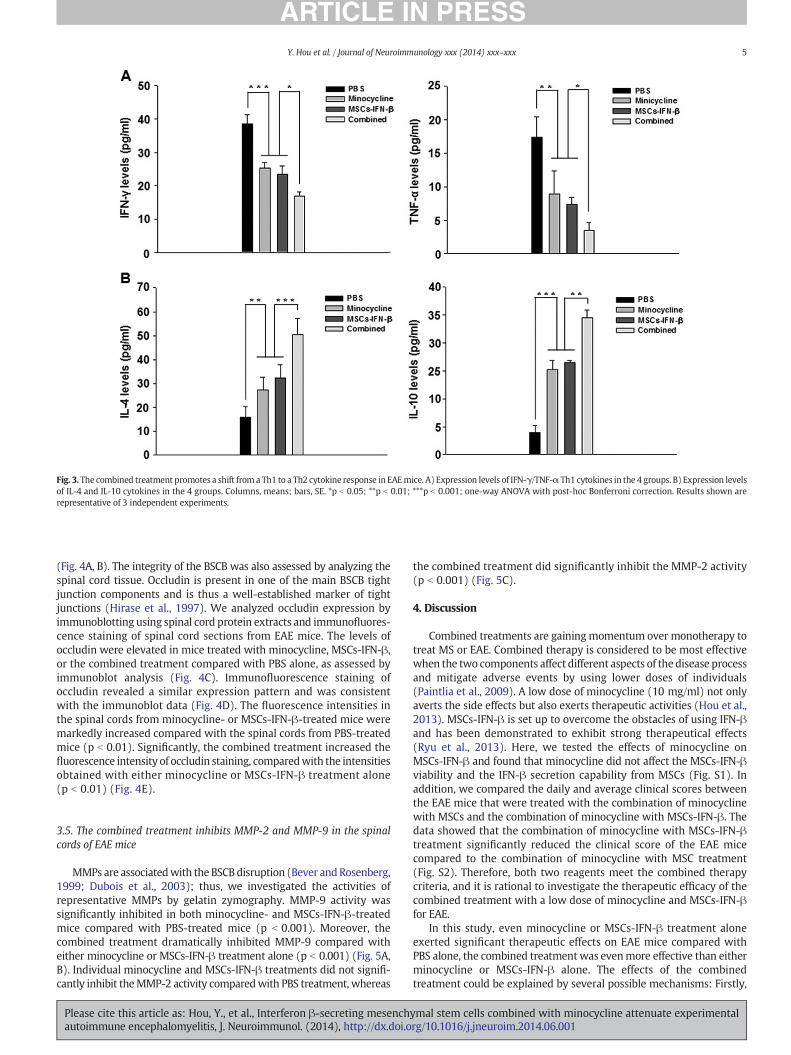
Fig. 3. The combined treatment promotes a shift from a Th1 to a Th2 cytokine response in EAEmice. A) Expression levels of IFN-γ/TNF-α Th1 cytokines in the 4 groups. B) Expression levelsof IL-4 and IL-10 cytokines in the 4 groups. Columns, means; bars, SE. *p b 0.05; **p b 0.01; ***p b 0.001; one-way ANOVA with post-hoc Bonferroni correction. Results shown arerepresentative of 3 independent experiments.
5Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
(Fig. 4A, B). The integrity of the BSCB was also assessed by analyzing thespinal cord tissue. Occludin is present in one of the main BSCB tightjunction components and is thus a well-established marker of tightjunctions (Hirase et al., 1997). We analyzed occludin expression byimmunoblotting using spinal cord protein extracts and immunofluores-cence staining of spinal cord sections from EAE mice. The levels ofoccludin were elevated in mice treated with minocycline, MSCs-IFN-β,or the combined treatment compared with PBS alone, as assessed byimmunoblot analysis (Fig. 4C). Immunofluorescence staining ofoccludin revealed a similar expression pattern and was consistentwith the immunoblot data (Fig. 4D). The fluorescence intensities inthe spinal cords from minocycline- or MSCs-IFN-β-treated mice weremarkedly increased compared with the spinal cords from PBS-treatedmice (p b 0.01). Significantly, the combined treatment increased thefluorescence intensity of occludin staining, comparedwith the intensitiesobtained with either minocycline or MSCs-IFN-β treatment alone(p b 0.01) (Fig. 4E).
3.5. The combined treatment inhibits MMP-2 and MMP-9 in the spinalcords of EAE mice
MMPs are associatedwith the BSCBdisruption (Bever andRosenberg,1999; Dubois et al., 2003); thus, we investigated the activities ofrepresentative MMPs by gelatin zymography. MMP-9 activity wassignificantly inhibited in both minocycline- and MSCs-IFN-β-treatedmice compared with PBS-treated mice (p b 0.001). Moreover, thecombined treatment dramatically inhibited MMP-9 compared witheither minocycline or MSCs-IFN-β treatment alone (p b 0.001) (Fig. 5A,B). Individual minocycline and MSCs-IFN-β treatments did not signifi-cantly inhibit theMMP-2 activity comparedwith PBS treatment, whereas
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
the combined treatment did significantly inhibit the MMP-2 activity(p b 0.001) (Fig. 5C).
4. Discussion
Combined treatments are gainingmomentum over monotherapy totreat MS or EAE. Combined therapy is considered to be most effectivewhen the two components affect different aspects of the disease processand mitigate adverse events by using lower doses of individuals(Paintlia et al., 2009). A low dose of minocycline (10 mg/ml) not onlyaverts the side effects but also exerts therapeutic activities (Hou et al.,2013). MSCs-IFN-β is set up to overcome the obstacles of using IFN-βand has been demonstrated to exhibit strong therapeutical effects(Ryu et al., 2013). Here, we tested the effects of minocycline onMSCs-IFN-β and found that minocycline did not affect the MSCs-IFN-βviability and the IFN-β secretion capability from MSCs (Fig. S1). Inaddition, we compared the daily and average clinical scores betweenthe EAE mice that were treated with the combination of minocyclinewith MSCs and the combination of minocycline with MSCs-IFN-β. Thedata showed that the combination of minocycline with MSCs-IFN-βtreatment significantly reduced the clinical score of the EAE micecompared to the combination of minocycline with MSC treatment(Fig. S2). Therefore, both two reagents meet the combined therapycriteria, and it is rational to investigate the therapeutic efficacy of thecombined treatment with a low dose of minocycline and MSCs-IFN-βfor EAE.
In this study, even minocycline or MSCs-IFN-β treatment aloneexerted significant therapeutic effects on EAE mice compared withPBS alone, the combined treatmentwas evenmore effective than eitherminocycline or MSCs-IFN-β alone. The effects of the combinedtreatment could be explained by several possible mechanisms: Firstly,
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001
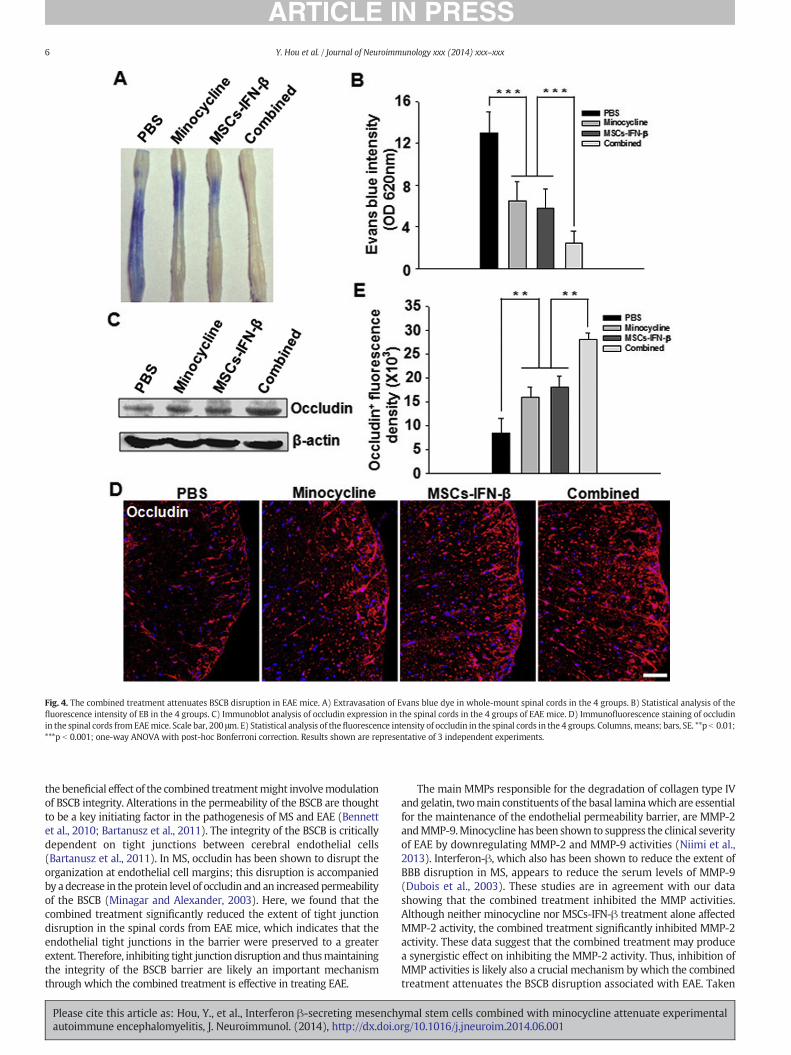
Fig. 4. The combined treatment attenuates BSCB disruption in EAE mice. A) Extravasation of Evans blue dye in whole-mount spinal cords in the 4 groups. B) Statistical analysis of thefluorescence intensity of EB in the 4 groups. C) Immunoblot analysis of occludin expression in the spinal cords in the 4 groups of EAE mice. D) Immunofluorescence staining of occludinin the spinal cords fromEAEmice. Scale bar, 200 μm. E) Statistical analysis of thefluorescence intensity of occludin in the spinal cords in the 4 groups. Columns, means; bars, SE. **p b 0.01;***p b 0.001; one-way ANOVA with post-hoc Bonferroni correction. Results shown are representative of 3 independent experiments.
6 Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
the beneficial effect of the combined treatmentmight involvemodulationof BSCB integrity. Alterations in the permeability of the BSCB are thoughtto be a key initiating factor in the pathogenesis of MS and EAE (Bennettet al., 2010; Bartanusz et al., 2011). The integrity of the BSCB is criticallydependent on tight junctions between cerebral endothelial cells(Bartanusz et al., 2011). In MS, occludin has been shown to disrupt theorganization at endothelial cell margins; this disruption is accompaniedby adecrease in the protein level of occludin and an increasedpermeabilityof the BSCB (Minagar and Alexander, 2003). Here, we found that thecombined treatment significantly reduced the extent of tight junctiondisruption in the spinal cords from EAE mice, which indicates that theendothelial tight junctions in the barrier were preserved to a greaterextent. Therefore, inhibiting tight junction disruption and thusmaintainingthe integrity of the BSCB barrier are likely an important mechanismthrough which the combined treatment is effective in treating EAE.
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
The main MMPs responsible for the degradation of collagen type IVand gelatin, twomain constituents of the basal laminawhich are essentialfor the maintenance of the endothelial permeability barrier, are MMP-2andMMP-9.Minocycline has been shown to suppress the clinical severityof EAE by downregulating MMP-2 and MMP-9 activities (Niimi et al.,2013). Interferon-β, which also has been shown to reduce the extent ofBBB disruption in MS, appears to reduce the serum levels of MMP-9(Dubois et al., 2003). These studies are in agreement with our datashowing that the combined treatment inhibited the MMP activities.Although neither minocycline nor MSCs-IFN-β treatment alone affectedMMP-2 activity, the combined treatment significantly inhibited MMP-2activity. These data suggest that the combined treatment may producea synergistic effect on inhibiting the MMP-2 activity. Thus, inhibition ofMMP activities is likely also a crucial mechanism by which the combinedtreatment attenuates the BSCB disruption associated with EAE. Taken
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001
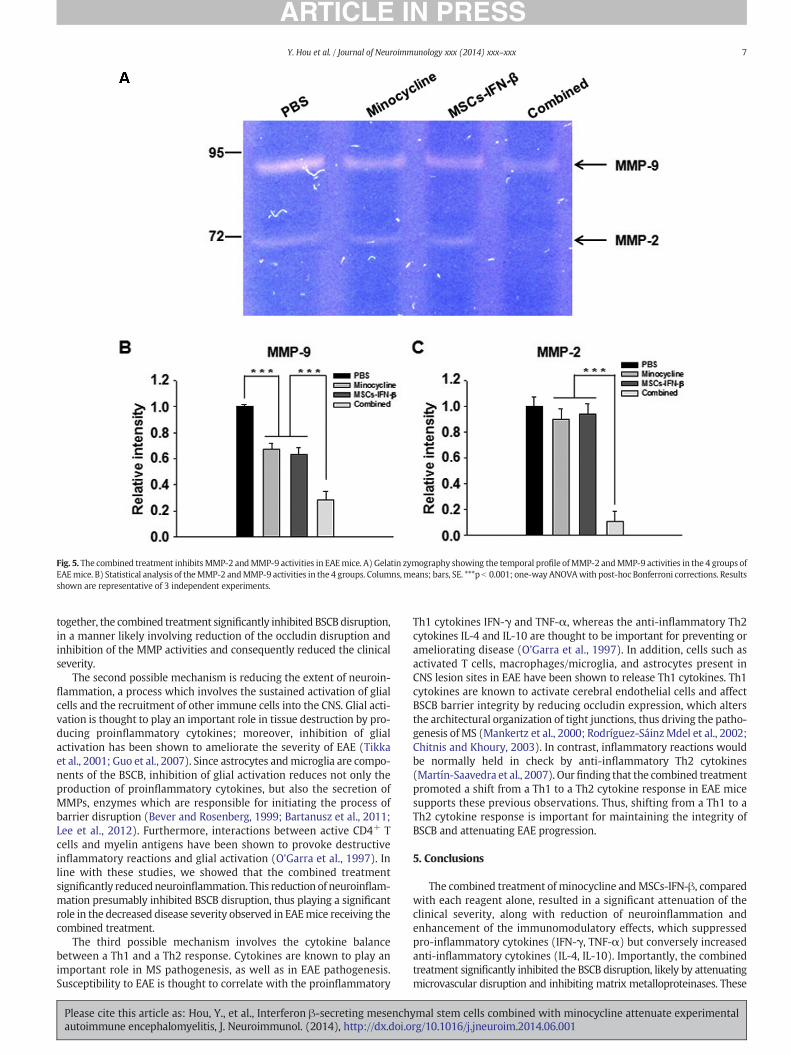
Fig. 5. The combined treatment inhibits MMP-2 andMMP-9 activities in EAEmice. A) Gelatin zymography showing the temporal profile ofMMP-2 andMMP-9 activities in the 4 groups ofEAEmice. B) Statistical analysis of theMMP-2 andMMP-9 activities in the 4 groups. Columns,means; bars, SE. ***p b 0.001; one-way ANOVAwith post-hoc Bonferroni corrections. Resultsshown are representative of 3 independent experiments.
7Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
together, the combined treatment significantly inhibited BSCB disruption,in a manner likely involving reduction of the occludin disruption andinhibition of the MMP activities and consequently reduced the clinicalseverity.
The second possible mechanism is reducing the extent of neuroin-flammation, a process which involves the sustained activation of glialcells and the recruitment of other immune cells into the CNS. Glial acti-vation is thought to play an important role in tissue destruction by pro-ducing proinflammatory cytokines; moreover, inhibition of glialactivation has been shown to ameliorate the severity of EAE (Tikkaet al., 2001; Guo et al., 2007). Since astrocytes andmicroglia are compo-nents of the BSCB, inhibition of glial activation reduces not only theproduction of proinflammatory cytokines, but also the secretion ofMMPs, enzymes which are responsible for initiating the process ofbarrier disruption (Bever and Rosenberg, 1999; Bartanusz et al., 2011;Lee et al., 2012). Furthermore, interactions between active CD4+ Tcells and myelin antigens have been shown to provoke destructiveinflammatory reactions and glial activation (O'Garra et al., 1997). Inline with these studies, we showed that the combined treatmentsignificantly reducedneuroinflammation. This reduction of neuroinflam-mation presumably inhibited BSCB disruption, thus playing a significantrole in the decreased disease severity observed in EAEmice receiving thecombined treatment.
The third possible mechanism involves the cytokine balancebetween a Th1 and a Th2 response. Cytokines are known to play animportant role in MS pathogenesis, as well as in EAE pathogenesis.Susceptibility to EAE is thought to correlate with the proinflammatory
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
Th1 cytokines IFN-γ and TNF-α, whereas the anti-inflammatory Th2cytokines IL-4 and IL-10 are thought to be important for preventing orameliorating disease (O'Garra et al., 1997). In addition, cells such asactivated T cells, macrophages/microglia, and astrocytes present inCNS lesion sites in EAE have been shown to release Th1 cytokines. Th1cytokines are known to activate cerebral endothelial cells and affectBSCB barrier integrity by reducing occludin expression, which altersthe architectural organization of tight junctions, thus driving the patho-genesis of MS (Mankertz et al., 2000; Rodríguez-SáinzMdel et al., 2002;Chitnis and Khoury, 2003). In contrast, inflammatory reactions wouldbe normally held in check by anti-inflammatory Th2 cytokines(Martín-Saavedra et al., 2007). Our finding that the combined treatmentpromoted a shift from a Th1 to a Th2 cytokine response in EAE micesupports these previous observations. Thus, shifting from a Th1 to aTh2 cytokine response is important for maintaining the integrity ofBSCB and attenuating EAE progression.
5. Conclusions
The combined treatment of minocycline andMSCs-IFN-β, comparedwith each reagent alone, resulted in a significant attenuation of theclinical severity, along with reduction of neuroinflammation andenhancement of the immunomodulatory effects, which suppressedpro-inflammatory cytokines (IFN-γ, TNF-α) but conversely increasedanti-inflammatory cytokines (IL-4, IL-10). Importantly, the combinedtreatment significantly inhibited the BSCB disruption, likely by attenuatingmicrovascular disruption and inhibiting matrix metalloproteinases. These
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001

8 Y. Hou et al. / Journal of Neuroimmunology xxx (2014) xxx–xxx
results suggest that this combined treatment has the potential to improvethe functional recovery of patients with MS.
Abbreviations
BSCB blood–spinal cord barrierEAE experimental autoimmune encephalomyelitisGFAP glial fibrillary acidic proteinIba1 ionized calcium-binding adaptor molecule 1IFN-γ interferon gammaIL-4 interleukin-4IL-10 interleukin-10MS multiple sclerosisMSCs-IFN-β interferon β (IFN-β)-secreting hBM-MSCsTdT terminal deoxynucleotidyl transferaseTNF-α tumor necrosis factor alpha
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.jneuroim.2014.06.001.
Acknowledgments
This studywas supported by grants from theKoreaHealth TechnologyR&DProject (A110330,HI09C1555),Ministry ofHealth&Welfare, Repub-lic of Korea and Research Fund of Seoul St. Mary's Hospital, The CatholicUniversity of Korea. The Catholic MASTER Cells supplied by CatholicInstitute of Cell Therapy (CIC, Seoul, Korea) were derived from humanbone marrow donated by healthy donors after informed consent.
ReferencesAdzemovic,M.Z., Zeitelhofer,M., Eriksson,U., Olsson, T., Nilsson, I., 2013. Imatinib ameliorates
neuroinflammation in a ratmodel ofmultiple sclerosis by enhancingblood–brainbarrierintegrity and by modulating the peripheral immune response. PLoS One 8, e56586.
Bartanusz, V., Jezova, D., Alajajian, B., Digicaylioglu, M., 2011. The blood–spinal cordbarrier: morphology and clinical implications. Ann. Neurol. 70, 194–206.
Bennett, J., Basivireddy, J., Kollar, A., Biron, K.E., Reickmann, P., Jefferies, W.A., McQuaid, S.,2010. Blood–brain barrier disruption and enhanced vascular permeability in themultiple sclerosis model EAE. J. Neuroimmunol. 229, 180–191.
Bever, T.B., Rosenberg, G.A., 1999. Matrix metalloproteinases in multiple sclerosis: targetsof therapy or markers of injury. Neurology 53, 1380–1381.
Brundula, V., Rewcastle, N.B., Metz, L.M., Bernard, C.C., Yong, V.W., 2002. Targetingleukocyte MMPs and transmigration: minocycline as a potential therapy for multiplesclerosis. Brain 125, 1297–1308.
Chen, X., Pi, R., Liu, M., Ma, X., Jiang, Y., Liu, Y., Mao, X., Hu, X., 2010. Combination ofmethylprednisolone andminocycline synergistically improves experimental autoimmuneencephalomyelitis in C57 BL/6 mice. J. Neuroimmunol. 226, 104–109.
Chitnis, T., Khoury, S.J., 2003. Cytokine shifts and tolerance in experimental autoimmuneencephalomyelitis. Immunol. Res. 28, 223–239.
Dubois, B., Leary, S.M., Nelissen, I., Opdenakker, G., Giovannoni, G., Thompson, A.J., 2003.Serum gelatinase B/MMP-9 in primary progressivemultiple sclerosis patients treatedwith interferon-beta-1a. J. Neurol. 250, 1037–1043.
Please cite this article as: Hou, Y., et al., Interferon β-secreting mesenchautoimmune encephalomyelitis, J. Neuroimmunol. (2014), http://dx.doi.o
Elkayam, O., Yaron, M., Caspi, D., 1999. Minocycline-induced autoimmune syndromes: anoverview. Semin. Arthritis Rheum. 28, 392–397.
Giuliani, F., Fu, S.A., Metz, L.M., 2005. Effective combination of minocycline and interferon-betain a model of multiple sclerosis. J. Neuroimmunol. 165, 83–91.
Guo, X., Nakamura, K., Kohyama, K., 2007. Inhibition of glial cell activation ameliorates theseverity of experimental autoimmune encephalomyelitis. Neurosci. Res. 59, 457–466.
Hirase, T., Staddon, J.M., Saitou, M., Ando-Akatsuka, Y., Itoh, M., 1997. Occludin as a possibledeterminant of tight junction permeability in endothelial cells. J. Cell Sci. 110,1603–1613.
Hollborn, M., Wiedemann, P., Bringmann, A., Kohen, L., 2010. Chemotactic and cytotoxiceffects of minocycline on human retinal pigment epithelial cells. Invest. Ophthalmol.Vis. Sci. 51, 2721–2729.
Hou, Y., Ryu, C.H., Park, K.Y., Kim, S.M., Jeong, C.H., Jeun, S.S., 2013. Effective combinationof human bone marrow mesenchymal stem cells and minocycline in experimentalautoimmune encephalomyelitis mice. Stem Cell Res. Ther. 4, 77.
Jeong, C.H., Kim, S.M., Lim, J.Y., Ryu, C.H., Jun, J.A., Jeun, S.S., 2014. Mesenchymal stem cellsexpressing brain-derived neurotrophic factor enhance endogenous neurogenesis inan ischemic stroke model. Biomed. Res. Int. 2014, 129145.
Lee, J.Y., Kim, H.S., Choi, H.Y., 2012. Fluoxetine inhibits matrix metalloprotease activationand prevents disruption of blood–spinal cord barrier after spinal cord injury. Brain135, 2375–2389.
Luccarini, I., Ballerini, C., Biagioli, T., 2008. Combined treatment with atorvastatin andminocycline suppresses severity of EAE. Exp. Neurol. 211, 214–226.
Mankertz, J., Tavalali, S., Schmitz, H., 2000. Expression from the human occludin promoteris affected by tumor necrosis factor alpha and interferon gamma. J. Cell Sci. 113,2085–2090.
Martín-Saavedra, F.M., Flores, N., Dorado, B., Eguiluz, C., Bravo, B., García-Merino, A.,Ballester, S., 2007. Beta-interferon unbalances the peripheral T cell proinflammatoryresponse in experimental autoimmune encephalomyelitis. Mol. Immunol. 44,3597–3607.
Minagar, A., Alexander, J.S., 2003. Blood–brain barrier disruption in multiple sclerosis.Mult. Scler. 9, 540–549.
Niimi, N., Kohyama, K., Matsumoto, Y., 2013. Minocycline suppresses experimentalautoimmune encephalomyelitis by increasing tissue inhibitors of metalloproteinases.Neuropathology 33, 612–620.
O'Garra, A., Steinman, L., Gijbels, K., 1997. CD4+ T-cell subsets in autoimmunity. Curr.Opin. Immunol. 9, 872–883.
Paintlia, A.S., Paintlia, M.K., Singh, I., Skoff, R.B., Singh, A.K., 2009. Combination therapy oflovastatin and rolipram provides neuroprotection and promotes neurorepair ininflammatory demyelination model of multiple sclerosis. Glia 57, 182–193.
Popovic, N., Schubart, A., Goetz, B.D., Zhang, S.C., Linington, C., Duncan, I.D., 2002. Inhibitionof autoimmune encephalomyelitis by a tetracycline. Ann. Neurol. 51, 215–223.
Ransohoff, R.M., Engelhardt, B., 2012. The anatomical and cellular basis of immunesurveillance in the central nervous system. Nat. Rev. Immunol. 12, 623–635.
Rodríguez-Sáinz Mdel, C., Sánchez-Ramón, S., de Andrés, C., Rodríguez-Mahou, M.,Muñoz-Fernández, M.A., 2002. Th1/Th2 cytokine balance and nitric oxide in cerebro-spinal fluid and serum from patients with multiple sclerosis. Eur. Cytokine Netw. 13,110–114.
Ryu, C.H., Park, K.Y., Hou, Y., Jeong, C.H., Kim, S.M., Jeun, S.S., 2013. Gene therapy ofmultiple sclerosis using interferon β-secreting human bone marrow mesenchymalstem cells. Biomed. Res. Int. 2013, 696738.
Schellenberg, A.E., Buist, R., Yong, V.W., 2007. Magnetic resonance imaging of blood–spinalcord barrier disruption in mice with experimental autoimmune encephalomyelitis.Magn. Reson. Med. 58, 298–305.
Sturkenboom, M.C., Meier, C.R., Jick, H., Stricker, B.H., 1999. Minocycline and lupuslikesyndrome in acne patients. Arch. Intern. Med. 159, 493–497.
Tikka, T., Fiebich, B.L., Goldsteins, G., Keinanen, R., Koistinaho, J., 2001. Minocycline, atetracycline derivative, is neuroprotective against excitotoxicity by inhibiting activationand proliferation of microglia. J. Neurosci. 21, 2580–2588.
Yong, V.W., Wells, J., Giuliani, F., Casha, S., Power, C., Metz, L.M., 2004. The promise ofminocycline in neurology. Lancet Neurol. 3, 744–751.
ymal stem cells combined with minocycline attenuate experimentalrg/10.1016/j.jneuroim.2014.06.001

















![Epac2 signaling at the β-cell plasma membrane920771/FULLTEXT01.pdf · small fraction of cells are pancreatic polypeptide-secreting PP-cells [6] and ghrelin-releasing ε-cells [7].](https://static.fdocument.org/doc/165x107/6065b034c80f1b4fbb7d2949/epac2-signaling-at-the-cell-plasma-membrane-920771fulltext01pdf-small-fraction.jpg)