
Biological evaluation and in silico docking study of γ-linolenic acid as a potential BACE1...
5
Short communications Biological evaluation and in silico docking study of γ-linolenic acid as a potential BACE1 inhibitor Kumju Youn a , Jinhyuk Lee b,c , Eun-Young Yun d , Chi-Tang Ho e , Mukund V. Karwe e , Woo-Sik Jeong f , Mira Jun a, * a Department of Food Science and Nutrition, Dong-A University, Busan 604-714, Republic of Korea b Korean Bioinformation Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon 305-806, Republic of Korea c Department of Bioinformatics, University of Sciences and Technology, Daejeon 305-350, Republic of Korea d Department of Agricultural Biology, National Academy of Agricultural Science RDA, Suwon 441-100, Republic of Korea e Department of Food Science, Rutgers University, New Brunswick, NJ 08901, USA f Department of Food & Life Science, College of Biomedical Science & Engineering, Inje University, Gimhae 621- 749, Republic of Korea ABSTRACT Sequential proteolytic cleavage of amyloid precursor protein (APP) by β-secretase (BACE1) is a crucial process in β-amyloid peptide (Aβ) generation, which further forms into neuro- toxic amyloid plaques that are considered to be a pivotal hallmark in the development and progress of Alzheimer’s disease (AD). Hence, the inhibition of BACE1 has emerged as a cred- ible target for the prevention and/or treatment of AD. In this study, γ-linolenic acid (GLA) was discovered as a novel BACE1 specific inhibitor. GLA non-competitively suppressed BACE1 activity with an IC50 value of 7.6 × 10 −5 M and Ki value of 3.5 × 10 −5 M. In addition, we dem- onstrated the calculated docking poses of GLA to human BACE1 and revealed the interac- tions of GLA with the allosteric site of the enzyme bound with the OH group of CYS359. Our findings provide a novel possibility of GLA to be efficacious for the prevention of AD and provide scaffolds to explore more potent natural BACE1 inhibitors. © 2014 Elsevier Ltd. All rights reserved. ARTICLE INFO Article history: Received 23 April 2014 Received in revised form 29 May 2014 Accepted 4 June 2014 Available online Keywords: Alzheimer’s disease β-secretase (BACE1) β-amyloid peptide (Aβ) γ-linolenic acid 1. Introduction Alzheimer’s disease (AD) is the most common form of age- related neurodegenerative disorders leading to dementia. Criti- cal pathological features of AD are characterized by generation of extracellular amyloid neuritic plaques and intracellular neu- rofibrillary tangles (NFT) composed of β-amyloid peptide (Aβ) and hyperphosphorylated tau proteins, respectively (Tong et al., 2005). Aβ, the central component of amyloid plaque, is gener- ated by the consecutive endoproteolytic cleavage of Aβ pre- cursor protein (APP) by two proteases, β- and γ-secretase. In the amyloidogenic pathway, β-site APP cleaving enzyme (BACE1) leaves APP at the N-terminal end of Aβ and then this cleav- age produces a secreted ectodomain of APP (sAPPβ) and the C99 fragment. γ-Secretase gives intramembranous cleavage on * Corresponding author. Tel.:+82 51 200 7323; fax: +82 51 200 7535. E-mail address: [email protected] (M. Jun). http://dx.doi.org/10.1016/j.jff.2014.06.005 1756-4646/© 2014 Elsevier Ltd. All rights reserved. journal of functional foods 10 (2014) 187–191 Available at www.sciencedirect.com ScienceDirect journal homepage: www.elsevier.com/locate/jff
Transcript of Biological evaluation and in silico docking study of γ-linolenic acid as a potential BACE1...

Short communications
Biological evaluation and in silico docking studyof γ-linolenic acid as a potential BACE1 inhibitor
Kumju Youn a, Jinhyuk Lee b,c, Eun-Young Yun d, Chi-Tang Ho e,Mukund V. Karwe e, Woo-Sik Jeong f, Mira Jun a,*a Department of Food Science and Nutrition, Dong-A University, Busan 604-714, Republic of Koreab Korean Bioinformation Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon 305-806,Republic of Koreac Department of Bioinformatics, University of Sciences and Technology, Daejeon 305-350, Republic of Koread Department of Agricultural Biology, National Academy of Agricultural Science RDA, Suwon 441-100, Republicof Koreae Department of Food Science, Rutgers University, New Brunswick, NJ 08901, USAf Department of Food & Life Science, College of Biomedical Science & Engineering, Inje University, Gimhae 621-749, Republic of Korea
A B S T R A C T
Sequential proteolytic cleavage of amyloid precursor protein (APP) by β-secretase (BACE1)
is a crucial process in β-amyloid peptide (Aβ) generation, which further forms into neuro-
toxic amyloid plaques that are considered to be a pivotal hallmark in the development and
progress of Alzheimer’s disease (AD). Hence, the inhibition of BACE1 has emerged as a cred-
ible target for the prevention and/or treatment of AD. In this study, γ-linolenic acid (GLA)
was discovered as a novel BACE1 specific inhibitor. GLA non-competitively suppressed BACE1
activity with an IC50 value of 7.6 × 10−5 M and Ki value of 3.5 × 10−5 M. In addition, we dem-
onstrated the calculated docking poses of GLA to human BACE1 and revealed the interac-
tions of GLA with the allosteric site of the enzyme bound with the OH group of CYS359.
Our findings provide a novel possibility of GLA to be efficacious for the prevention of AD
and provide scaffolds to explore more potent natural BACE1 inhibitors.
© 2014 Elsevier Ltd. All rights reserved.
A R T I C L E I N F O
Article history:
Received 23 April 2014
Received in revised form 29 May
2014
Accepted 4 June 2014
Available online
Keywords:
Alzheimer’s disease
β-secretase (BACE1)
β-amyloid peptide (Aβ)
γ-linolenic acid
1. Introduction
Alzheimer’s disease (AD) is the most common form of age-related neurodegenerative disorders leading to dementia. Criti-cal pathological features of AD are characterized by generationof extracellular amyloid neuritic plaques and intracellular neu-rofibrillary tangles (NFT) composed of β-amyloid peptide (Aβ)
and hyperphosphorylated tau proteins, respectively (Tong et al.,2005). Aβ, the central component of amyloid plaque, is gener-ated by the consecutive endoproteolytic cleavage of Aβ pre-cursor protein (APP) by two proteases, β- and γ-secretase. Inthe amyloidogenic pathway, β-site APP cleaving enzyme (BACE1)leaves APP at the N-terminal end of Aβ and then this cleav-age produces a secreted ectodomain of APP (sAPPβ) and theC99 fragment. γ-Secretase gives intramembranous cleavage on
* Corresponding author. Tel.:+82 51 200 7323; fax: +82 51 200 7535.E-mail address: [email protected] (M. Jun).
http://dx.doi.org/10.1016/j.jff.2014.06.0051756-4646/© 2014 Elsevier Ltd. All rights reserved.
j o u rna l o f f un c t i ona l f o od s 1 0 ( 2 0 1 4 ) 1 8 7 – 1 9 1
Available at www.sciencedirect.com
ScienceDirect
journal homepage: www.elsevier.com/ locate / j ff

the C99 fragment releasing Aβ of varying length which formaggregates that lead to be the predominant species in amyloidplaque (Hardy & Allsop, 1991).
Aβ is neurotoxic in several modes. It induces oxidative stresscausing generation of oxidized protein and DNA, lipidperoxidation, inflammatory responses, mitochondrial dysfunc-tion, and disruption of Ca2+ homeostasis (Butterfield, Castegna,Lauderback, & Drake, 2002). Aβ, furthermore, was proved to as-sociate with phosphorylation, cleavage and aggregation of tauprotein which is the other distinctive pathological character-istic in AD (De Felice et al., 2008).
In the amyloid cascade hypothesis, Aβ production is therate limiting step and modulation and/or clearance of brainAβ levels by inhibiting the key enzyme BACE1 is believed tobe one of the priority drug targets in AD prevention. It hasbeen demonstrated that the increased activity andoverexpression of BACE1 enhanced marked increases in sAPPβand Aβ peptide species levels, whereas the downregulationof BACE1 diminished Aβ production (Sinha et al., 1999). ThePhase I clinical trials showed that BACE1 inhibitors were ableto lower cerebrospinal fluid (CSF) Aβ levels (Jeppsson et al.,2012). On the other hand, both function and structure ofγ-secretase yet remain unknown and the enzyme, moreover,is involved in notch signaling pathway that controls cell fatedecision. γ-Secretase inhibitors are displayed to have neuro-toxic side effects in vitro and in vivo causing significant devel-opmental defects (Shimmyo, Kihara, Akaike, Niidome, &Sugimoto, 2008).
γ-Linolenic acid (GLA, 18:3) is the first derivative in the con-version of linoleic acid to arachidonic acid (AA) by the actionof enzyme Δ-6-desaturase. GLA is mostly found in plant seedoils of borage, evening primrose and black currant (Chang et al.,2010). Various studies suggested that GLA is unique among then-6 polyunsaturated fatty acid (PUFA) family members in itspotential to inhibit tumor growth, metastasis and inflamma-tory responses, which the end product AA apparently cannot(Janssen & Kiliaan, 2014; Montserrat-de la Paz, García-Giménez,Ángel-Martín, Marín-Aguilar, & Fernández-Arche, 2013). Recentfindings indicated that GLA exerted selective tumoricidal effectsin various cancers such as breast, pancreas, colon, and brain(Kapoor & Huang, 2006).
Moreover, GLA was proven to restore nerve blood flow, nor-malize nerve conduction velocity, and adjust the impaired nervefunction in diabetic animal models (Dines, Cameron, & Cotter,1995; Horrobin, 1997). It modulated interleukin-1β-inducedchanges in anxiety-like behavior, monoaminergeic neurotrans-mitters and brain inflammation in rats (Song, Manku, &Horrobin, 2008). In contrast to such various GLA studies on in-flammation, cancers, diabetes and neuropathy, no study hasexamined any potential impact or association of Alzheimer’sdisease with GLA.
In our continuous research for discovering natural BACE1inhibitors, encouraging series of previous works on GLAkept our attention. Furthermore, one of the fatty acids,oleic acid was proven to suppress in vitro human BACE1 ac-tivity in our previous study (Youn et al., 2014). Therefore,the possibility of GLA as a BACE1 inhibitor was evaluated andmoreover, in silico docking studies were performed to investi-gate the specific binding sites of the inhibitor in recombinantBACE1.
2. Materials and methods
2.1. General
Fluorescence and optical density were measured with a Bio-TEK ELISA microplate fluorescence reader FLx 800 and Bio-TEK ELx 808 (Winooski, VT, USA), respectively. A BACE1 assaykit was purchased from Pan Vera (Madison, WI, USA.).α-Secretase (TACE) and substrates were obtained from R&Dsystems (Minneapolis, MN, USA). Trypsin, chymotrypsin, elas-tase and their substrates were obtained from Sigma-Aldrich(St. Louis, MO, USA). GLA (>99% purity) was also purchased fromSigma-Aldrich. All other chemicals used were of the highestgrade commercially available.
2.2. Enzymatic assay of BACE1, TACE and otherserine proteases
For the solubility problem of high hydrophobicity of GLA, 5 nMof Tween 20 was added in each enzymatic assay that showedno effect on the enzyme activity. Fluorometric BACE1 and TACEassays were determined using the previously described method(Youn, Jeong, & Jun, 2013). Trypsin, chymotrypsin and elas-tase were assayed according to the protocol described in thereference (Youn et al., 2014).
2.3. Evaluation of the inhibition kinetics on BACE1
The mode of BACE1 inhibition was determined by the graphi-cal views of Dixon plot, Lineweaver–Burk plot, and its second-ary plot. The BACE1 inhibitory activity of GLA was measuredwith different concentrations of the substrates. The inhibi-tion constants (Ki) were determined by interpretation of theDixon plot, where the value of the x-axis implies −Ki. The datawere acquired as mean values of 1/V, the inverse of the in-crement of fluorescence intensity per min (min/mF.U) in theLineweaver–Burk plot (Tian, Bassit, Chau, & Li, 2010). Km andVmax values were obtained from intersection of abscissa andordinate, respectively. Graphs were plotted by using Sigma plotsoftware (Version 12.0).
2.4. In silico docking studies
For docking studies, the crystal structure (PDB ID: 2WJO)(Nicholls et al., 2010) used for target BACE1 and inhibitor wasprepared from the PubChem database (CID 5280933 for GLA).Marvin was used for drawing, displaying, and characterizingthe chemical structures [Marvin 5.11.4, 2012, ChemAxon (http://www.chemaxon.com)]. Pck software was used to search thebinding pocket residues of BACE1 and AutoDock Vina was usedfor protein-ligand docking simulations (Trott & Olson, 2010).The used parameters were as follows: xyz center coordinatesof the binding pocket residues; search space (15 Å) in each di-mension, and generation number (10) of each binding pocketresidue. All docking structures were clustered and catego-rized by lowest energy and largest number of clusters.
2.5. Statistical analysis
All experiments were performed in triplicate. Data of each ex-periment showed the mean ± SE. Significant differences were
188 j o u rna l o f f un c t i ona l f o od s 1 0 ( 2 0 1 4 ) 1 8 7 – 1 9 1
analyzed by Duncan’s multiple range tests using StatisticalAnalysis System (SAS) version 9.3.
3. Results and discussion
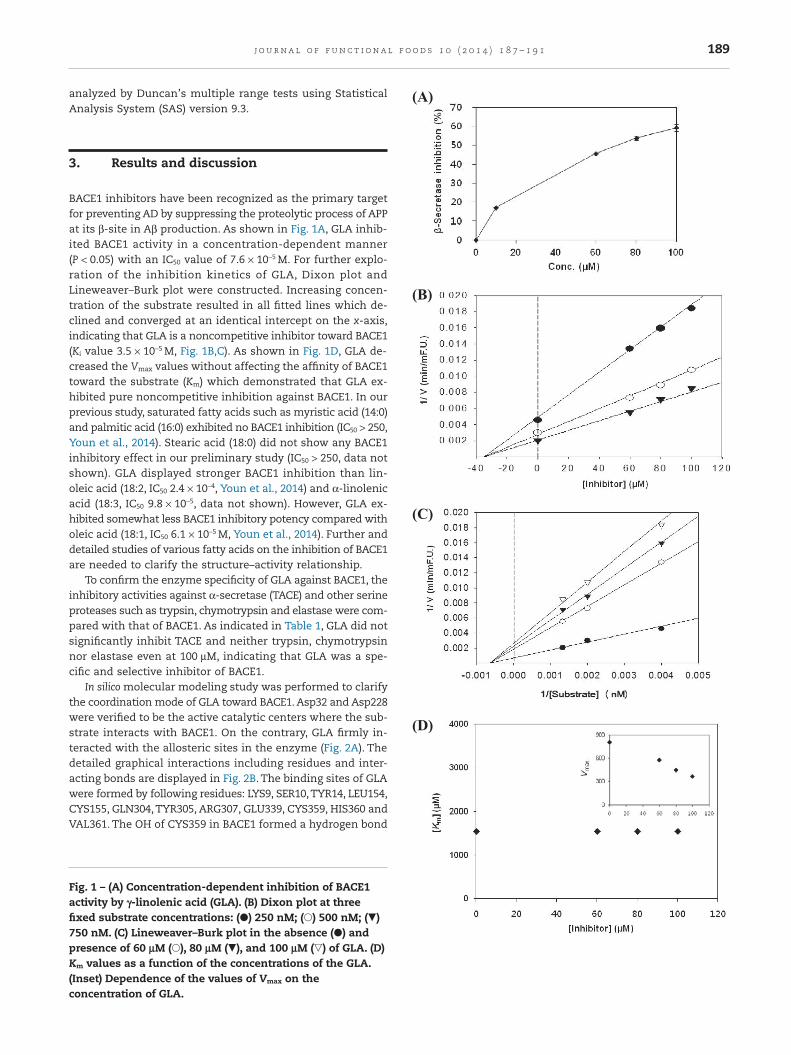
BACE1 inhibitors have been recognized as the primary targetfor preventing AD by suppressing the proteolytic process of APPat its β-site in Aβ production. As shown in Fig. 1A, GLA inhib-ited BACE1 activity in a concentration-dependent manner(P < 0.05) with an IC50 value of 7.6 × 10−5 M. For further explo-ration of the inhibition kinetics of GLA, Dixon plot andLineweaver–Burk plot were constructed. Increasing concen-tration of the substrate resulted in all fitted lines which de-clined and converged at an identical intercept on the x-axis,indicating that GLA is a noncompetitive inhibitor toward BACE1(Ki value 3.5 × 10−5 M, Fig. 1B,C). As shown in Fig. 1D, GLA de-creased the Vmax values without affecting the affinity of BACE1toward the substrate (Km) which demonstrated that GLA ex-hibited pure noncompetitive inhibition against BACE1. In ourprevious study, saturated fatty acids such as myristic acid (14:0)and palmitic acid (16:0) exhibited no BACE1 inhibition (IC50 > 250,Youn et al., 2014). Stearic acid (18:0) did not show any BACE1inhibitory effect in our preliminary study (IC50 > 250, data notshown). GLA displayed stronger BACE1 inhibition than lin-oleic acid (18:2, IC50 2.4 × 10−4, Youn et al., 2014) and α-linolenicacid (18:3, IC50 9.8 × 10−5, data not shown). However, GLA ex-hibited somewhat less BACE1 inhibitory potency compared witholeic acid (18:1, IC50 6.1 × 10−5 M, Youn et al., 2014). Further anddetailed studies of various fatty acids on the inhibition of BACE1are needed to clarify the structure–activity relationship.
To confirm the enzyme specificity of GLA against BACE1, theinhibitory activities against α-secretase (TACE) and other serineproteases such as trypsin, chymotrypsin and elastase were com-pared with that of BACE1. As indicated in Table 1, GLA did notsignificantly inhibit TACE and neither trypsin, chymotrypsinnor elastase even at 100 μM, indicating that GLA was a spe-cific and selective inhibitor of BACE1.
In silico molecular modeling study was performed to clarifythe coordination mode of GLA toward BACE1. Asp32 and Asp228were verified to be the active catalytic centers where the sub-strate interacts with BACE1. On the contrary, GLA firmly in-teracted with the allosteric sites in the enzyme (Fig. 2A). Thedetailed graphical interactions including residues and inter-acting bonds are displayed in Fig. 2B. The binding sites of GLAwere formed by following residues: LYS9, SER10,TYR14, LEU154,CYS155, GLN304, TYR305, ARG307, GLU339, CYS359, HIS360 andVAL361. The OH of CYS359 in BACE1 formed a hydrogen bond
Fig. 1 – (A) Concentration-dependent inhibition of BACE1activity by γ-linolenic acid (GLA). (B) Dixon plot at threefixed substrate concentrations: (●) 250 nM; (○) 500 nM; (▼)750 nM. (C) Lineweaver–Burk plot in the absence (●) andpresence of 60 μM (○), 80 μM (▼), and 100 μM (▽) of GLA. (D)Km values as a function of the concentrations of the GLA.(Inset) Dependence of the values of Vmax on theconcentration of GLA.
(A)
(B)
(C)
(D)
189j o u rna l o f f un c t i ona l f o od s 1 0 ( 2 0 1 4 ) 1 8 7 – 1 9 1

with the hydrogen atom of carboxylic acid moiety in GLA. Inaddition, the lowest energy conformation of the most pro-posed complex for GLA was −6.2 kcal/mol.
Previous studies in designing inhibitors of BACE1 havefocused on peptide-derived structures, which act as transition-state analogues based on the amino acid sequences at the APPcleavage sites of BACE1. Even though peptidomimetic species
displayed a nanomolar BACE1 inhibitory effect, lowbioavailability, high hydrophilicity, large molecular mass andrestricted brain penetration caused by binding affinity for trans-port protein have been their major obstacles and limitation tobe BACE1 inhibitors (Kang et al., 2013).
Based on efforts to discover natural molecules that con-trolled the generation of Aβ, several compounds have beenproven to exhibit their neuroprotection through suppressingthe amyloidogenic pathway. Rg1 from Panax notoginseng, greentea catechins, hispidin from Phellinus linteus, ellagic acid fromPunica granatum, isoflavones from Ficus benjamina, loganin fromCorni fructus, etc. (Dai, Shen, Yoshida, Parrish, & Williams, 2012;Jeon, Bae, Seong, & Song, 2003; Kwak et al., 2005; Park, Jeon,Lee, Kim, & Song, 2004; Wang & Du, 2009; Youn et al., 2013).Most of the previously discovered natural BACE1 inhibitorspossess small molecular weight and have similarity in theirstructures with aromatic rings including biphenyl, pentacyclic,stilbene/stilbene-like, benzopyran/benzofuran etc. (Jeon et al.,2007; Lv et al., 2008). However, in the current study we foundthat GLA, an extraordinary n-6 PUFA possessed a principal in-hibitory effect against BACE1.
Considering the discovery of therapeutics for AD, the com-pounds must traverse the blood–brain capillary wall whichforms blood–brain barrier (BBB). Only a limited class of com-pounds with low molecular weight less than 400–500 kDa withlipophilicility was demonstrated to cross BBB. The importmechanism of fatty acid into cells has been still controver-sial and only a few studies have investigated the mechanismof fatty acid transport across the BBB (Mitchell, Edmundson,Miller, & Hatch, 2009; Song et al., 2010). Indeed, recent encour-aging study revealed that PUFA is available to pass through theBBB using transport proteins such as FATP4, FABP5 and CD36(Mitchell, On, Del Bigio, Miller, & Hatch, 2011).
In conclusion, our results demonstrated that GLA exerteda significant and specific inhibitory effect against BACE1, andfirmly interacted with the allosteric sites in the enzyme. Eventhough comprehensive mechanistic studies need to be clari-fied, this study provides the scientific support for GLA as a po-tential BACE1 inhibitor, which furthermore might be a usefulreagent for studying both enzyme properties of BACE1 and non-synthetic BACE1 inhibitor in AD.
Acknowledgements
This research was supported by the Agenda Program fundedby Rural Development Administration (2012-PJ-008969), Re-public of Korea. The work was also partially supported by theBrain Busan 21 Program. Dr. Jinhyuk Lee was supported by agrant from the Korea Research Institute of Bioscience and Bio-technology (KRIBB) Research Initiative Program.
Table 1 – Inhibitory activity of γ-linolenic acid (GLA) against α-secretase (TACE) andother serine proteases.
Conc. (μM) TACE Trypsin Chymotrypsin Elastase
50 2.36 ± 0.19 1.62 ± 0.18 3.34 ± 1.18 1.25 ± 0.03100 4.83 ± 0.84 1.30 ± 0.37 0.21 ± 0.10 1.08 ± 0.17
(B)
(A)
Fig. 2 – In silico docking poses for γ-linolenic acid (GLA).Representative binding mode of the most stable dockingposes of GLA with human BACE1. (A) The complete viewof the docking poses of GLA. Human BACE1 is expressedas a gray solid ribbon diagram and GLA as a greenrepresentation. Asp32 and Asp228 are the active catalyticcenter residues marked in red color. (B) The close-up figureof GLA-BACE1 docking modes. Hydrogen bond interactionsbetween GLA and human BACE1 residues are displayed asa yellow dotted line. The structural ligands were performedby AutoDock Vina. (For interpretation of the references tocolor in this figure legend, the reader is referred to the webversion of this article.)
190 j o u rna l o f f un c t i ona l f o od s 1 0 ( 2 0 1 4 ) 1 8 7 – 1 9 1

R E F E R E N C E S
Butterfield, D. A., Castegna, A., Lauderback, C. M., & Drake, J.(2002). Evidence that amyloid beta-peptide-induced lipidperoxidation and its sequelae in Alzheimer’s disease braincontribute to neuronal death. Neurobiology of Aging, 23, 655–664.
Chang, C. S., Sun, H. L., Lii, C. K., Chen, H. W., Chen, P. Y., &Liu, K. L. (2010). Gamma-linolenic acid inhibits inflammatoryresponses by regulating NF-kappaB and AP-1 activation inlipopolysaccharide-induced RAW 264.7 macrophages.Inflammation, 33, 46–57.
Dai, J., Shen, D., Yoshida, W., Parrish, S., & Williams, P. (2012).Isoflavonoids from Ficus benjamina and their inhibitoryactivity on BACE1. Planta Medica, 78, 1357–1362.
De Felice, F. G., Wu, D., Lambert, M. P., Fernandez, S. J., Velasco, P.T., Lacor, P. N., Bigio, E. H., Jerecic, J., Acton, P. J., Shughrue, P. J.,Chen-Dodson, E., Kinney, G. G., & Klein, W. L. (2008).Alzheimer’s disease-type neuronal tau hyperphosphorylationinduced by Aβ oligomers. Neurobiology of Aging, 29, 1334–1347.
Dines, K. C., Cameron, N. E., & Cotter, M. A. (1995). Comparison ofthe effects of evening primrose oil and triglyceridescontaining γ-linolenic acid on nerve conduction and bloodflow in diabetic rats. The Journal of Pharmacology andExperimental Therapeutics, 273, 49–55.
Hardy, J., & Allsop, D. (1991). Amyloid deposition as the centralevent in the etiology of Alzheimer’s disease. TrendsPharmacological Science, 12, 383–388.
Horrobin, D. F. (1997). Essential fatty acids in the management ofimpaired nerve function in diabetes. Diabetes, 46, 590–593.
Janssen, C. I. F., & Kiliaan, A. J. (2014). Long-chain polyunsaturatedfatty acids (LCPUFA) from genesis to senescence: Theinfluence of LCPUFA on neural development, aging, andneurodegeneration. Progress in Lipid Research, 53, 1–17.
Jeon, S. Y., Bae, K., Seong, Y. H., & Song, K. S. (2003). Green teacatechins as a BACE1 inhibitor. Bioorganic & MedicinalChemistry Letters, 22, 3905–3908.
Jeon, S. Y., Kwon, S. H., Seong, Y. H., Bae, K., Hur, J. M., Lee, Y. Y.,Suh, D. Y., & Song, K. S. (2007). β-Secretase (BACE1)-inhibitingstilbenoids from Smilax Rhizoma. Phytomedicine: InternationalJournal of Phytotherapy and Phytopharmacology, 14, 403–408.
Jeppsson, F., Ekätjall, S., Janson, J., Karlström, S., Gustavsson, S.,Olsson, L. L., Radesätter, A. C., Pleoger, B., Cebers, G.,Kolmodin, K., Swahn, B. M., Berg, S., Bueters, T., & Fälting, J.(2012). Discovery of AZD3839, a potent and selective BACE1inhibitor clinical candidate for the treatment of Alzheimer’sdisease. The Journal of Biological Chemistry, 287, 41245–41257.
Kang, J. E., Cho, J. K., Curtis-Long, M. J., Ryo, H. W., Kim, J. H., Kim,H. J., Yuk, H. J., Kim, D. W., & Park, H. K. (2013). Inhibitoryevaluation of sulfonamide chalcones on β-secretase andacetylcholinesterase. Molecules, 18, 140–153.
Kapoor, R., & Huang, Y. S. (2006). Gamma linolenic acid: An anti-inflammatory omega-6 fatty acid. Current PharmaceuticalBiotechnology, 7, 531–534.
Kwak, H. M., Jeon, S. Y., Song, B. H., Kim, J. G., Lee, J. M., Lee, K. B.,Jeong, H. H., Hur, J. M., Kang, Y. H., & Song, K. S. (2005).β-Secretase (BACE1) inhibitors from pomegranate (Punicagranatum) husk. Archives of Pharmacal Research, 28, 1328–1332.
Lv, L., Yang, Q. Y., Zhao, Y., Yao, C. S., Sun, Y., Yang, E. J., Song, K.S., Jung, I. M., & Fang, W. S. (2008). BACE1 (β-secretase)inhibitory chromone glycosides from Aloe vera and Aloenobilis. Planta Medica, 74, 540–545.
Mitchell, R. W., Edmundson, C. L., Miller, D. W., & Hatch, G. M.(2009). On the mechanism of oleate transport across human
brain microvessel endothelial cells. Journal of Neurochemistry,110(3), 1049–1057.
Mitchell, R. W., On, N. H., Del Bigio, M. R., Miller, D. W., & Hatch, G.M. (2011). Fatty acid transport protein expression in humanbrain and potential role in fatty acid transport across humanbrain microvessel endothelial cells. Journal of Neurochemistry,117, 735–746.
Montserrat-de la Paz, S., García-Giménez, M. D., Ángel-Martín, M.,Marín-Aguilar, F., & Fernández-Arche, A. (2013). Dietarysupplementation evening primrose oil improves symptoms offibromyalgia syndrome. Journal of Functional Foods, 5(3), 1279–1287.
Nicholls, A., McGaughey, G. B., Sheridan, R. P., Good, A. C.,Warren, G., Mathieu, M., & Kelley, B. (2010). Molecular shapeand medicinal chemistry: A perspective. Journal of MedicalChemistry, 53(10), 3862–3886.
Park, I. H., Jeon, S. Y., Lee, H. J., Kim, S. I., & Song, K. S. (2004). Aβ-secretase (BACE1) inhibitor hispidin from the mycelialcultures of Phellinus linteus. Planta Medica, 70,143–146.
Shimmyo, Y., Kihara, T., Akaike, A., Niidome, T., & Sugimoto, H.(2008). Flavonols and flavones as BACE1 inhibitors: Structure-activity relationship in cell-free, cell-based and in silicostudies reveal novel pharmacophore features. Biochimica etBiophysica Acta, 1780, 819–825.
Sinha, S., Anderson, J. P., Barbour, R., Basi, G. S., Caccavello, R.,Davis, D., Doan, M., Dovey, H. F., Frigon, N., Hong, J., Jacobson-Croak, K., Jewett, N., Keim, P., Knops, J., Lieberburg, I., Power,M., Tan, H., Tatsuno, G., Tung, J., Schenk, D., Seubert, P.,Suomensaari, S. M., Wang, S., Walker, D., Zhao, J., McConlogue,L., & John, V. (1999). Purification and cloning of amyloidprecursor protein β-secretase from human brain. Nature, 402,537–540.
Song, B. J., Elbert, A., Rahman, T., Orr, S. K., Chen, C. T., Febbraio,M., & Bazinet, R. P. (2010). Genetic ablation of CD36 does notalter mouse brain polyunsaturated fatty acid concentrations.Lipids, 45, 291–299.
Song, C., Manku, M. S., & Horrobin, D. F. (2008). Long-chainpolyunsaturated fatty acids modulate interleukin-1β-inducedchanges in behavior, monoaminergic neurotransmitters, andbrain inflammation in rats. The Journal of Nutrition, 138, 954–963.
Tian, Y., Bassit, B., Chau, D., & Li, Y. (2010). An APP inhibitorydomain containing the Flemish mutation residue modulatesγ-secretase activity for Aβ production. Nature Structural &Molecular Biology, 17, 151–158.
Tong, Y., Zhou, W., Fung, V., Christensen, M. A., Qing, H., Sun, X., &Song, W. (2005). Oxidative stress potentiates BACE1 geneexpression and Aβ generation. The Journal of NeuralTransmission, 112, 455–469.
Trott, O., & Olson, A. J. (2010). AutoDock Vina: Improving thespeed and accuracy of docking with a new scoring function,efficient optimization, and multithreading. The Journal ofComputational Chemistry, 31, 455–461.
Wang, H. Y., & Du, G. H. (2009). Ginsenoside Rg1 inhibitsβ-secretase activity in vitro and protects against Aβ-inducedcytotoxicity in PC12 cells. The Journal of Asian Natural ProductsResearch, 11, 604–612.
Youn, K., Jeong, W. S., & Jun, M. (2013). β-Secretase (BACE1)inhibitory property of loganin isolated from Corni fructus.Natural Product Research, 27, 1471–1474.
Youn, K., Yun, E. Y., Lee, J., Kim, J. Y., Hwang, J. S., Jeong, W. S., &Jun, M. (2014). Oleic acid and linoleic acid from Tenebrio molitorlarvae inhibit BACE1 activity in vitro: Molecular dockingstudies. Journal of Medicinal Food, 17, 284–289.
191j o u rna l o f f un c t i ona l f o od s 1 0 ( 2 0 1 4 ) 1 8 7 – 1 9 1


















