
Structures of human primase reveal design of nucleotide ...
6
Structures of human primase reveal design of nucleotide elongation site and mode of Pol α tethering Mairi Louise Kilkenny, Michael Anthony Longo, Rajika L. Perera, and Luca Pellegrini 1 Department of Biochemistry, University of Cambridge, Cambridge CB2 1GA, United Kingdom Edited by Mike E. O’Donnell, The Rockefeller University, Howard Hughes Medical Institute, New York, NY, and approved August 21, 2013 (received for review June 12, 2013) Initiation of DNA synthesis in genomic duplication depends on primase, the DNA-dependent RNA polymerase that synthesizes de novo the oligonucleotides that prime DNA replication. Due to the discontinuous nature of DNA replication, primase activity on the lagging strand is required throughout the replication process. In eukaryotic cells, the presence of primase at the replication fork is secured by its physical association with DNA polymerase α (Pol α), which extends the RNA primer with deoxynucleotides. Our knowl- edge of the mechanism that primes DNA synthesis is very limited, as structural information for the eukaryotic enzyme has proved difficult to obtain. Here, we describe the crystal structure of hu- man primase in heterodimeric form consisting of full-length cata- lytic subunit and a C-terminally truncated large subunit. We exploit the crystallographic model to define the architecture of its nucleotide elongation site and to show that the small subunit integrates primer initiation and elongation within the same set of functional residues. Furthermore, we define in atomic detail the mode of association of primase to Pol α, the critical interaction that keeps primase tethered to the eukaryotic replisome. I t is of paramount importance to cellular life that the genetic blueprint encoded in its DNA is duplicated once, entirely and with utmost accuracy before each round of cell division. DNA duplication is performed by the replisome, a large and dynamic protein assembly that integrates and coordinates the enzymatic activities that are necessary to the task (1). Primase is a DNA- dependent RNA polymerase with an essential role in DNA replication (2, 3): after unwinding of the parental DNA at an origin site, primase assembles on each template strand a short RNA primer that is extended with deoxynucleotides by the replicative DNA polymerase. Given the discontinuous nature of DNA synthesis on the lagging strand, primase is required throughout replication to prime the synthesis of each Okazaki fragment. Despite the critical role of primase in DNA replica- tion, the structural basis for its function is still poorly understood. The requirement for a specialized polymerase that is able to prime DNA synthesis during genomic duplication is universal in all kingdoms of life. The evolutionarily related eukaryotic and archaeal primases are heterodimeric enzymes comprising a small, catalytic subunit (PriS) and a large, regulatory subunit (PriL) (4). Although the primary active site for RNA primer synthesis resides in the PriS subunit, PriL is important for regulation of primase activity and primer transfer to Pol α (5–8). Further in- sight into PriL function was provided by the observation that a conserved C-terminal Fe-S domain of PriL, PriL-CTD, is es- sential for initiation of primer synthesis, but dispensable for its subsequent elongation (9–13). Available structural evidence for the catalytic subunit of the eukaryotic primase is limited to eukaryotic-type archaeal PriS, in isolation and bound to PriL (14–16). The structural data dem- onstrate that the RNA priming activity of primase is encoded within a unique protein fold that is unrelated to that of other DNA or RNA polymerases. The polymerase fold of archaeal/ eukaryotic PriS has also been identified in an archaeal replicase with combined primase and polymerase activity (17), a bacterial plasmid primase (18), and the polymerase component of bacte- rial DNA repair ligase D (19). Conservation of aspartic residues in the PriS sequence and site-directed mutagenesis studies in- dicate that primase uses metal ion-dependent catalysis for nu- cleotide polymerization (20). However, the enzymatic requirements of the initiation step, when primase synthesizes the first dinucleotide of the RNA primer, likely require a larger set of residues, whose identity and role in catalysis remain incompletely understood. At the replication fork, primase is present in a constitutive complex with DNA polymerase α (Pol α), which extends the RNA primer with deoxynucleotides and makes the resulting RNA–DNA primer available to the leading- and lagging-strand polymerases, Pols e and δ, for processive elongation (21). On its turn, the Pol α/primase complex is anchored to the CMG heli- case by the conserved Ctf4 replication factor, which acts as a bridge between the GINS component of the helicase and the catalytic subunit of Pol α (22, 23). Thus, a chain of specific protein–protein interfaces links unwinding of the parental strands to priming of nucleic acid synthesis on template DNA. Our knowledge of the structure of the Pol α/primase complex and of the mechanism of RNA–DNA primer synthesis remains largely incomplete. Electron microscopy studies of a recombi- nant, unliganded version of the yeast Pol α/primase complex have shown that polymerase and primase reside in separate lobes of a flexible dumbbell-shaped particle (24). In agreement with the EM analysis, biochemical evidence shows that the interaction between primase and Pol α is mediated by the conserved C-terminal region of the catalytic subunit of Pol α (23, 24). Furthermore, a conserved motif at the C terminus of the catalytic subunit of Pol α is necessary and sufficient for association with Significance DNA synthesis during duplication of the genome depends on primase, the DNA-dependent RNA polymerase that initiates nucleotide polymerization by assembling short RNA primers on the unwound template DNA. The mechanism of DNA priming remains poorly understood, principally because of lack of high- resolution structural information. Our study begins to fill the gap in our knowledge by reporting crystal structures of human primase in unliganded form and bound to UTP. We exploit the atomic model of primase to map amino acids that are impor- tant for the catalytic function of the enzyme. In addition, we describe crystallographically the interaction between primase and DNA polymerase α, which keeps primase tethered to the replication fork. Author contributions: M.L.K. and L.P. designed research; M.L.K. performed research; M.A.L. contributed new reagents/analytic tools; M.L.K., R.L.P., and L.P. analyzed data; and M.L.K. and L.P. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Freely available online through the PNAS open access option. Data deposition: The atomic coordinates and structure factors have been deposited in the Protein Data Bank in Europe, www.ebi.ac.uk/pdbe [PDB ID codes 4BPU (primase), 4BPW (primase with bound UTP), and 4BPX (Pol α/primase complex)]. 1 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1311185110/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1311185110 PNAS | October 1, 2013 | vol. 110 | no. 40 | 15961–15966 BIOCHEMISTRY Downloaded by guest on December 18, 2021
Transcript of Structures of human primase reveal design of nucleotide ...

Structures of human primase reveal design ofnucleotide elongation site and mode of Pol α tetheringMairi Louise Kilkenny, Michael Anthony Longo, Rajika L. Perera, and Luca Pellegrini1
Department of Biochemistry, University of Cambridge, Cambridge CB2 1GA, United Kingdom
Edited by Mike E. O’Donnell, The Rockefeller University, Howard Hughes Medical Institute, New York, NY, and approved August 21, 2013 (received for reviewJune 12, 2013)
Initiation of DNA synthesis in genomic duplication depends onprimase, the DNA-dependent RNA polymerase that synthesizes denovo the oligonucleotides that prime DNA replication. Due to thediscontinuous nature of DNA replication, primase activity on thelagging strand is required throughout the replication process. Ineukaryotic cells, the presence of primase at the replication fork issecured by its physical association with DNA polymerase α (Pol α),which extends the RNA primer with deoxynucleotides. Our knowl-edge of the mechanism that primes DNA synthesis is very limited,as structural information for the eukaryotic enzyme has proveddifficult to obtain. Here, we describe the crystal structure of hu-man primase in heterodimeric form consisting of full-length cata-lytic subunit and a C-terminally truncated large subunit. Weexploit the crystallographic model to define the architecture ofits nucleotide elongation site and to show that the small subunitintegrates primer initiation and elongation within the same set offunctional residues. Furthermore, we define in atomic detail themode of association of primase to Pol α, the critical interactionthat keeps primase tethered to the eukaryotic replisome.
It is of paramount importance to cellular life that the geneticblueprint encoded in its DNA is duplicated once, entirely and
with utmost accuracy before each round of cell division. DNAduplication is performed by the replisome, a large and dynamicprotein assembly that integrates and coordinates the enzymaticactivities that are necessary to the task (1). Primase is a DNA-dependent RNA polymerase with an essential role in DNAreplication (2, 3): after unwinding of the parental DNA at anorigin site, primase assembles on each template strand a shortRNA primer that is extended with deoxynucleotides by thereplicative DNA polymerase. Given the discontinuous natureof DNA synthesis on the lagging strand, primase is requiredthroughout replication to prime the synthesis of each Okazakifragment. Despite the critical role of primase in DNA replica-tion, the structural basis for its function is still poorly understood.The requirement for a specialized polymerase that is able to
prime DNA synthesis during genomic duplication is universal inall kingdoms of life. The evolutionarily related eukaryotic andarchaeal primases are heterodimeric enzymes comprising a small,catalytic subunit (PriS) and a large, regulatory subunit (PriL) (4).Although the primary active site for RNA primer synthesisresides in the PriS subunit, PriL is important for regulation ofprimase activity and primer transfer to Pol α (5–8). Further in-sight into PriL function was provided by the observation thata conserved C-terminal Fe-S domain of PriL, PriL-CTD, is es-sential for initiation of primer synthesis, but dispensable for itssubsequent elongation (9–13).Available structural evidence for the catalytic subunit of the
eukaryotic primase is limited to eukaryotic-type archaeal PriS, inisolation and bound to PriL (14–16). The structural data dem-onstrate that the RNA priming activity of primase is encodedwithin a unique protein fold that is unrelated to that of otherDNA or RNA polymerases. The polymerase fold of archaeal/eukaryotic PriS has also been identified in an archaeal replicasewith combined primase and polymerase activity (17), a bacterialplasmid primase (18), and the polymerase component of bacte-rial DNA repair ligase D (19). Conservation of aspartic residues
in the PriS sequence and site-directed mutagenesis studies in-dicate that primase uses metal ion-dependent catalysis for nu-cleotide polymerization (20). However, the enzymatic requirementsof the initiation step, when primase synthesizes the first dinucleotideof the RNA primer, likely require a larger set of residues, whoseidentity and role in catalysis remain incompletely understood.At the replication fork, primase is present in a constitutive
complex with DNA polymerase α (Pol α), which extends theRNA primer with deoxynucleotides and makes the resultingRNA–DNA primer available to the leading- and lagging-strandpolymerases, Pols e and δ, for processive elongation (21). On itsturn, the Pol α/primase complex is anchored to the CMG heli-case by the conserved Ctf4 replication factor, which acts as abridge between the GINS component of the helicase and thecatalytic subunit of Pol α (22, 23). Thus, a chain of specificprotein–protein interfaces links unwinding of the parental strandsto priming of nucleic acid synthesis on template DNA.Our knowledge of the structure of the Pol α/primase complex
and of the mechanism of RNA–DNA primer synthesis remainslargely incomplete. Electron microscopy studies of a recombi-nant, unliganded version of the yeast Pol α/primase complexhave shown that polymerase and primase reside in separate lobesof a flexible dumbbell-shaped particle (24). In agreement withthe EM analysis, biochemical evidence shows that the interactionbetween primase and Pol α is mediated by the conservedC-terminal region of the catalytic subunit of Pol α (23, 24).Furthermore, a conserved motif at the C terminus of the catalyticsubunit of Pol α is necessary and sufficient for association with
Significance
DNA synthesis during duplication of the genome depends onprimase, the DNA-dependent RNA polymerase that initiatesnucleotide polymerization by assembling short RNA primers onthe unwound template DNA. The mechanism of DNA primingremains poorly understood, principally because of lack of high-resolution structural information. Our study begins to fill thegap in our knowledge by reporting crystal structures of humanprimase in unliganded form and bound to UTP. We exploit theatomic model of primase to map amino acids that are impor-tant for the catalytic function of the enzyme. In addition, wedescribe crystallographically the interaction between primaseand DNA polymerase α, which keeps primase tethered to thereplication fork.
Author contributions: M.L.K. and L.P. designed research; M.L.K. performed research; M.A.L.contributed new reagents/analytic tools; M.L.K., R.L.P., and L.P. analyzed data; and M.L.K.and L.P. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.
Data deposition: The atomic coordinates and structure factors have been deposited in theProtein Data Bank in Europe, www.ebi.ac.uk/pdbe [PDB ID codes 4BPU (primase), 4BPW(primase with bound UTP), and 4BPX (Pol α/primase complex)].1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1311185110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1311185110 PNAS | October 1, 2013 | vol. 110 | no. 40 | 15961–15966
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1

primase in vitro and for tethering primase to the replisome inyeast cells (25).A thorough understanding of the mechanism of DNA priming
in eukaryotic replication relies critically on high-resolutionstructural information about primase and its interaction with Polα. Such information is currently lacking due to a scarcity ofcrystallographic data. Here, we report the crystal structure ofhuman primase in heterodimeric form, consisting of PriS anda truncated version of PriL lacking the C-terminal Fe-S domain.We take advantage of the crystallographic model of primase todefine the architecture of its nucleotide elongation site, identi-fying residues that are important for primer initiation and ex-tension. In addition, we describe the cocrystal structure ofhuman primase in complex with the primase-binding motif of Polα and determine critical primase residues at the Pol α/primaseinterface that are responsible for tethering the enzyme to thereplication fork.
ResultsThree-Dimensional Architecture of Human Primase. To induce crys-tallization of human primase, we engineered a recombinant ver-sion of the enzyme that contained full-length PriS and a shorterversion of PriL (amino acids 1–253), lacking the C-terminal Fe-Sdomain. This recombinant enzyme, henceforth referred to simplyas primase, retains the ability to bind Pol α (25). The primasecrystallized in monoclinic space group P21, with two copies ofprimase in the asymmetric unit; the structure was solved bymultiple isomorphous replacement and refined to a resolution of2.7 Å (Table S1 and Fig. S1).The overall architecture of human primase, illustrating the
fold of the PriS and PriL subunits and their mode of interaction,is shown in Fig. 1. PriS adopts the expected “prim” fold char-acteristic of the prim-pol superfamily (26), fused to a smaller all-helical domain, which confers to PriS a rather flat, slab-like ap-pearance. The truncated PriL polypeptide partitions in a larger,globular, α-helical domain and a smaller, mixed α/β domain thatmediates the interaction with PriS. PriL docks on one side ofPriS, leading to a rather open and extended protein architecture,as already observed in the heterodimeric structure of the ar-chaeal primase (16).
PriS Fold and Active Site. The shape of the mixed α/β prim fold ofPriS is reminiscent of a Pac-Man character, where two three-strand β-sheets line the inside of the lower (strand β1 to β3) andupper (strands β4 to β6) jaws, each surrounded by α-helices onthe outside (Fig. 2). Human PriS can be superimposed on ar-chaeal PriS structures [Protein Data Bank (PDB) ID codes 1G71and 1ZT2], with overall rmsd of 2.5 Å for ∼180 aa (Fig. S2) (27).The presence of an invariant cluster of three aspartic acid
residues: D109, D111 on strand β4 and D306 immediately beforehelix α7, confirms that human primase relies on the two-metalions mode of nucleotide polymerization, as previously observedby site-directed mutagenesis of the mouse primase (20) (Fig.S3A). Equivalent sets of catalytic acidic residues have beenidentified in the PriS of archaeal primases (14–16), in the pri-mase–polymerase domain of an archaeal plasmid replicase (17)and in the polymerase domain of bacterial LigD (19). In additionto the catalytic triad of acidic amino acids, a conserved histidineresidue, corresponding to H166 in human primase, was high-lighted as essential for catalysis in all studies cited above.Beyond the adoption of the two-metal ions mode of nucleotide
polymerization, little is known about the mechanism of dinucleotideformation and primer synthesis by primase. We exploited ourcrystallographic model of human primase to identify residueswith specific functional roles in RNA priming. We performedextensive alanine scanning of highly conserved solvent-exposedresidues on both upper and lower β-sheets on the concave grooveof PriS. The targeted amino acids included the catalytic triad ofacidic residues D109, D111, D306, and invariant amino acids intheir immediate surroundings, D114, D116, H166, H315, K318,and H324. We also included in the analysis the more distantly
located residues E44, Y54, R56, and K77 in strands β1 to β3(lower jaw) of PriS, due to their strong evolutionary conservation(Fig. S3A).Each single-point mutant was assayed for its ability to either
initiate synthesis or to extend a templated primer (see Methodsfor details of the assay). Reaction products were analyzed bydenaturing acrylamide gel electrophoresis (Fig. 3). As expected,alanine mutants of the acidic residues in the catalytic triad D109,D111, and D306, lost all ability to synthesize RNA primers. Incontrast, the alanine mutants of conserved D114 and D116showed only a moderate reduction in primer synthesis, whichcould be remedied by increasing primase concentration in theassay. In the structure, D114 and D116 are located in the shorthelix α4 overlooking the active site and play structural roles inmaintaining the local conformation of the polypeptide chain.In addition to the catalytic triad of aspartic acid residues, a
unique feature of eukaryotic primases is the presence of threeconserved histidine residues: H166 on β5, H315 and H324 on theC-terminal segment of PriS that runs along the bottom of thecleft formed by the lower and upper β-sheets. Alanine mutationof H166 abolishes primer synthesis in human primase, as re-ported previously for the equivalent residue in archaeal andprokaryotic prim fold polymerases (17, 19). Interestingly, theH315A mutant is almost as impaired, leading to loss of primersynthesis. Alanine mutation of H324 has a strong negative effecton primer synthesis, which can be partially rescued by increasingprimase concentration.To provide a structural basis for the result of the alanine
scanning experiment, we soaked uridine 5′-triphosphate (UTP)
Fig. 1. Structure of the human primase, comprising full-length PriS anda truncated version of PriL, lacking its C-terminal Fe-S domain. Front and topviews of the primase are shown. The primase is drawn as a ribbon, coloredorange for PriS and blue for PriL. The position of the zinc atom in PriS is alsoshown. The position of the active site in PriS is indicated by a black arrow.
15962 | www.pnas.org/cgi/doi/10.1073/pnas.1311185110 Kilkenny et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1

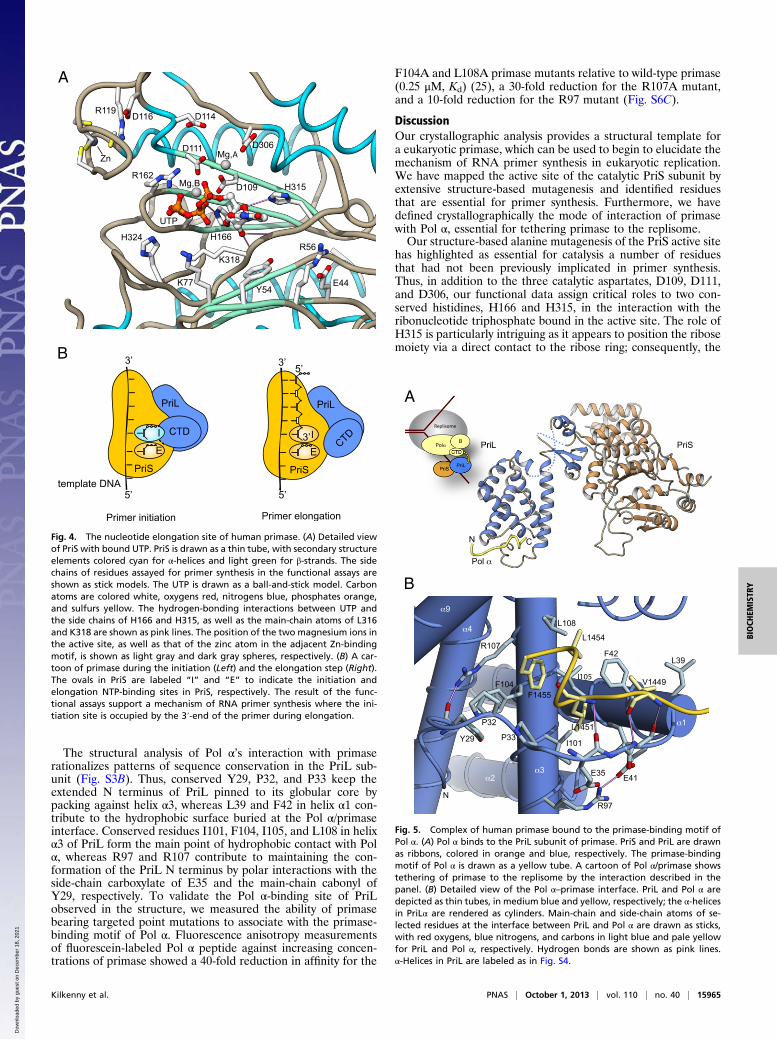
and MgCl2 in the crystals of human primase. A single-nucleotidetriphosphate (NTP) bound to each PriS chain in the asymmetricunit, suggesting that simultaneous binding of two nucleotides,necessary to synthesize the first dinucleotide, requires addition-ally the PriL Fe-S domain and/or the template DNA, which areabsent in our crystals. The NTP-binding site engaged by the UTPmoiety is likely to correspond to the so-called extension site ofthe primase, which is occupied by the NTP that will eventuallybecome the second nucleotide in the primer (28). The numberand location of the metal sites in the active site were confirmedby anomalous diffraction experiments of crystals soaked withnucleotide and MnCl2.In the structure, UTP binds in an extended conformation,
making extensive interactions with the upper β-sheet and theC-terminal segment of PriS that conjoins the lower and upperβ-sheets (Fig. 4A). The position of the triphosphate is de-termined by direct interactions with the 160-SGRRG-166 motiflinking strands β5 and β6 and by metal ion-mediated interactionswith D109 and D111 in strand β4. The conformation of the tri-phosphate is further consolidated by a direct hydrogen bond tothe β-phosphate by H166. The third catalytic aspartate D306 islikely to help coordinate a second magnesium ion, which acti-vates for nucleophilic attack the 3′-hydroxyl of the growing RNAprimer or of the first NTP bound at the initiation site.The ribose and base of the UTP pack against the 315-HLLK-
318 motif at the bottom of the active site groove, engaging inboth hydrophobic interactions with the side chains of leucines316 and 317 and in specific polar contacts. In particular, the sidechain of H315 may specify the position of the ribose by donatinga hydrogen bond to the oxygen of the ribose ring. Furthermore,the 2′-hydroxyl of the ribose is hydrogen bonded to the main-chain oxygen of L316 and the main-chain nitrogen of K318; thepresence of contacts involving the 2′-hydroxyl moiety might con-tribute to determining the preference of primase for nucleotidesover deoxynucleotides.Interactions between human primase and nucleotide are
completed by K318 and H324; K318 stretches its side chainunderneath the ribose and phosphate groups, thus assisting in
NTP binding by charge neutralization. The important role ofK318 in NTP binding is highlighted by the inability of the K318Amutant to perform RNA primer synthesis. The H324 side chainis oriented toward the UTP γ-phosphate, but beyond usefulrange for hydrogen bonding, which might be water mediated.The H324A mutant shows only partial impairment of primersynthesis, indicating a less critical role in NTP binding.Strong sequence conservation of solvent-exposed residues ex-
tends beyond the immediate surroundings of the NTP bindingsite. Alanine mutation of either K77 or Y54 has a clear negativeeffect on primer synthesis, which can be rescued by increasingprimase concentration in the assay; interestingly, however, theseprimase mutants remain defective in the ability to extend unit-length primers. Y54 occupies an analogous position to that ofF604 in the bacterial polymerase domain of LigD, which stacksagainst the guanine base of GTP (PDB ID code 2FAQ) (19), andmight therefore fulfill a similar functional role in human primase.Aligned with Y54 along the β1 strand, the side chain of R56 iskept in an upward thrust by an interstrand salt link with E44,seemingly poised for interaction. Strikingly, alanine mutation ofR56 abolishes RNA primer synthesis completely, pointing toa crucial functional role for this arginine residue. Alanine mu-tation of E44 is almost equally detrimental, although someresidual priming activity is detectable at higher primase con-centration. The mutant behavior is unlikely to be caused by lossof template DNA binding, which remains unaffected (Table S2).The essential roles of R56 and E44 uncovered by our experi-ments are presently unclear, but they might involve binding ofthe nucleotide at the initiation site.Interestingly, all of the mutants that showed reduced or ab-
rogated activity in the initiation assay demonstrated equally re-duced ability to extend an existing primer. Thus, it seems thathuman primase uses the same set of functional amino acids forsynthesis of the initial dinucleotide as well as for its successiveextension with ribonucleotides (Fig. 4B). To date, the only ex-ception to this rule is R306 in the Fe-S domain of PriL, which isnecessary for initiation of synthesis but dispensable for primerextension (9) (Fig. 3).
PriL Fold and Interface with PriS. The segment of large subunitpresent in the primase crystals folds in a two-domain structurecomprised of a larger, globular helical domain (PriLα) anda smaller mixed α/β domain (PriL–SBD) that binds to PriS (Fig.S4A). PriL–SBD docks onto the narrow side of the slab-like PriSshape, in such a way that the rest of PriL can reach over the sideof PriS that contains the active site (Fig. 1). Thus, PriL appearsto function as a mechanical arm, presumably to position itsC-terminal Fe-S domain, which has a crucial functional role ininitiation of synthesis, over the PriS active site. Comparison ofPriL position relative to PriS in the two primase heterodimers inthe asymmetric unit of the crystals reveals that, whereas PriL–SBD is docked firmly onto PriS, PriLα is tilted to a variabledegree relative to PriS (Fig. S4B); the apparent intersubunitflexibility of the PriS–PriL heterodimer might be a requiredstructural feature for efficient priming.The PriS–PriL interface buries about 1,600 Å2 of predom-
inantly hydrophobic surface, with contribution of polar contactson the outer edges of the interface. The side chains of PriSresidues F150, F152, V174, L177, V181, I185, and Y188 arearranged on the narrow side of the slab-like prim fold to forma contiguous hydrophobic surface, which is augmented by H209and P210 on the helical domain of PriS (Fig. S5A). InteractingPriL residues that become partially or completely buried at thehydrophobic PriS–PriL interface are F188, L192, F195, Y201,and L202 (Figs. S3B and S5B). PriS–PriL association relies onthe presence of two conserved glycine residues, G151 in PriS andG205 of PriL, which become closely juxtaposed at the PriS–PriLinterface. The interaction is fortified by prominent polar con-tacts across the interface, including hydrogen bonds betweenR198 of PriL and the main chain carbonyls of E187-Y188 inPriS, H209 in PriS and the main-chain carbonyl of V200 in PriL
Fig. 2. The prim fold of human primase. Only secondary structure elementsthat make up the prim fold are shown, as blue ribbons for α-helices and lightgreen arrows for β-strands. The small arrows indicate the points of insertionof the smaller, helical domain (omitted from the figure for clarity). Theposition of the active site in PriS is indicated by a black arrow.
Kilkenny et al. PNAS | October 1, 2013 | vol. 110 | no. 40 | 15963
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1

and main-chain to main-chain hydrogen bonding between car-bonyls of the E148-D149 in PriS and nitrogens in D204-G205of PriL.
Interaction with Pol α. Recruitment of primase to the replisomedepends critically on a conserved sequence motif at the extremeC terminus of Pol α (25). We decided to exploit the crystalstructure of human primase to investigate the structural basis forthe interaction with Pol α. Cocrystallization or soaking of a syn-thetic peptide corresponding to human Pol α sequence 1,445-GYSEVNLSKLFAGCAVKS-1,462 (C-end) in the crystals ofhuman primase did not provide a clear indication of the presenceof the bound peptide. We suspected that packing of primasemolecules in the crystal lattice might prevent partially or com-pletely the access of the Pol α peptide to its binding site. Wetherefore decided to attempt crystallization of a chimeric con-struct of human primase, where Pol α residues 1,445–1,462(C-end) had been fused with a long, flexible linker to the Nterminus of PriL. The Pol α/primase chimera crystallized in thesame lattice as the unliganded primase, but additional densitywas clearly visible in one primase molecule of the asymmetricunit, allowing us to trace amino acids 1,445–1,455 of the Polα sequence.
The PriL subunit is responsible for tethering primase to Pol α(Fig. 5A). The structure shows that Pol α binds PriL by dockingits C-terminal primase-binding motif onto a hydrophobic ledgeformed by the extended N terminus and helix α1 of PriL as theywrap around helix α3. Interestingly, in archaeal PriL structuresthe requirement to bury such an exposed hydrophobic surface issatisfied by a short helix within their N-terminal segment, whichfills the space occupied by the Pol α motif in the Pol α/primasecomplex (Fig. S6A). The stabilizing effect of the interaction withPol α is supported by thermal shifts experiments, which showincreased stability of primase in the presence of the primase-binding motif of Pol α (Fig. S6B).The primase-binding motif of Pol α adopts the predicted mode
of binding (25), as a short sequence motif that folds in a con-formation competent for binding in the presence of its in-teraction partner (Fig. 5B). In the Pol α/primase complex,residues 1,447-SEVN-1,450 of Pol α form an antiparallelβ-strand that pairs with residues 35-ENIS-38 of PriL, whereasresidues 1,451-LSKLF-1,455 fold in one turn of 310 helix thatpacks against the last two turns of PriL helix α3. Hot-spot resi-dues that were shown to be important for primase binding, suchas F1455 and L1451, become buried at the hydrophobic protein–peptide interface.
WTD109 R306 W
T D111 D114 D306 H166 WTD116 H315 K318 H324 W
T R56 E44 WT K77 Y54
A
B
WT W
T WT
D109R306R56E44
K77Y54
D111D114D116H166D306H315K318H324
Fig. 3. Functional analysis of active-site PriS resi-dues by structure-based alanine mutagenesis. A andB show the result of the primase initiation andelongation assays, respectively. The initiation assaymeasures the ability of primase to initiate primersynthesis in the presence of single-strand poly(dT)DNA and ATP. In the elongation assay, an oligo(A)primer is present in the reaction, in addition to poly(dT) DNA and ATP; the assay measures the ability ofprimase to initiate primer synthesis and/or extendan existing RNA primer. Briefly, reaction productswere separated by denaturing electrophoresis onpolyacrylamide gels and analyzed by phosphor-imaging. For each alanine mutant, two differentprimase concentrations were tested in the initiationassay. The R306A mutant refers to the residue in thePriL-CTD. See Methods for details.
15964 | www.pnas.org/cgi/doi/10.1073/pnas.1311185110 Kilkenny et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1
The structural analysis of Pol α’s interaction with primaserationalizes patterns of sequence conservation in the PriL sub-unit (Fig. S3B). Thus, conserved Y29, P32, and P33 keep theextended N terminus of PriL pinned to its globular core bypacking against helix α3, whereas L39 and F42 in helix α1 con-tribute to the hydrophobic surface buried at the Pol α/primaseinterface. Conserved residues I101, F104, I105, and L108 in helixα3 of PriL form the main point of hydrophobic contact with Polα, whereas R97 and R107 contribute to maintaining the con-formation of the PriL N terminus by polar interactions with theside-chain carboxylate of E35 and the main-chain cabonyl ofY29, respectively. To validate the Pol α-binding site of PriLobserved in the structure, we measured the ability of primasebearing targeted point mutations to associate with the primase-binding motif of Pol α. Fluorescence anisotropy measurementsof fluorescein-labeled Pol α peptide against increasing concen-trations of primase showed a 40-fold reduction in affinity for the
F104A and L108A primase mutants relative to wild-type primase(0.25 μM, Kd) (25), a 30-fold reduction for the R107A mutant,and a 10-fold reduction for the R97 mutant (Fig. S6C).
DiscussionOur crystallographic analysis provides a structural template fora eukaryotic primase, which can be used to begin to elucidate themechanism of RNA primer synthesis in eukaryotic replication.We have mapped the active site of the catalytic PriS subunit byextensive structure-based mutagenesis and identified residuesthat are essential for primer synthesis. Furthermore, we havedefined crystallographically the mode of interaction of primasewith Pol α, essential for tethering primase to the replisome.Our structure-based alanine mutagenesis of the PriS active site
has highlighted as essential for catalysis a number of residuesthat had not been previously implicated in primer synthesis.Thus, in addition to the three catalytic aspartates, D109, D111,and D306, our functional data assign critical roles to two con-served histidines, H166 and H315, in the interaction with theribonucleotide triphosphate bound in the active site. The role ofH315 is particularly intriguing as it appears to position the ribosemoiety via a direct contact to the ribose ring; consequently, the
A
B
Fig. 4. The nucleotide elongation site of human primase. (A) Detailed viewof PriS with bound UTP. PriS is drawn as a thin tube, with secondary structureelements colored cyan for α-helices and light green for β-strands. The sidechains of residues assayed for primer synthesis in the functional assays areshown as stick models. The UTP is drawn as a ball-and-stick model. Carbonatoms are colored white, oxygens red, nitrogens blue, phosphates orange,and sulfurs yellow. The hydrogen-bonding interactions between UTP andthe side chains of H166 and H315, as well as the main-chain atoms of L316and K318 are shown as pink lines. The position of the two magnesium ions inthe active site, as well as that of the zinc atom in the adjacent Zn-bindingmotif, is shown as light gray and dark gray spheres, respectively. (B) A car-toon of primase during the initiation (Left) and the elongation step (Right).The ovals in PriS are labeled “I” and “E” to indicate the initiation andelongation NTP-binding sites in PriS, respectively. The result of the func-tional assays support a mechanism of RNA primer synthesis where the ini-tiation site is occupied by the 3′-end of the primer during elongation.
A
B
Fig. 5. Complex of human primase bound to the primase-binding motif ofPol α. (A) Pol α binds to the PriL subunit of primase. PriS and PriL are drawnas ribbons, colored in orange and blue, respectively. The primase-bindingmotif of Pol α is drawn as a yellow tube. A cartoon of Pol α/primase showstethering of primase to the replisome by the interaction described in thepanel. (B) Detailed view of the Pol α–primase interface. PriL and Pol α aredepicted as thin tubes, in medium blue and yellow, respectively; the α-helicesin PriLα are rendered as cylinders. Main-chain and side-chain atoms of se-lected residues at the interface between PriL and Pol α are drawn as sticks,with red oxygens, blue nitrogens, and carbons in light blue and pale yellowfor PriL and Pol α, respectively. Hydrogen bonds are shown as pink lines.α-Helices in PriL are labeled as in Fig. S4.
Kilkenny et al. PNAS | October 1, 2013 | vol. 110 | no. 40 | 15965
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1

2′-hydroxyl of the ribose is within hydrogen bond distance withthe main-chain atoms of the primase. The additional contactsafforded to a ribose might contribute to the preference for NTPover dNTP by primase. Furthermore, our functional analysis hashighlighted the essential role of residues such as E44, Y54, andR56 that are located outside the NTP-binding site. Their func-tion in RNA primer synthesis is presently unknown, and theirrationalization will require further structural investigation of themechanism of priming.Current models of the mechanism of RNA primer synthesis
postulate that two distinct nucleotide-binding sites, the so-calledinitiation and elongation sites, need to be simultaneously occu-pied by NTPs, in preparation for the step of dinucleotide for-mation (2, 29). In eukaryotic primase, it has been proposed thatthe first nucleotide binds to the elongation site, followed bybinding to the initiation site of a second NTP, which will becomethe 5′-terminal nucleotide of the primer (28). The evidencepresented here shows that all PriS mutants that are important forprimer initiation by human primase are also required for primerextension. This observation indicates that PriS employs the sameset of catalytic residues for the addition of the first nucleotide tothe initiation nucleotide as for all subsequent nucleotide addi-tions. Thus, our experimental evidence provides support for theproposed model of priming, whereby the nucleotide at the ini-tiation site during dinucleotide synthesis would occupy the sameposition as the nucleotide at the 3′-end of a growing primer. Thecritical role of PriL in the initiation step is probably explained bythe additional interactions required to stabilize binding of theinitiation-site nucleotide during dinucleotide formation; indeed,R306 in PriL is the only residue identified to date that is essentialfor initiation but not for elongation (Fig. 3).The truncated form of heterodimeric primase analyzed in this
study crystallizes with two copies in the asymmetric unit, relatedby a pseudotwofold symmetry axis (Fig. S7). The two primasemolecules of the noncrystallographic dimer are held together byreciprocal contacts between the C-terminal segment of PriL anda protruding loop connecting strands β3 and β4 in PriS. Intriguingly,
in such a dimeric arrangement, the PriL-CTD of one primasewould be located opposite the PriS subunit of the second pri-mase and facing its active site, in a position that is compatiblewith priming in trans. We note that a model for in trans regula-tion of primer synthesis has been proposed for the prokaryoticprimase (30). Efforts to determine whether a dimer of eukaryoticprimase, such as observed in the crystals, is biochemicallycompetent for priming will require further experimental work.Priming of DNA synthesis in eukaryotic replication relies on
coordinated nucleotide polymerization by primase and Pol α:their constitutive association during DNA replication reflects therequirement for a tight functional integration of their catalyticactivities. Our structural analysis shows that this integration isphysically realized by means of a specific interaction between thehelical domain of the large subunit of primase and a short se-quence motif at the C terminus of Pol α. Tethering primase andPol α by a protein–peptide interaction provides the requiredspecificity and high affinity, while accommodating the large-scaleconformational changes that presumably take place within thecomplex during synthesis of the RNA-DNA primer. Given theconformationally dynamic nature of the eukaryotic replisome,we predict that further examples of association between repli-some components involving protein–peptide interactions willbe described.
MethodsDetails of all experimental procedures are described in SI Methods. All pri-mase constructs were produced in recombinant form in Escherichia coli. Thecrystal structure of human primase was solved by the multiple isomorphousreplacement method using heavy atom soaks.
ACKNOWLEDGMENTS. We thank Oscar Llorca and Karim Labib for helpfuldiscussions. We are grateful to Dimitri Chirgadze for assistance with initialcrystallographic experiments performed in the X-ray crystallographic facilityof the Department of Biochemistry, University of Cambridge. This work wasfunded by a Wellcome Trust senior research fellowship in basic biomedicalscience (to L.P.).
1. DePamphilis ML, Bell SD (2011) Genome Duplication (Garland Science, London).2. Frick DN, Richardson CC (2001) DNA primases. Annu Rev Biochem 70:39–80.3. Kuchta RD, Stengel G (2010) Mechanism and evolution of DNA primases. Biochim
Biophys Acta 1804(5):1180–1189.4. Iyer LM, Koonin EV, Leipe DD, Aravind L (2005) Origin and evolution of the archaeo-
eukaryotic primase superfamily and related palm-domain proteins: Structural insights
and new members. Nucleic Acids Res 33(12):3875–3896.5. Foiani M, Lindner AJ, Hartmann GR, Lucchini G, Plevani P (1989) Affinity labeling of
the active center and ribonucleoside triphosphate binding site of yeast DNA primase.
J Biol Chem 264(4):2189–2194.6. Copeland WC, Wang TS (1993) Enzymatic characterization of the individual mam-
malian primase subunits reveals a biphasic mechanism for initiation of DNA replica-
tion. J Biol Chem 268(35):26179–26189.7. Arezi B, Kirk BW, Copeland WC, Kuchta RD (1999) Interactions of DNA with human
DNA primase monitored with photoactivatable cross-linking agents: Implications for
the role of the p58 subunit. Biochemistry 38(39):12899–12907.8. Zerbe LK, Kuchta RD (2002) The p58 subunit of human DNA primase is important for
primer initiation, elongation, and counting. Biochemistry 41(15):4891–4900.9. Klinge S, Hirst J, Maman JD, Krude T, Pellegrini L (2007) An iron-sulfur domain of the
eukaryotic primase is essential for RNA primer synthesis. Nat Struct Mol Biol 14(9):
875–877.10. Weiner BE, et al. (2007) An iron-sulfur cluster in the C-terminal domain of the p58
subunit of human DNA primase. J Biol Chem 282(46):33444–33451.11. Sauguet L, Klinge S, Perera RL, Maman JD, Pellegrini L (2010) Shared active site ar-
chitecture between the large subunit of eukaryotic primase and DNA photolyase.
PLoS One 5(4):e10083.12. Vaithiyalingam S, Warren EM, Eichman BF, Chazin WJ (2010) Insights into eukaryotic
DNA priming from the structure and functional interactions of the 4Fe-4S cluster
domain of human DNA primase. Proc Natl Acad Sci USA 107(31):13684–13689.13. Agarkar VB, Babayeva ND, Pavlov YI, Tahirov TH (2011) Crystal structure of the C-terminal
domain of human DNA primase large subunit: Implications for the mechanism of the
primase-polymerase α switch. Cell Cycle 10(6):926–931.14. Augustin MA, Huber R, Kaiser JT (2001) Crystal structure of a DNA-dependent RNA
polymerase (DNA primase). Nat Struct Biol 8(1):57–61.
15. Ito N, Nureki O, Shirouzu M, Yokoyama S, Hanaoka F (2003) Crystal structure of thePyrococcus horikoshii DNA primase-UTP complex: Implications for the mechanism ofprimer synthesis. Genes Cells 8(12):913–923.
16. Lao-Sirieix SH, Nookala RK, Roversi P, Bell SD, Pellegrini L (2005) Structure of theheterodimeric core primase. Nat Struct Mol Biol 12(12):1137–1144.
17. Lipps G, Weinzierl AO, von Scheven G, Buchen C, Cramer P (2004) Structure of a bi-functional DNA primase-polymerase. Nat Struct Mol Biol 11(2):157–162.
18. Geibel S, Banchenko S, Engel M, Lanka E, Saenger W (2009) Structure and function ofprimase RepB′ encoded by broad-host-range plasmid RSF1010 that replicates exclu-sively in leading-strand mode. Proc Natl Acad Sci USA 106(19):7810–7815.
19. Zhu H, et al. (2006) Atomic structure and nonhomologous end-joining function of thepolymerase component of bacterial DNA ligase D. Proc Natl Acad Sci USA 103(6):1711–1716.
20. Copeland WC, Tan X (1995) Active site mapping of the catalytic mouse primasesubunit by alanine scanning mutagenesis. J Biol Chem 270(8):3905–3913.
21. Perera R, et al. (2013) Mechanism for priming DNA synthesis by yeast DNA polymerasealpha. Elife 2:e00482.
22. Gambus A, et al. (2009) A key role for Ctf4 in coupling the MCM2-7 helicase to DNApolymerase alpha within the eukaryotic replisome. EMBO J 28(19):2992–3004.
23. Tanaka H, et al. (2009) Ctf4 coordinates the progression of helicase and DNA poly-merase alpha. Genes Cells 14(7):807–820.
24. Núñez-Ramírez R, et al. (2011) Flexible tethering of primase and DNA Pol α in theeukaryotic primosome. Nucleic Acids Res 39(18):8187–8199.
25. Kilkenny ML, De Piccoli G, Perera RL, Labib K, Pellegrini L (2012) A conserved motif inthe C-terminal tail of DNA polymerase α tethers primase to the eukaryotic replisome.J Biol Chem 287(28):23740–23747.
26. Murzin AG, Brenner SE, Hubbard T, Chothia C (1995) SCOP: A structural classificationof proteins database for the investigation of sequences and structures. J Mol Biol247(4):536–540.
27. Irving JA, Whisstock JC, Lesk AM (2001) Protein structural alignments and functionalgenomics. Proteins 42(3):378–382.
28. Sheaff RJ, Kuchta RD (1993) Mechanism of calf thymus DNA primase: Slow initiation,rapid polymerization, and intelligent termination. Biochemistry 32(12):3027–3037.
29. Frick DN, Kumar S, Richardson CC (1999) Interaction of ribonucleoside triphosphateswith the gene 4 primase of bacteriophage T7. J Biol Chem 274(50):35899–35907.
30. Corn JE, Pease PJ, Hura GL, Berger JM (2005) Crosstalk between primase subunits canact to regulate primer synthesis in trans. Mol Cell 20(3):391–401.
15966 | www.pnas.org/cgi/doi/10.1073/pnas.1311185110 Kilkenny et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 18
, 202
1





![Nucleosid * DNA polymerase { ΙΙΙ, Ι } * Nuclease { endonuclease, exonuclease [ 5´,3´ exonuclease]} * DNA ligase * Primase.](https://static.fdocument.org/doc/165x107/56649cab5503460f9496ce53/nucleosid-dna-polymerase-nuclease-endonuclease-exonuclease.jpg)










![Calcareous sponge genomes reveal complex -carbonic … · 2017. 8. 29. · or characterize CA-proteins from the calcareous sponge S. ciliatum have not been successful [22]. Only recently,](https://static.fdocument.org/doc/165x107/60d35117c3bc180d086fdbcc/calcareous-sponge-genomes-reveal-complex-carbonic-2017-8-29-or-characterize.jpg)

