
Thesis Javier Infante Lourido
36
Rainbow trout (Oncorhynchus mykiss) monocytes in culture with cortisol: Expression of CD18, PU.1/Spi-1, TNF-α, IL-1β and Glucocorticoid Receptor (GR) Javier Infante Lourido September 2007 Tutors: Lluís Tort Simon Mackenzie Cellular biology, Physiology and Immunology Department Faculty of Biosciences Autonomous University of Barcelona
-
Upload
javierinfanteinfante -
Category
Documents
-
view
690 -
download
3
Transcript of Thesis Javier Infante Lourido

Rainbow trout (Oncorhynchus mykiss) monocytes in
culture with cortisol: Expression of CD18, PU.1/Spi-1,
TNF-α, IL-1β and Glucocorticoid Receptor (GR)
Javier Infante Lourido
September 2007
Tutors:
Lluís Tort
Simon Mackenzie
Cellular biology, Physiology and Immunology Department
Faculty of Biosciences
Autonomous University of Barcelona

1
INDEX
I. ABSTRACT................................................................................................................................2
1. INTRODUCTION.......................................................................................................................3
2. OBJECTIVES..........................................................................................................................10
3. MATERIAL AND METHODS..................................................................................................11
4. RESULTS AND DISSCUSION................................................................................................15
4.1 Cellular culture under different conditions with cortisol....................................15
4.2 CD18.........................................................................................................................17
4.3 Glucocorticoid receptor (GR)................................................................................19
4.4 Cytokines (TNF-αααα and IL-1ββββ)..................................................................................22
4.5 PU.1 (Spi-1)..............................................................................................................26
5. CONCLUSIONS......................................................................................................................28
6. REFERENCES........................................................................................................................30
ACKNOWLEDGMENTS............................................................................................................ 36

2
I. ABSTRACT
In this study primary cultures of rainbow trout monocytes/macrophages were treated
with the corticosteroid hormone, cortisol, simulating physiological concentrations during the
acute stress response. Cortisol at high concentrations (300ng/ml) affects the capacity of
monocytes to adhere to the culture plate surface therefore decreasing the number of
macrophages. Moreover, expression studies with one day culture monocytes show that cortisol
in high concentrations downregulates the expression of the inflammatory cytokines, TNF-α and
IL-1β and the transcription factor PU.1, which may indicate the activation and reprogramming of
homeostatic energy saving processes in the cultured cells. This may mimic a glucocorticiod-
induced acute stress response in vitro. However, the cellular response to conditions mimicking
chronic stress, 30ng/ml cortisol, is not observed indicating that such processes are only
activated at high cortisol concentrations. These results suggest that the induction of differential
regulatory mechanisms by cortisol in in vitro cultured monocytes/macrophages are
concentration and context-dependent (differentiation status of cells). This study provides the
basis of an in vitro cell culture model to investigate immune-endocrine interactions at the level of
the transcriptome.

3
1. INTRODUCTION
The characteristics of immune system of fishes are the results of the characteristics of
the environment were they live and the evolution of the immune system. The aquatic medium
allows easy transport and growth of microorganisms and this results in a high concentration of
prokaryotic cells in oceans, estimated in 1024 prokaryotic cells in oceans (Press and Evensen,
1999), and are very important in marine biomass, and viruses may account for 1010 in aquatic
habitats (Wilhelm and Suttle, 1999).
Fish immune system
Fish are the most diverse group in vertebrates, and are considered a crossroad
between the innate and adaptive immunity (Tort et al., 2003) inside a gradual transition from the
innate immune system of invertebrates to the recombinatorial immune system of higher
vertebrates (Khalturin et al., 2004).
Adaptive immunity requires an induction by the antigen, the differentiation of self from
non-self, the specificity towards the antigen and an immunological memory. A clonally diverse
anticipatory repertoire in which lymphocyte bears a unique antigen receptor is the central
feature of the adaptive immune response. All jawed vertebrates, assemble their antigen-
Fig 1: Evolution of inmune system (Tort et al., 2004).

4
receptor genes through recombinatorial rearrangement of different immunoglobulin or T cell
receptor gene segments (Cooper and Alder, 2006) and recently it has been discovered that
agnathans also have an adaptive immune system that is based on recombinatorial assembly of
a different type of modular genetic units to generate a highly diverse repertoire of lymphocytes,
each with a unique anticipatory receptor (Alder et al., 2005; Cooper and Alder, 2006; Pancer et
al., 2004). These facts suggest that lymphocyte progenitors evolved in the most basal
vertebrates or in a protochordate ancestor (Cooper and Alder, 2006).
Innate responses are induced by an antigen and differentiates the own from the
strange, witch allows to the innate immune system to detect the presence of an infectious
microbe, but also to determine the type of the infecting pathogen (Medzhitov and Janeway,
2000). The binding between TLRs (Toll-like receptors) and PAMPs (pathogen associated
molecular patterns) is the basis of this differentiation and triggers acute inflammation through
production of reactive oxygen and glucocorticoids and expression of cytokines and their
receptors and chemokines and their receptors (Stavitsky, 2007).
The majority of pluricellular organisms are able to maintain their integrity thanks to an
innate immune system based on cell phagocytosis and secretion of soluble antimicrobial
molecules (Tort et al., 2003). This innate immunity system is divided into physical and chemical
barriers, cellular and humoral components (Magnadottir, 2006). Physical barriers, like scales,
mucous surfaces of skin and the epidermis, are the first line of defence against pathogens
(Bernstein et al., 1996; Harding et al., 1990). Mucus possesses different substances including
antibacterial peptides, lysozyme, lectins and proteases (Ellis, 2001) and is very important as a
first barrier in trout (Haynes and McKinney, 1991; Rast and Litman, 1994) and in other teleost
species(McKinney et al., 1986; Pettey and McKinney, 1988; Walsh and Luer, 1998). Humoral
parameters include growth inhibitors, lytic enzymes, agglutinins, opsonins, interferon, lectins,
cytokines, chemokines, bacterial peptides and complement system, which is a multicomponent
system that involves 20-30 plasma proteins and several receptors (Magnadottir, 2006;
Magnadottir et al., 2005). Cellular components of innate immune system involve many cells,
including phagocytic cells, like granulocytes, mainly neutrophils, monocytes/macrophages, and
including B lymphocytes, which recently has been discovered that are capable of efficient
phagocytosis (Li et al., 2007).
The macrophages: origin and functions.
Macrophages perform pinocytosis and phagocytosis, and are cytotoxic against foreign
cells (McKinney et al., 1986; Walsh and Luer, 1998). They differentiate from monocytes
precursors after the expression of fms and L-plastin genes (Crowhurst et al., 2002). Another
important gene involved in the differentiation of myeloid cells is PU.1 (SPI-1), whose
concentration inside immunological stem cells determine the destination of this cells, that in

5
conditions of high concentration of the transcription factor of PU.1 turn into macrophages and in
low concentrations differentiate to B lymphocytes (DeKoter and Singh, 2000). In short,
macrophages are fundamental immune cells and are fundamental in defence against
pathogens, making diverse functions, like the phagocytosis, pathogen killing through respiratory
burst or antigen presentation (Secombes et al., 2001).
As regards to the origin of these immune cells, the thymus, kidney and spleen are the
major lymphoid organs in teleosts (Zapata et al., 2006). Rainbow trout, like the other teleost
species, do not have bone marrow or lymph nodes (Rombout et al., 2005; Zapata et al., 1995;
Zwollo et al., 2005). The head kidney (pronefros, anterior kidney) is the primary site for
haematopoiesis in teleosts, instead of bone marrow (Bromage et al., 2004; Fange, 1986;
Rombout et al., 2005; Zapata, 1979; Zwollo et al., 2005). The trunk kidney (opisthonephros,
posterior kidney) mainly consists in renal tissue (Zapata et al., 1995). Kidney is located in trout
ventrally across the backbone extending from the base of the cranium (head kidney) to the
caudal region (trunk kidney) (Wake, 1997).
In this organ takes place the multiplication and differentiation of hematopoietic stem
cells (Zapata et al., 1995). The head kidney has no renal function, does not have nephrons and
it is the place where hemopoiesis occurs. The hemopoietic tissue is interdigitated with adrenal-
Fig 2: Morphology and location of kidney.

6
like tissue, while trunk kidney posses renal and immune tissues (Grassi et al., 1997; Zapata,
1990). Furthermore, head kidney in teleosts has a system of macrophages that concentrate and
separate phagocytosed material, the reticuloendothelial system (Roberts, 1989). Spleen is
implicated in haematopoiesis, but its role is mainly limited to erythropoiesis and thrombopoiesis
(Rowley et al., 1999). In short, trout kidney is a complex organ that houses four different
functions, hemopoietic, reticuloendothelial, endocrine and excretory functions (Zapata, 1990).
Monocytopoiesis in teleosts usually occurs in the head kidney and/or spleen monocytes.
Cells are released to the blood stream and after arrival at the place of inflammation, increase
their phagocytic potential and enhance their antimicrobial activity (Reite and Evensen, 2006;
Rowley et al., 1999).
The mobilization of leucocytes from haematopoietic tissues to the place where the
antigenic particles are present is a very important episode in the inflammation process and is
essential for the development of a full immune response (Cuesta et al., 2006).
Inflammatory response.
Inflammation is a local response to protect the organism from pathogenic invasions, an
essential component of pathogen clearance, resolution of sepsis and repair of injury and many
diseases, and it is controlled by at least two different general mechanisms (Tracey, 2002)
During the more rapid, local, neural regulation, pro-inflammatory cytokines (IL-1β, IL-18, TNF-α)
are released. But inflammatory responses can be more dangerous than the original inciting
stimulus, so the activation is inhibited or limited by stimulation of the acetylcholine-cholinergic
pathway through the vagus nerve (Tracey, 2002). Anti-inflammatory cytokines, such as IL-10
and transforming growth factor-β (TGF-β) specifically inhibit the release of TNF and other
proinflammatory mediators. Glucocorticoids, adrenaline, α-melanocyte-stimulating hormone (α-
MSH) and other classical stress hormones inhibit cytokine synthesis and intracellular
transduction. The conjunction of the signals that arrive to the nervous system through sensory
systems is integrated in the brain and triggers appropriated responses, controlling inflammatory
mechanisms.
Adhesion molecules and migration process.
A very important success in immune is the migration of immune cells towards the
inflamed tissue, process in which adhesion molecules play a very important role. Inside this
group of molecules three fundamental classes are present: Selectins, Integrins and
immunoglobulin superfamily, and other like cadherins, hyaluronate receptors or the receptor
protein tyrosine phosphatases (Freemont and Hoyland, 1996).

7
Selectins are proteins that bind their intracellular domain to carbohydrates. There are
three major groups of selectins: L-selectin, E-selectin and P selectin, which are expressed
mainly in leucocytes, endothelial cells and platelets and endothelium respectively (Bevilacqua
and Nelson, 1993; Lasky, 1992).
Integrins are heterodimeric transmembrane glycoproteins consisting of non-covalently
associated α and β subunits and the distinctive in leucocytes is the expression of integrins
containing a β2 (CD18) chain paired with one of α chains (Harris et al., 2000). Immunoglobulin
superfamily is a large and diverse family of molecules that have one or more immunoglobulin-
like domains (Springer, 1990a) and include among others ICAMs (intercellular adhesion
molecules) VCAMs (vascular cell adhesion molecules), LFA-2, LFA-3 or MacCAM-1.
Adhesion molecules determine the leukocyte egress from blood to the extravascular
matrix, process that can be divided into four sequential steps which include adhesion and
signalling events (Pitzalis et al., 2002).
In the first step (the transient leukocyte adhesion) leukocytes leave the central stream
and start rolling along the vessel walls. The cell adhesion molecules (CAMs) that are involved in
this step are mainly the selectins. L-selectin is expressed mainly in leukocytes, whereas E and
P-selectin are expressed by endothelial cells and platelets, respectively. Usually, rolling cells
detach from the vessel walls under the influence of shear stress, but during inflammatory
reactions many more leukocytes arrest onto the endothelium (Atherton and Born, 1972). The
arrest of leukocytes onto epithelial cells is principally mediated by integrins, particularly β1 (VLA-
4) and β2 (LFA-1 and MAC-1) and their respective ligands in epithelial cells, VCAM-1 and ICAM-
1, ICAM-2 and ICAM-3 (Hogg, 1993; Springer, 1990b).
The second step in this process is the cell activation. Integrins are normally expressed
in a low-avidity and non functional state to prevent random adhesion in the circulation (Hogg
and Landis, 1993; Springer, 1990a) but can be activated by inflammatory mediators, particularly
proinflamatory chemokines (Baggiolini et al., 1997; Schall and Bacon, 1994).
Activated integrins can subsequently mediate the third step of leukocyte strong
adhesion to endothelial cells, forming bonds with specific endothelial receptors. Finally these
interactions make possible the fourth step, diapedesis, the transendothelial migration to the
inflamed tissues (Pitzalis et al., 2002), where macrophages perform their functions against the
pathogenic particles.

8
Immune responses to stress
Stress is a very important concept in physiology but is difficult to define and in fish has
been studied not only for theoretical interest, but also for the economic importance of intensive
fish culture. It could be defined as a real or symbolic (perception of a potential threat) state
produced by an environmental or other factor that extends the adaptive responses beyond its
normal physiological range (Brett, 1958).
From a physiologic point of view, a specific activation of stress system is useful to the
animal, because it allows restoring homeopathic balance of inner medium altered by a stressor.
The animal diverts energy from normal metabolism to adapt physiological systems to the
stressor. However, if the stress situation continues, the adaptive value could be lost and
originate problems to the individual. In these circumstances (maladaptation), stress could
originate problems in growth, reproduction, osmorregulation and immunity.
Stress produces a series of alterations, which are classified in three groups
(Wedemeyer and McLeay, 1981): Primary alterations, like the release of ACTH from
adenohypophysis or stress hormones (catecholamines and corticosteroids) from the interrenal.
Secondary alterations (physiological), like blood chemistry and hematological changes
(hyperglycemia, hyperlacticemia, hypochloremia, leucopenia, reduced blood clotting time),
tissue changes (depletion of liver glycogen, and inter-renal vitamin C), metabolic changes
(negative nitrogen balance and oxygen debt) or in the diuresis (blood electrolyte loss). Tertiary
effects affect to whole animal, like the impaired growth, parr-smolt transformation, spawning
success, migratory behaviour or increased disease incidence (Ottaviani and Franceschi, 1996).
Generally stress induces immunosuppresion; however different factors, like the nature
of the stressor and organism, type, intensity and duration of stress generate different
responses. Even so, it is clear that stressors of long or medium duration generate
immunosuppresion and maladaptation, especially if the stressor is chronic or repeated (Tort et
al., 2004).
The investigation of the evolutionary bases of stress argues that the basic mechanisms
and molecules involved in stress, like CRH, ACTH, glucocorticoids, biogenic amines and
cytokines are fundamentally similar and well preserved throughout evolution (Ottaviani and
Franceschi, 1996). One of the most important changes occurring in stress conditions is the
activation of hormonal axis, increasing plasma levels of cortisol (Herbert and Cohen, 1993).

9
The role of cortisol on immune response
The perception of a stressor activates the Central Nervous System (CNS), specifically in
the hypothalamus, where are the two mayor axis, the sympatico-chromafin axis, that innervates
chromafin cells in head kidney, involved in catecholamines synthesis, and the HPI
(Hypothalamus-Pituitary-Interrenal) axis, that through the synthesis of two hormones, CRH
(corticotrophin-releasing hormone) and TSH (Thyroid-releasing hormone) stimulates the
synthesis and liberation of cortisol in interrenal cells of head kidney.
Cortisol is the major stress steroid and plays multiple roles in organisms, like the
catabolic action of this glucocorticoid, that allow to the fish increases energetic availability to
maintain homeostasis (Schreck, 1981).
In short, cortisol causes multiple effects in the immune system (Table 1), but in seems
that cortisol could even act as an activator of the early immune response and thus it could help
to maintain key defence mechanisms, whereas other actions of cortisol at long term normally
would end in immunosuppression (Tort et al., 2004).
In this work we study the effects of cortisol on several indicators of the immune system,
particularly in adhesion and cellular differentiation of monocytic cells. To this purpose were
studied different molecules: the adhesion molecule CD-18, the transcription factor PU.1, cortisol
receptor (CR) and cytokines TNF-α and IL 1-β using the housekeeping gene 18S as reference.
Table 1: Effects of different stressors on immune indicators. Values are calculated as the reduction in percentage of each indicator compared to respective controls, which were set at 100%. PI = Phagocytic index. (Tort, 2004)

10
2. OBJECTIVES
The objective of this work is to determine the effect of cortisol in the expression of key
molecules, as the cytokines, adhesion proteins or transcription factors.
This general objective is split in the following experimental tasks:
1. Development of a primary cell culture to observe the behaviour of the macrophages
under diverse treatments with cortisol.
2. To determine the expression of the different genes of specific cytokines, the TNF-α and
Interleukin 1-β (IL 1-β) under conditions of high and low cortisol levels.
3. To determine the expression of the adhesion protein CD18 in conditions of high and low
cortisol concentration.
4. To determine the expression of the transcription factor PU.1 (spi1) under conditions of
high and low cortisol levels.

11
3. MATERIALS AND METHODS
2.1 Animals
Rainbow trout (Oncorhynchus mykiss) were obtained from the “Truchas Jose Andrés”
(Olot, Girona) and held in 150 litres tanks with recirculating freshwater at room temperature. The
tanks were provided with a biofiltration system, with oxygen, salinity, pH, temperature and
nitrogen compounds. The fish were subjected to 12 h light:dark cycle. The fish were fed with
commercial pellets provided by “Proaqua” at a daily ration of 0.5 % of the body weight. The fish
were at least 15 days under the above conditions prior experiments.
2.2 Cell culture
The fish were killed by overanesthetization in 2-phenoxy-ethanol and the head kidneys
were dissected and placed in sterile 200 µm cell strainer (BD Biosciences). The tissue was
squeezed through the cell strainer using sterile forceps into Dulbecco`s Modified Eagle Medium
(PAA laboratories) containing high glucose, 10% heat inactivated Foetal Bovine Serum (PAA
laboratories) and primocin (500ng, InvivoGen).
The samples were centrifuged for 1 minute at 1500 rpm (Hettich, D-78532 Tuttlingen),
and the pellet was taken and resuspended in DMEM medium with FBS and primocin. This
mixture of cells and medium were plated in 60mm of diameter plates (NUNCTM) and put in an
incubator (Sanyo MC0175), maintained at 15 ºC and 5% of CO2.
2.2.1 One day culture
After the cells were plated with medium with FBS and Primocin we added to each plate
cortisol, ethanol and/or RU-486 (cortisol antagonist) or none according to the condition in each
plate.
Conditions:
1. Control (nothing)
2. Ethanol (60µl in each plate).
3. Cortisol (30 ng/ml)
4. Cortisol (300 ng/ml)
5. Cortisol (300 ng/ml) + RU-486 (100ng/ml)
6. RU-486 (100ng/ml)
12
After stimulation the plates were stored during 24 hours and then start the RNA
extraction.
2.2.2 Five days culture
No adhering cells were removed at 24 hours and new medium was added and the cells
were incubated for an additional 4 days. After 48 hours no adhering cells were removed and the
medium was replaced.
At day 5 we took pictures of each culture condition to demonstrate the cortisol effects in
culture.
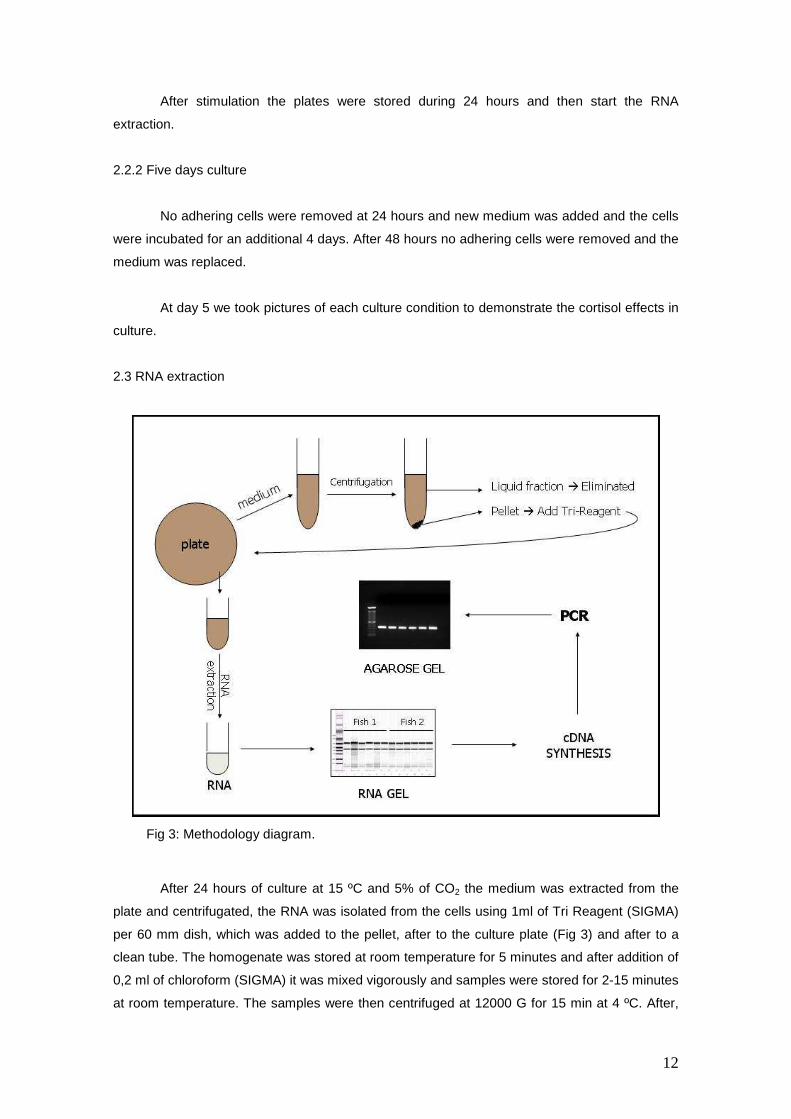
2.3 RNA extraction
After 24 hours of culture at 15 ºC and 5% of CO2 the medium was extracted from the
plate and centrifugated, the RNA was isolated from the cells using 1ml of Tri Reagent (SIGMA)
per 60 mm dish, which was added to the pellet, after to the culture plate (Fig 3) and after to a
clean tube. The homogenate was stored at room temperature for 5 minutes and after addition of
0,2 ml of chloroform (SIGMA) it was mixed vigorously and samples were stored for 2-15 minutes
at room temperature. The samples were then centrifuged at 12000 G for 15 min at 4 ºC. After,
Fig 3: Methodology diagram.

13
the aqueous phase was transferred to a clean tube, 0.5 ml of 2-propanol (SIGMA) was added
and mixed. The tubes were stored for 5-10 minutes at room temperature and centrifuged at
12000 G for 8 min (4-25 ºC). The pellet was washed with 1 ml of 75% ethanol and centrifuged at
7500 G for 5 min (4-25 ºC). The RNA pellet was air dried for 5-10 minutes and after, 10 µl of
DPC water was added. These samples were incubated at 55-60 ºC for 10 minutes to dissolve
the RNA pellet.
To measure the concentration of RNA and determine RNA quality a RNA gel was done.
2.4 Reverse transcription PCR
Total RNA from cells was reverse transcribed with SuperScript III reverse transcriptase
and an anchored oligo-dT primer (Biotools).
Reverse transcription was performed following the next protocol: Denaturation at 95 ºC
for 7 min followed by 30 cycles of: denaturation at 95 ºC for 1 min; annealing at 50 ºC for 1 min;
extension at 72 ºC for 1 min 30s. The final step was an extension at 72 ºC for 5 min. The cDNA
was stored al -20 ºC.
2.5 PCR reaction
The conditions for PCR were as follows: denaturation for 5 min at 94 ºC followed by 30
cycles of denaturation at 94 ºC for 45 s, annealing for 45 s and extension at 72 ºC for 30 s, with
a final extension at 72 ºC for 10 min.
The cDNA was then used to obtain partial cDNAs of different genes using Taq DNA
polymerase (Biotools). The genes used in this experiment were 18S, CD18, Cortisol Receptor
and SPI1.
CD18 primers, synthesized by Bonsai technologies group (Alcobendas, Madrid), were
designed based on rainbow trout CD18 partial sequence data generated (Goetz et al., 2004a)
(accession number CF753098). CD18 sense 5´- CAGGATTGGCTGGAGGAATA-3´and
antisense 5-´ GCCTATTGTAGGCAGCTTGG-3´ primers amplified a PCR product of length 347
bp. Annealing temperature in PCR was 58 ºC.
PCR Primers for PU.1 (SPI1), synthesized by Bonsai Technologies Group (Alcobendas,
Madrid), were designed based on rainbow trout SPI1 (PU.1) complete sequence (AY606036)
(Ribas et al, 2007). PCR temperature was 55 ºC.

14
Cortisol receptor primers were designed by Acerete (Acerete et al., 2007), forward 5′-
AGTGCTCCTGGCTGTTYCTNATG-3′ and reverse 5′-TTTCGGTAATTGGTTGCTGATGAT-3′.
Annealing temperature was 55ºC.
TNF-α and IL-1β primers were designed based on rainbow trout sequences (AJ277604)
and (AJ223954) respectively. TNF-α Forward: 5’ GGATCCATGGAGGGGTATGCGATG 3’ and
TNF-α Reverse: 5’ AAGCTTTCATAGTGCAAACACACC 3’. IL-1β Forward: 5’
GGAACCGAGTTCAAGGACAA 3’ and IL-1β Reverse: 5’ GGCTACAGGTCTGGCTTCAG 3’.
PCR products were visualized on 1% agarose gels containing 0,5 µg/mL ethidium
bromide.

15
4. RESULTS AND DISCUSSION.
4.1 Cellular culture under different conditions with cortisol
A macrophage culture was performed to observe changes induced by cortisol at high
and medium concentrations in macrophages.
The results obtained (FIG 4) show that in case of cortisol at 300 ng/ml there are less
number of cells than in the other plates, while after 30 ng/ml of cortisol this decrease of cell
number was not observed. At the same time, RU486, a cortisol antagonist blocks the effects of
cortisol in culture.
The presence of a lesser number of cells after 300 ng/ml cortisol treatment could be
caused by induction of apoptosis or due to a decrease of cellular adhesion molecules
expression.
Fig 4: Pictures of 5 days culture plates in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).

16
There are diverse studies that have shown that glucocorticoids enhance cellular
apoptosis (Deane et al., 2006; Walsh et al., 2002; Weyts et al., 1998a); however, other studies
showed that glucocorticoids, including cortisol, have a protective role by decreasing cellular
apoptosis (Deane et al., 2006; Liles et al., 1995; MacKenzie et al., 2006; Weyts et al., 1998b),
which agrees with the results performed in our laboratory, where apoptosis was not observed
(Mackenzie et al., personal observations).
In this way, the hypothesis after observing these results is that under conditions of high
concentrations of cortisol, similar to acute stress plasma cortisol concentrations, a
downregulation of cellular adhesion molecules could be induced leading to a lesser number of
cells. As a physiological response this could end up in an inhibition of leucocyte migration under
conditions of acute stress with de purpose of saving energy to be directed to respond to the
stressor and the stress situation.
However, under moderate concentrations of cortisol (30 ng/ml) no noticeable changes
were observed, which from a physiological point of view, could mean that under conditions of
chronic stress the cellular adhesion is not modified.
17
4.2 CD18
Integrins are cell surface receptors that mediate cell-
cell and cell-matrix interactions and bidirectional signal
transduction between extracellular matrix and cytoplasm
(Huhtala et al., 2005). Integrins bind to different ligands, like
extracellular matrix proteins, such as collagens, laminins and
fibronectin, cell surface immunoglobulin superfamily counter
receptors such intercellular adhesion molecules (ICAMs) and
blood proteins such as fibrinogen (Hynes, 2002).
Integrins are heterodimeric glycoproteins composed
by one α-chain and one β-chain and are classified in families
in function of their β chain. There are at least nine different β
chains (β1, β2, β3, β4, β5, β6, β7, β8, βu) which bind to
different α chains, so there are a great number of possible
combinations, of which 20 have been identified (Hynes,
1992; Postigo and Sanchez-Madrid, 1993).
β2 chain is CD18 and this subunit binds to different α chain to form different integrins, in
this sense β2 chain (CD18) binds to αL (CD11a), αM (CD11b) and αX (CD11c) and form LFA-1
Mac-1 and gp 150.95 integrins (Haynes and McKinney, 1991; Postigo and Sanchez-Madrid,
1993; Springer, 1990a).
Integrins are very important in cellular adhesion and migration to inflammated tissues,
particularly for leukocyte to endothelial cells (Pitzalis et al., 2002), a very important process for
the transport of immune cells to inflammated tissues.
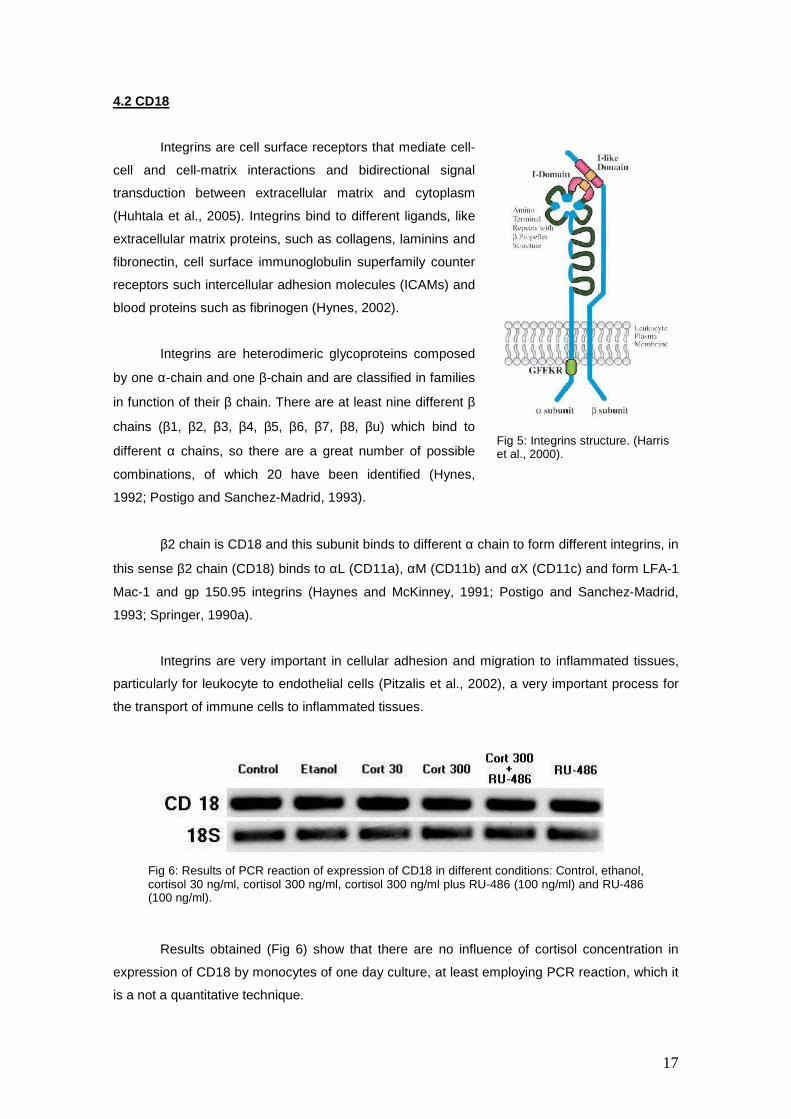
Results obtained (Fig 6) show that there are no influence of cortisol concentration in
expression of CD18 by monocytes of one day culture, at least employing PCR reaction, which it
is a not a quantitative technique.
Fig 6: Results of PCR reaction of expression of CD18 in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).
Fig 5: Integrins structure. (Harris et al., 2000).

18
These results do not support the initial theory that cortisol downregulates the expression
of cell adhesion molecules (CAMs) that was explained in anterior point, although this regulation
could be done on other adhesion molecules different of CD18.
Nevertheless the fact that cortisol does not induce noticeable variations in the
expression of this molecule does not mean that cortisol don’t have effects in cellular adhesion
process, since in addition to genetic mechanisms, cortisol triggers a set of non-genetic
mechanisms, which could have an important role. For example it is known that cellular adhesion
molecules, including integrins, are expressed in a low-binding avidity state to inhibit random
binding to endothelium, a state which quickly can change to high binding avidity (Hogg et al.,
1993; Hogg and Landis, 1993). Moreover, redistribution of cell adhesion molecules in cell
surface can modify the cellular adhesion process, and reorganizations are quickly possible
(Hogg et al., 1993; Hogg and Landis, 1993). In this way, there are experiments that show the
fast non-genetic mechanisms that modulate molecular adhesion processes, for example altering
binding avidity or surface distribution, (Pitzalis et al., 2002; Smith et al., 1988) and these
mechanisms could play an important role in the cellular adhesion process and could be inhibited
by glucocorticoids.

19
4.3 Glucocorticoid Receptor (GR)
Cortisol, binds to GR and the GC-GR complex moves to the nucleus where it has the
capacity to bind to specific DNA sequences called “glucocorticoid response elements”, inducing
the inhibition or stimulation of the relevant genes (Jewell et al., 1995; Pratt, 1993). To allow
binding of the ligand, the heat shock proteins Hsp90 and Hsp70 are essential as chaperones to
open the hydrophobic cleft of receptor (Morishima et al., 2000). So, the principal role of the GR
is as a transacting transcription factor.
This key role of GR in the cortisol action leads us to study this gene expression to help
us to better understand the mechanisms of action of cortisol in fish monocytes.
The GR structure and function is well established in mammals and conserved among
the vertebrates analyzed (Stolte et al., 2006). The GR belongs to the nuclear receptor
superfamily. This structure was predicted for the GR (Giguere et al., 1986) and consists in
several domains: N-terminal region, DNA-binding region, Hinge region and ligand-binding
region.
The N-terminal region (Fig 7: A/B) is the most variable and it is involved in
transactivation of downstream genes. The DNA-binding region (Fig 7: C) is the central domain
and binds to glucocorticoid responsive elements (GRE) in promoter regions to stimulate the
expression of glucocorticoid responsive genes. This region is strictly conserved in vertebrates
(Stolte et al., 2006). The hinge region (Fig 7: D) is involved in conformational changes during
receptor-ligand binding and it is quite variable in sequence and length. The ligand-binding
region (fig 7: E), situated at the C-terminus, is involved in joining to the ligand, including cortisol.
This region is relatively well conserved, what explains why multiple receptors can bind same
ligand and trigger a similar in vitro transactivation (Stolte et al., 2006).
Although the structure and function of GR is very well conserved in vertebrates there
are some differences. Unlike mammals, fish do not synthesize aldosterone, and it was
hypothesized that cortisol would act like a unique glucocorticoid to control mineralocorticoid
(MR) and glucocorticoid functions in fish, taking over also osmorregulatory functions (Dean et
al., 2003; Ducouret et al., 1995; Greenwood et al., 2003; Mommsen et al., 1999). As a result of
this MR and GR may act as cortisol receptors in fish species.
Fig 7: Glucocorticoid receptor cDNA schematic representation showing primers situation (Fw: Forward; Rv: Reverse). A/B, N-terminal region; C, DNA-binding region; D, Hinge region; E, ligand-binding region.

20
Moreover later investigations have modified the initial supposition of a unique cortisol
receptor, appearing duplicated genes (GR1 and GR2) and also splicing isoforms (Prunet et al.,
2006).
In rainbow trout concentrations of cortisol required to induce activation of downstream
genes are significantly different between the duplicate GR proteins, so GR2 requires lower
concentrations of cortisol than GR1a (with nine-amino-acids insert) to induce transactivation
(Bury et al., 2003). These differences could be involved in differential functions of different forms
in fish physiology.
The results obtained in this experiment were inconclusive because the huge differences
between the individuals analyzed, which prevent us to draw conclusions about the cortisol
action mechanisms.
In spite of the evidence on the mammalian GR regulation, there are few studies in fish.
In general, it has been shown that the GR expression and abundance is downregulated by the
presence of high concentrations of cortisol (Bellingham et al., 1992; Mommsen et al., 1999;
Pottinger et al., 2000; Rosewicz et al., 1988; Weyts et al., 1998c; Yudt and Cidlowski, 2002).
Moreover downregulation of GR expression, GR activity and half-life are modulated by GR
phosporylation (Webster et al., 1997), which is itself up-regulated by GRs (Orti et al., 1992).
This mechanism, with other influences, can modulate the stress response of organisms,
particularly when the stress is chronic (Sapolsky et al., 2000).
As regard to fishes in general there is an inverse correlation between GR abundance
and cortisol concentrations in fish. This fact was shown in different tissues and fish species, like
in peripheral blood leukocytes in carp (Cyprinus carpio L.) (Weyts et al., 1998c) and in liver
(Pottinger, 1990) and brain (Lee et al., 1992) of rainbow trout (Oncorhynchus mykiss).
However experiments performed in vitro (Sathiyaa and Vijayan, 2003) and in vivo
(Vijayan et al., 2003) with rainbow trout hepatocytes showed that, besides the downregulation of
GR there was an upregulation of GR mRNA, unlike the mammals. And other studies showed
Fig 4: Results of PCR reaction of expression of GR in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).

21
that the proteasome was involved in trout hepatocytes (Boone and Vijayan, 2002), which
coincide with mammal’s results (DeFranco, 2002; Wallace and Cidlowski, 2001).
In conclusion, abundance of GRs is highly variable between species, individuals and
from cell to cell (Bellingham et al., 1992). Moreover the differences between the results found by
the different authors in fish and the inconclusive dates obtained in this experiment stresses the
interest in elucidating the question of regulation of expression and abundance of GR in trout
monocytes/macrophages and posttranscriptional control mechanisms.

22
4.4 Cytokines (TNF-αααα and IL-1ββββ)
An inflammatory process triggers a cytokine cascade where Tumor necrosis factor α
(TNF-α) is released, followed by interleukin-1β (IL-1β) and then IL-6. This cascade triggers the
releasing of other cytokines, and globally functions like potent chemoattractans to induce
migration of neutrophils and macrophages to the site of infection (Secombes et al., 2001).
Cytokines are large (8–60 kDa) and soluble polypeptide mediators that regulate growth,
differentiation and function of different cellular types. The expression of these molecules and
their receptors is not restricted to cells of the immune system, and are expressed in other
tissues, like brain and endocrine glands (Turnbull and Rivier, 1999).
Moreover cytokines are involved in different physiological functions outside the
immunoregulation, like induce fever, sleep, anorexia, malaise and alterations in neuroendocrine
secretions. So, cytokines are key regulators of coordinated local and systemic responses to
tissue trauma, infection and disease.
Cytokines play an important role in interactions within and between nervous, endocrine
and immune systems. Cytokines are the molecules that act like messengers that stimulate the
hypothalamic-pituitary-adrenal (HPA) axis when the organism is under the action of a stressor
agent or an infection (Haddad et al., 2002; Harris and Bird, 2000; Liberman et al., 2007; Tracey,
2007).
IL-1 and TNF-α act as auto/paracrine or endocrine factors, regulating pituitary
development, cell proliferation, hormone secretion and feedback control of HPA axis (Hall et al.,
1985; Holland et al., 2002; Safieh-Garabedian et al., 2002a; Safieh-Garabedian et al., 2002b;
Woiciechowsky et al., 1999). Two events that confirm this question are the fact that intra-
peritoneal injection of recombinant trout IL-1β increase cortisol concentrations in plasma
(Holland et al., 2002) while cortisol is able to inhibit TNF-α preparation (Saeij et al., 2002).
These two cytokines, TNF-α and IL-1β, are two of the most important and the most well-
characterized of fish cytokines (Plouffe et al., 2005), and was taken to study their expression
under cortisol treatments.
4.4.1 Tumor necrosis factor αααα (TNF-αααα)
TNF-α is described as a “β- jellyroll” family of cytokines, in which eight antiparallel β-
strands form a sandwich 3D structure (Idriss and Naismith, 2000). TNF-α gene contains four
exons and three introns.

23
TNF are involved in diverse functions, like tumor cytotoxicity, broad-ranging
immunological activities, induction of many other cytokines, immunostimulant, or mediators of
the inflammatory response (Turnbull and Rivier, 1999).
TNF-α was identified at first time in Japanese flounder (Paralychthys olivaceus) (Hirono
et al., 2000), followed by mRNA identification in rainbow trout (Laing et al., 2001), brook trout
(Salvelinus fontinalis), carp (Saeij et al., 2003b) and sea bream (Garcia-Castillo et al., 2002).
The TNF molecule consists in rainbow trout of two forms, TNF-α y TNF-β, with two and
three isoforms respectively (Zou et al., 2002); nevertheless other authors (Goetz et al., 2004b)
affirm that TNF-β does not exist in fishes. Within TNF-α two genes presents in rainbow trout,
TNF1 and TNF2, TNF2 is highly expressed than TNF1. TNF-α in rainbow trout is expressed
constitutively in head kidney and gill and inducible in isolated head kidney cells (Laing et al.,
2001).
TNF-α is present in two different forms, a soluble 17 kDa form and a transmembrane 26
kDa form. Soluble 17 kDa form is synthesized by photolytic cleavage of the extracellular portion
of the 26 kDa form. The majority of actions mediated by TNF-α correspond to soluble and
mature 17 kDa form, which is found as homotrimers. However, the transmembrane form could
be too active and it may be responsible for some of local actions of TNF-α (Goetz et al., 2004b).
The results obtained (Fig 8) show a down regulation of TNF-α expression in conditions
of cortisol 300 ng/ml, however these results do not shown noticeable differences at the cortisol
concentration of 30 ng/ml.
These results are in concordance with data obtained by other authors, showing that
cortisol down-regulates the expression of TNF-α in different species and cells, like in human
macrophages (Joyce et al., 2001; Steer et al., 2000; Steer et al., 1998), rainbow trout
macrophages (MacKenzie et al., 2006), rainbow trout head kidney leukocytes (Holland et al.,
2003) and carp head kidney macrophages (Saeij et al., 2003a).
Fig 8: Results of PCR reaction of expression of TNF-α in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).

24
4.4.2 Interleukin 1-ββββ (IL 1-ββββ)
Interleukin have numerous and diverse immunoregulatory actions, some of them have
proinflamatory actions (IL-1α, IL-1β, IL-8, IL-9), while other interleukins have anti-inflammatory
effects (IL-1ra, IL-4, IL-10, IL-13) and IL also induce systemic aspects of acute phase response,
like the fever (Turnbull and Rivier, 1999).
IL-1β is a proinflamatory cytokine and is involved in induction of other
immunomodulatory cytokines and other transcripts of immune response, such as COX2 and
MHC class II (Major histocompatibility complex class II) (Plouffe et al., 2005).
Rainbow trout IL-1β was the first to be cloned in fish teleosts (Zou, 1999), and a second
IL-1β gene has been discovered in rainbow trout, and these two genes were called IL-1β1 and
IL-1β2, and both genes are expressed in head kidney leukocytes after LPS stimulation
(Secombes et al., 2001).
Unlike mammals, trout IL-1β haven’t got a clear caspase-1 cut site (Zou et al., 2002),
but it seems that trout IL-1B must be cleaved by interleukin converting enzyme (ICE) before to
be activated and secreted (Secombes et al., 2001).
Of the two IL-1β types receptors to IL-1: IL-1R type I and IL-1R type II, only the binding
of IL-1α or IL-1β to type 1 receptor work as antagonists by capturing excess of IL-1.
IL-1β plays an important role in interaction between immune, nervous and endocrine
systems, affecting directly to hypothalamic-pituitary-adrenal (HPA) axis function, stimulating
cortisol secretion, like it was demonstrated in rainbow trout with an intra-peritoneal injection of
recombinant trout IL-1β (Haddad et al., 2002; Holland et al., 2002).
The results obtained (Fig 9) show a down regulation of IL-1β expression in conditions of
cortisol 300 ng/ml, which simulates an acute stress condition, but do not show noticeable
differences at cortisol 30 ng/ml, corresponding to chronic conditions.
Fig 9: Results of PCR reaction of expression of IL 1-β in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).

25
These results are in concordance with the dates obtained by others authors, in which
cortisol acts as down-regulating expression of IL-1β in different conditions, like in human
leucocytes, rainbow trout culture leucocytes (MacKenzie et al., 2006; Zou et al., 2002) and carp
head kidney monocytes (Saeij et al., 2003a).

26
4.5 PU.1 (SPI-1)
PU.1 was identified as the putative oncogene Spi-1. The PU.1 locus is a high-frequency
integration site for spleen focus-forming virus (SFFV) in Friend murine acute erythroleukemias
and was called SFFV provirus integration site-1 (Spi-1) (Moreau-Gachelin et al., 1989; Moreau-
Gachelin et al., 1988).
PU.1 is a member of the transcription factor family Ets that plays an essential role in the
development and differentiation of both myeloid (granulocytes and monocytes/macrophages)
and lymphoid cells (Fisher and Scott, 1998; Hsu et al., 2004) and it is involved in expression
regulation of different genes, like heavy and light chains of immunoglobulins, cytokines (IL-1β,
MIP-1α, TNF-α, IL-1ra), complement receptors, growth factors, different components related
with phagocytosis and cellular development, enzymes, MHC class II and other genes
(Henderson and Calame, 1998; Lloberas et al., 1999).
During haematopoiesis PU.1 is up-regulated in myeloid development and down-
regulated in erythrocyte differentiation (Voso et al., 1994). In this way PU.1 deficient mouse
exhibits a group of functional deficiencies in macrophages, granulocytes and progenitors of B
and T lymphocytes (Scott et al., 1994). Different studies have shown that PU.1 blocks erythroid
differentiation by directly antagonizing GATA-1 activity. This reciprocal negative regulation
determines myeloid versus erythroid fate (Rekhtman et al., 2003; Rekhtman et al., 1999;
Rhodes et al., 2005; Zhang et al., 2000).
Moreover, recent studies have demonstrated that the concentration of PU.1
determinates haematopoietic progenitor cells fate to B lymphocytes or macrophages. Low
concentrations of PU.1 induces the B lymphocyte differentiation, while high concentrations of
PU.1 triggers macrophage differentiation and development and blocks B cells proliferation
(DeKoter and Singh, 2000).
The results obtained (Fig 10) show a down-regulation in expression of PU.1 (Spi-1)
gene in 300 ng/ml cortisol concentration, but do not reveal noticeable changes in 30 ng/ml of
cortisol condition.
Fig 10: Results of PCR reaction of expression of PU.1 in rainbow trout monocytes (SPI-1 in different conditions: Control, ethanol, cortisol 30 ng/ml, cortisol 300 ng/ml, cortisol 300 ng/ml plus RU-486 (100 ng/ml) and RU-486 (100 ng/ml).

27
These results suggest that cortisol repress the monocyte development and
differentiation, at least in conditions of high levels of cortisol, which simulate conditions of a
acute stress, but it seems that in conditions of moderate concentrations of cortisol, which are
similar to the plasma concentrations in chronic stress, this downregulation does not occur.
These results may help us to better understand the immune fish responses to stress. In
this sense, the downregulation of PU.1 gene in rainbow trout monocytes under high
concentrations of cortisol, and not the downregulation at moderate concentrations could be
explained as the fish response to an acute stress blocking myeloid differentiation and
development, and thus saving energy to cope with the acute stress situation.
Moreover, a possible interpretation for no downregulation of PU.1 at moderate
concentrations of cortisol could be that, in a chronic stress situation, blocking of myeloid and
lymphoid cells differentiation could not be beneficial in the long term, because it could
compromise immune system and their defence against pathogens.

28
5. CONCLUSIONS.
The effect of cortisol addition to trout macrophages culture shows that high
concentration of cortisol, similar than under conditions of acute stress, induces a reduced
number of cells compared with the conditions in which the cortisol is absent or it is counteracted
with its antagonist, RU-486. However, under conditions of cortisol at moderate concentrations,
similar than under chronic stress conditions, no noticeable variations are recorded respect to
control plates.
These results could be explained by two theories, the increment of cellular apoptosis or
the inhibition of cell adhesion to the matrix. Experiments performed in our laboratory show that
increase of apoptosis do not takes place, so the reduction of adhesion molecules could be the
cause of this effect.
From a physiological point of view the downregulation of cellular adhesion molecules in
conditions of acute stress has a functional sense, since the organism reduce the cellular
adhesion molecules expression decreasing cellular trafficking, with the consequent savings in
energy, which can be redirected to cope with the stress situation.
However, the results obtained to the adhesion molecule tested (CD18) reveal that
cortisol does not significantly affect the expression of this molecule, although this regulation
could exist in other adhesion molecules different than CD18.
Moreover, the lack of down-regulation of expression of CD18 does not mean that
cortisol doen not have effects in the cellular adhesion phenomenon. Cortisol could regulate the
expression of other genes, or through non-genetic mechanisms, like the changing of avidity
binding of cellular adhesion molecules or the alteration of their situation in cellular surface,
which could play an important role in cellular adhesion process.
As regards to glucocorticoid receptor (GR) the results obtained in this work were
inconclusive because of the enormous variation between de individuals. Nevertheless, this
question could be very interesting to study because it could provide a better understanding of
cortisol mechanisms of action.
Cortisol showed a clear effect on the expression levels of the cytokines TNF-α and IL-
1β, showing similar results than previously observed. This effect is opposite to the observed in
response to LPS in rainbow trout macrophages.
As regards to the PU.1 (Spi-1) gene, the results found show that cortisol down-regulates
the expression of this gene in conditions of high concentration of cortisol, but not under

29
moderate concentrations. These situations are very similar to the concentrations of cortisol in
plasma under acute and chronic stress conditions.
In this way, a possible functional and physiologic interpretation of these results is that in
conditions of acute stress PU.1 is downregulated, and so, differentiation and development of
monocytes to mature macrophages are blocked or attenuated. This fact could have the purpose
of save the energy of myeloid differentiation to use that energy to an overall systemic response
in front of an acute stressor.
Nevertheless, under conditions of chronic stress, the inhibition of myeloid differentiation
could be not adequate at long term, because it could compromise the defence of the organism
against pathogens, and for this reason the expression of this transcription factor (PU.1) could
not be downregulated in these conditions.
In short, the results obtained in this work support the conclusions obtained by other
authors (MacKenzie et al., 2006), who, using functional microarrays have arrived to the
conclusions that in general cortisol have generally contrary effects to LPS. These responses
included the cortisol cell protection of the suppression of genes involved in basic cellular
functions and maintenance of activities, including protection against apoptosis.
In conclusion, cortisol inhibits the expression of the inflammatory cytokines TNF-α and
IL-1β and the transcription factor PU.1, with a possible purpose of saving energy in order to
cope with the response to acute stress conditions; however, the same response to conditions of
chronic stress is not observed, as this downregulation could be negative in the long term. Our
results also indicate the induction of differential mechanisms involved in the regulatory roles of
cortisol which are depending on the concentration levels.

30
6. REFERENCES.
1. Acerete L., Balasch J. C., Castellana B., Redruello B., Roher N., Canario A. V., Planas J. V., Mackenzie S. and Tort L. (2007) Cloning of the glucocorticoid receptor (GR) in gilthead seabream (Sparus aurata) Differential expression of GR and immune genes in gilthead seabream after an immune challenge. Comp Biochem Physiol B Biochem Mol Biol. In press.
2. Alder M. N., Rogozin I. B., Iyer L. M., Glazko G. V., Cooper M. D. and Pancer Z. (2005) Diversity and function of adaptive immune receptors in a jawless vertebrate. Science 310, 1970-3.
3. Atherton A. and Born G. V. (1972) Quantitative investigations of the adhesiveness of circulating polymorphonuclear leucocytes to blood vessel walls. J Physiol 222, 447-74.
4. Baggiolini M., Dewald B. and Moser B. (1997) Human chemokines: an update. Annu Rev Immunol 15, 675-705.
5. Bellingham D. L., Sar M. and Cidlowski J. A. (1992) Ligand-dependent down-regulation of stably transfected human glucocorticoid receptors is associated with the loss of functional glucocorticoid responsiveness. Mol Endocrinol 6, 2090-102.
6. Bernstein R. M., Schluter S. F., Bernstein H. and Marchalonis J. J. (1996) Primordial emergence of the recombination activating gene 1 (RAG1): sequence of the complete shark gene indicates homology to microbial integrases. Proc Natl Acad Sci U S A 93, 9454-9.
7. Bevilacqua M. P. and Nelson R. M. (1993) Selectins. J Clin Invest 91, 379-87. 8. Boone A. N. and Vijayan M. M. (2002) Constitutive heat shock protein 70 (HSC70)
expression in rainbow trout hepatocytes: effect of heat shock and heavy metal exposure. Comp Biochem Physiol C Toxicol Pharmacol 132, 223-33.
9. Brett J. R. (1958) Implications and assessment of environmental stress. In H.R.MacMillan Lectures in Fisheries (Edited by Larking P. A.), p. 69-83, Univ. British Columbia., Vancouver.
10. Bromage E. S., Kaattari I. M., Zwollo P. and Kaattari S. L. (2004) Plasmablast and plasma cell production and distribution in trout immune tissues. J Immunol 173, 7317-23.
11. Bury N. R., Sturm A., Le Rouzic P., Lethimonier C., Ducouret B., Guiguen Y., Robinson-Rechavi M., Laudet V., Rafestin-Oblin M. E. and Prunet P. (2003) Evidence for two distinct functional glucocorticoid receptors in teleost fish. J Mol Endocrinol 31, 141-56.
12. Cooper M. D. and Alder M. N. (2006) The Evolution of Adaptive Immune Systems. Cell 124, 815-822.
13. Crowhurst M. O., Layton J. E. and Lieschke G. J. (2002) Developmental biology of zebrafish myeloid cells. Int J Dev Biol 46, 483-92.
14. Cuesta A., Salinas I., Rodriguez A., Esteban M. A. and Meseguer J. (2006) Injection of xenogeneic cells into teleost fish elicits systemic and local cellular innate immune responses. Cell Tissue Res 326, 93-9.
15. Dean B. D., Whitlow Z. W. and Borski R. J. (2003) Glucocorticoid receptor upregulation during seawater adaptation in a euryhaline teleost, the tilapia (Oreochromis mossambicus). General and Comparative Endocrinology 132, 112-118.
16. Deane E. E., Zhou L. and Woo N. Y. (2006) Cortisol can be pro- or anti-apoptotic in sea bream cells: potential role of HSP70 induction for cytoprotection. Mol Cell Endocrinol 259, 57-64.
17. DeFranco D. B. (2002) Navigating steroid hormone receptors through the nuclear compartment. Mol Endocrinol 16, 1449-55.
18. DeKoter R. P. and Singh H. (2000) Regulation of B lymphocyte and macrophage development by graded expression of PU.1. Science 288, 1439-41.
19. Ducouret B., Tujague M., Ashraf J., Mouchel N., Servel N., Valotaire Y. and Thompson E. B. (1995) Cloning of a teleost fish glucocorticoid receptor shows that it contains a deoxyribonucleic acid-binding domain different from that of mammals. Endocrinology 136, 3774-83.
20. Ellis A. E. (2001) Innate host defense mechanisms of fish against viruses and bacteria. Dev Comp Immunol 25, 827-39.
21. Fange R. (1986) Lymphoid organs in sturgeons (Acipenseridae). Vet Immunol Immunopathol 12, 153-61.
22. Fisher R. C. and Scott E. W. (1998) Role of PU.1 in hematopoiesis. Stem Cells 16, 25-37. 23. Freemont A. J. and Hoyland J. A. (1996) Cell adhesion molecules. Clin Mol Pathol 49, M321-
M330.

31
24. Garcia-Castillo J., Pelegrin P., Mulero V. and Meseguer J. (2002) Molecular cloning and expression analysis of tumor necrosis factor alpha from a marine fish reveal its constitutive expression and ubiquitous nature. Immunogenetics 54, 200-7.
25. Giguere V., Hollenberg S. M., Rosenfeld M. G. and Evans R. M. (1986) Functional domains of the human glucocorticoid receptor. Cell 46, 645-52.
26. Goetz F. W., Iliev D. B., McCauley L. A., Liarte C. Q., Tort L. B., Planas J. V. and Mackenzie S. (2004a) Analysis of genes isolated from lipopolysaccharide-stimulated rainbow trout (Oncorhynchus mykiss) macrophages. Mol Immunol 41, 1199-210.
27. Goetz F. W., Planas J. V. and MacKenzie S. (2004b) Tumor necrosis factors. Developmental & Comparative Immunology 28, 487-497.
28. Grassi E., Basari F. and Chimenti C. (1997) Adrenocortical and adrenomedullary homologs in eight species of adult and developing teleosts: morphology, histology, and immunohistochemistry. Gen Comp Endocrinol 108, 483-96.
29. Greenwood A. K., Butler P. C., White R. B., DeMarco U., Pearce D. and Fernald R. D. (2003) Multiple corticosteroid receptors in a teleost fish: distinct sequences, expression patterns, and transcriptional activities. Endocrinology 144, 4226-36.
30. Haddad J. J., Saade N. E. and Safieh-Garabedian B. (2002) Cytokines and neuro-immune-endocrine interactions: a role for the hypothalamic-pituitary-adrenal revolving axis. Journal of Neuroimmunology 133, 1-19.
31. Hall N. R., McGillis J. P., Spangelo B. L. and Goldstein A. L. (1985) Evidence that thymosins and other biologic response modifiers can function as neuroactive immunotransmitters. J Immunol 135, 806s-811s.
32. Harding F. A., Cohen N. and Litman G. W. (1990) Immunoglobulin heavy chain gene organization and complexity in the skate, Raja erinacea. Nucleic Acids Res 18, 1015-20.
33. Harris E. S., McIntyre T. M., Prescott S. M. and Zimmerman G. A. (2000) Minireview: The Leukocyte Integrins. J Biol Chem 275, 23409-23412.
34. Harris J. and Bird D. J. (2000) Modulation of the fish immune system by hormones. Vet Immunol Immunopathol 77, 163-76.
35. Haynes L. and McKinney E. C. (1991) Shark spontaneous cytotoxicity: characterization of the regulatory cell. Dev Comp Immunol 15, 123-34.
36. Henderson A. and Calame K. (1998) Transcriptional regulation during B cell development., Vol. 16, p. 163-200.
37. Herbert T. B. and Cohen S. (1993) Stress and immunity in humans: a meta-analytic review. Psychosom Med 55, 364-79.
38. Hirono I., Nam B. H., Kurobe T. and Aoki T. (2000) Molecular cloning, characterization, and expression of TNF cDNA and gene from Japanese flounder Paralychthys olivaceus. J Immunol 165, 4423-7.
39. Hogg N., Harvey J., Cabanas C. and Landis R. C. (1993) Control of leukocyte integrin activation. Am Rev Respir Dis 148, S55-9.
40. Hogg N. and Landis R. C. (1993) Adhesion molecules in cell interactions. Curr Opin Immunol 5, 383-90.
41. Hogg N., Landis, R.C. (1993) Adhesion molecules in cell interactions. Curr. Opinion Immunol. 5, 383-390.
42. Holland J. W., Gould C. R., Jones C. S., Noble L. R. and Secombes C. J. (2003) The expression of immune-regulatory genes in rainbow trout, Oncorhynchus mykiss, during a natural outbreak of proliferative kidney disease (PKD). Parasitology 126 Suppl, S95-102.
43. Holland J. W., Pottinger T. G. and Secombes C. J. (2002) Recombinant interleukin-1 beta activates the hypothalamic-pituitary-interrenal axis in rainbow trout, Oncorhynchus mykiss. J Endocrinol 175, 261-7.
44. Hsu K., Traver D., Kutok J. L., Hagen A., Liu T. X., Paw B. H., Rhodes J., Berman J. N., Zon L. I., Kanki J. P. and Look A. T. (2004) The pu.1 promoter drives myeloid gene expression in zebrafish. Blood 104, 1291-7.
45. Huhtala M., Heino J., Casciari D., de Luise A. and Johnson M. S. (2005) Integrin evolution: Insights from ascidian and teleost fish genomes. Matrix Biology 24, 83-95.
46. Hynes R. O. (1992) Integrins: versatility, modulation, and signaling in cell adhesion. Cell 69, 11-25.
47. Hynes R. O. (2002) Integrins: bidirectional, allosteric signaling machines. Cell 110, 673-87. 48. Idriss H. T. and Naismith J. H. (2000) TNF alpha and the TNF receptor superfamily: structure-
function relationship(s). Microsc Res Tech 50, 184-95.

32
49. Jewell C. M., Webster J. C., Burnstein K. L., Sar M., Bodwell J. E. and Cidlowski J. A. (1995) Immunocytochemical analysis of hormone mediated nuclear translocation of wild type and mutant glucocorticoid receptors. J Steroid Biochem Mol Biol 55, 135-46.
50. Joyce D. A., Gimblett G. and Steer J. H. (2001) Targets of glucocorticoid action on TNF-alpha release by macrophages. Inflamm Res 50, 337-40.
51. Khalturin K., Panzer Z., Cooper M. D. and Bosch T. C. (2004) Recognition strategies in the innate immune system of ancestral chordates. Mol Immunol 41, 1077-87.
52. Laing K. J., Wang T., Zou J., Holland J., Hong S., Bols N., Hirono I., Aoki T. and Secombes C. J. (2001) Cloning and expression analysis of rainbow trout Oncorhynchus mykiss tumour necrosis factor-alpha. Eur J Biochem 268, 1315-22.
53. Lasky L. A. (1992) Selectins: interpreters of cell-specific carbohydrate information during inflammation. Science 258, 964-9.
54. Lee P. C., Goodrich M., Struve M., Yoon H. I. and Weber D. (1992) Liver and brain glucocorticoid receptor in rainbow trout, Oncorhynchus mykiss: down-regulation by dexamethasone. Gen Comp Endocrinol 87, 222-31.
55. Li J., Barreda D. R., Zhang Y.-A., Boshra H., Gelman A. E., LaPatra S., Tort L. and Sunyer J. O. (2007) Complement and B cell cooperation in teleost fish: Role in phagocytosis and inflammation. Molecular Immunology 44, 205.
56. Liberman A. C., Druker J., Perone M. J. and Arzt E. (2007) Glucocorticoids in the regulation of transcription factors that control cytokine synthesis. Cytokine Growth Factor Rev 18, 45-56.
57. Liles W. C., Dale D. C. and Klebanoff S. J. (1995) Glucocorticoids inhibit apoptosis of human neutrophils. Blood 86, 3181-8.
58. Lloberas J., Soler C. and Celada A. (1999) The key role of PU.1/SPI-1 in B cells, myeloid cells and macrophages. Immunology Today 20, 184-189.
59. MacKenzie S., Iliev D., Liarte C., Koskinen H., Planas J. V., Goetz F. W., Molsa H., Krasnov A. and Tort L. (2006) Transcriptional analysis of LPS-stimulated activation of trout (Oncorhynchus mykiss) monocyte/macrophage cells in primary culture treated with cortisol. Mol Immunol 43, 1340-8.
60. Magnadottir B. (2006) Innate immunity of fish (overview). Fish Shellfish Immunol 20, 137-51. 61. Magnadottir B., Lange S., Gudmundsdottir S., Bogwald J. and Dalmo R. A. (2005) Ontogeny
of humoral immune parameters in fish. Fish Shellfish Immunol 19, 429-39. 62. McKinney E. C., Haynes L. and Droese A. L. (1986) Macrophage-like effector of spontaneous
cytotoxicity from the shark. Dev Comp Immunol 10, 497-508. 63. Medzhitov R. and Janeway C., Jr. (2000) Innate immune recognition: mechanisms and
pathways. Immunol Rev 173, 89-97. 64. Mommsen T. P., Vijayan M. M. and Moon T. W. (1999) Cortisol in teleosts: dynamics,
mechanisms of action, and metabolic regulation. Reviews in Fish Biology and Fisheries 9, 211-268.
65. Moreau-Gachelin F., Ray D., Mattei M. G., Tambourin P. and Tavitian A. (1989) The putative oncogene Spi-1: murine chromosomal localization and transcriptional activation in murine acute erythroleukemias. Oncogene 4, 1449-56.
66. Moreau-Gachelin F., Tavitian A. and Tambourin P. (1988) Spi-1 is a putative oncogene in virally induced murine erythroleukaemias. Nature 331, 277-80.
67. Morishima Y., Murphy P. J., Li D. P., Sanchez E. R. and Pratt W. B. (2000) Stepwise assembly of a glucocorticoid receptor.hsp90 heterocomplex resolves two sequential ATP-dependent events involving first hsp70 and then hsp90 in opening of the steroid binding pocket. J Biol Chem 275, 18054-60.
68. Orti E., Bodwell J. E. and Munck A. (1992) Phosphorylation of steroid hormone receptors. Endocr Rev 13, 105-28.
69. Ottaviani E. and Franceschi C. (1996) The neuroimmunology of stress from invertebrates to man. Prog Neurobiol 48, 421-40.
70. Pancer Z., Amemiya C. T., Ehrhardt G. R., Ceitlin J., Gartland G. L. and Cooper M. D. (2004) Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature 430, 174-80.
71. Pettey C. L. and McKinney E. C. (1988) Induction of cell-mediated cytotoxicity by shark 19S IgM. Cell Immunol 111, 28-38.
72. Pitzalis C., Pipitone N. and Perretti M. (2002) Regulation of leukocyte-endothelial interactions by glucocorticoids. Ann N Y Acad Sci 966, 108-18.

33
73. Plouffe D. A., Hanington P. C., Walsh J. G., Wilson E. C. and Belosevic M. (2005) Comparison of select innate immune mechanisms of fish and mammals. Xenotransplantation 12, 266-77.
74. Postigo A. A. and Sanchez-Madrid F. (1993) Adhesion and homing molecules. Transplant Proc 25, 65-9.
75. Pottinger T. G. (1990) The effect of stress and exogenous cortisol on receptor-like binding of cortisol in the liver of rainbow trout, Oncorhynchus mykiss. Gen Comp Endocrinol 78, 194-203.
76. Pottinger T. G., Carrick T. R., Appleby A. and Yeomans W. E. (2000) High blood cortisol levels and low cortisol receptor affinity: is the chub, Leuciscus cephalus, a cortisol-resistant teleost? Gen Comp Endocrinol 120, 108-17.
77. Pratt W. B. (1993) The role of heat shock proteins in regulating the function, folding, and trafficking of the glucocorticoid receptor. J Biol Chem 268, 21455-8.
78. Press C. M. and Evensen O. (1999) The morphology of the immune system in teleost fishes. Fish & Shellfish Immunology 9, 309-318.
79. Prunet P., Sturm A. and Milla S. (2006) Multiple corticosteroid receptors in fish: From old ideas to new concepts. General and Comparative Endocrinology 147, 17-23.
80. Rast J. P. and Litman G. W. (1994) T-cell receptor gene homologs are present in the most primitive jawed vertebrates. Proc Natl Acad Sci U S A 91, 9248-52.
81. Reite O. B. and Evensen O. (2006) Inflammatory cells of teleostean fish: A review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish & Shellfish Immunology 20, 192-208.
82. Rekhtman N., Choe K. S., Matushansky I., Murray S., Stopka T. and Skoultchi A. I. (2003) PU.1 and pRB interact and cooperate to repress GATA-1 and block erythroid differentiation. Mol Cell Biol 23, 7460-74.
83. Rekhtman N., Radparvar F., Evans T. and Skoultchi A. I. (1999) Direct interaction of hematopoietic transcription factors PU.1 and GATA-1: functional antagonism in erythroid cells. Genes Dev 13, 1398-411.
84. Rhodes J., Hagen A., Hsu K., Deng M., Liu T. X., Look A. T. and Kanki J. P. (2005) Interplay of pu.1 and gata1 determines myelo-erythroid progenitor cell fate in zebrafish. Dev Cell 8, 97-108.
85. Ribas L., Martínez M., Roher N, Doñate C., Goetz F.W., Iliev D., Planas J., Tort L. Mackenzie S. (2007) The role of the hematopoietic transcription factor PU.1 in the immune response in the rainbow trout (Oncorhynchus mykiss), cloning and characterization. Fish Shellfish Immunol.In Press.
86. Roberts R. (1989) Fish Pathology. Bailliere Tindall, London. 87. Rombout J. H., Huttenhuis H. B., Picchietti S. and Scapigliati G. (2005) Phylogeny and
ontogeny of fish leucocytes. Fish Shellfish Immunol 19, 441-55. 88. Rosewicz S., McDonald A. R., Maddux B. A., Goldfine I. D., Miesfeld R. L. and Logsdon C.
D. (1988) Mechanism of glucocorticoid receptor down-regulation by glucocorticoids. J Biol Chem 263, 2581-4.
89. Rowley A. F., Hunt T. C., Page M. and Mainwaring G. (1999) Vertebrate blood cells. Cambridge: University press., 18-127.
90. Saeij J. P., Stet R. J., de Vries B. J., van Muiswinkel W. B. and Wiegertjes G. F. (2003a) Molecular and functional characterization of carp TNF: a link between TNF polymorphism and trypanotolerance? Dev Comp Immunol 27, 29-41.
91. Saeij J. P., Van Muiswinkel W. B., Groeneveld A. and Wiegertjes G. F. (2002) Immune modulation by fish kinetoplastid parasites: a role for nitric oxide. Parasitology 124, 77-86.
92. Saeij J. P., Verburg-van Kemenade L. B., van Muiswinkel W. B. and Wiegertjes G. F. (2003b) Daily handling stress reduces resistance of carp to Trypanoplasma borreli: in vitro modulatory effects of cortisol on leukocyte function and apoptosis. Dev Comp Immunol 27, 233-45.
93. Safieh-Garabedian B., Dardenne M., Pleau J. M. and Saade N. E. (2002a) Potent analgesic and anti-inflammatory actions of a novel thymulin-related peptide in the rat. Br J Pharmacol 136, 947-55.
94. Safieh-Garabedian B., Poole S., Haddad J. J., Massaad C. A., Jabbur S. J. and Saade N. E. (2002b) The role of the sympathetic efferents in endotoxin-induced localized inflammatory hyperalgesia and cytokine upregulation. Neuropharmacology 42, 864-72.
95. Sapolsky R. M., Romero L. M. and Munck A. U. (2000) How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21, 55-89.

34
96. Sathiyaa R. and Vijayan M. M. (2003) Autoregulation of glucocorticoid receptor by cortisol in rainbow trout hepatocytes. Am J Physiol Cell Physiol 284, C1508-15.
97. Scott E. W., Simon M. C., Anastasi J. and Singh H. (1994) Requirement of transcription factor PU.1 in the development of multiple hematopoietic lineages. Science 265, 1573-7.
98. Schall T. J. and Bacon K. B. (1994) Chemokines, leukocyte trafficking, and inflammation. Curr Opin Immunol 6, 865-73.
99. Schreck C. B. (1981) Stress and compensation in teleostean fishes: response to social and physical
100. factors. In Stress and Fish (Edited by Pickering A. D.), p. 295-321. Academic Press, London. 101. Secombes C. J., Wang T., Hong S., Peddie S., Crampe M., Laing K. J., Cunningham C. and
Zou J. (2001) Cytokines and innate immunity of fish. Dev Comp Immunol 25, 713-23. 102. Smith M. D., Ahern M. J., Brooks P. M. and Roberts-Thomson P. J. (1988) The clinical and
immunological effects of pulse methylprednisolone therapy in rheumatoid arthritis. III. Effects on immune and inflammatory indices in synovial fluid. J Rheumatol 15, 238-41.
103. Springer T. A. (1990a) Adhesion receptors of the immune system. Nature 346, 425-34. 104. Springer T. A. (1990b) Adhesion receptors of the immune system. Nature 346, 425-434. 105. Stavitsky A. B. (2007) The innate immune response to infection, toxins and trauma evolved
into networks of interactive, defensive, reparative, regulatory, injurious and pathogenic pathways. Mol Immunol 44, 2787-99.
106. Steer J. H., Kroeger K. M., Abraham L. J. and Joyce D. A. (2000) Glucocorticoids suppress tumor necrosis factor-alpha expression by human monocytic THP-1 cells by suppressing transactivation through adjacent NF-kappa B and c-Jun-activating transcription factor-2 binding sites in the promoter. J Biol Chem 275, 18432-40.
107. Steer J. H., Ma D. T., Dusci L., Garas G., Pedersen K. E. and Joyce D. A. (1998) Altered leucocyte trafficking and suppressed tumour necrosis factor alpha release from peripheral blood monocytes after intra-articular glucocorticoid treatment. Ann Rheum Dis 57, 732-7.
108. Stolte E. H., van Kemenade B. M., Savelkoul H. F. and Flik G. (2006) Evolution of glucocorticoid receptors with different glucocorticoid sensitivity. J Endocrinol 190, 17-28.
109. Tort L., Balasch J. C. and Mackenzie S. (2003) Fish immune system. A crossroad between innate and adaptive responses. Inmunología 22, 277-286.
110. Tort L., Balasch J. C. and Mackenzie S. (2004) Fish health challenge after stress. Indicators of immunocompetence. Contributions to Science 2, 443-454.
111. Tracey K. J. (2002) The inflammatory reflex. Nature 420, 853-9. 112. Tracey K. J. (2007) Physiology and immunology of the cholinergic antiinflammatory pathway.
J Clin Invest 117, 289-96. 113. Turnbull A. V. and Rivier C. L. (1999) Regulation of the hypothalamic-pituitary-adrenal axis
by cytokines: actions and mechanisms of action. Physiol Rev 79, 1-71. 114. Vijayan M. M., Raptis S. and Sathiyaa R. (2003) Cortisol treatment affects glucocorticoid
receptor and glucocorticoid-responsive genes in the liver of rainbow trout. Gen Comp Endocrinol 132, 256-63.
115. Voso M. T., Burn T. C., Wulf G., Lim B., Leone G. and Tenen D. G. (1994) Inhibition of Hematopoiesis by Competitive Binding of Transcription Factor PU.1, Vol. 91, p. 7932-7936.
116. Wake M. H. (1997) Hyman’s comparative Vertebrate Anatomy. University of Chicago Press, Chicago.
117. Walsh C. J. and Luer C. A. (1998) Comparative phagocytic and pinocytic activities of leucocytes from peripheral blood and lymphomyeloid tissues of the nurse shark (Ginglymostoma cirratumBonaterre) and the clearnose skate (Raja eglanteriaBosc). Fish & Shellfish Immunology 8, 197-215.
118. Walsh C. J., Wyffels J. T., Bodine A. B. and Luer C. A. (2002) Dexamethasone-induced apoptosis in immune cells from peripheral circulation and lymphomyeloid tissues of juvenile clearnose skates, Raja eglanteria. Dev Comp Immunol 26, 623-33.
119. Wallace A. D. and Cidlowski J. A. (2001) Proteasome-mediated glucocorticoid receptor degradation restricts transcriptional signaling by glucocorticoids. J Biol Chem 276, 42714-21.
120. Webster J. C., Jewell C. M., Bodwell J. E., Munck A., Sar M. and Cidlowski J. A. (1997) Mouse glucocorticoid receptor phosphorylation status influences multiple functions of the receptor protein. J Biol Chem 272, 9287-93.
121. Wedemeyer G. A. and McLeay D. J. (1981) Methods for determining the tolerance of fishes to environmental stressors. In Stress and fish (Edited by Pickering A. D.), p. 248-275, London and New York.

35
122. Weyts F. A., Flik G., Rombout J. H. and Verburg-van Kemenade B. M. (1998a) Cortisol induces apoptosis in activated B cells, not in other lymphoid cells of the common carp, Cyprinus carpio L. Dev Comp Immunol 22, 551-62.
123. Weyts F. A., Flik G. and Verburg-van Kemenade B. M. (1998b) Cortisol inhibits apoptosis in carp neutrophilic granulocytes. Dev Comp Immunol 22, 563-72.
124. Weyts F. A., Verburg-van Kemenade B. M. and Flik G. (1998c) Characterisation of glucocorticoid receptors in peripheral blood leukocytes of Carp, Cyprinus carpio L. Gen Comp Endocrinol 111, 1-8.
125. Wilhelm S. W. and Suttle C. A. (1999) Viruses and nutrient cycles in the sea. Bioscience. 49, 781-788.
126. Woiciechowsky C., Schoning B., Daberkow N., Asche K., Stoltenburg G., Lanksch W. R. and Volk H. D. (1999) Brain-IL-1beta induces local inflammation but systemic anti-inflammatory response through stimulation of both hypothalamic-pituitary-adrenal axis and sympathetic nervous system. Brain Res 816, 563-71.
127. Yudt M. R. and Cidlowski J. A. (2002) The glucocorticoid receptor: coding a diversity of proteins and responses through a single gene. Mol Endocrinol 16, 1719-26.
128. Zapata A. (1979) Ultrastructural study of the teleost fish kidney. Dev Comp Immunol 3, 55-65. 129. Zapata A., Diez B., Cejalvo T., Gutierrez-de Frias C. and Cortes A. (2006) Ontogeny of the
immune system of fish. Fish Shellfish Immunol 20, 126-36. 130. Zapata A. G., Cooper, E.L. (1990) The immune System: Comparative Histopathology. Wiley,
Chichester, U.K. 131. Zapata A. G., Torroba M., Vicente A., Varas A., Sacedon R. and Jimenez E. (1995) The
relevance of cell microenvironments for the appearance of lympho-haemopoietic tissues in primitive vertebrates. Histol Histopathol 10, 761-78.
132. Zhang P., Zhang X., Iwama A., Yu C., Smith K. A., Mueller B. U., Narravula S., Torbett B. E., Orkin S. H. and Tenen D. G. (2000) PU.1 inhibits GATA-1 function and erythroid differentiation by blocking GATA-1 DNA binding. Blood 96, 2641-8.
133. Zou J., Wang T., Hirono I., Aoki T., Inagawa H., Honda T., Soma G. I., Ototake M., Nakanishi T., Ellis A. E. and Secombes C. J. (2002) Differential expression of two tumor necrosis factor genes in rainbow trout, Oncorhynchus mykiss. Developmental & Comparative Immunology 26, 161-172.
134. Zwollo P., Cole S., Bromage E. and Kaattari S. (2005) B cell heterogeneity in the teleost kidney: evidence for a maturation gradient from anterior to posterior kidney. J Immunol 174, 6608-16.


















