
Construction of amylolytic yeasts secreting xylanase and ...
7
Enzymatic modification of dough during bread making may improve bread quality attributes such as texture and taste. To develop amylolytic Saccharomyces cerevisiae baker’s yeasts producing and secreting xylanase and phytase, two integrative cassettes were constructed to allow co-expression of the Aspergillus awamori glucoamylase gene (GA1) and Debaryomyces castellii phytase gene (phytDc), and the Debaryomyces occidentalis α- amylase gene (AMY) and Bacillus subtilis xylanase gene (xynA), respectively. The recombinant baker’s yeast strain expressing GA1 and phytDc and the corresponding strain expressing AMY and xynA were co-cultured to secrete active glucoamylase and α-amylase, displaying both xylanase and phytase activities that degraded starch and consequently grew using it as the sole carbon source. The glucoamylase and α-amylase activities produced by the co-culture of these two yeast strains increased by 1.5-fold and 11-fold, respectively, relative to those in their monoculture with culture medium containing 2% (w/v) raw wheat starch within 5 days of growth. These new yeasts hydrolyzed 60% of the raw starch content during the same period. Keywords: Saccharomyces cerevisiae, amylases, baker’s yeast, phytase, wheat starch, xylanase Wheat flour used in bread making contains non-starch poly- saccharides, such as hemicelluloses, in relatively small amounts along with starch as the main polysaccharide; it also contains phytate as the major storage form of phosphate (Monfort et al., 1997; Lim et al., 2008). Unfortunately, Saccharomyces cerevisiae, including baker’s yeast and brewer’s yeast, is unable to meta- bolize starch, non-starch polysaccharides, and phytate. Addition of exogenous α-amylase and xylanase that modify starch and non-starch polysaccharides in the dough may delay the rate of staling and increase the volume and shelf-life of bread (Monfort et al., 1996). Further, on consumption, bread made with the addition of glucoamylase generates a melting feeling in the mouth (Saito et al., 1996). Intake of large amounts of foods that are rich in phytate may induce mineral deficiencies and phosphate pollution problems as phytate is an anti-nutritional factor that forms complexes with proteins and nutritionally important minerals. Phytase catalyzes the dephosphorylation of phytate to inositol and inorganic phosphate, thus alleviating phytate-related symptoms (Han et al., 1999). However, exogenous addition of these enzymes during bread making may result in the incidence of allergenic symptoms in bakers (Nieto et al., Korean Journal of Microbiology (2020) Vol. 56, No. 2, pp. 170-176 pISSN 0440-2413 DOI https://doi.org/10.7845/kjm.2020.0007 eISSN 2383-9902 Copyright ⓒ 2020, The Microbiological Society of Korea Construction of amylolytic yeasts secreting xylanase and phytase to improve bread quality Ja-Yeon Lee 1 , Young-Kum Im 2 , Jong-Eon Chin 3 , and Suk Bai 2 * 1 Department of Biological Sciences and Biotechnology Graduate School, Chonnam National University, Gwangju 61186, Republic of Korea 2 Department of Biological Sciences, College of Natural Sciences, Chonnam National University, Gwangju 61186, Republic of Korea 3 Department of Radiology, Donggang University, Gwangju 61200, Republic of Korea 빵의품질향상을위한 xylanase 와 phytase 분비전분분해능효모의개발 이자연 1 ・ 임영금 2 ・ 진종언 3 ・ 배 석 2 * 1 전남대학교 대학원 생물과학・생명기술학과, 2 전남대학교 자연과학대학 생물학과, 3 동강대학교 방사선과 (Received January 31, 2020; Revised March 7, 2020; Accepted March 9, 2020) *For correspondence. E-mail: [email protected]; Tel.: +82-62-530-3412; Fax: +82-62-530-3409
Transcript of Construction of amylolytic yeasts secreting xylanase and ...

Enzymatic modification of dough during bread making may
improve bread quality attributes such as texture and taste. To
develop amylolytic Saccharomyces cerevisiae baker’s yeasts
producing and secreting xylanase and phytase, two integrative
cassettes were constructed to allow co-expression of the Aspergillus
awamori glucoamylase gene (GA1) and Debaryomyces castellii
phytase gene (phytDc), and the Debaryomyces occidentalis α-
amylase gene (AMY) and Bacillus subtilis xylanase gene (xynA),
respectively. The recombinant baker’s yeast strain expressing
GA1 and phytDc and the corresponding strain expressing AMY
and xynA were co-cultured to secrete active glucoamylase and
α-amylase, displaying both xylanase and phytase activities that
degraded starch and consequently grew using it as the sole carbon
source. The glucoamylase and α-amylase activities produced
by the co-culture of these two yeast strains increased by 1.5-fold
and 11-fold, respectively, relative to those in their monoculture
with culture medium containing 2% (w/v) raw wheat starch
within 5 days of growth. These new yeasts hydrolyzed 60% of
the raw starch content during the same period.
Keywords: Saccharomyces cerevisiae, amylases, baker’s yeast,
phytase, wheat starch, xylanase
Wheat flour used in bread making contains non-starch poly-
saccharides, such as hemicelluloses, in relatively small amounts
along with starch as the main polysaccharide; it also contains
phytate as the major storage form of phosphate (Monfort et al.,
1997; Lim et al., 2008). Unfortunately, Saccharomyces cerevisiae,
including baker’s yeast and brewer’s yeast, is unable to meta-
bolize starch, non-starch polysaccharides, and phytate. Addition
of exogenous α-amylase and xylanase that modify starch and
non-starch polysaccharides in the dough may delay the rate of
staling and increase the volume and shelf-life of bread (Monfort et
al., 1996). Further, on consumption, bread made with the
addition of glucoamylase generates a melting feeling in the
mouth (Saito et al., 1996). Intake of large amounts of foods
that are rich in phytate may induce mineral deficiencies and
phosphate pollution problems as phytate is an anti-nutritional
factor that forms complexes with proteins and nutritionally
important minerals. Phytase catalyzes the dephosphorylation
of phytate to inositol and inorganic phosphate, thus alleviating
phytate-related symptoms (Han et al., 1999). However, exogenous
addition of these enzymes during bread making may result in
the incidence of allergenic symptoms in bakers (Nieto et al.,
Korean Journal of Microbiology (2020) Vol. 56, No. 2, pp. 170-176 pISSN 0440-2413DOI https://doi.org/10.7845/kjm.2020.0007 eISSN 2383-9902Copyright ⓒ 2020, The Microbiological Society of Korea
Construction of amylolytic yeasts secreting xylanase and phytase to
improve bread quality
Ja-Yeon Lee1, Young-Kum Im2, Jong-Eon Chin3, and Suk Bai2*
1Department of Biological Sciences and Biotechnology Graduate School, Chonnam National University, Gwangju 61186, Republic
of Korea 2Department of Biological Sciences, College of Natural Sciences, Chonnam National University, Gwangju 61186, Republic of
Korea3Department of Radiology, Donggang University, Gwangju 61200, Republic of Korea
빵의 품질향상을 위한 xylanase와 phytase 분비 전분 분해능 효모의 개발
이자연1 ・ 임영금2 ・ 진종언3 ・ 배 석2*1전남대학교 대학원 생물과학・생명기술학과, 2전남대학교 자연과학대학 생물학과, 3동강대학교 방사선과
(Received January 31, 2020; Revised March 7, 2020; Accepted March 9, 2020)
*For correspondence. E-mail: [email protected];
Tel.: +82-62-530-3412; Fax: +82-62-530-3409

Amylolytic baker’s yeasts secreting xylanase and phytase ∙ 171
Korean Journal of Microbiology, Vol. 56, No. 2
1999). To resolve this problem, several studies have been
conducted to develop baker’s yeasts that express Aspergillus α-
amylase, endoxylanase, and phytase to reduce or eliminate the
requirement for adding exogenous enzymes (Saito et al., 1996;
Monfort et al., 1997; Nieto et al., 1999; Li et al., 2009). In S.
cerevisiae, the usual way to maintain integrated foreign enzyme
genes is to use reiterated DNA sequences including δ-sequences
and ribosomal DNA as target sites for homologous recombination,
which results in multiple copies of the integrated genes, resulting
in higher expression levels and mitotic stability. Furthermore,
the unnecessary bacterial plasmid DNA sequences containing
the antibiotic resistance marker and the replication origin for
integrating foreign enzyme genes into yeast chromosomes should
be excised before transformation because recombinant yeasts
with bread enter our bodies orally (Lee and Da Silva, 1997;
Park et al., 2014). In this study, we constructed two linearized
δ-integrative vectors harboring the Aspergillus awamori raw
starch-degrading glucoamylase gene (GA1) (Lin et al., 1998)
and Debaryomyces castellii phytase gene (phytDc) (Lim et al.,
2008) or the Debaryomyces occidentalis α-amylase gene (AMY)
(Kang et al., 2003) and xylanase gene (xynA) from the GRAS
organism, Bacillus subtilis. This facilitated the development of
a baker’s yeast with glucoamylase and phytase activities, and a
yeast with α-amylase and xylanase activities. The growth, starch
utilization and enzyme activities of these new yeast strains
were analyzed and the efficacy of raw starch hydrolysis by their
co-culture was compared with that by the monoculture.
A baker’s yeast strain, Saccharomyces cerevisiae ATCC 6037
was used in the transformation experiments. Bacillus subtilis
ATCC 6633 was the source of the xynA gene. Escherichia coli
DH5α was used as the host for plasmid constructions. YIpδ
AURSAδ, YIpδAURDpδ, and YIpδAGSAδ (Kang et al., 2003;
Lim et al., 2008; Kim et al., 2010) served as the backbones
for the double δ-integrative system. All DNA manipulations
and the E. coli transformation were conducted using standard
protocols. Integrative transformation of yeast was conducted
using the method described by Gietz et al. (1992). The xylanase
gene was amplified by PCR using the oligonucleotides 5'-TCA
GGTACCATGTTTAAGTTTAAAAAGAAT-3' and 5'-CGAT
CTAGATTACCACACTGTTACGTTAGAACT-3'. These primers
were designed using the published nucleotide sequences of the
B. subtilis xylanase gene (xynA) (GenBank accession number
M36648). PCR resulted in 0.64 kb amplified DNA fragments
of the whole open reading frame from the genomic DNA of B.
subtilis ATCC 6633, which underwent digestion with KpnI and
XbaI, and was then inserted into the same sites downstream of
the ADC1 promoter (ADC1p) in YIpδAURSAδ lacking the AMY
gene, thereby generating YIpδAURXδ (7.2 kb). Furthermore,
to obtain expression of both the xynA and AMY genes, a 2.7 kb
DNA fragment containing the ADC1p-xynA cassette of YIpδ
AURXδ, which underwent digestion with ApaI and SacI, was
inserted using the same sites in YIpδAGSAδ lacking the GA1
gene to generate YIpδXSAδ (9.6 kb). To construct the δ-
integrative system containing the GA1 and phytDc genes, a 2.1
kb DNA fragment harboring the ADC1p-phytDc cassette was
excised from YIpδAURDpδ and inserted into the XbaI sites in
YIpδAGSAδ lacking the ADC1p-AMY cassette, thus generating
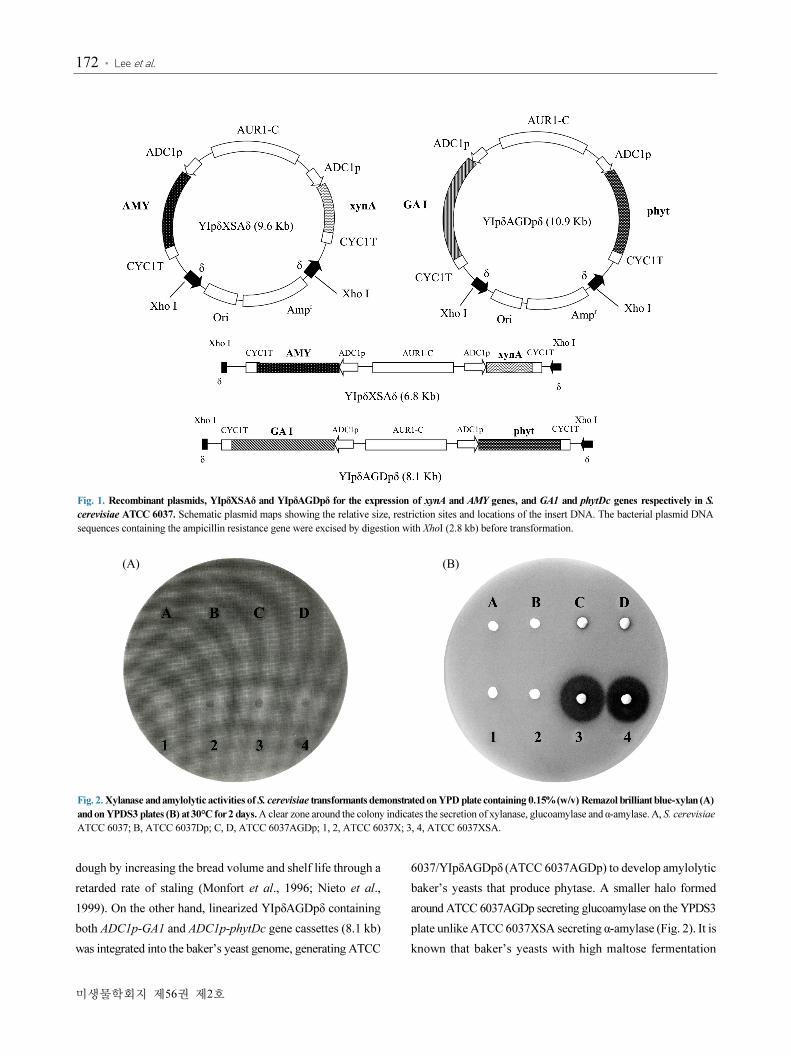
YIpδAGDpδ (10.9 kb) (Fig. 1).
To develop baker’s yeasts producing and secreting xylanase,
YIpδAURXδ was linearized by digesting the δ sequences with
XhoI, after which the fragments (4.4 kb) harboring the ADC1p-
xynA gene cassettes flanked by δ sequences were integrated
into the reiterated δ sequences dispersed throughout the baker’s
yeast’s genome (Lee and Da Silva, 1997; Kang et al., 2003).
The integrative cassette-harboring transformed yeasts were
designated ATCC 6037/YIpδAURXδ (ATCC 6037X). The
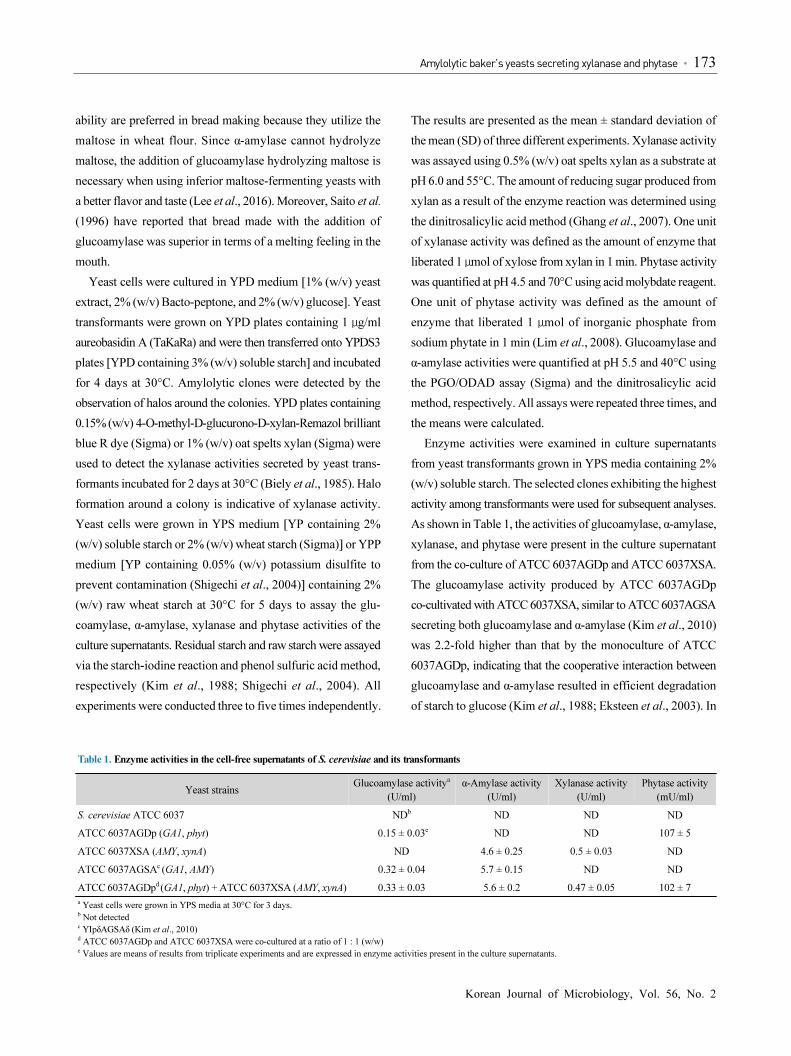
yeast transformants formed halos around their colonies on
YPD plates containing xylan whereas the untransformed baker’s
yeast strain displayed no halo (Fig. 2), indicating that the B.
subtilis xylanase gene is expressed under the control of yeast
ADC1 promoter and that xylanase is secreted under the control
of its own lead sequence in baker’s yeast (Ahn et al., 1992).
Then, linearized YIpδXSAδ (6.8 kb) (Fig. 1) was integrated
into the baker’s yeast genome, generating ATCC 6037/YIpδ
XSAδ that expresses both xynA and AMY genes (ATCC 6037XSA)
to develop amylolytic baker’s yeasts producing xylanase (Monfort
et al., 1997). This double δ integration system generates a
smaller unnecessary fragment (2.8 kb) containing an ampicillin
resistance marker, which was eliminated prior to transformation
(Kang et al., 2003). Large halos developed around ATCC
6037XSA secreting xylanase and α-amylase on YPD plates
containing xylan and YPDS3 plate, respectively (Fig. 2). It
is known that these two enzymes act synergistically on the
polysaccharides in wheat flour, resulting in improved bread
172 ∙ Lee et al.
미생물학회지 제56권 제2호
dough by increasing the bread volume and shelf life through a
retarded rate of staling (Monfort et al., 1996; Nieto et al.,
1999). On the other hand, linearized YIpδAGDpδ containing
both ADC1p-GA1 and ADC1p-phytDc gene cassettes (8.1 kb)
was integrated into the baker’s yeast genome, generating ATCC
6037/YIpδAGDpδ (ATCC 6037AGDp) to develop amylolytic
baker’s yeasts that produce phytase. A smaller halo formed
around ATCC 6037AGDp secreting glucoamylase on the YPDS3
plate unlike ATCC 6037XSA secreting α-amylase (Fig. 2). It is
known that baker’s yeasts with high maltose fermentation
Fig. 1. Recombinant plasmids, YIpδXSAδ and YIpδAGDpδ for the expression of xynA and AMY genes, and GA1 and phytDc genes respectively in S.
cerevisiae ATCC 6037. Schematic plasmid maps showing the relative size, restriction sites and locations of the insert DNA. The bacterial plasmid DNA
sequences containing the ampicillin resistance gene were excised by digestion with XhoI (2.8 kb) before transformation.
(A) (B)
Fig. 2. Xylanase and amylolytic activities of S. cerevisiae transformants demonstrated on YPD plate containing 0.15% (w/v) Remazol brilliant blue-xylan (A)
and on YPDS3 plates (B) at 30°C for 2 days. A clear zone around the colony indicates the secretion of xylanase, glucoamylase and α-amylase. A, S. cerevisiae
ATCC 6037; B, ATCC 6037Dp; C, D, ATCC 6037AGDp; 1, 2, ATCC 6037X; 3, 4, ATCC 6037XSA.
Amylolytic baker’s yeasts secreting xylanase and phytase ∙ 173
Korean Journal of Microbiology, Vol. 56, No. 2
ability are preferred in bread making because they utilize the
maltose in wheat flour. Since α-amylase cannot hydrolyze
maltose, the addition of glucoamylase hydrolyzing maltose is
necessary when using inferior maltose-fermenting yeasts with
a better flavor and taste (Lee et al., 2016). Moreover, Saito et al.
(1996) have reported that bread made with the addition of
glucoamylase was superior in terms of a melting feeling in the
mouth.
Yeast cells were cultured in YPD medium [1% (w/v) yeast
extract, 2% (w/v) Bacto-peptone, and 2% (w/v) glucose]. Yeast
transformants were grown on YPD plates containing 1 μg/ml
aureobasidin A (TaKaRa) and were then transferred onto YPDS3
plates [YPD containing 3% (w/v) soluble starch] and incubated
for 4 days at 30°C. Amylolytic clones were detected by the
observation of halos around the colonies. YPD plates containing
0.15% (w/v) 4-O-methyl-D-glucurono-D-xylan-Remazol brilliant
blue R dye (Sigma) or 1% (w/v) oat spelts xylan (Sigma) were
used to detect the xylanase activities secreted by yeast trans-
formants incubated for 2 days at 30°C (Biely et al., 1985). Halo
formation around a colony is indicative of xylanase activity.
Yeast cells were grown in YPS medium [YP containing 2%
(w/v) soluble starch or 2% (w/v) wheat starch (Sigma)] or YPP
medium [YP containing 0.05% (w/v) potassium disulfite to
prevent contamination (Shigechi et al., 2004)] containing 2%
(w/v) raw wheat starch at 30°C for 5 days to assay the glu-
coamylase, α-amylase, xylanase and phytase activities of the
culture supernatants. Residual starch and raw starch were assayed
via the starch-iodine reaction and phenol sulfuric acid method,
respectively (Kim et al., 1988; Shigechi et al., 2004). All
experiments were conducted three to five times independently.
The results are presented as the mean ± standard deviation of
the mean (SD) of three different experiments. Xylanase activity
was assayed using 0.5% (w/v) oat spelts xylan as a substrate at
pH 6.0 and 55°C. The amount of reducing sugar produced from
xylan as a result of the enzyme reaction was determined using
the dinitrosalicylic acid method (Ghang et al., 2007). One unit
of xylanase activity was defined as the amount of enzyme that
liberated 1 μmol of xylose from xylan in 1 min. Phytase activity
was quantified at pH 4.5 and 70°C using acid molybdate reagent.
One unit of phytase activity was defined as the amount of
enzyme that liberated 1 μmol of inorganic phosphate from
sodium phytate in 1 min (Lim et al., 2008). Glucoamylase and
α-amylase activities were quantified at pH 5.5 and 40°C using
the PGO/ODAD assay (Sigma) and the dinitrosalicylic acid
method, respectively. All assays were repeated three times, and
the means were calculated.
Enzyme activities were examined in culture supernatants
from yeast transformants grown in YPS media containing 2%
(w/v) soluble starch. The selected clones exhibiting the highest
activity among transformants were used for subsequent analyses.
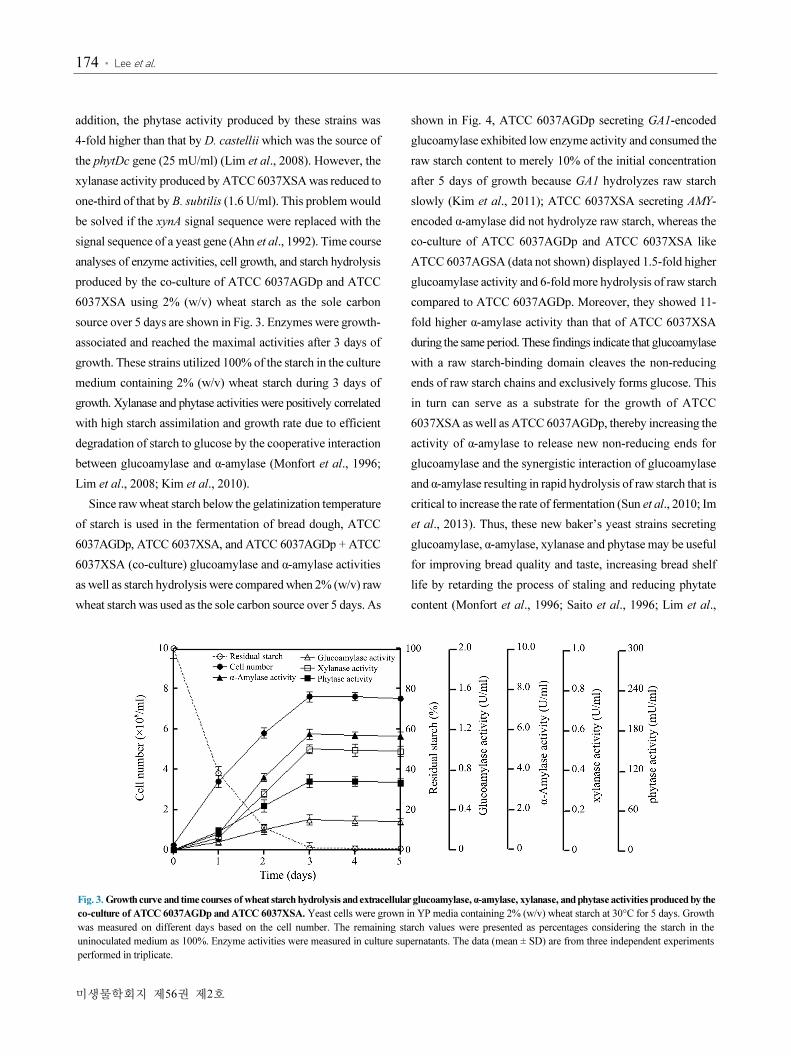
As shown in Table 1, the activities of glucoamylase, α-amylase,
xylanase, and phytase were present in the culture supernatant
from the co-culture of ATCC 6037AGDp and ATCC 6037XSA.
The glucoamylase activity produced by ATCC 6037AGDp
co-cultivated with ATCC 6037XSA, similar to ATCC 6037AGSA
secreting both glucoamylase and α-amylase (Kim et al., 2010)
was 2.2-fold higher than that by the monoculture of ATCC
6037AGDp, indicating that the cooperative interaction between
glucoamylase and α-amylase resulted in efficient degradation
of starch to glucose (Kim et al., 1988; Eksteen et al., 2003). In
Table 1. Enzyme activities in the cell-free supernatants of S. cerevisiae and its transformants
Yeast strainsGlucoamylase activitya
(U/ml)
α-Amylase activity
(U/ml)
Xylanase activity
(U/ml)
Phytase activity
(mU/ml)
S. cerevisiae ATCC 6037 NDb ND ND ND
ATCC 6037AGDp (GA1, phyt) 0.15 ± 0.03e ND ND 107 ± 5
ATCC 6037XSA (AMY, xynA) ND 4.6 ± 0.25 0.5 ± 0.03 ND
ATCC 6037AGSAc (GA1, AMY) 0.32 ± 0.04 5.7 ± 0.15 ND ND
ATCC 6037AGDpd (GA1, phyt) + ATCC 6037XSA (AMY, xynA) 0.33 ± 0.03 5.6 ± 0.2 0.47 ± 0.05 102 ± 7
a Yeast cells were grown in YPS media at 30°C for 3 days.b Not detectedc YIpδAGSAδ (Kim et al., 2010)d ATCC 6037AGDp and ATCC 6037XSA were co-cultured at a ratio of 1 : 1 (w/w) e Values are means of results from triplicate experiments and are expressed in enzyme activities present in the culture supernatants.
174 ∙ Lee et al.
미생물학회지 제56권 제2호
addition, the phytase activity produced by these strains was
4-fold higher than that by D. castellii which was the source of
the phytDc gene (25 mU/ml) (Lim et al., 2008). However, the
xylanase activity produced by ATCC 6037XSA was reduced to
one-third of that by B. subtilis (1.6 U/ml). This problem would
be solved if the xynA signal sequence were replaced with the
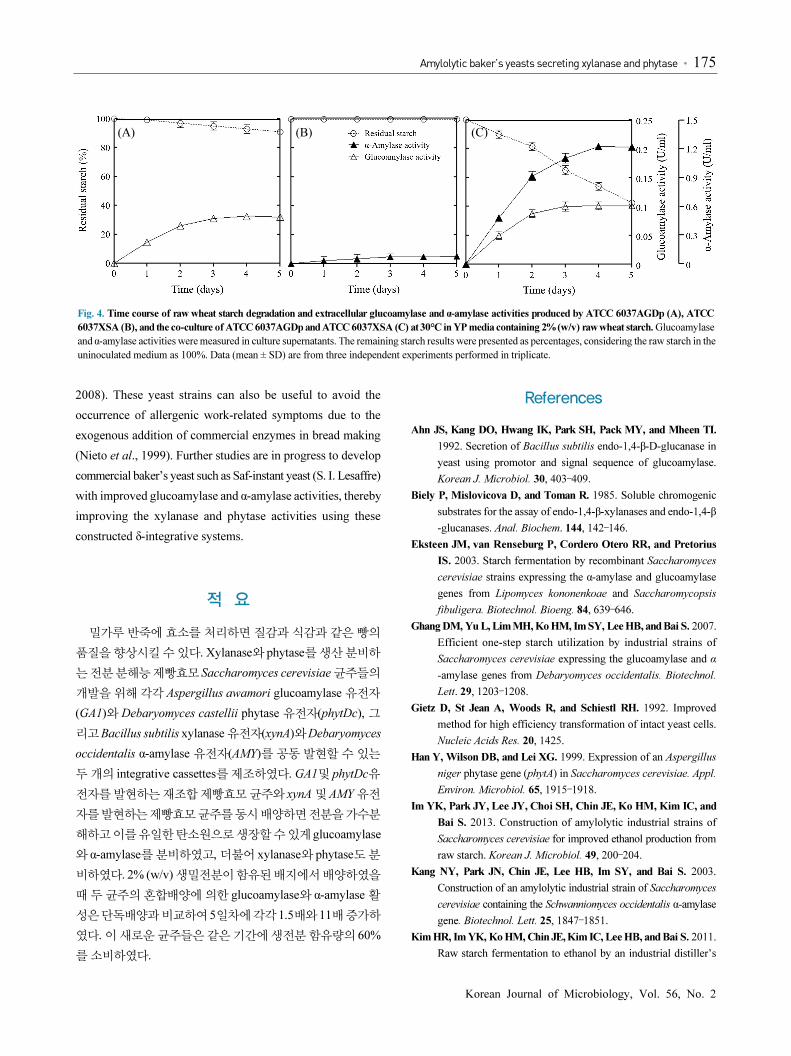
signal sequence of a yeast gene (Ahn et al., 1992). Time course
analyses of enzyme activities, cell growth, and starch hydrolysis
produced by the co-culture of ATCC 6037AGDp and ATCC
6037XSA using 2% (w/v) wheat starch as the sole carbon
source over 5 days are shown in Fig. 3. Enzymes were growth-
associated and reached the maximal activities after 3 days of
growth. These strains utilized 100% of the starch in the culture
medium containing 2% (w/v) wheat starch during 3 days of
growth. Xylanase and phytase activities were positively correlated
with high starch assimilation and growth rate due to efficient
degradation of starch to glucose by the cooperative interaction
between glucoamylase and α-amylase (Monfort et al., 1996;
Lim et al., 2008; Kim et al., 2010).
Since raw wheat starch below the gelatinization temperature
of starch is used in the fermentation of bread dough, ATCC
6037AGDp, ATCC 6037XSA, and ATCC 6037AGDp + ATCC
6037XSA (co-culture) glucoamylase and α-amylase activities
as well as starch hydrolysis were compared when 2% (w/v) raw
wheat starch was used as the sole carbon source over 5 days. As
shown in Fig. 4, ATCC 6037AGDp secreting GA1-encoded
glucoamylase exhibited low enzyme activity and consumed the
raw starch content to merely 10% of the initial concentration
after 5 days of growth because GA1 hydrolyzes raw starch
slowly (Kim et al., 2011); ATCC 6037XSA secreting AMY-
encoded α-amylase did not hydrolyze raw starch, whereas the
co-culture of ATCC 6037AGDp and ATCC 6037XSA like
ATCC 6037AGSA (data not shown) displayed 1.5-fold higher
glucoamylase activity and 6-fold more hydrolysis of raw starch
compared to ATCC 6037AGDp. Moreover, they showed 11-
fold higher α-amylase activity than that of ATCC 6037XSA
during the same period. These findings indicate that glucoamylase
with a raw starch-binding domain cleaves the non-reducing
ends of raw starch chains and exclusively forms glucose. This
in turn can serve as a substrate for the growth of ATCC
6037XSA as well as ATCC 6037AGDp, thereby increasing the
activity of α-amylase to release new non-reducing ends for
glucoamylase and the synergistic interaction of glucoamylase
and α-amylase resulting in rapid hydrolysis of raw starch that is
critical to increase the rate of fermentation (Sun et al., 2010; Im
et al., 2013). Thus, these new baker’s yeast strains secreting
glucoamylase, α-amylase, xylanase and phytase may be useful
for improving bread quality and taste, increasing bread shelf
life by retarding the process of staling and reducing phytate
content (Monfort et al., 1996; Saito et al., 1996; Lim et al.,
Fig. 3. Growth curve and time courses of wheat starch hydrolysis and extracellular glucoamylase, α-amylase, xylanase, and phytase activities produced by the
co-culture of ATCC 6037AGDp and ATCC 6037XSA. Yeast cells were grown in YP media containing 2% (w/v) wheat starch at 30°C for 5 days. Growth
was measured on different days based on the cell number. The remaining starch values were presented as percentages considering the starch in the
uninoculated medium as 100%. Enzyme activities were measured in culture supernatants. The data (mean ± SD) are from three independent experiments
performed in triplicate.
Amylolytic baker’s yeasts secreting xylanase and phytase ∙ 175
Korean Journal of Microbiology, Vol. 56, No. 2
2008). These yeast strains can also be useful to avoid the
occurrence of allergenic work-related symptoms due to the
exogenous addition of commercial enzymes in bread making
(Nieto et al., 1999). Further studies are in progress to develop
commercial baker’s yeast such as Saf-instant yeast (S. I. Lesaffre)
with improved glucoamylase and α-amylase activities, thereby
improving the xylanase and phytase activities using these
constructed δ-integrative systems.
적 요
밀가루 반죽에 효소를 처리하면 질감과 식감과 같은 빵의
품질을 향상시킬 수 있다. Xylanase와 phytase를 생산 분비하
는 전분 분해능 제빵효모 Saccharomyces cerevisiae 균주들의
개발을 위해 각각 Aspergillus awamori glucoamylase 유전자
(GA1)와 Debaryomyces castellii phytase 유전자(phytDc), 그
리고 Bacillus subtilis xylanase 유전자(xynA)와 Debaryomyces
occidentalis α-amylase 유전자(AMY)를 공동 발현할 수 있는
두 개의 integrative cassettes를 제조하였다. GA1및 phytDc유
전자를 발현하는 재조합 제빵효모 균주와 xynA 및 AMY 유전
자를 발현하는 제빵효모 균주를 동시 배양하면 전분을 가수분
해하고 이를 유일한 탄소원으로 생장할 수 있게 glucoamylase
와 α-amylase를 분비하였고, 더불어 xylanase와 phytase도 분
비하였다. 2% (w/v) 생밀전분이 함유된 배지에서 배양하였을
때 두 균주의 혼합배양에 의한 glucoamylase와 α-amylase 활
성은 단독배양과 비교하여 5일차에 각각 1.5배와 11배 증가하
였다. 이 새로운 균주들은 같은 기간에 생전분 함유량의 60%
를 소비하였다.
References
Ahn JS, Kang DO, Hwang IK, Park SH, Pack MY, and Mheen TI.
1992. Secretion of Bacillus subtilis endo-1,4-β-D-glucanase in
yeast using promotor and signal sequence of glucoamylase.
Korean J. Microbiol. 30, 403–409.
Biely P, Mislovicova D, and Toman R. 1985. Soluble chromogenic
substrates for the assay of endo-1,4-β-xylanases and endo-1,4-β
-glucanases. Anal. Biochem. 144, 142–146.
Eksteen JM, van Renseburg P, Cordero Otero RR, and Pretorius
IS. 2003. Starch fermentation by recombinant Saccharomyces
cerevisiae strains expressing the α-amylase and glucoamylase
genes from Lipomyces kononenkoae and Saccharomycopsis
fibuligera. Biotechnol. Bioeng. 84, 639–646.
Ghang DM, Yu L, Lim MH, Ko HM, Im SY, Lee HB, and Bai S. 2007.
Efficient one-step starch utilization by industrial strains of
Saccharomyces cerevisiae expressing the glucoamylase and α
-amylase genes from Debaryomyces occidentalis. Biotechnol.
Lett. 29, 1203–1208.
Gietz D, St Jean A, Woods R, and Schiestl RH. 1992. Improved
method for high efficiency transformation of intact yeast cells.
Nucleic Acids Res. 20, 1425.
Han Y, Wilson DB, and Lei XG. 1999. Expression of an Aspergillus
niger phytase gene (phytA) in Saccharomyces cerevisiae. Appl.
Environ. Microbiol. 65, 1915–1918.
Im YK, Park JY, Lee JY, Choi SH, Chin JE, Ko HM, Kim IC, and
Bai S. 2013. Construction of amylolytic industrial strains of
Saccharomyces cerevisiae for improved ethanol production from
raw starch. Korean J. Microbiol. 49, 200–204.
Kang NY, Park JN, Chin JE, Lee HB, Im SY, and Bai S. 2003.
Construction of an amylolytic industrial strain of Saccharomyces
cerevisiae containing the Schwanniomyces occidentalis α-amylase
gene. Biotechnol. Lett. 25, 1847–1851.
Kim HR, Im YK, Ko HM, Chin JE, Kim IC, Lee HB, and Bai S. 2011.
Raw starch fermentation to ethanol by an industrial distiller’s
(A) (B) (C)
Fig. 4. Time course of raw wheat starch degradation and extracellular glucoamylase and α-amylase activities produced by ATCC 6037AGDp (A), ATCC
6037XSA (B), and the co-culture of ATCC 6037AGDp and ATCC 6037XSA (C) at 30°C in YP media containing 2% (w/v) raw wheat starch. Glucoamylase
and α-amylase activities were measured in culture supernatants. The remaining starch results were presented as percentages, considering the raw starch in the
uninoculated medium as 100%. Data (mean ± SD) are from three independent experiments performed in triplicate.

176 ∙ Lee et al.
미생물학회지 제56권 제2호
strain of Saccharomyces cerevisiae expressing glucoamylase
and α-amylase genes. Biotechnol. Lett. 33, 1643–1648
Kim JH, Kim HR, Lim MH, Ko HM, Chin JE, Lee HB, Kim IC, and Bai
S. 2010. Construction of a direct starch-fermenting industrial
strain of Saccharomyces cerevisiae producing glucoamylase, α
-amylase and debranching enzyme. Biotechnol. Lett. 32, 713–
719.
Kim K, Park CS, and Mattoon JR. 1988. High-efficiency, one-step
utilization by transformed Saccharomyces cerevisiae cells which
secrete both yeast glucoamylase and mouse α-amylase. Appl.
Environ. Microbiol. 54, 966–971.
Lee JY, Chin JE, and Bai S. 2016. Construction of the recombinant
yeast strain with transformation of rice starch-saccharification
enzymes and its alcohol fermentation. Korean J. Microbiol. 52,
220–225.
Lee FWF and Da Silva NA. 1997. Improved efficiency and stability of
multiple cloned gene insertions at the δ sequences of Saccharomyces
cerevisiae. Appl. Microbiol. Biotechnol. 48, 339–345.
Li G, Yang S, Li M, Qiao Y, and Wang J. 2009. Functional analysis of
an Aspergillus ficuum phytase gene in Saccharomyces cerevisiae
and its root-specific, secretory expression in transgenic soybean
plants. Biotechnol. Lett. 31, 1297–1303.
Lim MH, Lee OH, Chin JE, Ko HM, Kim IC, Lee HB, Im SY, and Bai
S. 2008. Simultaneous degradation of phytic acid and starch by
an industrial strain of Saccharomyces cerevisiae producing
phytase and α-amylase. Biotechnol. Lett. 30, 2125–2130.
Lin LL, Ma YJ, Chien HR, and Hsu WH. 1998. Construction of an
amylolytic yeast by multiple integration of the Aspergillus
awamori glucoamylase gene into a Saccharomyces cerevisiae
chromosome. Enzyme Microb. Technol. 23, 360–365.
Monfort A, Blasco A, Prieto JA, and Sanz P. 1996. Combined
expression of Aspergillus nidulans endoxylanase X24 and
Aspergillus oryzae α-amylase in industrial baker’s yeasts and
their use in bread making. Appl. Environ. Microbiol. 62, 3712–
3715.
Monfort A, Blasco A, Prieto, JA, and Sanz P. 1997. Construction of
baker’s yeast strains that secrete different xylanolytic enzymes
and their use in bread making. J. Cereal Sci. 26, 195–199.
Nieto A, Prieto JA, and Sanz P. 1999. Stable high-copy number
integration of Aspergillus orizae α-amylase cDNA in an industrial
baker's yeast strain. Biotechnol. Prog. 15, 459–466.
Park JY, Lee JY, Choi SH, Ko HM, Chin JE, Kim IC, Lee HB, and Bai
S. 2014. Construction of dextrin and isomaltose-assimilating
brewer’s yeasts for production of low-carbohydrate beer. Biotechnol.
Lett. 36, 1693–1699.
Saito S, Mieno Y, Nagashima T, Kumagai C, and Kitamoto K. 1996.
Breeding of a new type of baker’s yeast by δ-integration for
overproduction of glucoamylase using a homothallic yeast. J.
Ferment. Bioeng. 81, 98–103.
Shigechi H, Koh J, Fujita Y, Matsumoto T, Bito Y, Ueda M, Satoh E,
Fukuda H, and Kondo A. 2004. Direct production of ethanol
from raw corn starch via fermentation by use of a novel surface-
engineered yeast strain codisplaying glucoamylase and α-amylase.
Appl. Environ. Microbiol. 70, 5037–5040.
Sun H, Zhao P, Ge X, Xia Y, Hao Z, Liu J, and Peng M. 2010. Recent
advances in microbial raw starch degrading enzymes. Appl.
Biochem. Biotechnol. 160, 988–1003.
















![Epac2 signaling at the β-cell plasma membrane920771/FULLTEXT01.pdf · small fraction of cells are pancreatic polypeptide-secreting PP-cells [6] and ghrelin-releasing ε-cells [7].](https://static.fdocument.org/doc/165x107/6065b034c80f1b4fbb7d2949/epac2-signaling-at-the-cell-plasma-membrane-920771fulltext01pdf-small-fraction.jpg)