
Receptor And Nucleotide Exchange Independent … · proposed bound to a “hot spot” on βγ...
23
Receptor And Nucleotide Exchange Independent Mechanisms For Promoting G Protein Subunit Dissociation * . Mousumi Ghosh ‡ , Yuri K. Peterson§, Stephen M. Lanier§ and Alan V. Smrcka ‡** *This work was supported by a grant from the National Institutes of Health GM60286 ‡Department of Pharmacology and Physiology, University of Rochester School of Medicine and Dentistry, 601 Elmwood Avenue, Rochester, NY 14642. §Department of Pharmacology and Experimental Therapeutics, Louisiana State University Health Sciences Center, New Orleans, LA 70112. **To whom correspondence should be addressed Phone: 585-275-0892 FAX: 585-273-2652 email: [email protected] Running title: Peptide mediated activation of G protein 1 Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc. JBC Papers in Press. Published on July 24, 2003 as Manuscript C300271200 by guest on July 15, 2018 http://www.jbc.org/ Downloaded from
Transcript of Receptor And Nucleotide Exchange Independent … · proposed bound to a “hot spot” on βγ...

Receptor And Nucleotide Exchange Independent Mechanisms For Promoting G Protein
Subunit Dissociation*.
Mousumi Ghosh‡, Yuri K. Peterson§, Stephen M. Lanier§ and Alan V. Smrcka‡**
*This work was supported by a grant from the National Institutes of Health GM60286
‡Department of Pharmacology and Physiology, University of Rochester School of Medicine and
Dentistry, 601 Elmwood Avenue, Rochester, NY 14642. §Department of Pharmacology and Experimental
Therapeutics, Louisiana State University Health Sciences Center, New Orleans, LA 70112.
**To whom correspondence should be addressed
Phone: 585-275-0892
FAX: 585-273-2652
email: [email protected]
Running title: Peptide mediated activation of G protein
1
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on July 24, 2003 as Manuscript C300271200 by guest on July 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Summary
Mechanisms for heterotrimeric G protein activation that do not rely on G protein coupled receptor
activation are becoming increasingly apparent. We recently identified βγ subunit binding peptides that we
proposed bound to a “hot spot” on βγ subunits, stimulating G protein dissociation without stimulating
nucleotide exchange, and activating G protein signaling in intact cells. AGS3, a member of the activators
of G protein signaling family of proteins, also activates G protein signaling in a nucleotide exchange
independent manner and AGS3 homologues are involved in asymmetric cell division during development.
These G protein activators have been proposed either to compete for G protein subunit assembly or to
actively promote subunit dissociation. Distinguishing between these mechanisms is critical to
understanding the active roles of these G protein regulators. Here we demonstrate that a consensus G
protein regulatory (GPR) peptide from AGS3 and related proteins is sufficient to induce G protein subunit
dissociation and that both the GPR and “hot spot” binding peptides promote dissociation to extents
comparable to a known G protein activator, AMF. Peptides derived from adenylyl cyclase 2 and GRK2
prevented formation of the heterotrimeric complex but did not alter the rate of α subunit dissociation from
βγ subunits. These data indicate that these nucleotide exchange independent G protein activator peptides
do not simply compete for α interactions with βγ subunits, but actively promote subunit dissociation.
Thus we propose two novel mechanisms for nucleotide exchange independent activation of G protein
signaling, one that involves conformational changes in the α subunit and one that involves conformational
changes in the βγ subunits.
2
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Heterotrimeric G proteins activated by G protein coupled receptors mediate a wide variety of
cellular processes (1). The mechanisms by which G protein coupled receptors activate G proteins
have not been fully defined, but involve interactions between the activated receptor, G protein α
subunits and perhaps G protein βγ subunits. This interaction leads to the exchange of GDP for GTP
on the G protein α subunit leading to a conformational change resulting in dissociation of the βγ
subunits from the α subunits (1;2). The free αGTP and βγ subunits interact with downstream targets
and regulate their activities.
Multiple mechanisms for G protein activation that do not rely on G protein coupled receptors
or even nucleotide exchange are becoming increasingly apparent. We recently identified a receptor
independent mechanism for activation of G protein βγ subunit signaling by peptides derived from a
random peptide phage display screen that we have proposed bind to a “hot spot” on βγ subunits (3;4).
Protein interaction hot spots are regions on protein surfaces thought to have unique characteristics
suited to driving protein-protein interactions that are often selected for in random peptide screens
(5;6). We propose that the ability of these peptides to selectively target this “hot spot” is a unique
characteristic that is involved in its ability to stimulate subunit dissociation and activate G protein βγ
subunit signaling in cells. We have shown these peptides increase the rate of subunit dissociation from
a preformed α-βγ complex and we proposed that the peptides cause a conformational change in the βγ
subunits to promote α subunit dissociation. A caveat to this idea is that G protein α subunits have two
major contacts with βγ subunits and peptides may simply compete for one of these contacts that might
dissociate transiently.
Activators of G protein signaling (AGS 1-3 proteins) (7;8) were isolated from a genetic screen
in yeast to look for activation of the βγ mediated mating pathway. AGS3 binds to α subunits and
activates the signaling pathway without stimulating nucleotide exchange on the Gα subunit (7).
Sequences similar to a 25-30 amino acid repeat region in AGS3 were found in multiple other proteins
and suggested to be a signature G protein regulatory (GPR) motif (7). This motif was also
3
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

independently postulated to be a G protein binding motif and termed the GoLoco motif (9). Synthetic
peptides representing this motif inhibit GDP release from α subunits (10-12) and have been co-
crystallized with G protein α subunits (13). It is still not clear, however, if the GPR peptides simply
block heterotrimer formation or actively promote G protein dissociation as part of the mechanism for
AGS3 mediated activation of cell signaling.
To determine if the GPR peptides and βγ “hot spot”-binding peptides uniquely target critical
sites on G protein subunits to promote subunit dissociation or if they are simply steric competitors of α
βγ binding, we analyzed and compared the effects of multiple peptides believed to interact at the βγ -α
subunit interface for their ability to induce the α subunit dissociation from βγ subunits. We conclude
that both the “hot spot” and GPR motif consensus peptides have the unique ability to dissociate
heterotrimers by a mechanism that most likely involves conformational changes in the βγ and α
subunits respectively.
4
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Experimental Procedures.
Peptides. SIGK, QEHA and βARK-ct peptide (643-670) were synthesized by Alpha Diagnostics
International, purified by HPLC to greater than 90% purity, and their identity was confirmed by mass
spectrometry analysis. The SIGK peptide was derived from the previously described SIRK
(SIRKALNILGYPDYD) peptide using a doping mutagenesis and rescreening strategy (14). Since SIGK
had an apparently higher affinity for βγ than SIRK but whose properties were otherwise similar to SIRK,
this peptide was used throughout the studies described here. The sequences of these peptides were as
follows: SIGK: SIGKAFKILGYPDYD; QEHA: QEHAQEPERQYMHIGTMVEFAYALVGK; βARK-ct
peptide: WKKELRDAYREAQQLVQRVPKMKNKPRS. The GPR consensus motif peptide
TMGEEDFFDLLAKSQSKRMDDQRVDLAG was synthesized and purified by Biosynthesis, Inc.
(Lewisville, TX). All the peptides were dissolved in water.
Preparation of biotinylated β1γ2 subunits. The cDNA for rat β1 subunit was subcloned into a
baculovirus transfer vector for expression of amino terminal fusions of a biotin acceptor peptide and the
biotinylated β1 subunit was expressed as described previously (7). Biotinylated-β1γ2 was purified from
Sf9 cells using hexahistidine tagged αi1 following the procedure described previously (15).
Measurement of α−βγ interactions by flow cytometry. Binding of fluorescein isothiocyanate
(FITC) labeled myristoylated αi1 (F-αi1) to biotinylated β1γ2 (b-βγ) subunits was measured using a
flow cytometry assay (4;16;17). F-αi1 was kindly provided by Dr. Richard Neubig and was prepared
by reacting purified myristoylated αi1 with FITC, followed by dialysis and repurification by βγ agarose
chromatography (16). The resulting F-αi1 has a specific activity of 11 pmol [35S] GTPγS bound/µg
and 0.9 mol of dye/mol of protein incorporated. Biotinylated-βγ (50 pM final concentration) was
mixed with streptavidin beads in HEDNMLG buffer (20 mM Hepes, pH 8.0, 1 mM EDTA, 1mM
DTT, 150 mM NaCl, 1.2 mM Mg2+, 0.1% C12E10, 10 µM GDP) at room temperature. After 30 min,
the beads were washed twice by centrifugation in a microcentrifuge with HEDNMLG buffer and
resuspended in the same buffer at 105 beads /mL (50 pM βγ). For α subunit dissociation experiments,
5
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

the beads with bound 50 pM βγ subunits were premixed with 300 pM F-αi for 10 min prior to the
addition of the different peptides or αi1. For equilibrium binding measurements 300 pM F-αi1 and
peptides or αi1 were added simultaneously. The amount of F-αi1 bound to beads with b-βγ was
assayed at the times indicated in the figure legends using a Becton-Dickinson FACs Calibur flow
cytometer. Non-specific binding, determined by the simultaneous addition of 300 pM F-αi1 and 50 nM
myristoylated αi1 subunits to the b-βγ bound beads was 10-20% of the total signal and was subtracted
from the mean channel numbers from each experiment unless otherwise indicated.
6
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Results
Comparison of the βγ ”hot spot” dependent dissociation mechanism with AMF induced subunit
dissociation. We wanted to determine if the βγ “hot spot” dependent mechanism for subunit
dissociation was similar in magnitude to classically described mechanisms for subunit dissociation.
AMF (AlF4- plus Mg2+) is a well-characterized mediator of G protein subunit dissociation. We chose
AMF for the comparison rather than GTP or GTPγS because the rate of dissociation of the G protein
subunits by AMF is not limited by the GDP release rate.
To measure αi binding and dissociation from βγ we used a flow cytometry assay developed by
Sarvazyan et al. (16) that measures protein α-βγ binding at concentrations of α and βγ near the Kd for
their interaction. In this assay, biotinylated βγ (b-βγ) was immobilized on the surface of beads and
fluorescein labeled αi1 (F-αi1) was added. F-αi1 that bound to b-βγ on the beads was detected by the
flow cytometer. The binding was concentration dependent and on and off rates could be measured.
Thus, this is an ideal method for measuring binding and dissociation of α subunits from βγ subunits.
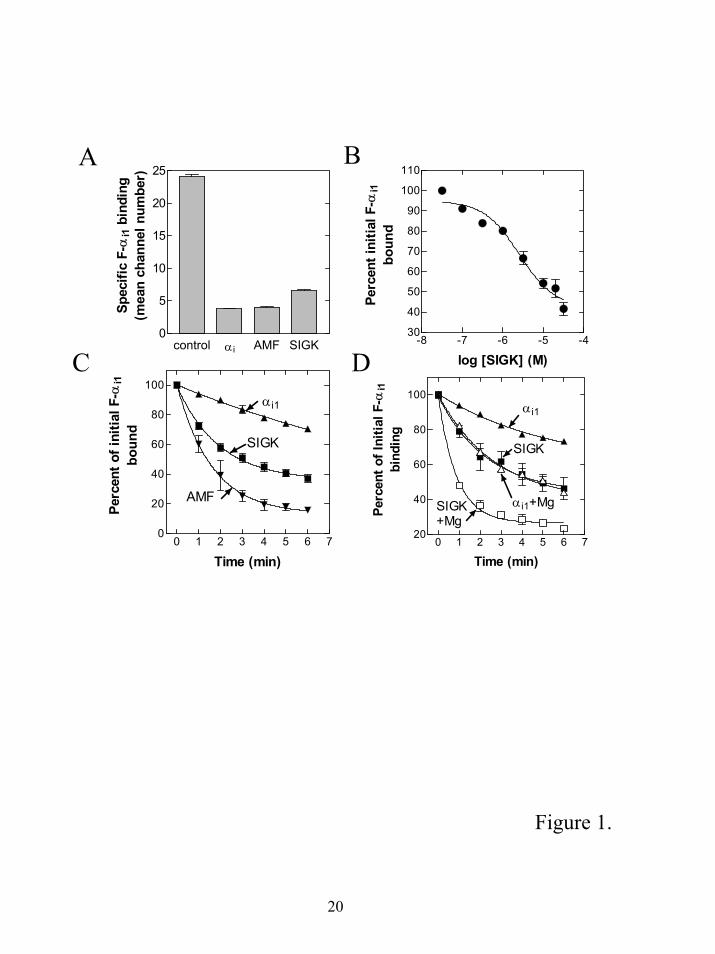
To demonstrate that the βγ “hot spot” binding peptide (SIGK) and AMF could inhibit α subunit
interactions with βγ, they were compared for their effects on the initial binding of F-αi1 to b-βγ.
Excess unlabeled myristoylated αi1 was used to measure non-specific binding. SIGK, AMF and αi all
inhibited formation of the heterotrimeric F-αi1βγ complex to comparable extents indicating they were
equally effective at preventing heterotrimer formation (Fig 1A).
Next we measured the effects of SIGK on subunit dissociation. First we determined the
concentration of SIGK required for dissociation of an F-αi1βγ complex (Fig. 1B). Increasing
concentrations of SIGK caused a progressive increase in disruption of the preformed complex with
maximal dissociation observed between 20 and 30 µM peptide. To compare the SIGK mediated
dissociation rates with AMF, we compared a maximally effective concentration of SIGK with a
standard concentration of AMF (30 µM AlCl3, 10 mM NaF, 10 mM MgCl2) that should be sufficient
7
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

to activate all of the α subunits (18). Both SIGK and AMF enhanced the release of α subunits from βγ
subunits relative to the intrinsic αi1 off rate (measured by addition of a 50 fold excess of unlabeled
myristoylated αi1) (Fig. 1C). The koff with AMF was 0.62 min-1 and with SIGK was 0.5 min-1
compared to the intrinsic off rate of 0.1 min-1. The intrinsic off rates are difficult to calculate
accurately because the extent of intrinsic dissociation in this short time course was only 20 %.
Significantly more intrinsic dissociation occurs with longer time courses but it is during this initial
dissociation phase that the greatest effects of the peptides are observed. Since the off rates induced by
SIGK and AMF are similar, it indicates that the βγ subunit binding peptide dependent dissociation
mechanism has the potential to increase the α-βγ dissociation rate to an extent comparable to the
classical G protein α subunit activation dependent dissociation.
Effect of MgCl2 on the rate of α subunit dissociation from the heterotrimeric complex. Magnesium is
known to profoundly affect the interaction of α and βγ subunits, and dissociation by activation of the
α subunit with either GTPγS or AlF4- is strongly dependent upon Mg2+concentration (1;19).
Magnesium stabilizes interactions of AlF4- and GTPγS at the nucleotide-binding site and directly
influences subunit dissociation through an undefined lower affinity site. For the experiment in figure
1C, SIGK mediated dissociation was measured with 1.2 mM MgCl2 and 1 mM EDTA (0.2 mM free
Mg2+). To determine if Mg2+ was required for subunit dissociation by SIGK, we measured SIGK
mediated dissociation in the absence of MgCl2, in the presence of 1 mM EDTA (Fig. 1D). SIGK was
able to significantly enhance subunit dissociation in the absence of Mg2+ indicating Mg2+ is not
required for this dissociation mechanism. Addition of Mg2+at a final concentration of 10 mM
increased the intrinsic off rate of F-αi as expected from 0.1 min-1 in the absence of Mg2+ to 0.37 min-1
with 10 mM Mg2+. The rate of α subunit dissociation by SIGK increased from 0.43 min-1 without
Mg2+ to 1.14 min-1 with 10 mM Mg2+.
Effect of βγ binding peptides on the interaction of F-αi1 with βγ. Since SIGK and other “hot spot”
binding peptides enhance the off rate of α from βγ subunits it suggests they do not act simply by
8
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

competition for reformation of spontaneously dissociating α-βγ complexes. Such a mechanism would
not increase the dissociation rate relative to the intrinsic off rate of α dissociation from βγ. Based on
this we have suggested that the peptides stabilize a conformation of βγ subunits that has a lower
affinity for α leading to an enhanced rate of α subunit dissociation (4). An alternative mechanism
might be that a small peptide could compete for one of the two major contacts of α subunits with the
sides and top of the β subunit torus during transient separation of one of these contacts, thereby
leading to an enhancement of the dissociation rate. If there were nothing unique about the “hot spot”
binding peptides and it were simply a competitor at the α-βγ interface this model would predict that
any peptide that bound at the α subunit interface with βγ should enhance the off rate.
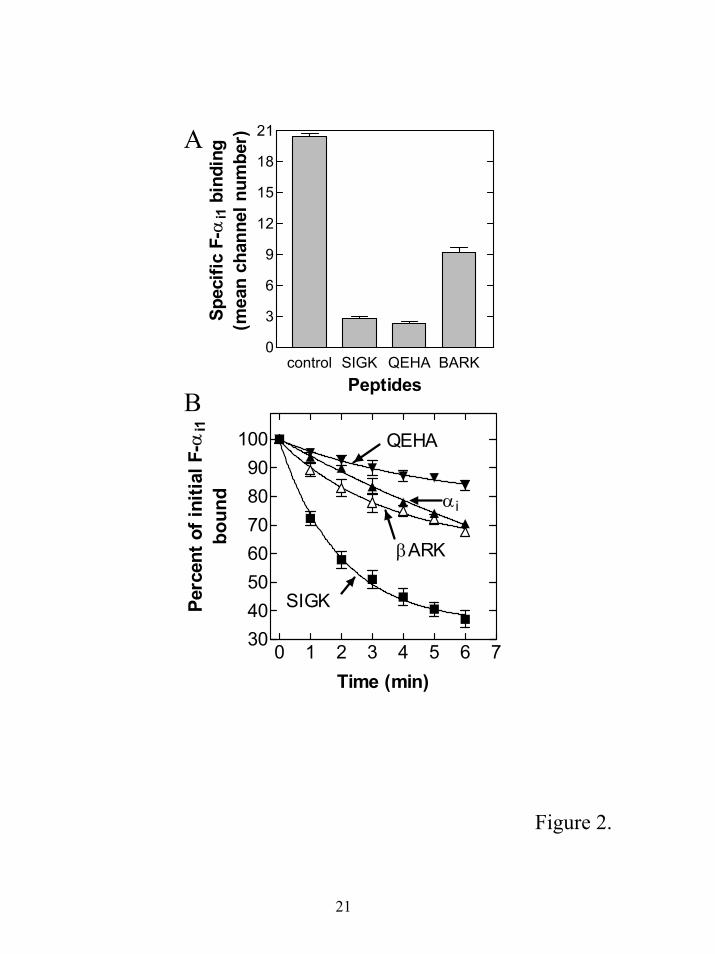
To try to distinguish between these two mechanisms, we tested two βγ binding peptides
thought to bind to βγ subunits at the α subunit interface. QEHA is a 27-residue peptide derived from
the second catalytic domain of adenylyl cyclase 2 (amino acids 956-982) and inhibits βγ regulation of
several effectors including K+ channels, phospholipase C-β and adenylyl cyclase (20). The IC50 for
QEHA effects on most processes was 50-200 µM. Cross-linking of the QEHA peptide to the beta
subunit is prevented by the α subunit suggesting it binds to βγ within the α subunit binding site
(20;21). Another peptide derived from the C-terminal region of GRK2 (βARK-ct, 643-670) (22) also
binds to βγ subunits, has an IC50 of 100 µM for its effects and has properties consistent with binding at
the α subunit binding site on βγ subunits. This notion is supported by the recently determined co-
crystal structure of α subunits with GRK2 (23) demonstrating binding of the region of GRK2
corresponding to this peptide to a region on βγ that overlaps with the α subunit binding site.
To confirm that these peptides block α-βγ interactions, the ability of these peptides to block
binding of F-αi1 to βγ was tested by flow cytometry. Both peptides inhibited the heterotrimer
formation with QEHA inhibiting by 90% and βARK-ct peptide (300µM) by more than 65% compared
to 85% by SIGK (Fig 2A). This supports the idea that QEHA and βARK-ct peptide can inhibit α βγ
interactions probably by directly competing for α binding to βγ.
9
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

To determine if QEHA and βARK-ct peptides can stimulate release of αi1 from the
heterotrimer, we measured the off rates in the presence of these peptides at concentrations that
significantly inhibited α binding to βγ. Neither of these peptides caused any significant enhancement
of the rate of dissociation of the α subunits from the preformed complex (Fig 2B). There are some
minor differences in the rate of dissociation for QEHA, βARK and the intrinsic dissociation rate but
these differences were not consistently reproducible and were not significant. Thus while being able
to compete for α-βγ interactions they are unable to promote dissociation of α from βγ. These data
strongly suggest that the βγ “hot spot”-binding peptides act through a unique mechanism that does not
involve simple steric occlusion of α binding sites on βγ subunits.
Effect of a GPR consensus motif peptide on association and dissociation of Gαβγ. A class of
proteins that stimulate G protein βγ subunit signaling by a nucleotide exchange independent
mechanism is the AGS proteins. AGS3 binds to α subunits, inhibits GDP release and promotes βγ
subunit signaling in yeast and is involved in establishing βγ and α subunit dependent asymmetric cell
division in Caenorhabditis elegans and Drosophila. A 28 amino acid (GPR) motif consensus peptide
derived from AGS3 and other related proteins is able to bind to α subunits and inhibit nucleotide
exchange in a manner comparable to larger protein fragments of AGS3 (10;24). Since a GPR like
peptide from RGS14 binds to α subunits near the α-βγ interface it was proposed that these peptides
enhance βγ subunit dependent processes by competing for rebinding of α to βγ by steric occlusion of
the βγ binding site on α (13).
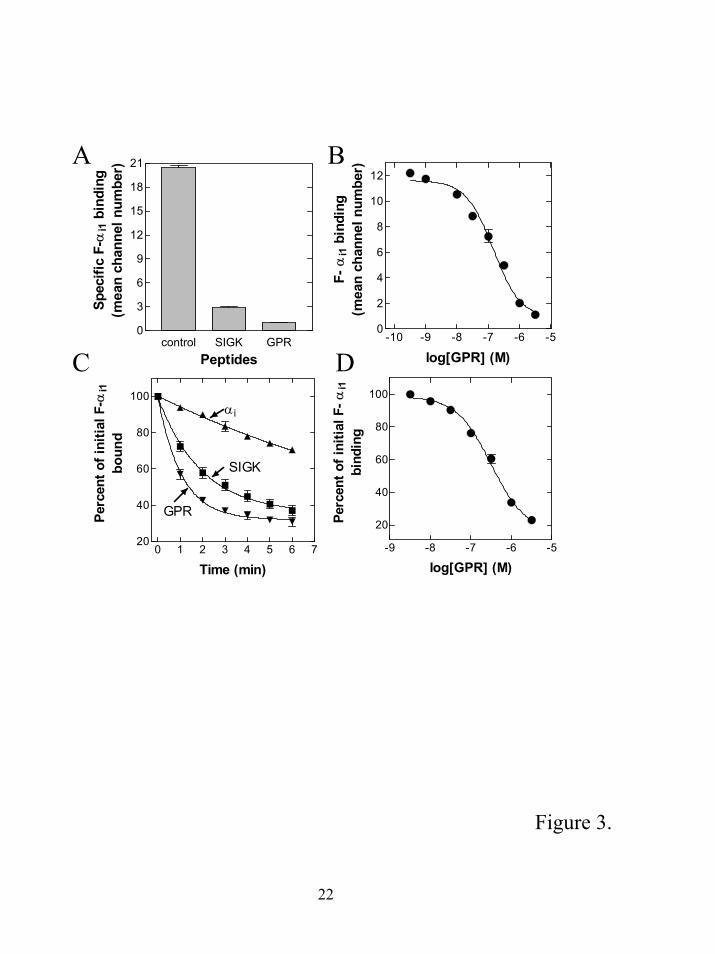
We directly tested if the GPR peptide could inhibit α-βγ binding and/or promote subunit
dissociation. 1 µM GPR peptide was able to inhibit α binding to βγ to an extent greater than that
observed with 10 µM SIGK (Fig. 3A). The IC50 for inhibition of α-βγ interactions was approximately
~ 250 nM (Fig. 3B) comparable to that observed for the ability of the peptide to inhibit GDP
dissociation from α subunits. Next we tested whether 1 µM GPR peptide could cause dissociation of a
10
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

preformed α-βγ complex. The GPR peptide caused a rapid dissociation of the G protein subunits,
about two-fold faster (0.95 min-1) than that observed with 25µM SIGK (0.5 min-1) and about 13 fold
higher than the intrinsic off rate of the F-αi1 subunit (Fig 3C). The GPR peptide very potently
promoted dissociation (Fig 3D) and was about 10 fold more potent than SIGK (compare figure 3D
with figure 1B). Thus, the GPR consensus peptide is a very potent and effective promoter of G protein
subunit dissociation. Since they dramatically increase the dissociation rate of α from βγ it strongly
suggests that the GPR peptides act by a non-competitive mechanism.
11
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Discussion
The standard model for G protein activation and subunit dissociation involves receptor
mediated release of GDP from the G protein α subunit and subsequent binding of GTP (1). This
binding of GTP to the active site of α subunits causes a conformational change in the switch
regions of the α subunit. In particular, conformational changes in the switch II region of the α
subunit at the α/βγ interface are thought to lead to subunit dissociation (2;25). Recent data from
several laboratories have identified receptor independent mechanisms for heterotrimeric G
protein activation (8;12). GPR motif peptides act as guanine nucleotide release inhibitors in
vitro and AGS proteins containing this motif are able to stimulate G protein signaling (8;10).
The precise mechanism for G protein activation has not been elucidated, but it has been proposed
that the GPR motif simply competes for α-βγ interactions by binding at the interface between
these two subunits leading to net G protein subunit dissociation (13). We identified peptides
using random peptide phage display that bind to a “hot spot” on the G protein βγ subunit surface
(3). These peptides can also activate G protein signaling in intact cells, and similar to AGS
proteins, do so in a nucleotide exchange independent fashion (4).
Here we have compared the effects of the GPR peptides and βγ “hot spot” binding
peptides with peptides that are thought to bind at the G protein βγ subunit-α subunit interface.
We show that AC2 and βARK derived peptides are capable of blocking βγ-α subunit interactions
consistent with previous data suggesting they bind at the α-βγ interface. These competitor
peptides were unable to enhance the rate of G protein subunit dissociation while SIGK and GPR
peptides significantly enhanced subunit dissociation. The enhanced rates of G protein subunit
dissociation by SIGK and GPR peptides were comparable to a known activator of G proteins,
AMF. This strongly suggests that neither the SIGK peptides nor the GPR peptides are simply
acting by preventing reassembly of dissociated subunits to lead to G protein activation. The
GPR and βγ binding peptides are unique in that they induce subunit dissociation, probably
through conformational alterations of α or βγ subunits respectively.
12
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

In the X-ray crystallographic structural model of a GPR like peptide from RGS 14 in a
complex with αi1 (13), conformations of switch I and II are altered relative to heterotrimeric αiGDP
βγ. These conformational differences could result in subunit dissociation via a mechanism analogous
to the GTP or AlF4- dependent conformational changes in the switch regions of the α subunit that
contribute to subunit dissociation (26;27).
GPR proteins play important roles in regulation of asymmetric cell division in Drosophila (28)
and in C. elegans (29;30). In particular, they appear to regulate the polarized distribution of free βγ
and α subunits derived from heterotrimers required for correct orientation of mitotic spindles. In α
subunit immunoprecipitates from Drosophila sensory organ precursor cells, Pins (An AGS3
homologue in Drosophila) and a peptide representing the GPR motif from Pins disrupted α-βγ
interactions when added during the immunoprecipitation (28). Our results that directly measure
subunit dissociation are consistent with these results.
The data presented also support a model where a conformational change in βγ subunits induced
by “hot spot” binding peptides results in destabilization of interactions with α subunits and an increase
in the koff for subunit dissociation. βγ subunits have indeed been shown to undergo conformational
changes upon binding of phosducin (31;32) although the functional significance of these changes are
not entirely understood. A mechanism for βγ binding-peptide mediated enhancement of subunit
dissociation, whatever the details of the mechanism, is clearly very distinct from other mechanisms
that exist for promoting subunit dissociation by either GPR peptides or nucleotide binding that involve
conformational alterations of the switch regions of the α subunits. That such a mechanism exists
suggests that it probably has been exploited by natural systems for promoting subunit dissociation.
Since some effectors of βγ subunits appear to bind at the same site as the βγ binding peptides perhaps
they stabilize a conformation required for their activation and that has a reduced affinity for αGDP.
Effectors that were more efficient at stabilizing this conformation might have an extended lifetime of
activation because αGDP subunits would not be readily able to turn off βγ subunit signaling.
13
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure Legends
Figure 1. The “hot spot” binding peptides are nearly as effective as AMF at promoting subunit
dissociation. A. SIGK inhibits the heterotrimeric Gαβγ complex formation. 300 pM F-αi was
preincubated with 50 nM unlabelled myristoylated -α subunit, 10 µM SIGK or AMF (60 µM AlCl3,
10 mM NaF, 10 mM MgCl2) in HEDNMLG buffer followed by incubation with biotinylated β1γ2 (50
pM) bound to streptavidin beads. After 30 minutes, F-αi1 binding to the b-βγ beads was measured
using flow cytometer. For each sample, 3000 beads were assessed. Data are means ± S.E.M. of the
pooled data from three experiments. B. SIGK concentration dependence for dissociation of
heterotrimeric complex. 300 pM F-αi1 was mixed with 50 pM b-βγ in the HEDMLNG buffer and
incubated for 15 minutes to allow the heterotrimer complex formation, followed by the addition of the
indicated concentrations of SIGK. The amount of F-αi1 bound was measured 2 minutes after addition
of the peptide. Nonspecific binding was measured in the presence of excess myristoylated -αi1 subunit
added simultaneously with the F-αi1 subunit and was subtracted from the total fluorescence for each of
the peptide concentrations used. Data are means ± S.E.M. of the pooled data from 3 experiments. The
curve was fit with a sigmoid dose response function using Graph Pad Prism data analysis software. C.
Comparison of the dissociation rates of F-α- b-βγ complex. 300 pM F-αi1 was preincubated with b-βγ
as in B and dissociation was initiated by the addition of 50 nM myristoylated αi1 subunits (▲), 25 µM
SIGK (■) or AMF (30 µM AlCl3, 10 mM NaF, 10 mM MgCl2) (▼). Intrinsic dissociation rates were
not significantly different if 25, 50 or 100 nM myristoylated αi1 were used. Data shown are means ±
S.E.M. of three experiments. Solid lines are fits of the data to single exponential dissociation
functions. D. Magnesium is not required for SIGK mediated dissociation but enhances the rate of
dissociation. The rate of F-αi1 dissociation from a preformed heterotrimeric F-αi1βγ complex was
measured as in Fig 1B and 1C. The additions were: 50 nM myristoylated αi1 subunits with 1 mM
EDTA and no added MgCl2 (▲) or 11 mM MgCl2 (∆); 25 µM SIGK with 1 mM EDTA and no added
MgCl2 (■) or 11 mM MgCl2 (□). Results are pooled data from three separate experiments and the
curves are fit to single exponential decay functions.
14
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 2. Binding of peptides to βγ at α subunit binding surfaces does not enhance the rate of
subunit dissociation. A. Competition for binding of F-αi1 to βγ in the presence of β-subunit
interacting peptides QEHA and βARK. Biotinylated-βγ was bound to streptavidin beads in
HEDNMLG buffer. QEHA, βARK-ct (643-670) (300 µM each) or 25 µM SIGK were added to b-βγ
bound beads followed by addition of 300 pM F-αi1, incubation for 30 minutes at room temperature,
and measurement of the bound αi1 by flow cytometry. Maximal binding of F-αi1 was determined in
the absence of any competitors (control). Non-specific binding, determined in the presence of 50 nM
unlabelled myristoylated αi1, was subtracted from the data. The data obtained are means ± S.E.M. of
three separate experiments. B. Comparison of kinetics of F-αi1 dissociation from βγ by SIGK, QEHA
and βARK peptides. Peptides or excess myristoylated αi1 subunits were added to a complex formed
with 300 pM F-αi1 and 50 pM βγ subunits as in Fig 1C. (▲) 100 nM myr-αi1, (▼) 300 µM QEHA,
(∆) 300 µM βARK and (■) 25 µM SIGK. Data are pooled from three separate experiments.
Figure 3. A GPR consensus peptide promotes G protein subunit dissociation. A. Influence of
GPR motif peptide on the assembly of α with βγ subunits. F-αi1 (300 pM) was added to b-βγ (50 pM)
in the presence of 1 µM GPR peptide or 25 µM SIGK, incubated for 30 minutes at room temperature,
and the extent of subunit association was determined by flow cytometry. Nonspecific binding in the
presence of 50 nM unlabelled myristoylated αi1 subunits was subtracted from the total binding. B.
Concentration requirement for GPR peptide inhibition of heterotrimer formation. The reaction
conditions as in A except the final concentration of the GPR motif peptide was varied from to 1.0 nM
to 3.0 µM. C. G protein subunit dissociation by GPR consensus peptide and SIGK. 1 µM GPR (▼) or
25 µM SIGK (■) or 50 nM unlabelled myristoylated αi (▲) were added to the preformed F-αi βγ
complex as in Figure 1C. The amount of nonspecific binding obtained in the presence of unlabeled
myristoylated αi1 subunits prior to the formation of the complex was subtracted from each value. D.
Concentration dependence of GPR peptide on the dissociation of G protein subunits. The amount of F-
15
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

αi1 remaining bound was measured after 2 minutes of incubation of the preformed F-αi1 βγ complex
with increasing concentrations of the GPR motif peptide and compared with the extent of initial
binding of F-αi1 to b-βγ in the absence of the GPR peptide. All data are the mean ± S.E.M. of three
individual experiments.
16
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgements. I would like to thank Dr. Richard Neubig for both his advice on performing the
flow cytometry experiments and for providing us with fluorescein labeled αi1.
17
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Literature Cited
1. Gilman, A. G. (1987) Ann.Rev.Biochem. 56, 615-649
2. Hamm, H. E. (1998) J.Biol.Chem. 273, 669-672
3. Scott, J. K., Huang, S. F., Gangadhar, B. P., Samoriski, G. M., Clapp, P., Gross, R. A., Taussig, R., and Smrcka, A. V. (2001) EMBO J. 20, 767-776
4. Goubaeva, F., Ghosh, M., Malik, S., Yang, J., Hinkle, P. M., Griendling, K. K., Neubig, R. R., and Smrcka, A. V. (2003) J.Biol.Chem. 278, 19634-19641
5. Delano, W. L., Ultsch, M. H., de Vos, A. M., and Wells, J. A. (2000) Science 287, 1279-1283
6. Delano, W. L. (2002) Curr.Opin.Struct.Biol. 12, 14-20
7. Takesono, A., Cismowski, M. J., Ribas, C., Bernard, M., Chung, P., Hazard, S., Duzic, E., and Lanier, S. M. (1999) J.Biol.Chem. 274, 33202-33205
8. Blumer, J. B. and Lanier, S. M. (2003) Receptors Channels 9, 195-204
9. Siderovski, D. P., Diverse-Pierluissi, M., and De Vries, L. (1999) Trends Biochem.Sci. 24, 340-341
10. Peterson, Y. K., Bernard, M. L., Ma, H. Z., Hazard, S., Graber, S. G., and Lanier, S. M. (2000) J.Biol.Chem. 275, 33193-33196
11. Peterson, Y. K., Hazard, S., Graber, S. G., and Lanier, S. M. (2002) J.Biol.Chem. 277, 6767-6770
12. De Vries, L., Fischer, T., Tronchere, H., Brothers, G. M., Strockbine, B., Siderovski, D. P., and Farquhar, M. G. (2000) Proc.Natl.Acad.Sci U.S.A. 97, 14364-14369
13. Kimple, R. J., Kimple, M. E., Betts, L., Sondek, J., and Siderovski, D. P. (2002) Nature 416, 878-881
14. Gallop, M. A., Barrett, R. W., Dower, W. J., Fodor, S. P., and Gordon, E. M. (1994) J.Med.Chem. 37, 1233-1251
15. Kozasa, T. and Gilman, A. G. (1995) J.Biol.Chem. 270, 1734-1741
16. Sarvazyan, N. A., Remmers, A. E., and Neubig, R. R. (1998) J.Biol.Chem. 273, 7934-7940
17. Sarvazyan, N. A., Lindau, M., and Neubig, R. R. (2002) Biochem. 41, 12858-12867
18
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

19
18. Higashijima, T., Ferguson, K. M., Sternweis, P. C., Ross, E. M., Smigel, M. D., and Gilman, A. G. (1987) J.Biol.Chem. 262, 752-756
19. Katada, T., Northup, J. K., Bokoch, G. M., Ui, M., and Gilman, A. G. (1984) J.Biol.Chem. 259, 3578-3585
20. Chen, J., DeVivo, M., Dingus, J., Harry, A., Li, J., Sui, J., Carty, D. J., Blank, J. L., Exton, J. H., Stoffel, R. H., Inglese, J., Lefkowitz, R. J., Logothetis, D. E., Hildebrandt, J., and Iyengar, R. (1995) Science 268, 1166-1169
21. Weng, G. Z., Li, J. R., Dingus, J., Hildebrandt, J. D., Weinstein, H., and Iyengar, R. (1996) J.Biol.Chem. 271, 26445-26448:
22. Koch, W. J., Inglese, J., Stone, W. C., and Lefkowitz, R. J. (1993) J.Biol.Chem. 268, 8256-8260
23. Lodowski, D. T., Pitcher, J. A., Capel, W. D., Lefkowitz, R. J., and Tesmer, J. J. G. (2003) Science 300, 1256-1262
24. Natochin, M., Lester, B., Peterson, Y. K., Bernard, M. L., Lanier, S. M., and Artemyev, N. O. (2000) J.Biol.Chem. 275, 40981-40985
25. Sprang, S. R. (1997) Ann.Rev.Biochem. 66, 639-678
26. Lambright, D. G., Noel, J. P., Hamm, H. E., and Sigler, P. B. (1994) Nature 369, 621-628
27. Wall, M. A., Coleman, D. E., Lee, E., Iniguez-Lluhi, J. A., Posner, B. A., Gilman, A. G., and Sprang, S. R. (1995) Cell 83, 1047-1058
28. Schaefer, M., Petronczki, M., Dorner, D., Forte, M., and Knoblich, J. A. (2001) Cell 107, 183-194
29. Gotta, M. and Ahringer, J. (2001) Nat.Cell Biol. 3, 297-300
30. Gotta, M., Dong, Y., Peterson, Y. K., Lanier, S. M., and Ahringer, J. (2003) Current Biology 13, 1029-1037
31. Loew, A., Ho, Y. K., Blundell, T., and Bax, B. (1998) Structure 6, 1007-1019
32. Gaudet, R., Bohm, A., and Sigler, P. B. (1996) Cell 87, 577-588
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
control αi AMF SIGK0
5
10
15
20
25
Spec
ific
F-α
i1 b
indi
ng(m
ean
chan
nel n
umbe
r)
-8 -7 -6 -5 -430
40
50
60
70
80
90
100
110
log [SIGK] (M)Pe
rcen
t ini
tial F
-αi1
boun
d
B
0 1 2 3 4 5 6 70
20
40
60
80
100αi1
SIGK
AMF
Time (min)
Perc
ent o
f ini
tial F
-αi1
boun
d
C
0 1 2 3 4 5 6 720
40
60
80
100αi1
SIGK
αi1+MgSIGK+Mg
Time (min)
Perc
ent o
f Ini
tial F
-αi1
bind
ing
D
A
Figure 1.
20
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
control SIGK QEHA BARK0
3
6
9
12
15
18
21
Peptides
Spec
ific
F-α
i1 b
indi
ng(m
ean
chan
nel n
umbe
r)
0 1 2 3 4 5 6 730405060708090100
SIGK
QEHA
βARK
αi
Time (min)
Perc
ent o
f ini
tial F
-αi1
boun
d
A
B
Figure 2.
21
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
control SIGK GPR0
3
6
9
12
15
18
21
Peptides
Spec
ific
F-α
i1 b
indi
ng(m
ean
chan
nel n
umbe
r)
-10 -9 -8 -7 -6 -50
2
4
6
8
10
12
log[GPR] (M)F-
αi1
bin
ding
(mea
n ch
anne
l num
ber)
0 1 2 3 4 5 6 720
40
60
80
100αi
SIGK
GPR
Time (min)
Perc
ent o
f ini
tial F
-αi1
boun
d
-9 -8 -7 -6 -5
20
40
60
80
100
log[GPR] (M)
Perc
ent o
f ini
tial F
-αi1
bind
ing
A B
C D
Figure 3.
22
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Mousumi Ghosh, Yuri K Peterson, Stephen M Lanier and Alan V Smrckasubunit dissociation
Receptor and nucleotide exchange independent mechanisms for promoting G protein
published online July 24, 2003J. Biol. Chem.
10.1074/jbc.C300271200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from











![Cyclic nucleotide phosphodiesterase 3B is …cAMP and potentiate glucose-induced insulin secretion in pancreatic islets and β-cells [3]. Cyclic nucleotide phosphodiesterases (PDEs),](https://static.fdocument.org/doc/165x107/5e570df60e6caf17b81f7d2a/cyclic-nucleotide-phosphodiesterase-3b-is-camp-and-potentiate-glucose-induced-insulin.jpg)






