
Specific roles of the αVβ1, αVβ3 and αVβ5 integrins in...
16
INTRODUCTION During early embryonic development, certain groups of cells, like neural crest cells, can transiently express locomotory prop- erties that allow them to migrate long distances from their sites of origin and populate other areas of the embryo where they undergo differentiation (Le Douarin, 1982; Newgreen and Erickson, 1986; Levi et al., 1990; Erickson and Perris, 1993). During migration to their final destination, neural crest cells penetrate extracellular matrices that are known to contain fibronectin, collagens, laminin, tenascin and a variety of pro- teoglycans (Thiery et al., 1982; Krotoski et al., 1986; Duband and Thiery, 1987; Tan et al., 1987; Mackie et al., 1988; Perris et al., 1991a,b). The role of these matrix components in migration has been analyzed in detail in the avian embryo essentially in in vitro approaches. Neural crest cells cultured in vitro adhere to and migrate efficiently on fibronectin, laminin, and type I, IV and VI collagens (Newgreen et al., 1982; Rovasio et al., 1983; Tucker and Erickson, 1984; Perris et al., 1989, 1991a, 1993a). In addition, antibodies to fibronectin or to the integrin β1 subunit and RGD peptides can impair neural crest cell migration on fibronectin substrata (Rovasio et al., 1983; Boucaut et al., 1984; Bronner-Fraser, 1985; Duband et al., 1986). Likewise, antibodies to the β1 or to the α1 subunit of integrins can affect neural crest cell adhesion to laminin or collagens (Lallier and Bronner-Fraser, 1992; Perris et al., 1993b). These studies thus provide strong evidence that avian neural crest cells can adhere and migrate in vitro on a variety of extracellular matrix molecules through β1 integrins. In vivo, injection of RGD-containing peptides or antibodies to fibronectin, to a laminin-proteoglycan complex or to the integrin β1 subunit into the cranial region of avian embryos 2687 Development 120, 2687-2702 (1994) Printed in Great Britain © The Company of Biologists Limited 1994 To identify potentially important extracellular matrix adhesive molecules in neural crest cell migration, the possible role of vitronectin and its corresponding integrin receptors was examined in the adhesion and migration of avian neural crest cells in vitro. Adhesion and migration on vitronectin were comparable to those found on fibronectin and could be almost entirely abolished by antibodies against vitronectin and by RGD peptides. Immunoprecip- itation and immunocytochemistry analyses revealed that neural crest cells expressed primarily the αVβ1, αVβ3 and αVβ5 integrins as possible vitronectin receptors. Inhibition assays of cellular adhesion and migration with function- perturbing antibodies demonstrated that adhesion of neural crest cells to vitronectin was mediated essentially by one or more of the different αV integrins, with a possible preeminence of αVβ1, whereas cell migration involved mostly the αVβ3 and αVβ5 integrins. Immunofluorescence labeling of cultured motile neural crest cells revealed that the αV integrins are differentially distributed on the cell surface. The β1 and αV subunits were both diffuse on the surface of cells and in focal adhesion sites in association with vinculin, talin and α-actinin, whereas the αVβ3 and αVβ5 integrins were essentially diffuse on the cell surface. Finally, vitronectin could be detected by immunoblotting and immunohistochemistry in the early embryo during the ontogeny of the neural crest. It was in particular closely associated with the surface of migrating neural crest cells. In conclusion, our study indicates that neural crest cells can adhere to and migrate on vitronectin in vitro by an RGD- dependent mechanism involving at least the αVβ1, αVβ3 and αVβ5 integrins and that these integrins may have specific roles in the control of cell adhesion and migration. Key words: neural crest, vitronectin, integrins, quail, cell adhesion, cell migration SUMMARY Specific roles of the αVβ1, αVβ3 and αVβ5 integrins in avian neural crest cell adhesion and migration on vitronectin Muriel Delannet 1 , Fabrice Martin 1 , Blaise Bossy 2,† , David A. Cheresh 3 , Louis F. Reichardt 2 and Jean-Loup Duband 1, * 1 Laboratoire de Biologie Cellulaire du Développement, Institut Jacques Monod, Université Paris 7, 2, place Jussieu, 75251 Paris Cedex 05, France 2 University of California and Howard Hughes Medical Institute, Parnassus and Third Avenues, San Francisco, California 94143- 0724, USA 3 Department of Immunology, The Scripps Research Institute, La Jolla, California 92037, USA *Author for correspondence at Institut Jacques Monod † Present address: Institut d’Embryologie Cellulaire et Moléculaire du CNRS et du Collège de France, 49bis, avenue de la Belle Gabrielle, 94736 Nogent-sur-Marne Cedex, France
Transcript of Specific roles of the αVβ1, αVβ3 and αVβ5 integrins in...

INTRODUCTION
During early embryonic development, certain groups of cells,like neural crest cells, can transiently express locomotory prop-erties that allow them to migrate long distances from their sitesof origin and populate other areas of the embryo where theyundergo differentiation (Le Douarin, 1982; Newgreen andErickson, 1986; Levi et al., 1990; Erickson and Perris, 1993).During migration to their final destination, neural crest cellspenetrate extracellular matrices that are known to containfibronectin, collagens, laminin, tenascin and a variety of pro-teoglycans (Thiery et al., 1982; Krotoski et al., 1986; Dubandand Thiery, 1987; Tan et al., 1987; Mackie et al., 1988; Perriset al., 1991a,b). The role of these matrix components inmigration has been analyzed in detail in the avian embryoessentially in in vitro approaches. Neural crest cells cultured in
vitro adhere to and migrate efficiently on fibronectin, laminin,and type I, IV and VI collagens (Newgreen et al., 1982;Rovasio et al., 1983; Tucker and Erickson, 1984; Perris et al.,1989, 1991a, 1993a). In addition, antibodies to fibronectin orto the integrin
β1 subunit and RGD peptides can impair neuralcrest cell migration on fibronectin substrata (Rovasio et al.,1983; Boucaut et al., 1984; Bronner-Fraser, 1985; Duband etal., 1986). Likewise, antibodies to the β1 or to the α1 subunitof integrins can affect neural crest cell adhesion to laminin orcollagens (Lallier and Bronner-Fraser, 1992; Perris et al.,1993b). These studies thus provide strong evidence that avianneural crest cells can adhere and migrate in vitro on a varietyof extracellular matrix molecules through β1 integrins.
In vivo, injection of RGD-containing peptides or antibodiesto fibronectin, to a laminin-proteoglycan complex or to theintegrin β1 subunit into the cranial region of avian embryos
2687Development 120, 2687-2702 (1994)Printed in Great Britain © The Company of Biologists Limited 1994
To identify potentially important extracellular matrixadhesive molecules in neural crest cell migration, thepossible role of vitronectin and its corresponding integrinreceptors was examined in the adhesion and migration ofavian neural crest cells in vitro. Adhesion and migration onvitronectin were comparable to those found on fibronectinand could be almost entirely abolished by antibodiesagainst vitronectin and by RGD peptides. Immunoprecip-itation and immunocytochemistry analyses revealed thatneural crest cells expressed primarily the αVβ1, αVβ3 andαVβ5 integrins as possible vitronectin receptors. Inhibitionassays of cellular adhesion and migration with function-perturbing antibodies demonstrated that adhesion ofneural crest cells to vitronectin was mediated essentially byone or more of the different αV integrins, with a possiblepreeminence of αVβ1, whereas cell migration involvedmostly the αVβ3 and αVβ5 integrins. Immunofluorescencelabeling of cultured motile neural crest cells revealed that
the αV integrins are differentially distributed on the cellsurface. The β1 and αV subunits were both diffuse on thesurface of cells and in focal adhesion sites in associationwith vinculin, talin and α-actinin, whereas the αVβ3 andαVβ5 integrins were essentially diffuse on the cell surface.Finally, vitronectin could be detected by immunoblottingand immunohistochemistry in the early embryo during theontogeny of the neural crest. It was in particular closelyassociated with the surface of migrating neural crest cells.In conclusion, our study indicates that neural crest cells canadhere to and migrate on vitronectin in vitro by an RGD-dependent mechanism involving at least the αVβ1, αVβ3and αVβ5 integrins and that these integrins may havespecific roles in the control of cell adhesion and migration.
Key words: neural crest, vitronectin, integrins, quail, cell adhesion,cell migration
SUMMARY
Specific roles of the αVβ1, αVβ3 and αVβ5 integrins in avian neural crest cell
adhesion and migration on vitronectin
Muriel Delannet1, Fabrice Martin1, Blaise Bossy2,†, David A. Cheresh3, Louis F. Reichardt2and Jean-Loup Duband1,*1Laboratoire de Biologie Cellulaire du Développement, Institut Jacques Monod, Université Paris 7, 2, place Jussieu, 75251 ParisCedex 05, France2University of California and Howard Hughes Medical Institute, Parnassus and Third Avenues, San Francisco, California 94143-0724, USA3Department of Immunology, The Scripps Research Institute, La Jolla, California 92037, USA
*Author for correspondence at Institut Jacques Monod†Present address: Institut d’Embryologie Cellulaire et Moléculaire du CNRS et du Collège de France, 49bis, avenue de la Belle Gabrielle, 94736 Nogent-sur-MarneCedex, France

2688
cause severe deficencies in neural crest cell migration (Boucautet al., 1984; Bronner-Fraser, 1985; Poole and Thiery, 1986;Bronner-Fraser and Lallier, 1991). However, the same anti-bodies fail to perturb neural crest cell migration in trunkregions although they are able to inhibit strongly myoblastmigration (Jaffredo et al., 1988; Bronner-Fraser, 1993). Thisindicates that, while cranial neural crest cells are likely tomigrate in vivo primarily on fibronectin and laminin using β1integrins, truncal neural crest cells may be able to interact withadditional extracellular matrix molecules for migration usingnon-β1 integrins, allowing them to overcome the inhibitoryeffect of the antibodies. Consistent with this, it has been shownrecently that cranial and trunk neural crest cells may differ intheir mechanisms of adhesion to selected extracellular matrixcomponents in vitro (Lallier et al., 1992).
Therefore, additional extracellular matrix components thatpromote truncal neural crest cell locomotion have to be deter-mined. A possible candidate is vitronectin, a multifunctionaladhesive glycoprotein of
Mr of about 70×103 (70K) found inthe circulation and in the extracellular matrix of various tissuesand which interacts with the surface of cells primarily throughthe αVβ3 integrin, also called vitronectin receptor (forreviews, see Preissner, 1991; Felding-Habermann andCheresh, 1993). Owing to its multidomain structure withbinding sites for various integrins, heparin, collagen, plas-minogen and plasminogen activator inhibitor 1, vitronectinplays a critical role in the substratum adhesion of a largevariety of cell types, in hemostasis and in immune defense.Surprisingly, although considerable information has accumu-lated regarding its structural and adhesive properties, theinvolvement of vitronectin as a possible regulatory extracellu-lar matrix molecule during development has been poorly inves-tigated. In the present report, we examine in vitro the adhesiveand migratory response of avian trunk neural crest cells to vit-ronectin. We also describe the distribution of vitronectin inneural crest cell migratory pathways and characterize the reper-toire and functions of the vitronectin-binding integrins in thesecells.
MATERIALS AND METHODS
Adhesive proteins and antibodiesVitronectin was purified from bovine serum by affinity chromatogra-phy on a heparin-Sepharose column as described by Yatohgo et al.(1988). Bovine plasma fibronectin was purified on a gelatin-Sepharose column as described previously (Rovasio et al., 1983).Rabbit polyclonal antibodies to chicken and bovine vitronectin werekindly provided by Dr M. Hayashi (Ochanomizu University, Tokyo,Japan) or have been described elsewhere (Neugebauer et al., 1991).A monoclonal antibody (mAb) to human vitronectin (clone VIT-2),rabbit antibodies to chicken α-actinin and RGDS peptides werepurchased from Sigma. GRDGS peptides that are known to lackinhibitory activity on vitronectin or on fibronectin were a kind gift ofDr K. M. Yamada (NIH, Bethesda, USA). Polyclonal antibodies tochicken fibronectin have been described previously (Duband andThiery, 1982). Polyclonal antibodies (2992) and the rat mAb ES46-8both directed against the chicken β1-integrin subunit were kindlyprovided by Dr K. M. Yamada. The CSAT hybridoma (anti-chickenβ1 integrin subunit) was kindly donated by Dr C. Buck (The WistarInstitute, Philadelphia, USA). The mAb Chav-1 (anti-chicken αVintegrin subunit) has been described previously (Neugebauer et al.,1991). Rabbit polyclonal antibodies (VNR-C3) raised against a
peptide corresponding to the cytoplasmic domain of the chick αVsubunit were described previously (Bossy and Reichardt, 1990).Rabbit polyclonal antibodies (344 and 4377) raised against peptidescorresponding to the cytoplasmic domains of the human β3 and β5subunits and cross-reacting with the avian β3 and β5 chains were akind gift of Dr K. Venstrom (University of California, San Francisco,USA). The mAbs LM609 and P3G2 to the human αVβ3 and αVβ5integrins, respectively, were described elsewhere (Cheresh and Spiro,1987; Wayner et al., 1991). Polyclonal antibodies to chicken vinculinwere a kind gift of Dr B. Geiger (The Weizmann Institute, Rehovot,Israel). Polyclonal antibodies to chicken talin were a gift of Dr S. Saga(Nagoya University School of Medicine, Nagoya, Japan). A mousemonoclonal antibody, called NC-1, identical to HNK-1 and specifi-cally recognizing neural crest cells has been described elsewhere(Tucker et al., 1984).
Embryos and cell culturesJapanese quail (Coturnix coturnix japonica) embryos were usedthroughout the study. Eggs were incubated at 38±1°C and stagedaccording to the number of somite pairs and to the duration of incu-bation. Trunk neural crest cell cultures were generated as describedpreviously (Duband et al., 1988a). Cells were cultured at 37°C in ahumidified 5% CO2 atmosphere in DMEM supplemented with 5%calf serum depleted in fibronectin. In a number of experiments though,cells were cultured in DMEM supplemented with 0.1% ovalbumin,transferrin and insulin, each at 10 µg/ml.
Assays for cellular adhesionCellular adhesion assays were performed in 10 cm bacteriologicalPetri dishes. The use of Petri dishes designed for cell culture wasavoided because they gave high non-specific cell adhesion. Smallareas of the dishes were incubated at 37°C for 1 hour with 50 µl ofvitronectin or fibronectin at various concentrations from 0.05 to 100µg/ml in calcium- and magnesium-free phosphate-buffered saline(PBS), followed by incubation with bovine serum albumin (BSA) inPBS at 10 mg/ml for 30 minutes and extensive washes in PBS. Neuralcrest cells were obtained from at least 50 neural tube explants culturedon fibronectin or vitronectin for 18 hours. Cells were harvested usingtreatment for 3 minutes at 37°C with an enzyme-free cell dissociationbuffer (Gibco-BRL). After addition of prewarmed DMEM, cells werecollected, sedimented at 1,000 revs/minute for 10 minutes and resus-pended in serum-free DMEM. Cells were counted and a 50 µl aliquotof cell suspension containing approximately 5×103 cells wasdeposited on the areas precoated with the proteins to be tested. Thedishes were then incubated at 37°C for 1 hour, rinsed in PBS toremove the non-adherent cells and fixed in a 3.7% formaldehydesolution in PBS. Attached and spread cells were counted under aNikon inverted phase contrast microscope.
Assays for cellular migrationCellular migration assays were performed either in 10 cm bacterio-logical Petri dishes or in Terasaki plates previously coated withadhesion-promoting proteins as for assays for cellular adhesion.Cultures were incubated in DMEM supplemented with either 5% calfserum or 0.1% ovalbumin and observed periodically with a Nikonphase contrast inverted microscope. The extent of migration of neuralcrest cells was estimated by measuring the linear distance between theneural tube explant and the front of migration of neural crest cells.This parameter was found to reflect accurately the degree of motilityof individual neural crest cells (Duband et al., 1991). For time-lapsevideomicroscopy, cultures were performed in Terasaki plates asdescribed previously (Dufour et al., 1988) and were observed with aNikon Diaphot inverted microscope in a heated plexiglass chamberequipped with a video camera (Hamamatsu, Japan) connected to a TVmonitor (Hitachi, Japan) and a time-lapse recorder (Mitsubishi,Japan). The total distance migrated by the selected cells was measuredand the speed of locomotion calculated. The degree of persistence of
M. Delannet and others

2689Avian crest cell migration on vitronectin
movement was defined as the ratio between the linear distance andthe total distance covered by the cells.
Histological sections and immunofluorescent stainingsEmbryos were routinely fixed at 4°C in 3.7% formaldehyde in PBSfor at least 12 hours, depending on the size of the embryos. Afterseveral extensive washes in PBS, embryos were embedded in asucrose solution in PBS (15% wt/vol) and subsequently frozen inCryo-M-Bed compound (Bright Instrument, Huntington, England) inliquid nitrogen. Sections were cut at 10-12 µm on a cryostat (BrightInstrument) and mounted on Superfrost/Plus slides (CML, Nemours,France). For immunofluorescent labeling of cell cultures, neural tubesor embryonic fibroblasts were explanted onto vitronectin-coated glasscoverslips in Petri dishes and cultured in DMEM supplemented with5% fibronectin-depleted serum. After washes in serum-free DMEM,cultures were fixed using different procedures: (i) in cold methanolfor 5 minutes, followed by cold acetone for 1 minute, (ii) in 3.7%formaldehyde in PBS for 1 hour at room temperature, (iii) in 3.7%formaldehyde-0.5% Triton X-100-5% sucrose in PBS for 5 minutesfollowed by a 1 hour incubation in 3.7% formaldehyde in PBS.Cultures were subsequently rinsed in PBS and, in some cases, theywere subjected to permeabilization with 0.5% Triton X-100 in PBSfor 3 minutes. Sections and cultures were subjected to immunofluo-rescent staining using biotinylated secondary antibodies and fluores-cein-conjugated streptavidin (Amersham). For control experiments,sections were treated with non-immune rabbit antibodies or unrelatedmAbs in place of the primary antibody, or with anti-vitronectin anti-bodies previously incubated with an excess of purified vitronectin.Preparations were observed with a Leica epifluorescence microscopeand photographed using TMX-400 Kodak film.
Immunoblottings and immunoprecipitationsFor immunoblotting, samples of embryonic tissues or cultured cellswere briefly homogenized and extracted at 90°C with SDS-samplebuffer under reducing or non-reducing conditions. The extracts wereclarified by centrifugation and subjected to SDS-PAGE followed byimmunoblotting analysis. PAGE was performed in Laemmli buffersystem on 7.5% polyacrylamide slab minigels. The protein bandswere electroblotted for 1 hour onto nitrocellulose in 50 mM Tris-glycine, 20% methanol buffer. The nitrocellulose membranes werethen incubated with a 4% bovine serum albumin solution in PBS for1 hour at ambient temperature, followed by incubation with antibodysolution for 12 hours at 4°C. The sheets were rinsed in PBS supple-mented with 1% Tween 20 and incubated first with biotinylatedsecondary antibodies for 2 hours at room temperature, then with 125I-labelled streptavidin (Amersham) for 1 hour at room temperature.After rinsing, the blot was subjected to autoradiography.
For immunoprecipitation experiments, cells were metabolicallylabeled with [35S]methionine for at least 8 hours, extracted in 2%Triton X-100 and processed for immunoprecipitation using protein-A-Sepharose as described previously (Duband et al., 1988b). Sampleswere subjected to SDS-PAGE on 7.5%-acrylamide gels under non-reducing conditions for detection of integrin complexes and on 10%-acrylamide gels under reducing conditions for detection of vitronectinfollowed by fluorography.
RESULTS
Distribution of vitronectin at the time of neural crestcell migrationWe have at first investigated whether vitronectin is expressedin the embryo at the time of neural crest cell migration and,more specifically, if it is associated with migrating neural crestcells, using immunoblotting and immunohistochemistry. Forthis purpose, we used three different antibodies directed
against vitronectin: two polyclonal antibodies against chickenvitronectin and characterized previously (Kitagaki-Ogawa etal., 1990; Neugebauer et al., 1991; Nagano et al., 1992) andthe mAb VIT-2 to human vitronectin from Sigma. All threeantibodies specifically recognized chicken serum vitronectin inimmunoblots (data not shown) and gave identical results inimmunohistochemical experiments. Fig. 1 shows that, inimmunoblotting experiments, the antibodies recognizedprimarily a band of about Mr of 68K and, occasionally, a minorband of 78K in extracts of embryos at day 2.5 of development(E2.5) as well as in E3.5 and E4.5 embryos. These bands cor-responded to the molecular masses expected for the two well-characterized forms of chicken vitronectin (Kitagaki-Ogawa etal., 1990). Another band of Mr of about 43K could also bedetected in all extracts of embryonic tissues and correspondedeither to one of the recently characterized forms of yolk vit-ronectin (Nagano et al., 1992) or to a degradation product ofvitronectin, this molecule being very susceptible to proteolyticcleavage.
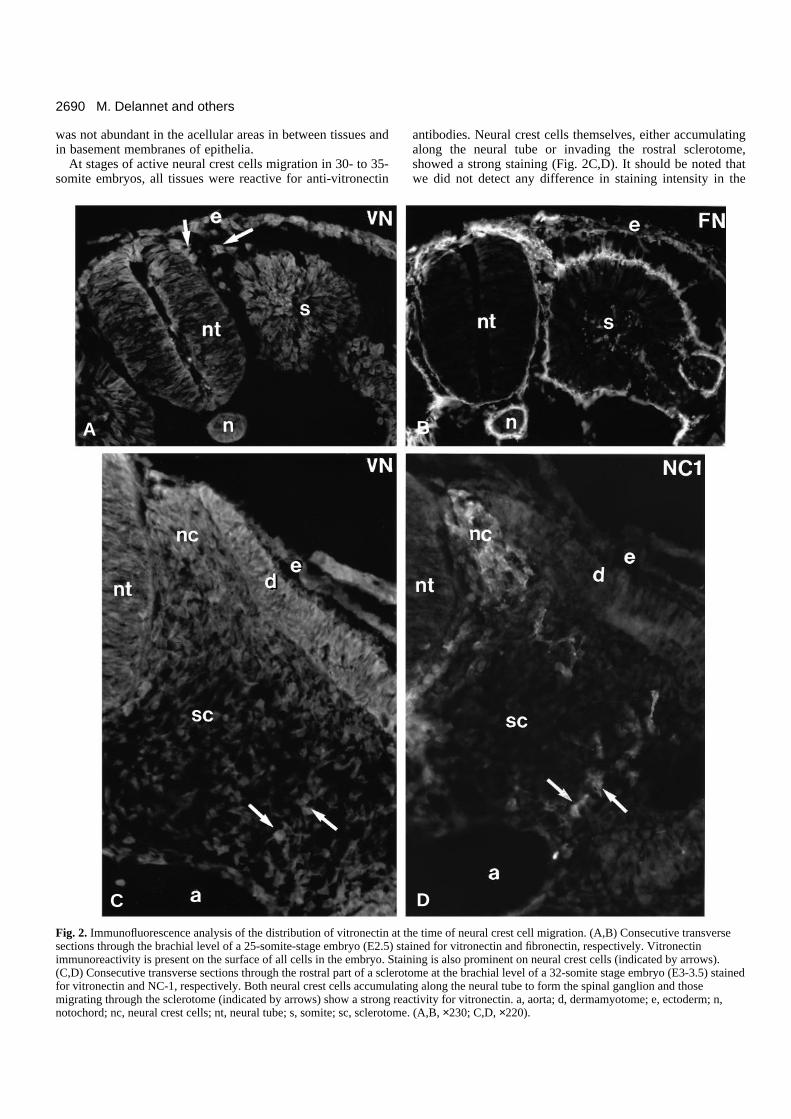
The distribution of vitronectin was analyzed in detail in thetrunk region of embryos at the time of neural crest cellmigration and differentiation, using immunofluorescencelabeling of frozen sections of formaldehyde-fixed embryos. Atthe onset of neural crest cell migration in 25-somite embryos,vitronectin immunoreactivity was found in all tissues of theembryo, including the somites, the lateral mesoderm, theneural tube, the notochord, the ectoderm and the endoderm(Fig. 2A). Early migrating neural crest cells showed an intensestaining for vitronectin on their surface. Interestingly, vit-ronectin exhibited a very distinct pattern of distribution fromfibronectin (Fig. 2B). In particular, it was intimately associatedwith the surface of both epithelial and mesenchymal cells and
Fig. 1. Immunoblotting analysis of vitronectin expression in embryosat the time of neural crest cell development. Extracts of truncal andcranial regions of embryos at E2.5 (lanes 1 and 2, respectively) and oftruncal regions of embryos at E3.5 and E4.5 (lanes 3 and 4,respectively) were subjected to immunoblotting using a monoclonalantibody to human vitronectin. Approximately the same amounts ofmaterial were loaded in the different lanes. Two major bands of aboutMr of 68K and 43K are found at very similar amounts in all embryoextracts. The upper band corresponds to vitronectin and the lowerband corresponds either to a degradation product of vitronectin or toone of the forms of yolk vitronectin (Nagano et al., 1992). The bandof Mr of 78K that can be seen in lane 4 is likely can be attributed tothe high molecular mass form of vitronectin (Kitagaki-Ogawa et al.,1990). Molecular mass standards are indicated on the left.
2690
was not abundant in the acellular areas in between tissues andin basement membranes of epithelia.
At stages of active neural crest cells migration in 30- to 35-somite embryos, all tissues were reactive for anti-vitronectin
antibodies. Neural crest cells themselves, either accumulatingalong the neural tube or invading the rostral sclerotome,showed a strong staining (Fig. 2C,D). It should be noted thatwe did not detect any difference in staining intensity in the
M. Delannet and others
Fig. 2. Immunofluorescence analysis of the distribution of vitronectin at the time of neural crest cell migration. (A,B) Consecutive transversesections through the brachial level of a 25-somite-stage embryo (E2.5) stained for vitronectin and fibronectin, respectively. Vitronectinimmunoreactivity is present on the surface of all cells in the embryo. Staining is also prominent on neural crest cells (indicated by arrows).(C,D) Consecutive transverse sections through the rostral part of a sclerotome at the brachial level of a 32-somite stage embryo (E3-3.5) stainedfor vitronectin and NC-1, respectively. Both neural crest cells accumulating along the neural tube to form the spinal ganglion and thosemigrating through the sclerotome (indicated by arrows) show a strong reactivity for vitronectin. a, aorta; d, dermamyotome; e, ectoderm; n,notochord; nc, neural crest cells; nt, neural tube; s, somite; sc, sclerotome. (A,B, ×230; C,D, ×220).
A B
C D
2691Avian crest cell migration on vitronectin
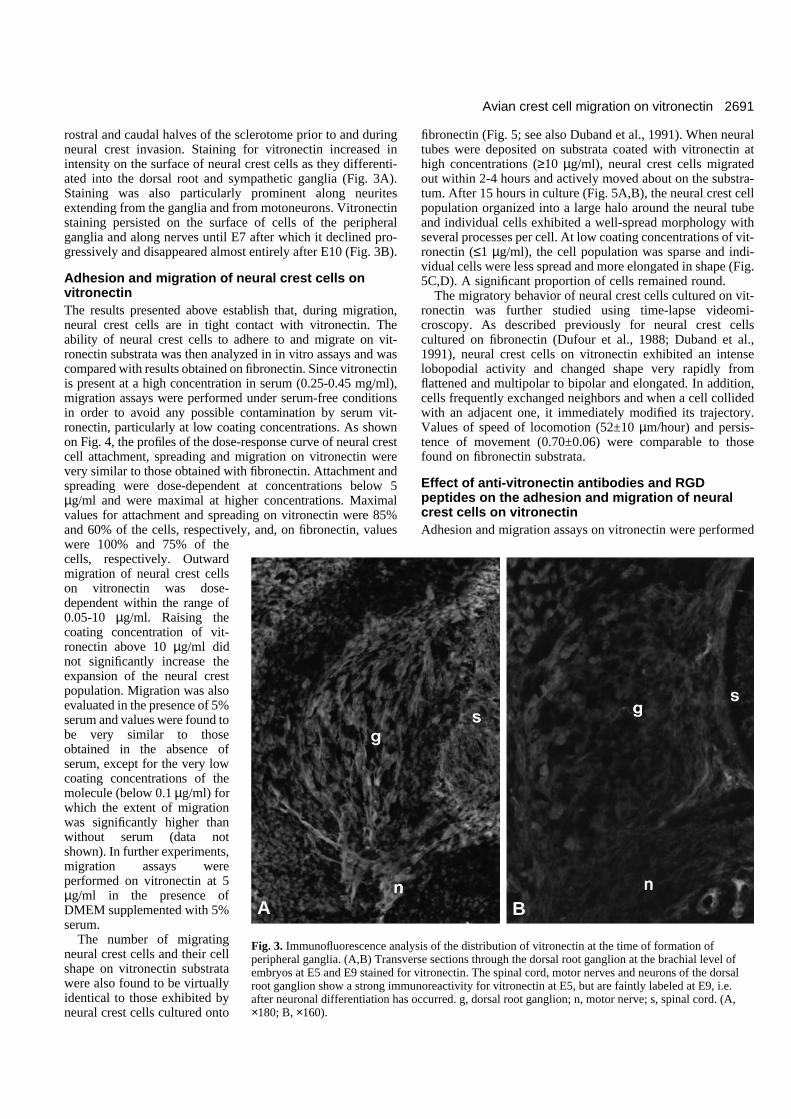
rostral and caudal halves of the sclerotome prior to and duringneural crest invasion. Staining for vitronectin increased inintensity on the surface of neural crest cells as they differenti-ated into the dorsal root and sympathetic ganglia (Fig. 3A).Staining was also particularly prominent along neuritesextending from the ganglia and from motoneurons. Vitronectinstaining persisted on the surface of cells of the peripheralganglia and along nerves until E7 after which it declined pro-gressively and disappeared almost entirely after E10 (Fig. 3B).
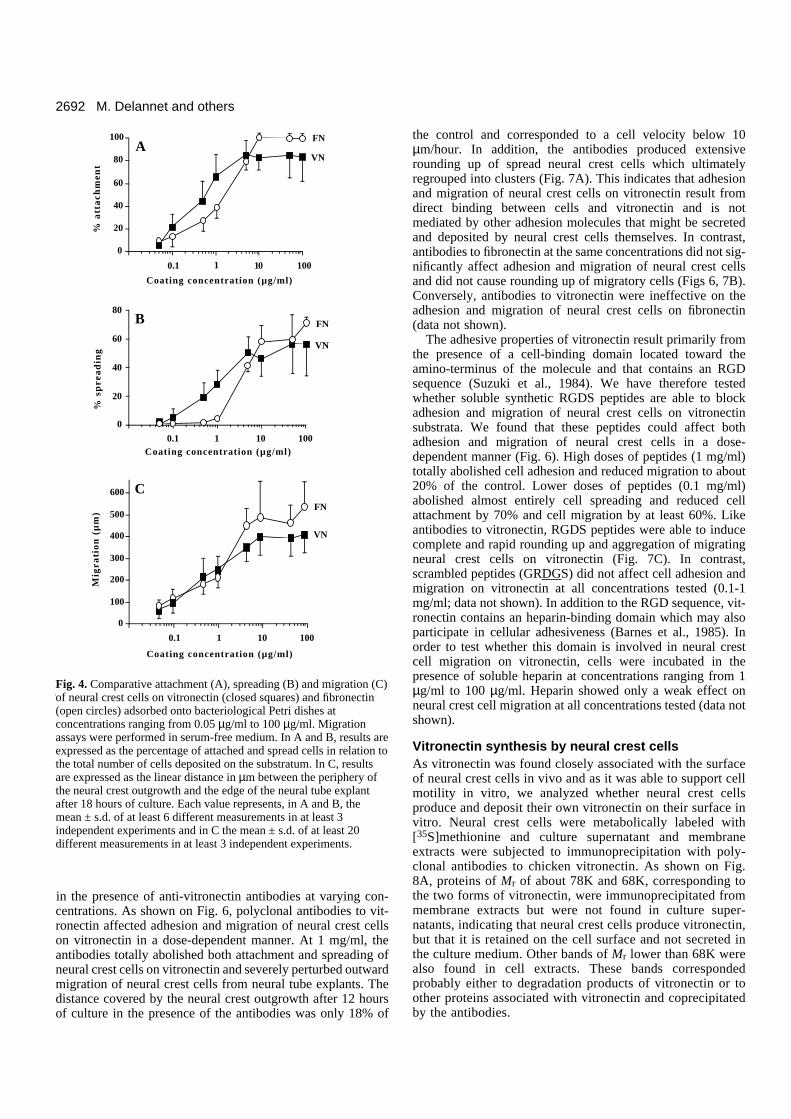
Adhesion and migration of neural crest cells onvitronectinThe results presented above establish that, during migration,neural crest cells are in tight contact with vitronectin. Theability of neural crest cells to adhere to and migrate on vit-ronectin substrata was then analyzed in in vitro assays and wascompared with results obtained on fibronectin. Since vitronectinis present at a high concentration in serum (0.25-0.45 mg/ml),migration assays were performed under serum-free conditionsin order to avoid any possible contamination by serum vit-ronectin, particularly at low coating concentrations. As shownon Fig. 4, the profiles of the dose-response curve of neural crestcell attachment, spreading and migration on vitronectin werevery similar to those obtained with fibronectin. Attachment andspreading were dose-dependent at concentrations below 5µg/ml and were maximal at higher concentrations. Maximalvalues for attachment and spreading on vitronectin were 85%and 60% of the cells, respectively, and, on fibronectin, valueswere 100% and 75% of thecells, respectively. Outwardmigration of neural crest cellson vitronectin was dose-dependent within the range of0.05-10 µg/ml. Raising thecoating concentration of vit-ronectin above 10 µg/ml didnot significantly increase theexpansion of the neural crestpopulation. Migration was alsoevaluated in the presence of 5%serum and values were found tobe very similar to thoseobtained in the absence ofserum, except for the very lowcoating concentrations of themolecule (below 0.1 µg/ml) forwhich the extent of migrationwas significantly higher thanwithout serum (data notshown). In further experiments,migration assays wereperformed on vitronectin at 5µg/ml in the presence ofDMEM supplemented with 5%serum.
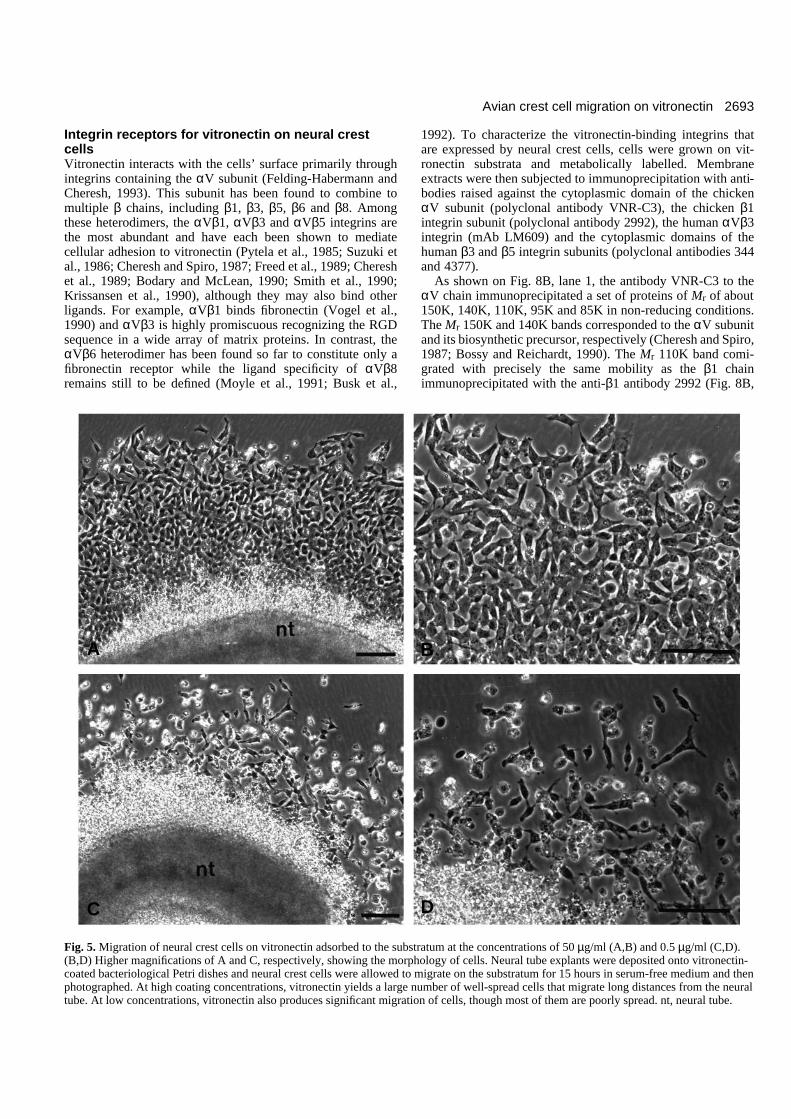
The number of migratingneural crest cells and their cellshape on vitronectin substratawere also found to be virtuallyidentical to those exhibited byneural crest cells cultured onto
fibronectin (Fig. 5; see also Duband et al., 1991). When neuraltubes were deposited on substrata coated with vitronectin athigh concentrations (≥10 µg/ml), neural crest cells migratedout within 2-4 hours and actively moved about on the substra-tum. After 15 hours in culture (Fig. 5A,B), the neural crest cellpopulation organized into a large halo around the neural tubeand individual cells exhibited a well-spread morphology withseveral processes per cell. At low coating concentrations of vit-ronectin (≤1 µg/ml), the cell population was sparse and indi-vidual cells were less spread and more elongated in shape (Fig.5C,D). A significant proportion of cells remained round.
The migratory behavior of neural crest cells cultured on vit-ronectin was further studied using time-lapse videomi-croscopy. As described previously for neural crest cellscultured on fibronectin (Dufour et al., 1988; Duband et al.,1991), neural crest cells on vitronectin exhibited an intenselobopodial activity and changed shape very rapidly fromflattened and multipolar to bipolar and elongated. In addition,cells frequently exchanged neighbors and when a cell collidedwith an adjacent one, it immediately modified its trajectory.Values of speed of locomotion (52±10 µm/hour) and persis-tence of movement (0.70±0.06) were comparable to thosefound on fibronectin substrata.
Effect of anti-vitronectin antibodies and RGDpeptides on the adhesion and migration of neuralcrest cells on vitronectinAdhesion and migration assays on vitronectin were performed
Fig. 3. Immunofluorescence analysis of the distribution of vitronectin at the time of formation ofperipheral ganglia. (A,B) Transverse sections through the dorsal root ganglion at the brachial level ofembryos at E5 and E9 stained for vitronectin. The spinal cord, motor nerves and neurons of the dorsalroot ganglion show a strong immunoreactivity for vitronectin at E5, but are faintly labeled at E9, i.e.after neuronal differentiation has occurred. g, dorsal root ganglion; n, motor nerve; s, spinal cord. (A,×180; B, ×160).
A B
2692
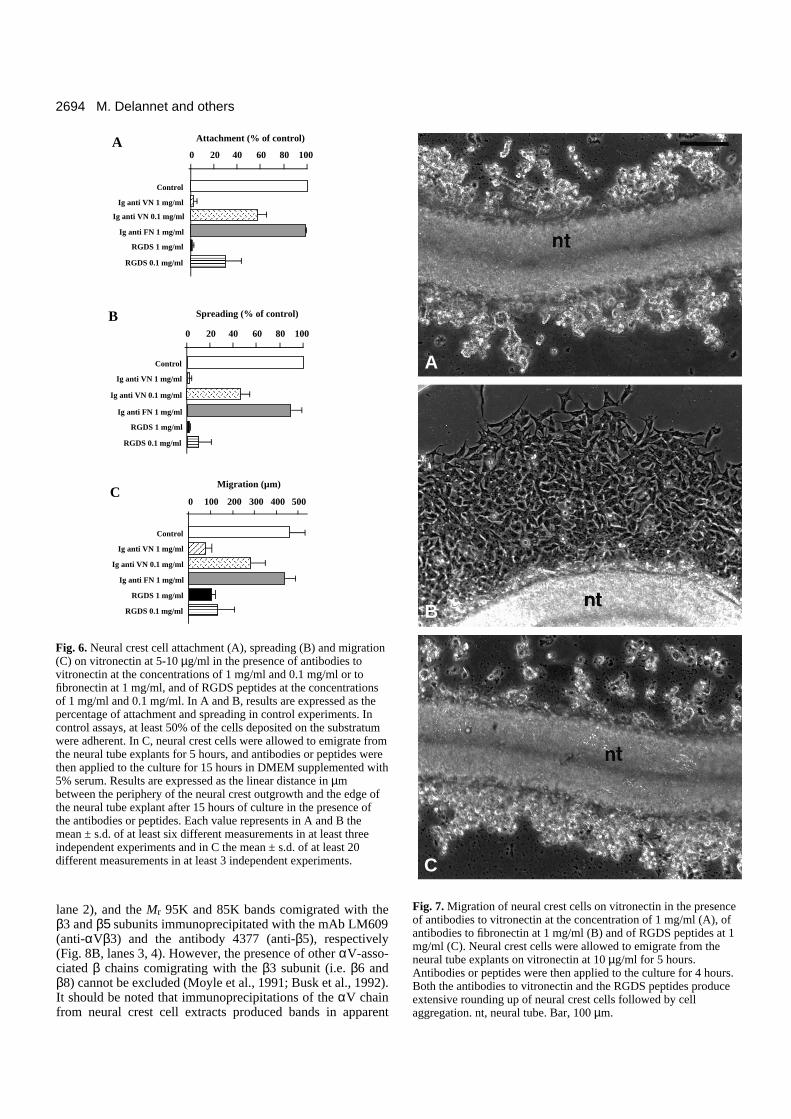
in the presence of anti-vitronectin antibodies at varying con-centrations. As shown on Fig. 6, polyclonal antibodies to vit-ronectin affected adhesion and migration of neural crest cellson vitronectin in a dose-dependent manner. At 1 mg/ml, theantibodies totally abolished both attachment and spreading ofneural crest cells on vitronectin and severely perturbed outwardmigration of neural crest cells from neural tube explants. Thedistance covered by the neural crest outgrowth after 12 hoursof culture in the presence of the antibodies was only 18% of
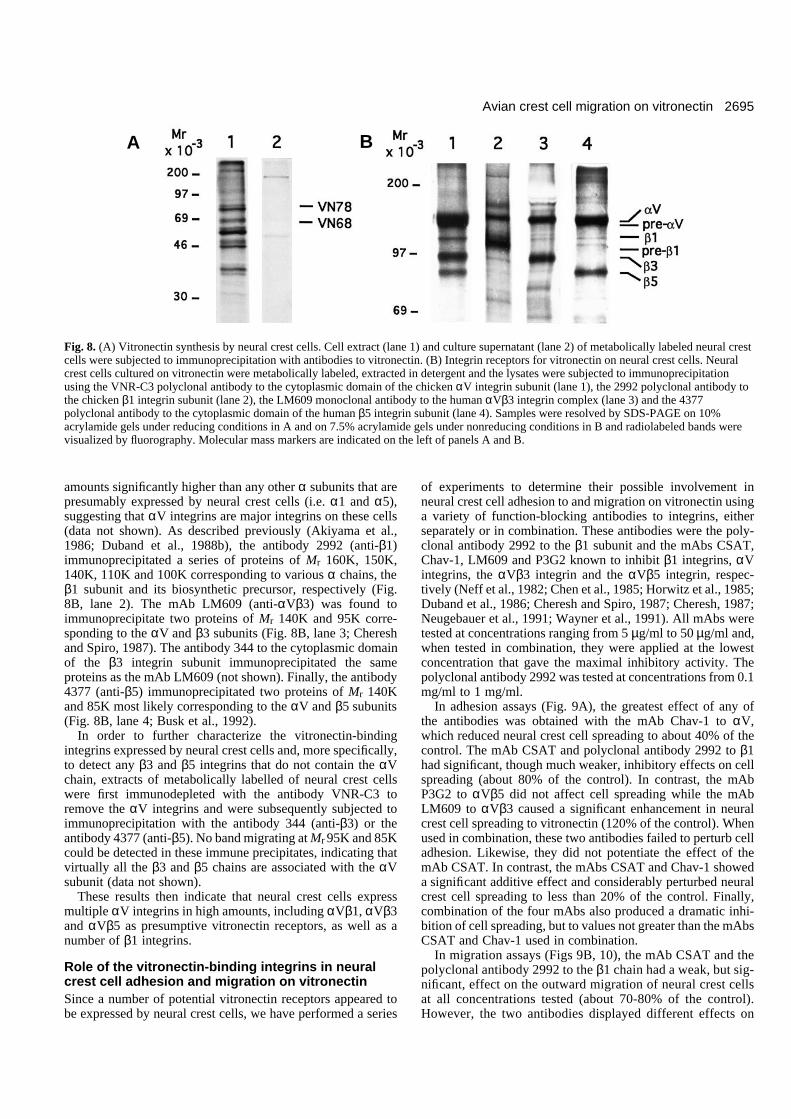
the control and corresponded to a cell velocity below 10µm/hour. In addition, the antibodies produced extensiverounding up of spread neural crest cells which ultimatelyregrouped into clusters (Fig. 7A). This indicates that adhesionand migration of neural crest cells on vitronectin result fromdirect binding between cells and vitronectin and is notmediated by other adhesion molecules that might be secretedand deposited by neural crest cells themselves. In contrast,antibodies to fibronectin at the same concentrations did not sig-nificantly affect adhesion and migration of neural crest cellsand did not cause rounding up of migratory cells (Figs 6, 7B).Conversely, antibodies to vitronectin were ineffective on theadhesion and migration of neural crest cells on fibronectin(data not shown).
The adhesive properties of vitronectin result primarily fromthe presence of a cell-binding domain located toward theamino-terminus of the molecule and that contains an RGDsequence (Suzuki et al., 1984). We have therefore testedwhether soluble synthetic RGDS peptides are able to blockadhesion and migration of neural crest cells on vitronectinsubstrata. We found that these peptides could affect bothadhesion and migration of neural crest cells in a dose-dependent manner (Fig. 6). High doses of peptides (1 mg/ml)totally abolished cell adhesion and reduced migration to about20% of the control. Lower doses of peptides (0.1 mg/ml)abolished almost entirely cell spreading and reduced cellattachment by 70% and cell migration by at least 60%. Likeantibodies to vitronectin, RGDS peptides were able to inducecomplete and rapid rounding up and aggregation of migratingneural crest cells on vitronectin (Fig. 7C). In contrast,scrambled peptides (GRDGS) did not affect cell adhesion andmigration on vitronectin at all concentrations tested (0.1-1mg/ml; data not shown). In addition to the RGD sequence, vit-ronectin contains an heparin-binding domain which may alsoparticipate in cellular adhesiveness (Barnes et al., 1985). Inorder to test whether this domain is involved in neural crestcell migration on vitronectin, cells were incubated in thepresence of soluble heparin at concentrations ranging from 1µg/ml to 100 µg/ml. Heparin showed only a weak effect onneural crest cell migration at all concentrations tested (data notshown).
Vitronectin synthesis by neural crest cellsAs vitronectin was found closely associated with the surfaceof neural crest cells in vivo and as it was able to support cellmotility in vitro, we analyzed whether neural crest cellsproduce and deposit their own vitronectin on their surface invitro. Neural crest cells were metabolically labeled with[35S]methionine and culture supernatant and membraneextracts were subjected to immunoprecipitation with poly-clonal antibodies to chicken vitronectin. As shown on Fig.8A, proteins of Mr of about 78K and 68K, corresponding tothe two forms of vitronectin, were immunoprecipitated frommembrane extracts but were not found in culture super-natants, indicating that neural crest cells produce vitronectin,but that it is retained on the cell surface and not secreted inthe culture medium. Other bands of Mr lower than 68K werealso found in cell extracts. These bands correspondedprobably either to degradation products of vitronectin or toother proteins associated with vitronectin and coprecipitatedby the antibodies.
M. Delannet and others
Figure 4: Delannet et al.
10
0
20
40
60
80
100
Coating concentration (µg/ml)
% a
tta
chm
ent
FN
VN
1 1000.1
A
0.1
0
20
40
60
80
Coating concentration (µg/ml)
% s
pre
ad
ing
FN
VN
1 10 100
B
C
0
100
200
300
400
500
600
Mig
rati
on
(µ
m)
FN
VN
0.1
Coating concentration (µg/ml)
1 10 100
Fig. 4. Comparative attachment (A), spreading (B) and migration (C)of neural crest cells on vitronectin (closed squares) and fibronectin(open circles) adsorbed onto bacteriological Petri dishes atconcentrations ranging from 0.05 µg/ml to 100 µg/ml. Migrationassays were performed in serum-free medium. In A and B, results areexpressed as the percentage of attached and spread cells in relation tothe total number of cells deposited on the substratum. In C, resultsare expressed as the linear distance in µm between the periphery ofthe neural crest outgrowth and the edge of the neural tube explantafter 18 hours of culture. Each value represents, in A and B, themean ± s.d. of at least 6 different measurements in at least 3independent experiments and in C the mean ± s.d. of at least 20different measurements in at least 3 independent experiments.
2693Avian crest cell migration on vitronectin
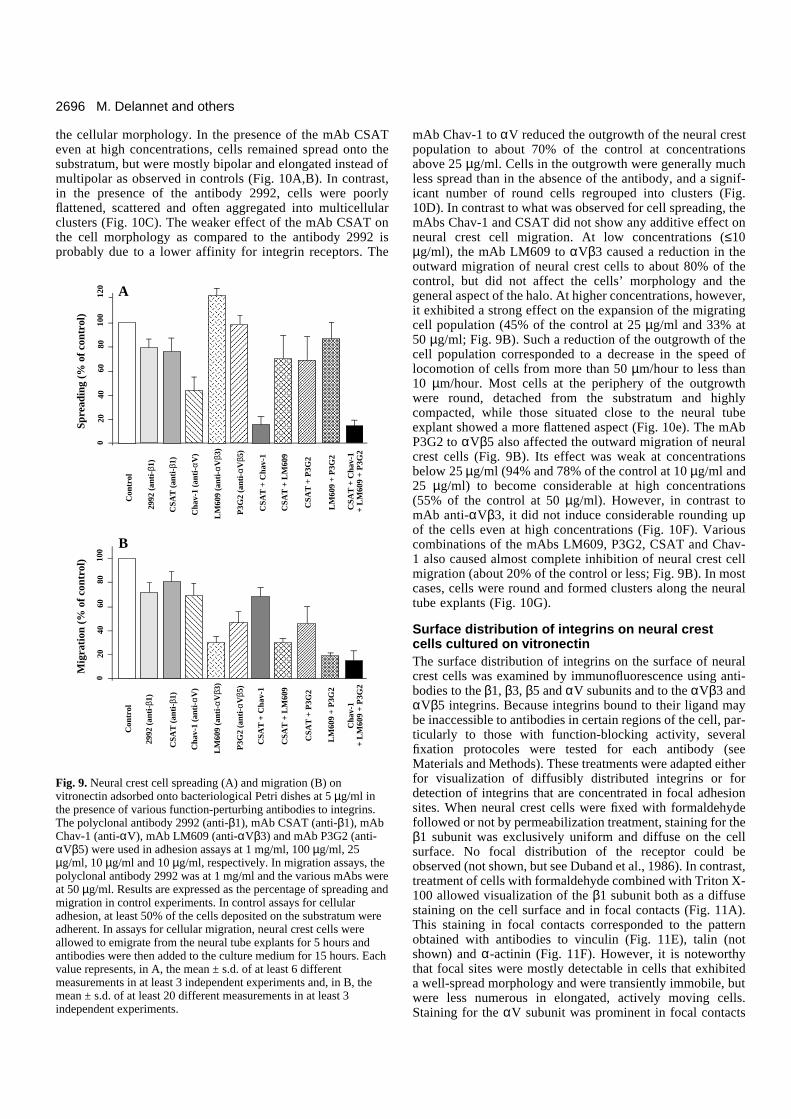
Integrin receptors for vitronectin on neural crestcellsVitronectin interacts with the cells’ surface primarily throughintegrins containing the αV subunit (Felding-Habermann andCheresh, 1993). This subunit has been found to combine tomultiple β chains, including β1, β3, β5, β6 and β8. Amongthese heterodimers, the αVβ1, αVβ3 and αVβ5 integrins arethe most abundant and have each been shown to mediatecellular adhesion to vitronectin (Pytela et al., 1985; Suzuki etal., 1986; Cheresh and Spiro, 1987; Freed et al., 1989; Chereshet al., 1989; Bodary and McLean, 1990; Smith et al., 1990;Krissansen et al., 1990), although they may also bind otherligands. For example, αVβ1 binds fibronectin (Vogel et al.,1990) and αVβ3 is highly promiscuous recognizing the RGDsequence in a wide array of matrix proteins. In contrast, theαVβ6 heterodimer has been found so far to constitute only afibronectin receptor while the ligand specificity of αVβ8remains still to be defined (Moyle et al., 1991; Busk et al.,
1992). To characterize the vitronectin-binding integrins thatare expressed by neural crest cells, cells were grown on vit-ronectin substrata and metabolically labelled. Membraneextracts were then subjected to immunoprecipitation with anti-bodies raised against the cytoplasmic domain of the chickenαV subunit (polyclonal antibody VNR-C3), the chicken β1integrin subunit (polyclonal antibody 2992), the human αVβ3integrin (mAb LM609) and the cytoplasmic domains of thehuman β3 and β5 integrin subunits (polyclonal antibodies 344and 4377).
As shown on Fig. 8B, lane 1, the antibody VNR-C3 to theαV chain immunoprecipitated a set of proteins of Mr of about150K, 140K, 110K, 95K and 85K in non-reducing conditions.The Mr 150K and 140K bands corresponded to the αV subunitand its biosynthetic precursor, respectively (Cheresh and Spiro,1987; Bossy and Reichardt, 1990). The Mr 110K band comi-grated with precisely the same mobility as the β1 chainimmunoprecipitated with the anti-β1 antibody 2992 (Fig. 8B,
Fig. 5. Migration of neural crest cells on vitronectin adsorbed to the substratum at the concentrations of 50 µg/ml (A,B) and 0.5 µg/ml (C,D).(B,D) Higher magnifications of A and C, respectively, showing the morphology of cells. Neural tube explants were deposited onto vitronectin-coated bacteriological Petri dishes and neural crest cells were allowed to migrate on the substratum for 15 hours in serum-free medium and thenphotographed. At high coating concentrations, vitronectin yields a large number of well-spread cells that migrate long distances from the neuraltube. At low concentrations, vitronectin also produces significant migration of cells, though most of them are poorly spread. nt, neural tube.
A B
C D
2694
lane 2), and the Mr 95K and 85K bands comigrated with theβ3 and β5 subunits immunoprecipitated with the mAb LM609(anti-αVβ3) and the antibody 4377 (anti-β5), respectively(Fig. 8B, lanes 3, 4). However, the presence of other αV-asso-ciated β chains comigrating with the β3 subunit (i.e. β6 andβ8) cannot be excluded (Moyle et al., 1991; Busk et al., 1992).It should be noted that immunoprecipitations of the αV chainfrom neural crest cell extracts produced bands in apparent
M. Delannet and others
Attachment (% of control)
0 20 40 60 80 100
AAAAAA
AAAAAAAAAAAA
Control
RGDS 1 mg/ml
RGDS 0.1 mg/ml
Ig anti VN 1 mg/ml
Ig anti FN 1 mg/ml
Figure 6: Delannet et al.
A
C
AAAA
AAAA
Control
RGDS 0.1 mg/ml
Ig anti VN 1 mg/ml
Ig anti FN 1 mg/ml
RGDS 1 mg/ml
B
0 100 200 300 400 500
Control
RGDS 0.1 mg/ml
Ig anti VN 1 mg/ml
Ig anti FN 1 mg/ml
RGDS 1 mg/ml
AAAAAAAAAAAA
Spreading (% of control)
Migration (µm)
AAAAAAAAA
0 20 40 60 80 100
AAAAAAAAAAAAAAAAAA
Ig anti VN 0.1 mg/ml
AAAAAAAAAA
Ig anti VN 0.1 mg/ml
AAAAAAAAAAAAAAAAAA
Ig anti VN 0.1 mg/ml
Fig. 6. Neural crest cell attachment (A), spreading (B) and migration(C) on vitronectin at 5-10 µg/ml in the presence of antibodies tovitronectin at the concentrations of 1 mg/ml and 0.1 mg/ml or tofibronectin at 1 mg/ml, and of RGDS peptides at the concentrationsof 1 mg/ml and 0.1 mg/ml. In A and B, results are expressed as thepercentage of attachment and spreading in control experiments. Incontrol assays, at least 50% of the cells deposited on the substratumwere adherent. In C, neural crest cells were allowed to emigrate fromthe neural tube explants for 5 hours, and antibodies or peptides werethen applied to the culture for 15 hours in DMEM supplemented with5% serum. Results are expressed as the linear distance in µmbetween the periphery of the neural crest outgrowth and the edge ofthe neural tube explant after 15 hours of culture in the presence ofthe antibodies or peptides. Each value represents in A and B themean ± s.d. of at least six different measurements in at least threeindependent experiments and in C the mean ± s.d. of at least 20different measurements in at least 3 independent experiments.
Fig. 7. Migration of neural crest cells on vitronectin in the presenceof antibodies to vitronectin at the concentration of 1 mg/ml (A), ofantibodies to fibronectin at 1 mg/ml (B) and of RGDS peptides at 1mg/ml (C). Neural crest cells were allowed to emigrate from theneural tube explants on vitronectin at 10 µg/ml for 5 hours.Antibodies or peptides were then applied to the culture for 4 hours.Both the antibodies to vitronectin and the RGDS peptides produceextensive rounding up of neural crest cells followed by cellaggregation. nt, neural tube. Bar, 100 µm.
A
B
C
2695Avian crest cell migration on vitronectin
amounts significantly higher than any other α subunits that arepresumably expressed by neural crest cells (i.e. α1 and α5),suggesting that αV integrins are major integrins on these cells(data not shown). As described previously (Akiyama et al.,1986; Duband et al., 1988b), the antibody 2992 (anti-β1)immunoprecipitated a series of proteins of Mr 160K, 150K,140K, 110K and 100K corresponding to various α chains, theβ1 subunit and its biosynthetic precursor, respectively (Fig.8B, lane 2). The mAb LM609 (anti-αVβ3) was found toimmunoprecipitate two proteins of Mr 140K and 95K corre-sponding to the αV and β3 subunits (Fig. 8B, lane 3; Chereshand Spiro, 1987). The antibody 344 to the cytoplasmic domainof the β3 integrin subunit immunoprecipitated the sameproteins as the mAb LM609 (not shown). Finally, the antibody4377 (anti-β5) immunoprecipitated two proteins of Mr 140Kand 85K most likely corresponding to the αV and β5 subunits(Fig. 8B, lane 4; Busk et al., 1992).
In order to further characterize the vitronectin-bindingintegrins expressed by neural crest cells and, more specifically,to detect any β3 and β5 integrins that do not contain the αVchain, extracts of metabolically labelled of neural crest cellswere first immunodepleted with the antibody VNR-C3 toremove the αV integrins and were subsequently subjected toimmunoprecipitation with the antibody 344 (anti-β3) or theantibody 4377 (anti-β5). No band migrating at Mr 95K and 85Kcould be detected in these immune precipitates, indicating thatvirtually all the β3 and β5 chains are associated with the αVsubunit (data not shown).
These results then indicate that neural crest cells expressmultiple αV integrins in high amounts, including αVβ1, αVβ3and αVβ5 as presumptive vitronectin receptors, as well as anumber of β1 integrins.
Role of the vitronectin-binding integrins in neuralcrest cell adhesion and migration on vitronectinSince a number of potential vitronectin receptors appeared tobe expressed by neural crest cells, we have performed a series
of experiments to determine their possible involvement inneural crest cell adhesion to and migration on vitronectin usinga variety of function-blocking antibodies to integrins, eitherseparately or in combination. These antibodies were the poly-clonal antibody 2992 to the β1 subunit and the mAbs CSAT,Chav-1, LM609 and P3G2 known to inhibit β1 integrins, αVintegrins, the αVβ3 integrin and the αVβ5 integrin, respec-tively (Neff et al., 1982; Chen et al., 1985; Horwitz et al., 1985;Duband et al., 1986; Cheresh and Spiro, 1987; Cheresh, 1987;Neugebauer et al., 1991; Wayner et al., 1991). All mAbs weretested at concentrations ranging from 5 µg/ml to 50 µg/ml and,when tested in combination, they were applied at the lowestconcentration that gave the maximal inhibitory activity. Thepolyclonal antibody 2992 was tested at concentrations from 0.1mg/ml to 1 mg/ml.
In adhesion assays (Fig. 9A), the greatest effect of any ofthe antibodies was obtained with the mAb Chav-1 to αV,which reduced neural crest cell spreading to about 40% of thecontrol. The mAb CSAT and polyclonal antibody 2992 to β1had significant, though much weaker, inhibitory effects on cellspreading (about 80% of the control). In contrast, the mAbP3G2 to αVβ5 did not affect cell spreading while the mAbLM609 to αVβ3 caused a significant enhancement in neuralcrest cell spreading to vitronectin (120% of the control). Whenused in combination, these two antibodies failed to perturb celladhesion. Likewise, they did not potentiate the effect of themAb CSAT. In contrast, the mAbs CSAT and Chav-1 showeda significant additive effect and considerably perturbed neuralcrest cell spreading to less than 20% of the control. Finally,combination of the four mAbs also produced a dramatic inhi-bition of cell spreading, but to values not greater than the mAbsCSAT and Chav-1 used in combination.
In migration assays (Figs 9B, 10), the mAb CSAT and thepolyclonal antibody 2992 to the β1 chain had a weak, but sig-nificant, effect on the outward migration of neural crest cellsat all concentrations tested (about 70-80% of the control).However, the two antibodies displayed different effects on
Fig. 8. (A) Vitronectin synthesis by neural crest cells. Cell extract (lane 1) and culture supernatant (lane 2) of metabolically labeled neural crestcells were subjected to immunoprecipitation with antibodies to vitronectin. (B) Integrin receptors for vitronectin on neural crest cells. Neuralcrest cells cultured on vitronectin were metabolically labeled, extracted in detergent and the lysates were subjected to immunoprecipitationusing the VNR-C3 polyclonal antibody to the cytoplasmic domain of the chicken αV integrin subunit (lane 1), the 2992 polyclonal antibody tothe chicken β1 integrin subunit (lane 2), the LM609 monoclonal antibody to the human αVβ3 integrin complex (lane 3) and the 4377polyclonal antibody to the cytoplasmic domain of the human β5 integrin subunit (lane 4). Samples were resolved by SDS-PAGE on 10%acrylamide gels under reducing conditions in A and on 7.5% acrylamide gels under nonreducing conditions in B and radiolabeled bands werevisualized by fluorography. Molecular mass markers are indicated on the left of panels A and B.
A B
2696
the cellular morphology. In the presence of the mAb CSATeven at high concentrations, cells remained spread onto thesubstratum, but were mostly bipolar and elongated instead ofmultipolar as observed in controls (Fig. 10A,B). In contrast,in the presence of the antibody 2992, cells were poorlyflattened, scattered and often aggregated into multicellularclusters (Fig. 10C). The weaker effect of the mAb CSAT onthe cell morphology as compared to the antibody 2992 isprobably due to a lower affinity for integrin receptors. The
mAb Chav-1 to αV reduced the outgrowth of the neural crestpopulation to about 70% of the control at concentrationsabove 25 µg/ml. Cells in the outgrowth were generally muchless spread than in the absence of the antibody, and a signif-icant number of round cells regrouped into clusters (Fig.10D). In contrast to what was observed for cell spreading, themAbs Chav-1 and CSAT did not show any additive effect onneural crest cell migration. At low concentrations (≤10µg/ml), the mAb LM609 to αVβ3 caused a reduction in theoutward migration of neural crest cells to about 80% of thecontrol, but did not affect the cells’ morphology and thegeneral aspect of the halo. At higher concentrations, however,it exhibited a strong effect on the expansion of the migratingcell population (45% of the control at 25 µg/ml and 33% at50 µg/ml; Fig. 9B). Such a reduction of the outgrowth of thecell population corresponded to a decrease in the speed oflocomotion of cells from more than 50 µm/hour to less than10 µm/hour. Most cells at the periphery of the outgrowthwere round, detached from the substratum and highlycompacted, while those situated close to the neural tubeexplant showed a more flattened aspect (Fig. 10e). The mAbP3G2 to αVβ5 also affected the outward migration of neuralcrest cells (Fig. 9B). Its effect was weak at concentrationsbelow 25 µg/ml (94% and 78% of the control at 10 µg/ml and25 µg/ml) to become considerable at high concentrations(55% of the control at 50 µg/ml). However, in contrast tomAb anti-αVβ3, it did not induce considerable rounding upof the cells even at high concentrations (Fig. 10F). Variouscombinations of the mAbs LM609, P3G2, CSAT and Chav-1 also caused almost complete inhibition of neural crest cellmigration (about 20% of the control or less; Fig. 9B). In mostcases, cells were round and formed clusters along the neuraltube explants (Fig. 10G).
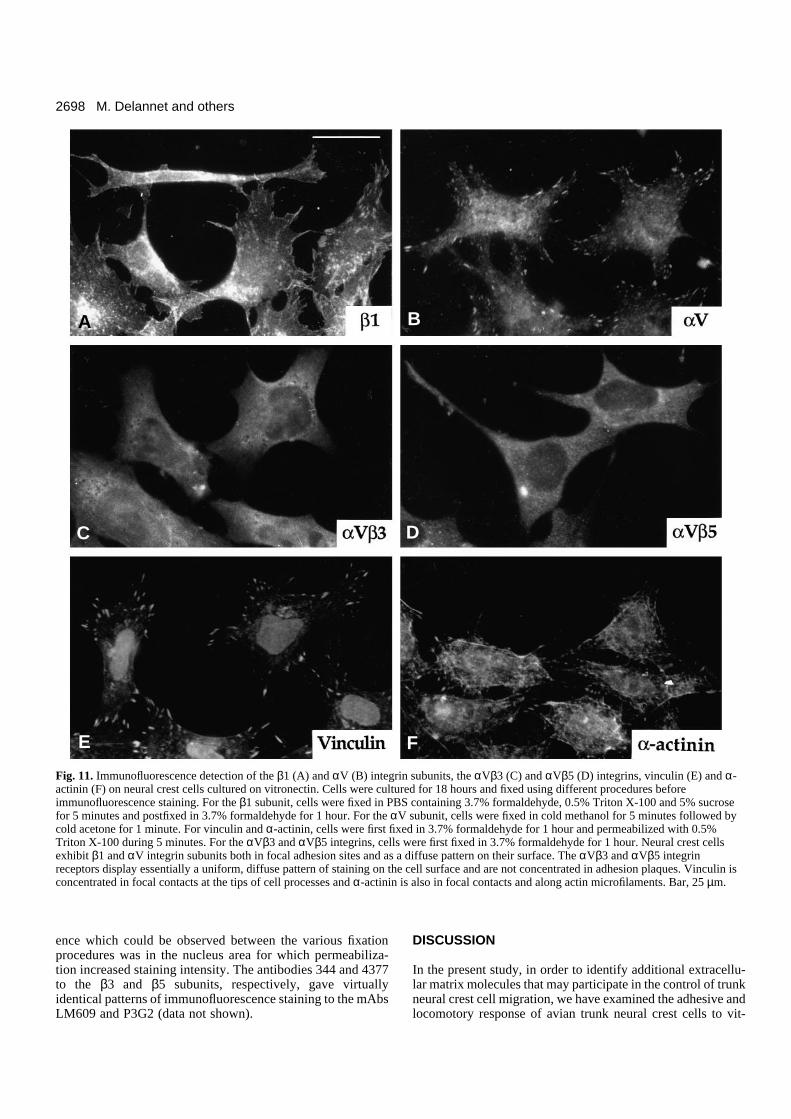
Surface distribution of integrins on neural crestcells cultured on vitronectinThe surface distribution of integrins on the surface of neuralcrest cells was examined by immunofluorescence using anti-bodies to the β1, β3, β5 and αV subunits and to the αVβ3 andαVβ5 integrins. Because integrins bound to their ligand maybe inaccessible to antibodies in certain regions of the cell, par-ticularly to those with function-blocking activity, severalfixation protocoles were tested for each antibody (seeMaterials and Methods). These treatments were adapted eitherfor visualization of diffusibly distributed integrins or fordetection of integrins that are concentrated in focal adhesionsites. When neural crest cells were fixed with formaldehydefollowed or not by permeabilization treatment, staining for theβ1 subunit was exclusively uniform and diffuse on the cellsurface. No focal distribution of the receptor could beobserved (not shown, but see Duband et al., 1986). In contrast,treatment of cells with formaldehyde combined with Triton X-100 allowed visualization of the β1 subunit both as a diffusestaining on the cell surface and in focal contacts (Fig. 11A).This staining in focal contacts corresponded to the patternobtained with antibodies to vinculin (Fig. 11E), talin (notshown) and α-actinin (Fig. 11F). However, it is noteworthythat focal sites were mostly detectable in cells that exhibiteda well-spread morphology and were transiently immobile, butwere less numerous in elongated, actively moving cells.Staining for the αV subunit was prominent in focal contacts
M. Delannet and others
Con
trol
020
4060
8010
0
Mig
rati
on (
% o
f co
ntro
l)Sp
read
ing
(% o
f co
ntro
l)
Con
trol
020
4060
8010
012
0 A
B
AAAAAAAAAAAAAAAAAA
CSA
T +
Cha
v-1
Cha
v-1
+ L
M60
9 +
P3G
2
CSA
T +
LM
609
AAAAAAAAA
CSA
T +
P3G
2
AAAAAAAAAAAAAAA
AAAAAAAAA
LM
609
+ P
3G2
LM
609
(ant
i-αV
β3)
CSA
T (
anti
-β1)
Cha
v-1
(ant
i-αV
)
AAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
AAAAAAAAAAAA
AAAAAAAAAAAAAAAAAAAAAAAA
CSA
T +
Cha
v-1
CSA
T +
Cha
v-1
+ L
M60
9 +
P3G
2
P3G
2 (a
nti-
αVβ5
)
CSA
T +
LM
609
AAAAAAAAAAAAAAAAAA
CSA
T +
P3G
2
AAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAAAAA
LM
609
+ P
3G2
AAAAAAAAAAAAAAA
2992
(an
ti-β
1)
AAAAAAAAA
LM
609
(ant
i-αV
β3)
CSA
T (
anti
-β1)
Cha
v-1
(ant
i-αV
)
P3G
2 (a
nti-
αVβ5
)
2992
(an
ti-β
1)
Fig. 9. Neural crest cell spreading (A) and migration (B) onvitronectin adsorbed onto bacteriological Petri dishes at 5 µg/ml inthe presence of various function-perturbing antibodies to integrins.The polyclonal antibody 2992 (anti-β1), mAb CSAT (anti-β1), mAbChav-1 (anti-αV), mAb LM609 (anti-αVβ3) and mAb P3G2 (anti-αVβ5) were used in adhesion assays at 1 mg/ml, 100 µg/ml, 25µg/ml, 10 µg/ml and 10 µg/ml, respectively. In migration assays, thepolyclonal antibody 2992 was at 1 mg/ml and the various mAbs wereat 50 µg/ml. Results are expressed as the percentage of spreading andmigration in control experiments. In control assays for cellularadhesion, at least 50% of the cells deposited on the substratum wereadherent. In assays for cellular migration, neural crest cells wereallowed to emigrate from the neural tube explants for 5 hours andantibodies were then added to the culture medium for 15 hours. Eachvalue represents, in A, the mean ± s.d. of at least 6 differentmeasurements in at least 3 independent experiments and, in B, themean ± s.d. of at least 20 different measurements in at least 3independent experiments.

2697Avian crest cell migration on vitronectin
when cold methanol-acetone fixation was used (Fig. 11B), butwas also found out of focal contacts when other fixation pro-cedures were employed. For mAbs LM609 and P3G2 to αVβ3
and αVβ5, all fixation procedures gave essentially a uniform,diffuse staining on the cell surface, which showed little colo-calization with focal contacts (Fig. 11C,D). The only differ-
Fig. 10. Migration of neural crest cells on vitronectin in the presence of various function-perturbing antibodies to integrins. Neural crest cellswere allowed to emigrate from the neural tube explants on vitronectin at the concentration of 10 µg/ml for 5 hours. Antibodies were thenapplied to the culture for 15 hours. (A) Control without antibody, (B) mAb CSAT (anti-β1) at 250 µg/ml, (C) polyclonal antibody 2992 (anti-β1) at 0.5 mg/ml, (D) mAb Chav-1 (anti-αV) at 50 µg/ml, (E) mAb LM609 (anti-αVβ3) at 50 µg/ml, (F) mAb P3G2 (anti-αVβ5) at 50 µg/mland (G) mAbs LM609 and P3G2 both at 50 µg/ml. nt, neural tube. Bar, 100 µm.
A
D
E
F
G
B
C
2698
ence which could be observed between the various fixationprocedures was in the nucleus area for which permeabiliza-tion increased staining intensity. The antibodies 344 and 4377to the β3 and β5 subunits, respectively, gave virtuallyidentical patterns of immunofluorescence staining to the mAbsLM609 and P3G2 (data not shown).
DISCUSSION
In the present study, in order to identify additional extracellu-lar matrix molecules that may participate in the control of trunkneural crest cell migration, we have examined the adhesive andlocomotory response of avian trunk neural crest cells to vit-
M. Delannet and others
Fig. 11. Immunofluorescence detection of the β1 (A) and αV (B) integrin subunits, the αVβ3 (C) and αVβ5 (D) integrins, vinculin (E) and α-actinin (F) on neural crest cells cultured on vitronectin. Cells were cultured for 18 hours and fixed using different procedures beforeimmunofluorescence staining. For the β1 subunit, cells were fixed in PBS containing 3.7% formaldehyde, 0.5% Triton X-100 and 5% sucrosefor 5 minutes and postfixed in 3.7% formaldehyde for 1 hour. For the αV subunit, cells were fixed in cold methanol for 5 minutes followed bycold acetone for 1 minute. For vinculin and α-actinin, cells were first fixed in 3.7% formaldehyde for 1 hour and permeabilized with 0.5%Triton X-100 during 5 minutes. For the αVβ3 and αVβ5 integrins, cells were first fixed in 3.7% formaldehyde for 1 hour. Neural crest cellsexhibit β1 and αV integrin subunits both in focal adhesion sites and as a diffuse pattern on their surface. The αVβ3 and αVβ5 integrinreceptors display essentially a uniform, diffuse pattern of staining on the cell surface and are not concentrated in adhesion plaques. Vinculin isconcentrated in focal contacts at the tips of cell processes and α-actinin is also in focal contacts and along actin microfilaments. Bar, 25 µm.
A B
C
E F
D

2699Avian crest cell migration on vitronectin
ronectin in vitro. Our major findings are the following: (i) vit-ronectin is widely expressed in the developing embryo and isfound on the surface of migrating neural crest cells; (ii) vit-ronectin promotes adhesion and migration of neural crest cellsthrough its RGD sequence; (iii) neural crest cells express atleast the αVβ1, αVβ3 and αVβ5 integrins as vitronectinreceptors; (iv) adhesion of neural crest cells to vitronectin canbe affected by antibodies to the αV integrin subunit, whereasmigration is almost totally abolished by antibodies to the αVβ3and αVβ5 integrins; (v) the various αV integrins are differen-tially distributed on the surface of neural crest cells.
Substratum-bound vitronectin has been shown to promoteadhesion of a large variety of cell lines as well as of severalprimary cultured cells, including smooth muscles, neurons,platelets and endothelial cells (Neugebauer et al., 1991, 1992;Grabham et al., 1992; Clyman et al., 1992; and reviewed inPreissner, 1991). In addition, in in vitro assays, vitronectin wasfound to stimulate keratinocyte and smooth muscle cellmotility (Brown et al., 1991; Clyman et al., 1992), melanomaand glioblastoma cell invasion (Seftor et al., 1991; Gladson andCheresh, 1991), and retinal neurite outgrowth (Neugebauer etal., 1991). However, apart from a few studies in the adultanimal (Hayman et al., 1983; and reviewed in Preissner, 1991),little is known about the tissular distribution of vitronectin andits possible involvement in morphogenetic events duringembryonic development. The present demonstration thatneural crest cells can adhere to and migrate on vitronectinsubstrata in vitro suggests that this molecule may play a sig-nificant role in the control of cellular adhesiveness and in cellmigrations during embryonic development. This possibility issubstantiated by the widespread occurence of vitronectin in theavian embryo.
A detailed analysis of the distribution of vitronectin intissues and in cultured cells reveals that it shares some commonfeatures with fibronectin. For example, vitronectin was foundin connective tissues and in cultured fibroblasts as a fibrillarpattern that overlaps frequently with fibronectin fibrils(Hayman et al., 1983; Delannet and Duband, unpublishedresults). These observations then suggest that fibronectin andvitronectin may play similar roles in substratum adhesion ofcells, consistent with the fact that their corresponding integrinreceptors, namely α5β1 and αVβ3, codistribute in focalcontacts of cells cultured on a mixed matrix of vitronectin andfibronectin (Singer et al., 1988; Dejana et al., 1988). However,both molecules were found to exhibit a number of strikingspecificities in their distribution during early embryonic devel-opment. Unlike fibronectin, vitronectin was neither concen-trated in the basement membrane of epithelial sheets nor foundabundantly in acellular areas adjacent to tissues. Instead, it wasclosely associated with the surface of cells, even in epitheliawhere it was present on the lateral surfaces of the cells. Thesedifferences in the cellular distributions of fibronectin and vit-ronectin may reflect significant differences in the mode ofaction of these two molecules in the control of cellular adhe-siveness during embryonic development and, more specifi-cally, during neural crest cell migration. Interestingly, neuralcrest cells themselves are the source of vitronectin present ontheir surface, at least in vitro, whereas fibronectin which is notproduced by neural crest cells is deposited ahead of them bycells lining the migration pathways (Newgreen and Thiery,1980; ffrench-Constant and Hynes, 1988). Thus, fibronectin, in
association with collagens, would serve as a scaffold ontowhich neural crest cells migrate. Conversely, vitronectinreleased by neural crest cells would be immediately trapped tothe matrix due to its binding domains to collagens and heparinor would spontaneously form multimers (see for example,Preissner, 1991; Stockmann et al., 1993). Consequently, itwould be immobilized in the vicinity of the cell surface,thereby generating a local environment that would facilitatecell locomotion. This particular distribution of vitronectin onthe surface of locomoting cells has been described in othersystems, supporting the notion that motile cells would providetheir own vitronectin substratum for migration. For example,vitronectin is the sole adhesive matrix component found inglioblastoma cells and it is specifically localized to the tumorduring invasion of normal brain (Gladstone and Cheresh,1991). Likewise, it is expressed in the retina adjacent to theoptic fiber layer at the time of neurite extension and disappearsfrom this site thereafter (Neugebauer et al., 1991).
Perturbation experiments using RGDS peptides, solubleheparin and antibodies against integrins indicate that neuralcrest cells interact with vitronectin primarily through integrinreceptors in an RGD-dependent manner. Furthermore,immunoprecipitation and immunofluorescence analysesrevealed that neural crest cells express, at least in vitro, theαVβ1, αVβ3 and αVβ5 integrins that have been shown pre-viously to constitute major vitronectin-binding receptors for avariety of cell types (Pytela et al., 1985; Suzuki et al., 1986;Cheresh and Spiro, 1987; Freed et al., 1989; Cheresh et al.,1989; Bodary and McLean, 1990; Smith et al., 1990; Kris-sansen et al., 1990). Consistent with these findings, experi-ments in progress in our laboratory indicate that, also in vivo,neural crest cells express at least the αV, β1 and β3 integrinsubunits. However, as suggested by immunoprecipitation andperturbation experiments, it cannot be excluded that otherintegrins are expressed by neural crest cells and may partici-pate to some extent in cell adhesion to vitronectin. Theseinclude β1 integrins containing an α chain distinct from αVand other αV integrins, e.g. αVβ6 or αVβ8.
The diversity of integrins expressed by neural crest cellsraises the problem of determining their exact functional speci-ficities. In other words, are these integrins interchangeable forpromoting adhesion and migration of neural crest cells to vit-ronectin or do they play distinct roles in these processes? Ininhibition assays of cellular adhesion using function-perturb-ing antibodies, we found that neural crest cell adhesion to vit-ronectin could be strongly impaired only in the presence of themAb Chav-1 to the αV subunit, suggesting that adhesion ismediated primarily by αV integrins. Moreover, the mAb to theβ1 subunit potentiated the mAb to the αV subunit suggestingthat, in addition to αV integrins, a β1 integrin containing an αchain distinct from αV may be involved in adhesion of neuralcrest cells to vitronectin. However, it should be mentioned that,apart from αVβ1, none of the β1 integrins described so farhave been found to bind vitronectin with significant efficiency.Therefore, if they exist, such integrins remain to be defined.Alternatively, a possible explanation to the additive effect ofthe mAbs to αV and β1 is that these antibodies when used sep-arately were not able to inhibit entirely the αVβ1 integrin andthat they were totally efficient only in combination. In contrast,adhesion of neural crest cells to vitronectin was not affectedby mAbs LM609 and P3G2 to the αVβ3 and αVβ5 integrins

2700
and only weakly by antibodies to the β1 integrins. This obser-vation would then suggest that the αVβ1, αVβ3 and αVβ5integrins might play only subsidiary roles in cell adhesion tovitronectin and that this process is essentially mediated by thetwo other αV integrins, αVβ6 and αVβ8. However, there isno indication yet that either αVβ6 or αVβ8 can function as vit-ronectin receptors. The αVβ6 integrin has only been found sofar to constitute a fibronectin receptor (Busk et al., 1992)whereas the ligand specificity of αVβ8 remains still to bedefined. Moreover, in contrast to the other αV integrins, αVβ8is expressed in low amounts by a very limited number of celltypes, suggesting a specialized function for this integrin(Moyle et al., 1991). Therefore, a more plausible explanationto the limited effect of antibodies to individual integrins is thatadhesion of neural crest cells to vitronectin is mediatedprimarily by the three major αV integrins (αVβ1, αVβ3 andαVβ5) and that these integrins can substitute for promoting celladhesion.
We have observed, however, that when the antibodies wereapplied to migrating neural crest cells that are already spreadonto vitronectin, only antibodies to the β1 chain and, in a lesserextent, to the αV chain could induce rounding up of the cells.The mAb P3G2 to αVβ5 did not affect much cell spreadingand the mAb LM609 to αVβ3 provoked rounding up only afterseveral hours. This indicates that, although these integrins maybe interchangeable in adhesion assays, αVβ1 would be pre-eminent over αVβ3 and αVβ5 for substratum anchorage.Therefore, there might exist a hierachy among αV integrins formediating cell adhesion, that would be apparent in locomotingcells but not in cells in suspension.
Concerning cell migration, our data demonstrate that, incontrast to β1 integrins, αVβ3 and αVβ5 appear to play acritical role in cell migration as the mAbs LM609 and P3G2reduced considerably the outward migration of neural crestcells. In addition, the fact that cell migration could beprevented by either the mAb to αVβ3 or the mAb to αVβ5 andthat these antibodies showed different effects on the cell mor-phology indicates that the αVβ3 and αVβ5 integrins cannotsubstitute for cell migration and that they may participate indifferent events of the locomotory process. In an apparent con-tradiction with these data, the mAb Chav-1 to the αV subunitshowed only a weak effect on neural crest cell migrationwhereas it affected cell spreading with high efficiently inadhesion assays. This surprising result can be explained by ahigher affinity of the antibody for αVβ1 than for αVβ3 orαVβ5. Alternatively, we have observed that the mAb Chav-1presents a relatively low affinity for the αV subunit (data notshown). Such a low affinity would make the antibody capableof binding to integrins that are readily accessible in cells in sus-pension, thereby preventing their further association with vit-ronectin, but would make it unable to displace the preexistingbounds between integrin and vitronectin molecules in alreadyspread cells.
Thus, it appears from our study that neural crest cells usedifferent integrins to perform the separate functions ofadhesion and migration: The αVβ1 integrin would be involvedessentially in cellular adhesion while the αVβ3 and αVβ5integrins would participate primarily in cell migration. Ourdata are in complete agreement with previous studies showingthat different integrins that bind common ligands may differ intheir functional specificities. The αVβ1 integrin, for example,
has been found to function as a fibronectin receptor, yet, incontrast to α5β1, it is not able to support fibronectin matrixassembly and cell motility (Zhang et al., 1993). Likewise, theαVβ3 and αVβ5 integrins, expressed on the same cell,promote distinct cellular responses to vitronectin substrata(Leavesley et al., 1992). Finally, vascular smooth muscle cellsdepend exclusively on β1 integrins for adhesion to extracellu-lar matrix components, but depend in a large extent on αVβ3for motility on the same molecules (Clyman et al., 1992).
The functional specificities of the αV integrins is reflectedby their distributions on the surface of neural crest cells andtheir possible modes of association with cytoskeletal elements.The αVβ3 and αVβ5 integrins were found essentially as apunctate distribution over much of the cell surface and werenot particularly concentrated in focal contacts. In contrast,although it was not possible to localize the αVβ1 integrin byitself, staining for the αV and β1 subunits suggests that it isabundant in focal contacts in association with talin, vinculinand α-actinin, in addition to a diffuse distribution over the cellsurface. As proposed previously (Duband et al., 1988a;Clyman et al., 1992), the diffuse location of the αVβ3 andαVβ5 integrins could favor transient cell-matrix interactionsnecessary for cell locomotion whereas concentration of αVβ1integrins in focal contacts would permit anchoring of the cellto the substratum.
It has been proposed that the β1 class of integrin receptorsis the prevailing category of integrins expressed by neural crestcells, based on the strong immunoreactivity of antibodies to β1integrins on neural crest cells and on their potent effect ininhibiting cell adhesion on fibronectin and laminin (Bronner-Fraser, 1985; Duband et al., 1986; Krotoski et al., 1986).However, except for α1, the identity of the α chains associatedwith the β1 subunit to compose the fibronectin and lamininreceptors on neural crest cells have not been defined yet(Lallier and Bronner-Fraser, 1992; Bronner-Fraser, 1993;Erickson and Perris, 1993). Our study demonstrates for the firsttime that, in addition to β1 integrins, non-β1 integrins, such asαVβ3 and αVβ5, may play a key role in neural crest cellmigration. Moreover, as a number of the vitronectin-bindingintegrins bind also fibronectin (i.e. αVβ1 and αVβ3) andlaminin (i.e. αVβ3), and as αV integrins are expressed in highamounts in neural crest cells as compared to α1β1 and α5β1,it may be proposed that these integrins could be also involvedin adhesion and migration of neural crest cells on fibronectinand laminin.
In summary, our study demonstrates that neural crest cellscan adhere to and migrate on vitronectin in vitro in amechanism involving distinct αV integrins that are differen-tially distributed on the cell surface, possibly resulting fromdifferences in their modes of association with the cytoskeletonand in the types of intracellular signals they transduce.Studying these specificities should help us to understand theprocess of cell motility in greater detail.
We are extremely grateful to Ken Yamada for useful discussionsand advice and for providing antibodies to β1 integrins so generously.Many thanks to Masa Hayashi for providing antibodies to vitronectinand for helpful discussions, to Kristine Venstrom for the antibodiesto the β5 subunit and to Clayton Buck for the CSAT hybridoma cellline. This work was supported by the Centre National de la RechercheScientifique (Programme ATIPE), the Ministère de la Recherche etde la Technologie (91.T.0011), the Association pour la Recherche
M. Delannet and others

2701Avian crest cell migration on vitronectin
contre le Cancer (ARC-6517), the Institut National de la Santé et dela Recherche Médicale (CRE 910705), the Association Françaisecontre les Myopathies and the Fondation pour la Recherche Médicale.M. D. is a recipient of a predoctoral fellowship of the Ministère del’Enseignement Supérieur et de la Recherche.
REFERENCES
Akiyama, S. K., Yamada, S. S. and Yamada, K. M. (1986). Characterizationof a 140-kD avian cell surface antigen as a fibronectin-binding molecule. J.Cell Biol. 102, 442-448.
Barnes, D. W., Reing, J. E. and Amos, B. (1985). Heparin-binding propertiesof human serum spreading factor. J. Biol. Chem. 260, 9117-9122.
Bodary, S. C. and McLean, J. W. (1990). The integrin β1 subunit associateswith the vitronectin receptor αv subunit to form a novel vitronectin receptorin a human embryonic kidney cell line. J. Biol Chem. 265, 5938-5941.
Bossy, B. and Reichardt, L. F. (1990). Chick integrin αV subunit molecularanalysis reveals high conservation of structural domains and association withmultiple β subunits in embryo fibroblasts. Biochemistry 29, 10191-10198.
Boucaut, J.-C., Darribère, T., Poole, T. J., Aoyama, H., Yamada, K. M. andThiery, J. P. (1984). Biological active synthetic peptides as probes ofembryonic development: A competitive peptide inhibitor of fibronectinfunction inhibits gastrulation in amphibian embryos and neural crest cellmigration in avian embryos. J. Cell Biol. 99, 1822-1830.
Bronner-Fraser, M. (1985). Alteration in neural crest migration by amonoclonal antibody that affects cell adhesion. J. Cell Biol. 101, 610-617.
Bronner-Fraser, M. (1993). Neural crest migration in the developing embryo.Trends Cell Biol. 3, 392-397.
Bronner-Fraser, M. and Lallier, T. (1991). A monoclonal antibody against alaminin-heparan sulfate proteoglycan complex perturbs cranial neural crestmigration in vivo. J. Cell Biol. 106, 1321-1329.
Brown, C., Stenn, K. S., Falk, R. J., Woodley, D. T. and O’Keefe, E. J.(1991). Vitronectin: Effects on keratinocyte motility and inhibition ofcollagen-induced motility. J. Invest. Dermatol. 96, 724-728.
Busk, M., Pytela, R. and Sheppard, D. (1992). Characterization of theintegrin αVβ6 as a fibronectin-binding protein. J. Biol. Chem. 267, 5790-5796.
Chen, W.-T., Hasegawa, E., Hasegawa, T., Weinstock, C. and Yamada, K.M. (1985). Development of cell surface linkage complexes in culturedfibroblasts. J. Cell Biol. 100, 1103-1114.
Cheresh, D. A. (1987). Human endothelial cells synthesize and express an Arg-Gly-Asp-directed adhesion receptor involved in attachment to fibrinogen andvon Willebrand factor. Proc. Natl. Acad. Sci. USA 84, 6471-6475.
Cheresh, D. A. and Spiro, R. C. (1987). Biosynthetic and functional propertiesof an Arg-Gly-Asp-directed receptor involved in human melenoma cellattachment to vitronectin, fibrinogen and von Willebrand factor. J. Biol.Chem. 262, 17703-17711.
Cheresh, D. A., Smith, J. W., Cooper, H. M. and Quaranta, V. (1989). Anovel vitronectin receptor integrin (αvβx) is responsible for distinct adhesiveproperties of carcinoma cells. Cell 57, 59-69.
Clyman, R. I., Mauray, F. and Kramer, R. H. (1992). β1 and β3 integrinshave different roles in the adhesion and migration of vascular smooth musclecells on extracellular matrix. Exp. Cell Res. 200, 272-284.
Dejana, E., Colella, S., Conforti, G., Abbadini, M., Gaboli, M. andMarchisio, P. C. (1988). Fibronectin and vitronectin regulate theorganization of their respective Arg-Gly-Asp receptors in cultured humanendothelial cells. J. Cell Biol. 107, 1215-1223.
Duband, J.-L. and Thiery, J. P. (1982). Distribution of fibronectin in the earlyphase of avian cephalic neural crest cell migration. Develop. Biol. 93, 308-323.
Duband, J.-L. and Thiery, J. P. (1987). Distribution of laminin and collagensduring avian neural crest development. Development 101, 461-478.
Duband, J.-L., Rocher, S., Chen, W.-T., Yamada, K. M. and Thiery, J. P.(1986). Cell adhesion and migration in the early vertebrate embryo: Locationand possible role of the putative fibronectin-receptor complex. J. Cell Biol.102, 160-178.
Duband, J.-L., Nuckolls, G. H., Ishihara, A., Hasegawa, T., Yamada, K.M., Thiery, J. P. and Jacobson, K. (1988a). The fibronectin receptorexhibits high lateral mobility in embryonic locomoting cells but is immobilein focal contacts and fibrillar streaks in stationary cells. J. Cell Biol. 107,1385-1396.
Duband, J.-L., Dufour, S., Yamada, K. M. and Thiery, J. P. (1988b). The
migratory behavior of avian embryonic cells does not requirephosphorylation of the fibronectin-receptor complex. FEBS Lett. 230, 181-185.
Duband, J.-L., Dufour, S., Yamada, S. S., Yamada, K. M. and Thiery, J. P.(1991). Neural crest cell locomotion induced by antibodies to β1 integrins: Atool for studying the roles of substratum molecular avidity and density inmigration. J. Cell Sci. 98, 517-532.
Dufour, S., Duband, J.-L., Humphries, M. J., Obara, M., Yamada, K. M.and Thiery, J. P. (1988). Attachement, spreading, and locomotion of avianneural crest cells are mediated by multiple adhesion sites on fibronectinmolecules. EMBO J. 7, 2661-2671.
Erickson, C.A. and Perris, R. (1993). The role of cell-cell and cell-matrixinteractions in the morphogenesis of the neural crest. Develop. Biol. 159, 60-74.
Felding-Habermann, B. and Cheresh, D. A. (1993). Vitronectin and itsreceptors. Curr. opin. Cell Biol. 5, 864-868.
ffrench-Constant, C. and Hynes, R. O. (1988). Pattern of fibronectin geneexpression and splicing during cell migration in chicken embryo.Development 104, 369-382.
Freed, E., Gailit, J., Van der Geer, P., Ruoslahti, E. and Hunter, T. (1989).A novel integrin β subunit is associated with the vitronectin receptor αsubunit (αV) in a human osteosarcoma cell line and is a substrate for proteinkinase C. EMBO J. 8, 2955-2965.
Gladson, C. L. and Cheresh, D. A. (1991). Glioblastoma expression ofvitronectin and the αVβ3 integrin: Adhesion mechanism of transformed glialcells. J. Clin. Invest. 88, 1924-1932.
Grabham, P. W., Gallimore, P. H. and Grand, R. J. A. (1992). Vitronectin isthe major serum protein essential for NGF-mediated neurite outgrowth fromPC12 cells. Exp. Cell Res. 202, 337-344.
Hayman, E. G., Pierschbacher, M. D., Ohgren, Y. and Ruoslahti, E. (1983).Serum spreading factor (vitronectin) is present at the cell surface and intissues. Proc. Natl. Acad. Sci. USA 80, 4003-4007.
Horwitz, A., Duggan, K., Greggs, R., Decker, C. and Buck, C. (1985). Thecell substrate attachment (CSAT) antigen has properties of a receptor forlaminin and fibronectin. J. Cell Biol. 101, 2134-2144.
Jaffredo, T., Horwitz, A. F., Buck, C. A., Rong, P.-M. and Dieterlen-Lièvre,F. (1988). Myoblast migration specifically inhibited in the chick embryo bygrafted CSAT hybridoma cells secreting an anti-integrin antibody.Development 103, 431-446.
Kitagaki-Ogawa, H., Yatohgo, T., Izumi, M., Hayashi, M., Kashiwagi, H.,Matsumoto, I. and Seno, N. (1990). Diversities in animal vitronectins.Differences in molecular weight, immunoreactivity and carbohydrate chains.Biochim. Biophys. Acta 1033, 49-56.
Krissansen, G. W., Elliott, M. J., Lucas, C. M., Stomski, F. C., Berndt, M.C., Cheresh, D. A., Lopez, A. F. and Burns, G. F. (1990). Identification ofa novel integrin β subunit expressed on cultured monocytes (macrophages).Evidence that one α subunit can associate with multiple β subunit. J. Biol.Chem. 265, 823-830.
Krotoski, D. M., Domingo, C. and Bronner-Fraser, M. (1986). Distributionof a putative cell surface receptor for fibronectin and laminin in the avianembryo. J. Cell Biol. 103, 1061-1071.
Lallier, T. and Bronner-Fraser, M. (1992). α1β1 integrin on neural crest cellsrecognizes some laminin substrata in a Ca2+-independent manner. J. CellBiol. 119, 1335-1345.
Lallier, T., Leblanc, G., Artinger, K. B. and Bronner-Fraser, M. (1992).Cranial and trunk neural crest cells use different mechanisms for attachmentto extracellular matrices. Development 116, 531-541.
Leavesley, D. I., Ferguson, G. D., Wayner, E. A. and Cheresh, D. A. (1992).Requirement of the integrin β3 subunit for carcinoma cell spreading ormigration on vitronectin and fibrinogen. J Cell Biol. 117, 1101-1107.
Le Douarin, N. M. (1982). The Neural Crest. Cambridge University Press.Cambridge.
Levi, G., Duband, J.-L. and Thiery, J. P. (1990). Modes of cell migration inthe vertebrate embryo. Int. Rev. Cytol. 123, 201-252.
Mackie, E. J., Tucker, R. P., Halfter, W., Chiquet-Ehrismann, R. andEpperlein, H. H. (1988). The distribution of tenascin coincides withpathways of neural crest cell migration. Development 102, 237-250.
Moyle, M., Napier, M. A. and McLean, J. W. (1991). Cloning and expressionof a divergent integrin subunit β8. J. Biol. Chem. 266, 19650-19658.
Nagano, Y., Hamano, T., Nakashima, N., Ishikawa, M., Miyazaki, K. andHayashi, M. (1992). Yolk vitronectin: Purification and differences from itsblood homologue in molecular size, heparin binding, collagen binding, andbound carbohydrate. J. Biol. Chem. 267, 24863-24870.
Neff, N. T., Lowrey, C., Decker, C., Tovar, A., Damsky, C., Buck, C. and

2702
Horwitz, A. F. (1982). A monoclonal antibody detaches embryonic skeletalmuscle from extracellular matrices. J. Cell Biol. 95, 654-666.
Neugebauer, K. M., Emmet, C. J., Venstrom, K. A. and Reichardt, L. F.(1991). Vitronectin and thrombospondin promote retinal neurite outgrowth:Developmental regulation and role of integrins. Neuron 6, 345-358.
Neugebauer, K. M., Venstrom, K. A. and Reichardt, L. F. (1992). Adhesionof a chick myeloblast cell line to fibrinogen and vitronectin through a β1-class integrin. J. Cell Biol. 116, 809-815.
Newgreen, D. F. and Thiery, J. P. (1980). Fibronectin in early avian embryos:Synthesis and distribution along the migration pathways of neural crest cells.Cell Tiss. Res. 211, 269-291.
Newgreen, D. F. and Erickson, C. A. (1986). The migration of neural crestcells. Int. Rev. Cytol. 103, 89-145.
Newgreen, D. F., Gibbins, I. L., Sauter, J., Wallenfels, B. and Wütz, R.(1982). Ultrastructural and tissue-culture studies on the role of fibronectin,collagen and glycosaminoglycans in the migration of neural crest cells in thefowl embryo. Cell Tiss. Res. 221, 521-549.
Perris, R., Paulsson, M. and Bronner-Fraser, M. (1989). Molecularmechanisms of avian neural crest cell migration on fibronectin and laminin.Develop. Biol. 136, 222-239.
Perris, R., Krotoski, D. and Bronner-Fraser, M. (1991a). Collagens in avianneural crest development: Distribution in vivo and migration-promotingability in vitro. Development 113, 969-984.
Perris, R., Krotoski, D., Lallier, T., Domingo, C., Sorrell, J. M. andBronner-Fraser, M. (1991b). Spatial and temporal changes in thedistribution of proteoglycans during avian neural crest development.Development 111, 583-599.
Perris, R., Kuo, H.-J., Glanville, R. W., Leibold, S. and Bronner-Fraser, M.(1993a). Neural crest cell interaction with type VI collagen is mediated bymultiple cooperative binding sites within triple-helix and globular domains.Exp. Cell Res. 209, 103-117.
Perris, R., Syfrig, J., Paulsson, M. and Bronner-Fraser, M. (1993b).Molecular mechanisms of neural crest cell attachment and migration on typesI and IV collagens. J. Cell Sci. 106, 1357-1368.
Poole, T. J. and Thiery, J. P. (1986). Antibodies and a synthetic peptide thatblock cell-fibronectin adhesion arrest neural crest cell migration in vivo.Prog. Clin. Biol. Res. 217B, 235-238.
Preissner, K. T. (1991). Structure and biological role of vitronectin. Annu. Rev.Cell Biol. 7, 275-310.
Pytela, R., Pierschbacher, M. D. and Ruoslahti, E. (1985). A 125/115-kDacell surface receptor specific for vitronectin interacts with the arginine-glycine-aspartic acid adhesion sequence derived from fibronectin. Proc.Natl. Acad. Sci. USA 82, 5766-5770.
Rovasio, R. A., Delouvée, A., Yamada, K. M., Timpl, R. and Thiery, J. P.(1983). Neural crest cell migration: Requirement for exogenous fibronectinand high cell density. J. Cell Biol. 96, 462-473.
Seftor, R. E. B., Seftor, E. A., Gehlsen, K. R., Stetler-Stevenson, W. G.,Brown, P. D., Ruoslahti, E. and Hendrix, M. J. C. (1991). Role of theαVβ3 integrin in human melanoma cell invasion. Proc. Natl. Acad. Sci. USA89, 1557-1561.
Singer, I. I., Scott, S., Kawka, D. W., Kazazis, D. M., Gailit, J. andRuoslahti, E. (1988). Cell surface distribution of fibronectin and vitronectinreceptors depends on substrate composition and extracellular matrixaccumulation. J. Cell Biol. 106, 2171-2182.
Smith, J. W., Vestal, D. J., Irwin, S. V., Burke, T. A. and Cheresh, D. A.(1990). Purification and functional characterization of integrin αVβ5: Anadhesion receptor for vitronectin. J. Biol. Chem. 265, 11008-11013.
Stockmann, A., Hess, S., Declerck, P., Timpl, R. and Preissner, K. T.(1993). Multimeric vitronectin: Identification and characterization ofconformation-dependent self-association of the adhesive protein. J. Biol.Chem. 268, 22874-22882.
Suzuki, S., Pierschbacher, M. D., Hayman, E. G., Nguyen, K., Öhgren, Y.and Ruoslahti, E. (1984). Domain structure of vitronectin: Alignment ofactive sites. J. Biol. Chem. 259, 15307-15314.
Suzuki, S., Argraves, W. S., Pytela, R., Arai, H., Krusius, T.,Pierschbacher, M. D. and Ruoslahti, E. (1986). cDNA and amino-acidsequences of the cell adhesion protein receptor recognizing vitronectinreveals a transmembrane domain and homologies with other adhesionprotein receptors. Proc. Natl. Acad. Sci. USA 83, 8614-8618.
Tan, S. S., Crossin, K. L., Hoffman, S. and Edelman, G. M. (1987).Asymmetric expression in somites of cytotactin and its proteoglycan ligandis correlated with neural crest cell distribution. Proc. Natl. Acad. Sci. USA 84,7977-7981.
Thiery, J. P., Duband, J.-L. and Delouvée, A. (1982). Pathways andmechanism of avian trunk neural crest cell migration and localization.Develop. Biol. 93, 324-343.
Tucker, G. C., Aoyama, H., Lipinski, M., Tursz, T. and Thiery, J. P. (1984).Identical reactivity of monoclonal antibodies HNK-1 and NC-1:Conservation in vertebrates on cells derived from the neural primordium andon some leukocytes. Cell Diff. 14, 223-230.
Tucker, R. P. and Erickson, C. A. (1984). Morphology and behavior of quailneural crest cells in artificial three-dimensional extracellular matrices.Develop. Biol. 104, 390-405.
Vogel, B. E., Tarone, G., Giancotti, F. G., Gailit, J. and Ruoslahti, E.(1990). A novel fibronectin receptor with an unexpected subunit composition(αvβ1). J. Biol. Chem. 265, 5934-5937.
Wayner, E. A., Orlando, R. A. and Cheresh, D. A. (1991). Integrins αVβ3and αVβ5 contribute to cell attachement to vitronectin but differentiallydistribute on the cell surface. J. Cell Biol. 113, 919-929.
Yatohgo, T., Izumi, M., Kashiwagi, H. and Hayashi, M. (1988). Novelpurification of vitronectin from human plasma by heparin affinitychromatography. Cell Struct. Funct. 13, 281-292.
Zhang, Z., Morla, A. O., Vuori, K., Bauer, J. S., Juliano, R. L. andRuoslahti, E. (1993). The αVβ1 integrin functions as a fibronectin receptorbut does not support fibronectin matrix assembly and cell migration onfibronectin. J. Cell Biol. 122, 235-242.
(Accepted 25 May 1994)
M. Delannet and others






![Effects of the RGD loop and C-terminus of rhodostomin on … · 2017. 5. 3. · RGD loop to regulate integrins recognition [8, 11, 16–22]. For example, Marcinkiewicz et al. reported](https://static.fdocument.org/doc/165x107/611e0fdbc7885320dd5190dc/effects-of-the-rgd-loop-and-c-terminus-of-rhodostomin-on-2017-5-3-rgd-loop.jpg)











