
Danuza Montijo Diniz - Grupo Santa Casa BH - DANUZA... · A Gracy companheira e amiga de todas as...
71
Danuza Montijo Diniz AÇÃO FARMACOLÓGICA DA TOXINA Phα1β ISOLADA DO VENENO DA ARANHA Phoneutria nigriventer EM MODELOS DE DOR VISCERAL EM CAMUNDONGOS Instituto de Ensino e Pesquisa Pós-Graduação da Santa Casa de Belo Horizonte Pós-Graduação em Medicina e Biomedicina BELO HORIZONTE 2012
Transcript of Danuza Montijo Diniz - Grupo Santa Casa BH - DANUZA... · A Gracy companheira e amiga de todas as...

1
Danuza Montijo Diniz
AÇÃO FARMACOLÓGICA DA TOXINA Phα1β ISOLADA DO VENENO DA
ARANHA Phoneutria nigriventer EM MODELOS DE DOR VISCERAL EM
CAMUNDONGOS
Instituto de Ensino e Pesquisa
Pós-Graduação da Santa Casa de Belo Horizonte
Pós-Graduação em Medicina e Biomedicina
BELO HORIZONTE
2012

2
Danuza Montijo Diniz
AÇÃO FARMACOLÓGICA DA TOXINA Phα1β ISOLADA DO VENENO DA
ARANHA Phoneutria nigriventer EM MODELOS DE DOR VISCERAL EM
CAMUNDONGOS
.
Orientador: Prof. Dr. Marcus Vinícius Gomez
Co-Orientadora: Profa
Dra
Alessandra Hübner de Souza
BELO HORIZONTE 2012
Dissertação submetida ao Curso de Pós-graduação em Medicina e Biomedicina do Instituto de Ensino e Pesquisa da santa Casa de Belo Horizonte como requisito parcial para obtenção do grau de Mestre em Ciências.

3
Este trabalho foi realizado no Laboratório da pós-graduação do Instituto de
Ensino e Pesquisa da Santa Casa de Belo Horizonte com o auxílio das
seguintes instituições:
-Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
-Coordenação de Aperfeiçoamento de Nível Superior (CAPES)
-Capes Toxinologia 1444/2011 -Capes AUXPE 2865/10 -Pronex -Fapemig

4
“Sábio é aquele que conhece os limites da própria ignorânciaa”
Sócrates

5
Dedico essa tese
A minha família, principal incentivadora de meus estudos.........
Aos meus pais, Kleber e Jara, meus imãos Arabelle e Bernardo
que sempre souberam guiar meus passos e sempre me
proporcionaram plenas condições para atingir meus objetivos.

6
AGRADECIMENTOS
Agradeço a Deus por me direcionar e me capacitar a alcançar mais esta
conquista.
Aos meus Pais, pelo apoio incondicional, amor, paciência, força,
otimismo.
Aos meus irmãos, pela lição de amor, e alegria diária.
A minha avó, tios e primos pelas orações, presença, torcida e risadas,
vocês são parte da minha felicidade.
Meu tio Sinésio Bastos, seu filho Sinesinho por suas sábias palavras e
me incentivar desde o começo, a todos da casa do meu tio pelas risadas e
tardes de cafes maravilhosas....
A Gracy companheira e amiga de todas as horas aqui em BH...
Ao Felipinho pelo carinho e muita paciência comigo...
Agradeço ao meu orientador, professor Marcus Vinícius pela
oportunidade que me deu para o meu crescimento profissional, mas também
pessoal, fundamentais para o alcançar essa conquista. Sua maneira de
conduzir a pesquisa faz com que todas as pessoas ao seu redor se tornem
apaixonadas pelo mundo científico! Obrigada por tudo!
A minha grande amiga e co-orientadora Alessandra Hübner de Souza,
obrigada por acreditar em mim, pelo carinho, dedicação e incentivo sempre.
Adoro vc guria!!
Aos professores do IEP da Santa Casa pela disponibilidade e acolhida...

7
A Juju (béeerra cabrita) pela amizade, carinho, paciência e
companheirismo. Pessoa que não mediu esforços para me ajudar em meus
experimentos... Adoro vc cabritinha!
Ao Célio, pela amizade sincera, pelo carinho e pela ajuda científica...
A Elizete pelo carinho, dedicação e pela ajuda científica...
A Luciene pela amizade, carinho e grande ajuda científica
Nanci exemplo de força e determinação obrigada pelo carinho e
amizade...
Fabíola exemplo de pesquisadora, obrigada pelo incentivo sempre...
Ao Xandoca pelo carinho, ensinamentos e amizade...
Ao meu grande amigo Bráulio pelos ensinamentos, amizade e me fazer
rir quando precisava mesmo que de longe...
Ao Cláudio pelo carinho e amizade...
Ao pessoal da santa Casa o Diogo, Aírton, Fabi, Níbia, Carol a todos
pelo carinho e amizade de todos os dias...
Aos meus amigos queridos de laboratório, em especial o grupo da
neurofarmacologia: Drica, Pat, Simone, Vítor, Alexandre magno (meu rei),
Jéssica, Cinthia, Lucélia, Priscila, Luíza, Gabriel, Flavinha, Daiane, Karen, Dani
a todos pelo companheirismo.
Às funcionárias: Nívea, Shirley, Zélia e as meninas da limpeza. Vocês
foram fundamentais para este trabalho.
Aos animais utilizados, meu respeito e gratidão.
Ao IEP da Santa Casa de Belo Horizonte e às entidades financiadoras
CAPES, CNPq, pelo suporte e incentivo na busca pela excelência da pesquisa
brasileira

8
SUMÁRIO
AGRADECIMENTOS ..................................................................................................... 6
LISTA DE ABREVIATURAS ........................................................................................... 10
Lista de Figuras .............................................................................................................. 11
Resumo ........................................................................................................................... 12
1 INTRODUÇÃO ........................................................................................................... 14
1.1 DOR E NOCICEPÇÃO ............................................................................................ 14 1.2 CLASSIFICAÇÃO DA DOR ................................................................................... 15
1.2.1 Dor visceral ................................................................................................... 17
1.3 MECANISMOS DA HIPERALGESIA VISCERAL- MECANISMOS
PERIFÉRICOS E CENTRAIS ....................................................................................... 18 1.3.1 SENSIBILIZAÇÃO CENTRAL ..................................................................................... 19
1.4 MECANISMOS NEURAIS DA DOR ...................................................................... 20
1.5 MEDIADORES QUÍMICOS NA VIA NOCICEPTIVA ......................................... 25 1.6 CANAIS DE CÁLCIO SENSÍVEIS À VOLTAGEM .............................................. 28
1.6.1 SUBUNIDADES MOLECULARES DOS CANAIS DE CÁLCIO ............. 29
1.7 CANAIS DE CÁLCIO SENSÍVEIS À VOLTAGEM (CCSV) E NOCICEPÇÃO .. 29 1.8 USO DE TOXINAS ANIMAIS COMO AGENTES TERAPÊUTICOS .................. 32
1.8.1 Phoneutria nigriventer .................................................................................. 32
2. OBJETIVOS ............................................................................................................... 37
2.1 OBJETIVO GERAL ................................................................................................. 37
2.2 OBJETIVOS ESPECÍFICOS .................................................................................... 37
3 MATERIAL E MÉTODOS ......................................................................................... 38
3.1 MATERIAIS ................................................................................................................ 38 DROGAS E TOXINAS ......................................................................................................... 38 ANIMAIS .......................................................................................................................... 38 3.2 MÉTODOS ............................................................................................................... 39
3.2.1 TESTES COMPORTAMENTAIS ................................................................ 40
4 RESULTADOS ........................................................................................................... 44
4.1 AVALIAÇÃO DO EFEITO ANTINOCICEPTIVO DA PH1 NAS CONTORÇÕES ABDOMINAIS
DE CAMUNDONGOS INDUZIDA POR ÁCIDO ÁCETICO. ........................................................ 44
4.2 AVALIAÇÃO DO EFEITO ANTINOCICEPTIVO DA MVIIA NAS CONTORÇÕES ABDOMINAIS
DE CAMUNDONGOS INDUZIDAS POR ÁCIDO ÁCETICO ....................................................... 46
4.3 EFEITO DAS TOXINAS PH1 E MVIIA NO CONTEUDO DE GLUTAMATO DO LIQUIDO
CEREBROESPINHAL DE CAMUNDONGOS SUBMETIDOS A DOR VISCERAL INDUZIDA POR
ÁCIDO ÁCETICO. ............................................................................................................... 48
4.4 EFEITO DAS TOXINAS PH1 E MVIIA NO CONTEUDO DE ROS DO LIQUIDO
CEREBROESPINHAL DE CAMUNDONGOS SUBMETIDO A DOR VISCERAL INDUZIDA POR ÁCIDO
ÁCETICO. ......................................................................................................................... 50 4.5 EFEITO DA CONCENTRAÇÃO DA CAPSAICINA INJETADA INTRACOLONICA NO
COMPORTAMENTO NOCICEPTIVO DE CAMUNDONGOS. ...................................................... 52

9
4.6 EFEITO ANTINOCICEPTIVO DA PH1 NA DOR VISCERAL INDUZIDA POR INJEÇÃO
INTRACOLONICA DE CAPASAICINA. .................................................................................. 54 4.7: EFEITO ANTINOCICEPTIVO DA MVIIA NA DOR VISCERAL INDUZIDA POR INJEÇÃO
INTRACOLONICA DE CAPASAICINA. .................................................................................. 56 5 DISCUSSÃO ............................................................................................................... 58
6 CONCLUSÃO ............................................................................................................. 61
7 REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................ 62

10
LISTA DE ABREVIATURAS
ω-ctx-MVIIA Ômega-conotoxina-MVIIA
AMPA α-amino-3-hidroxi-5-metil-4-isoxazolproprionato
Ca2+ Íons cálcio
CCSV Canais de cálcio sensíveis à voltagem
CGRP Peptídeo relacionado ao gene da calcitonina
cm Centímetros
g Gramas
GDH Glutamato desidrogenase
h Horas
i.t Administração por via intratecal
i.c Administração intracolônica
K+ Íon potássio
LCE Liquido cerebro espinhal
Na+ Íon sódio NMDA N-metil-D-aspartato PAG Substância cinzenta periaquedutal
P. nigriventer Phoneutria nigriventer
ROS Espécies reativas de oxigênio
s Segundos
SNC Sistema nervoso central
SP Substância P
Tx3-6 Toxina Tx3-6
RVM Medula rostral ventromcdial

11
Lista de Figuras
Figura I: O ciclo das vesículas sinápticas
Figura II: Modelo molecular da exocitose de vesículas sinápticas
Figura III: Aranha Phoneutria nigriventer
Figura 1: Avaliação do efeito antinociceptivo da Ph1 das contorções
abdominais de camundongo induzido por ácido ácetico.
Figura 2: Avaliação do efeito antinociceptivo da MVIIA das contorções
abdominais de camundongo induzido por ácido ácetico
Figura 3: Efeito das toxinas Ph1 e MVIIA no conteudo de glutamato do
liquido cerebroespinhal de camundongo submetido a dor visceral induzido por
ácido ácetico.
Figura 4: Efeito das toxinas Ph1 e MVIIA no conteudo de ROS do liquido
cerebroespinhal de camundongo submetido a dor visceral induzido por ácido
ácetico.
Figura 5: Efeito da concentração da capsaicina injetada intracolonica no
comportamento nociceptivo de camundongo.
Figura 6: Efeito antinociceptivo da Ph1 na dor visceral induzida por injeção
intracolonica de capasaicina.
Figura 7: Efeito antinociceptivo da MVIIA na dor visceral induzida por injeção
intracolonica de capasaicina.

12
Resumo
Os medicamentos utilizados atualmente no tratamento da dor estão
frequentemente associados a efeitos colaterais graves e desenvolvimento
rápido de tolerância e assim, existe a necessidade de novos fármacos mais
seletivos. O bloqueio de canais de cálcio sensíveis à voltagem (CCSVs) no
corno dorsal da medula espinhal tem se mostrado um alvo promissor, pois a
redução dos níveis de cálcio intracelular é um dos mecanismos de modulação
da informação nociceptiva. O veneno da aranha brasileira Phoneutria
nigriventer contém uma variedade de substâncias com ação
predominantemente neurotóxica, dentre elas a fração Phα1β foi recentemente
descrita como uma potente bloqueadora da neurotransmissão de canais de
cálcio sensíveis a voltagem tipo N. O objetivo deste estudo foi avaliar a
possível atividade antinociceptiva da toxina Tx3-6, purificada do veneno da
aranha Phoneutria nigriventer, uma vez que já foi estabelecida sua afinidade
por CCSVs do tipo-N,-P/Q e –R e comparar sua ação com as ω-conotoxina
MVIIA. O influxo de cálcio através de CCSVs media a informação nociceptiva
no corno dorsal da medula. A administração dos bloqueadores de CCSVs, tais
como a ω-conotoxina-MVIIA (ω-CTx-MVIIA), tem efeito analgésico e estreita
janela terapêutica apresentando muitos efeitos adversos. As toxinas foram
administrada por via intratecal e apresentaram efeito antinociceptivo nos
modelos de dor visceral. A Phα1β mostrou grande eficácia e uma analgesia
longa e demorada em um modelo de dor visceral, quando comparada com a ω-
CTx-MVIIA. Além do mais, a ação analgésica de ambas toxinas estão
relacionadas coma inibição da liberação de neurotransmissores pró-

13
nociceptivos evocados por cálcio, como o glutamato ocorrendo uma diminuição
da liberação de glutamato presente no Líquido cerebro espinhal (LCE). Assim,
o presente estudo demonstrou que a toxina Phα1β apresentou efeitos
analgésicos em modelos de dor visceral, sugerindo que esta toxina pode ter
potencial para ser usada como uma droga para o controle da dor visceral.

14
1 INTRODUÇÃO
1.1 DOR E NOCICEPÇÃO
Dor é uma qualidade sensorial complexa, puramente subjetiva, difícil de
ser definida, descrita ou interpretada. Segundo o Comitê de Taxonomia da
Associação Internacional para o Estudo da Dor, a dor pode ser conceituada
como uma sensação e experiência sensorial desagradável associada a um
dano tecidual atual ou potencial, ou descrita como tal dano (Merskey & Bogduk,
1994; Millan, 1997).
A dor é extensivamente influenciada por ansiedade, depressão,
expectativa e outras variáveis psicológicas. É uma experiência multifacetada,
um entrelaçamento das características físicas dos estímulos com as funções
motivacionais, afetivas e cognitivas do indivíduo. A dor desempenha o papel
de alerta, comunicando ao indivíduo que algo está errado, podendo gerar
estresse acentuado e incapacidade física. É sem sombra de dúvida, a maior
causa de afastamento do trabalho, gerando um enorme ônus para a nação
(Bruno, 2001).
As sensações dolorosas induzem respostas urgentes de seu alívio e nos
animais provocam comportamentos como massagear ou lamber a área lesada.
A dor manifesta-se com intensidade diferente entre os indivíduos, variando de
acordo com o sexo, idade, estado de humor (Ganong, 1988; Agnati et al., 1991;
Faucett & Levine, 1991; Berkley, 1999; Chapman & Gavrin, 1999; Turk &
Okifuji, 1999; Fillingen & Ness, 2000; Sharp, 2001).
A dor, além de uma sensação, é uma experiência. Isto é importante
porque as sensações possuem vias neuroanatômicas, com receptores

15
específicos que permitem a detecção e medida de um estímulo. Já as
experiências incorporam componentes sensoriais com influências pessoais e
ambientais importantes. No entanto, clínica e experimentalmente se faz
necessária a distinção entre a dor percebida e a resposta ao dano tecidual ou
nocicepção (Kandel et al., 2003). Assim, o termo nocicepção refere-se somente
à percepção do sinal no sistema nervoso central evocado pela ativação de
receptores sensoriais especializados (nociceptores), provenientes de um tecido
danificado (Fürst, 1999).
Os animais não são capazes de verbalizar os componentes subjetivos
da dor, ou seja, neles não se avalia dor mas nocicepção. Sendo assim, termos
como dor e analgesia são mais adotados para humanos e nocicepção e
antinocicepção para animais (Jones, 1992).
1.2 CLASSIFICAÇÃO DA DOR
A dor pode ser considerada como um sintoma ou manifestação de uma
doença ou afecção orgânica, mas também pode vir a constituir um quadro
clínico mais complexo. Existem muitas maneiras de se classificar a dor.
Considerando a duração da sua manifestação, ela pode se apresentar nas
formas transitória, aguda e crônica. Na dor transitória, a ativação dos
nociceptores acontece na ausência de qualquer dano tecidual, e contribui para
proteger o organismo de potenciais danos físicos causados pelo ambiente ou
por estresse de tecidos corporais. Contudo, a dor aguda, é uma resposta
causada por uma lesão de tecido com conseqüente ativação dos nociceptores
no local da lesão, se caracteriza por ser de curta duração, desaparecendo até

16
mesmo antes da cura do dano tecidual. Além disso, a dor aguda é geralmente
de fácil identificação e tratamento, possuindo também caráter protetor
(LOESER & MELZACK, 1999; BRENNAN et al., 2007). Já a dor crônica, é
causada por uma lesão tecidual ou doença, e geralmente ultrapassa o tempo
de recuperação do organismo, ou seja, este tipo de dor pode não desaparecer
mesmo quando o trauma inicial (lesão) foi resolvido. A dor crônica se estende
por meses ou anos, é de difícil identificação, sendo uma patologia que
geralmente necessita de tratamento complexo (BRENNAN et al., 2007;
TRACEY & MANTYH, 2007). Por fim, a dor crônica é causada por uma lesão
tecidual ou doença que geralmente ultrapassa o tempo de recuperação do
organismo, ou seja, este tipo de dor não desaparece mesmo quando o trauma
inicial (lesão) foi resolvido, sendo um importante fator de incapacidade e
sofrimento (Lotsch e Geisslinger, 2001; Loeser e Melzack, 1999). Não é apenas
a duração que distingue a dor aguda da dor crônica, mas a capacidade do
organismo de reparar o sítio da lesão e restaurar os disparos aferentes e o
processamento central normal (Loeser, 2000). Pode-se destacar ainda
alterações adaptativas como a neuroplasticidade em vários níveis do sistema
nervoso tais como sensibilização, desinibição dos neurônios inibitórios do corno
dorsal, reorganização do circuito neuronal do corno dorsal e alterações na
facilitação e inibição descendente da dor. Tendo em vista que estes eventos
são dependentes da intensidade e da duração do estímulo, quanto mais
persistente for o processo doloroso, mais difícil se torna o tratamento do quadro
patológico (Besson, 1999; Woolf e Salter, 2000; Zimmermann, 2001; Wang e
Wang, 2003).

17
1.2.1 Dor visceral
Dor visceral é a forma mais comum de dor para qual os pacientes
muitas vezes procuram atendimento médico. A grande variedade de dores
viscerais incluem cirurgias abdominais, pancreatites, dismenorréia, dores do
parto e desordens funcionais como síndrome do intestino irritado e dispepsia.
Apesar dos consideráveis avanços no conhecimento sobre os mecanismos
básicos subjacentes à dor visceral e hiperalgesia visceral, não há novas
terapias eficazes para a dor abdominal.
A dor visceral é mediada por fibras aferentes do sistema nervoso
autônomo, cujos receptores se localizam na parede das vísceras ocas e na
cápsula dos órgãos parenquimatosos. É originada quando se aumenta a
tensão da parede da víscera, seja por distensão, inflamação, isquemia ou
contração exagerada da musculatura. É uma sensação dolorosa profunda, e
mal localizada, de início gradual e de longa duração. É sentida na linha
mediana do abdome devido à inervação sensorial ser bilateral; fazem exceção
as vísceras duplas como rins e ureteres, anexos uterinos onde a dor tende a
ser do lado afetado, pois nestas as vias nervosas são unilaterais.
A sensação de dor é projetada em diferentes níveis de parede
abdominal, desde o epigástrio até o hipogástrio, na dependência da origem
embriológica da víscera afetada (intestino primitivo superior no epigástrio,
médio no mesogástrio ou inferior no hipogástrio). A dor visceral pode se
associar a hiperestesia cutânea e a hiperestesia muscular.
As vísceras raramente são expostas a estímulos externos mas são
alvos comuns de diversas doenças. A sensibilidade do tecido visceral a

18
estímulos térmicos, químicos e mecânicos difere significativamente. As
vísceras parecem mais sensíveis à distensão de órgãos cavitários de parede
muscular, sem dano tecidual, isquemia, e inflamação. A área sobre a qual o
estímulo acontece pode ser uma determinante crucial no desenvolvimento dos
tipos de dor. Os receptores mecânicos ou mecanorreceptores existentes na
musculatura lisa de todas as vísceras ocas são do tipo Aδ e C, e respondem a
estímulos mecânicos leves, tensão aplicada ao peritônio, contração e distensão
da musculatura lisa. O trato gastrintestinal possui receptores químicos e
mecânicos de adaptação lenta e rápida que são classificados em dois grupos:
o grupo de receptores de alto limiar para estímulos mecânicos leves, e o grupo
de baixo limiar para estímulos mecânicos que responde a estímulos agressivos
e não agressivos. O primeiro grupo é encontrado no esôfago, sistema biliar,
intestino delgado e cólon e o segundo, apenas, no esôfago e cólon. A relação
entre a intensidade do estímulo e a atividade nervosa é somente evocada após
a estimulação nociva. (LAMONT e TRANQUILLI, 2000; KRAYCHETE e
GUIMARÃES, 2003).
1.3 MECANISMOS DA HIPERALGESIA VISCERAL- MECANISMOS PERIFÉRICOS E CENTRAIS
Sensibilização Periférica dos Neurônios Aferentes dos Intestinos. Existe
uma grande quantidade de mediadores periféricos (bradicinina, citocinas,
prostaglandinas, serotonina, ATP, prótons H+) que agem diretamente nos
receptores nociceptivos gastrintestinais e inicia a transmissão dolorosa. Podem
ativar imunócitos locais ou outras células, como mastócitos e varicosidades
simpáticas, que liberam interleucinas e substâncias adrenérgicas, perpetuando

19
e facilitando, assim, a hiperexcitabilidade neuronal. Na sensibilização periférica
ocorre, então, redução da intensidade dos estímulos necessários para iniciar a
despolarização neuronal e aumento do número ou da amplitude de descarga
neuronal, em resposta a certos estímulos químicos ou mecânicos.
1.3.1 Sensibilização Central
A sensibilização central é um processo resultante da atividade
sustentada que acontece na fibra aferente primária, após a sensibilização
periférica, favorecendo a liberação de neurotransmissores excitatórios. Estes
aumentam a 4-eficácia da transmissão sináptica entre os neurônios aferentes
primários e os do corno dorsal, envolvendo, portanto, receptores pré e pós-
sinápticos específicos.
Embora o mecanismo de sensibilização visceral central não seja
totalmente conhecido, acredita-se que alguns mediadores como a substância
P, CGRP, aspartato, glutamato, neurocininas, somatostatina e VIP estejam
envolvidos no desenvolvimento e manutenção da sensibilização central
induzida pela inflamação. A ação desses neuromediadores em receptores
específicos ionotrópicos (AMPA, cainato) e metabotrópicos (NMDA) ativa
segundos mensageiros (cAMP, PKC, fosfatidilinositol, fosfolipase C) para
abertura de canais de cálcio e entrada dessas substâncias para o interior das
membranas celulares. Ocorre então produção de outros mediadores (óxido
nítrico e metabólitos do ácido araquidônico) e formação de oncogenes (cfos,
fos B, C jun, jun B e D), que provavelmente alteram a transmissão do potencial
de ação e ultra-estrutura dos nervos e suas sinápses, sensibilização medular e
fenômeno de wind up (aumento da duração da resposta de certos neurônios).

20
Acredita-se, também, que as conexões entre estruturas espinhais e
supraespinhais, as chamadas projeções supraespinhais, estejam envolvidas no
processo de sensibilização central para hiperalgesia viscera
1.4 MECANISMOS NEURAIS DA DOR
A transmissão da dor envolve uma interação complexa de estruturas
periféricas e centrais desde a pele, vísceras e outros tecidos até o córtex
cerebral. Um nervo periférico consiste em axônios de três diferentes tipos de
neurônios: sensoriais primários, motores e pós-ganglionares simpáticos. As
terminações livres de fibras aferentes primárias sensíveis a estímulos nocivos
são chamados de nociceptores (ou receptores da dor) (MILLAN, 1999).
A sensibilização dos nociceptores se deve a diferentes estímulos, tais
como mudança de temperatura (estímulo nocivo térmico), diferença osmótica
ou distensão tecidual (estímulo nocivo mecânico), hipóxia ou lesão tecidual
seguida de inflamação (estímulo nocivo químico). (Julius e Basbaum, 2001).
Esses estímulos nocivos ativam fibras aferentes sensoriais delgadas do tipo C
e A. Além destas, outro tipo de fibra que pode estar envolvida na transmissão
do estímulo sensorial são as fibras do tipo A, que normalmente respondem a
estímulos inócuos aplicados à pele, porém em condições especiais são
capazes de conduzir rapidamente o estímulo doloroso (30-100 m/s). As fibras
A são mielinizadas e de grande diâmetro. Já as fibras do tipo A, de condução
intermediária (12 a 30 m/s), são pobremente mielinizadas, enquanto as fibras
do tipo C são não mielinizadas e transmitem o estímulo nociceptivo de forma
mais lenta (0,5 a 2,0m/s) (Figura I) (JULIUS & BASBAUM, 2001).

21
As fibras aferentes primárias C e A transmitem a informação nociceptiva
da periferia até o corno dorsal da medula espinhal, e suas terminações
encontram-se principalmente nas lâminas I (zona marginal) e lâmina II
(substância gelatinosa). No corno dorsal da medula espinhal neurônios de
primeira ordem fazem sinapse com neurônios de segunda ordem, que
compreendem as vias ascendentes. Assim esses neurônios recebem seus
sinais sensoriais pela liberação de glutamato e SP dos neurônios aferentes
primários; além disso, este processo excitatório também envolve canais de
cálcio, sendo os principais reguladores da liberação de neurotransmissores
(Hill, 2001).
Figura I: Representação esquemática dos diferentes tipos de neurônio
sensoriais primários responsáveis pela condução do sinal nociceptivo da
periferia ao SNC (Adaptado a partir de Julius e Basbaum, 2001).

22
Os neurônios de segunda ordem cruzam a medula espinhal para ascender
ao trato espinotalâmico, projetando seus corpos celulares ao tálamo. No
tálamo, neurônios de terceira ordem emitem axônios através da cápsula interna
do córtex somatosensor, onde a somatização do estímulo nocivo ocorre, ou
emitem axônios ao giro cingulado anterior responsável pelo componente
emocional da dor (Russo e Brose, 1998).
O tálamo e o córtex são regiões finais da projeção das vias de
nocicepção. O tálamo é um dos responsáveis por informar que existe sensação
nociceptiva, e o córtex é responsável pela discriminação do tipo de sensação
nociceptiva e por identificar, de forma pouco fiel, de onde provém (Guyton,
1992).
Além dessa modulação ascendente, existe uma modulação descendente
da nocicepção (Figura II). Existem casos em que pessoas sujeitas a ferimentos
dolorosos, aparentemente, não sentem dor. As vias descendentes originam-se
no tronco cerebral e outras estruturas como hipotálamo, córtex, tálamo, núcleo
magno da rafe (NMR), substância cinzenta periaquedutal (PAG) e estruturas
adjacentes da medula rostroventromedial (RVM), que exercem importante
papel na integração e modulação das mensagens nociceptivas no corno dorsal
da medula espinhal (MILLAN, 2002; VANEGAS & SCHAIBLE, 2004). Os
mecanismos descendentes modulam a resposta nociceptiva por exercer suas
ações em nociceptores presentes nas fibras aferentes primárias, bem como em
neurônios intrínsecos do corno dorsal, como interneurônios excitatórios,
interneurônios inibitórios e neurônios de projeção (MILLAN, 2002). Uma das
descobertas mais interessantes a respeito do circuito modulatório da dor é que
este pode tanto facilitar quanto inibir a transmissão nociceptiva (JULIUS &

23
BASBAUM, 1999; PORRECA et al., 2002). Por exemplo, na RVM (medula
rostral ventromcdial) estão presentes dois tipos de neurônios, as chamadas
células “liga” (on) e as células “desliga “(off), as quais estão envolvidas na
modulação nociceptiva. É proposto que as células “liga” (on) medeiam a
facilitação da condução de estímulos nociceptivos quando ativadas, e as
células “desliga” (off) medeiam a inibição da transmissão nociceptiva,
provocada pela estimulação da substância cinzenta periaquedutal (PAG). De
maneira geral, a substância cinzenta periaquedutal deve excitar as células off e
inibir as células on na medula rostroventromedial (FIELDS et al., 2006). Logo, o
balanço entre a ativação dessas duas subpopulações de neurônios determina a
resposta a um estímulo nociceptivo periférico. No entanto, em situações de dor
persistente, alterações na neuroplasticidade podem resultar em uma
estimulação facilitatória sustentada, o que ocasiona respostas persistentes e
exageradas à dor (REN & DUBNER, 2002; PORRECA et al., 2002).
Além da modulação descendente da informação nociceptiva envolver
uma série de estruturas cerebrais, como mencionado anteriormente, os
sistemas de neurotransmissores também estão envolvidos nesta conexão. Os
neurotransmissores envolvidos serotonina, noradrenalina e opióides
endógenos, parecem inibir a excitação de neurônios de segunda ordem na
presença de estímulo nocivo (FURST 1999; FIELDS, 2006).
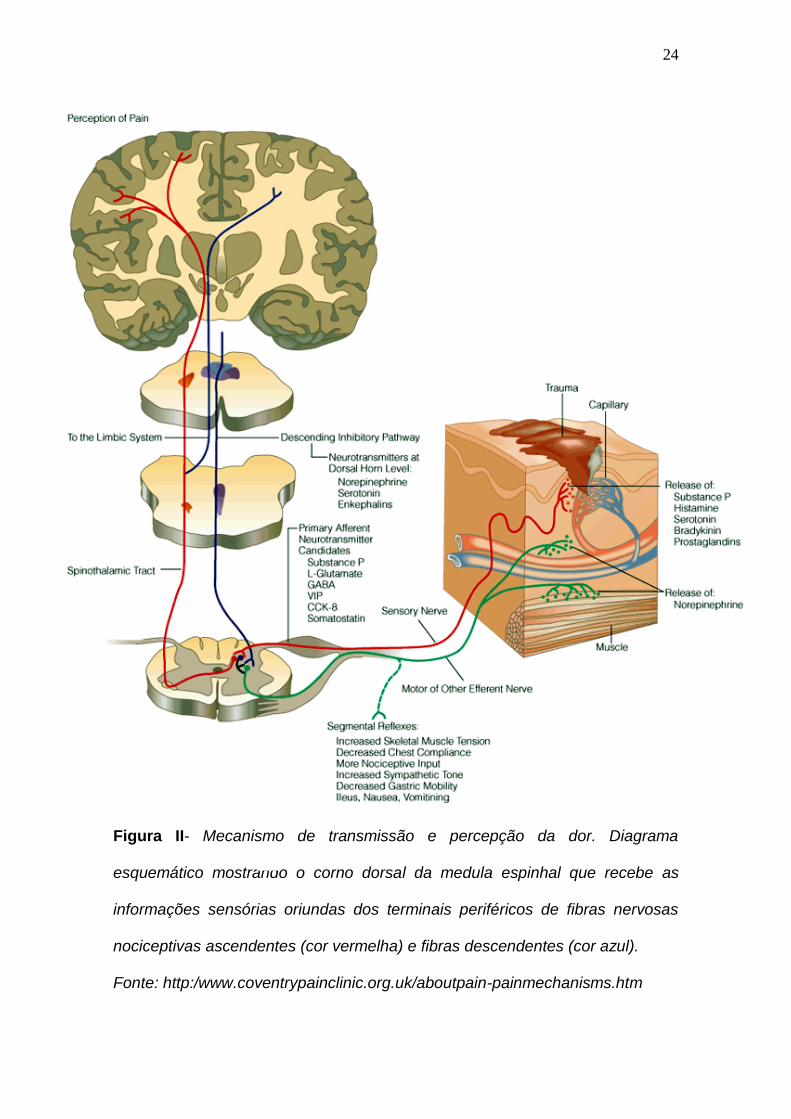
24
Figura 2- Mecanismo de transmissão e percepção da dor. Diagrama
esquemático mostrando o corno dorsal da medula espinhal que recebem as
informações sensórias oriundas dos terminais periféricos de fibras nervosas
nociceptivas ascendentes (cor vermelha) e fibras descendentes (cor azul).
Figura II- Mecanismo de transmissão e percepção da dor. Diagrama
esquemático mostrando o corno dorsal da medula espinhal que recebe as
informações sensórias oriundas dos terminais periféricos de fibras nervosas
nociceptivas ascendentes (cor vermelha) e fibras descendentes (cor azul).
Fonte: http:/www.coventrypainclinic.org.uk/aboutpain-painmechanisms.htm

25
1.5 MEDIADORES QUÍMICOS NA VIA NOCICEPTIVA
A atividade dos nociceptores é mediada pela ação de substâncias
algogênicas que são liberadas e/ou sintetizadas em elevada concentração no
ambiente tecidual na decorrência de processos inflamatórios (Cotran et al.,
1994). Substâncias endógenas como prostaglandinas, neuropeptídeos, cininas,
aminoácidos excitatórios, entre outros, são produzidas e/ou liberadas pelo
tecido lesionado e estimulam os receptores presentes na membrana dos
neurônios. Além disso, os mediadores inflamatórios liberados facilitam a
neurotransmissão e sensibilizam o nociceptor (Björkman, 1995).
Existem várias fontes importantes de mediadores químicos que
participam da resposta dolorosa, entre as quais se destacam os tecidos
lesionados e adjacentes, sistema vascular, células do sistema imunológico,
nervos simpáticos e sensoriais, entre outros. Assim, há liberação local de
diversos mediadores químicos celulares resultantes de lesão tecidual que são
capazes de desencadear uma reação inflamatória local, atraindo macrófagos e
linfócitos. Essas células liberam mediadores inflamatórios como: cininas
(bradicina e calidina); produtos das células imunes (citocinas, como as
interleucinas e o fator de necrose tumoral); aminas (serotonina, histamina) e
prostanóides. Desta forma, a injúria celular e a reação inflamatória que advém
de tal injúria expõe as fibras aferentes primárias (FaPs) a um grande número
de substâncias capazes de estimular o nociceptor. O aferente primário
transmite o impulso nociceptivo a neurônios específicos para nocicepção (EN)
ou a neurônios que também transmitem outras sensações (WDR) no corno
dorsal da medula (Millan, 1997, 1999; Calixto et al., 2000, 2001).

26
A transferência sináptica de informação nociceptiva é comandada pela
liberação de neurotransmissores, como glutamato que está presente em todos
os tipos de aferentes primários. O glutamato, aminoácido excitatório é
encontrado em proporções consideráveis na medula espinhal, originado de
fibras aferentes primárias mielinizadas e não-mielinizadas, em adição a
interneurônios intrínsecos e projeção de neurônios (Battaglia & Rustioni, 1988).
O glutamato e alguns neuropeptídeos são liberados juntos de terminais
aferentes primários e têm ações fisiológicas distintas nos neurônios pós-
sinápticos, atuando coordenadamente para regular as propriedades desses
neurônios (Kandel et al., 2000).
As ações do glutamato é mediada principalmente por receptores
ionotrópicos NMDA e não-NMDA. Os receptores não-NMDA consistem de dois
receptores, AMPA e cainato (Dickenson, 1995). A transmissão de fibras C,
depois de estímulos agudos mecânicos ou térmicos, parece envolver
receptores AMPA para produzir excitações curtas e constantes. Os estímulos
são mantidos e/ou sua freqüência é aumentada pela liberação de
transmissores que contribuem para a transmissão nociceptiva quando o
receptor NMDA é ativado, aumentando resposta de hiperalgesia (Dickenson,
1995). Uma ação excitatória direta do glutamato nas fibras aferentes primárias,
é consistente com a evidência que a ativação dos receptores NMDA causa a
liberação de substância P por seus terminais centrais (Liu et al., 1994). Essa
ação do NMDA pode ser mediada pelo óxido nítrico (NO) dos terminais das
fibras aferentes primárias (Sorkin, 1993), e o NO poderia também interferir nas
ações periféricas mediadas pelas FaPs (Jackson et al., 1995).

27
O glutamato induz também a liberação de SP (substancia P) das FaPs
(definir) de terminais simpáticos, de tal forma que este peptídeo também pode
estar envolvido na nocicepção induzida por glutamato. Da mesma forma que o
glutamato, a SP pode ativar as FAPs em vários tecidos (Inoeu et al., 1995;
Carlton et al., 1996) e, conseqüentemente, causar dor (Edvinsson et al., 1997).
O controle inibitório descendente da transmissão da dor também envolve
neurotransmissores serotonina, noradrenalina, entre outros e opioides
endogenos que parecem inibir a excitação dos neurônios do corno dorsal da
medula na presença do estímulo nocivo (Russo & Brose, 1998; Furst, 1998).
Assim, os sinais dolorosos podem ser suprimidos antes de chegar ao cérebro.
O sistema descendente inibitório estimula neurônios medulares a secretar
opióides endógenos, que causam a inibição pré-sináptica e pós-sináptica das
fibras C e A (Besson, 1999), portanto, o mecanismo pelo qual provocam essa
analgesia descendente envolve a liberação de opióides na medula espinhal.
Provavelmente, o mecanismo que produz essa analgesia é através do bloqueio
de canais de Ca²+. Como o aumento da concentração do Ca²+ intracelular é o
mecanismo de ação do neurotransmissor glutamato, o bloqueio dos canais de
Ca²+ resultaria em inibição pré-sináptica (Kuraishi et al.,1985)
Estudos farmacológicos, eletrofisiológicos e anatômicos contribuiram
para o descobrimento de múltiplos mediadores químicos envolvidos na dor,
facilitando o entendimento dos mecanismos de ação dos neurotransmissores e
das drogas envolvidas na modulação central e periférica da dor (Levine e
Taiwo, 1994; Wood e Docherty, 1997; Millan, 1999).
Com o objetivo de encontrar drogas eficazes, com baixos efeitos
colaterais, muitos trabalhos de pesquisa estudam os mecanismos celulares e

28
moleculares envolvidos na origem da dor. De fato, atualmente, não existe
tratamento satisfatório e nem medidas adequadas e específicas para o controle
da dor (Kingery, 1997; Woolf e Mannion, 1999; Mendell e Sahenk, 2003).
Cresce o interesse pelo esclarecimento da possível participação de canais
de cálcio na dor neuropática. O íon cálcio entra nas terminações nervosas
através de canais de cálcio e regulam muitas funções, incluindo proteínas
relacionadas ao crescimento.
1.6 CANAIS DE CÁLCIO SENSÍVEIS À VOLTAGEM
Os canais de cálcio sensíveis à voltagem (CCSV) foram primeiramente
identificados em músculo de crustáceo por Paul Fatt e Bernard Katz (1953).
Estes canais exercem papel importante em vários eventos fisiológicos (Caterall,
1995; Dunlap e cols., 1995). Transcrição gênica, contração muscular, liberação
de neurotransmissores e regulação da excitabilidade neuronal são todas
respostas celulares mediadas por Ca2+ (Doering e Zamponi, 2003; Klugbauer e
cols., 2003). Além do envolvimento em eventos fisiológicos, os canais de Ca2+
também estão envolvidos em numerosas doenças, tais como, epilepsia,
hipertensão, isquemia e alguns tipos de arritmias (Jen, 1999; Dworakowska e
Dolowy, 2000; Snutch e cols., 2001).

29
1.6.1 SUBUNIDADES MOLECULARES DOS CANAIS DE CÁLCIO
Os canais de Ca2+ são membros de uma superfamília de canais
catiônicos sensíveis à voltagem (Richards e cols., 2004). Eles são proteínas
complexas compostas por 4 ou 5 diferentes subunidades: α1, β1, α2-δ e γ ver
figura 3). A subunidade α1 é a maior das subunidades com peso molecular de
190-250 kDa (revisto por Herlitze e cols., 2003). Ela incorpora o poro condutor,
o sensor de voltagem, o aparato de gating e os sítios de regulação do canal por
segundos messageiros, drogas e toxinas. A subunidade α1 é estruturalmente
organizada em 4 domínios homólogos (domínio I a IV), cada um contendo 6
segmentos transmembrana (S1-S6). A alça do poro entre os segmentos S5 e
S6 de cada domínio determina a condutância iônica e a seletividade. Assim,
mudanças em 3 aminoácidos na alça do poro entre os domínios I, III e IV
convertem um canal seletivo à sódio para um canal seletivo à cálcio (Herlitze e
cols., 2003; Klugbauer e cols., 2003). As subunidades acessórias ou
auxiliares não formam poros, mas são capazes de modificar várias
propriedades do canal formado pela subunidade 1.
1.7 CANAIS DE CÁLCIO SENSÍVEIS À VOLTAGEM (CCSV) E NOCICEPÇÃO
Entre outros fatores a regulação da liberação de neurotransmissores
rege o desenvolvimento de agentes terapêuticos direcionados contra subtipos
específicos de canais de Ca2+ sensíveis à voltagem. A liberação de
neurotransmissores no SNC depende de múltiplos subtipos de canais de Ca2+ e

30
a complementaridade de ação desses canais varia de uma população de
sinapses para a outra (Wheeler et al., 1994).
A diversidade de genes que codificam canais de Ca2+ gera diferentes
subtipos de canais com diferentes funções fisiológicas. Isso sugere a
possibilidade de uso de antagonistas subtipo-específicos para esses canais,
como novos agentes no tratamento para algumas neuropatologias (Miljanich
and Ramachandran, 1995). Em estudos com humanos avalia-se o efeito
desses agentes na prevenção da degeneração neuronal oriúnda de traumas
cerebrais isquêmicos e na persistência da dor.
Vários autores relatam o envolvimento dos íons cálcio na regulação de
diferentes processos biológicos como a excitabilidade das membranas,
nocicepção e antinocicepção (Zamponi & Snutch, 1998. Substâncias que
modulam a ação destes canais têm sido investigadas no estudo da nocicepção
e antinocicepção. Os canais de cálcio do tipo L e N estão diretamente
implicados na liberação de neurotransmissores e de neuromoduladores, como
o CGRP (Calcitonin gene related peptide), nos neurônios sensoriais da medula
espinhal. O bloqueio dos canais de cálcio sensíveis à voltagem do tipo N e P/Q,
mas não do tipo L, reduz os sinais comportamentais da dor de origem
neuropática em modelos animais de lesão nervosa, porém, a ausência de
antagonistas seletivos para os diferentes tipos de canais de Ca2+ torna difícil o
entendimento de suas ações nos processos de dor crônica (Matthews e
Dickenson, 2001; Sekizawa et al., 2000; Dogrul et al, 2003). Os canais do tipo
N estão presentes no terminal pré-sináptico de neurônios nociceptivos no corno
dorsal da medula espinhal, onde regulam a liberação de neurotransmissores
pró-nociceptivos como o glutamato e a substância P (WEN et al., 2005).

31
Experimentos realizados com antagonistas de CCVS indicam que os
canais dos tipos L, N, P/Q e T estão envolvidos na nocicepção (PRADO, 2001).
Atualmente, cresce o interesse pelo esclarecimento da participação dos canais
de cálcio nos processos de dor crônica. O íon Ca2+ entra nas terminações
nervosas através de canais e regula muitas funções, incluindo proteínas
relacionadas ao crescimento.
Pesquisas da última década descrevem o papel de inibidores de canal de
cálcio tipo N para o desenvolvimento de novas drogas analgésicas. (Altier and
Zamponi, 2004). Demonstrou-se que bloqueadores de CCSV tipo N, como as
-conotoxinas, previnem ou atenuam a dor. A -conotoxina- MVIIA (-Ctx-
MVIIA), obtida do Conus Magnus, é inibidora seletiva de CCSV tipo N, foi a
primeira toxina a ser aprovada pelo FDA para o tratamento de dor crônica
intratável. Prialt® é o nome comercial da -Ctx-MVIIA (também conhecida como
ziconotida e SNX-111), teve seu uso clínico aprovado em Dezembro de 2004
nos Estados Unidos e em Fevereiro de 2005 na Europa (Hogg, 2006). A
ziconotida produz antinocicepção em modelos de dor aguda e crônica em
animais quando administrada por via intratecal (i.t) (Malmberg and Yaksh,
1994; Malmberg and Yaksh, 1995; Bowersox et al., 1996). Entretanto, a
administração i.t de ziconotida causa efeitos adversos (Penn and Paice, 2000).
Devido a isso, há a necessidade de investigar outros inibidores de CCSV para
o tratamento da dor com menores efeitos adversos.

32
1.8 USO DE TOXINAS ANIMAIS COMO AGENTES TERAPÊUTICOS
Estudos indicam que peptideos naturais de venenos de diferentes
espécies de animais teriam aplicações terapeuticas. Além de possíveis alvos
terapêuticos, esses peptídeos são usados como ferramentas farmacológicas e
dessa forma modulam ou auto- regulam os canais iônicos (Rajendra, 2004).
O veneno do caramujo marinho Conus magnus contém uma mistura
diversa de agentes farmacologicamente ativos que exercem suas funções em
receptores e canais iônicos (Jones e cols., 2000). Entre eles a conotoxina SNX-
111, bloqueadora dos canais de cálcio do tipo N, apresenta propriedades
analgésicas e neuroprotetoras atribuídas à inibição da liberação de
neurotransmissores e a supressão de conseqüências secundárias do influxo
excessivo de cálcio. O análogo sintético dessa toxina, a Ziconotida, apresentou
eficácia na dor neuropática e isquemia cerebral(Bowersox e cols., 1998). O
análogo sintético dessa toxina, a Ziconotida, apresentou eficácia na dor
neuropática e isquemia cerebral (Bowersox e cols., 1998).
1.8.1 Phoneutria nigriventer
A aranha Phoneutria nigriventer tem ocorrência desde o sul do Rio de
Janeiro até o Uruguai. É a aranha mais comumente envolvida em
envenenamento humano no Brasil (Eickstedt, 1983 e Lucas, 1988), é muito
agressiva e é conhecida como aranha armadeira pela posição que toma ao se
sentir ameaçada, possui hábitos noturnos e permanece refugiada durante o
dia. Não constrói teia e seu sucesso como predadora pode ser explicado pela
potência das diversas toxinas presentes em seu veneno (Gomez e cols., 2002).

33
Acidentes com picadas da aranha P. nigriventer causam severa dor e
sintomas tóxicos tais como, cãimbras, tremores, convulsão tônica, paralisia
espástica, priapismo, arritmias, distúrbios visuais e sudorese (Lucas, 1998).
Esses sintomas são mais severos em crianças e a intoxicação pode levar a
morte se não tratada.
Baseado nos sintomas observados em humanos e nos resultados de
injeções intracérebro-ventriculares em animais experimentais, viu-se que o
veneno de Phoneutria é, sobretudo, neurotóxico. Parte do efeito neurotóxico do
veneno parece estar relacionado à sua ação sobre canais de Na+ regulados por
voltagem (Araujo et al., 1993; Romano-Silva et al., 1993). Entretanto, outras
atividades farmacológicas, relacionadas à ação em canais iônicos, podem ser
encontradas em todo o veneno. Essa observação pode ser mais facilmente
comprovada pelo fracionamento do veneno, em que se separa as frações com
ações farmacológicas distintas.
1.9 AÇÕES DAS TOXINAS PURIFICADAS DO VENENO DA PHONEUTRIA
NIGRIVENTER
As toxinas obtidas do veneno da aranha Phoneutria nigriventer têm sido
extensivamente investigadas e cerca de 17 peptídeos com atividade tóxica já
foram descritos na literatura (revisado por Gomez e cols., 2002). As toxinas são
basicamente polipetídeos com peso molecular entre 3500 e 9000 muitos deles
interagindo com canais iônicos. Assim, algumas classes de toxinas podem
afetar o funcionamento de canais de sódio (Na+), cálcio (Ca2+), potássio (K+),

34
dentre outros (Grishin, 1999). A caracterização bioquímica e farmacológica de
neurotoxinas do veneno desta aranha é ferramenta de investigação das
funções dos canais iônicos em níveis moleculares e celulares (Diniz e cols.,
1990; Araújo e cols., 1993; Romano-Silva e cols., 1993; Cassola e cols., 1998).
Investiga-se o veneno de P. nigriventer por sua habilidade em alterar grande
número de sistemas fisiológicos, em particular os relacionados a dor e a
inflamação (Costa e cols., 2002).
Um dos efeitos principais do veneno de P. nigriventer é a sua ação sobre
os diferentes subtipos de canais de Ca2+. Trabalhos publicados descrevem a
ação de algumas toxinas de P. nigriventer na inibição das correntes de Ca2+
(Cassola e cols., 1998; Leão e cols., 2000) ou no bloqueio do influxo deste íon
em terminais nervosos (Prado e cols., 1996; Guatimosim e cols., 1997; Miranda
e cols., 1998). A outra fração tóxica, PhTx3, administrada i.c.v. em ratos
causou paralisia flácida (Rezende Junior et al., 1991), possivelmente em
consequência da ação inibitória sobre a liberação de neurotransmissores
(Gomez et al., 1995; Prado et al., 1996). Seis diferentes toxinas (Tx3-1 a 6)
foram purificadas a partir da fração PhTx3 (Cordeiro Mdo et al., 1993) e pelo
menos 3 delas (Tx3-3, Tx3-4 e Tx3-6) bloquearam o influxo de Ca2+ induzido
por KCl em terminais nervosos (Prado et al., 1996; Guatimosim et al., 1997;
Miranda et al., 1998), sugerindo que essas toxinas bloqueiam canais de Ca2+ e
são, portanto, novas ω-conotoxinas. Além de ter toxinas que interagem com
canais de Na+ e Ca2+, uma das toxinas da Phoneutria, a toxina PhTx3-1 é uma
bloqueadora potente e seletiva de canais de K+ Tipo-A, aumentando a
frequência de oscilações de Ca2+ em células GH3 (Kushmerick et al., 1999).

35
Outra toxina, a toxina PhTx3-2, é capaz de bloquear, ainda que parcialmente,
canais de Ca2+ Tipo-L (Kalapothakis et al., 1998).
Das toxinas bloqueadoras de canais de Ca2+, isoladas do pool PhTx3, a
toxina PhTx3-3 foi uma das primeiras a ser estudada. Dados farmacológicos
iniciais sugeriram que essa toxina inibe a liberação de aceltilcolina em fatias de
córtex e em neurônios do plexo mientérico, mas essa ação não possuia um
efeito aditivo com a ω-agatoxina IVA na inibição da liberação de ACh (Gomez
et al., 1995). Além disso, a PhTx3-3 é bloqueadora eficaz da liberação de
glutamato (Prado et al., 1996). Através do uso dos marcadores fluorescentes
Fura2-AM e FM1-43 em sinaptosomas cérebro-corticais, para monitorar
medidas de Ca2+ interno e de liberação de vesículas, respectivamente,
observou-se que a PhTx3-3 inibe com alta potência (IC50 0,9nM) canais de Ca2+
que regulam a entrada de Ca2+ em sinaptosomas e também bloqueia a
exocitose de vesículas sinápticas, sendo que esse efeito se dá em canais de
Ca2+ que também são inibidos pela ω-agatoxina IVA (Guatimosim et al., 1997).
Análises de mais dados demonstraram que a PhTx3.4 bloqueia canais
de cálcio subtipo N (Cassola et al.,1998; Dos Santos et.al, 2002) e P/Q (Dos
Santos e cols., 2002). Esta toxina parece ligar-se em vários sítios de ligação,
sendo parcialmente deslocada pelas ω-conotoxinas GVIA e/ou MVIIC (Dos
Santos e cols., 2002). A toxina PhTx3-4 possui um intrigante mecanismo de
ação, uma vez que essa toxina também diminui a liberação de glutamato
dependente e independente de Ca2+ em teminações nervosas de ratos (Reis e
cols., 1999) e atuan sobre certas condições neuropatológicas como na
isquemia in vitro. Os resultados (Pinheiro et al., 2006) apontam para novas
toxinas eficazes no controle da degeneração neuronal após insultos isquêmicos

36
vinda de diversos estímulos nóxicos. Também foi investigado a interação entre
a toxina Phα1β (PhTx3-6) e os vários canais de Ca2+, determinando a potência,
seletividade e o possível mecanismo de ação da toxina, estudado em sistemas
heterólogos que expressavam canais de Ca2+ recombinantes.
A Phα1β inibe de maneira reverssível as correntes dos canais de calcio
dos subtipos L-, N-, P/Q- e R- em ordem de potência N> R> P/Q>L e valores de
IC50 = 122nM; 136nM; 263 nM e 607 nM, respectivamente. Seu efeito parece
atuar como uma -conotoxina (Vieira et al, 2005). Como é demonstrado que os
bloqueadores do subtipo N têm utilidade farmacológica no tratamento da dor, a
toxina Phα1β possui potencial analgésico interessante.

37
2. OBJETIVOS
2.1 OBJETIVO GERAL
Investigar a ação antinociceptiva espinhal da
toxina Ph1 em camundongos utilizando modelos de dor visceral periferica.
2.2 OBJETIVOS ESPECÍFICOS
1) Avaliar o efeito dose-dependente da administração intratecal das toxinas
Ph1 e -Ctx-MVIIA sobre a nocicepção química utilizando o modelo de
contorções abdominais induzidas por ácido acético;
2) Avaliar o efeito dose-dependente da administração intratecal das toxinas
Ph1 e -Ctx-MVIIA sobre a nocicepção química usando o modelo de
contorçoes abdominais induzidas por capsaicina ( intracolônico).
3) Avaliar o efeito do tratamento com Ph1 e MVIIA sobre os niveis de
glutamato e ROS do liquido cerebro espinhal de camundongos
submetidos a dor visceral induzida por acido ácetico intraperitonial.

38
3 MATERIAL E MÉTODOS
3.1 Materiais
Drogas e Toxinas
A toxina Ph1 purificada do veneno da aranha Phoneutria nigrivienter
foi purificada nos laboratórios da FUNED em colaboração com a Dra. Marta N.
Cordeiro. A ω-conotoxina MVIIA (-Ctx-MVIIA) foi obtida da Latoxan (Valence,
França). As soluções estoque das drogas foram preparadas utilizando solução
de PBS q.s.p. em tubos plásticos (eppendorf) siliconizados e foram mantidas a
-20ºC, bem como diluídas para a concentração desejada imediatamente antes
do uso. Capsaicina (8-methyl N-vanilil-6-noneamida, Sigma, EUA) foi dissolvida
em veículo composto por etanol (10%), Tween 80 (10%) e PBS q.s.p. Ácido
acético glacial 100% (Merck, EUA) foi dissolvido em PBS q.s.p. para a
concentração final de 0,6%.
Animais
Foram utilizados camundongos Swiss (20-25g) adultos, machos
provenientes do Biotério Central da Universidade Federal de Minas Gerais.
Esses animais foram mantidos em ciclo 12 h luz/escuro com temperatura
ambiente constante em 22±1ºC e com livre acesso à água e comida. Este
trabalho foi submetido e aprovado pelo Comitê de Ética do IEP/ Santa Casa de
Belo Horizonte numero 179 e os experimentos foram realizados em
concordância com as recomendações dos cuidados com animais em
experimentação (Zimmermann,1983).

39
3.2 MÉTODOS
Administração Intratecal (i.t)
As injeções i.t foram realizadas de acordo com o método descrito por
Hylden e Wilcox (1980) para camundongos. Brevemente, um volume de 5 L
para camundongos foi administrado com uma microseringa Hamilton, enquanto
o animal foi gentilmente imobilizado para ser mantido na posição necessária
para administração intratecal entre as vértebras L5-L6. A perfuração da dura foi
indicada comportamentalmente por um rápido movimento da cauda.
Administração intracolônica (i.c.)
A administração i.c. foi realizada conforme descrito por Laird et al. (2001)
em camundongos. O volume de 25 µL foi administrado pela introdução de uma
cânula com a extremidade arredondada (0.61 mm de diâmetro externo e 4 cm
de comprimento com quatro orifícios distais para melhor dispersão da
substância) no cólon descendente a 3.5 cm do ânus. Foi realizado um
treinamento do examinador para garantir que o volume seria injetado na área
de interesse sem lesionar o animal. Também, Azul de Evans foi injetado i.c. e ,
em seguida, o animal foi sacrificado e dissecado, confirmando que o volume foi
administrado no cólon descendente.

40
3.2.1 TESTES COMPORTAMENTAIS
Teste das contorções abdominais induzidas por ácido acético:
o teste de contorção abdominal em camundongos é um método muito
utilizado para se avaliar a atividade analgésica de substâncias contra dor
visceral de origem inflamatória, onde o ácido acético na concentração de 0,6%
(v/v), induz lesão no abdômen do camundongo, suficiente para provocar os
espasmos traduzidos como contorção (Koster & Anderson, 1959).
Para avaliar se a Ph1 e -Ctx-MVIIA previnem o aparecimentodas
contorções e da nocicepção, os Camundongos receberam administração
intratecal de Ph1 (10-200 pmol/sítio), -Ctx-MVIIA (1-100 pmol/sítio) ou
veículo (PBS, 5L/sítio). Após 2 horas os animais receberam uma injeção
intraperitoneal de ácido acético 0,6% (v/v; 0.1ml/10g peso corporal). Após a
injeção do ácido acético os camundongos foram colocados em caixas de
acrilico individuais, e o número de contorções abdominais foi quantificado
cumulativamente durante um período de 30 min. As contorções abdominais
consistem na contração da musculatura abdominal juntamente com a extensão
de uma das patas posteriores de acordo com o método descrito anteriormente
(Collier et al., 1968; Santos et al., 1999 a; Le Bars et al., 2001).

41
Mensuração do comportamento nociceptivo relacionado a dor visceral
por estimulação química do cólon
Para avaliar outro modelo de dor visceral os Camundongos receberam
administração intratecal de Ph1 (10-200 pmol/sítio), -Ctx-MVIIA (1-100
pmol/sítio) ou veículo (PBS, 5L/sítio). Após 2 horas os animais foram
submetidos à administração intracolônica de 25L de capsaicina 0,3% e, em
seguida os camundongos foram colocados em caixas de acrilico individuais, e
o comportamento nociceptivo foi observado, diretamente, por um período de 30
min. Foi considerado como comportamento relacionado a nocicepção as
seguintes posturas: 1) lambidas do abdômen (o comportamento de lambedura,
neste modelo foi definida com o camundongo fazendo uma flexão para a frente,
trazendo o focinho mais perto do abdômen e lambendo esta região), 2)
estiramento do abdômen, 3) esmagamento do abdômen contra o fundo da
caixa e 4) retrações abdominais (Laird et al., 2001)
Dosagem de Glutamato no Líquido cerebroespinhal
Para avaliar se a Ph1 é capaz de inibir a liberação de glutamato após
o estímulo nociceptivo já instalado, camundongos receberam por via intratecal
Ph1 (100 pmol/sítio), -Ctx-MVIIA (30 pmol/sítio) ou veículo (PBS, 5
L/sítio) 2 horas antes da administração i.p de acido acetico 0,6% (v/v;
0.1ml/10g peso corporal). Após 30 minutos o animal foi sacrificado com
halotano e em seguida seu líquido cerebroespinhal foi retirado com uma
punção na cisterna magna. Os níveis de glutamato foram dosados
enzimaticamente a partir do aumento da fluorescência devido à produção de

42
NADPH+ na presença de glutamato desidrogenase (GDH) e NADP+, de acordo
com metodo descrito por (Nicholls et al., 1987). Incialmente adicionou-se
NADP+ (1.0 mM) e 50 U de GDH pelo menos 10 minutos antes da adição das
amostras de liquido cerebro espinhal (LCE). A excitação foi fixada em 360 nm e
a emissao em 450 nm utilizando um espectroflurimetro marca Shimadzu,
modelo RF-5301.
Avaliação de ROS do líquido cerebroespinhal após indução da dor viceral
Para verificar se o efeito antinociceptivo da Ph1 está relacionado com
uma redução de ROS das amostras de líquido cerebroespinhal, camundongos
receberam por via intratecal Ph1 (100 pmol/sítio), -Ctx-MVIIA (30
pmol/sítio) ou veículo (PBS, 5 L/sítio) 2 horas antes da administração i.p de
acido acetico 0,6% (v/v; 0.1ml/10g body weight). Após 20 minutos o animal foi
sacrificado com halotano e em seguida seu líquido cerebroespinhal foi retirado
com uma punção na cisterna magna. Para avaliar a variação de fluorescência
em função do tempo decorrente da oxidação da diclorofluoresceína cuja
fluorescência aumenta em virtude da presença de espécies reativas de
oxigênio (ROS). Aproximadamente 20 µL de líquido cerebroespinhal eram
retirados por animal. Após retirado o líquido cerebroespinhal era centrifugado a
10000 rpm por 1 minuto e o sobrenadante era coletado e utilizado para os
testes de ROS. O método para dosagem de ROS foi baseado em Loechutinati
e cols (2011), com pequenas adaptações. Basicamente, 5uL da amostra de
líquido cerebroespinhal eram adicionados a 2 mL de solução PBS (Phosphate
Saline Buffer), transferidos para cubeta de acrílico e levados para leitura de
fluorescência ao longo do tempo em espectrofluorímetro marca Shimadzu,

43
modelo RF-5301 (Excitação 502nm, emissão 523nm), com frequência de
leitura de 1 leitura por segundo. Após estabelecer a linha de base era
adicionado Diclorofluoresceína (concentração final de 100nM). Os níveis de
fluorescência desse composto são elevados devida a sua oxidação, causada
principalmente pelas espécies reatívas de oxigênio da amostra. A fluorescência
foi registrada continuamente por 8 minutos após adição da diclorofluoresceína.
Os níveis de fluorescência dos 2 últimos segundos da amostra de líquido
cerebroespinhal vindo do grupo de animais que recebeu ácido acético
intraperitoneal e PBS intratecal, foram considerados como 100% de produção
de ROS, enquanto que o basal de produção de ROS, aqui considerado como
0%, foi o final da leitura de fluorescência do líquido cerebroespinhal no grupo
de animais que recebeu PBS intraperitoneal. A comparação estatística dos
grupos foi feita pelo teste ANOVA 1 via seguido do teste de Tukey.

44
4 RESULTADOS
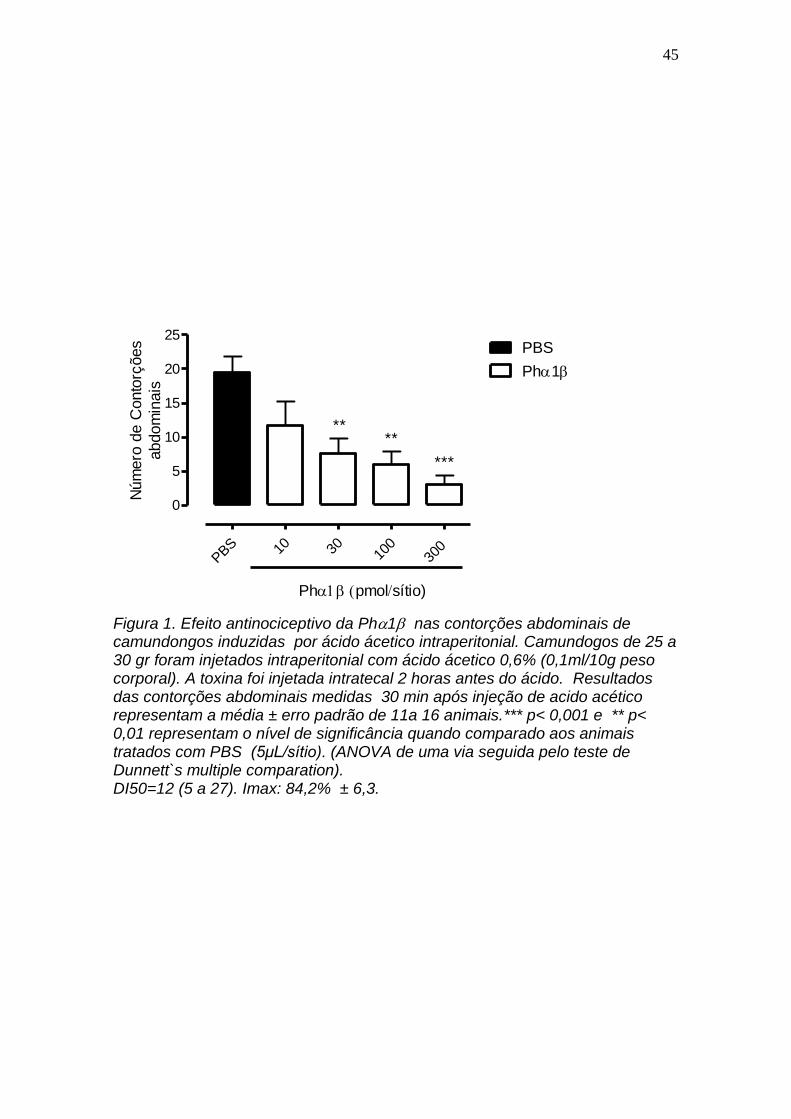
4.1 Avaliação do efeito antinociceptivo da Ph1 nas contorções abdominais de camundongos induzidas por ácido ácetico intraperitonial
A figura 1 mostra que o efeito inibitório da toxina Ph1 nas contorções
abdominais em camundongos induzidas por ácido acético é dependente da
concentração. Desse modo, nas concentrações de 10, 30, 100 e 300 pmol a
toxina Ph1 inibiu em torno de 31, 61, 69 e 84%, respectivamente, as
contorções abdominais induzidas pelo acido acético injetado intraperitonial. A
dose inibitoria 50% foi 12 (5 a 27). A inibição máxima foi de 84,2 6.3 na
concentração de 300 pmol. Na concentração de 10 pmol o efeito inibitório da
toxina não foi estatisticamente significante P > 0.05.
45
PBS 10 30 10
030
0
0
5
10
15
20
25
****
***
PBS
Ph1
Phpmolsítio)
Núm
ero
de C
onto
rções
abdom
inais
Figura 1. Efeito antinociceptivo da Ph1 nas contorções abdominais de camundongos induzidas por ácido ácetico intraperitonial. Camundogos de 25 a 30 gr foram injetados intraperitonial com ácido ácetico 0,6% (0,1ml/10g peso corporal). A toxina foi injetada intratecal 2 horas antes do ácido. Resultados das contorções abdominais medidas 30 min após injeção de acido acético representam a média ± erro padrão de 11a 16 animais.*** p< 0,001 e ** p< 0,01 representam o nível de significância quando comparado aos animais tratados com PBS (5μL/sítio). (ANOVA de uma via seguida pelo teste de Dunnett`s multiple comparation). DI50=12 (5 a 27). Imax: 84,2% ± 6,3.

46
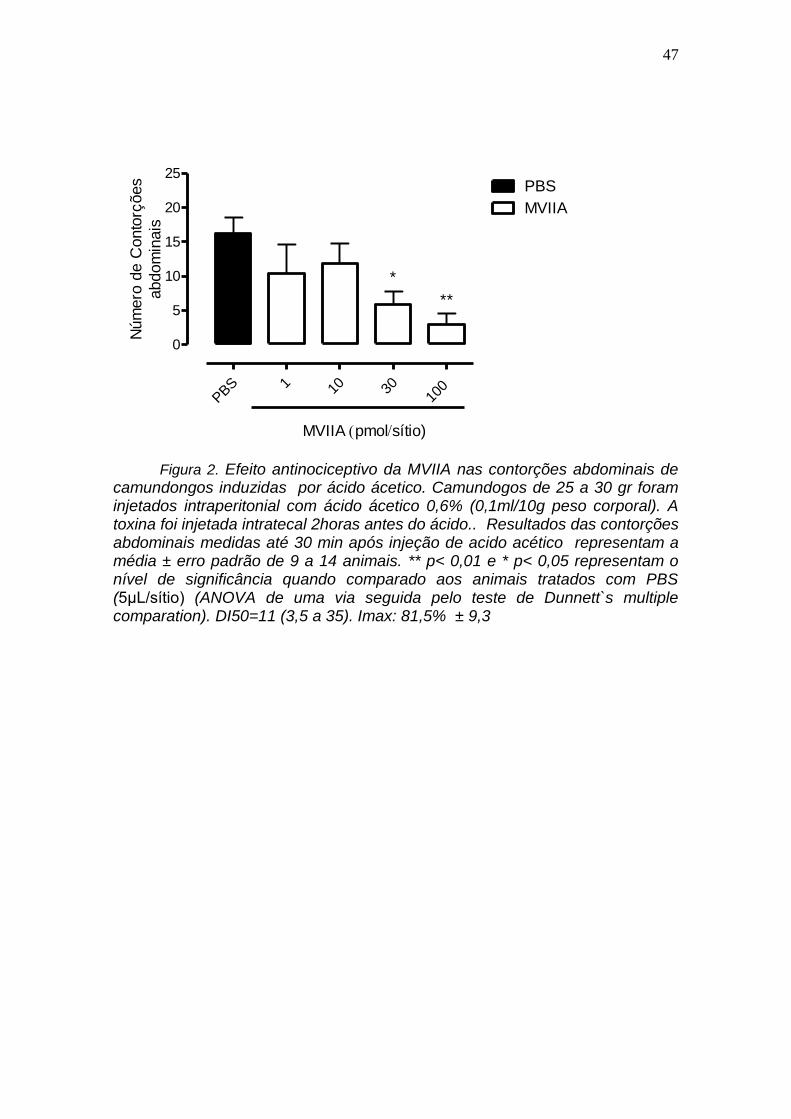
4.2 Avaliação do efeito antinociceptivo da MVIIA das contorções abdominais de camundongo induzido por ácido ácetico
A figura 2 mostra que o efeito inibitório da toxina -conotoxina MVIIA nas
contorções abdominais em camundongos induzidas por acido acético é
tambem dependente da concentração. Nas concentrações 1,0, 10, 30 e 100
pmol a toxina -conotoxina MVIIA inibiu em torno de 36, 29, 64,5 e 81,5,
respectivamente ác. acé, as contorções abdominais, induzidas por tico. A dose
inibitoria 50% foi de 11 (3,5 a 35) e a inibição máxima 81.5 9,3. Não foi
estatisticamente sigficante as inibições com as concentrações de 1 e 10 pmol
de -conotoxina MVIIA.
47
PBS 1 10 30
100
0
5
10
15
20
25
*
**
PBS
MVIIA
MVIIApmolsítio)
Núm
ero
de C
onto
rções
abdom
inais
Figura 2. Efeito antinociceptivo da MVIIA nas contorções abdominais de camundongos induzidas por ácido ácetico. Camundogos de 25 a 30 gr foram injetados intraperitonial com ácido ácetico 0,6% (0,1ml/10g peso corporal). A toxina foi injetada intratecal 2horas antes do ácido.. Resultados das contorções abdominais medidas até 30 min após injeção de acido acético representam a média ± erro padrão de 9 a 14 animais. ** p< 0,01 e * p< 0,05 representam o nível de significância quando comparado aos animais tratados com PBS (5μL/sítio) (ANOVA de uma via seguida pelo teste de Dunnett`s multiple comparation). DI50=11 (3,5 a 35). Imax: 81,5% ± 9,3

48
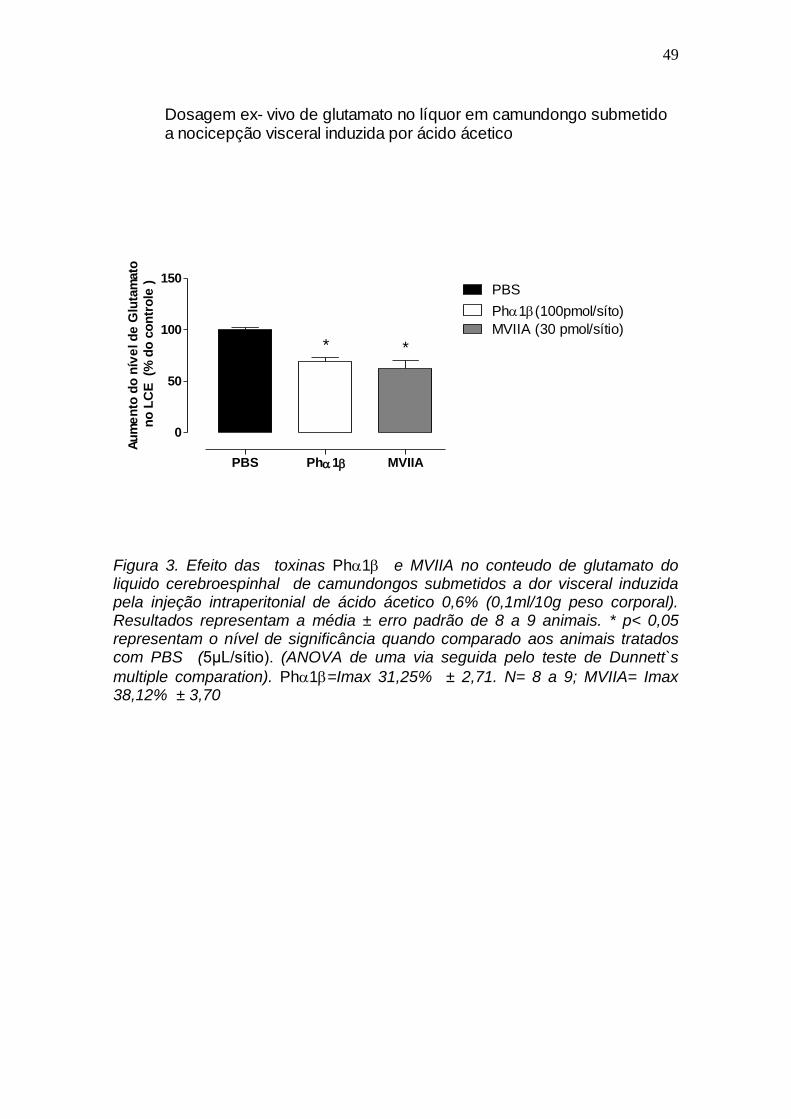
4.3 Efeito das toxinas Ph1 e MVIIA no conteudo de glutamato do liquido cerebroespinhal de camundongos submetidos a dor visceral induzidas pela injeção intraperitonial de ácido ácetico.
A figura 3 mostra o efeito das toxinas Ph1 (100 pmol) e -conotoxina
MVIIA (30 pmol) nos niveis de glutamato do liquido cerebroespinhal de
camundongos submetidos a dor visceral induzida por ácido acético. Ambas
toxinas diminuiram a liberação de glutamato com Imax de 31,25% 2,7 para a
Ph1 e 38,1% 3,7 para a -conotoxina MVIIA, P < 0,05.
49
PBS Ph1 MVIIA
0
50
100
150
**
PBS
Ph1(100pmol/síto)
MVIIA (30 pmol/sítio)
Dosagem ex- vivo de glutamato no líquor em camundongo submetidoa nocicepção visceral induzida por ácido ácetico
Au
me
nto
do
nív
el d
e G
luta
mato
no
LC
E (%
do
co
ntr
ole
)
Figura 3. Efeito das toxinas Ph1 e MVIIA no conteudo de glutamato do liquido cerebroespinhal de camundongos submetidos a dor visceral induzida pela injeção intraperitonial de ácido ácetico 0,6% (0,1ml/10g peso corporal). Resultados representam a média ± erro padrão de 8 a 9 animais. * p< 0,05 representam o nível de significância quando comparado aos animais tratados com PBS (5μL/sítio). (ANOVA de uma via seguida pelo teste de Dunnett`s
multiple comparation). Ph1=Imax 31,25% ± 2,71. N= 8 a 9; MVIIA= Imax 38,12% ± 3,70

50
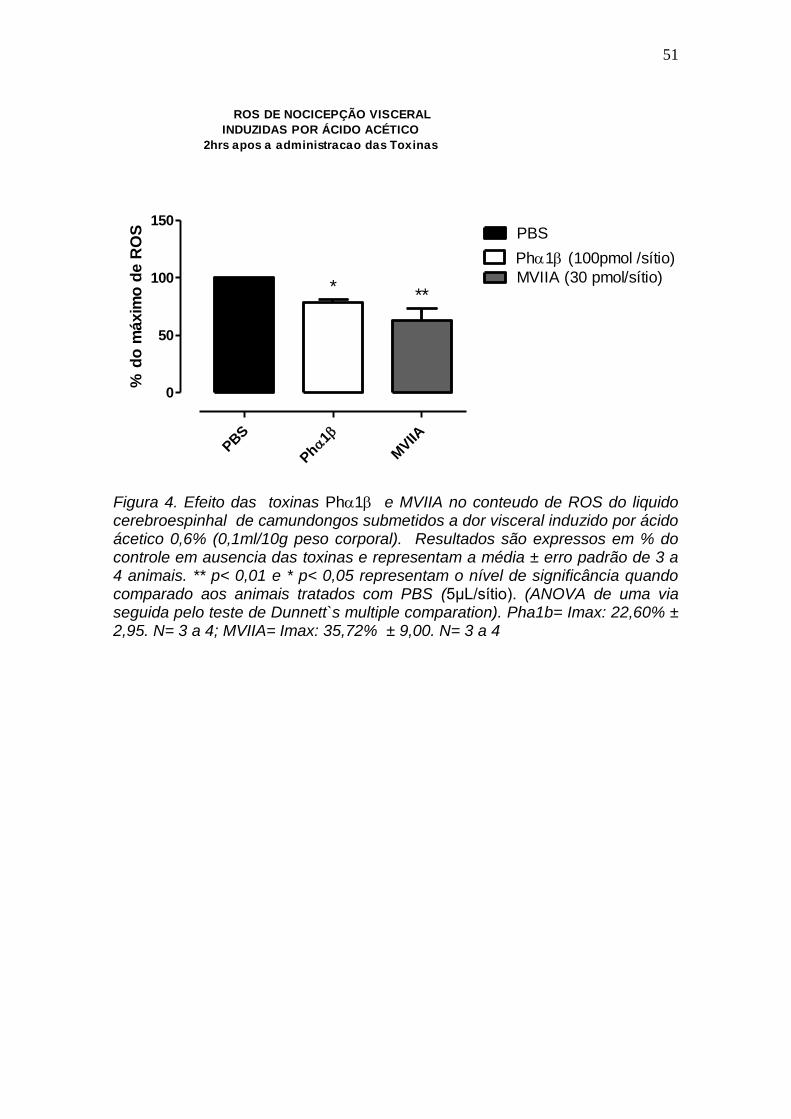
4.4 Efeito das toxinas Ph1 e MVIIA no conteudo de ROS do liquido cerebroespinhal de camundongo submetido a dor visceral induzido por ácido ácetico.
A figura 4 mostra o efeito das toxinas Ph1 (100 pmol) e -conotoxina
MVIIA (30 pmol) nos niveis de ROS do liquido cerebroespinhal de
camundongos submetidos a dor visceral induzida por ácido acético,
evidenciando que ambas as toxinas , Ph1 e -conotoxina MVIIA reduzem
em 22,6% 2.9 e 35,7% 9,0 a concentração de ROS, P < 0.05.
51
PBS 1
Ph M
VIIA
0
50
100
150
* **
PBS
Ph1 (100pmol /sítio)
MVIIA (30 pmol/sítio)
ROS DE NOCICEPÇÃO VISCERAL
INDUZIDAS POR ÁCIDO ACÉTICO
2hrs apos a administracao das Toxinas%
do
máxim
o d
e R
OS
Figura 4. Efeito das toxinas Ph1 e MVIIA no conteudo de ROS do liquido cerebroespinhal de camundongos submetidos a dor visceral induzido por ácido ácetico 0,6% (0,1ml/10g peso corporal). Resultados são expressos em % do controle em ausencia das toxinas e representam a média ± erro padrão de 3 a 4 animais. ** p< 0,01 e * p< 0,05 representam o nível de significância quando comparado aos animais tratados com PBS (5μL/sítio). (ANOVA de uma via seguida pelo teste de Dunnett`s multiple comparation). Pha1b= Imax: 22,60% ± 2,95. N= 3 a 4; MVIIA= Imax: 35,72% ± 9,00. N= 3 a 4

52
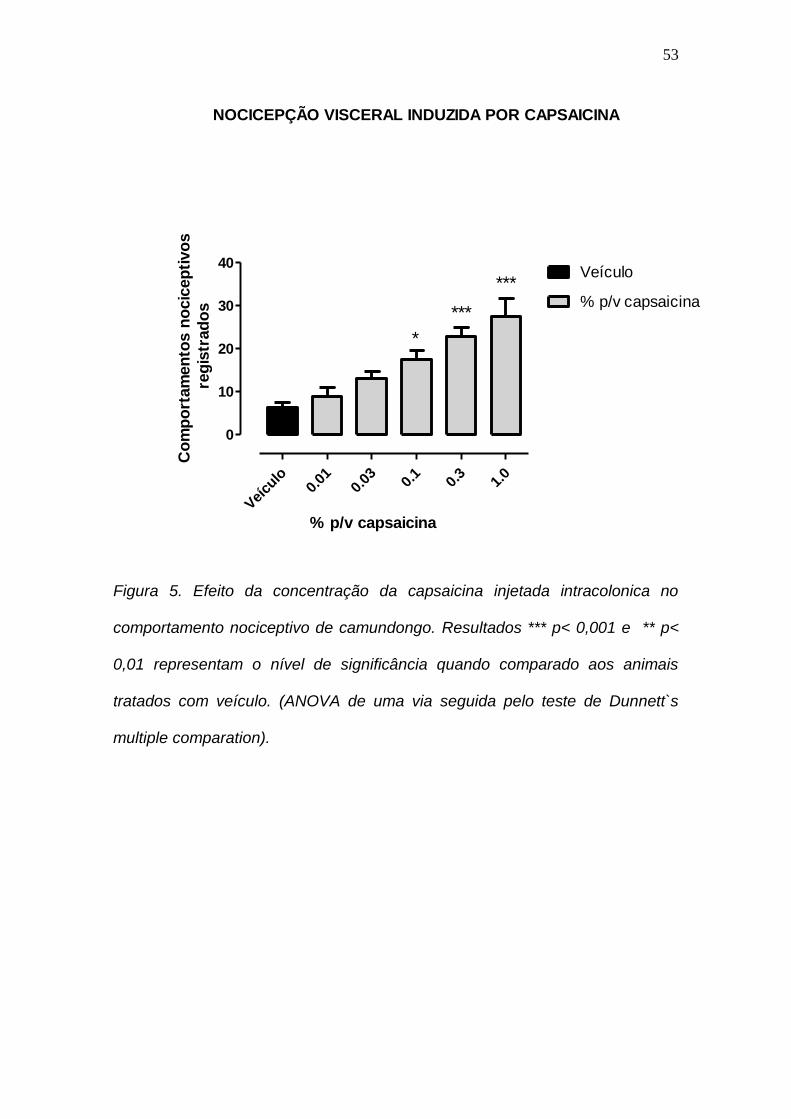
4.5 Efeito da concentração da capsaicina injetada intracolonica no comportamento nociceptivo de camundongo.
Recentemente, Laird et al. 2001 desenvolveram novo metodo de dor
visceral usando administração de capsaicina no colon de ratos. Resolvemos
testar esse metodo em nossos experimentos. A figura 5 refere-se à
administração intracolonica (intcol) de capsaicina nas concentrações de 0,01,
0.03, 0,1 e 1,0%. Nestas concentrações a capsaicina aumentou a hiperalgesia
dos camundongos em 33, 93, 166, 246 e 350%, respectivamente. Observamos
assim uma relação direta entre o aumento da concentração de capsaicina e do
comportamento nociceptivo dos animais. Nesse outro modelo de dor visceral
resolvemos testar o efeito antinociceptivo da Ph1 e -conotoxina MVIIA.
53
NOCICEPÇÃO VISCERAL INDUZIDA POR CAPSAICINA
Veí
culo
0.01
0.03 0.
10.
31.
0
0
10
20
30
40Veículo
% p/v capsaicina
*
***
***
% p/v capsaicina
Co
mp
ort
am
en
tos n
ocic
ep
tivo
s
reg
istr
ad
os
Figura 5. Efeito da concentração da capsaicina injetada intracolonica no
comportamento nociceptivo de camundongo. Resultados *** p< 0,001 e ** p<
0,01 representam o nível de significância quando comparado aos animais
tratados com veículo. (ANOVA de uma via seguida pelo teste de Dunnett`s
multiple comparation).

54
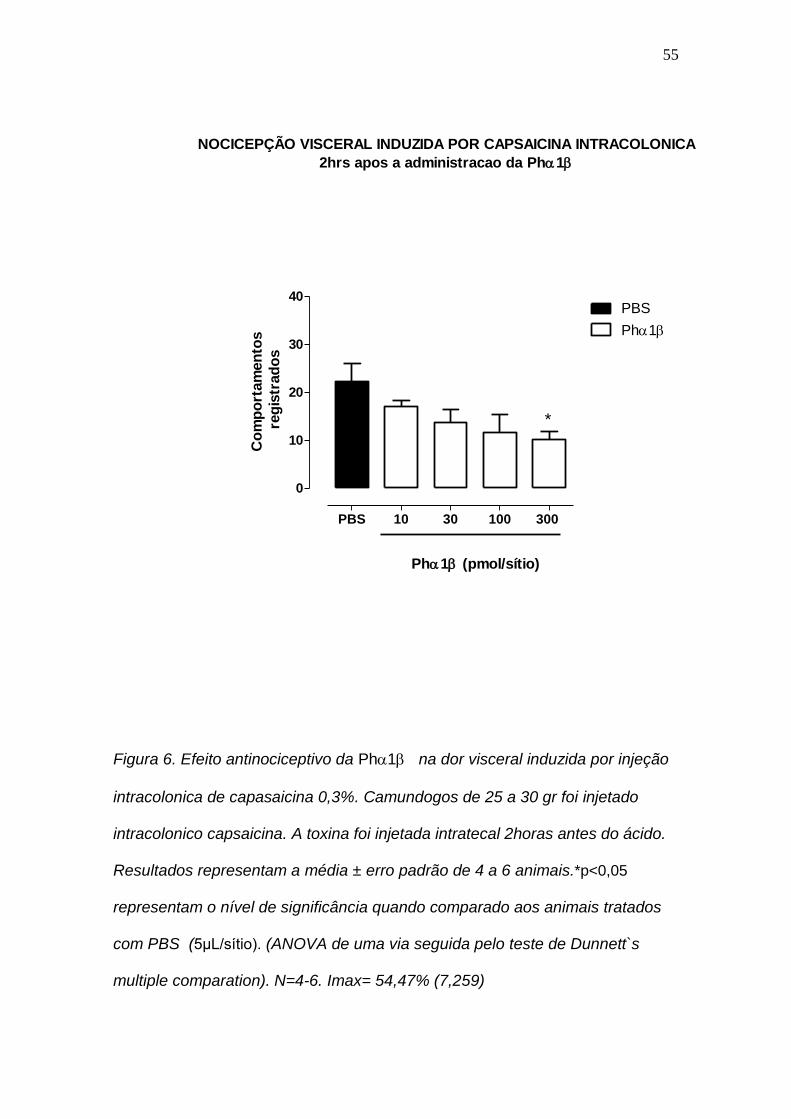
4.6 Efeito antinociceptivo da Ph1 na dor visceral induzida por injeção intracolonica de capasaicina.
A figura 6 mostra curva de concentração do efeito da toxina Ph1 no
modelo de dor visceral induzido pela capsaicina intracolônica. Observou-se que
nas concentrações de 10, 30, 100 e 300 nmol/site a Ph1 causou inibição do
efeito nociceptivo em 25, 39, 49 e 54% respectivamente. Somente na
concentração de 300 nmol a toxinou inibiu significativamente a nocicepção
induzida pela aplicação intracolônica da capsaicina, P < 0.05. A Imax foi igual
a54,4 7,2.
55
NOCICEPÇÃO VISCERAL INDUZIDA POR CAPSAICINA INTRACOLONICA
2hrs apos a administracao da Ph1
PBS 10 30 100 300
0
10
20
30
40
Ph1 (pmol/sítio)
*
PBS
Ph1
Co
mp
ort
am
en
tos
reg
istr
ad
os
Figura 6. Efeito antinociceptivo da Ph1 na dor visceral induzida por injeção
intracolonica de capasaicina 0,3%. Camundogos de 25 a 30 gr foi injetado
intracolonico capsaicina. A toxina foi injetada intratecal 2horas antes do ácido.
Resultados representam a média ± erro padrão de 4 a 6 animais.*p<0,05
representam o nível de significância quando comparado aos animais tratados
com PBS (5μL/sítio). (ANOVA de uma via seguida pelo teste de Dunnett`s
multiple comparation). N=4-6. Imax= 54,47% (7,259)

56
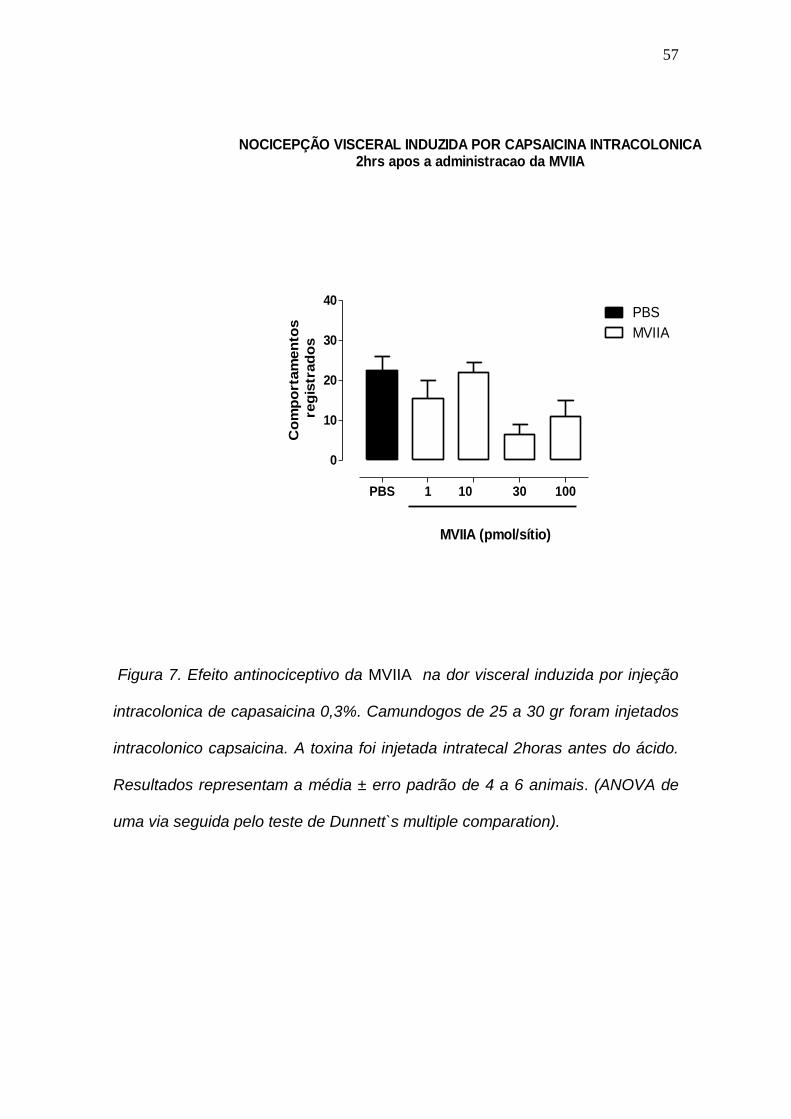
4.7: Efeito antinociceptivo da MVIIA na dor visceral induzida por injeção intracolonica de capasaicina.
A figura 7 mostra o efeito da concentração da -conotoxina MVIIA na
nocicepção induzida pela aplicação intracolôncia da capsaicina. Apesar de em
algumas concentrações a toxina causou maior efeito inibitório os resultados
não foram significativos P > 0,05 impossibilitanto o calculo da IC50. Os dados
do experimento impossibilitaram o calculo da IC50
57
NOCICEPÇÃO VISCERAL INDUZIDA POR CAPSAICINA INTRACOLONICA2hrs apos a administracao da MVIIA
PBS 1 10 30 100
0
10
20
30
40
MVIIA (pmol/sítio)
PBS
MVIIA
Co
mp
ort
am
en
tos
reg
istr
ad
os
Figura 7. Efeito antinociceptivo da MVIIA na dor visceral induzida por injeção
intracolonica de capasaicina 0,3%. Camundogos de 25 a 30 gr foram injetados
intracolonico capsaicina. A toxina foi injetada intratecal 2horas antes do ácido.
Resultados representam a média ± erro padrão de 4 a 6 animais. (ANOVA de
uma via seguida pelo teste de Dunnett`s multiple comparation).

58
5 DISCUSSÃO
Em animais várias substâncias algogenicas como acido acético,
capsaicina, mostarda, ciclofosfamida aplicadas em estruturas viscerais
provocam provocar dor. Estas substâncias algogenicas são capazes de induzir
a dor, bem como a reação inflamatória em neuronios aferentes viscerais ou
neurónios centrais. Na busca de novos substâncias naturais, que possuam
propriedade antinociceptiva visceral, foram utilizados os modelos de dor
visceral induzida por administração intraperitoneal de ácido acético ou injeção
intracolónica de capsaicina. A atividade antinociceptiva das toxinas Phα1β e ω-
conotoxina MVIIA foi avaliada nos dois modelos acima. O modelo químico de
nocicepção de contorções abdominais em camundongos baseia-se na
contagem das contorções da parede abdominal seguidas de torção do tronco e
extensão dos membros posteriores, como resposta reflexa à peritonite
produzidas pela injeção intraperitoneal de ácido acético (Whittle, 1964).
O processo inflamatório é deflagrado por um agente agressor ou
flogístico que provoca lesão tecidual. Esta lesão induz liberação de histamina
dos grânulos localizados nos mastócitos (células teciduais), a qual é
responsável pela gênese do processo inflamatório, que culmina com a
liberação de prostaglandina, o agente causador da dor inflamatória (Goodman
et al., 1996). Assim sendo, o teste de contorção abdominal em camundongos é
um método muito utilizado para se avaliar a atividade analgésica de
substâncias contra dor de origem inflamatória visceral, onde o ácido acético na
concentração de 0,6% (v/v), induz lesão no abdômen do camundongo, o que é
suficiente para provocar os espasmos traduzidos como contorção (Koster &
Anderson, 1959).O teste das contorções abdominais induzidas pelo ácido

59
acético é descrito como um modelo para avaliar a dor visceral de origem
inflamatória, com pouca especificidade, mas boa sensibilidade, podendo ser
considerado uma ferramenta para avaliação da atividade analgésica e
antiinflamatória de novos produtos (Le Bars et al., 2001).
Recentemente, Feng et al. (2003) demonstraram que a injeção intraperitoneal
de ácido acético induz aumento da concentração de glutamato e aspartato no
liquido cerebroespinhal. Nossos resultados confirmam o aumento de glutamato
na dor visceral induzida por acido acético ( FIG. 4) e a diminuição da liberação
deste neurotransmissor quando as toxinas Phα1β e ω-conotoxina MVIIA foram
aplicadas intratecal antes da indução das contorções abdominais. O
mecanismo de ação das toxinas envolve inibição dos canais de calcio. Nesse
particular a Phα1β tem ação nos canais tipo L, P/Q e maior atividade no
canal de calcio N (Vieira et al., 2005) enquanto a ω-conotoxina MVIIA inibe
especificamente o canal de calcio do tipo N. A inibição das toxinas na dor
visceral induzidas por injeção intraperitonial de ac. acético e intracolônica por
capsaicina foi dose dependente e tiveram praticamente o mesmo efeito
inibitório. Além de inibir a nocicepção as toxinas foram tambem eficazes em
inibir o conteudo de ROS do liquido cerebro espinhal de animais submetidos a
dor visceral induzida por ac. acético. Nesse metodo o ác. acético induz
processo inflamatório e com o aumento do processo inflamatorio e da dor
ocorre o aumento de ROS. Radicais livres, ROS estão implicados no
desenvolvimento de dor persistente resultante de injuria ou de insulto
inflamatório (Gao et al., 2007). Agentes que reduzem ROS tem ação em reduzir
a hiperalgesia (Yowtak et al., 2011). Nossos resultados confirmam que as

60
toxinas com sua ação antinociceptiva na dor visceral induzem diminuição de
ROS do liquido cerebroespinhal.
Com o objetivo de melhor caracterizar a atividade antinociceptiva
envolvida na dor visceral, foi utilizado o teste da capsaicina administrada
intracolônica em camundongos. Esse modelo químico de nocicepção fornece
uma resposta mais específica da dor visceral do que aquele induzida pela
injeção intraperitonial de ácido acético (Shibata, 1989). Além disso o modelo de
dor visceral induzido pela administração intracolônica da capsaicina é
considerado atualmente o modelo que mais se aproxima da dor clínica (Tjolsen
& Hole, 1997). Nossos resultados demonstram a atividade antinociceptiva da
toxina Phα1β inibindo a dor visceral, induzida pela administração intracolônica
da capsaicina. Os resultado com ω-conotoxina MVIIA mostram que a inibição
induzida pela mesma não foi estatisticamente significante, talvez pela grande
variação no grupo, tornando-se necessário aumentar o número de amostras do
efeito da ω-conotoxina MVIIA. Finalmente os dados assim obtidos confirmam o
potencial terapeutico antinociceptivo da Phα1β.

61
6 CONCLUSÃO
Diante dos nossos resultados é possível concluir que:
A Phα1β produz efeito antinociceptivo em modelos de dor visceral
induzido por ácido acético 0,6% intraperitonial e capsaicina 0,3% intracolônical
nas doses testadas (30, 100 e 300 pmol/sítio).;
A ω-CTx-MVIIA produz efeito antinociceptivo em modelos de dor visceral
induzido por ácido acético 0,6% intraperitonial e capsaicina 0,3% intracolônical
nas doses testadas (30 e 100 pmol/sítio) porém ja nas doses de 30 pmol/site ja
apresentava toxicidade nos animais;
O efeito antinociceptivo da Phα1β esta relacionado com a depleção nos
níveis de glutamato liberado, isso ocorre através da redução do influxo de
cálcio devido ao bloqueio dos CCSV do tipo N presentes no corno dorsal da
medula, pela Ph1 .
Alem de apresentar efeito antinoceceptivo no teste de contorção abdominal
induzida por acido acético a Ph1 e a A ω-CTx-MVIIA. Também reduzem a
liberação de ROS no liquido cerebroespinhal.

62
7 REFERÊNCIAS BIBLIOGRÁFICAS
ALTIER C, ZAMPONI GW (2004). Targeting Ca2+ channels to treat pain: T-type versus N-type. Trends Pharmacol Sci. 25:465-469. ARAÚJO DAM, CORDEIRO MN, DINIZ CR, BEIRÃO OS (1993). Effects of a toxic fraction, PhTx2, from the spider Phoneutria nigriventer on the sodium current. Naunyn Schmiedebergs Arch Pharmacol. 347:205-208. BATTTAGLIA G, RUSTIONI (1988). The coexistence of glutamate and substance P in dorsal root ganglion cells of the rat and monkey. J Comp Neurol. 277:302-312. BRUNO, A. A Abordagem clínica na dor crônica. Rev. Bras. Med.; 58: 1-10, 2001. BERKLEY, K. J. Sex differences in pain. Behav Brain Sci 20: 371-380
BESSON JM (1999) The neurobiology of pain. Lancet 353: 1610-1615, 1999.
BJÖRKMAN R (1995). Central antinociceptive effect of non-steroidal anti-inflammatory drugs and paracetamol. Acta Anesthesiol Scand. 39 ( supplement 103):1-44. BOWERSOX SS, GADBOIS T, SINGH T, PETTUS M, WANG YX, LUTHER RR (1996) Selective N-type neuronal voltage-sensitive calcium channel blocker, SNX-111, produces spinal antinociception in rat models of acute, persistent and neuropathic pain. J Pharmacol Exp Ther 279:1243-1249.
BRENNAN F, CARR DB, COUSINS M (2007). Pain management: a fundamental human right. Anesth Analg. 105:205-221.

63
CALIXTO JB, CABRINE DA, FERREIRA J, CAMPOS MM (2000). Kinins in pain andinflammation. Pain 87:1-5. CALIXTO JB, CABRINI DA, FERREIRA J, CAMPOS MM (2001). Inflammatory pain: kinins and antagonists. Curr Opin Anesthesiol. 14:519-526.
CARLTON SM, ZHOU S, COGESHALL RE (1996). Localization and activation of substance P receptors in unmyelinated azons of rat glabrous skin. Brain Res 734: 103-108.
CASSOLA AC, JAFFE H, FALES HM, CASTRO AFECHE S, MAGNOLI F AND CIPOLLA-NETO J (1998) omega-phonetoxin-IIA: a calcium channel blocker from the spider Phoneutria nigriventer. Pflugers Arch 436:545-52.
CHAPMAN CR, GAVRIN J (1999) Suffering: the contributions of persistent pain.
Lancet 353: 2233-2237.
COSTA SK, MORENO RA, ESQUISATTO LC, JULIANO L, BRAIN SD, DE NUCCI, ANTUNES E (2002). Role of kinins and sensory neurons in the rat pleural leokocyte migration induced by Phoneutria nigriventer spider venom. Neurosci. Lett. 318:158-162.
COTRAN C, KUMAR T, ROBBINS A (1994) Patologia estrutural e funcional. Rio de Janeiro: Guanabara-Koogan, p. 35-71.
DICKENSON A H (1995). Central acute pain mechanisms. Ann. Med. 27:223-227.
DINIZ, CR, CORDEIRO, MN, JUNIOR, LR, KELLY, P, FISCHER, S, REIMANN, F, OLIVEIRA, E B AND RICHARDSON, M (1990) The purification and amino acid sequence of the lethal neurotoxic Tx1 from the venom of the Brazilian “armed”spider Phoneutria nigriventer. FEBS Lett 263: 251-253.

64
COLLIER, H. D. J.; DINNIN, L. C.; JOHNSON, C. A.; SCHNEIDER, C(1968). The abdominal response and its suppression by analgesic drug in the mouse. Br. J. Pharmacol. Chemother.; 32: 295-310. DOS SANTOS RG, VAN RENTERGHEM C, MARTIN-MOUTOT N, MANSUELLE P, CORDEIRO MN, DINIZ CR, MORI Y, DE LIMA ME AND SEAGAR M (2002) Phoneutria nigriventer omega-phonetoxin II A blocks the Cav2 family of calcium channels and interacts with omega-conotoxin-binding sites. J Biol Chem 277:13856-13862. EDVINSSON L, CANTERA L, JANSEN-OLENSEN I. ET AL (1997). Expresion of calcitonin gene-related peptide, receptor mRNA in humam trigeminal ganglia and cerebral arteries. Neurosci Lett 229: 209-211.
EICKSTEDT VRD (1983). Considerações sobre a sistemática das espécies amazônicas de Phoneutria (Araneae, Ctenidae). Brasil Zoologia 1:183-191.
FAUCETT JA, LEVINE JD (1991) The contributions of interpersonal conflict to chronic pain in the presence or absence of organic pathology. Pain 44: 35-43.
FENG, Z. P.; DOERING, C. J.; WINKFEIN, R. J.; BEEDLE, A. M.; SPAFFORD, J. D.; ZAMPONI, G. W (2003). Determinants of inhibition of transiently expressed voltage-gated calcium channels by omega-conotoxins GVIA and MVIIA. J. Biol. Chem.; 278: 20171-20178. FIELDS HL, BASBAUM AL, HEINRICHER MM (2006). Central nervous system mechanisms of pai modulation. In: McMahon S, Koltzenburg M. (eds). Walls, Melzack . Textbook of pain. 5a ed. Edinburg: Churchill Livingstone, cap 125-142. FILLINGEN RB, NESS TJ (2000) Sex-related hormonal influences on pain and
analgesic responses. Neurosci Beh R 26: 485-501.
FÜRST, S (1998). Transmitters involved in antinociception in the spinal cord. Brain Res. Bull.; 48: 129-141.

65
GANONG W (1988) The stress response: a dynamic overview. Hospital Practice, 23(6): 155-190. GAO X, KIM HK, CHUNG JM, CHUNG K. Reactive oxygen species (ROS) are involved in enhancement of NMDA-receptor phosphorylation in animal models of pain.Pain 2007;131:262–71, GOMEZ MV, KALAPOTHAKIS E, GUATIMOSIM C, PRADO MA (2002). Phoneutria nigriventer venom: a cocktail of toxins that affect ion channels. Cell. Mol. Neurobiol. 22:579-588. GRISHIN E (1999). Polypeptide neurotoxins from spider venoms. Eur. J. Biochem. 264: 276-280. GUATIMOSIM C, ROMANO-SILVA MA, CRUZ JS, BEIRÃO PSL, KALAPOTHAKIS E,MORAES-SANTOS T, CORDEIRO MN, DINIZ CR, GÓMEZ MV, PRADO MAM (1997). Atoxin from the spider Phoneutria nigriventer that blocks calcium channels coupled to exocytosis. Br. J. Pharmacol. 122:591-597. GOODMAN, L. S.; GILMAN, A. G.; RALL, T. W.; SCHLEIFER, L. S (1996). The Pharmacological Basis of Therapeutics, 442-443. HERLITZE S, XIE M, HAN J, HÜMER A, MELNIK-MARTINEZ KV, MONERO R, MAERK MD (2003). Targeting mechanisms of high voltage-activated Ca2+ channels. J.Bionerg. Biomembr. 35: 621-637. HILL, R. G (2001). Molecular basis for the perception of pain. Trends Neurosci.; 7: 282 -292. HOGG, R. C (2006). Novel approaches to pain relief using venom-derived peptides. Curr. Med. Chem.; 13: 3191-3201. HYLDEN, J. L.; WILCOX, G. L(1980). Intrathecal morphine in mice: a new technique. Eur. J. Pharmacol.; 67: 313-316. IKEDA, Y.; UENO, A.; NARABA, H.; OH-ISHI, S (2001). Involvement of vanilloid receptor VR1 and prostanoids in the acid-induced writhing responses of mice. Life Science. 69: 2911-2919.

66
INOEU K, NAKASAWA K, INUOEU K. et al (1995) Nonseletctive cátion channels coupled with tackykinin receptors in rat sensory neurons. J Neurophysiol (73): 736-742.
JACKSON DL, GRAFF CB, RICHARDSON, JD. et al (1995) Glutamate participates in the peripheral modulation of thermal hyperalgesia in rats. Eur J Pharmacol 284: 321-325.
JIANG MC, GEBHART GF. Development of mustard oil-induced hyperalgesia in rats. Pain 1998;77(3):305-13. JIN MO CHUNG (2011). Reactive oxygen species contribute to neuropathic pain by reducing spinal GABA release. Pain 152 844–852 JONES, S. L (1992). Anatomy of pain. in: Sinatra RS, Hord, AH, Ginsberg B, Preble l. Acute Pain: Mechanisms & Management. Mosby–Year Book: St. Louis. JULIUS, D.; BASBAUM, A. I (2001). Molecular mechanisms of nociception. Nature; 413: 203-210. KALAPOTHAKIS E, PENAFORTE CL, BEIRÃO PSL, ROMANO-SILVA MA, CRUZ JS, PRADOMAM, GUIMARÃES PEM, GOMEZ MV, PRADO VF (1998). Cloning of cDNA senconding neurotoxic peptides from the spider Phoneutria nigriventer. Toxicon. 36:1843-1850. KANDEL, E. R.; SCHWARTZ, J. H.; JESSEL, T. M (2003). Principios da neurosciencia. Manole: São Paulo. KOSTER, R.; ANDERSON, M.; BEER, E. J (1959). Acetic acid for analgesic screening. Fed. Proc.; 18: 412-,. KURAISHI Y, HIROTA N, SOTA Y, ET AL (1985) Noradrenergic inhibition of the release of substance P from the primary afferents in the rabit spinal dorsal horn. Br Res 359: 177-182.
KUSHMERICK C, KALAPOTHAKIS E, BEIRÃO PSL, PENAFORTE CL, PRADO VF, CRUZ JS, DINIZ CR, CORDEIRO MN, GOMEZ MV, ROMANO-SILVA MA, PRADO MAM (1999).Phoneutria nigriventer toxin tx3-1 blocks A-type K+ currents controlling Ca2+oscillation frequency in GH3 cells. J

67
Neurochem. 72:1472-1481. LE BARS, D.; GOZARIU, M.; CADDEN, S. W (2001). Animal models of nociception. Pharmacol. Rev.; 53: 597-652. LAIRD JM, MARTINEZ-CARO L, GARCIA-NICAS E, CERVERO F. (2001). A new model of visceral pain and referred hyperalgesia in the mouse.. Pain. Jun;92(3):335-42
LEVINE, J. D.; TAIWO, Y (1994). Inflammatory Pain, in: Wall PD, Melzack R, Bonica JJ - Textbook of Pain. 3rd ed., Churchill Livingstone: Edinburgh. LIU H, WANG H, SHENG M, JAN LY, JAN YN, BASBAUM AI (1994). Evidence for presynaptic N-methyl-D-aspartate autoreceptors in the spinal cord dorsal horn. Proc. Nat. Acad. Sci. USA. 91:8383-8387. LOESER JD, MELZACK R (1999). Pain: An Overview. Lancet 353:1607-1609. LUCAS S (1988). Spiders in Brazil. Toxicon 26:759-772. MALMBERG A, YAKSH TL (1994). Voltage-sensitive calcium channels in spinal nociceptive processing: Blockade of N- and P-type channels inhibits formalininduced nociception. J. Neurosci. 14:4882-4890.
-conopeptides, N-type calcium-channel blockers, on behavior and antinociception in the formalin and hot-plate tests in rats. Pain; 60: 83-90, 1995. MATTHEWS EA, DICKENSON AH (2001). Effects of ethosuximide, a T-type Ca(2+) channel blocker, on dorsal horn neuronal responses in rats. Eur. J. Pharmacol. 415:141-149. MERSKEY, H.; BOGDUK, N (1994). Classification of chronic pain: description of chronic pain syndromes and definitions of pain terms. IASP: Press Seattle. MENDELL JR, SAHENK Z (2003). Painful sensory neuropathy. N Engl J Med. 348:1243-1255. MILLAN, M. J (1999). The induction of pain: an integrative review. Prog Neurobiol 57, 1-164,.

68
MILLAN, M. J (1997). The role of descending noradrenergic and serotoninergic pathways in the modulation of nociception: focus on receptor multiplicity. In: Dickenson A, Besson J-M. The Pharmacology of Pain. Handbook of Experirmental Pharmacology. Springer Verlag: Berlin,. MILJANICH GP, RAMACHANDRAN J (1995). Antagonists of neuronal calcium channels: structure, function, and therapeutic implications. Annu Rev Pharmacol Toxicol. 35:707-34. MIRANDA DM, ROMANO-SILVA MA, KALAPOTHAKIS E, DINIS CR, CORDEIRO MN, MORAES-SANTOS T, PRADO MAM, GOMEZ MV (1998). Phoneutria nigriventer toxinsblock tityustoxin-induced calcium influx in synaptosomes. Neuroreport. 9:1371-1373. NICHOLLS, D. G.; SIHRA, T. S.; SANCHEZ-PRIETO (1987), J. Calcium dependent and independent release of glutamate from synatosomes monitored by continuous fluorometry. J. Neurochem.; 49: 50-57. OLIVAR T, LAIRD JM (1999). Cyclophosphamide cystitis in mice: behavioural characterisation and correlation with bladder inflammation.Eur J Pain;3(2):1419.
PINHEIRO AC, GOMEZ RS, MASSENSINI AR, CORDEIRO MN, RICHARDSON M, ROMANO-SILVA MA, PRADO MA, DE MARCO LA, GÓMEZ, MV (2006). Neuroprotective effecton injury by neurotoxins from the spider Phoneutria nigriventer. Neurochem Int.49:543-547. PENN RD, PAICE JA (2000). Adverse effects associated with the intrathecal administration of ziconotide. Pain 85:291-296. PORRECA F, OSSIPOV MH, GEBHART GF (2002). Chronic pain and medullarydescending facilitation. Trends Neurosci. 25:319-325. PRADO, W. A (2001). Involvement of calcium in pain and antinociception. Braz. J. Med. Biol. Res.; 34:449–461,. REIS, HJ, PRADO, MAM, KALAPTOTHAKIS, E, COREIRO, MN, DINIZ, CR, DE MARCO, LA, GOMEZ, MV AND ROMANO-SILVA, MA (1999) Inhibition of glutamate uptake by a polypeptide toxin (Phoneutriatoxin 3-4) from the spider Phoneutria nigriventer. Biochem J 343:413-418.

69
REZENDE, LJ CORDEIRO MN, OLIVEIRA EB AND DINIZ CR (1991) Isolation of neurotoxic peptides from the venom of the „armed‟ spider Phoneutria nigriventer. Toxicon 29:1225-1233. RIBEIRO, R. A.; VALE, M. L.; FERREIRA, S. H.; CUNHA, F. Q (2000).; Analgesiceffect of thalidomide on inflammatory pain. Eur J Pharmacol.; 391(1/2):97-103,. RIEDEL, W. NEECK, G (2001). Nociception, pain and antinociception: current concepts. Z. Reumathol.; 60: 404 – 415. ROMANO-SILVA MA, RIBEIRO SANTOS R, RIBEIRO AM, GOMEZ MV, DINIZ CR, CORDEIRO MN E BRAMMER MJ (1993). Rat cortical synaptosomes have more than one mechanism for Ca2+ entry linked to rapid glutamate release: studies using 77 the Phoneutria nigriventer toxin PhTx2 and potassium depolarization. Biochem J. 296:313-319. RUSSO, C. M.; BROSE, W. G (1998). Chronic pain. Annu. Rev. Med.; 49: 123-133,.
SANTOS, A. R. S.; MIGUEL, O. G.; YUNES, R. A.; CALIXTO, J. B(1999). Antinociceptive properties of the new alkaloid, cis-8, 10-di-N-propyllobelidiol hydrochloride dehydrate isolated from Siphocampylus verticillatus: evidence for the mechanism of action. J. Pharmacol. Experim. Therapeutics; 289: 417-426, SHARP TJ (2001) Chronic pain: a reformulation of the cognitive-behavioral model.Beh Res Ther 39: 787-800. SOUZA, A. H.; FERREIRA, J.; CORDEIRO, M. N.; VIEIRA, L. B.; DE CASTRO, C. J.; TREVISAN, G.; REIS, H.; SOUZA, I. A.; RICHARDISON, M.; PRADO, M. A. M.; PRADO, V. F.; GOMEZ, M. V. Analgesic effect in rodents of native and recombinant Phα1β toxin, a high-voltage-activated calcium channel blocker isolated from armed spider venom. Pain; 140: 115-126, 2008. SNUTCH TP, SUTTON KG, ZAMPONI GW (2001). Voltage-dependent calcium channels-beyond the dihydropyridine antagonists. Curr Opin Pharmacol. 1:11-16.

70
SORKIN LS, MCDOO DJ, WILLIS WD (1993). Raphe magnus stimulation-induced antinociception in the cat is associated with release of aminonacids as well as serotonin in the lumbar dorsal horn. Brain Res. 618:95-108 TRACEY J, MANTYH PW (2007). The cerebral signature for pain perception and itsmodulation. Neuron 55: 377-91. TURK, DC, OKIFUJI A (1999) Assessment of patients‟ reporting of pain: an
integrated perspective. Lancet 353: 1784-1788.
VANEGAS H, SCHEIBLE (2004). Descending control of persistent pain: inhibitory or facilitatory? Brain Res Rev. 46:295-309. VIEIRA LB, KUSHMERICK C, HILDEBRAND ME, GARCIA E, STEA A, CORDEIRO MN, RICHARDSON M, GOMEZ MV, SNUTCH TP (2005). Inhibition of high voltage-activated calcium channels by spider toxin PnTx3-6. J Pharmacol Exp Ther 314(3):1370-1377.
. WEN, L.; YANG, S.; ZHOU WX.; ZHANG YX.; ZHOU XW.; HUANG PT( 2005).; Involvement of N-type calcium channels in pain and antinociception. Sheng LiKe Xue Jin Zhan. 36: 23-28, WHEELER DB, SATHER WA, RANDALL A, TSIEN RW (1994). Distintive properties of a neuronal calcium channel and its contribution to excitatory synaptic transmission in the central nerve system. Adv. Second Messenger phosphoprotein Res. ISSN: 1040-7952, 29:155-171. WOOLF CJ, MANNION RJ (1999). Neuropathic pain: etiology symptoms, mechanisms and management. Pain 353:1959-1964., ZAMPONI, G. W.; SNUTCH, T. P (1998). Modulation of voltage-dependent calcium channels by G proteins. Curr. Opin. Neurobiol.; 8: 4089-4097,. ZIMMERMANN, M (1983). Ethical guidelines for investigations of experimental pain in conscious animals. Pain; 16: 109-110,.

71


















