Expression of adenovirus type 12 Elb 58-kDa protein in Escherichia coli and production of antibodies...
8
Gene, 53 (1987) 173-180 Elsevier 173 GEN 01971 Expression of adenovirus type 12 Elb 5%kDa protein in Escherichia coli and production of antibodies raised against a S&kDa : : fi-galactosidase fusion protein (R~omb~~t DNA; gene cloning; prokaryotic vector; ~munoprecipitation; DNA-dependent translation system) Klaus SchugharP, Brigitte von Wilcken-Bergm~b and Helmut Esche” a Z~stitute of Molecular Biology (Cancer Research), Universjtyof Essen, D-4300 Essen I, and b Znstit~te of Genetics, U~~vers~~ of Cologne, D-5000 Cologne (F.R.G.) Tel. 49-0221-4702386 Received 14 October 1986 Revised 6 January 1987 Accepted 21 January 1987 SUMMARY DNA fragments coding for the N-terminal 185 amino acids (aa) and for the entire coding region of the adenovirus (Ad)12 Elb 58-kDa protein have been cloned in a prokaryotic expression vector. The N-terminal region of the 58kDa viral protein (aa 2 l-205) is expressed as a j?-galactosidase @Gal) fusion protein encoded by plasmid pBS8Ngal. E~/zetic/zia coli strains transformed with this plasmid synthesize a full-length fusion protein of 150-kDa and two truncated proteins: a MO-kDa protein cont~~ng aa 64-205 and a 120-kDa polypeptide containing aa 158-205 of the Elb 58-kDa protein. Antibodies raised against purified fusion proteins specifically immunoprecipitate the Elb 58kDa protein from AdlZinfected and transformed cells. Bacteria transformed with plasmid pB58 carrying the entire Elb 58-kDa coding region (minus the first N-terminal 20 aa which are replaced by 4 aa of /IGal) showed dramatically reduced growth properties after induction of 58K gene expression. We have not been able to detect substantial amounts of the 58-kDa protein in these cells. However, the viral 58-kDa polypeptide could be synthesized in vitro from plasmid pB58 in a DNA-dependent translation system from E. coli. I~RODU~IO~ The transforming region of human Ad is located at the left-hand end of the viral genome (Graham et al., 1974) and consists of two tr~sc~ption units, Ela and Elb. In the case of Ad12, the nucleotide sequence of both regions has been determined (Sugisaki et al., 1980; Bos et al., 198 1; Kimura et al., Corresgondenee ro: Dr. H. Esche, Institute of Molecular Biology (Cancer Research), University of Essen, D-4300 Essen 1 (F.R.G.) Tel. 49-0201-79913415. pyranoside; kb, kilobase or 1000 bp; nt, nucleotide(s); PAGE, polyacrylamide-gel electrophoresis; p.i., post infection; PolIk, Klenow (large) fragment of E. coli DNA polymerase I; R, resistance; SDS, sodium dodecyl sulfate; Tc, tetracycline; ::, Abbreviations: aa, ammo acid(s): Ad, adenovirus; /?Gal, fl-galac- tosidase; bp, base pair(s); IPTG, isopropyl-~-~thiog~acto- novel joint. 0378-I 119/87/%03.50 0 1987 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript of Expression of adenovirus type 12 Elb 58-kDa protein in Escherichia coli and production of antibodies...

Gene, 53 (1987) 173-180 Elsevier
173
GEN 01971
Expression of adenovirus type 12 Elb 5%kDa protein in Escherichia coli and production of antibodies raised against a S&kDa : : fi-galactosidase fusion protein
(R~omb~~t DNA; gene cloning; prokaryotic vector; ~munoprecipitation; DNA-dependent translation system)
Klaus SchugharP, Brigitte von Wilcken-Bergm~b and Helmut Esche”
a Z~stitute of Molecular Biology (Cancer Research), Universjty of Essen, D-4300 Essen I, and b Znstit~te of Genetics, U~~vers~~ of Cologne, D-5000 Cologne (F.R.G.) Tel. 49-0221-4702386
Received 14 October 1986
Revised 6 January 1987
Accepted 21 January 1987
SUMMARY
DNA fragments coding for the N-terminal 185 amino acids (aa) and for the entire coding region of the adenovirus (Ad)12 Elb 58-kDa protein have been cloned in a prokaryotic expression vector. The N-terminal region of the 58kDa viral protein (aa 2 l-205) is expressed as a j?-galactosidase @Gal) fusion protein encoded by plasmid pBS8Ngal. E~/zetic/zia coli strains transformed with this plasmid synthesize a full-length fusion protein of 150-kDa and two truncated proteins: a MO-kDa protein cont~~ng aa 64-205 and a 120-kDa polypeptide containing aa 158-205 of the Elb 58-kDa protein. Antibodies raised against purified fusion proteins specifically immunoprecipitate the Elb 58kDa protein from AdlZinfected and transformed cells. Bacteria transformed with plasmid pB58 carrying the entire Elb 58-kDa coding region (minus the first N-terminal 20 aa which are replaced by 4 aa of /IGal) showed dramatically reduced growth properties after induction of 58K gene expression. We have not been able to detect substantial amounts of the 58-kDa protein in these cells. However, the viral 58-kDa polypeptide could be synthesized in vitro from plasmid pB58 in a DNA-dependent translation system from E. coli.
I~RODU~IO~
The transforming region of human Ad is located at the left-hand end of the viral genome (Graham
et al., 1974) and consists of two tr~sc~ption units, Ela and Elb. In the case of Ad12, the nucleotide sequence of both regions has been determined (Sugisaki et al., 1980; Bos et al., 198 1; Kimura et al.,
Corresgondenee ro: Dr. H. Esche, Institute of Molecular Biology (Cancer Research), University of Essen, D-4300 Essen 1 (F.R.G.) Tel. 49-0201-79913415.
pyranoside; kb, kilobase or 1000 bp; nt, nucleotide(s); PAGE, polyacrylamide-gel electrophoresis; p.i., post infection; PolIk, Klenow (large) fragment of E. coli DNA polymerase I; R, resistance; SDS, sodium dodecyl sulfate; Tc, tetracycline; ::,
Abbreviations: aa, ammo acid(s): Ad, adenovirus; /?Gal, fl-galac- tosidase; bp, base pair(s); IPTG, isopropyl-~-~thiog~acto-
novel joint.
0378-I 119/87/%03.50 0 1987 Elsevier Science Publishers B.V. (Biomedical Division)

174
1981), the transcripts have been mapped (Sawada and Fujinaga, 1980; Bos et al., 1981; Saito et al., 1983), and the gene products have been identified (Jochemsen et al., 1981; Esche et al., 1984).
Region Elb of Ad12 codes for two proteins of 19 kDa and 58 kDa translated in two different read- ingframes from a 2.2-kb mRNA (Bos et al., 1981) at early times after infection. Elb 58K gene functions have been reported to play an important role during adenovirus infection and transformation of cells. In addition to Elb 19-kDa and Ela protein functions, the Elb 58K gene products are responsible for the expression of a fully transformed cell phenotype (Houweling et al., 1980; Van den Elsen et al., 1982) and for the high oncogenicity of AdlZtransformed cells in rodents (Bernards et al., 1983). The 58-kDa protein is also required to increase the half life of a cellular 53-kDa protein (Zantema et al., 1985) which is thought to be involved in cell-cycle regulation (Reich and Levine, 1984; Mercer et al., 1984).
It has been demonstrated in AdS-infected cells that functions of the Elb 58K coding region are necessary for efficient shutoff of transport and trans- lation ofhost-cell mRNAs late after infection (Babiss et al., 1985). Different functions of the Elb 58-kDa protein might be exhibited in Adl2-infected cells where the protein is also required for viral DNA replication (Shiroki et al., 1986).
Less is known, however, about the mechanism of the 58-kDa protein functions. As limited amounts of 58-kDa protein are synthesized in infected and transformed cells, we decided to establish a pro- karyotic expression system which would allow the synthesis of large quantities of the Elb 58-kDa pro- tein as well as 58-kDa : : /?Gal fusion proteins.
In this report we demonstrate the expression of an Elb 58-kDa: :fiGal fusion protein in E. coli. Anti- bodies raised against fusion proteins precipitate specifically the 58-kDa protein from Adl2-infected and transformed cells. The entire 58-kDa poly- peptide seems to be toxic for growth of the bacteria in vivo and could not be detected in these cells. However, the protein could be synthesized in vitro using a DNA-dependent in vitro translation system prepared from E. coli.
MATERIALS AND METHODS
(a) Bacterial strains, cells and virus
E. coli strain K-12/71-18 (Messing et al., 1977) and strain BR17 (von Wilcken-Bergmann et al., 1986) were used for transformation and expression of proteins. Human KB cells were obtained from the American Type Culture Collection. The HA12/7 cell line was derived from primary Syrian hamster embryo cells by transformation with Ad12 (zur Hausen, 1973). Infection of cells and purification of Ad12 have been described by Doerfler (1969).
(b) Construction and analysis of recombinant plas-
mids
Standard procedures (Maniatis et al., 1982) were used for agarose gel electrophoresis, plasmid prepa- rations, construction and transformation of recombi- nant plasmids. Dideoxysequencing of DNA was performed as described by Sanger et al. (1977).
(c) Purification of fusion proteins
Bacteria carrying plasmid pB58Ngal were grown overnight in YT-medium (Maniatis et al., 1982) containing 2 mM IPTG. Cells were harvested and incubated in 40 ~1 lysozyme solution (40 mM Tris - HCl pH 8; 5 mM EDTA; 0.3 mg/ml lysozyme) per ml culture for 10 min at 0’ C. Extracts were then treated with DNase for 10 min at 0” C (40 ~1 of 20 mM Tris - HCl pH 7.5; 20 mM MgCl; 10 pg/ml of DNase I) and 1~1 of 10% deoxycholate was added. Lysates were centrifuged for 10 min at 10000 x g and the resulting pellets were extracted with 4 M urea solution (4 M urea; 40 mM Tris * HCl pH 8; 5 mM EDTA; 0.5% Nonidet P40). The remaining 4 M urea-insoluble fraction was dissolved at 37°C for 1 h in SDS sample buffer (8 M urea; 50mM TriseHCI pH6.8; 2% SDS; 5% 2-mer- captoethanol; 10% glycerol), and fusion proteins from this fraction were further purified by prepara- tive PAGE (Laemmli, 1970).
(d) Expression of the 5%kDa protein in Escherichia coli extracts
Plasmid DNA (2.5 pg) was added to DNA- dependent E. coli translation extracts (Radiochemi-

175
cal Centre, Amersham, U.K.) and proteins were labeled with [ 35S]methionine as described by Zubay (1973).
(e) Immunization, immunoprecipitation and West-
em blotting
Purified fusion proteins (150-200 pg per injec- tion) were used to immunize 3-month-old rabbits. Injections and preparation of antisera had been per- formed as described previously (Schughart et al.,
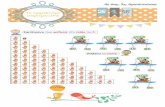
1775 Ad12 Elb 2457 //1,1//11 //////If//l//.C?l @
L” ll I
t t Hind III Taq I
I---- M13mplO BamHI+Accl
IacZa
Eco Rws:? l Act I
BamHI I ’
Eco
B PCS219 IacZoc
Hind III
F%
Hind III
1985). Anti-/?Gal antibodies were purchased from Atlanta (Heidelberg, F.R.G.); 35S-labeled anti- mouse Ig antibodies from Amersham. Cells were labeled for 1 h in the presence of 100 @i/ml [ 35S]methionine (Amersham) in methionine-free Dulbecco’s medium containing 5 % fetal calf serum. AdlZinfected KB cells were labeled 20 h post infec- tion. Preparation of cell extracts and immuno- precipitations have been described earlier (Schughart et al., 1985). Western blots were performed as described by Griesser et al. (1983).
Ad12 Elb 23s 3582 /,,,,,,,,/,,/,,,//,,//// /,,,
I 1 t Hind Ill TGA Pst I
Q pB58Ngal Hind Ill + Pst I
IacZ
Fig. 1. Construction of expression plasmids pB58Ngal (A) and
pB58 (B). (Panel A) A Sau3A-TaqI fragment was isolated from
a cloned EcoRI-C fragment of Ad12 (from A.J. van der Eb,
University of Leiden, Netherlands) and cloned between the
BamHI and AccI sites of vector M13mplO (Messing, 1983),
resulting in phage pKS36. Phage pKS36 DNA was cut with Sal1
and the protruding ends were partially (two out of four residues)
tilled in with dTTP and dCIP (T + C) using PolIk. Plasmid
pKS36 DNA was then cut with EcoRI and the resulting 703-bp
viral fragment was isolated. The newly constructed protruding
end at the Sal1 site was cohesive to a protruding end, partially
tilled in with dGTP and dATP (G + A), at the BamHI site of
vector pUC8 (Vieira and Messing, 1982). The pUC8 plasmid had
to be cut in addition with EcoRI to allow cloning of the 703-bp
viral fragment in frame to the a-domain of the 1acZ gene,
obtaining vector pCS219. As the 5’ end of the viral sequence in
plasmid pCS219 still contained a noncoding region of Elb, a
PvuII-PstI fragment was isolated from plasmid pCS219 and
cloned in frame into the 1acZ gene of the expression vector
piwiT15, which had been cut before at the Hind111 site, filled in,
and then cut with PstI. a and b mark the Elb 58K::piwiT15
junction sites shown in detail in Fig. 2. (Panel B) A 1264-bp
HindIII-PstI fragment of region Elb was isolated from the Ad12
EcoRI-C plasmid and inserted into Hind111 + PstI-cut plasmid
pB58Ngal resulting in plasmid pB58; PO,, synthetic promotor/
operator sequence.

176
RESULTS AND DISCUSSION
(a) Construction of expression plasmids pB58Ngal and pB58
The expression plasmid pB58Ngal directs syn- thesis of a fusion protein in which N-terminal 185th aa of the Ad12 Elb 5%kDa protein have been fused to BGal (Fig. lA, 2). Plasmid pB58 codes for the entire Elb 58-kDa protein minus the first N-terminal 20 aa (Fig. 1B).
We have used plasmid piwiT (a derivative of vector piwiT10; von Wilcken-Bergmann, 1986) as expression vector, which allows transcription of the IacZ gene from a synthetic promoter under the con-
trol of the Lac repressor (Fig. lA, 2). A codon frame shift within the multiple cloning site at the N-terminal codons of the 1acZ gene of piwiT permits easy identification of bacterial clones which synthesize fusion proteins after insertion of foreign DNA.
A 554-bp viral PvuII-TaqI Ad12 Elb fragment (nt 1903-2457) has been inserted in frame into the multiple cloning site of piwiT (Fig. 1A). The resulting construct, pB58Ngal, codes for a fusion protein of a theoretically predicted size of 140 kDa. This fusion protein contains 4 aa of piwiT at the N-terminus followed by 185 aa (21-205) of the Ad12 Elb 58-kDa protein which are fused to the N ter- minus of /KM (Fig. 1 A, 2). To join the viral fragment in frame to the pGal coding sequence, several sub-
Sequence a- _: -----CTCGAGATAAAAAATTTATTTGCTTTCAGGTAC
AATTCTTGATATAATATTATCATCTAGTAATTGTGAGCGCTCACAATTACTAGATAATAA
1904 AAATTTAAGGGATATCTTATGACAAGCTCTGTGGAGCT~TGTGGAGGGC----------------
MT S S 'J E G - - - - - - 21 22 23 (amino acid No.)
. . . . . . . . . . . piWiT15_........><....... Ad12 Elb . . . . . .._.
Sequence b: _~__. 2460
-----------------GTTGCTTTTAGATGTCGA ----- -VAFRCR (amino acid No.) 200 205 .._................... Ad12 Elb_...>
TCCGTCGACCTGCAGCAAGATCTACCCGGGAATTCACTGGCC------------ s v 3 LQQDLP G N S LA-----
< I . M13mplO .><....... PGal . .._........_....._.__....
Fig. 2. Nucleotide and amino acid sequences at the Elb 58K:: piwiT junction sites. Sequence (a) shows the promoter/operator region
with the 5’ region of Elb 58K gene fused to the first four N-terminal codons of piwiT15-lacZ gene in plasmids pB58Ngal and pB58.
Sequence (b) outlines the 3’ region of the Elb 58K fragment joined to the 5’ end of the BGaf-coding region in plasmid pB58Ngal.
Numbers refer to nucleotide and amino acid positions in the Ad12 genome.

177
cloning steps had to be performed as described in the legend of Fig. 1A.
The correct structure of the expression plasmids has been verified by restriction mapping and sequencing of the 5 ’ and 3’ Elb 58K: :lucZ junctions (Fig. 2).
Expression plasmid pB58 could be constructed in a single cloning step by inserting a HindIII-PstI fragment (nt 2318-3582) of the Ad12 Elb region into plasmid pB58Ngal (Fig. 1B). This fragment contains the remaining 3’ portion of the Elb 58-kDa coding region. Plasmid pB58 codes for the entire 58-kDa protein minus the first N-terminal 20 aa which have been replaced by 4 aa of vector piwiT (Figs. 1B and 2).
(b) Expression, characterization and purification of
El b 5%kDa : : BGal fusion proteins
Total protein extracts of transformed E. coli strains K-12/71-18 and BR17 were prepared after induction of the synthetic promoter with IPTG and analysed on Coomassie brilliant blue stained SDS-polyacrylamide gels (Fig. 3). pB58NgaLtrans- formed bacteria synthesized large amounts of 120-kDa and smaller amounts of 140-kDa and
a b .c d e f
150-kDa proteins (lanes c, g) constituting approx. 30-40% of total cellular protein in transformed BR17 bacteria. Transformation of E. coli strain BR17 with plasmid pB58Ngal resulted in clones producing about five times more of the described proteins than transformed K-12/71-18 bacteria (compare lanes c and g).
The 150-kDa, 140-kDa and the 120-kDa proteins were the only polypeptides synthesized at higher levels after induction with IPTG. In addition, these proteins could be detected with monoclonal anti- bodies against /IGal in Western blots (not shown), indicating that all three bands represent /?Gal fusion proteins.
Because limited amounts of the 150-kDa protein were synthesized in E. coli, it was not possible to purify sufficient quantities of this protein and to perform N-terminal amino acid sequencing. We con- clude from its M, that it represents the full-length fusion protein.
The 140-kDa and 120-kDa 58-kDa: : flGal fusion proteins, purified as described in MATERIALS AND
METHODS, section c (Fig. 4), were analysed in detail by amino acid sequencing. The first N-terminal 7 aa of the purified 140-kDa protein had been identified by Edman degradation as: M E Q Q V Q E . This
g h i
,150kDa 140kDa 120kDa
69kDa
46kDa
* L 2
71-16 BR17
Fig. 3. Expression of 5%kDa::BGal fusion proteins in pB58Ngal transformed bacteria. Total protein extracts from transformed E. coli
strains K-12/71-18 (lanes a-d) and BR17 (lanes e-h) were analysed on 0.2% SDS-7.5% polyacrylamide gels and stained with Coomassie brilliant blue. Lanes: a and e, extracts from untransformed strains (+ IPTG); b and f, extracts from pB%Ngal-transformed strains (-IPTG); c and g, extracts from pB58Ngal-transformed cells ( + IPTG); d and h, extracts from piwiT15-transformed strains ( + IPTG). Proteins of 150-kDa, 140-kDa and 120-kDa size observed in lanes b, c, f and g represent Elb 58-kDa::BGal fusion proteins. Lane i, size marker (BSA, 69 kDa; ovalbumin, 46 kDa).

d e i:
$50 kDa 14DkDa 12OkDa
&___._ -..-
Fig. 4. Purification of 5%kDa::/IGal fusion proteins. Bacterial lysates were fractionated (as described in MATERIALS AND METHODS, section c), into soluble fractions (lane a), 4 M urea extracts of the insoluble fraction (lane b), and the remaining fractions which are insoluble in 4 M urea (lane c) Further separa- tion of individual fusion proteins from fractions insoluble in 4 M urea (lane c) on preparative polyacrylamide gels resulted in pure preparations of 140-kDa (lane d) and 120-kDa (lane e) fusion proteins.
sequence represents aa 48-54 of the full-length 58kDa : : @Gal fusion protein, which implies that the 140-kDa protein lacks the first 47 aa of the fusion protein and contains aa 64-205 of the Elb 58-kDa polypeptide. The amino acid sequence obtained of the 120-kDa protein revealed the following N ter- minus: M IS A F A K . This sequence corresponds to aa 142-146 of the predicted 58-kDa: : /?Gal pro- tein, indicating that the 120~kDa protein lacks the first 140 aa of the full-len~h fusion protein and carries only aa 158-205 of the Ad12 Elb 58-kDa protein. Based on the nucleotide sequence (Bos et al., 198 l), an isoleucine residue would be expected instead of the methionine residue at position 141 of the fusion protein {aa 157 of the 58-kDa protein). We cannot explain this contradiction at the moment. Differences might be due to the use of DNA from different Ad12 strains or to a base-pair exchange during plasmid amplification in E. coli. Because of their truncated N termini, the 120-kDa and 140-kDa proteins might be derived from specific degradation of the 150-kDa protein or from usage of secondary translation initiation codons within the Elb coding region. The latter possibility seems to be more likely, because both protein sequences start with a methio- nine residue. In addition, Shine-D~g~o-Ike sequences (Scherer et al., 1980) are present upstream from the potential AUG start codons.
(c) Expression of the Elb 58-kDa protein in
Escherichia coli
We have not been able to observe substantial amounts of Elb 58-kDa protein in total protein extracts of pB58-transformed bacteria at various time points after induction with IPTG. Moreover, pB58-transformed bacteria showed greatly reduced growth after induction of the 58K gene (not shown), suggesting that the presence of the viral protein interferes with essential biological processes in E. coli. Similar observations have been made by Steinberg et al., 1986. As shown for plasmid pB58Nga1, there is some basal expression from the promoter of piwiT in bacteria grown in the absence of IPTG (Fig. 3). This basal expression might also be expected in pB58-transformed bac- teria; nevertheless, these cells grow reasonably well in the absence of IPTG. This observation could be explained by a high metabolic instability of the Elb 58”kDa polypeptide in E. co&. After induction of the 58K gene expression by IPTG, however, the amount of newly synthesized 58-kDa protein may reach a level which is toxic for bacterial growth. The amount of 58-kDa protein responsible for the growth inhibi- tion is still below the level of detection.
However, the 58-kDa polypeptide could be
a b c d e
69kDa-
46 kDa-
30kDa-
Elb - 58kDa
Fig. 5. Synthesis of the 58-kDa protein in DNA-dependent translation extracts from E. co/i. DNA of plasmid pAT153 (lane c), pB58 (lanes d and e), or no DNA (lane b) was added to DNA-dependent in vitro translation extracts from E. coli 35S-labeled proteins were analysed on O.Zu/, SDS-IS% poly- acrylamide gels. Lanee shows the immunoprecipitated Elb 5%kDa protein, expressed in these extracts, using anti-140-kDa antiserum. Lane a, size markers (BSA, 69 kDa; ovaibumin, 46 kDa; carbonic anhydrase, 30 kDa).

179
a b c d e f 9 h i
30kDa--_
Fig. 6. Immunoprecipitation of Elb 5%kDa protein from Adl2-infected and transformed cells. [35S]Methionine-labeled protein extracts
were prepared from mock infected or AdlZ-infected KB cells (20 h p.i.) and AdlZ-transformed HAl2/7 cells. Extracts of HAl2/7 cells
were incubated with preimmune serum (lane b), anti-140-kDa antiserum (lane c), anti-peptide antiserum (lane d), anti-j?Gal antiserum
(lane e), or anti-120-kDa antiserum (lane f). Extracts of mock-infected KB cells were incubated with anti-140-kDa antiserum (lane g),
and extracts of infected cells were precipitated with preimmune serum (lane h) or anti-140-kDa antiserum (lane i). Immunoprecipitated
proteins were analysed on 0.2% SDS-15% polyacrylamide gels. Lane a: size markers (see Fig. 5).
synthesized in DNA-dependent cell-free translation extracts from E. coli (Fig. 5, lanes d,e). It could be immunoprecipitated by anti-Elb 5%kDa antibodies (Fig. 5, lane e) and has the same M, as that immuno- precipitated from labeled extracts of infected cells. These results proved that the predicted structure of plasmid pB58 is correct. The 58-kDa protein synthe- sized in vitro will allow additional studies on the biological functions of the viral protein, although it might not be correctly modified post-translationally in these extracts.
(d) Immunoprecipitation of Ad12 Elb 5%kDa pro- tein from infected and transformed cells using anti- bodies raised against 58-kDa : : fiGal fusion proteins
Purified 140-kDa and 120-kDa fusion proteins (Fig. 4, lanes d,e) have been used to immunize rabbits. Antisera raised against the 140-kDa fusion protein specifically precipitated Elb 5%kDa protein from extracts of [ 35S]methionine-labeled Ad12- infected KB and transformed HA12/7 cells (Fig. 6, lanes c and i). Antisera raised against the 120-kDa fusion protein reacted only weakly with the Elb 58-kDa protein (lane f). Previously described anti- bodies directed against an octapeptide correspond- ing to the C-terminal region of Elb 58-kDa protein
(Schughart et al., 1985) precipitated the same Elb 58-kDa protein (Fig. 6, lane d). There are some indi- cations, together with results of recent immuno- precipitation experiments using anti-peptide anti- bodies (Schughart et al., 1985), for additional pro- teins of 70-kDa and 54-kDa coprecipitating in infected cells and for a 54-kDa protein co-precipi- tating in Adl2-transformed cells. At present we do not know whether these co-precipitated proteins are of viral or cellular origin.
The ‘monospecific’ antibodies described here were directed against a defined N-terminal region of Ad12 58-kDa protein (aa 64 to 205). These antibodies will allow immunoaffinity purification of the Elb 58-kDa protein from infected and transformed cells, investi- gations of functional properties in previously described Elb-dependent in vitro replication sys- tems (Shiroki et al., 1986), and studies on possible 58-kDa protein functions in in vitro translation systems.
ACKNOWLEDGEMENTS
We acknowledge the excellent help of H. Reinke, K. Neifer and Dr. K. Beyreuther (Institute of Genetics, Cologne) in amino acid sequencing of the fusion proteins.

180
REFERENCES
Babiss, L.E., Ginsberg, H.S. and Damell, J.E.: Adenovirus ElB proteins are required for accumulation of late viral mRNA and for effects on cellular mRNA translation and transport. Mol. Cell. Biol. 5 (1985) 2552-2558.
Bernards, R., Schrier, PI., Bos, J.L. and Van der Eb, A.J.: Role of adenovirus types 5 and 12 early region 1 B tumor antigens in oncogenic transformation. Virology 127 (1983) 45-53.
Bos, J.L., Polder, L.J., Bemards, R., Schrier, P.I., Van den Elsen, P.J., Van der Eb, A.J. and Van Ormondt, H.: The 2.2-kb ElB mRNA of human Ad12 and Ad5 codes for two tumor anti- gens starting at different AUG triplets. Cell 27 (1981) 121-131.
Doerfler, W.: Nonproductive infection of baby hamster kidney cells (BHK21) with adenovirus type 12. Virology 38 (1969) 587-606.
Esche, H., Reuther, M. and Schughart, K.: Early and late pro- teins of adenovirus type 12: translation mapping with RNA isolated from infected and transformed cells. Curr. Top. Microbial. Immunol. 111 (1984) 91-106.
Graham, F.L., Van der Eb, A.J. and Heyneker, H.L.: Size and location of the transforming region in human adenovirus type 5 DNA. Nature 25 (1974) 687-691.
Griesser, H., Mtiller-Hill, B. and Overath, P.: Characterization ofb-galactosidase-lactose-permease chimaeras in Escherikhiu
coli. Eur. J. Biochem. 137 (1983) 567-572. Houweling, A., Van den Elsen, P.J. and Van der Eb, A.J.: Partial
transformation of primary rat cells by the leftmost 4.5% fragment of adenovirus 5 DNA. Virology 105 (1980) 537-550.
Jochemsen, H., Daniels, G.S.G., Hertoghs, J.J.L., Schrier, P.I., Van den Elsen, P.J. and Van der Eb, A.J.: Identification of adenovirus type 12 gene products involved in transformation and oncogenesis. Virology 122 (1982) 15-28.
Kimura, T., Sawada, Y., Shinawawa, M., Shimizu, Y., Shiroki, K., Shimojo, H., Sugisaki,T.,Takanami, M., Uemizu,Y. and Fujinaga, K.: Nucleotide sequence of the transforming early region ElB of adenovirus type 12 DNA: structure and gene organization, and comparison with those of adenovirus type 5 DNA. Nucl. Acids Res. 9 (1981) 6571-6589.
Laemmli, U.K.: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227 (1970) 680-685.
Maniatis, T., Fritsch, E.F. and Sambrook, J.: Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982.
Mercer, W.E., Avignolo, C. and Baserga, R.: Role of the ~53 protein in cell proliferation as studied by microinjection of monoclonal antibodies. Mol. Cell. Biol. 4 (1984) 276-281.
Messing, J., Gronenborn, B., Mttller-Hill, B. and Hofschneider, P.H.: Filamentous coliphage Ml3 as a cloning vehicle: inser- tion of a Hind111 fragment of the lac regulatory region into Ml3 replicative form in vitro. Proc. Natl. Acad. Sci. USA 74 (1977) 3641-3646.
Messing, J.: New Ml3 vectors for cloning. Methods Enzymol. 101 (1983) 20-78.
Reich, N.C. and Levine, A.J.: Growth regulation of a cellular tumor antigen, ~53, in nontransformed cells. Nature 308 (1984) 199-201.
Sanger, F., Nicklen, S. and Coulson, A.R.: DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 74 (1977) 5463-5467.
Saito, I., Shiroki, K. and Shimojo, H.: mRNA species and proteins of adenovirus type 12 transforming regions: identiti- cation of proteins translated from multiple coding stretches in 2.2 kb region 1 B mRNA in vitro and in vivo. Virology 127 1983) 272-289.
Sawada, Y. and Fujinaga, K.: Mapping of adenovirus 12 mRNA’s transcribed from the transforming region. J. Virol. 36 (1980) 639-65 1.
Scherer, G.F.E., Walkinshaw, M.D., Arnott, S. and Morre, D.J.: The ribosome binding sites recognized by E. co6 ribosomes have regions with signal character in both the leader and protein coding segments. Nucl. Acids Res. 8 (1980) 3895-3907.
Schughart, K., Bause, E. and Esche, H.: Structure and expres- sion of adenovirus type 12 ElB 58K protein in infected and transformed cells: studies using antibodies directed against a synthetic peptide. Virus Res. 3 (1985) 41-56.
Shiroki, K., Ohishima, K., Fukui, Y. and Ariga, H.: The adeno- virus type 12 early region 1B SSOOO-ill, gene product is required for viral DNA synthesis and for initiation of cell transformation. J. Virol. 57 (1986) 792-801.
Steinberg, D.A., Watson, R.J. and Maiese, W.M.: Synthesis of fused glycoprotein D of Herpes simplex virus type 1 but not type 2 inhibits Escherichiu coli hosts. Gene 43 ( 1986) 3 1 l-3 17.
Sugisaki, H., Sugimoto, K., Takanami, M., Shiroki, K., Saito, I., Shimojo, H., Sawada, Y., Uemizu, Y., Uesugi, S.J. and Fujinaga, K.: Structure and gene organization in the trans- forming HindIII-G fragment of Ad12. Cell 20 (1980) 777-786.
Van den Elsen, P., De Pater, S., Houweling, A., Van der Veer, J. and Van der Eb, A.J.: The relationship between region Ela and El b of human adenoviruses in cell transformation. Gene
18 (1982) 175-185. Vieira, J. and Messing, J.: The pUC plasmid, an Ml3mp7-derived
system for insertion mutagenesis and sequencing with synthetic universal primers. Gene 19 (1982) 259-268.
von Wilcken-Bergmann, B., Tils, D., Sartorius, J., Auerswald, E.A., Schroder, W. and Mttller-Hill, B.: A synthetic operon containing 14 bovine pancreatic trypsin inhibitor genes is expressed in E. coli. EMBO J. 5 (1986) 3219-3225.
Zantema, A., Schrier, P.I., Davis-Oliver, A., Van Laar, T., Vaessen, R.T.M.J. and Van der Eb, A.J.: Adenovirus serotype determines association and localization of the large Elb tumor antigen with cellular ~53 in transformed cells. Mol. Cell. Biol. 5 (1985) 3084-3091.
Zubay, G.: In vitro synthesis of protein in microbial systems. Annu. Rev. Genet. 7 (1973) 267-287.
zur Hausen, H.: Interactions of adenovirus type 12 with host cell chromosomes. Prog. Exp. Tumor Res. 18 (1973) 240-259.
Communicated by H. van Ormondt.