Integrin β1 Is Required for Dermal Homeostasis
8
Integrin b1 Is Required for Dermal Homeostasis Shangxi Liu 1 and Andrew Leask 1,2 Integrins are heterodimers that mediate cell attachment to the extracellular matrix. Previously, we used fibroblast- specific knockout mice to show that the integrin b1 subunit was required in vivo for dermal fibrogenesis and cutaneous tissue repair. Here, we show that integrin b1 expression by fibroblasts is required for dermal homeostasis; at 8 weeks after deletion of integrin b1, the dermis is significantly thinner, expressing less collagen and displaying reduced generation of reactive oxygen species (ROS). Cultured integrin b1–deficient fibroblasts show reduced rac1 activation and ROS generation. Overexpressing rac1 in integrin b1–deficient fibroblasts restored ROS generation; adding hydrogen peroxide to integrin b1–deficient fibroblasts restored Col1a2 (collagen, type I, a2) and a-smooth muscle actin (a-SMA) mRNA expression and a-SMA protein expression and stress fiber formation. Thus integrin b1 expression by fibroblasts is required for the maintenance of dermal connective tissue via a rac/ROS pathway. Journal of Investigative Dermatology (2013) 133, 899–906; doi:10.1038/jid.2012.438; published online 29 November 2012 INTRODUCTION The mammalian skin comprises two primary layers, the epidermis and the dermis. The epidermis, the outermost layer, serves as a protective barrier. The dermis, the layer below the epidermis, consists of highly elastic soft connective tissue that not only cushions the body from stress and strain but also provides the epidermis with mechanical support. The dermal layer closest to the epidermis is called the papillary (or upper) dermis, which has thin collagen fibers. Below that, and above subcutaneous fat, is the reticular (or deep) dermis, which has dense collagen fibers. Although tissues exist under a mechan- ical tension, resident dermal fibroblasts are protected from external loads by the mechanical properties of the surrounding matrix and are thus considered to be ‘‘stress-shielded’’ by the extracellular matrix (ECM) that they deposit and remodel (Tomasek et al., 2002). In response to tissue injury, this protective structure is lost and fibroblasts begin to acquire contractile stress fibers that are first composed of cytoplasmic actins that are localized at adhesive contacts at specific regions within the plasma membrane (i.e., focal adhesions; Hinz and Gabbiani, 2003; Hinz, 2010). In response to mechanical strain and growth factors such as transforming growth factor-b, fibroblasts differentiate into myofibroblasts, which are a hallmark of both granulation tissue and fibrocontractive diseases (Hinz, 2010). Myofibroblasts contain stress fibers comprising a-smooth muscle actin (a-SMA) that are connected to the ECM via developed, complex, ‘‘super-mature’’ focal adhesions (Hinz, 2010). Cell attachment to ECM via focal adhesions is essential for cell migration and ECM contraction, which are key features of tissue repair and fibrosis (Eckes et al., 2000). The essential cell surface ECM receptors that cluster within focal adhesions in response to mechanical stress are the integrins. Integrins, heterodimers of an a- and a b-subunit, are well characterized as to their specificity for different ECM components (Hynes, 1992). The following are integrin heterodimers and are known to be collagen receptors: a1b1, a2b1, a3b1, a10b1, and a11b1 (Gullberg et al., 1992; Yamamoto et al., 1995; Camper et al., 1998; Gardner, 1999; Tiger et al., 2001; Barczyk et al., 2010). Integrin b1 is overexpressed in fibrotic fibroblasts isolated from scars of diffuse scleroderma patients, and is required for the excessive adhesive and contractile phenotype of these cells (Shi-Wen et al., 2007). Integrin b1 knockout mice have an embryonic lethal phenotype (Fassler and Meyer, 1995; Stephens et al., 1995). To circumvent this issue, we have generated mice in which b1 integrin has been deleted in adult fibroblasts (Liu et al., 2009a, 2010). These mice are resistant to bleomycin- induced skin scleroderma (Liu et al., 2009a). Moreover, these mice display delayed cutaneous tissue repair and reduced collagen and a-SMA expression, associated with a failure to activate latent transforming growth factor-b1 in response to ECM contraction (Liu et al., 2010). Collectively, these data indicate a role for b1 integrin in mediating tissue repair and fibrosis in the dermis. Numerous reports using epithelial cells have implicated integrins, integrin-associated proteins, or downstream integrin effectors in the regulation of cellular processes other than ORIGINAL ARTICLE 1 Department of Dentistry, Schulich School of Medicine and Dentistry, University of Western Ontario, London, Ontario, Canada and 2 Department of Physiology and Pharmacology, Schulich School of Medicine and Dentistry, University of Western Ontario, London, Ontario, Canada Correspondence: Andrew Leask, Department of Dentistry, Physiology, and Pharmacology, Schulich School of Medicine and Dentistry, University of Western Ontario, Dental Sciences Building, London, Ontario, Canada, N6A 5C1. E-mail: [email protected] Received 18 June 2012; revised 11 September 2012; accepted 5 October 2012; published online 29 November 2012 Abbreviations: DHE, dihydroethidium; ECM, extracellular matrix; ROS, reactive oxygen species; a-SMA, a-smooth muscle actin & 2013 The Society for Investigative Dermatology www.jidonline.org 899
Transcript of Integrin β1 Is Required for Dermal Homeostasis

Integrin b1 Is Required for Dermal HomeostasisShangxi Liu1 and Andrew Leask1,2
Integrins are heterodimers that mediate cell attachment to the extracellular matrix. Previously, we used fibroblast-specific knockout mice to show that the integrin b1 subunit was required in vivo for dermal fibrogenesis andcutaneous tissue repair. Here, we show that integrin b1 expression by fibroblasts is required for dermalhomeostasis; at 8 weeks after deletion of integrin b1, the dermis is significantly thinner, expressing less collagenand displaying reduced generation of reactive oxygen species (ROS). Cultured integrin b1–deficient fibroblastsshow reduced rac1 activation and ROS generation. Overexpressing rac1 in integrin b1–deficient fibroblastsrestored ROS generation; adding hydrogen peroxide to integrin b1–deficient fibroblasts restored Col1a2(collagen, type I, a2) and a-smooth muscle actin (a-SMA) mRNA expression and a-SMA protein expression andstress fiber formation. Thus integrin b1 expression by fibroblasts is required for the maintenance of dermalconnective tissue via a rac/ROS pathway.
Journal of Investigative Dermatology (2013) 133, 899–906; doi:10.1038/jid.2012.438; published online 29 November 2012
INTRODUCTIONThe mammalian skin comprises two primary layers, theepidermis and the dermis. The epidermis, the outermost layer,serves as a protective barrier. The dermis, the layer below theepidermis, consists of highly elastic soft connective tissue thatnot only cushions the body from stress and strain but alsoprovides the epidermis with mechanical support. The dermallayer closest to the epidermis is called the papillary (or upper)dermis, which has thin collagen fibers. Below that, and abovesubcutaneous fat, is the reticular (or deep) dermis, which hasdense collagen fibers. Although tissues exist under a mechan-ical tension, resident dermal fibroblasts are protected fromexternal loads by the mechanical properties of the surroundingmatrix and are thus considered to be ‘‘stress-shielded’’ by theextracellular matrix (ECM) that they deposit and remodel(Tomasek et al., 2002). In response to tissue injury, thisprotective structure is lost and fibroblasts begin to acquirecontractile stress fibers that are first composed of cytoplasmicactins that are localized at adhesive contacts at specificregions within the plasma membrane (i.e., focal adhesions;Hinz and Gabbiani, 2003; Hinz, 2010). In response tomechanical strain and growth factors such as transforminggrowth factor-b, fibroblasts differentiate into myofibroblasts,
which are a hallmark of both granulation tissue andfibrocontractive diseases (Hinz, 2010). Myofibroblastscontain stress fibers comprising a-smooth muscle actin(a-SMA) that are connected to the ECM via developed,complex, ‘‘super-mature’’ focal adhesions (Hinz, 2010).Cell attachment to ECM via focal adhesions is essential forcell migration and ECM contraction, which are key features oftissue repair and fibrosis (Eckes et al., 2000).
The essential cell surface ECM receptors that cluster withinfocal adhesions in response to mechanical stress are theintegrins. Integrins, heterodimers of an a- and a b-subunit,are well characterized as to their specificity for different ECMcomponents (Hynes, 1992). The following are integrinheterodimers and are known to be collagen receptors: a1b1,a2b1, a3b1, a10b1, and a11b1 (Gullberg et al., 1992;Yamamoto et al., 1995; Camper et al., 1998; Gardner,1999; Tiger et al., 2001; Barczyk et al., 2010). Integrin b1 isoverexpressed in fibrotic fibroblasts isolated from scars ofdiffuse scleroderma patients, and is required for the excessiveadhesive and contractile phenotype of these cells (Shi-Wenet al., 2007). Integrin b1 knockout mice have an embryoniclethal phenotype (Fassler and Meyer, 1995; Stephens et al.,1995). To circumvent this issue, we have generated mice inwhich b1 integrin has been deleted in adult fibroblasts (Liuet al., 2009a, 2010). These mice are resistant to bleomycin-induced skin scleroderma (Liu et al., 2009a). Moreover, thesemice display delayed cutaneous tissue repair and reducedcollagen and a-SMA expression, associated with a failure toactivate latent transforming growth factor-b1 in response toECM contraction (Liu et al., 2010). Collectively, these dataindicate a role for b1 integrin in mediating tissue repair andfibrosis in the dermis.
Numerous reports using epithelial cells have implicatedintegrins, integrin-associated proteins, or downstream integrineffectors in the regulation of cellular processes other than
ORIGINAL ARTICLE
1Department of Dentistry, Schulich School of Medicine and Dentistry,University of Western Ontario, London, Ontario, Canada and 2Department ofPhysiology and Pharmacology, Schulich School of Medicine and Dentistry,University of Western Ontario, London, Ontario, Canada
Correspondence: Andrew Leask, Department of Dentistry, Physiology, andPharmacology, Schulich School of Medicine and Dentistry, University ofWestern Ontario, Dental Sciences Building, London, Ontario, Canada,N6A 5C1. E-mail: [email protected]
Received 18 June 2012; revised 11 September 2012; accepted 5 October2012; published online 29 November 2012
Abbreviations: DHE, dihydroethidium; ECM, extracellular matrix; ROS,reactive oxygen species; a-SMA, a-smooth muscle actin
& 2013 The Society for Investigative Dermatology www.jidonline.org 899
adhesion (Brakebusch and Fassler, 2005; Watt and Fujiwara,2011). Although the role of integrins in epidermal homeostasishas been well established; the role of integrins, includingintegrin b1, in dermal homeostasis is almost wholly unknown(Brakebusch and Fassler, 2005; Watt and Fujiwara, 2011).Moreover, the signaling pathways operating downstream ofintegrin b1 in fibroblasts have yet to be fully elucidated.
Here, we use mice deleted for b1 integrin in fibroblasts toassess the role of b1 integrin in dermal homeostasis. More-over, we use fibroblasts derived from these mice to investigatethe signaling mechanisms operating downstream of b1 integ-rin in fibroblasts. Our results provide useful insights into therole that b1 integrin has in maintaining connective tissue.
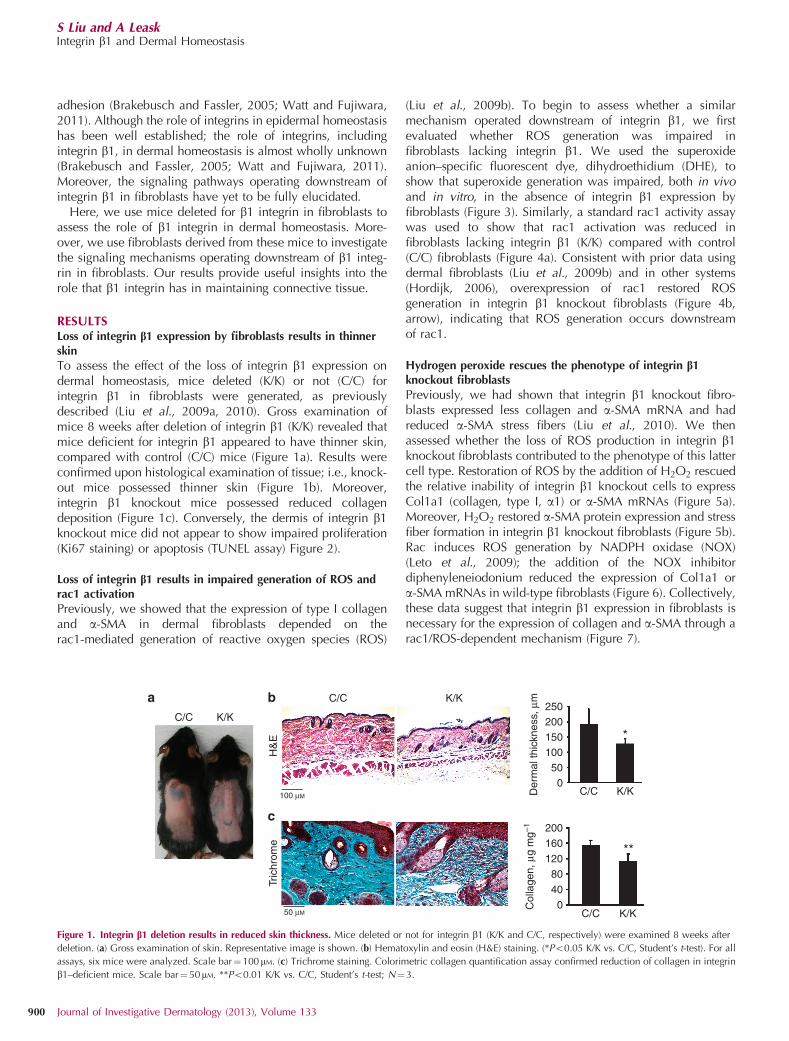
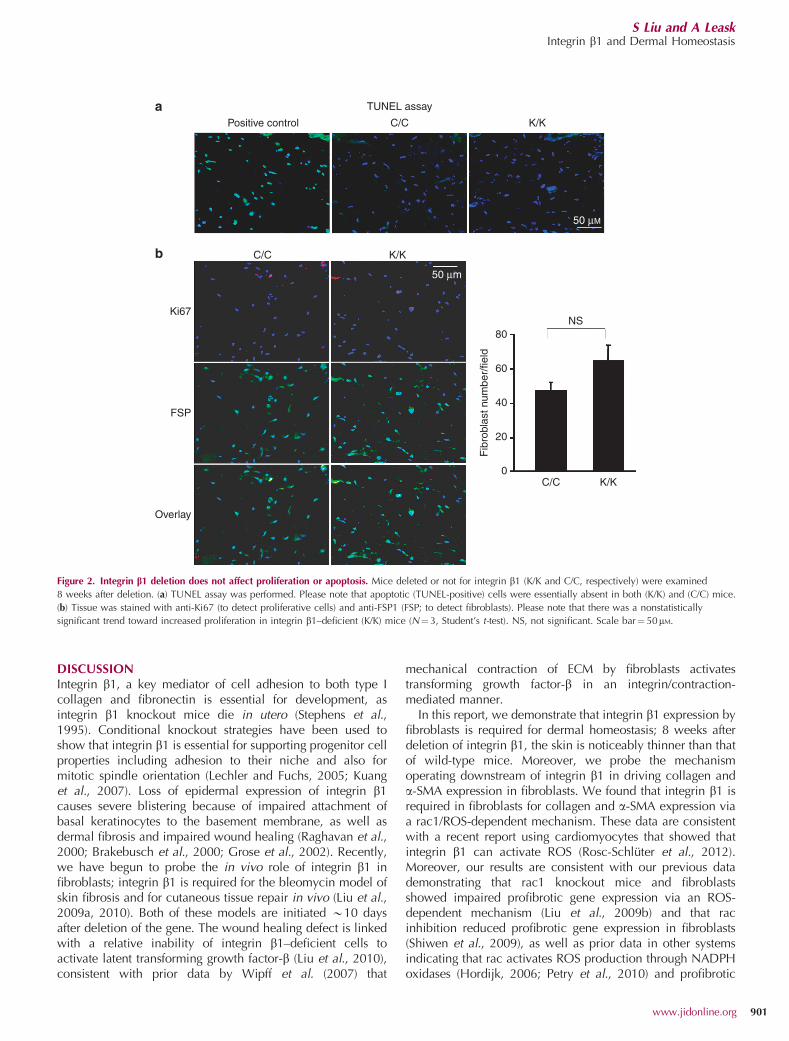
RESULTSLoss of integrin b1 expression by fibroblasts results in thinnerskinTo assess the effect of the loss of integrin b1 expression ondermal homeostasis, mice deleted (K/K) or not (C/C) forintegrin b1 in fibroblasts were generated, as previouslydescribed (Liu et al., 2009a, 2010). Gross examination ofmice 8 weeks after deletion of integrin b1 (K/K) revealed thatmice deficient for integrin b1 appeared to have thinner skin,compared with control (C/C) mice (Figure 1a). Results wereconfirmed upon histological examination of tissue; i.e., knock-out mice possessed thinner skin (Figure 1b). Moreover,integrin b1 knockout mice possessed reduced collagendeposition (Figure 1c). Conversely, the dermis of integrin b1knockout mice did not appear to show impaired proliferation(Ki67 staining) or apoptosis (TUNEL assay) Figure 2).
Loss of integrin b1 results in impaired generation of ROS andrac1 activation
Previously, we showed that the expression of type I collagenand a-SMA in dermal fibroblasts depended on therac1-mediated generation of reactive oxygen species (ROS)
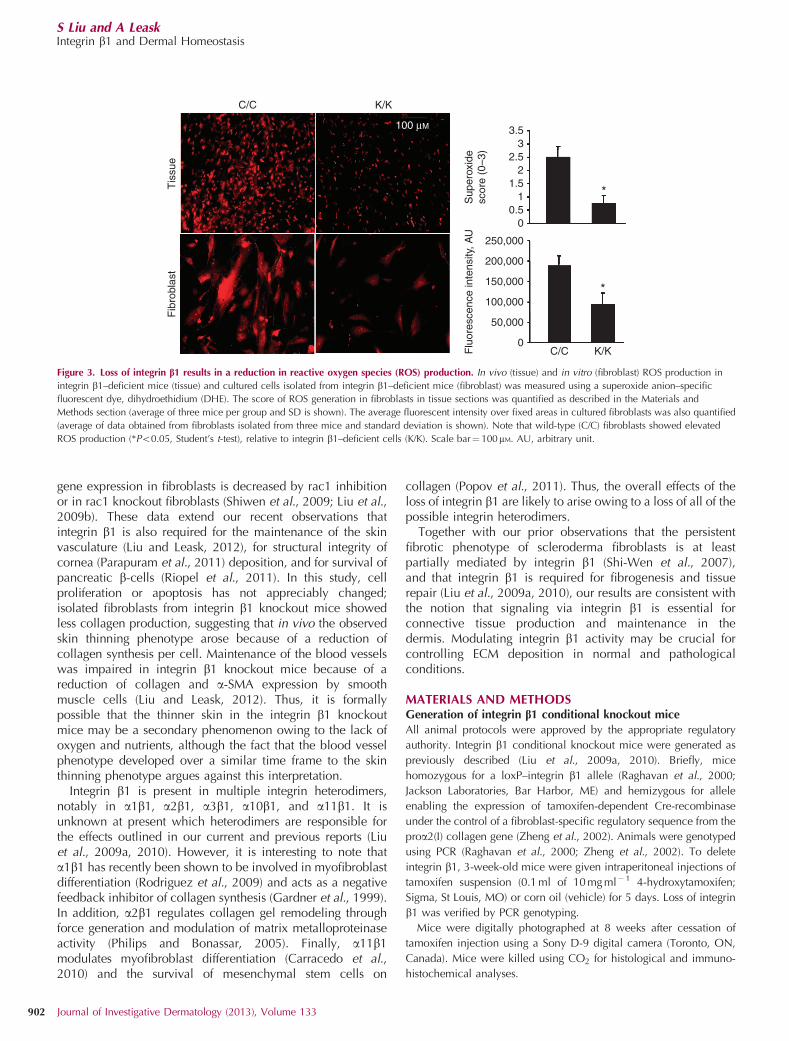
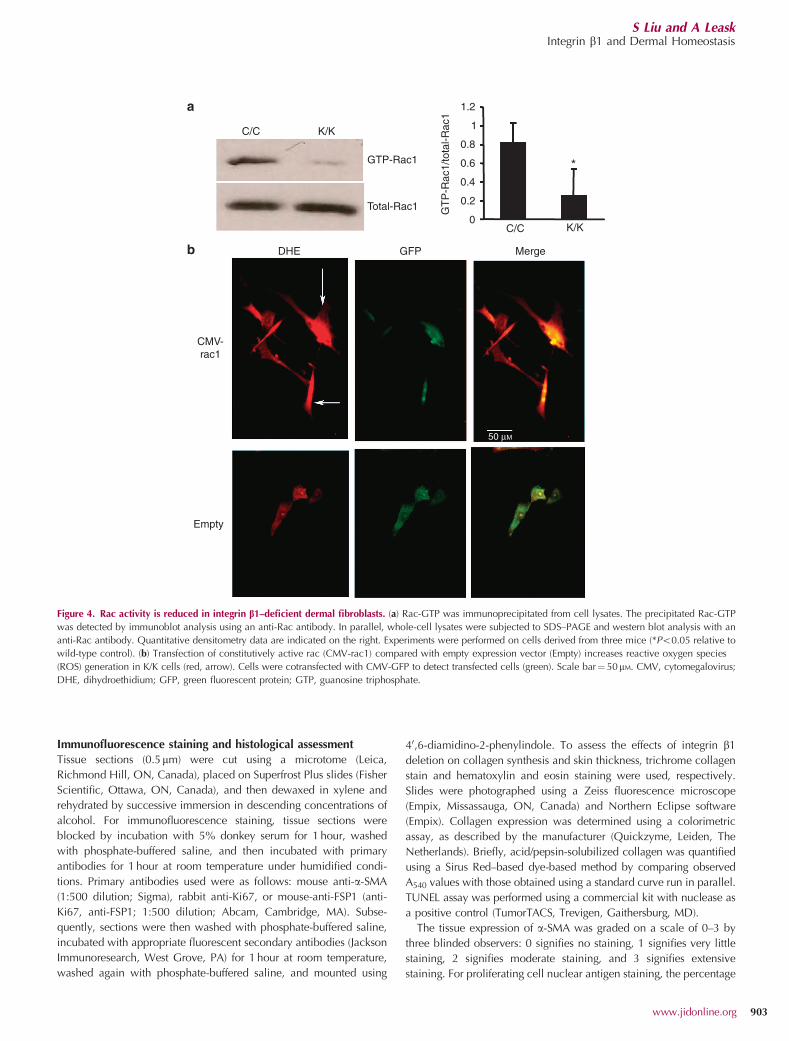
(Liu et al., 2009b). To begin to assess whether a similarmechanism operated downstream of integrin b1, we firstevaluated whether ROS generation was impaired infibroblasts lacking integrin b1. We used the superoxideanion–specific fluorescent dye, dihydroethidium (DHE), toshow that superoxide generation was impaired, both in vivoand in vitro, in the absence of integrin b1 expression byfibroblasts (Figure 3). Similarly, a standard rac1 activity assaywas used to show that rac1 activation was reduced infibroblasts lacking integrin b1 (K/K) compared with control(C/C) fibroblasts (Figure 4a). Consistent with prior data usingdermal fibroblasts (Liu et al., 2009b) and in other systems(Hordijk, 2006), overexpression of rac1 restored ROSgeneration in integrin b1 knockout fibroblasts (Figure 4b,arrow), indicating that ROS generation occurs downstreamof rac1.
Hydrogen peroxide rescues the phenotype of integrin b1knockout fibroblasts
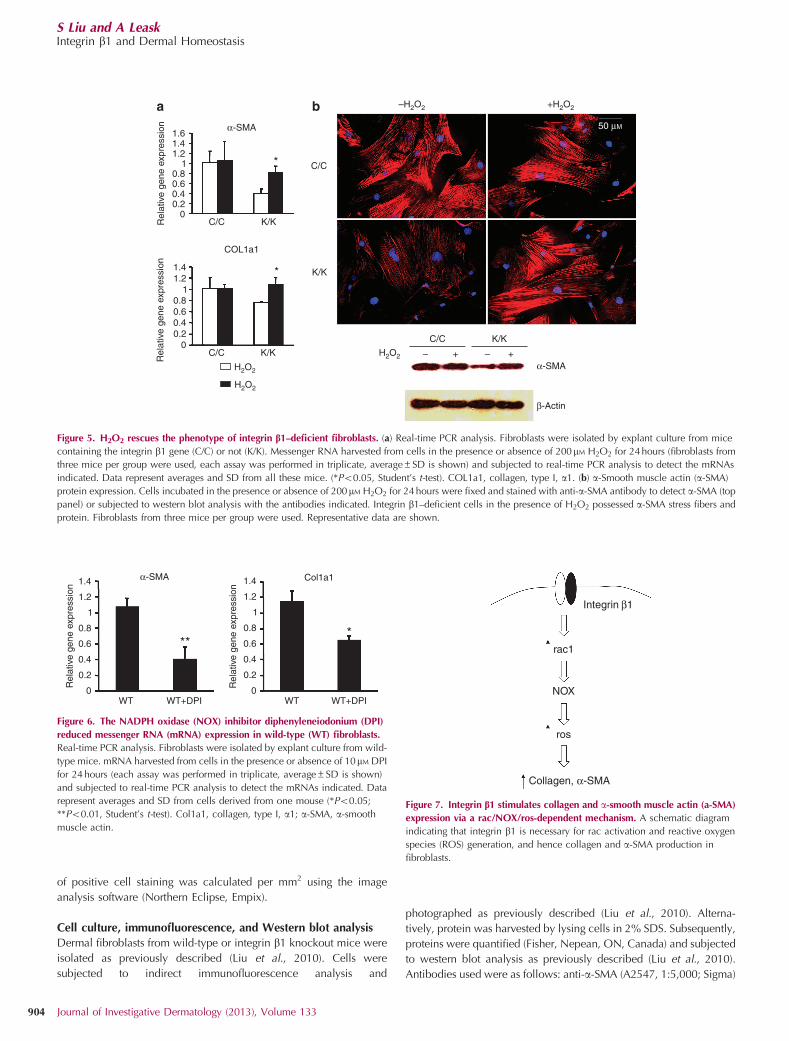
Previously, we had shown that integrin b1 knockout fibro-blasts expressed less collagen and a-SMA mRNA and hadreduced a-SMA stress fibers (Liu et al., 2010). We thenassessed whether the loss of ROS production in integrin b1knockout fibroblasts contributed to the phenotype of this lattercell type. Restoration of ROS by the addition of H2O2 rescuedthe relative inability of integrin b1 knockout cells to expressCol1a1 (collagen, type I, a1) or a-SMA mRNAs (Figure 5a).Moreover, H2O2 restored a-SMA protein expression and stressfiber formation in integrin b1 knockout fibroblasts (Figure 5b).Rac induces ROS generation by NADPH oxidase (NOX)(Leto et al., 2009); the addition of the NOX inhibitordiphenyleneiodonium reduced the expression of Col1a1 ora-SMA mRNAs in wild-type fibroblasts (Figure 6). Collectively,these data suggest that integrin b1 expression in fibroblasts isnecessary for the expression of collagen and a-SMA through arac1/ROS-dependent mechanism (Figure 7).
C/C K/K
C/Ca
c
b
100 μM
50 μM
K/K
C/C K/K
C/C K/K
*
**
250200150100500
Der
mal
thic
knes
s, μ
m
200
160
120
80
40
0Col
lage
n, μ
g m
g–1
H&
ETr
ichr
ome
Figure 1. Integrin b1 deletion results in reduced skin thickness. Mice deleted or not for integrin b1 (K/K and C/C, respectively) were examined 8 weeks after
deletion. (a) Gross examination of skin. Representative image is shown. (b) Hematoxylin and eosin (H&E) staining. (*Po0.05 K/K vs. C/C, Student’s t-test). For all
assays, six mice were analyzed. Scale bar¼100mM. (c) Trichrome staining. Colorimetric collagen quantification assay confirmed reduction of collagen in integrin
b1–deficient mice. Scale bar¼ 50mM. **Po0.01 K/K vs. C/C, Student’s t-test; N¼3.
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
900 Journal of Investigative Dermatology (2013), Volume 133
DISCUSSIONIntegrin b1, a key mediator of cell adhesion to both type Icollagen and fibronectin is essential for development, asintegrin b1 knockout mice die in utero (Stephens et al.,1995). Conditional knockout strategies have been used toshow that integrin b1 is essential for supporting progenitor cellproperties including adhesion to their niche and also formitotic spindle orientation (Lechler and Fuchs, 2005; Kuanget al., 2007). Loss of epidermal expression of integrin b1causes severe blistering because of impaired attachment ofbasal keratinocytes to the basement membrane, as well asdermal fibrosis and impaired wound healing (Raghavan et al.,2000; Brakebusch et al., 2000; Grose et al., 2002). Recently,we have begun to probe the in vivo role of integrin b1 infibroblasts; integrin b1 is required for the bleomycin model ofskin fibrosis and for cutaneous tissue repair in vivo (Liu et al.,2009a, 2010). Both of these models are initiated B10 daysafter deletion of the gene. The wound healing defect is linkedwith a relative inability of integrin b1–deficient cells toactivate latent transforming growth factor-b (Liu et al., 2010),consistent with prior data by Wipff et al. (2007) that
mechanical contraction of ECM by fibroblasts activatestransforming growth factor-b in an integrin/contraction-mediated manner.
In this report, we demonstrate that integrin b1 expression byfibroblasts is required for dermal homeostasis; 8 weeks afterdeletion of integrin b1, the skin is noticeably thinner than thatof wild-type mice. Moreover, we probe the mechanismoperating downstream of integrin b1 in driving collagen anda-SMA expression in fibroblasts. We found that integrin b1 isrequired in fibroblasts for collagen and a-SMA expression viaa rac1/ROS-dependent mechanism. These data are consistentwith a recent report using cardiomyocytes that showed thatintegrin b1 can activate ROS (Rosc-Schluter et al., 2012).Moreover, our results are consistent with our previous datademonstrating that rac1 knockout mice and fibroblastsshowed impaired profibrotic gene expression via an ROS-dependent mechanism (Liu et al., 2009b) and that racinhibition reduced profibrotic gene expression in fibroblasts(Shiwen et al., 2009), as well as prior data in other systemsindicating that rac activates ROS production through NADPHoxidases (Hordijk, 2006; Petry et al., 2010) and profibrotic
C/CPositive control K/K
C/C
Ki67
FSP
Overlay
K/K
C/C
80NS
60
40
20
Fib
robl
ast n
umbe
r/fie
ld0
K/K
50 μM
50 μm
a
b
TUNEL assay
Figure 2. Integrin b1 deletion does not affect proliferation or apoptosis. Mice deleted or not for integrin b1 (K/K and C/C, respectively) were examined
8 weeks after deletion. (a) TUNEL assay was performed. Please note that apoptotic (TUNEL-positive) cells were essentially absent in both (K/K) and (C/C) mice.
(b) Tissue was stained with anti-Ki67 (to detect proliferative cells) and anti-FSP1 (FSP; to detect fibroblasts). Please note that there was a nonstatistically
significant trend toward increased proliferation in integrin b1–deficient (K/K) mice (N¼ 3, Student’s t-test). NS, not significant. Scale bar¼ 50mM.
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
www.jidonline.org 901
gene expression in fibroblasts is decreased by rac1 inhibitionor in rac1 knockout fibroblasts (Shiwen et al., 2009; Liu et al.,2009b). These data extend our recent observations thatintegrin b1 is also required for the maintenance of the skinvasculature (Liu and Leask, 2012), for structural integrity ofcornea (Parapuram et al., 2011) deposition, and for survival ofpancreatic b-cells (Riopel et al., 2011). In this study, cellproliferation or apoptosis has not appreciably changed;isolated fibroblasts from integrin b1 knockout mice showedless collagen production, suggesting that in vivo the observedskin thinning phenotype arose because of a reduction ofcollagen synthesis per cell. Maintenance of the blood vesselswas impaired in integrin b1 knockout mice because of areduction of collagen and a-SMA expression by smoothmuscle cells (Liu and Leask, 2012). Thus, it is formallypossible that the thinner skin in the integrin b1 knockoutmice may be a secondary phenomenon owing to the lack ofoxygen and nutrients, although the fact that the blood vesselphenotype developed over a similar time frame to the skinthinning phenotype argues against this interpretation.
Integrin b1 is present in multiple integrin heterodimers,notably in a1b1, a2b1, a3b1, a10b1, and a11b1. It isunknown at present which heterodimers are responsible forthe effects outlined in our current and previous reports (Liuet al., 2009a, 2010). However, it is interesting to note thata1b1 has recently been shown to be involved in myofibroblastdifferentiation (Rodriguez et al., 2009) and acts as a negativefeedback inhibitor of collagen synthesis (Gardner et al., 1999).In addition, a2b1 regulates collagen gel remodeling throughforce generation and modulation of matrix metalloproteinaseactivity (Philips and Bonassar, 2005). Finally, a11b1modulates myofibroblast differentiation (Carracedo et al.,2010) and the survival of mesenchymal stem cells on
collagen (Popov et al., 2011). Thus, the overall effects of theloss of integrin b1 are likely to arise owing to a loss of all of thepossible integrin heterodimers.
Together with our prior observations that the persistentfibrotic phenotype of scleroderma fibroblasts is at leastpartially mediated by integrin b1 (Shi-Wen et al., 2007),and that integrin b1 is required for fibrogenesis and tissuerepair (Liu et al., 2009a, 2010), our results are consistent withthe notion that signaling via integrin b1 is essential forconnective tissue production and maintenance in thedermis. Modulating integrin b1 activity may be crucial forcontrolling ECM deposition in normal and pathologicalconditions.
MATERIALS AND METHODSGeneration of integrin b1 conditional knockout mice
All animal protocols were approved by the appropriate regulatory
authority. Integrin b1 conditional knockout mice were generated as
previously described (Liu et al., 2009a, 2010). Briefly, mice
homozygous for a loxP–integrin b1 allele (Raghavan et al., 2000;
Jackson Laboratories, Bar Harbor, ME) and hemizygous for allele
enabling the expression of tamoxifen-dependent Cre-recombinase
under the control of a fibroblast-specific regulatory sequence from the
proa2(I) collagen gene (Zheng et al., 2002). Animals were genotyped
using PCR (Raghavan et al., 2000; Zheng et al., 2002). To delete
integrin b1, 3-week-old mice were given intraperitoneal injections of
tamoxifen suspension (0.1 ml of 10 mg ml� 1 4-hydroxytamoxifen;
Sigma, St Louis, MO) or corn oil (vehicle) for 5 days. Loss of integrin
b1 was verified by PCR genotyping.
Mice were digitally photographed at 8 weeks after cessation of
tamoxifen injection using a Sony D-9 digital camera (Toronto, ON,
Canada). Mice were killed using CO2 for histological and immuno-
histochemical analyses.
C/C
3.53
2.52
*
*
1.51
0.50
250,000
200,000
150,000
100,000
50,000
0C/C K/K
Fib
robl
ast
Tis
sue
Flu
ores
cenc
e in
tens
ity, A
US
uper
oxid
esc
ore
(0–3
)
K/K
100 μM
Figure 3. Loss of integrin b1 results in a reduction in reactive oxygen species (ROS) production. In vivo (tissue) and in vitro (fibroblast) ROS production in
integrin b1–deficient mice (tissue) and cultured cells isolated from integrin b1–deficient mice (fibroblast) was measured using a superoxide anion–specific
fluorescent dye, dihydroethidium (DHE). The score of ROS generation in fibroblasts in tissue sections was quantified as described in the Materials and
Methods section (average of three mice per group and SD is shown). The average fluorescent intensity over fixed areas in cultured fibroblasts was also quantified
(average of data obtained from fibroblasts isolated from three mice and standard deviation is shown). Note that wild-type (C/C) fibroblasts showed elevated
ROS production (*Po0.05, Student’s t-test), relative to integrin b1–deficient cells (K/K). Scale bar¼ 100mM. AU, arbitrary unit.
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
902 Journal of Investigative Dermatology (2013), Volume 133
Immunofluorescence staining and histological assessment
Tissue sections (0.5mm) were cut using a microtome (Leica,
Richmond Hill, ON, Canada), placed on Superfrost Plus slides (Fisher
Scientific, Ottawa, ON, Canada), and then dewaxed in xylene and
rehydrated by successive immersion in descending concentrations of
alcohol. For immunofluorescence staining, tissue sections were
blocked by incubation with 5% donkey serum for 1 hour, washed
with phosphate-buffered saline, and then incubated with primary
antibodies for 1 hour at room temperature under humidified condi-
tions. Primary antibodies used were as follows: mouse anti-a-SMA
(1:500 dilution; Sigma), rabbit anti-Ki67, or mouse-anti-FSP1 (anti-
Ki67, anti-FSP1; 1:500 dilution; Abcam, Cambridge, MA). Subse-
quently, sections were then washed with phosphate-buffered saline,
incubated with appropriate fluorescent secondary antibodies (Jackson
Immunoresearch, West Grove, PA) for 1 hour at room temperature,
washed again with phosphate-buffered saline, and mounted using
40,6-diamidino-2-phenylindole. To assess the effects of integrin b1
deletion on collagen synthesis and skin thickness, trichrome collagen
stain and hematoxylin and eosin staining were used, respectively.
Slides were photographed using a Zeiss fluorescence microscope
(Empix, Missassauga, ON, Canada) and Northern Eclipse software
(Empix). Collagen expression was determined using a colorimetric
assay, as described by the manufacturer (Quickzyme, Leiden, The
Netherlands). Briefly, acid/pepsin-solubilized collagen was quantified
using a Sirus Red–based dye-based method by comparing observed
A540 values with those obtained using a standard curve run in parallel.
TUNEL assay was performed using a commercial kit with nuclease as
a positive control (TumorTACS, Trevigen, Gaithersburg, MD).
The tissue expression of a-SMA was graded on a scale of 0–3 by
three blinded observers: 0 signifies no staining, 1 signifies very little
staining, 2 signifies moderate staining, and 3 signifies extensive
staining. For proliferating cell nuclear antigen staining, the percentage
C/C
a
b
K/K
C/C K/K
GTP-Rac1
GT
P-R
ac1/
tota
l-Rac
1
Total-Rac1
DHE
CMV-rac1
Empty
50 μM
GFP Merge
Figure 4. Rac activity is reduced in integrin b1–deficient dermal fibroblasts. (a) Rac-GTP was immunoprecipitated from cell lysates. The precipitated Rac-GTP
was detected by immunoblot analysis using an anti-Rac antibody. In parallel, whole-cell lysates were subjected to SDS–PAGE and western blot analysis with an
anti-Rac antibody. Quantitative densitometry data are indicated on the right. Experiments were performed on cells derived from three mice (*Po0.05 relative to
wild-type control). (b) Transfection of constitutively active rac (CMV-rac1) compared with empty expression vector (Empty) increases reactive oxygen species
(ROS) generation in K/K cells (red, arrow). Cells were cotransfected with CMV-GFP to detect transfected cells (green). Scale bar¼ 50mM. CMV, cytomegalovirus;
DHE, dihydroethidium; GFP, green fluorescent protein; GTP, guanosine triphosphate.
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
www.jidonline.org 903
of positive cell staining was calculated per mm2 using the image
analysis software (Northern Eclipse, Empix).
Cell culture, immunofluorescence, and Western blot analysisDermal fibroblasts from wild-type or integrin b1 knockout mice were
isolated as previously described (Liu et al., 2010). Cells were
subjected to indirect immunofluorescence analysis and
photographed as previously described (Liu et al., 2010). Alterna-
tively, protein was harvested by lysing cells in 2% SDS. Subsequently,
proteins were quantified (Fisher, Nepean, ON, Canada) and subjected
to western blot analysis as previously described (Liu et al., 2010).
Antibodies used were as follows: anti-a-SMA (A2547, 1:5,000; Sigma)
K/K*
* C/C
–H2O2
H2O2
+H2O2
50 μM
H2O2
H2O2
C/C K/K
C/C K/K
C/C
– –+ +
K/K
α-SMA
a b
α-SMA
2
β-Actin
COL1a1
Rel
ativ
e ge
ne e
xpre
ssio
nR
elat
ive
gene
exp
ress
ion
Figure 5. H2O2 rescues the phenotype of integrin b1–deficient fibroblasts. (a) Real-time PCR analysis. Fibroblasts were isolated by explant culture from mice
containing the integrin b1 gene (C/C) or not (K/K). Messenger RNA harvested from cells in the presence or absence of 200mM H2O2 for 24 hours (fibroblasts from
three mice per group were used, each assay was performed in triplicate, average±SD is shown) and subjected to real-time PCR analysis to detect the mRNAs
indicated. Data represent averages and SD from all these mice. (*Po0.05, Student’s t-test). COL1a1, collagen, type I, a1. (b) a-Smooth muscle actin (a-SMA)
protein expression. Cells incubated in the presence or absence of 200mM H2O2 for 24 hours were fixed and stained with anti-a-SMA antibody to detect a-SMA (top
panel) or subjected to western blot analysis with the antibodies indicated. Integrin b1–deficient cells in the presence of H2O2 possessed a-SMA stress fibers and
protein. Fibroblasts from three mice per group were used. Representative data are shown.
WT WT+DPI WT WT+DPI
α-SMA
***
Col1a1
Rel
ativ
e ge
ne e
xpre
ssio
n
Rel
ativ
e ge
ne e
xpre
ssio
n
Figure 6. The NADPH oxidase (NOX) inhibitor diphenyleneiodonium (DPI)
reduced messenger RNA (mRNA) expression in wild-type (WT) fibroblasts.
Real-time PCR analysis. Fibroblasts were isolated by explant culture from wild-
type mice. mRNA harvested from cells in the presence or absence of 10mM DPI
for 24 hours (each assay was performed in triplicate, average±SD is shown)
and subjected to real-time PCR analysis to detect the mRNAs indicated. Data
represent averages and SD from cells derived from one mouse (*Po0.05;
**Po0.01, Student’s t-test). Col1a1, collagen, type I, a1; a-SMA, a-smooth
muscle actin.
Integrin β1
rac1
NOX
ros
Collagen, α-SMA
Figure 7. Integrin b1 stimulates collagen and a-smooth muscle actin (a-SMA)
expression via a rac/NOX/ros-dependent mechanism. A schematic diagram
indicating that integrin b1 is necessary for rac activation and reactive oxygen
species (ROS) generation, and hence collagen and a-SMA production in
fibroblasts.
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
904 Journal of Investigative Dermatology (2013), Volume 133

and anti-b-actin (1:5,000, A3853; Sigma). When indicated, cells were
treated in 0.5% fetal bovine serum in the presence or absence of
200mM H2O2 or 10mM diphenyleneiodonium (Sigma) for 24 hours.
Real-time PCR
Real-time PCR was performed essentially as described previously (Liu
et al., 2010; Thompson et al., 2010). Briefly, cells were cultured until
80% confluence and total RNA was isolated (Trizol, Invitrogen,
Burlington, ON, Canada). Integrity of the RNA was verified by gel
electrophoresis and by spectrophotometry. Total RNA (25 ng) was
reverse transcribed and amplified (using One-step mastermix and
TaqMan Assays on Demand probes; Applied Biosystems, Foster City,
CA) using the ABI Prism 7900 HT sequence detector (Perkin-Elmer-
Cetus, Vaudreuil, QC, Canada). Triplicates of each sample were run,
and expression values were standardized to values obtained with
control 18S RNA primers using the DDCt method.
Measurement of ROS production
In vivo and in vitro ROS production was measured essentially as
previously described, using a superoxide anion–specific fluorescent
dye, DHE (Sigma) (Wedgwood et al., 2001; Liu et al., 2009a). DHE is
oxidized on reaction with superoxide to ethidium bromide, which
binds to DNA in the nucleus and fluoresces red. For in vitro assays,
freshly frozen, enzymatically intact, 10-mm-thick sections from mice
were incubated with DHE (10mM) in phosphate-buffered saline for
30 minutes at 37 1C, protected from light. Sections were fixed in 4%
paraformaldehyde and photographed. The production of superoxide
in tissue was graded on a scale of 0–3 by three blinded observers: 0
signifies no staining, 1 signifies very little staining, 2 signifies
moderate staining, and 3 signifies extensive staining. For in vitro
assays, cells were incubated in the dark with 10mM DHE (30 minutes).
Cells were fixed and photographed, and fluorescent intensity in
individual cells was quantified by the Northern Eclipse (Empix)
software.
Rac activity assay
A commercially available rac activity assay was used, essentially as
previously described (Leask et al., 2008; Upstate Biotechnology, Lake
Placid, NY). Briefly, fibroblasts were grown in 60-mm dishes and
protein was harvested by lysis in buffer containing NP-40. Agarose
beads to which a p21-activated kinase–glutathione S-transferase
(PAK-GSH) fusion protein was conjugated was added. Active Rac
(Rac-GTP), which binds PAK-GSH, was recovered through repeated
centrifugation, washing of the agarose beads, and by removing bound
Rac, which was identified by boiling beads in Laemmli buffer. The
resultant extracts were then subjected to western blot analysis using
anti-Rac antibody. For the rac rescue experiment, expression vector
encoding constitutively active rac (V12rac1) under the control of the
cytomegalovirus promoter (courtesy Alan Hall, University College
London, London, UK) or empty expression vector was transfected into
cells using Fugene (Roche, Laval, QC, Canada) in a ratio of 2mg
DNA/3ml Fugene. Cells were cotransfected with an expression vector
encoding green fluorescent protein (Clontech, Mountain View, CA);
transfected cells were detected using a microscope.
CONFLICT OF INTERESTThe authors state no conflict of interest.
ACKNOWLEDGMENTSThis work is supported by grants from the Canadian Foundation for Innovation,the Canadian Institutes of Health Research, and the Scleroderma Society ofOntario.
REFERENCES
Barczyk M, Carracedo S, Gullberg D (2010) Integrins. Cell Tissue Res339:269–80
Brakebusch C, Grose R, Quondamatteo F et al. (2000) Skin and hair follicleintegrity is crucially dependent on beta 1 integrin expression onkeratinocytes. EMBO J 19:3990–4003
Brakebusch C, Fassler R (2005) b1 integrin function in vivo: adhesion,migration and more. Cancer Metastasis Rev 24:403–11
Camper L, Hellman U, Lundgren-Akerlund E (1998) Isolation, cloning, andsequence analysis of the integrin subunit alpha10, a beta1-associatedcollagen binding integrin expressed on chondrocytes. J Biol Chem273:20383–9
Carracedo S, Lu N, Popova SN et al. (2010) The fibroblast integrin alpha11-beta1 is induced in a mechanosensitive manner involving activin A andregulates myofibroblast differentiation. J Biol Chem 285:10434–45
Eckes B, Zigrino P, Kessler D et al. (2000) Fibroblast-matrix interactions inwound healing and fibrosis. Matrix Biol 19:325–32
Fassler R, Meyer M (1995) Consequences of lack of b1 integrin gene expressionin mice. Genes Dev 9:1896–908
Gardner HA (1999) Integrin signaling in fibrosis and scleroderma. CurrRheumatol Rep 1:28–33
Gardner H, Broberg A, Pozzi A et al. (1999) Absence of integrin alpha1beta1 inthe mouse causes loss of feedback regulation of collagen synthesis innormal and wounded dermis. J Cell Sci 112:263–72
Grose R, Hutter C, Bloch W et al. (2002) A crucial role of beta 1 integrins forkeratinocyte migration in vitro and during cutaneous wound repair.Development 129:2303–15
Gullberg D, Gehlsen KR, Turner DC et al. (1992) Analysis of alpha 1 beta 1,alpha 2 beta 1 and alpha 3 beta 1 integrins in cell–collagen interactions:identification of conformation dependent alpha 1 beta 1 binding sites incollagen type I. EMBO J 11:3865–73
Hinz B (2010) The myofibroblast: paradigm for a mechanically active cell.J Biomech 43:146–55
Hinz B, Gabbiani G (2003) Cell-matrix and cell-cell contacts of myofibroblasts:role in connective tissue remodeling. Thromb Haemost 90:993–1002
Hordijk PL (2006) Regulation of NADPH oxidases: the role of Rac proteins.Circ Res 98:453–62
Hynes RO (1992) Integrins: versatility, modulation, and signaling in celladhesion. Cell 69:11–25
Kuang S, Kuroda K, Le Grand F et al. (2007) Asymmetric self-renewal andcommitment of satellite stem cells in muscle. Cell 129:999–1010
Leask A, Shi-Wen X, Khan K et al. (2008) Loss of protein kinase Cepsilon resultsin impaired cutaneous wound closure and myofibroblast function. J CellSci 121(Part 20):3459–67
Lechler T, Fuchs E (2005) Asymmetric cell divisions promote stratification anddifferentiation of mammalian skin. Nature 437:275–80
Leto TL, Morand S, Hurt D et al. (2009) Targeting and regulation of reactiveoxygen species generation by Nox family NADPH oxidases. AntioxidRedox Signal 11:2607–19
Liu S, Kapoor M, Denton CP et al. (2009a) Loss of b1 integrin in mousefibroblasts results in resistance to skin scleroderma in a mouse model.Arthritis Rheum 60:2817–21
Liu S, Kapoor M, Leask A (2009b) Rac1 expression by fibroblasts is required fortissue repair in vivo. Am J Pathol 174:1847–56
Liu S, Leask A (2012) Integrin b1 is required for maintenance of vascular tonein postnatal mice. J Cell Commun Signal 6:175–80
Liu S, Xu SW, Blumbach K et al. (2010) Expression of integrin beta1 byfibroblasts is required for tissue repair in vivo. J Cell Sci 123:3674–82
Margadant C, Charafeddine RA, Sonnenberg A (2010) Unique and redundantfunctions of integrins in the epidermis. FASEB J 24:4133–52
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
www.jidonline.org 905

Parapuram SK, Huh K, Liu S et al. (2011) Integrin b1 is necessary for the
maintenance of corneal structural integrity. Invest Ophthalmol Vis Sci
52:7799–806
Petry A, Weitnauer M, Gorlach A (2010) Receptor activation of NADPH
oxidases. Antioxid Redox Signal 13:467–87
Phillips JA, Bonassar LJ (2005) Matrix metalloproteinase activity synergizes
with alpha2beta1 integrins to enhance collagen remodeling. Exp Cell Res
310:79–87
Popov C, Radic T, Haasters F et al. (2011) Integrins a2b1 and a11b1 regulate
the survival of mesenchymal stem cells on collagen I. Cell Death Dis
2:e186
Raghavan S, Bauer C, Mundschau G et al. (2000) Conditional ablation of beta1
integrin in skin. Severe defects in epidermal proliferation, basement
membrane formation, and hair follicle invagination. J Cell Biol
150:1149–60
Riopel M, Krishnamurthy M, Li J et al. (2011) Conditional b1-integrin-deficient
mice display impaired pancreatic b cell function. J Pathol 224:45–55
Rodriguez A, Karen J, Gardner H et al. (2009) Integrin alpha1beta1 is involved
in the differentiation into myofibroblasts in adult reactive tissues in vivo. J
Cell Mol Med 13:3449–62
Rosc-Schluter BI, Hauselmann SP, Lorenz V et al. (2012) NOX2-derived
reactive oxygen species are crucial for CD29-induced pro-survival
signalling in cardiomyocytes. Cardiovasc Res 93:454–62
Shi-Wen X, Renzoni EA, Kennedy L et al. (2007) Endogenous endothelin-1
signaling contributes to type I collagen and CCN2 overexpression in
fibrotic fibroblasts. Matrix Biol 26:625–32
Shiwen X, Liu S, Eastwood M et al. (2009) Rac inhibition reverses thephenotype of fibrotic fibroblasts. PLoS One 4:e7438
Stephens LE, Sutherland AE, Klimanskaya IV et al. (1995) Deletion of beta 1integrins in mice results in inner cell mass failure and peri-implantationlethality. Genes Dev 9:1883–95
Thompson K, Hamilton DW, Leask A (2010) ALK5 inhibition blocks TGF�-induced CCN2 expression in gingival fibroblasts. J Dent Res 89:1450–4
Tiger CF, Fougerousse F, Grundstrom G et al. (2001) alpha11beta1 integrin is areceptor for interstitial collagens involved in cell migration and collagenreorganization on mesenchymal nonmuscle cells. Dev Biol 237:116–29
Tomasek JJ, Gabbiani G, Hinz B et al. (2002) Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat Rev Mol Cell Biol 3:349–63
Watt FM, Fujiwara H (2011) Cell-extracellular matrix interactions in normaland diseased skin. Cold Spring Harb Perspect Biol 3:4
Wedgwood S, Dettman RW, Black SM (2001) ET-1 stimulates pulmonaryarterial smooth muscle cell proliferation via induction of reactive oxygenspecies. Am J Physiol Lung Cell Mol Physiol 281:L1058–67
Wipff PJ, Rifkin DB, Meister JJ et al. (2007) Myofibroblast contraction activateslatent TGF-beta1 from the extracellular matrix. J Cell Biol 179:1311–23
Yamamoto M, Yamato M, Aoyagi M et al. (1995) Identification of integrinsinvolved in cell adhesion to native and denatured type I collagens and thephenotypic transition of rabbit arterial smooth muscle cells. Exp Cell Res219:249–56
Zheng B, Zhang Z, Black CM et al. (2002) Ligand-dependent geneticrecombination in fibroblasts: a potentially powerful technique for inves-tigating gene function in fibrosis. Am J Pathol 160:1609–17
S Liu and A LeaskIntegrin b1 and Dermal Homeostasis
906 Journal of Investigative Dermatology (2013), Volume 133