
Targeting of E. coli β-galactosidase to the nucleus in yeast
9
Cell,Vol. 36, 1057-1065, April 1984, CopyrightQ 1984 by MIT Targeting of E. coli ,&Galactosidase to the Nucleus in Yeast Michael N. Hall,* Lynna Hereford,’ and ha HerskowiW *Department of Biochemistry and Biophysics University of California, San Francisco San Francisco, California 94143 +Dana Farber Cancer Research Center Harvard Medical School Boston. Massachusetts 02115 Summary In order to identify determinants governing nuclear protein localization, we constructed a set of hybrid genes by fusing the S. cerevisiae gene, MATa2, coding for a presumptive nuclear protein, and the E. coli gene, /acZ, coding for B-galactosidase. The re- sultant hybrid proteins contain 3, 13, 25, 67, or all 210 amino acids of wild-type a2 protein at the amino terminus and a constant, enzymatically active por- tion of @-galactosidase at the carboxy terminus. Indirect immunofluorescence and subcellular frac- tionation studies with yeast cells containing the a2- LacZ hybrid proteins indicate that the a2 segment can direct localization of jI-galactosidase to the nu- cleus. A segment as small as 13 amino acids from a2 is sufficient for this localization. Comparison of amino acid sequences of other nuclear proteins with this region of a2 reveals a sequence that may be necessary for nuclear targeting. Production of some a2-LacZ hybrid proteins causes cell death, perhaps as a result of improper or incomplete localization. These studies also indicate that the a2 protein, argued on genetic grounds to be a negative regula- tor, acts in the yeast nucleus. Introduction The nucleus contains a discrete set of proteins synthesized in the cytoplasm (Gurdon, 1970; Bonner, 1975b; Feldherr, 1975; Merriam and Hill, 1976; De Robertis et al., 1978). The molecular basis by which these proteins accumulate in the nucleus remains to be elucidated (see Bonner, 1978; De Robertis, 1983 for review). Two models of nuclear localization are commonly considered. First, all proteins freely diffuse into the nucleus with subsequent retention of nuclear proteins by binding to a nondiffusible substrate. Second, certain proteins are selectively translocated across the nuclear envelope. Substantial evidence sug- gests that small macromolecules can diffuse into the nu- cleoplasm through pores in the nuclear envelope (Paine et al., 1975; Feldherr, 1965; Gurdon, 1970; Bonner, 1975a; Rechsteiner and Kuehl, 1979; Unwin and Milligan, 1982; Franke et al., 1981). Diffusion through a simple pore as the sole route of entry, though, is not sufficient to account for large proteins found in the nucleus. Nuclear pores have a functional radius of 45 A and an exclusion size limit of 0092~sS74/84/041057-0!3 !$02.00/0 approximately 70 kd for a globular protein. The kinetics of transport observed for large nuclear proteins (Feldherr, 1975; Bonner, 1975a; Yamaizumi et al., 1978; De Robertis et al., 1978) are difficult to reconcile with these pore dimensions. Nuclear localization of nuclear proteins may therefore involve more than the first model. The second model implies the existence of a “signal” in a nuclear protein that identifies it as a protein destined to be selectively translocated across the nuclear envelope. Dingwall et al. (1982) have recently presented evidence for such a signal. These investigators have shown that nucleoplasmin, a large, abundant protein of the Xenopus oocyte nucleus, can be cleaved into two fragments, one of which contains a determinant required for entry into the nucleus. Such a determinant is a candidate for the nuclear “signal se- quence” stipulated by the second model. In order to study how proteins are localized to the nucleus, we have taken a genetic approach with the yeast S. cerevisiae that has previously proven useful in bacteria (Bassford et al., 1979; Hall et al., 1982) the construction of gene fusions. We have constructed a set of gene fusions that code for hybrid proteins containing varying amounts of the yeast protein a2 at the amino terminus and a constant portion of E. coli P-galactosidase at the carboxy terminus. The a2 protein is a negative regulator of expres- sion of two sets of genes (Strathern et al., 1981; Sprague and Herskowitz, 1981; Nasmyth et al., 1981; Jensen et al., 1983; K. Wilson, personal communication) and hence was a candidate for a nuclear protein. Our rationale for identify- ing a localization determinant made the assumption that if a2 is a nuclear protein and fl-galactosidase is nonnuclear, then fusion of the two proteins could convert /3-galactosid- ase to a nuclear protein. Our analysis indicates that this assumption is correct. Localization studies demonstrate that a2 contains a signal at its amino terminus that is sufficient for targeting the heterologous @-galactosidase to the nucleus. Results Construction of MATaP-/acZ Hybrid Genes Fusions of the E. coli &galactosidase gene, /acZ, and the yeast gene MATol2 were constructed. The MATa gene was chosen for the following reasons. First, MATa has been extensively characterized (Strathern et al., 1981; Sprague and Herskowitz, 1981; Astell et al., 1981; Tatchell et al., 1981) and its product is possibly a nuclear protein. In a cells, a2 acts to turn off expression of a-specific genes, and in a/a cells it acts with the al product of MATa to turn off expression of a variety of genes (see Klar et al., 1981 and Jensen et al., 1983). Second, the MATa gene product is nonessential for growth. Third, the MATa gene is regulated-its expression is 5-fold higher in a haploid a cell than in an a/a diploid (Nasmyth et al., 1981). The use of a conditionally expressed gene allows one to test for a possible lethal phenotype caused by hybrid proteins, as previously observed in bacterial systems (Hall et al., 1982).
Transcript of Targeting of E. coli β-galactosidase to the nucleus in yeast

Cell, Vol. 36, 1057-1065, April 1984, Copyright Q 1984 by MIT
Targeting of E. coli ,&Galactosidase to the Nucleus in Yeast
Michael N. Hall,* Lynna Hereford,’ and ha HerskowiW *Department of Biochemistry and Biophysics University of California, San Francisco San Francisco, California 94143 +Dana Farber Cancer Research Center Harvard Medical School Boston. Massachusetts 02115
Summary
In order to identify determinants governing nuclear protein localization, we constructed a set of hybrid genes by fusing the S. cerevisiae gene, MATa2, coding for a presumptive nuclear protein, and the E. coli gene, /acZ, coding for B-galactosidase. The re- sultant hybrid proteins contain 3, 13, 25, 67, or all 210 amino acids of wild-type a2 protein at the amino terminus and a constant, enzymatically active por- tion of @-galactosidase at the carboxy terminus. Indirect immunofluorescence and subcellular frac- tionation studies with yeast cells containing the a2- LacZ hybrid proteins indicate that the a2 segment can direct localization of jI-galactosidase to the nu- cleus. A segment as small as 13 amino acids from a2 is sufficient for this localization. Comparison of amino acid sequences of other nuclear proteins with this region of a2 reveals a sequence that may be necessary for nuclear targeting. Production of some a2-LacZ hybrid proteins causes cell death, perhaps as a result of improper or incomplete localization. These studies also indicate that the a2 protein, argued on genetic grounds to be a negative regula- tor, acts in the yeast nucleus.
Introduction
The nucleus contains a discrete set of proteins synthesized in the cytoplasm (Gurdon, 1970; Bonner, 1975b; Feldherr, 1975; Merriam and Hill, 1976; De Robertis et al., 1978). The molecular basis by which these proteins accumulate in the nucleus remains to be elucidated (see Bonner, 1978; De Robertis, 1983 for review). Two models of nuclear localization are commonly considered. First, all proteins freely diffuse into the nucleus with subsequent retention of nuclear proteins by binding to a nondiffusible substrate. Second, certain proteins are selectively translocated across the nuclear envelope. Substantial evidence sug- gests that small macromolecules can diffuse into the nu- cleoplasm through pores in the nuclear envelope (Paine et al., 1975; Feldherr, 1965; Gurdon, 1970; Bonner, 1975a; Rechsteiner and Kuehl, 1979; Unwin and Milligan, 1982; Franke et al., 1981). Diffusion through a simple pore as the sole route of entry, though, is not sufficient to account for large proteins found in the nucleus. Nuclear pores have a functional radius of 45 A and an exclusion size limit of
0092~sS74/84/041057-0!3 !$02.00/0
approximately 70 kd for a globular protein. The kinetics of transport observed for large nuclear proteins (Feldherr, 1975; Bonner, 1975a; Yamaizumi et al., 1978; De Robertis et al., 1978) are difficult to reconcile with these pore dimensions. Nuclear localization of nuclear proteins may therefore involve more than the first model. The second model implies the existence of a “signal” in a nuclear protein that identifies it as a protein destined to be selectively translocated across the nuclear envelope. Dingwall et al. (1982) have recently presented evidence for such a signal. These investigators have shown that nucleoplasmin, a large, abundant protein of the Xenopus oocyte nucleus, can be cleaved into two fragments, one of which contains a determinant required for entry into the nucleus. Such a determinant is a candidate for the nuclear “signal se- quence” stipulated by the second model.
In order to study how proteins are localized to the nucleus, we have taken a genetic approach with the yeast S. cerevisiae that has previously proven useful in bacteria (Bassford et al., 1979; Hall et al., 1982) the construction of gene fusions. We have constructed a set of gene fusions that code for hybrid proteins containing varying amounts of the yeast protein a2 at the amino terminus and a constant portion of E. coli P-galactosidase at the carboxy terminus. The a2 protein is a negative regulator of expres- sion of two sets of genes (Strathern et al., 1981; Sprague and Herskowitz, 1981; Nasmyth et al., 1981; Jensen et al., 1983; K. Wilson, personal communication) and hence was a candidate for a nuclear protein. Our rationale for identify- ing a localization determinant made the assumption that if a2 is a nuclear protein and fl-galactosidase is nonnuclear, then fusion of the two proteins could convert /3-galactosid- ase to a nuclear protein. Our analysis indicates that this assumption is correct. Localization studies demonstrate that a2 contains a signal at its amino terminus that is sufficient for targeting the heterologous @-galactosidase to the nucleus.
Results
Construction of MATaP-/acZ Hybrid Genes Fusions of the E. coli &galactosidase gene, /acZ, and the yeast gene MATol2 were constructed. The MATa gene was chosen for the following reasons. First, MATa has been extensively characterized (Strathern et al., 1981; Sprague and Herskowitz, 1981; Astell et al., 1981; Tatchell et al., 1981) and its product is possibly a nuclear protein. In a cells, a2 acts to turn off expression of a-specific genes, and in a/a cells it acts with the al product of MATa to turn off expression of a variety of genes (see Klar et al., 1981 and Jensen et al., 1983). Second, the MATa gene product is nonessential for growth. Third, the MATa gene is regulated-its expression is 5-fold higher in a haploid a cell than in an a/a diploid (Nasmyth et al., 1981). The use of a conditionally expressed gene allows one to test for a possible lethal phenotype caused by hybrid proteins, as previously observed in bacterial systems (Hall et al., 1982).

Cell 1058
A
Figure 1. Construction of ~%fAT~24acZ Hybrid Genes
The 3.0 kb Sal I /acZ DNA fragment contains no transcription- or translation- initiation signals (Casadaban et al., 1983); these are provided by the gene, MATa2, into which this fragment is inserted. Wavy lines represent the MATal and MATa transcripts. The following restriction sites are indicated: Hind III, (H); Sal I, (S); and Xho I, (X). The three Xho I sites are on separate plasmids.
The E. coli IacZ gene was chosen for the following reasons. The /?-galactosidase monomer, of 116 kd, is larger than the exclusion size limit of nuclear pores. We therefore presume that entry of @-galactosidase into the nucleus cannot be attributed to diffusion. Second, the IacZ gene is procaryotic. The choice of a eucaryotic cytoplasmic protein would have required making the assumption that such a protein does not contain a determinant limiting it to the cytoplasm.
The MATcy24acZ gene fusions coding for hybrid proteins with a2 at the amino terminus and @galactosidase at the carboxy terminus were constructed in vitro (see Experi- mental Procedures). A Sal I restriction fragment containing the IacZ gene was inserted into the MATa gene at each of three differently situated Xho I sites (see Figure 1). These Xho I sites were generated by linker mutagenesis (Tatchell et al., 1981) and are unique to the plasmid. In two of these cases, the resultant constructs, 2%2-/acZ and 67a2-/acZ, are in-frame fusions encoding hybrid pro- teins in which the amino terminus of ,&galactosidase is replaced by the 25 and 67 amino terminal amino acids of (~2, respectively. In the third case, the MATa and IacZ sequences were out-of-frame. The reading frames were properly aligned by introducing a small Sl -mediated dele- tion at the fusion joint. The DNA sequence of one pre- sumptive in-frame fusion, 27fh2-/acZ, was determined and shown to contain an 11 bp deletion that properly aligns the MATol2 and /acZ reading frames (see Figure 2). The 210 amino acids of a2 at the amino terminus of the 21 Oa2-LacZ hybrid protein constitute the entire cu2 protein (as determined at the nucleic acid level); the fusion joint falls within the termination codon of the MATcu2 gene.
Hybrids containing fewer than 25 amino acids of (~2 were obtained by treating the fusion joint of 2%2-/acZ with exonuclease Bal 31. Deletions that extend only into MATa DNA but not into IacZ were screened by restriction analysis (see Experimental Procedures). The DNA se-
I 2 1 k 9
3a.?-tocz GAAbAA G& t& Ah A:G Gh & Gk I , BanHI-
MATa2’ @ ‘/ocr
Figure 2. DNA Sequences of the Fusion Joints of MATa2-/acZ Hybrid Genes
MATa and /acZ are underlined and labeled. The DNA that is not underlined is polylinker DNA. Amino acids are indicated by the single-letter code. The number below an amino acid indicates the position of the amino acid in the respective wild-type protein. (A) The MATa and the /acZ reading frames in the 2100124acZ hybrid genes were properly aligned by introducing an Sl-mediated 11 bp deletion at the Sma I site. (6) The 3n24acZ and 13012~/acZ fusions were generated by Bal 31 -mediated, unidirectional dele-
tions of 90 and SO bp, respectively, from the Sma I site of the 250124acZ hybrid.
quence determination of two shorter fusions, 3a2-IacZ and 73&acZ, revealed deletions of 90 and 60 bp, resulting in hybrid proteins that contain 3 and 13 amino acids of a2, respectively (see Figure 2). In all five fusions described above, the fusion joint within /acZ is constant, falling within the eighth codon. In the case of the hybrid proteins, the amino-terminal eight amino acids of /3-galactosidase are replaced by the indicated number of amino-terminal a2 amino acids: 3, 13, 25, 67, or 210 (see Figure 3).
The construction of fusions as described above results in hybrid genes with the following features. Since the fusion is generated by insertion, the 3’ end of the MATcu2 gene is still present. Consequently, the proper transcription- termination signals of the MATa gene are still available; translation terminates at the /acZ termination codons (see Figure 1). Also available is DNA homology at both ends of the hybrid gene with regard to the wild-type MATcY~ gene (see Figure l), which facilitates gene replacement (see below).
Regulation of MATaP-lacZ Fusions In order to confirm the previously observed regulation of MATa (Nasmyth et al., 1981) and to determine whether MATol24acZ hybrid genes are regulated, isogenic aIcy, (Y, and a strains containing plasmid-borne fusions were as-
Nuclear Protein Localization 1059
a2 N+’ a2 210 Coo” 3a2-LacZ 2 LrlCZ
13a2-LocZ 2 LCCZ
25c&LacZ = Lacz
67a2-LCCZ 67 LOCZ
2loa2-Lacz no Lacz
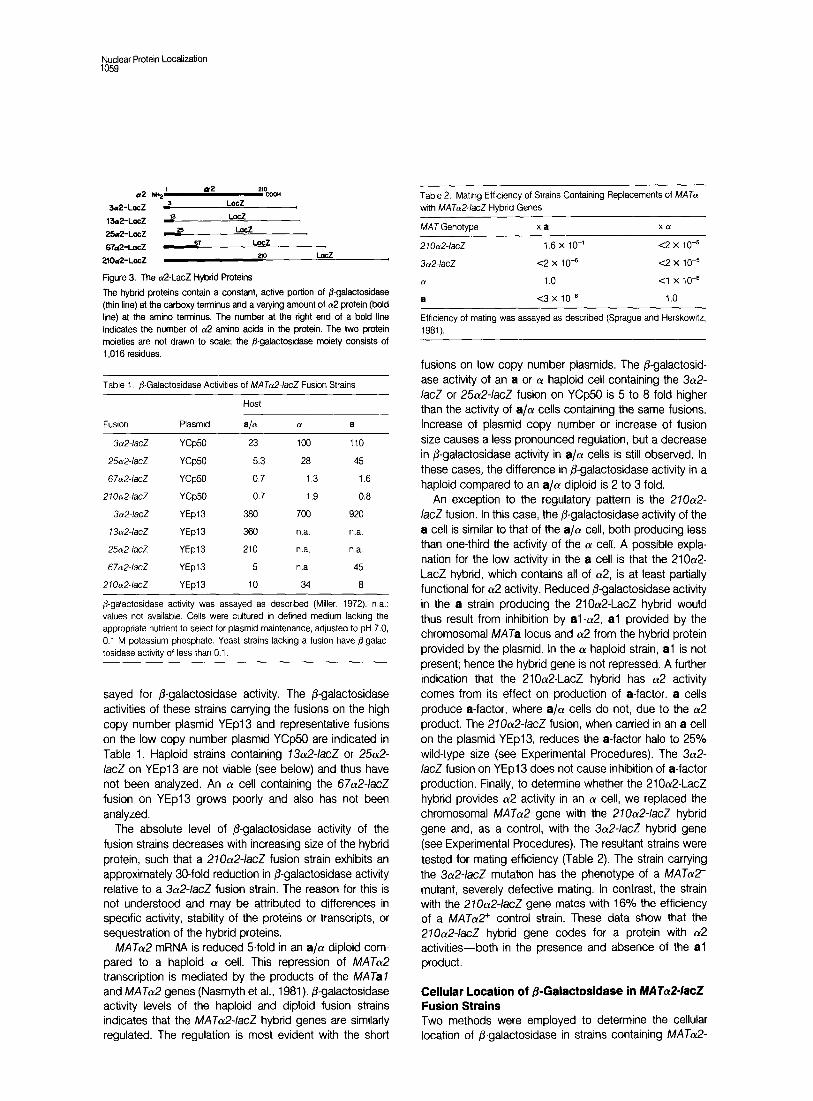
Figure 3. The UP-LacZ Hybrid Proteins
The hybrid proteins contain a constant, active portion of flgatactosidase (thin line) at the carboxy terminus and a varying amount of a2 protein (bold line) at the amino terminus. The number at the right end of a bold line indicates the number of a2 amino acids in the protein. The two protein moieties are not drawn to scale; the @-gafactosidase moiety consists of 1 ,016 residues.
Table 1 /$Galactosidase Actrvities of MAT&-/acZ Fusion Strains
Host
Fusron
3&/acZ
25ru2-/acz
67GlacZ
2 1 Oa2-/acZ
3Lu2-/acZ
1302./acZ
25dlacZ
67&/acZ
210&lacZ
Plasmid
YCp50
YCp50
YCp50
YCp50
YEpI
YEpl3
YEp13
YEpI
YEp13
a/a a a
23 100 110
5.3 28 45
0.7 1.3 1.6
0.7 1.9 0.8
380 700 920
360 n.a. na.
210 n.a. n.a.
5 n.a. 45
10 34 0
&galactosidase actrvity was assayed as described (Miller, 1972). n.a.: values not available. Cells were cultured in defined medium lacking the appropriate nutrient to select for plasmid maintenance, adjusted to pH 7.0, 0.1 M potassrum phosphate. Yeast strains lacking a fusion have @-galac- tosidase actrvity of less than 0.1.
sayed for 6-galactosidase activity. The 6-galactosidase activities of these strains carrying the fusions on the high copy number plasmid YEpI and representative fusions on the low copy number plasmid YCp50 are indicated in Table 1. Haploid strains containing 73a2-/acZ or 25012- /acZ on YEpI are not viable (see below) and thus have not been analyzed. An (Y cell containing the 67a2-/acZ fusion on YEpl3 grows poorly and also has not been analyzed.
The absolute level of P-galactosidase activity of the fusion strains decreases with increasing size of the hybrid protein, such that a 270~12~/acZ fusion strain exhibits an approximately 30-fold reduction in /3-galactosidase activity relative to a 3a2-/acZ fusion strain. The reason for this is not understood and may be attributed to differences in specific activity, stability of the proteins or transcripts, or sequestration of the hybrid proteins.
MATa mRNA is reduced 5fold in an a/a! diploid com- pared to a haploid (Y cell. This repression of MATol2 transcription is mediated by the products of the MATal and MATa genes (Nasmyth et al., 1981). P-galactosidase activity levels of the haploid and diploid fusion strains indicates that the MATot24acZ hybrid genes are similarly regulated. The regulation is most evident with the short
Table 2. Mating Efficiency of Strains Containing Replacements of MATa wrth MATaP-/acZ Hybrid Genes
MAT Genotype xa Xl?
270&lacZ 1.6 x IO-’ <2 x 10-S
3&/acZ c2 x 10-S -=2x w
a 1 .o <I x 104
a <3x 104 1 .o
Efficiency of mating was assayed as described (Sprague and Herskowrtz, 1981).
fusions on low copy number plasmids. The P-galactosid- ase activity of an a or 01 haploid cell containing the 3012- /acZ or 25x2~/acZ fusion on YCp.50 is 5 to 8 fold higher than the activity of a/cu cells containing the same fusions. Increase of plasmid copy number or increase of fusion size causes a less pronounced regulation, but a decrease in @galactosidase activity in a/a cells is still observed. In these cases, the difference in /?-galactosidase activity in a haploid compared to an a/a diploid is 2 to 3 fold.
An exception to the regulatory pattern is the 21&G /acZ fusion. In this case, the 6-galactosidase activity of the a cell is similar to that of the a/a cell, both producing less than one-third the activity of the (Y cell. A possible expla- nation for the low activity in the a cell is that the 210a2- LacZ hybrid, which contains all of a2, is at least partially functional for a2 activity. Reduced /3-galactosidase activity in the a strain producing the 210a2-LacZ hybrid would thus result from inhibition by ak2, al provided by the chromosomal MATa locus and ot2 from the hybrid protein provided by the plasmid. In the (Y haploid strain, al is not present; hence the hybrid gene is not repressed. A further indication that the 210a2-LacZ hybrid has (~2 activity comes from its effect on production of a-factor. a cells produce a-factor, where a/a cells do not, due to the (~2 product. The 210a24acZ fusion, when carried in an a cell on the plasmid YEpl3, reduces the a-factor halo to 25% wild-type size (see Experimental Procedures). The 3a2- 1acZ fusion on YEpl3 does not cause inhibition of a-factor production. Finally, to determine whether the 210~y2-LacZ hybrid provides 012 activity in an (Y cell, we replaced the chromosomal MATa gene with the 2lOa2-/acZ hybrid gene and, as a control, with the 3a24acZ hybrid gene (see Experimental Procedures). The resultant strains were tested for mating efficiency (Table 2). The strain carrying the &2-/acZ mutation has the phenotype of a MAToG’- mutant, severely defective mating. In contrast, the strain with the 270GlacZ gene mates with 16% the efficiency of a MATcY~+ control strain. These data show that the 210GlacZ hybrid gene codes for a protein with (~2 activities-both in the presence and absence of the al product.
Cellular Location of @Galactosidase in MATa24acZ Fusion Strains Two methods were employed to determine the CSllUlSr
location of @galactosidase in strains containing MATot2-

Cell 1060
Figure 4. Indirect lmmunofluorescent Staining of Cells with Anti-&Galactosidase IgG
Representative distribution of @-gatactosidase in cells containing gene fusions 210a2-lacZ (A); 25012~/acZ (C); 13012~/acZ (E); 3a2-/acZ (G). (B- H) Nuclear fluorescence of DNA-staining DALI. The 27O~&/acZ and 3a2-/acZ fusions are in strain HR125-5Da. The 25n2-/acZ and 73&/acZ fu- sions are in strain HR1255Da/a. The plasmid carrying the fusions is YEpl3. Primary antibody concentration is 30 pg/ml. Bar is equivalent to IO
P.
/acZ fusions: immunofluorescence and cell fractionation. Strains containing the fusion on a high copy number 2 p plasmid and a low copy number centromere plasmid were examined by both methods. The copy number of the plasmid bearing the MATa2-/acZ fusion did not affect the results. Haploid cell type or a/a diploidy of the host strain also did not affect the results.
The results of the immunofluorescence experiments are shown in Figure 4. Cells were simultaneously treated with DAPI to stain nuclei and with antibodies to detect the hybrid proteins (see Experimental Procedures). Approxi- mately 50 stained cells from each fusion strain were examined and all yielded a similar result for a particular fusion. The strains containing hybrids 21 Oa2-LacZ, 67a2- LacZ (data not shown), 25a2-LacZ, and 13a2-LacZ exhibit
fluorescence localized to the nucleus. The 3a2-LacZ hybrid protein is dispersed throughout the cell with no evident concentration in the nucleus. These results indicate that the 13 amino-terminal residues of the a2 protein are suffi- cient for targeting E. coli P-galactosidase to the nucleus.
The cellular location of /3-galactosidase was also deter- mined by fractionating cells and comparing P-galactosid- ase activity in the cytosol and nuclear fractions. Nuclei were separated from the cytosol by centrifuging a lysate through a Percoll, isopycnic gradient under conditions in which the cytosol remains at the top of the gradient and the nuclei form a discrete, visible band at an intermediate density (see Experimental Procedures). The Percoll gra- dient was fractionated and individual fractions were as- sayed for /I-galactosidase activity (see Figure 5). The p-
Nuclear Protein Localization 1081
cyto. WC.
_t t Table 3. Germination Frequencies of Spores Containing MATcr.2/acZ Fusions II A
E 2’oa2-Locz
2 B :: 25cr2-LocZ &
FL 3a2-Lscz
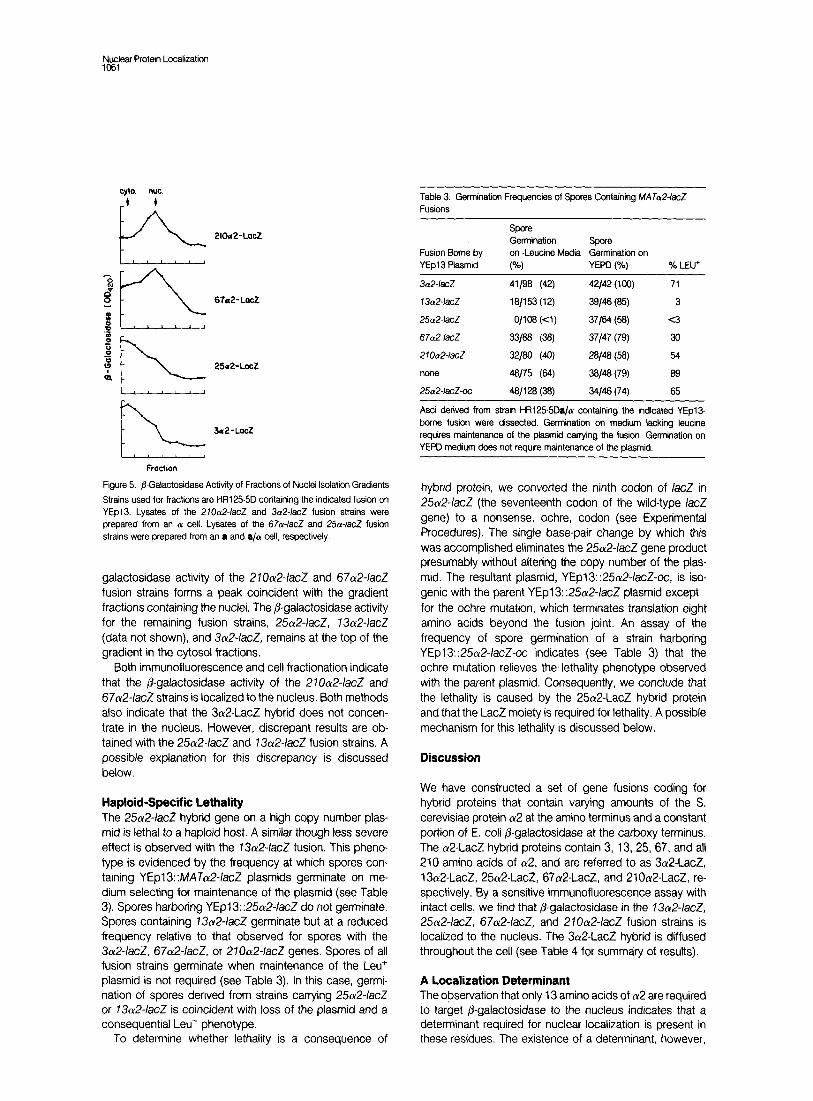
Figure 5. @Gatactosidase Activity of Fractrons of Nuclei Isolation Gradients
Strains used for fractions are t-R1255D containing the indicated fusion on YEpl3. Lysates of the 210&lacZ and 3a2-/acZ fusion strains were prepared from an a cell. Lysates of the 67a-/acZ and 2.5~~lacZ fusion strains were prepared from an a and a/a cell, respectively.
galactosidase activity of the 270a2-IacZ and 67a2-IacZ fusion strains forms a peak coincident with the gradient fractions containing the nuclei. The P-galactosidase activity for the remaining fusion strains, 25ot2-/acZ, 13a2-/acZ (data not shown), and 3~12~/acZ, remains at the top of the gradient in the cytosol fractions.
Both immunofluorescence and cell fractionation indicate that the /3-galactosidase activity of the 21Oa2-IacZ and 67a2-/acZ strains is localized to the nucleus. Both methods also indicate that the 3Lu2-LacZ hybrid does not concen- trate in the nucleus. However, discrepant results are ob- tained with the 2&2-/acZ and 73a2-IacZ fusion strains. A possible explanation for this discrepancy is discussed below.
Haploid-Specific Lethality The 25a2-IacZ hybrid gene on a high copy number plas- mid is lethal to a haploid host. A similar though less severe effect is observed with the 13a2-/acZ fusion. This pheno- type is evidenced by the frequency at which spores con- taining YEpl3: MA Jot24acZ plasmids germinate on me- dium selecting for maintenance of the plasmid (see Table 3). Spores harboring YEpl3::2%2-IacZ do not germinate. Spores containing 13a2-/acZ germinate but at a reduced frequency relative to that observed for spores with the 3a2-/acZ, 67ct2-/acZ. or 270a2-/acZ genes. Spores of all fusion strains germinate when maintenance of the Leu+ plasmid is not required (see Table 3). In this case, germi- nation of spores derived from strains carrying 25a2-/acZ or 13Lu2-/acZ is coincident with loss of the plasmid and a consequential Leu- phenotype.
To determine whether lethality is a consequence of
Fusion Borne by YEpI Plasmid
Spore Germination Spore on -Leucine Media Germination on
(“4 YEPD (%) % LEu+
3~x2~far9 41/C@ (42) 42/42 (100) 71
13~x2~lacZ 18/153 (12) 39/48 (85) 3
2&2-/acZ o/108 (Cl) 37164 (W <3
67c&/acZ 33188 (W 37147 (79) 30
2lOCx2-IaCZ 32/80 (40) 28/B W) 54
none 48/75 (W w* (7% 89
2&x2-lacZuc 48/l 28 (38) 34146 (74) 65
Asci derived from strain HR125-5Da/a! containing the indicated YEpl3- borne fusion were dissected. Germination on medium lacking leucine requires maintenance of the plasmid carrying the fusion. Germination on YEPD medium does not require maintenance of the plasmid.
hybrid protein, we converted the ninth codon of IacZ in 25a24acZ (the seventeenth codon of the wild-type IacZ gene) to a nonsense, ochre, codon (see Experimental Procedures). The single base-pair change by which this was accomplished eliminates the 25a2-IacZ gene product presumably without altering the copy number of the plas- mid. The resultant plasmid, YEpl3::25u2-IacZ-oc, is iso- genie with the parent YEpl3: :25a2-/acZ plasmid except for the ochre mutation, which terminates translation eight amino acids beyond the fusion joint. An assay of the frequency of spore germination of a strain harboring YEpl3::25&lacZ-oc indicates (see Table 3) that the ochre mutation relieves the lethality phenotype observed with the parent plasmid. Consequently, we conclude that the lethality is caused by the 25a2-LacZ hybrid protein and that the LacZ moiety is required for lethality. A possible mechanism for this lethality is discussed below.
Discussion
We have constructed a set of gene fusions coding for hybrid proteins that contain varying amounts of the S. cerevisiae protein a2 at the amino terminus and a constant portion of E. coli fi-galactosidase at the carboxy terminus, The a2-LacZ hybrid proteins contain 3, 13, 25, 67, and all 210 amino acids of (~2, and are referred to as 3a2-LacZ, 13a2-LacZ, 25a2-LacZ, 67a2-LacZ, and 21 Oa2-LacZ, re- spectively. By a sensitive immunofluorescence assay with intact cells, we find that @-galactosidase in the 73cu2-/acZ, 25a2-/acZ, 67o12-/acZ, and 210&/acZ fusion strains is localized to the nucleus. The 3a2-LacZ hybrid is diffused throughout the cell (see Table 4 for summary of results).
A Localization Determinant The observation that only 13 amino acids of a2 are required to target P-galactosidase to the nucleus indicates that a determinant required for nuclear localization is present in these residues, The existence of a determinant, however,
CM 1062
Table 4. Summary of Properties of Hybrid Proteins
Nuclear Localization: Haploid
Hybrid MW (kd) IIF Fractionation Lethality
3a2-LacZ 116 - -
13a2-LacZ 117 + +
25&LacZ 119 + +
67&LacZ 123 + +
210012.LacZ 137 + + -
Nuclear localization of hybrid protein was determined by two methods, indirect immunofluorescence (IF) was subcellular fractionation.
does not allow us to distinguish between the two models of nuclear localization. According to a model of selective uptake, the determinant within the 13 amino acids is a signal recognized by the nuclear envelope in some man- ner. A signal of this type is analogous to signal sequences commonly found at the amino terminus of extracytoplasmic proteins (Kreil, 1981). There are no sequence similarities between these signal sequences and the amino-terminal 13 residues at (~2. According to a model of passive diffusion and selective retention by substrate binding, the amino-terminal 13 residues of cu2 would be involved in binding a substrate once the protein has diffused into the nucleoplasm.
Although we cannot rule out either model of nuclear protein localization, the following points should be consid- ered. First, the localized hybrids, ranging in molecular weight from 117 to 137 kd, are larger than the exclusion size limit of nuclear pores (70 kd). Assuming that yeast nuclear pores have the same physical characteristics as the pores in the cells from which they have been charac- terized, the hybrids should not be able to diffuse freely into the nucleus. Little is known about nuclear pores in yeast other than that they exist (Robinow and Marak, 1966; Byers, 1981). Second, only 13 amino acids of a2 are necessary for localization. According to a retention model, the 13a2-LacZ hybrid must bind a nuclear substrate de spite missing 197 of 210 a2 amino acids, which is possible but not expected.
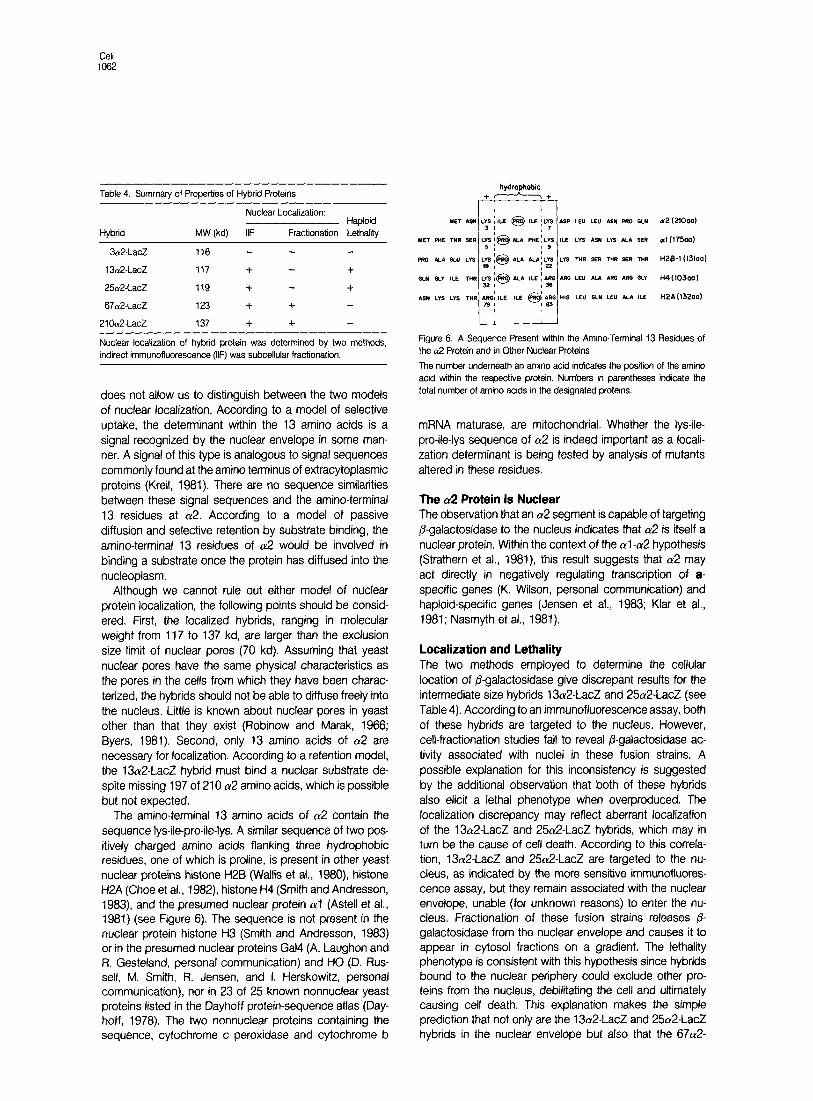
The amino-terminal 13 amino acids of a2 contain the sequence lys-ile-pro-ile-lys. A similar sequence of two pos- itively charged amino acids flanking three hydrophobic residues, one of which is proline, is present in other yeast nuclear proteins histone H2B (Wallis et al., 1980) histone H2A (Choe et al., 1982) histone H4 (Smith and Andresson, 1983) and the presumed nuclear protein (~1 (Astell et al., 1981) (see Figure 6). The sequence is not present in the nuclear protein histone H3 (Smith and Andresson, 1983) or in the presumed nuclear proteins Gal4 (A. Laughon and Ft. Gesteland, personal communication) and HO (0. Rus- sell, M. Smith, Ft. Jensen, and I. Herskowitz, personal communication), nor in 23 of 25 known nonnuclear yeast proteins listed in the Dayhoff protein-sequence atlas (Day- hoff, 1978). The two nonnuclear proteins containing the sequence, cytochrome c peroxidase and cytochrome b
hydrophobic +-+
a2 (2l000)
al U75aal
HZ&1 (131aol
H4(103ool
H2A(132aa)
Figure 6. A Sequence Present within the Amino-Terminal 13 Residues of the a2 Protein and in Other Nuclear Proteins
The number underneath an amino acid indicates the position of the amino acid within the respective protein. Numbers in parentheses indicate the total number of amino acids in the designated proteins.
mRNA maturase, are mitochondrial. Whether the lys-ile- pro-ile-lys sequence of (~2 is indeed important as a locali- zation determinant is being tested by analysis of mutants altered in these residues.
The (r2 Protein Is Nuclear The observation that an (~2 segment is capable of targeting @-galactosidase to the nucleus indicates that (~2 is itself a nuclear protein. Within the context of the al-n2 hypothesis (Strathern et al., 1981) this result suggests that (~2 may act directly in negatively regulating transcription of a- specific genes (K. Wilson, personal communication) and haploid-specific genes (Jensen et al., 1983; Klar et al., 1981; Nasmyth et al., 1981).
Localization and Lethality The two methods employed to determine the cellular location of /I-galactosidase give discrepant results for the intermediate size hybrids 13a2-LacZ and 25cu2-LacZ (see Table 4). According to an immunofluorescence assay, both of these hybrids are targeted to the nucleus. However, cell-fractionation studies fail to reveal /I-galactosidase ac- tivity associated with nuclei in these fusion strains. A possible explanation for this inconsistency is suggested by the additional observation that both of these hybrids also elicit a lethal phenotype when overproduced. The localization discrepancy may reflect aberrant localization of the 13a2-LacZ and 25a2-LacZ hybrids, which may in turn be the cause of cell death. According to this correla- tion, 13a2-LacZ and 25cY2-LacZ are targeted to the nu- cleus, as indicated by the more sensitive immunofluores- cence assay, but they remain associated with the nuclear envelope, unable (for unknown reasons) to enter the nu- cleus. Fractionation of these fusion strains releases p- galactosidase from the nuclear envelope and causes it to appear in cytosol fractions on a gradient. The lethality phenotype is consistent with this hypothesis since hybrids bound to the nuclear periphery could exclude other pro- teins from the nucleus, debilitating the cell and ultimately causing cell death. This explanation makes the simple prediction that not only are the 13a2-LacZ and 25a2-LacZ hybrids in the nuclear envelope but also that the 67cu2-

Nuclear Protein Localization 1063
LacZ and 210a2-LacZ are in the nucleoplasm. A method more sensitive than immunofluorescence with intact cells is required to test this prediction. Genetic evidence sup- porting the contention that at least 2lOcu2-LacZ is in the nucleoplasm is the observation that it exhibits (~2 activity (see Results), assuming that the (~2 activity is not the result of 21 Oa2-LacZ proteolysis.
A second explanation for the lethal phenotype does not require that there be any difference in localization between the intermediate size hybrids and the larger hybrids (67~~2- LacZ and 210a2-LacZ). Perhaps the intermediate size hybrids result in cell death simply because they are pro- duced at a higher level than the larger hybrids, not because they are intrinsically different. Lethality might result from disruption of the nuclear envelope due to the overproduc- tion of a harmful protein. This explanation, however, does not account for the discrepant localization results obtained with the 1302-LacZ and 2&2-LacZ hybrids.
In a parallel analysis (R. Moreland and L. Hereford, unpublished), it has been observed that a yeast histone H2B-/I-galactosidase hybrid protein is localized to the nu- cleus. The use of fusions between /3-galactosidase and a2, histones, and other nuclear proteins ought to make it possible to determine whether these proteins are localized to the nucleus according to similar rules.
Strains, Plasmids, and Media The yeast strains used in all experiments, unless otherwise indicated, are from an isogenic a/a, a, and a set of HR1255D: leu2 ura3-52 trpl his3 his4 (Ft. Jensen, Ph.D. thesis, University of Oregon, 1983). W301-30A is MATa leu2-3.7 12 trpl-1 canl-100 ura3-1 his3-11.15 ade2-1 SUP40 (R. Rothstein). The bacterial strains used are as follows. MC1061 is a&739 @a, leu)7697 lacX74 ga/U galK hsr rpsL (MC strains are described in Casadaban et al., 1983). MHl, afaD lacX74 ga/U gal/C hsr rpsL, is a Leu+ Pl transductant of MClC61. MH3, MHl frpC9830. is derived from MC1064 MH4, lacX74 ga/fJ ga/K hsr rp.sL /euB600, is derived from MC1065 MH7901 is F’kZU7 18 Tn5/MS365 (Hall and Silhavy, 1981). The relevant features of the yeast plasmids used are as follows. YRp7 is amp’ TRP7 A/El (Struhl et al., 1979). YEpl3 is amp’ LEU2 2~ (Broach et al., 1979). YCpSO is amp’ WA3 CEr’v4 A/W (C. Mann, personal communi- cation).
Bacterial media and manipulations are described by Miller (1972). Kan- amycin was used at a concentration of 40 pg/ml. Yeast media and manipulations are described by Sherman et al. (1979). Medra containing 5 Bromo-4-chloro-3-indolyl-@-D-galactosidase (XG) was prepared as de- scribed by Rose et al. (1981).
Assays Production of a-factor was measured by a halo assay (L. Blair, personal communication). Strains to be tested for a-factor were replica-plated onto medium previously spread with a strain RC757 (Chan and Otte, 1982) that is hypersensitive to the mating factor of the opposite mating type. The amount of a-factor produced by a strain is indicated by the size of the halo, a zone of growth inhibition, surrounding the strain being tested. Efficiency of mating assays were performed as described by Sprague and Herskowitz (1981). P-galactosidase activity was assayed as described by Miller (1972). Cells to be assayed for @-galactosidase activity were cultured in the appropriate medium adjusted to pH 7.0, 0.1 M KPO,, and permeabilized with chloroform and sodium dodecyl sulfate (SDS).
Genetic Manipulations The /acZ ochre mutation U118 was recombined in VIVO onto the MAT25a2- /acZ fusron in E. coli as follows. The F’lacZUI 18 Tn5 from strain MH7901
was transferred into MC1061 harboring YEpl3::25a2-/acZ. Exconjugants were selected as resistant to both kanamycin and ampicillin. A Kan’ Amp’ exconjugant was then used as a donor in a mating with led?- strain MH4. Exconjugants from this mating were selected as Leu* and screened for a Lac- (white) phenotype on M63 glucose media containing XG. The Leu+ selection required that YEpl3: :25a2-/acZ be mobilized as a cointegrant with the F’lscZUl78 Tn5. Exconjugants were screened for white colony color since such a colony would resuft from recombination of the U118 ochre mutation from the F’ onto the /acZ of YEpl3::25&/acZ. White exconjugants appeared at an approximate frequency of 0.1%. Plasmid DNA was prepared from these exconjugants. introduced into bacterial strain MHI, reisolated, and shown to have the same restriction pattern as the Lac+ parent plasmid. The presence of the ochre mutation was verified by transforming a SUP4 ochre suppressor yeast strain (W30-30A) with YEpl3: :25&/acZUl18. The resultant transformants exhibited a Lac+ phe- notype (blue colony color on XG indicator medium). Transformants of an isogenic sup4+ strain were Lac-.
The MATa gene of HR125-5Da was replaced with a MAT&/acZ hybrid gene by the one-step gene-disruption method (Rothstein, 1983) with the following modification. Hind Ill-digested YRp7::MAT&/acZ (see Figure 1) was cotransformed with intact Leu+ plasmid YEpl3. Leu+ transformants were selected, collected in suspension, and spread on medium containing both leucine and XG. Stable blue transformants comprised approximately 1% of all transformants. Presence of the MAT&lad fusion at MAT was confirmed by allelism tests and by a defective matrng phenotype.
Hybrid Gene Constructii The MAr~G/acz hybrid genes were constructed as illustrated in Figures 1 and 2. A 3 kb Sal I /acZ fragment from pMC1871 (Casadaban et al., 1983) was ligated into the MATa gene at each of three Xho I sites. Each recipient YRp7mafa2::Xho I plasmid in this manipulation contained a unique and differently Xho I site within the MATa gene since each site had been generated by Xho I linker mutagenesis (Tatchell et al., 1981). The 270a2- /acZ, 67012-/acZ, and 25&/acZ fusions were made by ligation into the ax75 ax38, and ax183 Xho I linker insertion mutations, respectively. Insertion of the /acZ Sal I fragment into the Xho I sites ax38 and 01x183 results in in-frame MAT&/acZ fusions, whrch were detected as blue Lac+ Trp’ colonies subsequent to transformation of MH3. Insertion of the /acZ Sal I fragment into the Xho I site ~1x75, however, does not yield an in-frame fusion. Consequently, the initial construct conferred a white Lac- phenotype In MH3. The MATct2 and /acZ reading frames were aligned by introducing a small deletion at the fusion joint of the MATaP and /acZ DNAs. The deletion was produced by cleavage of the Lac- YRp7-ax75-/acZ plasmid at a unique Sma I sate within the fusion joint (see Figure 2) digestion with nuclease Si, ligation, and transformation of MHl. Plasmids bearing presum- ably in-frame 210a2-/acZ fustons were detected as blue Lac+ transformants on medium containing XG. Candidates were checked for loss of the Sma I site. The DNA sequence of the fusion jornt of one candidate was subse- quently determined (see Figure 2).
The 13012./acZ and 3a2-/acZ fusions were produced by introducing deletions at the fusion joint of 25a2-/acZ. YRp7::25&/acZ was cleaved at the Sma I site within the fusion joint, treated with Bal 31, ligated, and transformed into MHI. Plasmrd DNA was prepared from transformants that exhibited a blue Lac+ phenotype on medium containing XG. In order to detect deletions that extended only into MAk2, the DNA was screened for loss of the Eco RI site and conservation of a Bam HI site within the fusion joint (see Figure 2). Two candidates that met these prerequisites and contained deletions of approximately the desired sizes were further char- acterized by DNA sequence determination of the fusion joints.
DNA sequences were determined by the dideoxy method @anger et al., 1977). Hpa I-Eco RV fragments spanning the fusion joints were isolated for use as templates, A pentadecamer (gift of Joe Hedgpeth) that hybridizes to the region of /acZ corresponding to the twelfth through seventeenth amino acid of wild-type 8-galactosidase was used as a primer.
The MATaP-/acZ hybrid genes were transferred to YEpl3 and YCp50 from YRp7 by the Hand Ill restriction sates flanking MATO (see Figure 1).
Fluorescence Microscopy Fluorescence microscopy was performed as described by Kilmartrn et al. (1982) with the following modifications (M. Rozdziel and R. Moreland,

Cell 1064
personal communication). Cells in early log phase (2-4 x 106 cells/ml) were harvested by centrifugation, washed once with 1.2 M sorbitol, and digested for 1.5 hr at 3o°C with Zymolyase 5006 (0.5 mg/ml) and Glusulasa (20 As/ml). An equal volume of 6.7% formaldehyde in 1.2 M sorbffol was added. Spheroplasts were incubated at 0°C on ice for 10 min, centrifuged, washed once with 1.2 M sorbitol, and resuspended at approximately IO9 cells/ml in 1.2 M sorbitol. All subsequent steps (preparation of slides, fixation of cells, incubation with antibodies, and purification of primary and secondary antibodies) were performed according to the procedure of Kilmartin et al. (1982). Following incubation with the secondary antibody, the cells were incubated for 5 min with 4,6diamidino-2-phenyfindole @API) (0.5 mg/ml) in PBS (0.5 M KPD,, pH 7, 0.15 M NaCI, 0.1% azide, 1% bovine serum albumin), washed once with PBS, and then mounted in 95% glycerol, 0.01% PBS, pH 9, containing 0.5 mg/ml pphenylenediamine.
Rabbit anti-p-galactosidase antibody was the generous gift of Beth Rasmussan. FITC-anti-rabbit goat IgG was obtained from Cappel Labora- tories
lsolatlon of fduclei Nuclei were isolated as described (Ide and Saunders, 1981; Nagy et al., 1982). Cells harboring a MAT&/scZ gene fusion on YEpI or YCp50 were grown to O.Dem of 1.0 at 30°C in defined medium lacking the appropriate nutrient (leucine and uracil, respectively) to maintain the plas- mid. Lysates were loaded on 30% Percoll gradients. Gradients were fractionated into 1 ml fractions and assayed for j+galactosidase activity as described (Miller, 1972). Fractions containing nuclei were identified by their turbidity. Cytosol material remains at the top of the gradient.
We would like to thank Kelly Tatchell and Malcolm Casadaban for gener- ously providing strains and plasmids, Joe Hedgpeth for teaching dideoxy sequencing, Mike Stern, Sandy Johnson, Stan Fields, Flora Banuett, and Douglass Forbes for comments on the manuscript, and Sabine Carrere and Wendy Blankfort-Doyle for help in the preparation of the manuscript. Lynna Hereford would like to thank Moshe Rozdziel and John Kilmarfin for help with developing the immunofluorescence assay. The research was supported by Research Grants from the Public Health Service to I. H. and to L. H. M. N. H. was the recipient of a Helen Hay Whitney Fellowship.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
Received December 9, 1983; revised February 3. 1984
References
Astell, C. R., Ahlstrom-Jonasson, L., Smith, M., Tatchell, K., Nasmyth, K. A., and Hall, 8. D. (1981). The sequence of the DNAs coding for the mating- type loci of Saccharomyces cerevisiae. Cell 27, 15-23.
Bassford. P. J., Jr., Silhavy, T. J., and Beckwith. J. R. (1979). Use of gene fusion to study secretion of maltose-binding proteins into Escherichia co/i periplasm. J. Bacterial. 139, 19-31.
Banner, W. M. (1975a). Protein migration into nuclei. I. Frog occyte nuclei in vivo accumulate microinjected histones. allow entry to small proteins, and exclude large proteins. J. Cell Biol. 64, 421-430.
Bonner, W. M. (1975b). Protein migration into nuclei. II. Frog oocyte nuclei accumulate a class of microinjected oocyte nuclear proteins and exclude a class of microinjected oocyte cytoplasmic proteins. J. Cell Biol. 64, 431- 437.
Bonner, W. M. (1978). Protein migration and accumulation in nulcei. In The Cell Nucleus, 6, H. Busch, ed. (New York: Academic Press), pp. 97-148.
Broach, J. R., Strathern, J. N., and Hicks, J. B. (1979). Transformation in yeast: development of a hybrid cloning vector and isolation of the CAN1 gene. Gene 8, 121-133.
Byers, B. (1981). Cytology of the yeast life cycle. In The Molecular Biology of Yeast Saccharomyces: Life Cycle and Inheritance. J. N. Strathern, E. W.
Jones, and J. R. Broach, eds. (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory), pp. 59-96.
Casadaban, M. J., Martinez-Arias, A, Shapira, S. K., and Chou, J. (1983). 8galactosidase gene fusion for analyzing gene expression in Escherichia co/i and yeast. Meth. Enzymol. 100, 293-308.
Chan, R. K., and Dtte, C. A. (1982). Isolation and genetic analysis of Saccbaromyces cefevisise mutants supersensitive to Gi arrest by a-factor and a-factor pheromones. Mol. Cell. Biol. 2, 11-20.
Choe, J., Kolodrubetz, D., and Grunstein, M. (1982). The two yeast histone H2A genes encode similar protein subtypes, Proc. Nat. Acad. Sci. USA 79, 1484-1487.
Dayhoff, M. 0. (1978). Atlas of protein sequence and structure. Nat. Biomed. Res. Found. Washington, D.C. vol. 5 suppl. 3.
De Robertis, E. M. (1983). Nucleocytoplasmic segregation of proteins and RNAs. Cell 32, 1021-1025.
De Robertis, E. M., Longthorne, R. F., and Gurdon, J. B. (1978). Intracellular migration of nuclear proteins in Xenopus ocytes. Nature 272, 254-256.
Dingwall, C., Sharnick, S. V., and Laskey, R. A. (1982). A polypeptide domain that specifies migration of nucleoplasmin into the nucleus. Cell 30, 449-458.
Feldherr. C. M. (1965). The effect of the electron-opaque pore material on exchanges through the nuclear annuli. J. Cell Biol. 25, 43-53.
Feldherr, C. M. (1975). The uptake of endogenous proteins by oocyte nuclei. Exp. Cell Res. 93, 41 l-419.
Franke, W. W., Scheer, U., Krohne, G., and Jarasch, E. D. (1981). The nuclear envelope and the architecture of the nuclear periphery. J. Cell Biol. 91,39s-50s.
Gurdon, J. 8. (1970). Nuclear transplantation and the control of gene activity in animal development. Proc. Roy. See. Lond. B. 776, 303-314.
Hall, M. N., and Silhavy. T. J. (1981). The on@ locus and the regulation of the major outer membrane porin proteins of Escherichia co/i K12. J. Mol. Biol. 146, 23-43.
Hall, M. N., Schwartz M., and Silhavy, T. J. (1982). Sequence information within the lamb6 gene is required for proper routing of the bacteriophage X receptor protein to the outer membrane of Escherichia co/i K-12. J. Mol. Biol. 756, 93-l 12.
Ide, G. J., and Saunders, C. A. (1981). Rapid isolation of yeast nuclei. Cun. Genet. 4, 85-90.
Jensen, R., Sprague, G. F., Jr., and Herskowitz, I. (1983). Regulation of yeast mating-type interconversion: feedback control of HO gene expression by the mating-type locus. Proc. Nat. Acad. Sci. USA 80, 3035-3039.
Kilmartin, J. V., Wright, B., and Milstein. C. (1982). Rat monoclonal antitu- bulin antibodies derived by using a new nonsecreting rat cell line. J. Cell Biol. 93, 576-582.
Klar, A. J. S., Strathern, J. N., Broach, J. R., and Hicks, J. B. (1981). Regulation of transcription in expressed and unexpressed mating type cassettes of yeast. Nature 289, 239-244.
Kreil, G. (1981). Transfer of proteins across membranes. Ann. Rev. Bio- them. 50, 317-348.
Merriam, R. W.. and Hill, R. J. (1976). The germinal vesicle nucleus of Xenopos laevis oocytes as a selective storage receptacle for proteins. J. Cell Biol. 69, 659-668.
Miller, J. H. (1972). Experiments in Molecular Genetics. (Cold Spring Harbor, New York: Cold Spring Harbor Laboratcry).
Nagy, M., Laport, J., Penverne, 8.. and Herve, G. (1982). Nuclear locatiza- tion of aspartate transcarbamoylase in Saccharomyces cerevisiae. J. Cell Biol. 92, 790-794.
Nasmyth, K. A., Tatchell, K., Hall, B. D., Astell, C., and Smith, M. (1981). Physical analysis of mating-type loci in Sacchsromyces cerevisise. Cold Spring Harbor Symp. C&ant. Biol. 45, 961-981.
Paine, P. L., Moore, L. C., and Horowitz, S. B. (1975). Nuclear envelope permeabikty. Nature 254, 109-l 14.
Rechsteiner, M., and Kuehl. L. (1979). Microinjection of the nonhistone chromosomal protein HMGI into bovine fibroblasts and HeLa cells. Cell 16.901-908.

Nuclear Protein Localization 1065
Robinow, C. F., and Marak. J. (1966). A fiber apparatus in the nucleus of the yeast cell. J. Cell Biol. 29, 129-151.
Rose, M., Casadaban, M. J., and Botstein, D. (1981). Yeast genes fused to j%galactosidase in Escherichia co/i can be expressed normally in yeast. Proc. Nat. Acad. SCI. USA 78, 2460-2464.
Rothstein, R. J. (1983). One-step gene disruption in yeast. Meth. Enzymol. 701,202-211.
Sanger, F., Nrcklen, S., and Coulson, A. R. (1977). DNA sequencing with charn-terminating inhibitors. Proc. Nat. Acad. Sci. USA 74, 54635467.
Sherman, F., Fink, G. Ft., and Lawrence, C. W. (1979). Methods in Yeast Genetics. (Cold Spring Harbor, New York Cold Spring Harbor Laboratory).
Smith, M. M., and Andresson, 0. S. (1983). The DNA sequences of yeast H3 and H4 histone genes from two non-allelic gene sets encode identical H3 and H4 proterns. J. Mol. Biol. 769, 663-690.
Sprague, G. F., Jr., and Herskowitz, I. (1981). Control of yeast cell type by the mating type locus. I. Identification and control of expression of the a- specific gene BARl. J. Mol. Biol. 153. 305-321.
Strathern. J., Hicks, J., and Herskowitz, I. (1981). Control of cell type in yeast by the mating type locus: the ~~l-n2 hypothesis, J. Mol. Biol. 147, 357-372.
Struhl, K., Stinchcomb. D. T., Scherer, S., and Davis, R. W. (1979). High- frequency transformation of yeast: autonomous replication of hybrid DNA molecules. Proc. Nat. Acad. Sci. USA 76, 1035-1039.
Tatchell, K., Nasmyth, K. A., Hall, B. D.. Astell, C., and Smith, M. (1981). In vitro mutahon analysis of the mating-type locus in yeast. Cell 27, 25-35.
Unwin, P. N. T., and Milligan, R. A. (1982). A large particle associated with the perimeter of the nuclear pore complex. J. Cell. Biol. 93, 63-75.
Walks, J. W., Hereford, L., and Grunstein, M. (1980). Histone H2B genes of yeast encode two different proteins. Cell 22, 799-805.
Yamaizumi. M., Uchida, T., Okada, Y., Furusawa, M., and Mitsui, H. (1978). Rapid transfer of non-histone chromosomal proteins to the nucleus of living cells. Nature 273, 782-784.


















