
Activation of the oxidative stress pathway by HIV-1 Vpr ... · PDF fileproduction of...
21
Induction of HIF-1α by Vpr Deshmane et al. 1 Activation of the oxidative stress pathway by HIV-1 Vpr leads to induction of hypoxia inducible factor 1 alpha expression Satish L. Deshmane 1* , Ruma Mukerjee 1*§ , Shongshan Fan 1‡ , Luis Del Valle 1 , Carine Michiels 2 , Thersa Sweet 1,3 , Inna Rom 1 , Kamel Khalili 1 , Jay Rappaport 1 , Shohreh Amini 1,3 , and Bassel E Sawaya 1§¶ . 1 Department of Neuroscience & Center for Neurovirology, Temple University School of Medicine, 1900 North 12 th Street, Philadelphia, PA 19122. 2 Unité de Recherche en Biologie Cellulaire, University of Namur-FUNDP, Namur, Belgium. 3 Department of Biology, College of Science and Technology, Temple University, Philadelphia, PA 19122. * Equal Contribution ¶ Corresponding Author: Bassel E. Sawaya, PhD, Department of Neuroscience & Center for Neurovirology, Temple University School of Medicine, 1900 North 12 th Street, Philadelphia, PA 19122. § Present Address: Department of Neurology, Laboratory of Molecular Virology, Temple University School of Medicine, 3400 North Broad Street, Philadelphia, PA 19140. Phone: 215-707 5446; Fax: 215-707 5948; E-mail: [email protected]. ‡ Department of Pathology and laboratory Medicine, Temple University School of Medicine, 3401 North Broad Street, Philadelphia, PA 19140. ABSTRACT The detection of biomarkers of oxidative stress in brain tissue and cerebrospinal fluid of patients with HIV-1 dementia indicates the involvement of stress pathways in the neuropathogenesis of AIDS. While the biological importance of oxidative stress on events involved in AIDS neuropathogenesis and the HIV-1 proteins responsible for oxidative stress remain to be elucidated, our results point to the activation of hypoxia-inducible factor 1 (HIF-1) upon HIV-1 infection and its elevation in brain cells of AIDS patients with dementia. HIF-1 is a transcription factor that is responsive to oxygen. Under hypoxic conditions, HIF-1α becomes stable and translocates to the nucleus where it dimerizes with ARNT and modulates gene transcription. Activation of HIF-1 can also be mediated by the HIV-1 accessory protein, Vpr. In addition, cellular events including reactive oxygen species (ROS) contribute to the induction of HIF-1α. Our results show that Vpr induces ROS by increasing H 2 O 2 production, which can contribute to HIF-1α accumulation. Interestingly, increased levels of HIF-1α stimulated HIV-1 gene transcription through HIF-1 association with HIV-1 LTR DNA. These observations point to the existence of a positive feedback interplay between HIF-1α and Vpr and that, by inducing oxidative stress via activation of HIF-1, Vpr can induce HIV-1 gene expression and dysregulate multiple host cellular pathways. INTRODUCTION Oxidative stress (OS) is a phenomenon occurring in cells when production of oxygen radicals exceeds antioxidant capacity (1). An excess of free radicals damages essential macromolecules of the cell, leading to abnormal gene expression, disturbances in receptor activity, cell proliferation, cell death, or immune perturbation. OS is the major player in numerous human diseases such as cancer, ocular degeneration, and neurodegenerative diseases including AIDS-associated neuropathies (2). OS is involved in many aspects of HIV disease pathogenesis, including viral replication (3), inflammatory responses, decreased immune cell proliferation, loss of immune function, chronic weight loss and increased sensitivity to drug toxicity (4). Active replication of HIV in macrophages and microglia represents a reservoir for virus and an important step for HIV neuropathogenesis (5). This process leads to production of inflammatory products and free radical species (6). Because macrophages and microglia function as a long-term reservoir for HIV-1, these cells must possess a mechanism to protect themselves against the toxic effects of superoxide anions. In this regard, HIV-1 Tat has been shown to induce neuronal death via TNF-α induction and activation of the OS pathway (7, 8). In another study, injection of HIV-1 Tat in the rat striatum was shown to lead to activation of the OS pathway (9). In addition to Tat, HIV-1 gp120 protein has also been shown to cause reactive oxygen species (ROS) production in glial cells leading to neurodegeneration and apoptosis (10). Finally, in a recent study, HIV-1 Vpr protein was shown to be involved in OS pathway (11). During contact with HIV-infected macrophages or microglia, endothelial cells might contribute to their own damage as a result of the http://www.jbc.org/cgi/doi/10.1074/jbc.M809266200 The latest version is at JBC Papers in Press. Published on February 9, 2009 as Manuscript M809266200 Copyright 2009 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on May 25, 2018 http://www.jbc.org/ Downloaded from
Transcript of Activation of the oxidative stress pathway by HIV-1 Vpr ... · PDF fileproduction of...

Induction of HIF-1α by Vpr Deshmane et al.
1
Activation of the oxidative stress pathway by HIV-1 Vpr leads to induction of hypoxia inducible factor 1 alpha expression
Satish L. Deshmane1*, Ruma Mukerjee1*§, Shongshan Fan1‡, Luis Del Valle1, Carine Michiels2, Thersa Sweet1,3,
Inna Rom1, Kamel Khalili1, Jay Rappaport1, Shohreh Amini1,3, and Bassel E Sawaya1§¶.
1Department of Neuroscience & Center for Neurovirology, Temple University School of Medicine, 1900 North 12th Street, Philadelphia, PA 19122. 2Unité de Recherche en Biologie Cellulaire, University of Namur-FUNDP, Namur, Belgium. 3Department of Biology, College of Science and Technology, Temple University, Philadelphia, PA 19122.
* Equal Contribution
¶ Corresponding Author: Bassel E. Sawaya, PhD, Department of Neuroscience & Center for Neurovirology, Temple University School of Medicine, 1900 North 12th Street, Philadelphia, PA 19122.
§ Present Address: Department of Neurology, Laboratory of Molecular Virology, Temple University School of Medicine, 3400 North Broad Street, Philadelphia, PA 19140. Phone: 215-707 5446; Fax: 215-707 5948; E-mail: [email protected]. ‡ Department of Pathology and laboratory Medicine, Temple University School of Medicine, 3401 North Broad Street, Philadelphia, PA 19140. ABSTRACT
The detection of biomarkers of oxidative stress in brain tissue and cerebrospinal fluid of patients with HIV-1 dementia indicates the involvement of stress pathways in the neuropathogenesis of AIDS. While the biological importance of oxidative stress on events involved in AIDS neuropathogenesis and the HIV-1 proteins responsible for oxidative stress remain to be elucidated, our results point to the activation of hypoxia-inducible factor 1 (HIF-1) upon HIV-1 infection and its elevation in brain cells of AIDS patients with dementia. HIF-1 is a transcription factor that is responsive to oxygen. Under hypoxic conditions, HIF-1α becomes stable and translocates to the nucleus where it dimerizes with ARNT and modulates gene transcription. Activation of HIF-1 can also be mediated by the HIV-1 accessory protein, Vpr. In addition, cellular events including reactive oxygen species (ROS) contribute to the induction of HIF-1α. Our results show that Vpr induces ROS by increasing H2O2 production, which can contribute to HIF-1α accumulation. Interestingly, increased levels of HIF-1α stimulated HIV-1 gene transcription through HIF-1 association with HIV-1 LTR DNA. These observations point to the existence of a positive feedback interplay between HIF-1α and Vpr and that, by inducing oxidative stress via activation of HIF-1, Vpr can induce HIV-1 gene expression and dysregulate multiple host cellular pathways. INTRODUCTION
Oxidative stress (OS) is a phenomenon occurring in cells when production of oxygen
radicals exceeds antioxidant capacity (1). An excess of free radicals damages essential macromolecules of the cell, leading to abnormal gene expression, disturbances in receptor activity, cell proliferation, cell death, or immune perturbation. OS is the major player in numerous human diseases such as cancer, ocular degeneration, and neurodegenerative diseases including AIDS-associated neuropathies (2).
OS is involved in many aspects of HIV disease pathogenesis, including viral replication (3), inflammatory responses, decreased immune cell proliferation, loss of immune function, chronic weight loss and increased sensitivity to drug toxicity (4). Active replication of HIV in macrophages and microglia represents a reservoir for virus and an important step for HIV neuropathogenesis (5). This process leads to production of inflammatory products and free radical species (6). Because macrophages and microglia function as a long-term reservoir for HIV-1, these cells must possess a mechanism to protect themselves against the toxic effects of superoxide anions. In this regard, HIV-1 Tat has been shown to induce neuronal death via TNF-α induction and activation of the OS pathway (7, 8). In another study, injection of HIV-1 Tat in the rat striatum was shown to lead to activation of the OS pathway (9). In addition to Tat, HIV-1 gp120 protein has also been shown to cause reactive oxygen species (ROS) production in glial cells leading to neurodegeneration and apoptosis (10). Finally, in a recent study, HIV-1 Vpr protein was shown to be involved in OS pathway (11).
During contact with HIV-infected macrophages or microglia, endothelial cells might contribute to their own damage as a result of the
http://www.jbc.org/cgi/doi/10.1074/jbc.M809266200The latest version is at JBC Papers in Press. Published on February 9, 2009 as Manuscript M809266200
Copyright 2009 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
2
production of ROS (12). ROS including superoxide (O2
-), hydrogen peroxide (H2O2), hydroxyl radical (OH), and reactive nitrogen species, such as nitric oxide (NO) and peroxynitrite (NO3
-) are biologically active species that are increasingly recognized to play major roles in vascular biology through redox signaling (13). Cytokines could form a pivotal link in ROS-dependent pathways leading to the activation of redox-sensitive transcription factors, such as the hypoxia inducible factor 1 (HIF-1), whose upregulation determines the specificity of cellular responses to oxidative stress (14).
HIF is an oxygen sensor and master regulator in unicellular and multicellular organisms (15), and is involved in the response to low oxygen levels (hypoxia), inducing changes in gene expression (16). HIF regulates glycolysis, mitochondrial oxygen consumption, erythropoiesis, angiogenesis, and cellular survival (17). HIF is a heterodimer of two basic helix-loop-helix/PAS proteins containing HIF-1α or HIF-2α and the aryl hydrocarbon nuclear translocator (ARNT or HIF-1β) (18). HIF heterodimers bind to the hypoxia response element (HRE), a 5’-RCGTG-3’ consensus sequence (19). HIF-1 activation is modulated by TNF-α, ROS and nitric oxide and/or NO-derived species (NOS). ROS contributes to the accumulation and stabilization of HIF-1α (20, 21). Under hypoxic conditions, TNF-α promotes a signaling cascade, which depends on the translocation of NF-κB and leads to accumulation of HIF-1α protein (22). Finally, HIF-1α is phosphorylated (23). In this regard, HIF-1α has been shown to be a substrate for various protein kinase pathways including the mitogen-activated protein kinases (MAPKs) pathway.
Viral protein R, Vpr is a highly conserved HIV-1 accessory protein, involved in viral replication, transactivation of long terminal repeat (LTR), nuclear localization of the pre-integration complex in non-dividing cells, cell cycle arrest, DNA damage and apoptosis (24-26). Vpr causes apoptosis through suppression of NF-κB activity or through rapid dissipation of the mitochondrial transmembrane potential, through its association with ANT leading to the release of cytochrome c, and activation of caspase 3 (27, 28). Finally, Vpr was recently shown to cause neuronal death through convergent pathogenic mechanisms with ensuing in vivo neurodegeneration (29).
We have now identified Vpr as one of the HIV-1 proteins involved in triggering free radical
production. This occurs via increased production of ROS leading to HIF-1α accumulation in microglia. Accumulation of HIF-1α then upregulates the HIV-1 promoter. These data reveal a new function for Vpr regarding its ability to induce oxidative stress (OS) and shed light on the balance that governs the mechanisms used by HIV-1 to activate the OS pathway, and its involvement in neuroAIDS. MATERIALS AND METHODS
Plasmids. HIV-1 LTR Luciferase reporter plasmid and Vpr expression plasmid were previously described (30). CMV-HIF-1α was a gift from Dr. Steve McKnight (University of Texas Southwestern Medical Center, Dallas, TX). The HIF-1α promoter fused to Luciferase reporter plasmid was previously described (31, 32).
Cell Culture, Transfection and Transduction Assays. The human microglial cell line (33) was maintained in DMEM + 10% FBS. Cells were transfected with 0.5µg of reporter plasmid (LTR-Luc or HIF-1α-Luc) or co-transfected with 0.5µg of various expression cDNAs as indicated (34). Separately, the cells were transduced with adeno-Vpr or adeno-null. The amount of DNA used for each transfection was normalized with pcDNA3 vector plasmid. Each transfection was repeated at least three times with different plasmid preparations. Cell extracts were prepared 48h after transfection, and luciferase assays were performed (Promega, Madison, WI).
Recombinant adenoviruses. Vpr cDNA (288 bp) was excised from pcDNA3-Vpr and cloned into the EcoRI and NheI sites of the adenovirus-shuttle plasmid pDC515 under the control of the murine cytomegalovirus promoter (purchased from Microbix Inc. Ontario, Canada). Adeno-Vpr recombinant shuttle containing Vpr sequence (pDC515-Vpr) was transfected into HEK-293 cells with pBHGfrt (del) E1, 3FLP, and a plasmid that provides adenovirus type-5 genome deleted in E1 and E3 genes. Plaques of recombinant adenovirus arising as a result of frt/FLP recombination were isolated, grown and purified by cesium chloride density equilibrium banding as previously described (35). Empty shuttle plasmid, pDC515, was used to construct control adenoviral vector (Adeno-null, a virus without a transgene). Adeno-Vpr or adeno-Null was used at an MOI of 5 plaques forming units per cell. [MOI= multiplicity of infection].
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
3
Hydrogen peroxide production assay. Microglia were grown in DMEM with 10% FBS and seeded in 6 well plates. Cells were transduced with adeno-null or adeno-Vpr virus at an MOI of 5 for 48h. Uninfected cells were also collected as a control. Cells were lysed by freeze and thaw method three times. Protein was measured using Biorad protein assay reagent. Equal amounts of protein were used in H2O2 assays with the Quantichrom Peroxide assay kit (DIOX-250) from Bioassay systems. The kit is designed to measure peroxide concentration in biological samples without any pretreatment. The method utilizes the chromogenic Fe3+ xynol orange reaction, in which a purple complex formed when Fe2+ provided in the reagent is oxidized to Fe3+ by peroxides present in the sample. The intensity of the color, measured at 540-610nm, is an accurate measure of peroxide level in the sample. A standard curve was generated as recommended by the manufacturer using known amounts of H2O2. (Conversions: 1µM H2O2 equals 34ng/ml).
Western blot. Microglia were infected with adeno-null or adeno-Vpr, or -Tat virus at an MOI of 5 at 37°C. Twenty-four hours post-infection, 30µg of cell extracts were subjected to Western blot analysis using anti-HIF-1α, -Vpr or anti-Grb2, antibodies using 1/1000 dilution. For HIF-1α detection, two different anti-HIF-1α antibodies were used. The HIF-1α antibodies were purchased either from BD Biosciences, (San Jose, CA) or from Abcam (Cambridge, MA). Interestingly and according to the manufacturer catalogue, antibody purchased from BD Biosciences detects two bands of HIF-1α, while Abcam antibody detects only one band of HIF-1α. Note that all Western blot assays were performed at least three times.
RNA interference. Transient knockdown of HIF-1α was performed with a HIF-1α-specific siRNA (Dharmacon Research Inc., Lafayette, CO). Twenty-four hours after cell plating, microglial cells were rinsed once with Optimem. siRNA was added at a final concentration of 50nM. Western blot was performed with protein extracts from untransfected cells or cells transfected with HIF-1α specific siRNA using anti-HIF-1α antibody. For a loading control, anti-Grb2 antibody was used. Control non-targeting siRNA was also used (Dharmacon).
Tissue Acquisition. Archival autopsy samples from the frontal lobe of HIV-Encephalitis cases
and matching normal brain samples have been collected from the Manhattan Brain Bank at the Mount Sinai School of Medicine, New York, under Institutional Review Board (IRB) approval by Temple University Office for Human Subjects Protection.
Immunohistochemistry. The tissue, which was formalin fixed and paraffin embedded, was sectioned at 5µm thickness and placed on electromagnetically charged glass slides. Sections were processed for immunohisto-chemistry as previously described (36). A rabbit polyclonal anti-HIF-1α antibody was utilized as primary antibody. After rinsing with PBS, sections were incubated for one hour at room temperature with biotinylated anti-rabbit secondary antibody, followed by one hour of incubation with Avidin-Biotin-Peroxidase complexes (ABC Elite Kit, Vector Laboratories, Burlingame, California) and developed with Diaminobenzydine (DAB), counterstained with Hematoxylin and mounted for histological evaluation.
ROS measurement. The trafficking of 2,3,4,5, 6-pentafluorodihydrotetramethylfosamine (PF-H2TMROS or Redox Sensor Red CC-1; Molecular Probes-Invitrogen, Carlsbad, CA) was used to detect reactive oxygen intermediates (37). Redox Sensor Red CC-1 is oxidized in the presence of O2
– and H2O2. Briefly, microglial cells mock-, ad-null- or adeno-Vpr infected were loaded at 37°C for 20 min with 1µM of Redox Sensor Red CC-1 and a mitochondria-specific dye, MitoTracker
green FM (50nM; Molecular Probes). Culture slides were washed with PBS and visualized with a Nikon fluorescence microscope (Nikon Eclipse E800) equipped with a triple-filter cube and
charge-coupled device camera (Nikon DXM1200). Note that all ROS measurement experiments were performed at least three times.
Chromatin Immunoprecipitation (ChIP) Assay. HeLa 3T1 (HL3T1) cells were grown overnight in 100-mm dishes to 60-70% confluency; cells were then transfected with 50 nM of siRNA-HIF-1α and/or transduced with Ad-Vpr or Ad-Null at an MOI of 5 using lipofectamine transfection reagent (Roche Applied Sciences). Plates were returned to the incubator for 40-48 h. Cells were cross-linked with formaldehyde, harvested, and ChIP was performed. For these studies, only 5 x 106 cells were used per immunoprecipitation reaction because the plasmid is present at a high copy number. The remainder of the procedure followed
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
4
standard protocols for ChIP analysis, as recommended by the manufacturer (Upstate Biotechnology, address). The resulting DNA was analyzed by PCR reactions using the following HIV-LTR primers: (coding)(-120/+66) 5’ aactggtac catcgagcttgct 3’, and (non-coding) (+66/-120) 5’ ttgaggatccagcagtgggttc 3’. Antibody used in the ChIP procedure was against HIF-1α (purchased from BD Biosciences) as well as rabbit anti-mouse IgG. All ChIP assays were performed twice.
RNA Extraction and Northern blot analysis. RNA was purified using the RNeasy kit from Ambion (Austin, TX) and used for Northern blot analysis as described previously (38). For generation of probes for Northern blot analysis, cDNA of human full length HIF-1α (~2.48 kb) or 5.8S rRNA cDNAs were labeled using a random primed labeling reaction with Klenow enzyme and α-[32P]-dCTP. The Northern blot experiment was performed three times.
HIV-1 infection. The human U-937 monocytic cell line was maintained in RPMI + 10% FBS, 100 units/ml penicillin, 50µg/ml streptomycin-G. Cells in log phase were infected with pNL4-3, pNL4-3 ΔVpr or JR-FL strains of HIV-1 as follows. Fifty nanograms of p24-containing virus stock were added per 1x 106 cells. Cells were incubated with virus stock in a small volume of serum-free medium for 2h at 37˚C. The cells were then washed twice with PBS and fresh medium containing 2% of FBS was added (500,000 cells /ml). All infection experiments were performed three times.
Statistical analyses. Densitometric analyses were performed using EZQuant-Gel software (EZQuant Biology, Israel). The obtained data are expressed as the mean ± SE. Statistical analysis of parameters was performed using the one-way ANOVA test, followed by the unpaired Student’s t test, with P value less than 0.05 considered statistically significant
RESULTS
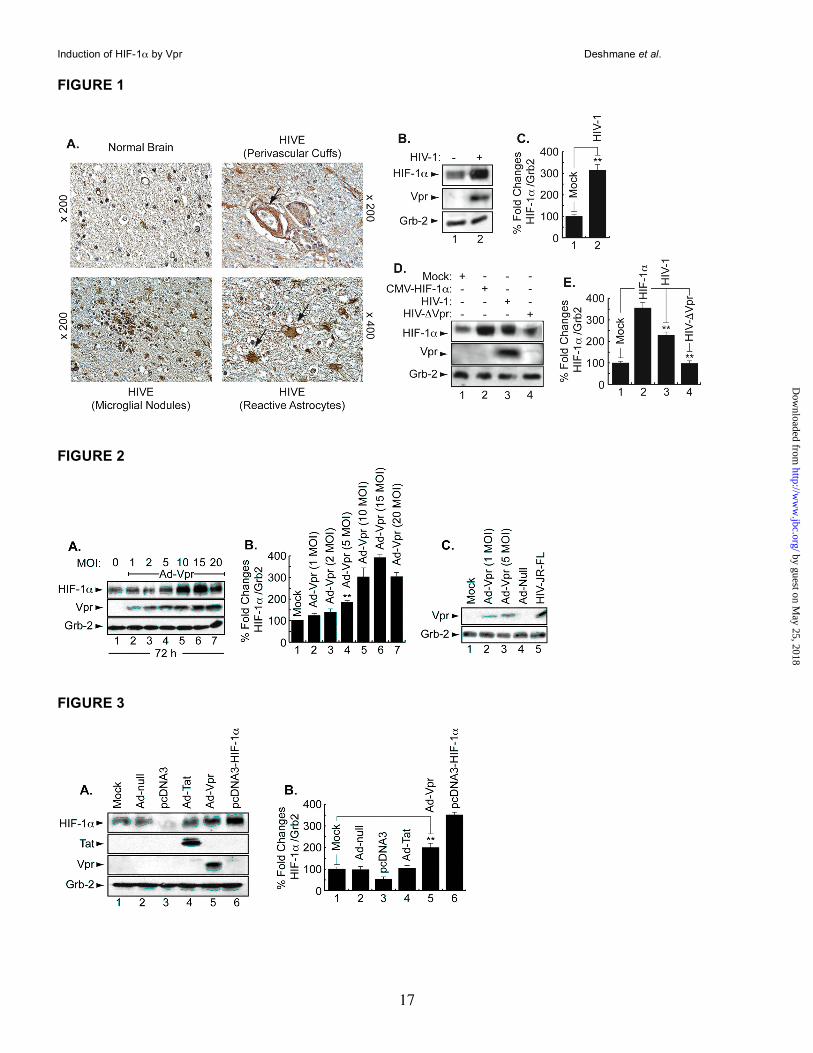
Induction of HIF-1α protein in the brain of HIV-1-infected patients. First, we examined the level of HIF-1α in brain tissue from HIVE patients. Figure 1A shows white matter of an HIVE case, where numerous reactive astrocytes with enlarged nuclei and abundant eosinophilic cytoplasm are present (arrows). HIF-1α was found in low levels in the cytoplasm of astrocytes
in normal brain, compared to the robust immunolabeling that was observed in the cytoplasm of reactive astrocytes and in macrophages and microglial nodules in HIVE white matter indicating increased levels of HIF-1α in HIVE.
Induction of HIF-1α in HIV-1 infected microglia. Next, we examined the levels of HIF-1α in HIV-infected microglia. Primary cultures of human microglial cells (1x106) were infected with 50ng (p24) of the JR-FL strain of HIV-1 in 200µl of serum free medium for 2 h at 37°C. Cells were washed twice with PBS and maintained in fresh medium containing 2% FBS added (500,000 cells/ml). After 5 days, cells were harvested and total protein extracts were prepared and analyzed by Western blot. As shown in Figure 1B, HIF-1α was induced in HIV-1-infected cells compared to the mock-infected cells. Infection of the cells with HIV-1 was verified, in parallel, by Western blot using anti-Vpr antibody. The levels of the housekeeping protein Grb-2 served as internal controls for equal protein loading. Densitometry was used to quantitate the fold changes in HIF-1α protein prepared from uninfected or infected cells observed in Panel B and is presented in Panel C.
Expression of HIF-1α in the absence of Vpr in HIV-1 infected cells. To investigate the importance of Vpr in HIF-1α accumulation during the course of HIV-1 infection, human microglial cells were infected with HIV-1pNL4-3 or HIV-1pNL4-
3ΔVpr [HIVΔVpr] (where Vpr is deleted) or transfected with pcDNA3-HIF-1α. After 48h, protein extracts were prepared and analyzed (30µg of lysate) by Western blot. HIF-1α was induced in HIV-infected or HIF-transfected cells (Figure 1D, HIF panel, compare lane 1 to lanes 2 and 3). HIF-1α was weakly induced in cells infected with Vpr-deleted HIV-1 [HIVΔVpr] (lane 4), indicating a small role for the other viral proteins, such as Tat, in HIF-1α induction. The efficiency of infection was monitored by Western blot using anti-Vpr antibody. As shown in Figure 1D, Vpr was detected only in extracts from HIV-1 infected cells and not from mock infected or HIVΔVpr cells (Vpr panel, lane 3). Grb2 levels are shown as a loading control. Note that HIV-1pNL4-3 is a T-cell tropic virus and cannot infect primary microglia. However, our experiment was performed in a microglial cell line, and the ability of HIV-1pNL4-3 to replicate in this particular microglial cell line was confirmed by PCR using
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
5
several LTR primers (data not shown). Differences in the levels of expression of HIF-1α protein were further evaluated by densitometric analysis using Scion image software (Scion Image Corps). As shown in Figure 1 (Panels C and E), HIF-1α protein levels increased by almost 3- (Panel C) and 2- (Panel E) fold in HIV-infected cells, respectively. Almost no change was observed in the HIF-1α protein levels in Cells infected with HIV-1 that lacks Vpr (Panel E, lane 4).
Induction of HIF-1α by Adenovirus-Vpr. Next, we examined whether induction of HIF-1α by Vpr is dose-dependent. Microglial cells were infected with different Ad-Vpr concentrations (1, 2, 5, 10, 15, and 20 MOI) for 72h after which the cells were washed and cellular proteins were collected and analyzed by Western blot. As shown in Figure 2A, HIF-1α was induced by Ad-Vpr at an MOI as low as 1 (lane 2). Almost no difference was observed in HIF-1α induction in cells infected with Ad-Vpr at MOIs of 5, 10, 15 and 20 (compare lane 4 to lanes 5-7). Differences in the levels of expression of HIF-1α protein were further evaluated by densitometric analysis (Panel B). Since Ad-Vpr at an MOI of 5 was enough to induce HIF-1α, it was necessary to examine whether this concentration is physiologically significant. To this end, microglial cells were infected with of Ad-Vpr at an MOI of 1 and 5, Ad-Null at an MOI of 5, while U937 cells were infected with JR-FL strain of HIV-1. Cellular proteins were prepared from Ad-null or Ad-Vpr-infected cells 48 post-infection, while collected 8 days post-infection from U937 cells infected with HIV-1. Thirty micrograms of extracts were subjected to Western blot analysis using anti-Vpr antibody. As shown in Figure 2C, the amount of Vpr detected in extracts collected from HIV-1 infected cells was considerably higher than the amount of Vpr detected in cells infected with Ad-Vpr (compare lane 5 to lanes 2 and 3). Vpr was not detected in mock or Ad-null-infected cells (lanes 1 and 4). Based on these results, we used Ad-Vpr at an MOI of 5 in all experiments.
Expression of HIF-1α in the presence of Tat and Vpr. In light of the previous results showing the ability of Tat and Vpr to induce oxidative stress (6, 11, 39, 40), we examined the effect of Tat (101 aa) and Vpr on HIF-1α expression in microglial cells. Human microglia were transduced with Ad-Tat, Ad-Vpr, or Ad-Null at an MOI of 5.
After 48h, cells were harvested and analyzed by Western blot using anti-HIF-1α antibody (Abcam). Mock-infected cells or cells transfected with pcDNA3 were used as a negative control, while pcDNA3-HIF1α-transfected cells were used as a positive control (Figure 3A, lanes 3 and 6, respectively). HIF-1α was weakly detected in control cells (lanes 1-3). Infection of cells with Ad-Tat had no significant effect on HIF-1α in microglial cells (lane 4), whereas the expression of Vpr significantly increased HIF-1α production (lane 5). Anti-Tat and -Vpr antibodies were used as controls to show expression of both proteins. Anti-Grb2 was used as a control for equal protein loading. Differences in the levels of expression of HIF-1α protein were further evaluated by densitometric analysis (Panel B).
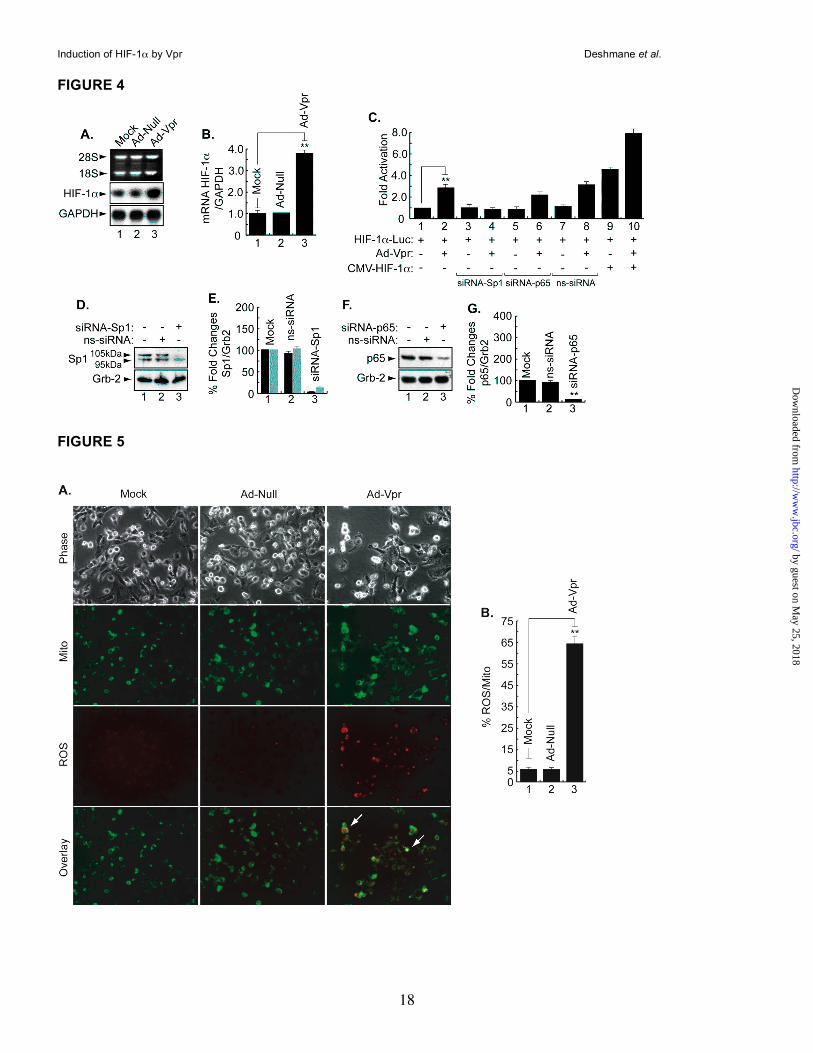
Vpr induces HIF-1α at the level of transcription. Induction of HIF-1α by Vpr gave us a rationale to investigate the ability of Vpr to affect expression of HIF-1α at the RNA level. To this end, microglial cells were infected with Ad-null or Ad-Vpr for 48h at an MOI of 5 after which total RNA was prepared and analyzed by Northern blot using HIF-1α cDNA as a probe. As shown in Figure 4A, HIF-1α RNA was induced in Vpr-infected cells but not in mock or Ad-null infected cells (compare lane 3 to lanes 1 and 2). GAPDH was used as the RNA loading control. Panel B displays the fold changes in mRNA prepared from uninfected or infected cells as indicated. Next, we examined the ability of Vpr to induce the activity of the HIF-1α promoter. As shown in Figure 4C, infection of microglial cells with Ad-Vpr at an MOI of 5 stimulated the HIF-1α promoter (lane 2). In light of earlier studies suggesting the cooperativity of Vpr with Sp1 and NF-κB (26, 27), and the presence of both Sp1 and NF-κB binding sites within the HIF-1α promoter, we investigated the importance of Sp1 and the p65 subunit of NF-κB in Vpr activation of the HIF-1α promoter. We used siRNA (50nM) to suppress expression of Sp1 and NF-κB-p65 in the transfected cells and found that the presence of Sp1, and to a lesser degree, p65 is essential for Vpr activation of the HIF-1α promoter (compare lane 2 with lanes 4 and 6). As a control, non-targeting siRNA (50 nM) was used (ns-siRNA). Addition of ns-siRNA did not affect the ability of Vpr to activate the HIF-1α promoter (compare lanes 2 and 8). Finally, we determined whether addition of HIF-1α affects the ability of Vpr to regulate the HIF-1α promoter.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
6
Interestingly, the activation of the HIF-1α promoter by HIF-1α (lane 9) is increased by the presence of Vpr (lane 10). These results point to a functional association between Vpr and HIF-1α and that Vpr activates HIF-1α at transcriptional and translational levels. Panels 4D and F illustrate results from Western blots showing suppression of Sp1 and p65 in cells treated with siRNA for targeting Sp1 and p65, respectively. Non-targeting siRNA was also used as a control. Differences in the levels of expression of Sp1 and p65 proteins were further evaluated by densitometric analysis (Panels E and G).
Vpr induces mitochondrial ROS production in microglia. As the generation of excess mitochondrial ROS induces a number of redox reactions that can induce HIF-1α expression, in the next series of studies, we investigated the effect of Vpr on mitochondrial ROS (41). Microglia were infected with Ad-Null and Ad-Vpr virus at an MOI of 5 for 48h as described in Materials and Method and then treated with the redox-sensitive
probe Redox Sensor Red CC-1 and the mitochondria-specific dye MitoTracker green FM. As shown in Figure 5A, only Vpr-infected cells exhibit bright yellow/orange fluorescence in mitochondria due to the co-localization of oxidized Red CC-1 and Mito Tracker green, whereas oxidation of Red CC-1 in cytoplasm shows red/orange fluorescence, indicative of increased ROS production in the mitochondria and cytosolic compartments (compare Vpr panels to mock or adeno-null panels). These results point to the ability of Vpr to induce mitochondrial ROS in microglia. Figure 5B shows quantification of the ROS levels. In comparison with mock- or Ad-null-infected cells, ROS production was approximately 15-fold higher in the cells infected with Ad-Vpr. Infection of the cells with Ad-null had no effect on ROS production.
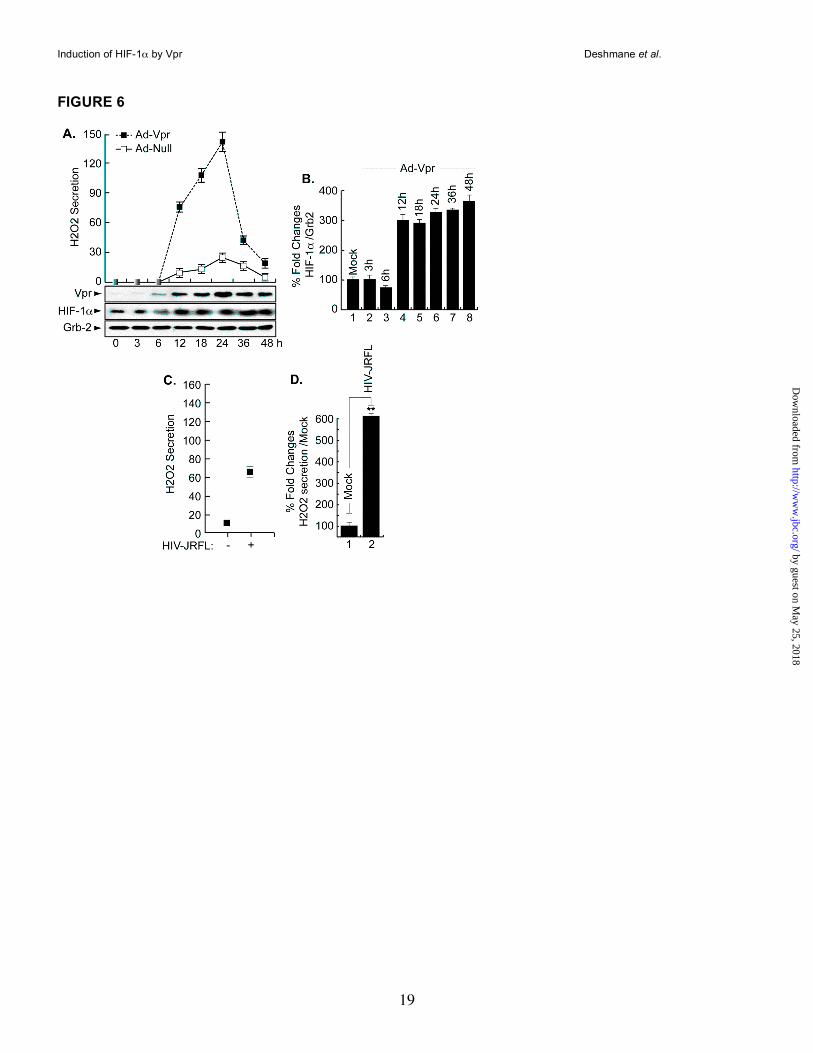
Vpr induces H2O2 production in microglia. As a component of ROS, H2O2 has been shown to cause activation of the HIV-1 LTR in patients with latent HIV-1 (13, 42). Further, exogenous H2O2 was shown to prevent HIF-1α accumulation in the liver cell line Hep3B (43). Therefore, we investigated the effect of Vpr on endogenous H2O2 secretion and whether this secretion will affect endogenous levels of HIF-1α. To this end, microglial cells were infected in duplicate with Ad-Null and Ad-Vpr virus at an MOI of 5 and at various times post-infection (3 - 48h). The level of
H2O2 was determined from one set of cells using the Quantichrom Peroxide assay kit (DIOX-250, Bioassay systems), which is designed to measure H2O2. The method utilizes a chromogenic Fe3+ xynol orange reaction, in which a purple complex formed when Fe2+ provided in the reagent is oxidized to Fe3+ by peroxides present in the sample. The intensity of the color (540-610 nm) measures H2O2. A standard curve was generated according to the manufacturer’s protocol using known amounts of H2O2. As shown in Figure 6A, in the presence of Vpr, the level of H2O2 production was increased at 12 hours post-infection and reached a maximum level at 24 hours, then decreased at later time points (36 and 48 h). Note that we observed an unexplained “boost” in H2O2 secretion/production at 48 h (data not shown [Studies in progress]). Infection of the cells with Ad-Null did not lead to any increase in the production of H2O2 (Panel A). The other set of cells was used for Western analysis. Briefly, the cell lysates were prepared from these cells and subjected to Western blot analysis using anti-Vpr, anti-HIF-1α and anti-Grb2 antibodies. As shown in Figure 6A, Vpr was detected as early as 6 h post-infection (Vpr Panel). It peaks at 24 h, relatively decreases as of 36 h and remains constant at 48 h. Endogenous levels of HIF-1α were induced as early as 12 h post-infection (HIF-1α Panel) and remained invariable up until 48 h. Differences in the levels of expression of HIF-1α protein were further evaluated by densitometric analysis (Panel B). These data led us to conclude that secretion of H2O2 did promote HIF-1α accumulation in microglial cells and not as previously reported in Hep3B (43).
To investigate the ability of HIV-1 infection to induce H2O2 secretion, microglial cells (1 x106) were infected with JR-FL strain of HIV-1 (50ng of p24-containing virus stock). Our results showed an increase in the level of H2O2 upon HIV-1 infection at 24h post-infection (Panel C), suggesting that expression of Vpr induces H2O2 production during the course of HIV-1 infection of microglial cells.
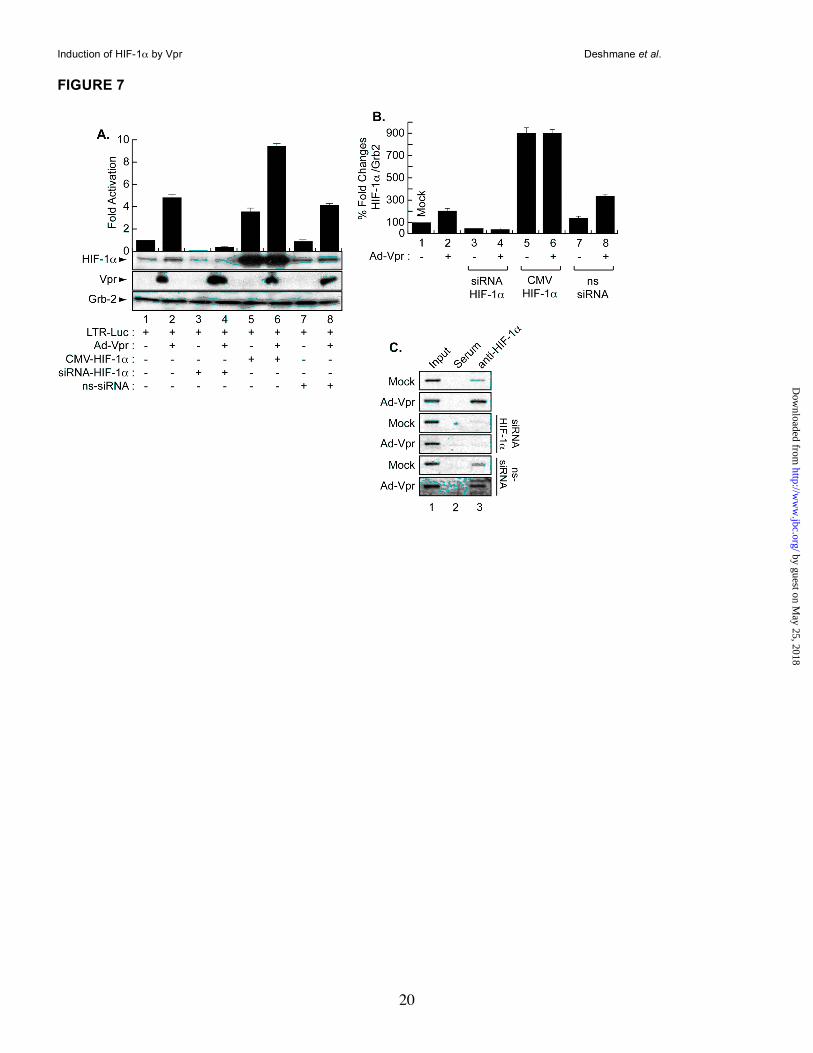
Regulation of the HIV-1 promoter by HIF-1α and/or Vpr in microglia. To examine the impact of HIF-1α induction on HIV-1, microglial cells were transfected in triplicate with HIV-1-Luc alone or with 1.0µg of CMV-HIF-1α and Ad-Vpr. After 48h, the cells were processed for luciferase assay while one set was analyzed by Western blot. As
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
7
shown in Figure 7A, transfection of HIF-1α (lane 5) or infection of the cells with Ad-Vpr (lane 2) activated the HIV-1 LTR promoter when compared to the LTR alone (lane 1). Similar to the results obtained with the HIF-1α promoter, activation of the LTR by HIF-1α was further increased in cells that were expressing Vpr (lane 6), suggesting cooperativity between HIF-1α and Vpr upon LTR transcription. To further demonstrate the specificity of this functional interaction, and to examine the involvement of HIF-1α in Vpr-mediated activation of the HIV-1 promoter, the LTR was transfected with 50 nM of siRNA-HIF-1α alone or in the presence of Ad-Vpr. Interestingly, inhibition of HIF-1α led to silencing of the LTR in the presence and absence of Vpr (compare lanes 1 and 2 to lanes 3 and 4). To further confirm the dependency of Vpr on the presence of HIF-1α in order to activate the HIV-1 promoter, the cells were transfected with non-targeting siRNA in the presence or absence of Vpr. As shown in Figure 7A, activation of the LTR by Vpr was not affected by the presence of ns-siRNA (compare lanes 1 and 2 to lanes 7 and 8). In parallel, protein extracts were prepared from one set of the cells and analyzed by Western blot using anti-HIF-1α, anti-Vpr and anti-Grb2 antibodies. As expected, infection of the cells with Vpr led to an increase in the levels of HIF-1α (lane 2). Vpr failed to induce endogenous levels of HIF-1α in the presence of siRNA-HIF-1α (lane 4) but not in the presence of ns-siRNA (lane 8). Vpr was only detected in lanes where it was introduced (Vpr Panel). Anti-Grb2 was used to control equal protein loading. Differences in the levels of expression of HIF-1α protein were further evaluated by densitometric analysis (Panel B). The functional interplay between HIF-1α and Vpr prompted us to examine whether there is a physical interaction between the two proteins. Microglia were infected with Ad-Vpr or Ad-Null at an MOI of 5 and lysates harvested after 48h. Three hundred micrograms of extracts were then subjected to immunoprecipitation/Western blot analysis using anti-Vpr or anti-HIF-1α antibodies. These experiments failed to show an interaction between these two proteins (data not shown).
Identification of hypoxia response element (HRE) within the HIV-LTR. Since HIF-1 activates the HIV-1 LTR, we next investigated that mechanism. First, we examined if HIF-1 can activate the LTR directly since the LTR contains a
predicted HRE located between -71 and -67 [RCGTG, where R could be an A or G] (16). Therefore, we performed chromatin immunoprecipitation assay (ChIP) to examine if HIF-1 can bind to this site. As shown in Figure 7C, microglial cells were transfected with 0.5 µg of the HIV-1 promoter alone or in the presence of 50nM of siRNA-HIF-1α, ns-siRNA and/or infected with Ad-Vpr (lanes 3) or Ad-Null (data not shown) at an MOI of 5. After 48h, cells were cross-linked with formaldehyde, harvested and ChIP was performed. The remainder of the procedure followed standard protocols for ChIP analysis as previously described (34). The resulting DNA was analyzed by PCR reactions with HIV-LTR primers. Anti-HIF-1α antibody (lanes 3) and rabbit anti-mouse IgG (lanes 2) were used in the procedure. Endogenous HIF-1 binds to HIV-LTR DNA. Western blot showed expression of Vpr and HIF-1α (data not shown). Interestingly, addition of Vpr increases the association of HIF-1 with the DNA (Vpr Panel, lane 3). As expected, addition of siRNA-HIF-1α but not ns-siRNA abolishes HIF-1 interaction with the LTR DNA in the absence or presence of Vpr. DISCUSSION
Recent advances in the field of oxidative stress (OS) have highlighted the importance of reduction-oxidation (redox)-sensitive transcription factors in cellular regulation. Redox-sensitive transcription factors are often associated with the development and progression of many human disease states. For example, OS occurs in brain tissues of patients undergoing HIV infection and is implicated in cell death of both astroglia and neurons and hence to play a role in the pathogenesis of neuroAIDS. For this reason, OS is a potential target for therapeutic interventions for possible clinical applications. HIF-1 is a crucial transcription factor that is a master regulatory element in sensing hypoxic conditions and integrating an adaptive response via gene expression of redox-sensitive enzymes and cofactors. The signal transduction components that link the availability of oxygen to the activation of HIF-1α are poorly defined. Thus, a better understanding of the processes that trigger and modulate free radical formation in the brain tissue of AIDS patients requires answers to the following questions: What are the mechanisms that trigger free radical overproduction in neuroAIDS? What is the impact of HIV-1 infection on HIF-1α
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
8
accumulation? Which viral protein(s) induces OS and accumulation of HIF-1α in CNS and what are the mechanisms used? Does induction of HIF-1α by HIV-1 protein(s) happen at the transcriptional or post-transcriptional level? What is the impact of OS induction and HIF-1α accumulation on HIV-1 gene expression and replication?
In order to address these questions, we examined the involvement of viral (Vpr), and cellular (HIF-1α) regulatory proteins in oxidative stress-induced neuropathogenesis of HIV-1 infection. Our data indicate that Vpr induced HIF-1α accumulation and ROS secretion in microglia. In turn, HIF-1 binds and upregulates the HIV-1 promoter.
Microglial cells were chosen because hypoxia has been shown to induce microglia activation, and animal studies have also shown that neuronal cell death is correlated with microglial activation in other situations, e.g., following cerebral ischemia (18). Thus, it is likely that toxic inflammatory mediators produced by activated microglia, e.g., HIV-1- or Ad-Vpr-infection, under hypoxic conditions may exacerbate neuronal injury following cerebral ischemia via induction of HIF-1α. Although the role of HIF-1 in microglia activation under hypoxia remains to be elucidated, it has been shown that exposure of primary rat microglial cultures, as well as an established microglial cell line, to hypoxia induced accumulation of HIF-1α. In turn, HIF-1 induces the expression of inducible NO synthase (iNOS) and ROS secretion, indicating that hypoxia could lead to the inflammatory activation of microglia (18). Thus, during cerebral ischemia, hypoxia may not only directly damage neurons, but may also promote neuronal injury indirectly via microglial activation.
Earlier work has shown that Vpr causes cell death through its association with the adenine nucleotide translocase protein (ANT), which promotes the permeability of the mitochondrial membrane, leading to the release of cytochrome c and activation of the caspase-3 pathway (44). These events are involved in inducing OS, suggesting the possibility of involvement of Vpr in the induction of OS. Recent studies demonstrated that the dose-dependent H2O2 stress response promotes increased survival for Schizosaccharomyces pombe cells expressing HIV-1 Vpr (11). These results are in conflict with our data. However, several studies demonstrated that ROS might play different roles (pro- or anti-
apoptosis) according to the cell types. Our data presented in Figure 5 indicate that
Vpr may induce NADPH oxidase ROS production but this possibility remains to be elucidated. It has been demonstrated that, in addition to its involvement in phagocytosis, NADPH oxidase-derived ROS is also implicated in facilitating cell-to-cell communication (45). Therefore, activation of NADPH oxidase pathway could point to a novel mechanism used by Vpr to facilitate virus trafficking. Thus, it is possible that Vpr helps in creating a suitable environment for HIV-1 to “travel” between the cells (studies in progress). However, our preliminary results demonstrated that Vpr failed to induce production of internal calcium from microglia (data not shown) leading to the activation of the IP3 pathway and preventing cell connection. Both pathways are under investigation.
In addition to the oxidative stress pathway, the TNFα and p38MAPK pathways have also been shown to induce HIF-1α (16, 23). We further confirmed this relation by examining the ability of Vpr to induce these pathways leading to HIF-1α upregulation. Interestingly, Vpr was capable of activating TNFα at the transcriptional and translational levels leading to HIF-1α induction. Further, we found that Vpr promotes phosphorylation of the p38MAPK protein leading to accumulation of HIF-1α (data not shown). Upregulation of HIF-1α by Vpr through p38MAPK pathway was further confirmed using inhibitor of p38MAPK (data not shown).
Activation and accumulation of HIF-1α by a viral protein is not without a precedent. In this regard, it has been shown that HIF-1α can be induced and accumulated in cells infected with HTLV-1 and HHV8 (46, 47). Further, HIF-1α was also shown to be suppressed in cells infected with a temperature-sensitive Moloney murine leukemia virus mutant- (MoMuLV-ts1) (48). In addition to its diverse regulation by different viruses, HIF-1α has also been shown to be regulated positively as well negatively in different cell lines. In this regard, it has been shown that H2O2 blocks accumulation of HIF-1α in the liver cell line, Hep3B (43). These latest results differ from our data presented in Figure 6. However, this difference could be due to variation between the cell types used as well as to the presence of HIV-1 Vpr protein, which may have the ability to overcome the negative effect of H2O2 (study in progress).
On the other hand, although, we found that
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
9
Vpr is involved in ROS secretion and HIF-1α accumulation, the involvement of other viral proteins such as gp120 cannot be ruled out. In this regard, gp120 has been shown to induce ROS secretion in monocytic cells that can produce neuronal damage (49). ROS produced in glial cells by gp120 exposure have been shown to cause neurodegeneration by inducing the synthesis of IL-1β (50). In addition to Vpr and gp120, HIV-1 Tat has been shown to induce ROS secretion by inhibiting expression of Mn-dependent superoxide dismutase (Mn-SOD) in HeLa and T-cells via a pathway that involved NF-κB (51). Tat also induces apoptosis of hippocampal neurons by a mechanism that involves oxidative stress (52). Oxidative damage has also been demonstrated to be induced by the injection of HIV-1 Tat protein in the rat striatum (9). Finally Tat protein was found to induce oxidative and inflammatory pathways in brain endothelium (53). Hence, these observations, in addition to our data, suggest a role for gp120 and Tat proteins, in addition to Vpr, on HIF-1α accumulation and ROS secretion.
Further, activation of the HIF-1α promoter by Vpr through Sp1 is not without a precedent. Several groups including ours demonstrated the ability of Vpr to activate the HIV-1 LTR and the p21WAF1 promoters through its association with Sp1 transcription factor (26, 54 and references within). In this regard, since no physical association between Vpr and HIF-1α was observed, it will be interesting to examine whether Sp1 mediates the interaction of Vpr with HIF-1α in these cells. Furthermore, Vpr has previously been shown to
cause apoptosis through suppression of NF-κB (27). Thus, association of HIF-1α with NF-κB within the HIV-1 LTR might facilitate the ability of Vpr to activate the HIV-1 promoter and to induce apoptosis. In order to examine this last possibility, we performed studies using microglial cells where we examined the status of cleaved caspase 3 in the presence and absence of Vpr and/or HIF-1α. Interestingly, over-expression of HIF-1α prevented Vpr from cleaving caspase 3 (data not shown). Therefore, we concluded that accumulation of HIF-1α in microglial cells might explain the lack of apoptosis observed in these cells in the presence of HIV-1.
These observations, in addition to the results presented in this manuscript, lead us to suggest that Vpr plays a key role in inducing OS in HIV-1 infected patients. In addition, accumulation of HIF-1α in microglia following Vpr addition might have a positive impact on cell survival and might explain the inability of Vpr to cause microglial cell death. In future studies, we plan to identify the mechanisms used by Vpr to induce OS leading to HIF-1α accumulation and to examine the impact of this induction/accumulation on Vpr functions and on HIV-1 gene expression and replication. Further, since Vpr can be released and taken up by neurons, and since neurons undergo degeneration, interplay between HIF-1α and Vpr should also be examined in neurons. Completion of these studies will help in developing successful therapeutic approaches against HIV infection and against neurodegeneration often observed in AIDS patients.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
10
ACKNOWLEDGEMENTS
The authors wish to thank past and present members of the Department of Neuroscience and Center for
Neurovirology for their support, and sharing of reagents and ideas. We also thank Dr. Martyn White for his
comments on the manuscript. This work was supported by grants awarded by NIH to BES and SA.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
11
REFERENCES 1. Blomgren, K., Leist, M., and Groc, L. (2007) Apoptosis 12, 993-1010
2. Sacktor, N., Haughey, N., Cutler, R., Tamara, A., Turchan, J., Pardo, C., Vargas, D., and Nath, A. (2004) J. Neuroimmunol. 157, 176-184
3. Hervouet, E., Simonnet, H., and Godinot, C. (2007) Biochimie 89, 1080-1088
4. Kimura, H., Sawada, T., Oshima, S., Kozawa, K., Ishioka, T., and Kato, M. (2005) Curr. Drug Targets Inflamm. Allergy 4, 489-495
5. Mollace, V., Nottet, H.S., Clayette, P., Turco, M.C., Muscoli, C., Salvemini, D., and Perno, C.F. (2001) Trends Neurosci. 24, 411-416
6. Price, T.O., Uras, F., Banks, W.A., and Ercal, N. (2006) Exp. Neurol. 201, 193-202
7. New, D.R., Maggirwar, S.B., Epstein, L.G., Dewhurst, S., and Gelbard, H.A. (1998) J. Biol. Chem. 273, 17852-17858
8. Shi, B., Raina, J., Lorenzo, A., Busciglio, J., and Gabuzda, D. (1998) J. Neurovirol. 4, 281-290
9. Aksenov, M.Y., Hasselrot, U., Bansal, A.K., Wu, G., Nath, A., Anderson, C., Mactutus, C.F., and Booze, R.M. (2001) Neurosci. Lett. 305, 5-8
10. Israel, N., and Gougerot-Pocidalo, M.A. (1997) Cell. Mol. Life Sci. 53, 864-870
11. Antal, J., and Pesti, M. (2006) Folia Microbiol. (Praha). 51, 406-412
12. Flores, S.C., Marecki, J.C., Harper, K.P., Bose, S.K., Nelson, S.K., and McCord, J.M. (1993) Proc. Natl. Acad. Sci. USA. 90, 7632-7636
13. Wu, W.S. (2006) Cancer Metastasis Rev. 25, 695-705
14. Chin, B.Y., Jiang, G., Wegiel, B., Wang, H.J., Macdonald, T., Zhang, X.C., Gallo, D., Cszimadia, E., Bach, F.H., Lee, P.J., and Otterbein, L.E. (2007) Proc. Natl. Acad. Sci. USA. 104, 5109-5114
15. Gordan, J.D., and Simon, M.C. (2007) Opin. Genet. Dev. 17, 71-77
16. Semenza, G.L. (2006) Expert Opin. Ther. Targets. 10, 267-280
17. Vaux, E.C., Metzen, E., Yeates, K.M., and Ratcliffe, P.J. (2001) Blood 98, 296-302
18. Makino, Y., Cao, R., Svensson, K., Bertilsson, G., Asman, M., Tanaka, H., Cao, Y., Berkenstam, A., and Poellinger, L. (2001) Nature 414, 550-554
19. Schweppe, R.E., Cheung, T.H., and Ahn, N.G. (2006) J. Biol. Chem. 281, 20993-21003
20. Simon, M.C. (2006) Adv. Exp. Med. Biol. 588, 165-170
21. Haddad, J.J. (2004) Int. Immunopharmacol. 4, 1249-1285
22. Zhou, J., Schmid. T., and Brune, B. (2003) Mol. Biol. Cell. 14, 2216-2225
23. Ke, Q., and Costa, M. (2006) Mol. Pharmacol. 70, 1469-1480
24. Amini, S., Khalili, K., and Sawaya, B.E. (2004) DNA Cell Biol. 23, 249-60
25. Nitahara-Kasahara, Y., Kamata, M., Yamamoto, T., Zhang, X., Miyamoto, Y., Muneta, K., Iijima, S., Yoneda, Y., Tsunetsugu-Yokota, Y., and Aida, Y. (2007) J. Virol. 81, 5284-5293
26. Sawaya, B.E., Khalili, K., Mercer, W.E., Denisova, L., and Amini, S. (1998) J. Biol. Chem. 273, 20052-20057
27. Ayyavoo, V., Mahboubi, A., Mahalingam, S., Ramalingam, R., Kudchodkar, S., Williams, W.V., Green, D.R., and Weiner, D.B. (1997) Nat. Med. 3, 1117-1123
28. Jacotot, E., Ravagnan, L., Loeffler, M., Ferri, K.F., Vieira, H.L., Zamzami, N., Costantini, P., Druillennec, S., Hoebeke, J., Briand, J.P., Irinopoulou, T., Daugas, E., Susin, S.A., Cointe, D., Xie, Z.H., Reed, J.C., Roques, B.P., and Kroemer, G. (2000) J. Exp. Med. 191, 33-46
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
12
29. Jones, G.J., Barsby, N.L., Cohen, E.A., Holden, J., Harris, K., Dickie, P., Jhamandas, J., and Power, C. (2007) J. Neurosci. 27, 3703-3711
30. Sawaya, B.E., Khalili, K., Gordon, J., Taube, R., and Amini, S. (2000) J. Biol. Chem. 275, 35209-35214
31. Bonello, S., Zähringer, C., BelAiba, R.S., Djordjevic, T., Hess, J., Michiels, C., Kietzmann, T., and Görlach, A. (2007) Arterioscler. Thromb. Vasc. Biol. 27, 755-761
32. Minet, E., Ernest, I., Michel, G., Roland, I., Remacle, J., Raes, M., and Michiels, C. (1999) Biochem. Biophys. Res. Commun. 261, 534-540
33. Janabi, N., Peudenier, S., Héron, B., Ng, K.H., and Tardieu, M. (1995) Neurosci. Lett. 195, 105-108
34. Mukerjee, R., Sawaya, B.E., Khalili, K., and Amini, S. (2007) J. Cell. Biochem. 100, 1210-1216
35. Eldeen, M.B., Deshmane, S.L., Simbiri, K., Khalili, K., Amini, S., and Sawaya, B.E. (2006) J. Neuroimmunol. 176, 174-180
36. Saunders, M., Eldeen, M.B., Valle, L.D., Reiss, K., Peruzzi, F., Mameli, G., Gelman, B.B., Khalili, K., Amini, S., and Sawaya, B.E. (2005) Apoptosis 10, 1419-1431
37. Chintapalli, J., Yang, S., Opawumi, D., Goyal, S.R., Shamsuddin, N., Malhotra, A., Reiss, K., and Meggs, L.G. (2007) Am. J. Physiol. Renal Physiol. 292, F523-530
38. Sweet, T., Khalili, K., Sawaya, B.E., and Amini, S. (2003) J. Cell. Biochem. 90, 884-891
39. Valcour, V., and Shiramizu, B. (2004) Mitochondrion 4, 119-129
40. Wallace, D.R., Dodson, S., Nath, A., and Booze, R.M. (2006) Synapse 59, 51-60
41. Le Bras, M., Clement, M.V., Pervaiz, S., and Brenner, C. (2005) Histol Histopathol. 20, 205-219
42. Kurata, S. (1996) J. Biol. Chem. 271, 21798-21802
43. Huang, L.E., Arany, Z., Livingston, D.M., and Bunn, H.F. (1996) J. Biol. Chem. 271, 32253-32259
44. Jacotot, E., Ferri, K.F., El Hamel, C., Brenner, C., Druillennec, S., Hoebeke, J., Rustin, P., Metivier, D., Lenoir, C., Geuskens, M., Vieira, H.L., Loeffler, M., Belzacq, A.S., Briand, J.P., Zamzami, N., Edelman, L., Xie, Z.H., Reed, J.C., Roques, B.P., and Kroemer, G. (2001) J. Exp. Med. 193, 509-519
45. Sun, G.Y., Horrocks, L.A., and Farooqui, A.A. (2007) J. Neurochem. 103, 1-16
46. Tomita, M., Semenza, G.L., Michiels, C., Matsuda, T., Uchihara, J.N., Okudaira, T., Tanaka, Y., Taira, N., Ohshiro, K., and Mori, N. (2007) Biochem. J. 406, 317-323.
47. Cai, Q., Murakami, M., Si, H., and Robertson, E.S. (2007) J. Virol. 81, 10413-10423.
48. Lungu, G.F., Stoica, G., and Wong, P.K. (2008) J. Neurovirol. 14, 239-251.
49. Foga, I.O., Nath, A., Hasinoff, B.B., and Geiger, J.D. (1997) J. AIDS Hum. Retrovirol. 16, 223-229
50. Viviani, B., Corsini, E., Binaglia, M., Galli, C.L., and Marinovich, M. (2001) Neuroscience 107, 51-58
51. Westendorp, M.O., Shatrov, V.A., Schulze-Osthoff, K., Frank, R., Kraft, M., Los, .M, Krammer, P.H., Droge, W., and Lehmann, V. (1995) EMBO J. 14, 546-54
52. Kruman, I.I., Nath, A., and Mattson, M.P. (1998) Exp. Neurol. 154, 276-288
53. Toborek M, Lee YW, Pu H, Malecki A, Flora G, Garrido R, Hennig B, Bauer HC, Nath A. (2003) J. Neurochem. 84, 169-179
54. Amini, S., Saunders, M., Kelley, K., Khalili, K., and Sawaya, B.E. (2004) J. Biol. Chem. 279, 46046-46056.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
13
FIGURE LEGENDS Figure 1. Induction of HIF-1α in AIDS patients with HIVE. A. White matter of a normal brain showing
small normal astrocytes. In contrast, the white matter of an HIVE case shows numerous reactive astrocytes
with enlarged nuclei and abundant stellar cytoplasm. HIF-1α was weakly detected in the cytoplasm of
normal astrocytes. In a case of HIVE, there is robust immunoreactivity for HIF-1α in the cytoplasm of
enlarged reactive astrocytes as well as in macrophages (perivascular cuffs) (x 400) and in microglial
nodules. B, D. Representative Western blots. HIV-1 infection enhances HIF-1α levels. Cell lysates were
prepared from uninfected (lane 1) and HIV-infected (lane 2) microglial cultures and 30 µg of cell lysate were
analyzed for HIF-1α using anti-HIF-1α, -Vpr, and -Grb2 antibodies (B). Induction of HIF-1α in HIV- but not in
HIVΔVpr-infected cells. Cell lysates were prepared from microglial cells, uninfected (lane 1), HIF-transfected
(lane 2), HIV-infected (lane 3) or HIVΔVpr-infected cells (lane 4). Thirty micrograms of cell lysate were used
(D). C, E. Histograms represent the variations in HIF-1α levels obtained by Western assays and measured
relatively to Grb2 using densitometric analysis (EZQuant-Gel software) (mean ± SE; n=3; one-way analysis
of variance and ANOVA test; ** compared to the Mock control group, p <0.05). The control was set
arbitrarily at 100%.
Figure 2. Induction of HIF-1α by Vpr is dose and time dependent. A. Representative Western blot. Cell
lysates were prepared from microglial cells, uninfected, or infected with Ad-Vpr-, Ad-null- or HIV-infected.
Thirty micrograms of cell lysate prepared from microglial cells were analyzed for HIF-1α and Vpr. Induction
of HIF-1α was measured relative to Grb2 and the values obtained with EZQuant-Gel software are presented
in panel B. The control was set arbitrarily at 100%, (mean ± SE; n=5). For the statistical analysis, the results
were analyzed using the Student’s t test statistical significance level, p < 0.05 is indicated by “**”, **
compared to the Mock control group (one-way analysis of variance and ANOVA test). C. Microglial cells
were mock-infected or infected with two concentrations of Ad-Vpr as well with Ad-null or HIV-1 as indicated.
Cell lysates were prepared and analyzed by Western blot as described in A. The arrows indicate the
position of each protein. Anti-Grb2 antibody was used to show equal protein loading.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
14
Figure 3. The HIV-1 protein, Vpr, enhances HIF-1α levels. A. Representative Western blot. Cell lysates
were prepared from uninfected (lane 1) or Ad-Null (lane 2), Ad-Tat (lane 4), Ad-Vpr (lane 5) -infected
microglial cultures, as well as from pcDNA3 (lane 3) or pcDNA3-HIF1α (lane 6) -transfected cells and
analyzed. Anti-Tat and Vpr antibodies were used as a control to show expression of both proteins. Anti-
Grb2 antibody was used to show equal protein loading. B. Histograms represent the values of HIF-1α
bands obtained by Western assays as measured relatively to Grb2 using densitometric analysis using
EZQuant-Gel software (mean ± SE; n=3). For the statistical analysis, the results were analyzed using the
Student’s t test statistical significance level, p < 0.05 is indicated by “**”, ** compared to the Mock control
group (one-way analysis of variance and ANOVA test).
Figure 4. Transcriptional regulation of HIF-1α by Vpr. A, B. Representative Northern blot. Microglia
were infected with Ad-null or Ad-Vpr at MOI of 5 or transfected with 0.5 µg of HIF-1α-luc alone or in
combination with 5 MOI of Ad-Vpr or 1 µg of HIF-1α expression plasmid, 50 nM of siRNA-Sp1 or p65 as
described. Cells infected with Ad-null or Ad-Vpr were analyzed using 20 µg of total RNA. Induction of HIF-
1α was measured relative to GAPDH with EZQuant-Gel software and the values are presented in panel B,
(mean ± SE; n=3; one-way analysis of variance and ANOVA test; ** compared to the Mock control group, p
<0.05). C. Transfected cells were subjected to luciferase assay 48h after being transfected with the HIF-1α
promoter construct. Data represent the mean value of five separate transfection experiments performed
with several DNA preps. For the statistical analysis, the results were analyzed using the Student’s t test
statistical significance level, p < 0.05 is indicated by “**”, ** compared to the Mock control group (one-way
analysis of variance and ANOVA test). D, F. Representative Western blots. Cells were transfected with 40
nM of siRNA-Sp1, siRNA-p65, or non-targeting siRNA3 (ns-siRNA) as indicated. Thirty micrograms of
extract were assayed with anti-Sp1, -p65, or anti-Grb2 antibodies. Arrows indicate the positions of each
protein. E, G. Histograms represent the induction of Sp1 and p65, respectively, relatively measured to Grb2
using densitometric analysis (EZQuant-Gel software) (mean ± SE; n=3; one-way analysis of variance and
ANOVA test; ** compared to the Mock control group, p <0.05). Note that in panel E, the 105 and 95 kDa
isoforms (bands) of Sp1 are shown in black and gray, respectively.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
15
Figure 5. Induction of ROS production by Vpr at 48 hr. A. Microglia were infected with Ad-null, Ad-Vpr or
mock-infected. Cells were loaded with the redox-sensitive dye Redox Sensor Red CC-1 (red panels) and
the mitochondrial-specific dye MitoTracker green FM (green panels). Induction of ROS production and its
localization in the mitochondria (yellow/orange and arrows) or in the cytoplasm (red/orange and arrows) is
also shown. B. Total ROS fluorescence was averaged from individual cells and expressed as a percentage
of Mito fluorescence. Measurements were made from 100 cells per field. The columns (means ± SE)
represent the average of five fields. Statistically significant differences were seen between experimental
values and controls: (p < 0.05, ** compared to the Mock control group, Student's t test). The experiment
was carried out 3 times, with similar results.
Figure 6. Induction of H2O2 in Vpr-infected cells in the absence of HIF-1α. Representative Western
blots. Assay for H2O2 showing production of H2O2 in Vpr-infected microglia (A) or in HIV-1-infected cells (C).
Expression of Vpr, HIF-1α and Grb2 was analyzed as indicated. Induction of HIF-1α was measured relative
to Grb2 and the values are presented in panel B. [Conversions: 1µM H2O2 equals 34 ng/ml]. The
experiments were carried out 3 times, with similar results. D. Data shown here are from a single,
representative experiment that was replicated three times and statistically significant. For the statistical
analysis, the results of the H2O2 secretion were analyzed using the Student’s t test statistical significance
level, p < 0.05 is indicated by “**” in panel D only, ** compared to the Mock control group (one-way analysis
of variance and ANOVA test). Even though not labeled, note that the results presented in panel B were also
statistically significant.
Figure 7. Transcription regulation of the HIV-1 promoter by Vpr and HIF-1α. A. Microglia were
transfected in triplicate with 0.5 µg of LTR-luc in the presence or absence of 1 µg of CMV-HIF-1α and/or
infected with Ad-Vpr at an MOI of 5 as indicated or Ad-Null at an MOI of 5 (data not shown). siRNA-HIF-1α
and ns-siRNA were also transfected alone or in the presence of Ad-Vpr (lanes 5, 6, 7 and 8). Data
represent the mean value of three separate transfection experiments. Luciferase activity was determined
after 48 h. Expression of Vpr, HIF-1α and Grb2 was analyzed by Western blot as indicated using one set of
transfected/infected cells (mean ± SE; n=5; one-way analysis of variance and ANOVA test). B. Data shown
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Induction of HIF-1α by Vpr Deshmane et al.
16
here are from a single, representative experiment that was replicated five times and statistically significant.
For the statistical analysis, the results were analyzed using the Student’s t test statistical significance level,
p < 0.05 is indicated by “**”, ** compared to the Mock control group (one-way analysis of variance and
ANOVA test). C. Interaction of HIF-1α with HIV-LTR DNA by ChIP in Vpr-infected microglia in the presence
and absence of siRNA-HIF-1α or ns-siRNA. Anti-HIF-1α (lanes 3), pre-immune serum (lanes 2), and no
antibody (lanes 1) were used.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Induction of HIF-1α by Vpr Deshmane et al.
17
FIGURE 1
FIGURE 2
FIGURE 3
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Induction of HIF-1α by Vpr Deshmane et al.
18
FIGURE 4
FIGURE 5
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Induction of HIF-1α by Vpr Deshmane et al.
19
FIGURE 6
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Induction of HIF-1α by Vpr Deshmane et al.
20
FIGURE 7
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SawayaThersa Sweet, Inna Rom, Kamel Khalili, Jay Rappaport, Shohreh Amini and Bassel E.
Satish L. Deshmane, Ruma Mukerjee, Shongshan Fan, Luis Del Valle, Carine Michiels,inducible factor 1 alpha expression
Activation of the oxidative stress pathway by HIV-1 Vpr leads to induction of hypoxia
published online February 9, 2009J. Biol. Chem.
10.1074/jbc.M809266200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from


![Calcareous sponge genomes reveal complex -carbonic … · 2017. 8. 29. · or characterize CA-proteins from the calcareous sponge S. ciliatum have not been successful [22]. Only recently,](https://static.fdocument.org/doc/165x107/60d35117c3bc180d086fdbcc/calcareous-sponge-genomes-reveal-complex-carbonic-2017-8-29-or-characterize.jpg)











![contrast - Southeast Asian Linguistics Societyjseals.org/seals23/cooper2013case.pdf · Modern Burmese is said to have a dental fricative [θ] Acoustic studies reveal it to be a dental](https://static.fdocument.org/doc/165x107/5e0840be171fc366cc12d0fd/contrast-southeast-asian-linguistics-modern-burmese-is-said-to-have-a-dental-fricative.jpg)


