
Impaired uptake of β-carotene by Caco-2 human intestinal cells in the presence of iron
11
Impaired uptake of b-carotene by Caco-2 human intestinal cells in the presence of iron ANTON BENGTSSON, NATHALIE SCHEERS, THOMAS ANDLID, MARIE LARSSON ALMINGER, ANN-SOFIE SANDBERG & ULF SVANBERG Department of Chemical and Biological Engineering, Food Science, Chalmers University of Technology, Go ¨teborg, Sweden Abstract At present, there are conflicting data regarding whether or not b-carotene has a positive effect on iron absorption. This study was undertaken to evaluate possible interactions involved in the uptake of b-carotene and iron in a human intestinal cell model (Caco-2). The Caco-2 cells were incubated with test solutions containing different amounts of ferrous chloride (1050 mM) and b-carotene (0.32.5 mM) incorporated in synthetic micelles. In the absence of iron, cellular accumulation of b-carotene from synthetic micelles was proportional (r 2 0.97, P B0.001) to the b-carotene concentration in the test solution. However, with addition of ferrous chloride (30 mM), the b-carotene uptake was significantly reduced (P B0.05), on average by 22%. There was also an inverse relationship between the b-carotene uptake and iron concentration in the test solution (r 2 0.93, P B0.05). Iron provided in physiological amounts inhibited the uptake of b-carotene in the in vitro Caco-2 cell system. Keywords: Caco-2 cells, b-carotene, iron uptake, micelles, ferritin, high-performance liquid chromatography Introduction Both vitamin A deficiency and anaemia are public health problems that affect millions of people worldwide, particularly in developing countries (World Health Organization 2001; Sommer and Davidson 2002). While vitamin A deficiency essentially is caused by an insufficient intake of vitamin A and provitamin A carotenoids from foods, anaemia may be a result of several disorders such as iron deficiency, folate deficiency, and infectious diseases including malaria and intestinal parasites (Lee 1983; Viteri 1997; van den Broek and Letsky 2000). Vitamin A deficiency is also recognized as a cause of anaemia (Bloem 1995). The mechanisms by which vitamin A deficiency induces anaemia are not clear, but impaired iron metabolism and erythropoiesis as well as decreased resistance to infection have been suggested in previous reviews (Fishman et al. 2000; Semba and Bloem 2002). Vitamin A and provitamin A (b-carotene) have also been proposed to influence iron absorption. In two Venezuelan Correspondence: Anton Bengtsson, Department of Chemical and Biological Engineering, Food Science, Chalmers University of Technology, SE-412 96 Go ¨teborg, Sweden. Fax: 46 31 772 38 30. E-mail: [email protected] ISSN 0963-7486 print/ISSN 1465-3478 online # 2009 Informa UK Ltd DOI: 10.1080/09637480802641270 International Journal of Food Sciences and Nutrition, September 2009; 60(S5): 125135 Int J Food Sci Nutr Downloaded from informahealthcare.com by University of Connecticut on 10/28/14 For personal use only.
Transcript of Impaired uptake of β-carotene by Caco-2 human intestinal cells in the presence of iron

Impaired uptake of b-carotene by Caco-2 humanintestinal cells in the presence of iron
ANTON BENGTSSON, NATHALIE SCHEERS, THOMAS ANDLID,
MARIE LARSSON ALMINGER, ANN-SOFIE SANDBERG &
ULF SVANBERG
Department of Chemical and Biological Engineering, Food Science, Chalmers University of
Technology, Goteborg, Sweden
AbstractAt present, there are conflicting data regarding whether or not b-carotene has a positive effect oniron absorption. This study was undertaken to evaluate possible interactions involved in theuptake of b-carotene and iron in a human intestinal cell model (Caco-2). The Caco-2 cells wereincubated with test solutions containing different amounts of ferrous chloride (10�50 mM) andb-carotene (0.3�2.5 mM) incorporated in synthetic micelles. In the absence of iron, cellularaccumulation of b-carotene from synthetic micelles was proportional (r2�0.97, PB0.001) tothe b-carotene concentration in the test solution. However, with addition of ferrous chloride(30 mM), the b-carotene uptake was significantly reduced (PB0.05), on average by 22%. Therewas also an inverse relationship between the b-carotene uptake and iron concentration inthe test solution (r2�0.93, PB0.05). Iron provided in physiological amounts inhibited theuptake of b-carotene in the in vitro Caco-2 cell system.
Keywords: Caco-2 cells, b-carotene, iron uptake, micelles, ferritin, high-performance liquid
chromatography
Introduction
Both vitamin A deficiency and anaemia are public health problems that affect millions
of people worldwide, particularly in developing countries (World Health Organization
2001; Sommer and Davidson 2002). While vitamin A deficiency essentially is caused
by an insufficient intake of vitamin A and provitamin A carotenoids from foods,
anaemia may be a result of several disorders such as iron deficiency, folate deficiency,
and infectious diseases including malaria and intestinal parasites (Lee 1983; Viteri
1997; van den Broek and Letsky 2000). Vitamin A deficiency is also recognized as a
cause of anaemia (Bloem 1995). The mechanisms by which vitamin A deficiency
induces anaemia are not clear, but impaired iron metabolism and erythropoiesis as
well as decreased resistance to infection have been suggested in previous reviews
(Fishman et al. 2000; Semba and Bloem 2002). Vitamin A and provitamin A
(b-carotene) have also been proposed to influence iron absorption. In two Venezuelan
Correspondence: Anton Bengtsson, Department of Chemical and Biological Engineering, Food Science,
Chalmers University of Technology, SE-412 96 Goteborg, Sweden. Fax: �46 31 772 38 30. E-mail:
ISSN 0963-7486 print/ISSN 1465-3478 online # 2009 Informa UK Ltd
DOI: 10.1080/09637480802641270
International Journal of Food Sciences and Nutrition,
September 2009; 60(S5): 125�135
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

studies, supplementation of iron-fortified cereal meals with either vitamin A or
b-carotene enhanced the absorption of non-heme iron in humans (Layrisse et al.
1997; Garcia-Casal et al. 1998). On the contrary, in a Swiss/Swedish study no effect of
vitamin A was noticed on iron absorption in adults who consumed test meals of corn
bread (Walczyk et al. 2003). In addition, erythrocyte incorporation of iron decreased
significantly from a maize porridge supplemented with vitamin A given to iron-
deficient and vitamin A-deficient children in Cote d’Ivoire (Davidsson et al. 2003).
The contradictory findings in these studies might be explained by differences in
vitamin A, iron or disease status between the study groups. To what extent food iron
might influence the absorption of vitamin A or provitamin A carotenoids is not known.
Hence, there is an obvious lack of knowledge of the possible interactions between
vitamin A, b-carotene, iron, and their respective uptake systems during absorption
and the subsequent metabolism of these nutrients. The metabolic interactions of these
two micronutrients may have implications for the design of intervention programs
aimed at populations with both vitamin A deficiency and iron-deficiency anaemia.
The following study was designed to evaluate possible interactions involved in the
uptake of b-carotene and iron in a Caco-2 human intestinal cell model under
conditions of iron deficiency. The Caco-2 cell line is a human colon adenocarcinoma
cell line that differentiates into polarized duodenal-like cells in culture, thus displaying
many characteristics of normal enterocytes (Pinto et al. 1983; Hidalgo et al. 1989).
The Caco-2 cell line is recognized as an established model for studies of intestinal iron
uptake (Fairweather-Tait et al. 2005) and it has also been used to measure the
incorporation of b-carotene and other carotenoids (Garrett et al. 1999; During et al.
2002). The specific objective of the present study was to explore the effects of
interactions*direct or indirect*between b-carotene in synthetic micelles and iron on
the uptake of these nutrients by Caco-2 cells.
Materials and methods
Chemicals
Synthetic crystalline b-carotene (]95% all-trans), lysophosphatidylcholine, sodium
glycocholate, sodium glycodeoxycholate, sodium taurocholate, sodium taurodeox-
ycholate, cholesterol, butylated hydroxytoluene, minimal essential medium (MEM),
and radioimmunoprecipitation assay lysis buffer were purchased from Sigma-Aldrich
(Schnelldorf, Germany). High-purity (99%) monoolein, oleic acid, and phosphati-
dylcholine were obtained from Larodan Fine Chemicals (Malmo, Sweden). Media,
supplements, and other reagents for cell culture maintenance were purchased from
Fischer Scientific GTF (Vastra Frolunda, Sweden).
Cell culture
Caco-2 cells (HTB-37) were obtained from American Type Culture Collection
(Rockville, MD, USA) at passage 18. Stock cultures were maintained in Dulbecco’s
modified Eagle medium supplemented with 16% (v/v) foetal bovine serum, 1% (v/v)
non-essential amino acids, and antibiotic solution (100 units/ml penicillin and 100 mg/
ml streptomycin) at 378C in a humidified atmosphere of 95% air and 5% CO2. The
growth medium was changed every second to third day. Monolayers were subcultured
at �80% confluence using 0.5 g/l trypsin with 0.5 mM ethylenediamine tetraacetic
126 A. Bengtsson et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

acid (EDTA) in Dulbecco’s phosphate-buffered saline (PBS). Cells at passages 27�35
were seeded at a density of 50,000 cells/cm2 in collagen-treated 12-well plates
(Corning Incorporated Life Sciences, Lowell, MA, USA).
Preparation of synthetic micelles containing b-carotene
Synthetic micelles were prepared based on several studies (Canfield et al. 1990;
Chitchumroonchokchai et al. 2004; Reboul et al. 2005) with some minor modifica-
tions. Stock solutions of monoolein, oleic acid, phosphatidylcholine, and cholesterol in
chloroform, and lysophosphatidylcholine in chloroform:methanol (9:1, v/v) were
prepared, blanketed with nitrogen and stored at �208C. A stock solution of all-
trans-b-carotene was prepared in n-hexane. Aliquots of these compounds were
combined in a 20 ml glass vial to obtain the following final concentrations: 0.5 mM
monoolein, 1.5 mM oleic acid, 0.1 mM phosphatidylcholine, 0.3 mM lysophosphati-
dylcholine, and 0.15 mM cholesterol. b-Carotene was added to yield micellar
concentrations ranging from 0.3 to 2.5 mM in the final test solutions. The transfer
efficiency of crystalline b-carotene into micelles was lower with higher added
concentrations of b-carotene in the preparation of micelles (data not shown). Solvents
were evaporated by a stream of nitrogen at room temperature. MEM containing
25 mM 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES) and 0.85 mM
glycocholate, 0.45 mM glycodeoxycholate, 0.45 mM taurocholate, and 0.25 mM
taurodeoxycholate was added to the vial and the mixture was sonicated in a water
bath (Branson Ultrasonics Corporation, 8510; Danbury, CT, USA) at room
temperature for 30 min. The mixture was filtered and sterilized by passage through a
0.22 mm filter to remove microcrystalline b-carotene particles, and the resulting
micellar solution was optically clear. All b-carotene preparations were protected from
light. High-performance liquid chromatography (HPLC) analysis was used to
determine the b-carotene content in the final micellar solutions. No significant
degradation of b-carotene was detected after sonication and filtration. In the uptake
experiments with added iron, no differences were observed in b-carotene content
between micellar test solutions containing iron added at different concentrations (10�50 mM) measured after 2 h incubation. Micellar solutions containing b-carotene were
prepared immediately prior to the uptake experiments.
Uptake experiments
Iron and b-carotene uptake experiments were conducted 14 days post seeding. Two
days before the uptake experiment, the growth medium was exchanged for MEM
supplemented with 3.5 g/l glucose and antibiotic solution (100 units/ml penicillin and
100 mg/ml streptomycin) in order to induce iron deficiency in the cells. This medium
did not contain detectable levels of iron (B0.5 mM) as determined by a high-
performance ion chromatography method developed by Fredriksson et al. (2002).
The mean background ferritin level in the cells after 2 days of iron-free incubation was
5.5 ng ferritin/mg protein. Prior to the experiments, the integrity of the monolayers
was examined by optical microscopy. To initiate the experiments, cells were washed
once in fresh MEM before adding 1 ml test solution containing 0.5 ml MEM and
0.5 ml micellar solution. The total b-carotene concentrations in the different test
solutions varied between 0.3 and 2.5 mM, and iron as ferrous chloride (FeCl2�4H2O)
was added at a concentration of 30 mM. Ferrous iron is commonly provided as
Impaired Caco-2 cell uptake of b-carotene 127
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.
a dietary fortification agent. It is also extensively used in human absorption studies as
well as in Caco-2 cell studies. Cells incubated with test solutions devoid of added iron
were used to obtain baseline ferritin levels. The pH of the test solutions was �7.2 and
remained stable during the incubation period. The incubation with test solutions was
carried out for 2 h at 378C in a humidified atmosphere of 95% air and 5% CO2. After
this incubation period, the test solutions were removed from the wells. The cells were
washed once with PBS containing 1% (w/v) bovine serum albumin to reduce non-
specific adsorption of micelles and twice with PBS only. Fresh MEM was added to the
cells before the plates were returned to the incubator for overnight incubation (22 h)
to allow for ferritin formation.
A separate experiment was performed to test the stability of incorporated b-carotene
up to 24 h by comparing cells harvested both directly after the 2 h incubation with the
test solutions and after the additional 22 h incubation with fresh medium (MEM).
Four different concentrations of iron were used in this experiment (0, 10, 30, and
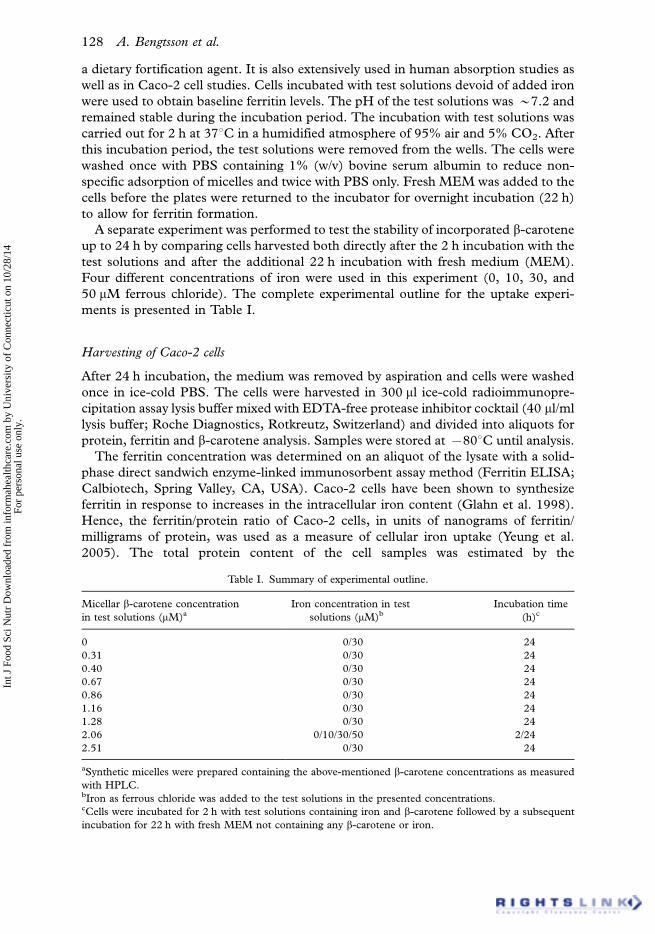
50 mM ferrous chloride). The complete experimental outline for the uptake experi-
ments is presented in Table I.
Harvesting of Caco-2 cells
After 24 h incubation, the medium was removed by aspiration and cells were washed
once in ice-cold PBS. The cells were harvested in 300 ml ice-cold radioimmunopre-
cipitation assay lysis buffer mixed with EDTA-free protease inhibitor cocktail (40 ml/ml
lysis buffer; Roche Diagnostics, Rotkreutz, Switzerland) and divided into aliquots for
protein, ferritin and b-carotene analysis. Samples were stored at �808C until analysis.
The ferritin concentration was determined on an aliquot of the lysate with a solid-
phase direct sandwich enzyme-linked immunosorbent assay method (Ferritin ELISA;
Calbiotech, Spring Valley, CA, USA). Caco-2 cells have been shown to synthesize
ferritin in response to increases in the intracellular iron content (Glahn et al. 1998).
Hence, the ferritin/protein ratio of Caco-2 cells, in units of nanograms of ferritin/
milligrams of protein, was used as a measure of cellular iron uptake (Yeung et al.
2005). The total protein content of the cell samples was estimated by the
Table I. Summary of experimental outline.
Micellar b-carotene concentration
in test solutions (mM)a
Iron concentration in test
solutions (mM)b
Incubation time
(h)c
0 0/30 24
0.31 0/30 24
0.40 0/30 24
0.67 0/30 24
0.86 0/30 24
1.16 0/30 24
1.28 0/30 24
2.06 0/10/30/50 2/24
2.51 0/30 24
aSynthetic micelles were prepared containing the above-mentioned b-carotene concentrations as measured
with HPLC.bIron as ferrous chloride was added to the test solutions in the presented concentrations.cCells were incubated for 2 h with test solutions containing iron and b-carotene followed by a subsequent
incubation for 22 h with fresh MEM not containing any b-carotene or iron.
128 A. Bengtsson et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

bicinchoninic acid protein assay (Pierce Biotechnology, Rockford, IL, USA) using
bovine serum albumin as a standard.
Extraction and analysis of b-carotene
An aliquot of the cell suspension was extracted three times with 1.5 ml n-hexane and
0.5 ml methanol including 0.1% (w/v) butylated hydroxytoluene. Samples were
vortexed and centrifuged at 2,500 x g between each extraction to facilitate phase
separation. The combined hexanic phases were evaporated to dryness in a water bath
at 358C under a stream of nitrogen. The residues were dissolved in 100 ml mobile
phase (methanol:methyl tert-butyl ether, 1:1 (v/v)). b-Carotene was analysed by
reversed-phase HPLC using a Waters 600 system equipped with auto sampler
injector, degasser, pump and a Waters 996 UV�visible photodiode array detector
operating at 450 nm. The data were stored and processed by means of Millennium
4.00 Software (Waters, Stockholm, Sweden). Absorption spectra were recorded
between 250 and 500 nm. Separations were carried out on a C30 carotenoid column
(5 mm, 250 mm�4.6 mm i.d.: YMC Europe GMBH, Schermbeck, Germany). The
mobile phase used for isocratic elution consisted of methanol:methyl tert-butyl
ether:water (55:41:4, v/v/v). The flow rate was 1 ml/min and the injection volume
40 ml. Quantification was achieved using calibration curves with b-carotene at
450 nm. b-Carotene could not be detected in cells that had been exposed to micellar
solutions without any carotenoids.
Statistical analysis
The data for the b-carotene uptake as a function of different b-carotene concentra-
tions in the test solutions with and without iron are represented by the mean9
standard deviation (SD) of four incubations for each concentration. Incubations
were performed on separate plates and on two different days. The significance of
differences in b-carotene uptake was calculated by Student’s t-test. The data for the
b-carotene uptake as a function of different iron concentrations in the test solutions
are represented by the mean9SD of six incubations from two separate occasions.
These data were analysed by analysis of variance, and determination of significance of
differences was obtained by Tukey’s HSD test. The Pearson coefficients and their
significance levels were obtained from linear regression analysis. The statistical
evaluation was performed with SPSS (version 14.0; SPSS Inc., Chicago, IL, USA)
and PB0.05 was considered significant.
Results and discussion
b-Carotene uptake in the absence of iron
Synthetic micelles were used as the delivery vehicle for b-carotene to Caco-2 cells since
micelles facilitate the transfer of carotenoids in vivo across the unstirred water layer to
the intestinal brush border membrane, and thereby also the subsequent uptake
through the membrane (Erdman et al. 1993; Yonekura and Nagao 2007). In the
present study, cellular b-carotene content was directly proportional (r2�0.97, PB
0.001) to the b-carotene concentration in the test solutions without added ferrous
iron (Figure 1). The incorporated b-carotene content varied between 20.6 and
Impaired Caco-2 cell uptake of b-carotene 129
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.
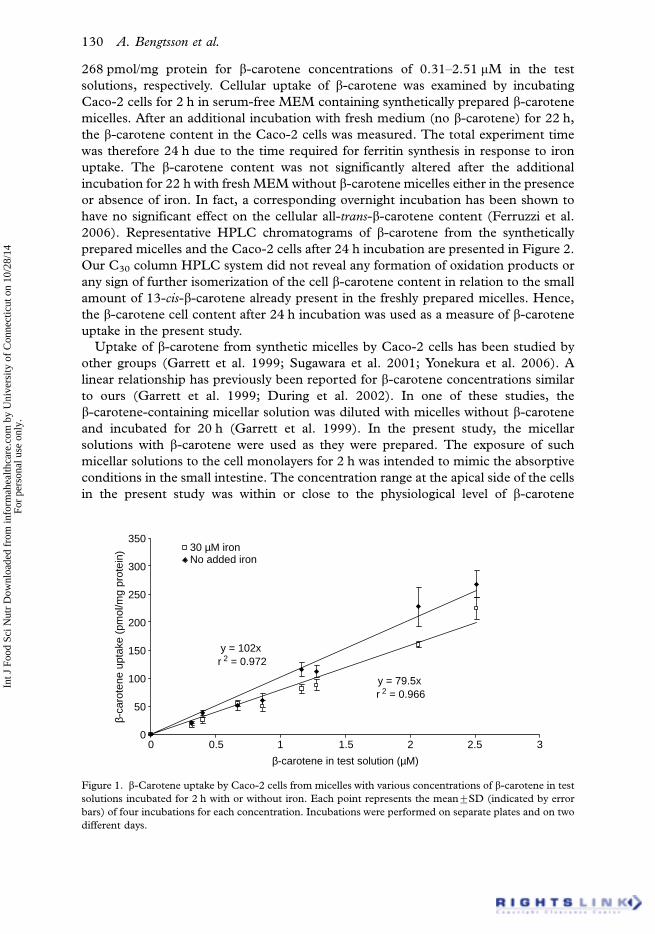
268 pmol/mg protein for b-carotene concentrations of 0.31�2.51 mM in the test
solutions, respectively. Cellular uptake of b-carotene was examined by incubating
Caco-2 cells for 2 h in serum-free MEM containing synthetically prepared b-carotene
micelles. After an additional incubation with fresh medium (no b-carotene) for 22 h,
the b-carotene content in the Caco-2 cells was measured. The total experiment time
was therefore 24 h due to the time required for ferritin synthesis in response to iron
uptake. The b-carotene content was not significantly altered after the additional
incubation for 22 h with fresh MEM without b-carotene micelles either in the presence
or absence of iron. In fact, a corresponding overnight incubation has been shown to
have no significant effect on the cellular all-trans-b-carotene content (Ferruzzi et al.
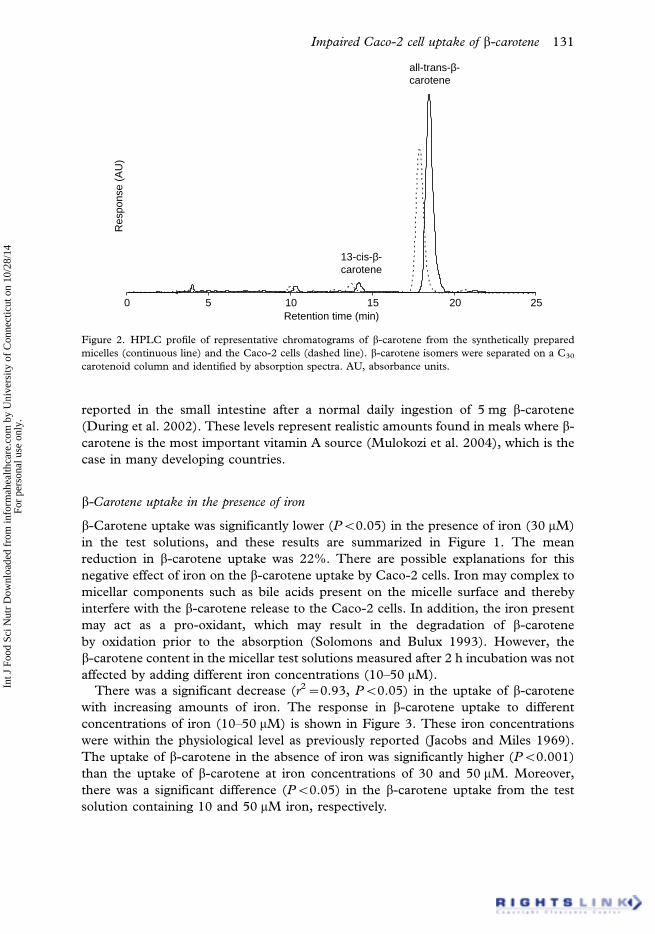
2006). Representative HPLC chromatograms of b-carotene from the synthetically
prepared micelles and the Caco-2 cells after 24 h incubation are presented in Figure 2.
Our C30 column HPLC system did not reveal any formation of oxidation products or
any sign of further isomerization of the cell b-carotene content in relation to the small
amount of 13-cis-b-carotene already present in the freshly prepared micelles. Hence,
the b-carotene cell content after 24 h incubation was used as a measure of b-carotene
uptake in the present study.
Uptake of b-carotene from synthetic micelles by Caco-2 cells has been studied by
other groups (Garrett et al. 1999; Sugawara et al. 2001; Yonekura et al. 2006). A
linear relationship has previously been reported for b-carotene concentrations similar
to ours (Garrett et al. 1999; During et al. 2002). In one of these studies, the
b-carotene-containing micellar solution was diluted with micelles without b-carotene
and incubated for 20 h (Garrett et al. 1999). In the present study, the micellar
solutions with b-carotene were used as they were prepared. The exposure of such
micellar solutions to the cell monolayers for 2 h was intended to mimic the absorptive
conditions in the small intestine. The concentration range at the apical side of the cells
in the present study was within or close to the physiological level of b-carotene
y = 102xr 2 = 0.972
y = 79.5xr 2 = 0.966
0
50
100
150
200
250
300
350
0 0.5 1 1.5 2 2.5 3
β-carotene in test solution (µM)
β-ca
rote
ne u
ptak
e (p
mol
/mg
prot
ein)
30 µM ironNo added iron
Figure 1. b-Carotene uptake by Caco-2 cells from micelles with various concentrations of b-carotene in test
solutions incubated for 2 h with or without iron. Each point represents the mean9SD (indicated by error
bars) of four incubations for each concentration. Incubations were performed on separate plates and on two
different days.
130 A. Bengtsson et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.
reported in the small intestine after a normal daily ingestion of 5 mg b-carotene
(During et al. 2002). These levels represent realistic amounts found in meals where b-
carotene is the most important vitamin A source (Mulokozi et al. 2004), which is the
case in many developing countries.
b-Carotene uptake in the presence of iron
b-Carotene uptake was significantly lower (PB0.05) in the presence of iron (30 mM)
in the test solutions, and these results are summarized in Figure 1. The mean
reduction in b-carotene uptake was 22%. There are possible explanations for this
negative effect of iron on the b-carotene uptake by Caco-2 cells. Iron may complex to
micellar components such as bile acids present on the micelle surface and thereby
interfere with the b-carotene release to the Caco-2 cells. In addition, the iron present
may act as a pro-oxidant, which may result in the degradation of b-carotene
by oxidation prior to the absorption (Solomons and Bulux 1993). However, the
b-carotene content in the micellar test solutions measured after 2 h incubation was not
affected by adding different iron concentrations (10�50 mM).
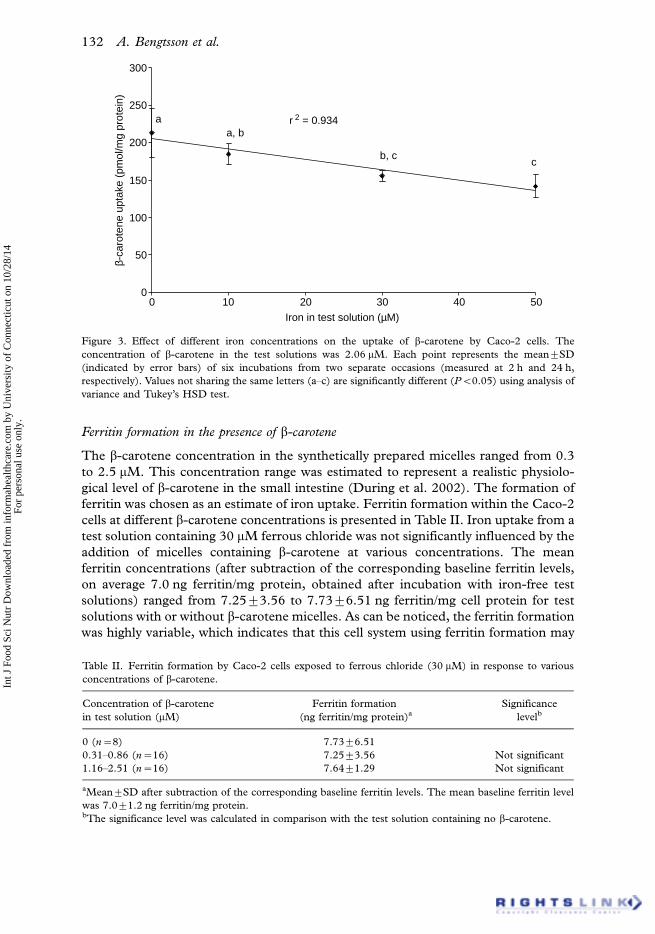
There was a significant decrease (r2�0.93, PB0.05) in the uptake of b-carotene
with increasing amounts of iron. The response in b-carotene uptake to different
concentrations of iron (10�50 mM) is shown in Figure 3. These iron concentrations
were within the physiological level as previously reported (Jacobs and Miles 1969).
The uptake of b-carotene in the absence of iron was significantly higher (PB0.001)
than the uptake of b-carotene at iron concentrations of 30 and 50 mM. Moreover,
there was a significant difference (PB0.05) in the b-carotene uptake from the test
solution containing 10 and 50 mM iron, respectively.
0 5 10 15 20 25Retention time (min)
Res
pons
e (A
U)
all-trans-β-carotene
13-cis-β-carotene
Figure 2. HPLC profile of representative chromatograms of b-carotene from the synthetically prepared
micelles (continuous line) and the Caco-2 cells (dashed line). b-carotene isomers were separated on a C30
carotenoid column and identified by absorption spectra. AU, absorbance units.
Impaired Caco-2 cell uptake of b-carotene 131
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.
Ferritin formation in the presence of b-carotene
The b-carotene concentration in the synthetically prepared micelles ranged from 0.3
to 2.5 mM. This concentration range was estimated to represent a realistic physiolo-
gical level of b-carotene in the small intestine (During et al. 2002). The formation of
ferritin was chosen as an estimate of iron uptake. Ferritin formation within the Caco-2
cells at different b-carotene concentrations is presented in Table II. Iron uptake from a
test solution containing 30 mM ferrous chloride was not significantly influenced by the
addition of micelles containing b-carotene at various concentrations. The mean
ferritin concentrations (after subtraction of the corresponding baseline ferritin levels,
on average 7.0 ng ferritin/mg protein, obtained after incubation with iron-free test
solutions) ranged from 7.2593.56 to 7.7396.51 ng ferritin/mg cell protein for test
solutions with or without b-carotene micelles. As can be noticed, the ferritin formation
was highly variable, which indicates that this cell system using ferritin formation may
r 2 = 0.934
0
50
100
150
200
250
300
0 10 20 30 40 50
Iron in test solution (µM)
β-ca
rote
ne u
ptak
e (p
mol
/mg
prot
ein)
aa, b
b, cc
Figure 3. Effect of different iron concentrations on the uptake of b-carotene by Caco-2 cells. The
concentration of b-carotene in the test solutions was 2.06 mM. Each point represents the mean9SD
(indicated by error bars) of six incubations from two separate occasions (measured at 2 h and 24 h,
respectively). Values not sharing the same letters (a�c) are significantly different (PB0.05) using analysis of
variance and Tukey’s HSD test.
Table II. Ferritin formation by Caco-2 cells exposed to ferrous chloride (30 mM) in response to various
concentrations of b-carotene.
Concentration of b-carotene
in test solution (mM)
Ferritin formation
(ng ferritin/mg protein)a
Significance
levelb
0 (n�8) 7.7396.51
0.31�0.86 (n�16) 7.2593.56 Not significant
1.16�2.51 (n�16) 7.6491.29 Not significant
aMean9SD after subtraction of the corresponding baseline ferritin levels. The mean baseline ferritin level
was 7.091.2 ng ferritin/mg protein.bThe significance level was calculated in comparison with the test solution containing no b-carotene.
132 A. Bengtsson et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

not be a suitable model for the investigation of possible effects of b-carotene on iron
absorption. However, with a modified Caco-2 cell system and with lower iron
concentrations, which may be the case in plant-based diets from developing countries,
any influence of b-carotene on iron uptake might be revealed.
One previous study addresses the question of whether b-carotene may influence
iron uptake by Caco-2 cells (Garcia-Casal et al. 2000). In contrast to the present
study, iron uptake was significantly increased in the presence of 0.9�6.0 mM
b-carotene. However, a number of differences in the experimental procedures make
a direct comparison difficult. In the study by Garcia-Casal et al. (2000), iron uptake
was measured as increased levels of radio iron in the incubated Caco-2 cells, while in
our study the formation of ferritin was used as a measure of cellular iron uptake.
Furthermore, cold water-soluble beadlets were used as the delivery vehicle for
b-carotene to the medium by Garcia-Casal et al. (2000). Micellar media prepared
with these water-soluble beadlets have been shown to be unstable, rendering orange
precipitations and hence a low micellar yield (Garrett et al. 1999). In addition, the
preparation of beadlets also contained additives such as sucrose and a-tocopherol,
which could possibly affect the uptake of nutrients in Caco-2 cells. In the present
study, b-carotene was presented to the Caco-2 cells in synthetically prepared micelles,
which more closely reflects the conditions in the duodenum.
To conclude, the presence of b-carotene, in concentrations ranging from 0.3 to
2.5 mM did not affect the uptake of iron in our cell assay. Our interpretation is that
there is no effect of b-carotene on the uptake of iron in cultured Caco-2 cells, and that
b-carotene converted to vitamin A is more likely to modulate iron utilization at
another level of iron metabolism. Animal studies have shown an impaired incorpora-
tion of iron into erythrocytes in vitamin A-deficient rats, suggesting that during this
condition iron is trapped in the liver and spleen and is not effectively released for
erythropoiesis by red bone marrow (Gardner et al. 1979; Mejıa et al. 1979).
At the chosen pH, suggested to be within the physiological pH range at the site for
absorption (Woodtli and Owyang 1995; Wright et al. 2008), there is a possibility of
some degree of iron precipitation, which may have compromised the effect of
b-carotene on iron uptake.
Conclusion
To our knowledge, the present study provides the first assessment of the impact of iron
on uptake of micellar b-carotene by Caco-2 cells. The uptake of b-carotene was
negatively affected by the presence of iron in the test solution provided to the cells,
and higher iron concentrations resulted in lower b-carotene uptake. However,
b-carotene in synthetic micelles did not increase iron uptake under the experimental
conditions employed in the present study. Our in vitro results that measures
accumulation of b-carotene in Caco-2 cells warrant further studies in humans using
stable isotope techniques that measures both uptake and consecutive metabolism of
b-carotene.
Acknowledgements
This work was supported by The Swedish Agency for Research Cooperation in
Developing Countries (Grant SWE-2004-005), The Swedish Council for Environment,
Impaired Caco-2 cell uptake of b-carotene 133
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

Agricultural Sciences and Spatial Planning (Formas) (Grant No. 222-2004-1889),
and Wilhelm och Martina Lundgrens stiftelse.
References
Bloem MW. 1995.Interdependence of vitamin A and iron: An important association for programmes of
anaemia control Proc Nutr Soc 54.
Canfield WK, Fritz TA, Tarara TE. 1990.Incorporation of b-carotene into mixed micelles Methods
Enzymol 189.
Chitchumroonchokchai C, Schwartz SJ, Failla ML. 2004.Assessment of lutein bioavailability from meals
and a supplement using simulated digestion and Caco-2 human intestinal cells J Nutr 134.
Davidsson L, Adou P, Zeder C, Walczyk T, Hurrell RF. 2003.The effect of retinyl palmitate added to iron-
fortified maize porridge on erythrocyte incorporation of iron in African children with vitamin A deficiency
Br J Nutr 90.
During A, Hussain MM, Morel DW, Harrison EH. 2002.Carotenoid uptake and secretion by CaCo-2 cells:
b-Carotene isomer selectivity and carotenoid interactions J Lipid Res 43.
Erdman JW Jr, Bierer TL, Gugger ET. 1993.Absorption and transport of carotenoids Ann NY Acad Sci
691.
Fairweather-Tait S, Lynch S, Hotz C, Hurrell R, Abrahamse L, Beebe S, Bering S, Bukhave K, Glahn R,
Hambidge M, et al. 2005.The usefulness of in vitro models to predict the bioavailability of iron and zinc:
A consensus statement from the HarvestPlus expert consultation Int J Vitam Nutr Res 75.
Ferruzzi MG, Lumpkin J, Schwartz SJ, Failla M. 2006.Digestive stability, micellarization, and uptake of
b-carotene isomers by Caco-2 human intestinal cells J Agric Food Chem 54.
Fishman SM, Christian P, West KP Jr.. 2000.The role of vitamins in the prevention and control of anaemia
Public Health Nutr 3.
Fredriksson M, Carlsson NG, Almgren A, Sandberg AS. 2002.Simultaneous and sensitive analysis of Cu,
Ni, Zn, Co, Mn, and Fe in food and biological samples by ion chromatography J Agric Food Chem 50.
Garcia-Casal MN, Layrisse M, Solano L, Baron MA, Arguello F, Llovera D, Ramirez J, Leets I, Tropper E.
1998.Vitamin A and b-carotene can improve nonheme iron absorption from rice, wheat and corn by
humans J Nutr 128.
Garcia-Casal MN, Leets I, Layrisse M. 2000.b-Carotene and inhibitors of iron absorption modify iron
uptake by Caco-2 cells J Nutr 130.
Gardner R, Hodges R, Rucker R. 1979.Fate of erythrocyte iron in vitamin A deficient rats Fed Proc 38.
Garrett DA, Failla ML, Sarama RJ, Craft N. 1999.Accumulation and retention of micellar b-carotene and
lutein by Caco-2 human intestinal cells J Nutr Biochem 10.
Glahn RP, Lee OA, Yeung A, Goldman MI, Miller DD. 1998.Caco-2 cell ferritin formation predicts
nonradiolabeled food iron availability in an in vitro digestion/Caco-2 cell culture model J Nutr 128.
Hidalgo IJ, Raub TJ, Borchardt RT. 1989.Characterization of the human colon carcinoma cell line
(Caco-2) as a model system for intestinal epithelial permeability Gastroenterology 96.
Jacobs A, Miles PM. 1969.Intraluminal transport of iron from stomach to small-intestinal mucosa Br Med J
4.
Layrisse M, Garcia-Casal MN, Solano L, Baron MA, Arguello F, Llovera D, Ramirez J, Leets I, Tropper E.
1997.The role of vitamin A on the inhibitors of nonheme iron absorption: Preliminary results J Nutr
Biochem 8.
Lee GR. 1983. The anemia of chronic disease Semin Hematol 20.
Mejıa LA, Hodges RE, Rucker RB. 1979.Clinical signs of anemia in vitamin A-deficient rats Am J Clin Nutr
32.
Mulokozi G, Hedren E, Svanberg U. 2004.In vitro accessibility and intake of b-carotene from cooked green
leafy vegetables and their estimated contribution to vitamin A requirements Plant Food Hum Nutr 59.
Pinto M, Robine-Leon S, Appay M-D, Kedinger M, Triadou N, Dussaulx E, Lacroix B, Simon-Assmann P,
Haffen K, Fogh J, Zweibaum A. 1983.Enterocyte-like differentiation and polarization of the human colon
carcinoma cell line Caco-2 in culture Biol Cell 47.
Reboul E, Abou L, Mikail C, Ghiringhelli O, Andre M, Portugal H, Jourdheuil-Rahmani D, Amiot M-J,
Lairon D, Borel P. 2005.Lutein transport by Caco-2 TC-7 cells occur partly by a facilitated process
involving the scavenger receptor class B type 1 (SR-BI) Biochem J 387.
Semba RD, Bloem MW. 2002.The anemia of vitamin A deficiency: Epidemiology and pathogenesis Eur J
Clin Nutr 56.
134 A. Bengtsson et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.

Solomons NW, Bulux J. 1993.Plant sources of provitamin A and human nutriture Nutr Rev 51.
Sommer A, Davidson FR. 2002.Assessment and control of vitamin A deficiency: The Annecy accords
J Nutr 132.
Sugawara T, Kushiro M, Zhang H, Nara E, Ono H, Nagao A. 2001.Lysophosphatidylcholine enhances
carotenoid uptake from mixed micelles by Caco-2 human intestinal cells J Nutr 131.
Walczyk T, Davidsson L, Rossander-Hulthen L, Hallberg L, Hurrell RF. 2003.No enhancing effect of
vitamin A on iron absorption in humans Am J Clin Nutr 77.
van den Broek NR, Letsky EA. 2000.Etiology of anemia in pregnancy in south Malawi Am J Clin Nutr 72.
Viteri FE. 1997.Iron supplementation for the control of iron deficiency in populations at risk Nutr Rev 55.
Woodtli W, Owyang C. 1995.Duodenal pH governs interdigestive motility in humans Am J Physiol
Gastrointest Liver Physiol 268.
World Health Organization. 2001. Iron deficiency anaemia: Assessment, prevention and control. A guide for
programme managers. Geneva: World Health Organization. p 15.
Wright AJ, Pietranglo C, MacNaugthon A. 2008.Influence of simulated upper intestinal parameters on the
efficiency of beta carotene micellarisation using an in vitro model of digestion Food Chem 107.
Yeung CK, Glahn RP, Miller DD. 2005.Inhibition of iron uptake from iron salts and chelates by divalent
metal cations in intestinal epithelial cells J Agric Food Chem 53.
Yonekura L, Nagao A. 2007.Intestinal absorption of dietary carotenoids Mol Nutr Food Res 51.
Yonekura L, Tsuzuki W, Nagao A. 2006.Acyl moieties modulate the effects of phospholipids on b-carotene
uptake by Caco-2 cells Lipids 41.
This paper was first published online on iFirst on 4 February 2009.
Impaired Caco-2 cell uptake of b-carotene 135
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Con
nect
icut
on
10/2
8/14
For
pers
onal
use
onl
y.


















