







γλώσσες
Σελίδες
Νομικός

1
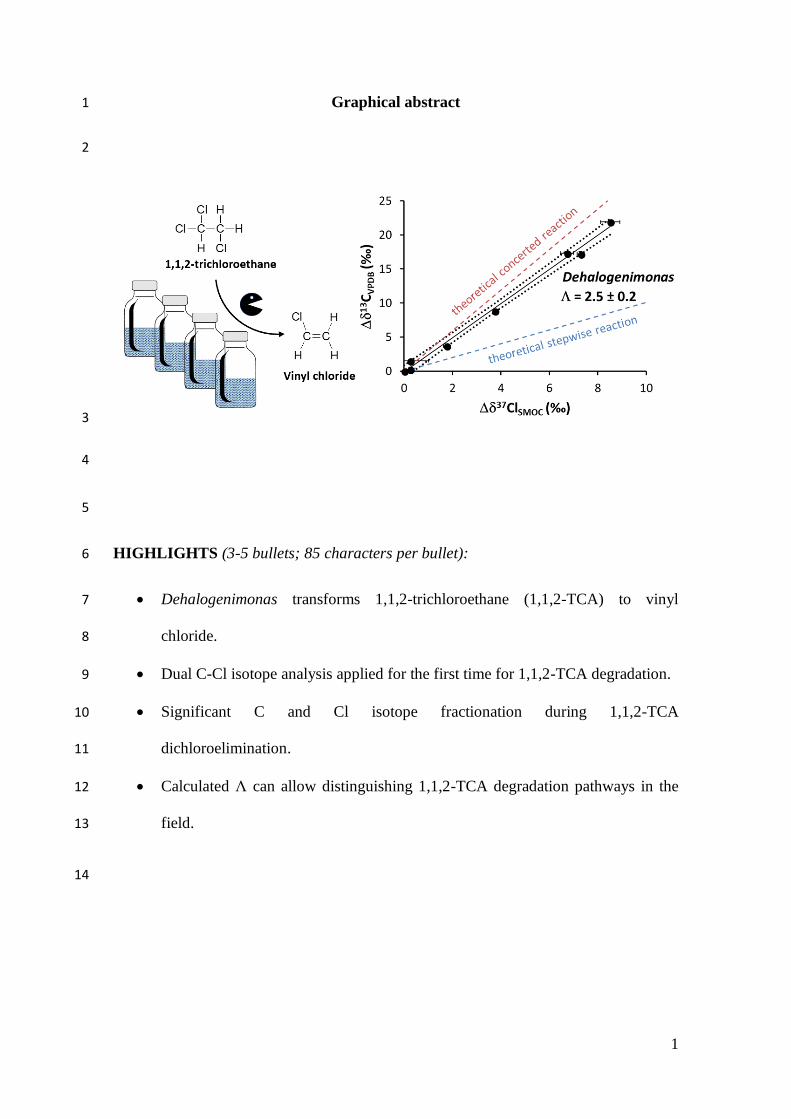
Graphical abstract 1
2
3
4
5
HIGHLIGHTS (3-5 bullets; 85 characters per bullet): 6
Dehalogenimonas transforms 1,1,2-trichloroethane (1,1,2-TCA) to vinyl 7
chloride. 8
Dual C-Cl isotope analysis applied for the first time for 1,1,2-TCA degradation. 9
Significant C and Cl isotope fractionation during 1,1,2-TCA 10
dichloroelimination. 11
Calculated Λ can allow distinguishing 1,1,2-TCA degradation pathways in the 12
field. 13
14

2
Dual carbon - chlorine isotope fractionation during 15
dichloroelimination of 1,1,2-trichloroethane by an enrichment 16
culture containing Dehalogenimonas sp. 17
18
Mònica Rosell1, Jordi Palau*1,2, Siti Hatijah Mortan3,a, Gloria Caminal4, Albert Soler1, 19
Orfan Shouakar-Stash5,6, Ernest Marco-Urrea3 20
21
1 Grup MAiMA, Mineralogia Aplicada, Geoquímica i Geomicrobiologia, Departament 22
de Mineralogia, Petrologia i Geologia Aplicada, Facultat de Ciències de la Terra, 23
Universitat de Barcelona (UB), Martí Franquès s/n, 08028 Barcelona, Spain. 24
2 Institute of Environmental Assessment and Water Research (IDAEA), CSIC, and 25
Hydrogeology Group (UPC−CSIC), Jordi Girona 18-26, 08034 Barcelona, Spain 26
3 Departament d’Enginyeria Química, Biològica i Ambiental, Universitat Autònoma de 27
Barcelona (UAB), 08193 Bellaterra, Barcelona, Spain. 28
4 Institut de Química Avançada de Catalunya (IQAC), CSIC, Jordi Girona 18-26, 08034 29
Barcelona, Spain 30
5 Department of Earth and Environmental Sciences, University of Waterloo, Waterloo, 31
Ontario N2L 3G1, Canada. 32
6 Isotope Tracer Technologies Inc., Waterloo, Ontario N2 V 1Z5, Canada 33
a Current address: Faculty of Chemical and Natural Resources Engineering, Universiti 34
Malaysia Pahang, Lebuhraya Tun Razak, Gambang, 26300 Kuantan, Pahang, Malaysia. 35
36
*Corresponding author: [email protected] 37
38
39

3
ABSTRACT 40
41
Chlorinated ethanes are frequent groundwater contaminants but compound specific 42
isotope analysis (CSIA) has been scarcely applied to investigate their degradation 43
pathways. In this study, dual carbon and chlorine isotope fractionation was used to 44
investigate for the first time the anoxic biodegradation of 1,1,2-trichloroethane (1,1,2-45
TCA) using a Dehalogenimonas-containing culture. The isotopic fractionation values 46
obtained for the biodegradation of 1,1,2-TCA were ɛC = -6.9 ± 0.4‰ and ɛCl = -2.7 ± 47
0.3‰. The detection of vinyl chloride (VC) as unique byproduct and a closed carbon 48
isotopic mass balance corroborated that dichloroelimination was the degradation 49
pathway used by this strain. Combining the values of δ13C and δ37Cl resulted in a dual 50
element C-Cl isotope slope of Λ=2.5 ± 0.2‰. Investigation of the apparent kinetic 51
isotope effects (AKIEs) expected for cleavage of a C-Cl bond showed an important 52
masking of the intrinsic isotope fractionation. Theoretical calculation of Λ suggested 53
that dichloroelimination of 1,1,2-TCA was taking place via simultaneous cleavage of 54
two C-Cl bonds (concerted reaction mechanism). The isotope data obtained in this study 55
can be useful to monitor natural attenuation of 1,1,2-TCA via dichloroelimination and 56
provide insights into the source and fate of VC in contaminated groundwaters. 57
58
Keywords: Dehalogenimonas; dual isotope fractionation; dichloroelimination; 59
organohalide-respiring bacteria; 1,1,2-trichloroethane. 60
61
62

4
1. Introduction 63
1,1,2-Trichloroethane (1,1,2-TCA) has been widely used as a solvent and chemical 64
intermediate in the industry (Pankow and Cherry, 1996). Improper storage and 65
accidental spills have contributed to 1,1,2-TCA being a frequent detected contaminant 66
in groundwater at industrial facilities (ATSDR, 1989). In the United States, it is ranked 67
166 out of 275 substances on the Priority List of Hazardous Substances based on a 68
combination of its frequency, toxicity, and potential for human exposure (ATSDR, 69
2015). 70
Quantification of the distribution and fate of chlorinated contaminants and 71
degradation products in the subsurface is a complex task since biological, chemical, and 72
physical processes may affect them (Němeček et al., 2017). Biological transformation 73
of 1,1,2-TCA is influenced by the intrinsic heterogeneity of natural environments that 74
allows for different redox conditions to occur either spatially or temporally separated. 75
Under anoxic conditions, reductive dechlorination is expected to be the prevailing 76
mechanism to transform 1,1,2-TCA by two different biodegradation pathways: 77
hydrogenolysis and dichloroelimination. In the case of dichloroelimination, two vicinal 78
C-Cl bonds of 1,1,2-TCA are cleaved to produce vinyl chloride (VC), whereas during 79
hydrogenolysis 1,1,2-TCA is sequentially transformed by single C-Cl bond cleavage to 80
1,2-dichloroethane (1,2-DCA) and monochloroethane (Moe et al., 2016; Zhao et al., 81
2015) (Fig. 1). The key organisms catalyzing hydrogenolysis and dichloroelimination 82
are organohalide-respiring bacteria (OHRB), which can use 1,1,2-TCA as respiratory 83
electron acceptor (Leys et al., 2013). To date, dichloroelimination of 1,1,2-TCA has 84
been described for OHRB belonging to the genus Dehalobacter and Dehalogenimonas 85
(Grostern and Edwards, 2006; Mortan et al., 2017; Yan et al., 2009), but 86
hydrogenolysis only for Desulfitobacterium (Zhao et al., 2015). Under oxic conditions, 87

5
no bacteria are currently known to use 1,1,2-TCA as growth substrate, but 88
cometabolism of 1,1,2-TCA can occur during aerobic oxidation of methane, propane, 89
butane, n-pentane, n-hexane or ammonia (Frascari et al., 2006, 2008, 2013; Vannelli 90
et al., 1990). The only byproducts identified for aerobic cometabolism of 1,1,2-TCA 91
include chloroacetic acid (which was sequentially oxidized to glyoxylic acid) and minor 92
amounts of VC in microcosms containing a Pseudomonas sp. (Castro and Belser, 93
1990). Abiotic transformation of 1,1,2-TCA can produce a wide array of byproducts, 94
including VC (Patterson et al., 2016), ethane (Song and Carraway, 2005) or 1,1-95
dichloroethene (1,1-DCE) (Pagana et al., 1998) (Fig.1). 96
Knowledge on degradation pathways occurring in an aquifer contaminated with 97
1,1,2-TCA is a key aspect to design suitable bioremediation strategies. However, this is 98
a challenge when the site contains multiple chlorinated aliphatic hydrocarbons because 99
the same daughter products of 1,1,2-TCA dechlorination can be formed from other 100
precursors (i.e., VC is produced from anaerobic reductive dechlorination of DCE 101
isomers or 1,2-DCA) (Hunkeler et al., 2002). It is important to note that VC, produced 102
during biotic or abiotic reductive dichloroelimination of 1,1,2-TCA, is even much more 103
toxic than 1,1,2-TCA. 104
Compound-specific isotope analysis (CSIA) has emerged in recent years as a 105
technique with great potential to elucidate specific reaction pathways even if no 106
products are detected (Elsner, 2010). The magnitude of carbon and chlorine kinetic 107
isotope effects (KIEs) during contaminant degradation relies on the observation that 108
lighter stable isotopes (i.e., 12C, 35Cl) react at faster rates than the heavier ones (i.e., 13C, 109
37Cl). For a given compound and reaction, single element isotope fractionationvalues 110
(are determined in laboratory degradation experiments according to the Rayleigh 111
equation. However, values associated to biodegradation cannot be accurately 112

6
measured in the field because other processes such as sorption or mixing through 113
dispersion also affect contaminant concentration. 114
Two-dimensional CSIA brings the potential to overcome the limitation of single 115
element isotope fractionation values to identify contaminant degradation pathways in 116
the field. Combined changes in isotope ratios of two elements (i.e., 13C and 37Cl) 117
for a given reactant generally correlate in a dual element isotope plot obtaining a slope 118
( = 13C / 37Cl) that reflects the isotope effects of both elements. Hence, values 119
may act as direct indicator for different initial reaction mechanisms. To interpret dual 120
element CSIA data sets obtained from contaminated field sites, it is necessary to know 121
experimental carbon and chlorine isotope enrichment factors and values derived from 122
microbial strains catalyzing known transformation reactions (Cretnik et al., 2013; 123
Kuntze et al., 2016). However, to our knowledge, chlorine isotope fractionation (Cl) 124
and values are not available for 1,1,2-TCA. Hunkeler et al. (2002) showed that 125
dichloroelimination of 1,1,2-TCA to VC in anaerobic microcosms inoculated with 126
contaminated groundwater was accompanied of a relatively weak carbon isotopic 127
fractionation of 1,1,2-TCA (εC = -2.0 ± 0.2‰). Recently, in a laboratory flow-through 128
column experiment consisting of both biodegradable organic carbon and zero valent 129
iron, εC changed from -14.6±0.7‰ to -0.72±0.12‰, being this last value assigned to 130
anaerobic biodegradation (Patterson et al., 2016). 131
The main aims of this research were to measure for the first time dual C-Cl 132
isotope fractionation and to determine the resultant value during biodegradation of 133
1,1,2-TCA with an anaerobic bacterial culture containing a Dehalogenimonas sp. This 134
is valuable information i) to investigate the fate of 1,1,2-TCA in future biodegradation 135
field studies and ii) to get insight into the underlying reaction mechanism involved in 136

7
the dechlorination of 1,1,2-TCA. In addition, carbon isotope values of VC were 137
measured to determine the product isotope pattern during biodegradation of 1,1,2-TCA. 138
139
2. Materials and methods 140
2.1. Biodegradation batch experiments 141
A stable enrichment culture containing a Dehalogenimonas sp. described previously 142
that transforms 1,1,2-TCA to VC via dichloroelimination (Martín-González et al., 143
2015) was used in batch experiments. Each microcosm consisted of 100 mL glass serum 144
bottles containing 65 mL of a sterilized anoxic synthetic medium previously used to 145
grow Dehalococcoides mccartyi strain CBDB1 (Adrian et al., 2000). This medium 146
contained vitamins, trace elements, Na2S × 9 H2O and L-cysteine (0.2 mM each) as 147
reducing agent, and as carbon source either sodium acetate (5 mM) or pyruvate (5 mM) 148
as indicated. The serum bottles were sealed with Teflon-coated butyl rubber septa and 149
aluminum crimp caps and gassed with N2/CO2 (4:1, v/v, 0.2 bar overpressure) and H2 150
(added to an overpressure of 0.4 bar). 1,1,2-TCA was added with a syringe from a stock 151
solution in acetone to give an initial aqueous phase concentration of 20 mol L-1; 152
higher concentrations appeared to be inhibitory for this Dehalogenimonas-containing 153
culture. 154
A total of 16 parallel incubations from the same inoculum were prepared at the 155
same time. Half of these cultures contained acetate and the other half pyruvate as carbon 156
source. Cultures were incubated at 25°C in the dark without shaking. Samples were 157
collected for isotopic and concentration analyses at different extents of 1,1,2-TCA 158
dechlorination. In order to control losses, abiotic transformations, and the transfer of 159
compounds with the inoculum (previous growth in 1,2-dichloropropane, 1,2-DCP) or 160
potential impurities from the stock solution, two types of controls were included in 161

8
triplicate: (i) live controls without 1,1,2-TCA and (ii) abiotic controls containing the 162
growth medium with 1,1,2-TCA but without inoculum. 163
2.2. Analytical methods 164
2.2.1. Concentration and isotopic measurements. 1,1,2-TCA and VC concentrations 165
in serum bottles were monitored along the experiment by taking 0.5 mL headspace (HS) 166
samples with a 1.0 mL pressure-lock precision analytical syringe (Vici, U.S.) and 167
injecting them in a gas chromatograph (GC) model 6890N (Agilent Technologies) 168
equipped with a DB-624 column (30 m × 0.32 mm with 0.25 μm film thickness; Agilent 169
Technologies) and a flame ionization detector (FID), as described elsewhere (Palau et 170
al., 2017). Depending on the measured concentrations (expressed in μmol L–1 of liquid 171
volume) the bottles were sacrificed at different extent of degradation stopping biological 172
activity by adding 12 mL of an oxic, saturated H2SO4/Na2SO4 solution (pH=1). 173
Compound-specific carbon and chlorine isotope analyses were performed by 174
HS-solid-phase micro-extraction (HS-SPME)-GC-isotope ratio mass spectrometry (GC-175
IRMS) as described elsewhere (Palau et al., 2017). δ13C analyses were performed in the 176
Centres Científics i Tecnològics de la Universitat de Barcelona (CCiT-UB), Spain, 177
while δ37Cl were carried out at Isotope Tracer Technologies Inc. (IT2), Canada. For 178
analyzing chlorine isotope ratios of 1,1,2-TCA, the two most abundant fragment ions 179
(m/z 97 and 99) were used, which correspond to isotopologue pairs (i.e., [35Cl212C2
1H3]+ 180
and [37Cl35Cl12C21H4]
+, respectively) that differ by one heavy chlorine isotope. For 181
1,1,2-TCA, the intensities of the most abundant fragment ion peaks are much higher 182
than those of the parent ion peaks. The raw δ37Cl values were calibrated to the standard 183
mean ocean chloride (SMOC) scale using a two-point linear calibration. The standards 184
were dissolved in water and measured similarly to the samples interspersed in the same 185

9
sequence. Duplicate samples and standards were analyzed. The precision (1σ) on the 186
analysis of standards was ≤0.5‰ for δ13C and ≤0.2‰ for δ37Cl. 187
2.2.2. Isotope data evaluation. Carbon and chlorine isotope ratios of 1,1,2-TCA were 188
measured at natural abundance and were expressed using the -notation in per mil (eq. 189
1), 190
191
δ Esample = h (
R( E/ E) l
h
sample
R( E/ E) l
h
standard− 1) ∙ 1000 (1) 192
193
where R is the isotope ratio of heavy (hE) to light (lE) isotopes of an element “E “(e.g., 194
13C/12C and 37Cl/35Cl). The relationship between isotope fractionation and the extent of 195
1,1,2-TCA biodegradation in laboratory experiments was evaluated by a modified form 196
of the Rayleigh distillation equation (2) 197
198
ln (δ ES+1
h 000
δ ES0+1000 h ) =
εbulk
1000∙ ln f (2) 199
200
where hES0 is the initial isotopic composition of element “E” in a substrate “S” and 201
hES is the isotopic composition at a remaining fraction “f” (f = CS/CS0). The 202
compound-average isotope fractionation values (εbulk) were quantified by least squares 203
linear regression of eq. 2 without forcing the regression through the origin (Scott et al., 204
2004) and the uncertainty corresponds to the 95% confidence interval (C.I.) derived 205
from the standard deviation of the regression slope. The Rayleigh equation can also be 206
applied to calculate the isotopic fractionation of chlorine despite the higher natural 207
abundance of 37Cl compared to 13C (Elsner and Hunkeler, 2008). 208

10
To evaluate the product carbon isotope fractionation pattern, the 13C of VC that 209
was produced was calculated using eq. 3, where 13CP is the isotopic composition of the 210
product “P” (i.e., VC) and εbulk is the estimated carbon isotopic fractionation of 1,1,2-211
TCA (eq. 2) (Cretnik et al., 2014; Hunkeler et al., 2005). 212
213
δ CP = δ CS0 13
13 −
εbulk ∙ f ∙ ln f
1 − f (3) 214
215
For a given substrate, intrinsic KIEs during compound transformation are 216
position specific whereas bulk values are calculated from compound-average isotope 217
data (eq. 2). Therefore, observable bulk values must be converted into apparent KIEs 218
(AKIEs) in order to obtain information about the underlying reaction mechanisms 219
(Elsner et al., 2005). For the calculation and interpretation of AKIEs a hypothesis about 220
the reaction mechanism, or assumed reaction mechanism, is necessary. The effects of 221
non-reacting positions within the molecule, as well as of intramolecular competition, are 222
then taken into account using equations 4 and 5, respectively (Elsner et al., 2005), 223
224
εrp ≈ n
x∙ εbulk (4) 225
AKIEC,Cl =1
z∙(εrp
1000)+1
(5) 226
227
where rp is the isotopic fractionation at the reactive position, “n” is the number of 228
atoms of the element considered, “x” is the number of these atoms at reactive sites (i.e., 229
atoms that would experience isotope effects in the given reaction) and “z” the number of 230
identical reactive sites undergoing intramolecular competition. These equations assume 231
the absence of secondary isotope effects. For carbon, secondary isotope effects are 232

11
usually insignificant (Elsner et al., 2005). For dichloroelimination of 1,1,2-TCA to VC, 233
if the two C-Cl bonds are broken in sequence (i.e., stepwise dichloroelimination, single 234
C-Cl bond cleavage at the first reaction step), assuming that the first bond cleavage is 235
the rate determining step, then n = x = z = 2 and n = x = z = 3 for C and Cl, respectively, 236
as all C and Cl atoms are in equivalent position and compete for reaction. On the other 237
hand, if the two C-Cl bonds are broken simultaneously (i.e., concerted 238
dichloroelimination), the average AKIEC and AKIECl for the two reacting positions 239
were calculated since there is no intramolecular competition between them, n = x = 2, z 240
= 1 and n = 3, x = 2, z = 1 for C and Cl, respectively. AKIEs that were calculated 241
assuming stepwise or concerted dichloroelimination are referred hereafter as 242
“AKIEstepwise” and “AKIEconcerted” and their uncertainty was calculated by error 243
propagation. 244
For a given substrate and reaction, the dual C-Cl isotope slope () obtained from 245
13C vs 37Cl isotope plots can be expressed as follows (Elsner, 2010 and references 246
herein): 247
248
ΛC−Cl =∆δ C
13
∆δ Cl 37
≈ε bulk
C
ε bulkCl ≈
(x
n)
C
(x
n)
Cl
∙(A)KIEC−1
(A)KIECl−1∙
1+(A)KIEC∙(zC−1)
1+(A)KIECl∙(zCl−1) (6) 249
250
3. Results and discussion 251
3.1. Concentration and isotope patterns 252
3.1.1. Dechlorination of 1,1,2-TCA by a Dehalogenimonas-containing culture. The 253
anaerobic microcosms amended with pyruvate and acetate as carbon source lasted 254
approximately 7 and 15 days, respectively, at which point the initial 1,1,2-TCA was 255
transformed to VC via dichloroelimination. The concentration of 1,1,2-TCA in the 256

12
abiotic controls (19.0 ± 0.5 mol L-1, ± 1, n = 5) remained at the initial concentration 257
along the experiments, which indicates that compound losses through the caps during 258
incubation were insignificant. The difference in the lag phase between acetate and 259
pyruvate amended microcosms is not probably associated with the carbon source but to 260
the inoculum source that was more enriched in the microcosms with pyruvate. No other 261
volatile organic compounds were detected, especially 1,2-DCA was absent discarding 262
1,1,2-TCA hydrogenolysis. At different stages of 1,1,2-TCA degradation, isotope 263
signatures of 1,1,2-TCA (δ13C and δ37Cl) and VC (δ13C) were measured for all the 264
samples to determine the corresponding isotopic fractionation values of 1,1,2-TCA (εC 265
and εCl) and the carbon isotope pattern of produced VC. 266
3.1.2. Carbon isotope pattern of 1,1,2-TCA. The δ13C of 1,1,2-TCA in the abiotic 267
controls remained constant through both experiments, with a total average value of -268
36.3 ± 0.6‰. In contrast, carbon isotopic composition of 1,1,2-TCA in the cultures 269
became progressively enriched in 13C during its degradation reaching a δ13C value up to 270
-14.3‰ when 96% of 1,1,2-TCA was degraded in both acetate- and pyruvate-containing 271
media (Fig. 2). These results show that despite the differences in the lag phase and the 272
inoculum source, no statistical difference in concentrations and carbon isotope values 273
was observed for the experiments prepared with either acetate or pyruvate as carbon 274
source. Isotopic data from both experiments were combined and the total carbon isotope 275
composition of 1,1,2-TCA followed a Rayleigh trend (r2=0.9901, Fig. 3A) with an εC 276
value of -6.9 ± 0.4‰ (95% C.I., n=16). 277
The similar isotope fractionation of 1,1,2-TCA for the microcosms amended 278
with either acetate or pyruvate agrees with recent studies investigating isotopic 279
fractionation of trichloroethene (TCE) under different growth conditions. Harding et 280
al. (2013) showed that carbon isotope fractionation during TCE degradation by 281

13
Dehalococcoides-containing cultures remained consistent despite a variety of 282
temperature, nutrient, and cofactor-limiting conditions investigated. In addition, 283
Buchner et al. (2015) studied the potential effects of metabolic adaptation on carbon 284
and chlorine isotope fractionation of TCE during biodegradation by Desulfitobacterium 285
hafniesne Y51. These authors reported similar bulk values for C and Cl isotopes under 286
different growth conditions (i.e., cultures pre-grown with fumarate or TCE as electron 287
acceptors) and enzyme quantity per cell and suggested that isotope fractionation was not 288
affected. 289
3.1.3. Carbon isotope pattern of VC. In parallel to 1,1,2-TCA transformation, the δ13C 290
of its degradation product (i.e., VC) was monitored. The δ13C of VC was initially 291
depleted in 13C, in agreement with the normal isotope effect of 1,1,2-TCA, and shifted 292
toward more positive values during the course of reaction reaching the initial value of 293
1,1,2-TCA once this was completely degraded (Fig. 2). As observed for 1,1,2-TCA, the 294
carbon isotope data of VC from the experiments with acetate and pyruvate showed 295
similar values (Fig. 2). This figure also shows that δ13C values of VC fitted very well 296
with the expected product isotope trend determined according to eq. 3. The closed 297
isotopic mass balance confirmed the absence of other relevant degradation products. 298
Moreover, δ13C of VC never overpass the initial δ13C of 1,1,2-TCA suggesting that VC 299
is not further degraded to non-chlorinated compounds such as ethene or ethane, which is 300
consistent to its accumulation. 301
A different product isotope pattern was observed for degradation of 1,1,2-TCA 302
in a previous study with microcosms constructed with aquifer material and groundwater 303
(Hunkeler et al., 2002). These authors observed δ13C values of VC very enriched in 13C 304
compared to those of 1,1,2-TCA towards the end of reaction, which was indicative of 305
further degradation of VC to ethene via reductive dechlorination. Therefore, the results 306

14
of the present study and Hunkeler et al., 2002 illustrate the potential of the product 307
carbon isotope pattern to investigate the fate of VC in sites impacted with 1,1,2-TCA. 308
Analysis of ethene concentration can be used to evaluate the fate of VC in groundwater, 309
provided that other potential precursors of ethene such as 1,2-DCA are not present at the 310
site. However, assessing the fate of VC based solely on ethene concentration can be 311
difficult because ethene can be transformed under both oxic and anoxic conditions to 312
carbon dioxide and ethane, respectively (Mundle et al., 2012), highlighting the benefit 313
of VC isotope analysis as complementary data. 314
3.1.4. Chlorine isotope pattern of 1,1,2-TCA and dual C-Cl isotope approach. 315
Chlorine isotope data of 1,1,2-TCA (δ37Cl) were obtained from the pyruvate amended 316
microcosms. The δ37Cl of 1,1,2-TCA in the abiotic controls (-0.88 ± 0.2‰) did not 317
change significantly during the experiment, while an enrichment in the heavy isotope 318
(37Cl) during 1,1,2-TCA degradation following a Rayleigh trend (εCl = -2.7 ± 0.3‰, 319
n=8, 95% C.I., Fig. 3B) was observed in the cultures. Chlorine isotope fractionation was 320
much lower than for carbon, in agreement with the large primary carbon isotope effects 321
expected for C-Cl bond cleavage (Elsner et al., 2005). The measurement of chlorine 322
isotope ratios enabled for the first time a dual C-Cl isotope approach for biodegradation 323
of 1,1,2-TCA. A very good linear correlation (r2 = 0.994) was obtained when δ13C and 324
δ37Cl were combined in a dual element isotope plot showing a slope (Λ) of 2.5 ± 0.2 325
(95% C.I., Fig. 4). 326
A recent study on 1,2-DCA showed different Λ values during 327
dichloroelimination by Dehalogenimonas- and Dehalococcoides-containing cultures, 328
suggesting that a dual C-Cl isotope approach could help to identify the microbial taxa 329
responsible for anaerobic biodegradation of 1,2-DCA in the field (Palau et al., 2017). 330
This information is particularly important for 1,1,2-TCA given that, in contrast to 331

15
Desulfitobacterium (Zhao et al., 2015) (Fig. 1), its degradation by Dehalogenimonas 332
can result in an accumulation of the highly toxic VC in groundwater. Therefore, 333
comparison of the Λ value obtained for Dehalogenimonas in the present study with 334
those obtained for 1,1,2-TCA degradation by other bacteria in future studies might help 335
to investigate the fate of 1,1,2-TCA and to predict potential accumulation of VC in 336
contaminated sites. 337
3.2. Isotope effects and insight into dichloroelimination mechanisms of 1,1,2-TCA 338
Significant variation on reported bulk carbon isotope fractionation during 339
biodegradation of 1,1,2-TCA is observed (Table 1). The εC value of -6.9 ± 0.4‰ 340
determined in this study is significantly larger than that previously reported, -2.0 ± 341
0.2‰ from microcosms constructed with anaerobic aquifer material and groundwater 342
(Hunkeler et al., 2002). In addition, a much lower εC value of -0.7 ± 0.1‰ was 343
determined by Patterson et al. 2016, which was attributed to biodegradation in a 344
laboratory column consisted of both zero valent iron Fe(0) and biodegradable organic 345
carbon. Interestingly, microbiological data from this laboratory column suggested that a 346
co-culture composed by Desulfitobacterium and Dehalococcoides was responsible for 347
the sequential degradation of 1,1,2-TCA to ethene. The enzymatic mechanism of 348
Desulfitobacterium sp. strain PR to transform 1,1,2-TCA to 1,2-DCA via 349
hydrogenolysis differs from the production of VC via dichloroelimination in our 350
Dehalogenimonas-containing culture which could explain the difference on carbon 351
isotope fractionation observed in both studies (Fig. 1). A simultaneous cleavage of two 352
C-Cl bonds via concerted dichloroelimination of 1,1,2-TCA might result theoretically in 353
a larger bulk εC value compared to hydrogenolysis, where a single C-Cl bond is broken 354
at the initial reaction step. However, the occurrence of isotope-masking leading to 355
smaller εC values cannot be excluded. In this case, if preceding (rate-limiting) steps 356

16
exhibit small or no isotope fractionation, the observable isotope effect will be smaller 357
(i.e., masked) than the intrinsic isotope effect. 358
To address in more detail whether dichloroelimination of 1,1,2-TCA by 359
Dehalogenimonas proceeds via a stepwise or concerted mode, AKIE values were 360
calculated according to eq. 4 and 5 as it was previously done with the same 361
Dehalogenimonas containing enrichment for 1,2-DCP (Martín-González et al., 2015) 362
or 1,2-DCA (Palau et al., 2017, see also Table 1). Assuming stepwise or concerted 363
mode, carbon AKIEs obtained for 1,1,2-TCA (AKIECstepwise = 1.0138 ± 0.0008 and 364
AKIECconcerted = 1.0069 ± 0.0004, respectively) were much below the Streitweiser limit 365
of KIEC for complete C-Cl bond cleavage (1.057) and the realistic value of 50% bond 366
cleavage (1.029) (Elsner et al., 2005), making both modes feasible, but showing 367
important masking of intrinsic isotope fractionation. For chlorine, AKIEs determined 368
for both mechanisms (AKIEClstepwise = 1.0082 ± 0.0009 and AKIECl
concerted = 1.0041 ± 369
0.0005), were also below the Streitweiser limit for C-Cl bond cleavage (1.013). 370
Apart from theoretical Streitweiser limits, isotopic fractionation values and 371
derived AKIEs from abiotic reactions are often considered closest to the intrinsic 372
isotope effects. Abiotic reductive dechlorination of 1,1,2-TCA was suggested in the 373
same above-mentioned Fe(0) column study but without the organic carbon amendment 374
(Patterson et al. 2016). In that case, an AKIE for stepwise mode of 1.0246 can be 375
calculated from the reported εC value (−12 ± 5‰). This AKIE value is within the range 376
(AKIECstepwise = 1.0158 to 1.0326) previously available for abiotic reductive 377
dechlorination of 1,1,1-TCA and other polychlorinated ethanes, 1,1,2,2-378
tetrachloroethane (1,1,2,2-TeCA), pentachloroethane (PCA) and hexachloroethane 379
(HCA) by Cr(II), Fe(0) and Cu and Fe mixtures (Elsner et al., 2007; Hofstetter et al., 380
2007; Palau et al., 2014). Chlorine isotope effects (AKIEClstepwise = 1.0125 to 1.0207) 381

17
were also reported by Hofstetter et al., 2007 and Palau et al., 2014. The reported 382
carbon and chlorine AKIEs for abiotic reductive dechlorination of chlorinated ethanes 383
(via single C-Cl bond cleavage at the first reaction step) are higher than those 384
determined for 1,1,2-TCA dichloroelimination in this study assuming either stepwise or 385
concerted scenarios. Therefore, mechanistic interpretations are challenged by the 386
relatively low observable bulk isotope effects of 1,1,2-TCA. The occurrence of isotope-387
masking effects can sometimes complicate the identification of the underlaying reaction 388
mechanism since derived AKIEs may then be no longer characteristic of a certain 389
reaction (Elsner et al., 2005). However, an improved interpretation might be possible 390
by comparing dual C-Cl isotope slopes (see below). 391
A large isotope fractionation masking such that of 1,1,2-TCA during degradation 392
by Dehalogenimonas in this study was also observed for Dehalobacter-containing 393
mixed culture degrading 1,1,1-TCA versus 1,1-DCA (Sherwood Lollar et al., 2010). In 394
particular, the large intrinsic kinetic isotope effect expected for cleavage of a C-Cl bond 395
was almost completely masked during 1,1,1-TCA biodegradation by both whole cells 396
and cell-free extracts, while for 1,1-DCA the reduction was only roughly 50%. These 397
effects were not attributable to transport effects across the cell membrane, rather than to 398
significant differences in the kinetics of the enzymes catalyzing chlorinated ethane 399
degradation. 400
3.3. Reaction mechanism insight from dual C-Cl plot. 401
An important advantage of values compared to bulk values (and derived AKIEs) is 402
that the magnitude of the latter can be significantly affected by isotope-masking 403
processes. Since isotope-masking affect both elements to a similar extent, the dual 404
element isotope slopes remain largely unaltered (Elsner, 2010). For 1,1,2-TCA, the lack 405
of degradation studies including both carbon and chlorine isotope data makes not 406

18
possible a comparison of the value determined for Dehalogenimonas in this study 407
with values for different reactions (biotic and abiotic) and microbial strains. However, 408
values for a new compound like 1,1,2-TCA can be predicted based on the expected 409
KIEs for carbon and chlorine according to eq 6, and it can be then compared to the 410
experimentally determined value for Dehalogenimonas. 411
Assuming concerted dichloroelimination of 1,1,2-TCA (n = x = 2, z = 1 and n = 412
3, x = 2, z = 1 for C and Cl, respectively, see above), the carbon and chlorine isotope 413
effects determined in a recent study (Palau et al., 2017) for reductive 414
dichloroelimination of 1,2-DCA by Dehalogenimonas were used in eq. 6 (AKIECconcerted 415
= 1.024 ± 0.003 and AKIEClconcerted = 1.0121 ± 0.0008, see Table 1). These authors 416
postulated a concerted character of the reaction based on determined carbon isotope 417
effects. As a result, a value of 2.98 was obtained, which is similar to the experimental 418
value of 2.5 ± 0.2 (Fig. 4). In contrast, if a stepwise dichloroelimination of 1,1,2-TCA is 419
assumed (n = x = z = 2 and n = x = z = 3 for C and Cl, respectively), a very different 420
value of 1.01 is obtained. In this case, the average carbon and chlorine isotope effects 421
for 1,1,2,2-TeCA, PCA and HCA during abiotic dichloroelimination by Cr(II) via 422
sequential -elimination of two chlorine atoms were considered (AKIECstepwise = 1.026 ± 423
0.005 and AKIEClstepwise = 1.017 ± 0.004, see Table 1) (Hofstetter et al., 2007). In 424
addition, a smaller value of 0.66 was obtained in case the AKIEs estimated for 425
reduction of 1,1,1-TCA by Fe(0) via single electron transfer are used in the calculations 426
(AKIECstepwise = 1.0158 ± 0.0008 and AKIECl
stepwise = 1.0160 ± 0.0006, see Table 1). 427
Therefore, the comparison of the experimental value of 1,1,2-TCA with those 428
expected for stepwise and concerted mechanisms according to eq. 6 suggests that a 429
concerted dichloroelimination is more likely, highlighting the benefit of using a dual 430
C−Cl isotope approach. This result is in agreement with previous studies of 1,2-DCP 431

19
and 1,2-DCA biodegradation by Dehalogenimonas suggesting a concerted 432
dichloroelimination pathway (Martín-González et al., 2015; Palau et al., 2017). 433
Identification of the underlying transformation mechanism controlling isotope 434
fractionation can be valuable information to improve the characterization of reductive 435
dehalogenases. In addition, an eventual identification of different dichloroelimination 436
mechanisms of 1,1,2-TCA (i.e., concerted vs stepwise) by distinct microbial strains 437
might indicate the existence of diverse reductive dehalogenases with similar function 438
but likely different structure. For 1,2-DCA, the isotopic differences observed by Palau 439
et al. (2017) between Dehalogenimonas and Dehalococcoides containing cultures on 440
the concerted dichloroelimination mechanism were associated to a distinct interaction 441
mode between cobalamin dependent enzymes rather than two different reaction 442
pathways (i.e., stepwise vs concerted). The same isotopic results and conclusions were 443
validated by Franke et al. (2017) with two pure Dehalococcoides mccartyi strains (195 444
and BTF08). 445
446
4. Conclusions 447
1,1,2-TCA is a frequent groundwater contaminant but surprisingly only few studies 448
applying CSIA have been reported so far. Our work provides the first application of 449
dual isotope fractionation to investigate the anaerobic biodegradation of 1,1,2-TCA. The 450
stable isotope data obtained in this study during the dichloroelimination of 1,1,2-TCA 451
can be potentially helpful in monitoring the fate of this pollutant in contaminated 452
environments. In addition, the carbon isotope pattern of VC obtained in our enrichment 453
enlightens its potential use to identify the dominant VC production mechanism and 454
predict further transformation of this toxic compound. The single element kinetic 455
isotope effects could not provide conclusive information about the reaction mechanism 456

20
involved in 1,1,2-TCA dichloroelimination (concerted or stepwise); however, the dual-457
element approach can reduce interpretation bias due to isotope-masking effects 458
overcoming this limitation and pointing to more likely concerted mechanism. Further 459
investigations on carbon and chlorine isotope fractionation with bacteria catalyzing 460
alternate degradation pathways (i.e., hydrogenolysis) will allow the comparison 461
between microbial dechlorination reactions of 1,1,2-TCA. 462
463
5. Acknowledgements 464
This research has been supported by the Spanish Ministry of Economy and 465
Competitiveness, MINECO (CTM2013-48545-C2-1-R, CTM2016-75587-C2-1-R, 466
CGL2014-57215-C4-1-R and CGL2017-87216-C4-1-R projects) co-financed by the 467
European Union through the European Regional Development Fund (ERDF). This work 468
was also partly supported by the Generalitat de Catalunya through the consolidate 469
research groups (2017-SGR-14 and 2017SGR-1733). M. Rosell acknowledges a Ramón 470
y Cajal contract (RYC-2012-11920) from MINECO and S.H. Mortan support from the 471
Ministry of Education Malaysia (SLAI-UMP Scholarship) for a predoctoral fellowship. 472
The Departament d’Enginyeria Química, Biològica i Ambiental of the Universitat 473
Autònoma de Barcelona is a member of the Xarxa de Referència en Biotecnologia de la 474
Generalitat de Catalunya. We thank technical support from CCiT-UB. 475
476
477

21
Figure 1. Degradation pathways of 1,1,2-TCA: (a) dehydrochlorination, (b) 478
hydrogenolysis, (c) dichloroelimination, (d) hydrolysis. Numbers indicate the 479
dechlorinating agent: (1) base mediated abiotic reaction in aqueous solution (Pagana et 480
al. 1998); (2) Desulfitobacterium sp. strain PR (Zhao et al. 2015); (3) Dehalobacter and 481
Dehalogenimonas spp (Grostern and Edwards, 2006; Mortan et al. 2017, Yan et al. 482
2009); (4) nanosized zero-valent iron (Song and Carraway, 2005); (5) zero valent iron 483
and zinc (Patterson et al. 2016); (6) Pseudomonas sp. (Castro and Belser, 1990). Bold 484
arrow: biotic reaction; dashed arrow: abiotic reaction. 485
486
487
488
22
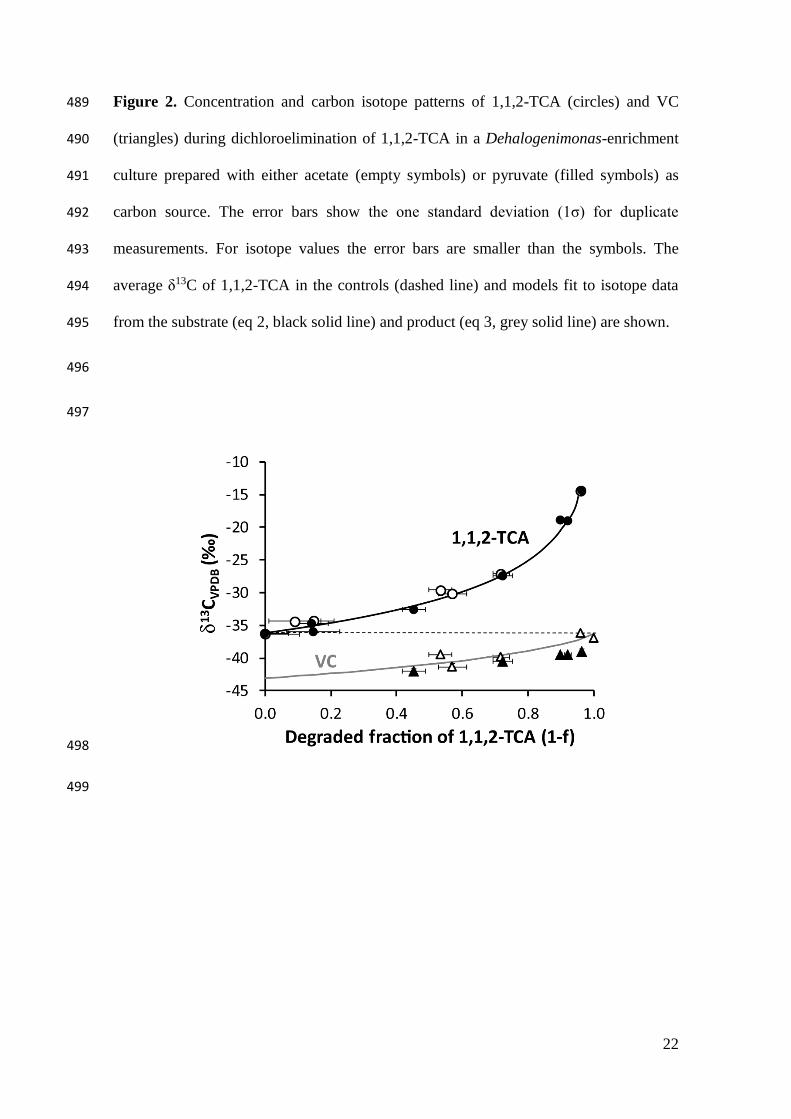
Figure 2. Concentration and carbon isotope patterns of 1,1,2-TCA (circles) and VC 489
(triangles) during dichloroelimination of 1,1,2-TCA in a Dehalogenimonas-enrichment 490
culture prepared with either acetate (empty symbols) or pyruvate (filled symbols) as 491
carbon source. The error bars show the one standard deviation (1σ) for duplicate 492
measurements. For isotope values the error bars are smaller than the symbols. The 493
average δ13C of 1,1,2-TCA in the controls (dashed line) and models fit to isotope data 494
from the substrate (eq 2, black solid line) and product (eq 3, grey solid line) are shown. 495
496
497
498
499
23
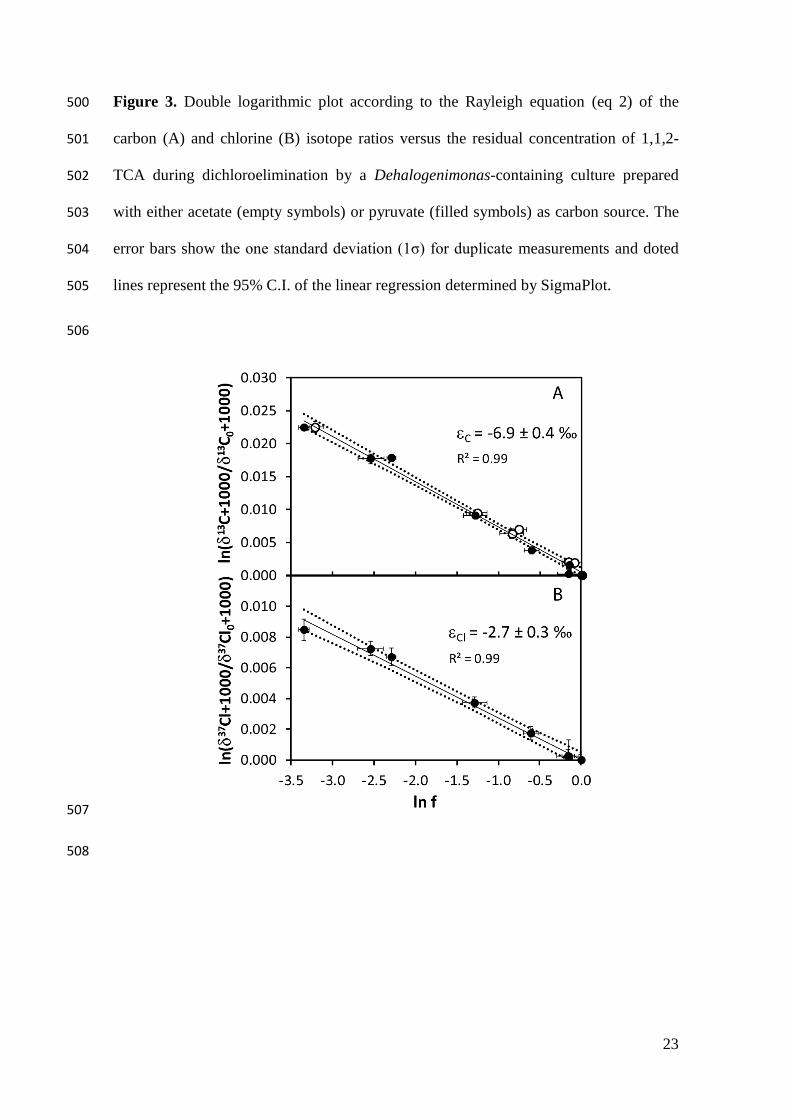
Figure 3. Double logarithmic plot according to the Rayleigh equation (eq 2) of the 500
carbon (A) and chlorine (B) isotope ratios versus the residual concentration of 1,1,2-501
TCA during dichloroelimination by a Dehalogenimonas-containing culture prepared 502
with either acetate (empty symbols) or pyruvate (filled symbols) as carbon source. The 503
error bars show the one standard deviation (1σ) for duplicate measurements and doted 504
lines represent the 95% C.I. of the linear regression determined by SigmaPlot. 505
506
507
508
24
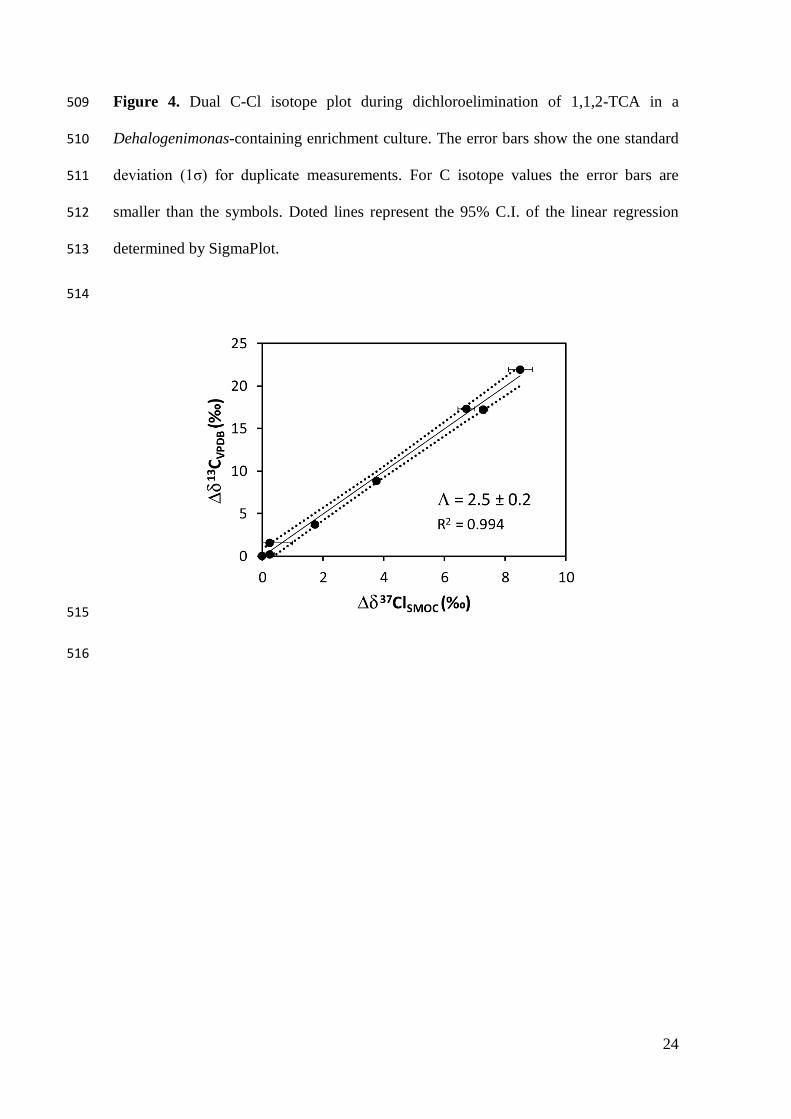
Figure 4. Dual C-Cl isotope plot during dichloroelimination of 1,1,2-TCA in a 509
Dehalogenimonas-containing enrichment culture. The error bars show the one standard 510
deviation (1σ) for duplicate measurements. For C isotope values the error bars are 511
smaller than the symbols. Doted lines represent the 95% C.I. of the linear regression 512
determined by SigmaPlot. 513
514
515
516
25
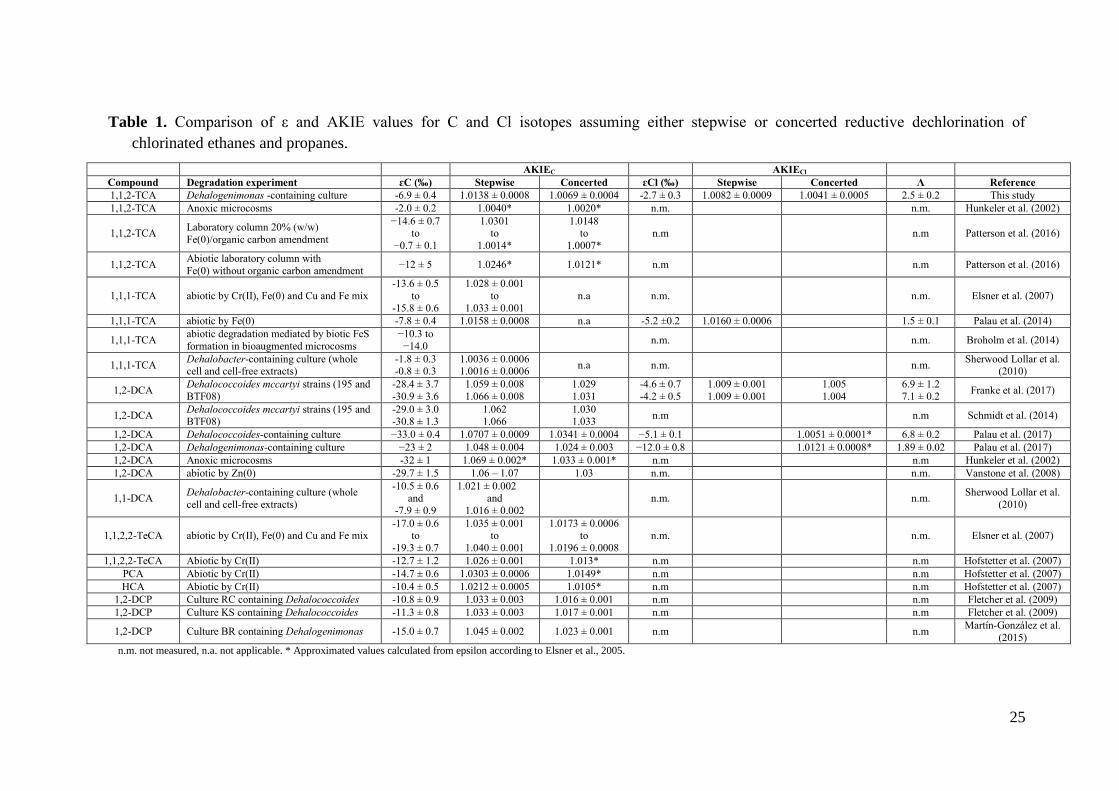
Table 1. Comparison of ε and AKIE values for C and Cl isotopes assuming either stepwise or concerted reductive dechlorination of
chlorinated ethanes and propanes.
AKIEC AKIECl
Compound Degradation experiment ɛC (‰) Stepwise Concerted ɛCl (‰) Stepwise Concerted Λ Reference
1,1,2-TCA Dehalogenimonas -containing culture -6.9 ± 0.4 1.0138 ± 0.0008 1.0069 ± 0.0004 -2.7 ± 0.3 1.0082 ± 0.0009 1.0041 ± 0.0005 2.5 ± 0.2 This study
1,1,2-TCA Anoxic microcosms -2.0 ± 0.2 1.0040* 1.0020* n.m. n.m. Hunkeler et al. (2002)
1,1,2-TCA Laboratory column 20% (w/w)
Fe(0)/organic carbon amendment
−14.6 ± 0.7 to
−0.7 ± 0.1
1.0301 to
1.0014*
1.0148 to
1.0007*
n.m n.m Patterson et al. (2016)
1,1,2-TCA Abiotic laboratory column with
Fe(0) without organic carbon amendment −12 ± 5 1.0246* 1.0121* n.m n.m Patterson et al. (2016)
1,1,1-TCA abiotic by Cr(II), Fe(0) and Cu and Fe mix
-13.6 ± 0.5
to
-15.8 ± 0.6
1.028 ± 0.001
to
1.033 ± 0.001
n.a n.m. n.m. Elsner et al. (2007)
1,1,1-TCA abiotic by Fe(0) -7.8 ± 0.4 1.0158 ± 0.0008 n.a -5.2 ±0.2 1.0160 ± 0.0006 1.5 ± 0.1 Palau et al. (2014)
1,1,1-TCA abiotic degradation mediated by biotic FeS
formation in bioaugmented microcosms
−10.3 to
−14.0 n.m. n.m. Broholm et al. (2014)
1,1,1-TCA Dehalobacter-containing culture (whole cell and cell-free extracts)
-1.8 ± 0.3 -0.8 ± 0.3
1.0036 ± 0.0006 1.0016 ± 0.0006
n.a n.m. n.m. Sherwood Lollar et al.
(2010)
1,2-DCA Dehalococcoides mccartyi strains (195 and
BTF08)
-28.4 ± 3.7
-30.9 ± 3.6
1.059 ± 0.008
1.066 ± 0.008
1.029
1.031
-4.6 ± 0.7
-4.2 ± 0.5
1.009 ± 0.001
1.009 ± 0.001
1.005
1.004
6.9 ± 1.2
7.1 ± 0.2 Franke et al. (2017)
1,2-DCA Dehalococcoides mccartyi strains (195 and
BTF08) -29.0 ± 3.0
-30.8 ± 1.3
1.062
1.066
1.030
1.033 n.m n.m Schmidt et al. (2014)
1,2-DCA Dehalococcoides-containing culture −33.0 ± 0.4 1.0707 ± 0.0009 1.0341 ± 0.0004 −5.1 ± 0.1 1.0051 ± 0.0001* 6.8 ± 0.2 Palau et al. (2017)
1,2-DCA Dehalogenimonas-containing culture −23 ± 2 1.048 ± 0.004 1.024 ± 0.003 −12.0 ± 0.8 1.0121 ± 0.0008* 1.89 ± 0.02 Palau et al. (2017)
1,2-DCA Anoxic microcosms -32 ± 1 1.069 ± 0.002* 1.033 ± 0.001* n.m n.m Hunkeler et al. (2002)
1,2-DCA abiotic by Zn(0) -29.7 ± 1.5 1.06 – 1.07 1.03 n.m. n.m. Vanstone et al. (2008)
1,1-DCA Dehalobacter-containing culture (whole
cell and cell-free extracts)
-10.5 ± 0.6 and
-7.9 ± 0.9
1.021 ± 0.002 and
1.016 ± 0.002
n.m. n.m. Sherwood Lollar et al.
(2010)
1,1,2,2-TeCA abiotic by Cr(II), Fe(0) and Cu and Fe mix -17.0 ± 0.6
to
-19.3 ± 0.7
1.035 ± 0.001 to
1.040 ± 0.001
1.0173 ± 0.0006 to
1.0196 ± 0.0008
n.m. n.m. Elsner et al. (2007)
1,1,2,2-TeCA Abiotic by Cr(II) -12.7 ± 1.2 1.026 ± 0.001 1.013* n.m n.m Hofstetter et al. (2007)
PCA Abiotic by Cr(II) -14.7 ± 0.6 1.0303 ± 0.0006 1.0149* n.m n.m Hofstetter et al. (2007)
HCA Abiotic by Cr(II) -10.4 ± 0.5 1.0212 ± 0.0005 1.0105* n.m n.m Hofstetter et al. (2007)
1,2-DCP Culture RC containing Dehalococcoides -10.8 ± 0.9 1.033 ± 0.003 1.016 ± 0.001 n.m n.m Fletcher et al. (2009)
1,2-DCP Culture KS containing Dehalococcoides -11.3 ± 0.8 1.033 ± 0.003 1.017 ± 0.001 n.m n.m Fletcher et al. (2009)
1,2-DCP Culture BR containing Dehalogenimonas -15.0 ± 0.7 1.045 ± 0.002 1.023 ± 0.001 n.m n.m Martín-Gonzalez et al.
(2015)
n.m. not measured, n.a. not applicable. * Approximated values calculated from epsilon according to Elsner et al., 2005.

26
References
Adrian, L., Szewzyk, U., Wecke, J., Görisch, H., 2000. Bacterial dehalorespiration with
chlorinated benzenes. Nature 408, 580-583. DOI:10.1038/35046063
ATDSR, 1989. Toxicological Profile for 1,1,2-trichloroethane, Agency for Toxic
Substances and Disease Registry. U.S. Department of Health and Human
Services,Public Health Service., Atlanta, GA.
ATSDR, Agency for Toxic Substances and Disease Registry. 2015 Substance Priority
List Resource Page. https://www.atsdr.cdc.gov/spl/resources/index.html (accessed on
23.05.2018).
Broholm, M. M., Hunkeler, D., Tuxen, N., Jeannottat, S., Scheutz, C., 2014. Stable
carbon isotope analysis to distinguish biotic and abiotic degradation of 1,1,1-
trichloroethane in groundwater sediments. Chemosphere 108, 265−273. DOI:
10.1016/j.chemosphere.2014.01.051
Buchner, D., Behrens, S., Laskov, C., Haderlein, S. B., 2015. Resiliency of stable
isotope fractionation (13C and 37Cl) of trichloroethene to bacterial growth physiology
and expression of key enzymes. Environ. Sci. Technol. 49, 13230-13237. DOI:
10.1021/acs.est.5b02918.
Castro, E., Belser, N.O., 1990. Biodehalogenation: Oxidative and reductive metabolism
of 1,1,2-trichloroethane by Pseudomonas Putida—Biogeneration of vinyl chloride.
Environ. Toxicol. Chem. 9, 707-714. DOI: https://doi.org/10.1002/etc.5620090603

27
Cretnik, S., Thoreson, K.A., Bernstein, A., Ebert, K., Buchner, D., Laskov, C.,
Haderlein, S., Shouakar-Stash, O., Kliegman, S., McNeill, K,, Elsner, M., 2013.
Reductive dechlorination of TCE by chemical model systems in comparison to
dehalogenating bacteria: insights from dual element isotope analysis (C-13/C-12, Cl-
37/Cl-35). Environ. Sci. Technol. 47, 6855-6863. DOI: 10.1021/es400107n
Cretnik, S., Bernstein, A., Shouakar-Stash, O., Loffler, F., Elsner, M., 2014. Chlorine
isotope effects from isotope ratio mass spectrometry suggest intramolecular C-Cl bond
competition in trichloroethene (TCE) reductive dehalogenation. Molecules 19, 6450-
6473. DOI: 10.3390/molecules19056450.
Elsner, M., Zwank, L., Hunkeler, D., Schwarzenbach, R.P., 2005. A new concept
linking observable stable isotope fractionation to transformation pathways of organic
pollutants. Environ. Sci. Technol. 39, 6896-6916. DOI: 10.1021/es0504587
Elsner, M., Cwiertny, D.M., Roberts, A.L., Lollar, B.S., 2007. 1,1,2,2-tetrachloroethane
reactions with OH-, Cr(II), granular iron, and a copper-iron bimetal: Insights from
product formation and associated carbon isotope fractionation. Environ. Sci. Technol.
41, 4111-4117. DOI: 10.1021/es063040x
Elsner, M., Hunkeler, D., 2008. Evaluating chlorine isotope effects from isotope ratios
and mass spectra of polychlorinated molecules. Anal. Chem. 80, 4731-4740. DOI:
10.1021/ac702543y
Elsner, M., 2010. Stable isotope fractionation to investigate natural transformation
mechanisms of organic contaminants: principles, prospects and limitations. J. Environ.
Monit. 12, 2005-2031. DOI: 10.1039/c0em00277a

28
Fletcher, K.E., Loffler, F.E., Richnow, H.H., Nijenhuis, I., 2009. Stable carbon isotope
fractionation of 1,2-dichloropropane during dichloroelimination by Dehalococcoides
populations. Environ. Sci. Technol. 43, 6915−6919. DOI: 10.1021/es900365x
Franke, S., Lihl, C., Renpenning, J., Elsner, M., Nijenhuis, I., 2017. Triple-element
compound-specific stable isotope analysis of 1,2-dichloroethane for characterization of
the underlying dehalogenation reaction in two Dehalococcoides mccartyi strains. FEMS
Microbiol. Ecol. 93, fix137. DOI: https://doi.org/10.1093/femsec/fix137
Frascari, D., Fraraccio, S., Nocentini, M., Pinelli, D., 2013. Aerobic/anaerobic/aerobic
sequenced biodegradation of a mixture of chlorinated ethenes, ethanes and methanes in
batch bioreactors. Bioresour Technol. 128, 479-486. DOI:
https://doi.org/10.1016/j.biortech.2012.10.026
Frascari, D., Pinelli, D., Nocentini, M., Baleani, E., Cappelletti, M., Fedi, S., 2008. A
kinetic study of chlorinated solvent cometabolic biodegradation by propane-grown
Rhodococcus sp PB1. Biochem. Engin. J. 42, 139-147. DOI:
https://doi.org/10.1016/j.bej.2008.06.011
Frascari, D., Pinelli, D., Nocentini, M., Fedi, S., Pii, Y., Zannoni, D., 2006. Chloroform
degradation by butane-grown cells of Rhodococcus aetherovorans BCP1. Appl.
Microbiol. Biotechnol. 73, 421-428. DOI: 10.1007/s00253-006-0433-3.
Grostern A., Edwards EA. 2006. Growth of Dehalobacter and Dehalococcoides spp.
during degradation of chlorinated ethanes. Appl. Environ. Microbiol. 72:428-436. DOI:
10.1128/AEM.72.1.428-436.2006
Harding, K.C., Lee, P.K., Bill, M., Buscheck, T.E., Conrad, M.E., Alvarez-Cohen, L.,
2013. Effects of varying growth conditions on stable carbon isotope fractionation of

29
trichloroethene (TCE) by tceA-containing Dehalococcoides mccartyi strains. Environ.
Sci. Technol. 47, 12342-12350. DOI: 10.1021/es402617q
Hofstetter, T.B., Reddy, C.M., Heraty, L.J., Berg, M., Sturchio, N.C., 2007. Carbon and
chlorine isotope effects during abiotic reductive dechlorination of polychlorinated
ethanes. Environ. Sci. Technol. 41, 4662-4668. DOI: 10.1021/es0704028
Hunkeler, D., Aravena, R., Berry-Spark, K., Cox, E., 2005. Assessment of degradation
pathways in an aquifer with mixed chlorinated hydrocarbon contamination using stable
isotope analysis. Environ. Sci. Technol. 39, 5975-5981. DOI: 10.1021/es048464a
Hunkeler, D., Aravena, R., Cox, E., 2002. Carbon isotopes as a tool to evaluate the
origin and fate of vinyl chloride: laboratory experiments and modeling of isotope
evolution. Environ. Sci. Technol. 36, 3378-3384. DOI: 10.1021/es011479d
Kuntze, K., Kozell, A., Richnow, H.H., Halicz, L,, Nijenhuis, I., Gelman, F., 2016. Dual
carbon–bromine stable isotope analysis allows distinguishing transformation pathways
of ethylene dibromide. Environ. Sci. Technol. 50, 9855-9863. DOI:
10.1021/acs.est.6b01692
Leys, D., Adrian, L., Smidt, H., 2013. Organohalide respiration: microbes breathing
chlorinated molecules. Phil. Trans. R. Soc. B. 368 20120316. DOI:
10.1098/rstb.2012.0316
Martín-González, M., Mortan, S.H., Rosell, M., Parladé, E., Martínez-Alonso, M., Gaju,
N., Caminal, G., Adrian, L., Marco-Urrea, E., 2015. Stable carbon isotope fractionation
during 1,2-dichloropropane-to-propene transformation by an enrichment culture
containing Dehalogenimonas strains and a dcpA gene. Environ. Sci. Technol. 49, 8666-
8674. DOI: 10.1021/acs.est.5b00929

30
Moe, W.M., Rainey, F.A., Yan, J., 2016. The genus Dehalogenimonas, in: Adrian, L.,
Löffler, F.E. (Eds.), Organohalide-respiring bacteria. Springer-Verlag Berlin
Heidelberg. DOI 10.1007/978-3-662-49875-0_7
Mortan, S.H., Martín-González, L., Vicent, T., Caminal, G., Nijenhuis, I., Adrian, L.,
Marco-Urrea, E., 2017. Detoxification of 1,1,2-trichloroethane to ethene in a bioreactor
co-culture of Dehalogenimonas and Dehalococcoides mccartyi strains. J. Hazard.
Mater. 331, 218-225. DOI: 10.1016/j.jhazmat.2017.02.043
Mundle, S.O.C., Johnson, T., Lacrampe-Couloume, G., Perez-de-Mora, A., Duhamel,
M., Edwards, E.A., McMaster, M.L., Cox, E., Revesz, K., Lollar, B.S., 2012.
Monitoring biodegradation of ethene and bioremediation of chlorinated ethenes at a
contaminated site using compound-specific isotope analysis (CSIA). Environ. Sci.
Technol. 46, 1731-1738. DOI: 10.1021/es202792x
Němeček, J., Dolinová, I., Macháčková, J., Špánek, R., Ševců, A., Lederen, T., Černík,
M., 2017. Stratification of chlorinated ethenes natural attenuation in an alluvial aquifer
assessed by hydrochemical and biomolecular tools. Chemophere 184, 1157-1167. DOI:
https://doi.org/10.1016/j.chemosphere.2017.06.100
Pagana, M., Cooperab, W.J., Joensa, J.A., 1998. Kinetic studies of the homogeneous
abiotic reactions of several chlorinated aliphatic compounds in aqueous solution. Appl.
Geochem. 13, 779–785. DOI: https://doi.org/10.1016/S0883-2927(98)00005-5
Palau, J., Shouakar-Stash, O., Hunkeler, D., 2014. Carbon and chlorine isotope analysis
to identify abiotic degradation pathways of 1,1,1-trichloroethane. Environ. Sci. Technol.
48, 14400-14408. DOI: 10.1021/es504252z

31
Palau, J., Yu, R., Hatijah Mortan, S., Shouakar-Stash, O., Rosell, M., Freedman, D.L.,
Sbarbati, C., Fiorenza, S., Aravena, R., Marco-Urrea, E., Elsner, M., Soler, A.,
Hunkeler, D., 2017. Distinct dual C-Cl isotope fractionation patterns during anaerobic
biodegradation of 1,2-dichloroethane: potential to characterize microbial degradation in
the field. Environ Sci Technol. 51, 2685-2694. DOI: 10.1021/acs.est.6b04998
Pankow J.F., Cherry J.A. Dense chlorinated solvents and other DNAPLs in
groundwater: history, behavior, and remediation. Portland, OR: Waterloo Press; 1996.
Patterson, B.M., Lee, M., Bastow, T.P., Wilson, J.T., Donn, M.J., Furness, A.,
Goodwin, B., Manefield, M., 2016. Concentration effects on biotic and abiotic
processes in the removal of 1,1,2-trichloroethane and vinyl chloride using carbon-
amended ZVI. J. Contam. Hydrol. 188, 1-11. DOI: 10.1016/j.jconhyd.2016.02.004
Sherwood Lollar, B., Hirschorn, S., Mundle, S.O., Grostern, A., Edwards, E.A.,
Lacrampe-Couloume, G., 2010. Insights into enzyme kinetics of chloroethane
biodegradation using compound specific stable isotopes. Environ. Sci. Technol. 44,
7498–7503. DOI: 10.1021/es101330r
Schmidt, M., Lege, S., Nijenhuis, I., 2014. Comparison of 1,2-dichloroethane,
dichloroethene and vinyl chloride carbon stable isotope fractionation during
dechlorination by two Dehalococcoides strains. Water Res. 52, 146-154. DOI:
10.1016/j.watres.2013.12.042.
Scott, K.M., Lu, X., Cavanaugh, C.M., Liu, J.S., 2004. Optimal methods for estimating
kinetic isotope effects from different forms of the Rayleigh distillation equation.
Geochim. Cosmochim. Ac. 68, 433-442. DOI: https://doi.org/10.1016/S0016-
7037(03)00459-9

32
Song, H., Carraway, E.R., 2005. Reduction of chlorinated ethanes by nanosized zero-
valent iron: kinetics, pathways, and effects of reaction conditions. Environ. Sci.
Technol. 39, 6237-6245. DOI: 10.1021/es048262e
Vannelli, T., Logan, M., Arciero, D.M., Hopper, A.B., 1990. Degradation of
halogenated aliphatic compounds by the ammonia-oxidizing bacterium Nitrosomonas
europaea. Appl. Environ. Microbiol. 56, 1169-1171.
Vanstone, N., Elsner, M., Lacrampe-Couloume, G., Mabury, S., Sherwood Lollar, B.,
2008. Potential for identifying abiotic chloroalkane degradation mechanisms using
carbon isotopic fractionation. Environ. Sci. Technol. 42, 126–132. DOI:
10.1021/es0711819
Yan, J., Rash, B.A., Rainey, F.A., Moe, W.M., 2009. Isolation of novel bacteria within
the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ.
Microbiol. 11, 833-843. DOI: https://doi.org/10.1111/j.1462-2920.2008.01804.x
Zhao, S., Ding, C., He, J., 2015. Detoxification of 1,1,2-Trichloroethane to ethene by
Desulfitobacterium and identification of its functional reductase gene. PLoS ONE 10,
e0119507. DOI: https://doi.org/10.1371/journal.pone.0119507
Top Related