
Small doses of 5α-dihydrotestosterone mimic the effects of 5α-androstane-3β,17β-diol on acid...
7
Molecular and Cellular Endocrinology, 22 (1981) 223-229 Elsevier/North-Holland Scientific Publishers, Ltd. 223 SMALL DOSES OF Sa-DIHYDROTESTOSTERONE MIMIC THE EFFECTS OF Sa-ANDROSTANE-3&17&DIOL ON ACID PHOSPHATASE ACTIVITY IN THE ADULT RAT PROSTATE GLAND M.P. TENNISWOOD *, P.P. ABRAHAMS, C.E. BIRD and A.F. CLARK ** Departments of Biochemistry and Medicine. Queen’s University and Kingston General Hospital, Kingston, Ont. K7L 3iV4 (CanadaJ Received 4 September 1980; accepted 28 January 1981 These studies were designed to further investigate whether 5a-androstane-3p,l7pdiol was exerting unique effects on rat prostate acid phosphatase activity or could possibly be exerting its actions by a small peripheral conversion to 5cudihydrotestosterone. Intraperitoneal adminis- tration of 5odihydrotestosterone in doses of 1 mg, 100 gg or 50 wg per day starting 7 days after castration led to the restoration of normal characteristics of acid phosphatase activity. However, when 5adihydrotestosterone was given in a dose of only 25 ug per day starting 7 days after castration, the changes in acid phosphatase activity were indistinguishable from those found when 5a-androstane-3@,17pdiol was administered in a dose of 2 mg per day. This suggests that the effects of So-androstane-3p,l7Pdiol can be explained by its conversion to small amounts of 5cudihydrotestosterone. Keywords: 5cu-androstane-3@,17P_diol; Sadihydrotestosterone; androgens; prostatic acid phosphatase. Previous studies in our laboratory established that certain characteristics of acid phosphatase (EC 3.1.3.2) activity are good indicators for monitoring androgen effects in the rat prostate (Tenniswood et al., 1976, 1978a). Extension of these studies into the effects of Sa-androstane3/3,17P-diol (3fl-androstanediol) led to results which suggested that this steroid had actions distinctly different from those of the classical androgens, testosterone, 17/3-hydroxy-So-androstan-3-one (SLY- dihydrotestosterone) and Sor-androstane-3a,l7P-diol (3oandrostanediol) (Tennis- wood et al., 1978b). While 3&androstanediol had no effects on prostatic weight of adult rats castrated 7 days previously, it caused the androgen-dependent secretory acid phosphatase to return earlier than did the classical androgens. The acid phos- phatase specific activity and its percentage inhibition by L-tartrate increase follow- ing castration, reaching a maximum level on day 7, following which they decrease to a new steady state by day 15. The intraperitoneal administration of pharmacologic doses of testosterone, Scr-dihydrotestosterone or 3a-androstanediol starting 7 days * Current address: Medical Research Council, National Institute for Medical Research, The Ridgeway, Mill Hill London, NW7 1AA (Great Britain) ** To whom correspondence should be addressed. 0303-7207/81/0000-0000/$02.50 0 Elsevier/North-Holland Biomedical Press
Transcript of Small doses of 5α-dihydrotestosterone mimic the effects of 5α-androstane-3β,17β-diol on acid...

Molecular and Cellular Endocrinology, 22 (1981) 223-229 Elsevier/North-Holland Scientific Publishers, Ltd.
223
SMALL DOSES OF Sa-DIHYDROTESTOSTERONE MIMIC THE EFFECTS OF
Sa-ANDROSTANE-3&17&DIOL ON ACID PHOSPHATASE ACTIVITY IN THE
ADULT RAT PROSTATE GLAND
M.P. TENNISWOOD *, P.P. ABRAHAMS, C.E. BIRD and A.F. CLARK ** Departments of Biochemistry and Medicine. Queen’s University and Kingston General Hospital, Kingston, Ont. K7L 3iV4 (CanadaJ
Received 4 September 1980; accepted 28 January 1981
These studies were designed to further investigate whether 5a-androstane-3p,l7pdiol was exerting unique effects on rat prostate acid phosphatase activity or could possibly be exerting its actions by a small peripheral conversion to 5cudihydrotestosterone. Intraperitoneal adminis- tration of 5odihydrotestosterone in doses of 1 mg, 100 gg or 50 wg per day starting 7 days after castration led to the restoration of normal characteristics of acid phosphatase activity. However, when 5adihydrotestosterone was given in a dose of only 25 ug per day starting 7 days after castration, the changes in acid phosphatase activity were indistinguishable from those found when 5a-androstane-3@,17pdiol was administered in a dose of 2 mg per day. This suggests that the effects of So-androstane-3p,l7Pdiol can be explained by its conversion to small amounts of 5cudihydrotestosterone.
Keywords: 5cu-androstane-3@,17P_diol; Sadihydrotestosterone; androgens; prostatic acid phosphatase.
Previous studies in our laboratory established that certain characteristics of acid phosphatase (EC 3.1.3.2) activity are good indicators for monitoring androgen effects in the rat prostate (Tenniswood et al., 1976, 1978a). Extension of these studies into the effects of Sa-androstane3/3,17P-diol (3fl-androstanediol) led to results which suggested that this steroid had actions distinctly different from those of the classical androgens, testosterone, 17/3-hydroxy-So-androstan-3-one (SLY- dihydrotestosterone) and Sor-androstane-3a,l7P-diol (3oandrostanediol) (Tennis- wood et al., 1978b). While 3&androstanediol had no effects on prostatic weight of adult rats castrated 7 days previously, it caused the androgen-dependent secretory acid phosphatase to return earlier than did the classical androgens. The acid phos- phatase specific activity and its percentage inhibition by L-tartrate increase follow-
ing castration, reaching a maximum level on day 7, following which they decrease to a new steady state by day 15. The intraperitoneal administration of pharmacologic doses of testosterone, Scr-dihydrotestosterone or 3a-androstanediol starting 7 days
* Current address: Medical Research Council, National Institute for Medical Research, The Ridgeway, Mill Hill London, NW7 1AA (Great Britain) ** To whom correspondence should be addressed.
0303-7207/81/0000-0000/$02.50 0 Elsevier/North-Holland Biomedical Press

224 M.P. Tenniswood, P.P. Abmhams, C.E. Bird, A.F. Clark
after castration cause both parameters to return to normal. When 3fl-androstanediol was administered, both parameters remained elevated.
Baulieu (1975) had suggested that at the level of the target tissue, testosterone is transformed to metabolites which have different complementary activities and which together explain the overall actions of testosterone. Thus it was proposed that prostate proliferation is primarily controlled by 5cY-dihydrotestosterone while features such as maintenance of epithelial height and the stimulation of prostatic secretion are governed by 3/I-androstanediol (Lasnitzki, 1970; Baulieu et al., 1975). However the possibility still exists that 3fi-androstanediol might exert its actions through prior conversion to 5a-dihydrotestosterone and its effects might be due solely to small amounts of the latter. While this possibility is quite remote in tissue culture since prostatic 3fl-hydroxysteroid dehydrogenase is primarily directed toward synthesis of 3P-androstanediol (Robe1 et al., 1971) and may be irreversible (Levy et al., 1974) there is little doubt that 5cy-dihydrotestosterone can be pro- duced from 3fl-androstanediol by the testis (Wiebe, 1976) liver (Lisboa and Gustafsson, 1970) and erythrocytes (Heyns and De Moor, 1974). Furthermore when [3H]3@androstanediol was infused into either intact or castrated male rats small amounts of [3 H] 50c-dihydrotestosterone accumulated in the prostate (Tennis- wood et al., 1980).
This paper reports results obtained with a series of 5ol-dihydrotestosterone doses to determine if small doses would mimic the effects of 3fl-androstanediol on rat prostatic acid phosphatase characteristics. The doses ranged from ones which would stimulate to those which were well below that needed to initiate growth of the prostate in castrated rats (Lesser and Bruchovsky, 1973). 3&Androstanediol did not initiate prostate growth in our initial studies (Tenniswood et al., 1978b). The acid phosphatase characteristics examined were polyacrylamide gel electrophoretic pat- tern, extent of inhibition by L-tartrate and specific activity.
MATERIALS AND METHODS
Steroids 3fl-Androstanediol dipropionate and Sol-dihydrotestosterone propionate were ob-
tained from Steraloids (Wilton, NH, U.S.A.). The steroids were pure as judged by thin-layer chromatography on silica gel G and detection with iodine vapors. Injec- tions were given intraperitoneally starting 7 days after castration. In all cases the dose was given in 0.2 ml of sesame oil.
All the procedures used have been detailed by Tenniswood et al. (1978a). Enzyme assays were performed using p-nitrophenylphosphate as substrate. To determine the per cent inhibition L-tartrate was added to a final assay concentra- tion of 14 mM in parallel incubations. Polyacrylamide gel electrophoresis was per- formed using 7% acrylamide gels which were run in /3-alanine-acetic acid buffer, pH 4.5 for 3 h at 4 mA/gel at 4°C. The enzyme activity was localized on the gels using

a-naphthylphosphate as the substrate and Fast Garnet GBC salt as the coupling reagent. Protein assays were performed using the Miller modification (1959) of the method of Lowry et al. (1951 j.
The organ wei~t~body weight ratios were calculated as previously described (Jackson et al., 1977) using the equation:
oW,BW = organ weight in milligrams
body weight in grams
Experiments
To determine whether the response of prostatic acid phosphatase to 3&andro- stanediol was a dosage phenomenon of Sa-dihydrotestosterone or a unique response
to 3P-androstanediol 240 rats were randomly divided into 5 groups and castrated. 7 days later replacement therapy was started with one of the following steroid dosages: 3&androstanediol dipropionate (2 mgfdaylratj or So-dihydrotestosterone prop~onate (dosages of 1 mg, 100 pg, SO yg and 2.5 ~~/day/rat). Rats were killed on selected days over the next 14 days.
RESULTS
Administration of 5~.d~ydrotestosterone to castrated rats, starting 7 days after castration, affected the prostatic weight in a dose-dependent manner as shown in Fig. 1A. On day 7 the prostate was small with an OW/BW ratio of 0.18 i: 0.03; on treatment with 5o-dihydrotestosterone at a dosage of 1 mg/day the OW/BW ratio increased, until by day 17 the ratio was 0.85 + 0.04 which is near the normal value seen for intact rats. So-~hydrotestosterone at dosages of 100, 50 and 25 pg/day did not stimulate prostate growth, and the OW/BW ratio remained consistently low at 0.20 t 0.05 for the length of the experiment. 3&Androstanediol at a dosage of 2 mglday did not cause any change in the OW/BW ratio.
Fig. 1B shows the changes in the specific activity of prostatic acid phosphatase after administration of the different dosages of So-d~ydrotestosterone and 3P-androstanediol. Administration of %dihydrotestosterone caused a dose-depen- dent decrease in the specific activity. The dosage of 1 trig/day Scr-dihydrotestoster- one caused the specific activity to decrease from the level of 9.50 + 0.80 pmoles/h/ mg protein seen on day 7 to 4.86 + 1.60 fimoles/h/mg protein. The 100 and 50 pg/ day dosages caused a decrease of the specific activity to 6.86 ? 0.48 and to 7.59 + 0.27 ~molesjh/mg protein on day 17 respectively. The lowest dosage of Sa-dihy- drotestosterone (25 @g/day) administered did not cause a decrease in the specific activity of prostatic acid phosphatase and it remained elevated at 10.20 _C 0.31 pmoles/h/mg protein. This result was not significantly different from that seen when 3~-androst~ediol is administered at a dosage of 2 mglday, where the specific activity of acid phosphatase remains eievated at IO.50 rt 0.39 ~molesihjmg protein
on day 17.
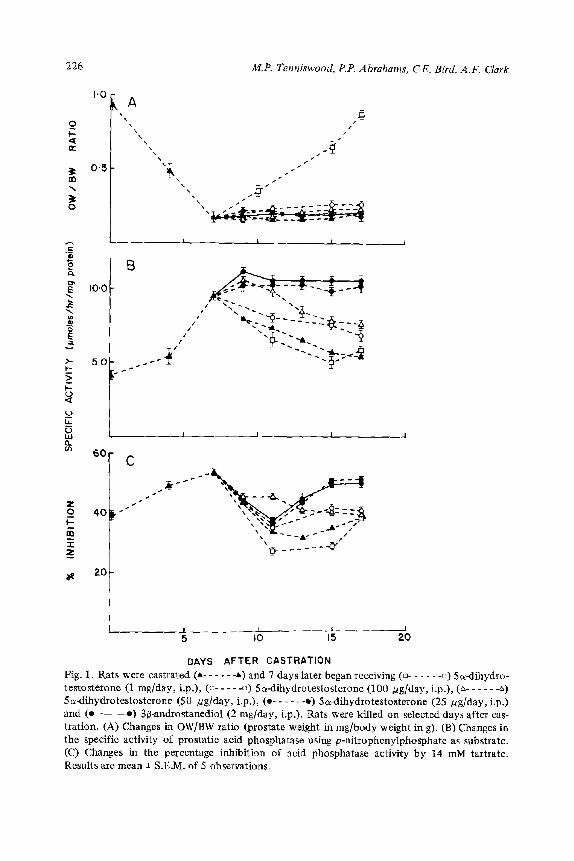
226 M.P. Tenniswood, P.P. Abrahams, CE. Bird, A.F. Clark
I I 1 * I
5 10 15 20
DAYS AFTER CASTRATION
Fig. 1. Rats were castrated (A- - - - - 4) and 7 days later began receiving (n- - - - - -0) Socdihydro- testosterone (1 mglday, i.p.), (o- - - - -0) Soldihydrotestosterone (100 ug/day, i.p.), (A- - - - - -A) Soldihydrotestosterone (SO ug/day, i.p.), (o- - - - - -0) 5ordihydrotestosterone (25 fig/day, i.p.) and (0~) 3p-androstanediol (2 mg/day, i.p.). Rats were killed on selected days after cas-
tration. (A) Changes in OW/BW ratio (prostate weight in mg/body weight in g). (B) Changes in the specific activity of prostatic acid phosphatase using p-nitrophenylphosphate as substrate. (C) Changes in the percentage inhibition of acid phosphatase activity by 14 mM tartrate.
Results are mean + S.E.M. of 5 observations.

Androgenic control of rat prostatic acid phosphate 221
For normal rats there are two bands of acid phosphatase activity on polyacryl- amide gels - the lysosomal and secretory bands with the latter being androgen- dependent in that it disappears within 4 days following castration. When castrated rats were treated with the various dosages of So-dihydrotestosterone starting on
day 7 the secretory acid phosphatase was restored on polyacrylamide gel electro- phoresis. The high dose (1 mg/day) of Sa-dihydrotestosterone restored the secre- tory acid phosphatase on day 17, while the 100 pg/day dose restored the secretory acid phosphatase on day 15. The secretory enzyme reappeared on day 11 when 2.5 pg/day or 50 pg/day of Sol-dihydrotestosterone is administered. 3/3-Androstanediol restored the secretory acid phosphatase on day 13. Thus all the dosages of Sol-dihy- drotestosterone were capable of restoring acid phosphatase as seen on gels, although the smaller dosages were more effective in achieving a rapid return of secretory acid phosphatase.
Fig. 1C shows that following the administration of Sa-dihydrotestosterone at a dosage of 1 mg/day there was a drop in the per cent inhibition of acid phosphatase activity by L-tartrate from a level of 53.5 -+ 1 .O% on day 7 to a level of 27.0 + 1 .O% on day Il. The per cent inhibition then increased to a level of 37.5 ir 1.2% by day 17. The administration of 5cr-dihydrotestosterone at dosages of 100 pg/day or 50 pg/day resulted in a decrease before the per cent inhibition levels off at a plateau of 40.5 + 1 .O% by day 17. Thus these 3 doses of Sa-dihydrosterone when administered starting 7 days after castration resulted in a per cent inhibition which is not signifi- cantly different from values for intact male rats. Administration of Sa-dihydrotest- osterone at a dosage of 25 pg/day, one the other hand, resulted in a decrease from 53.5 + 1 .O% to 35.5 t 1.5% on day 11. The per cent inhibition then rose to a level
of 50.5 + 1 .O% between days 15 and 17.3/3-Androstanediol administration resulted in a decrease in the per cent inhibition to 35.5 1 1 .l% on day 11, followed by a rise between days 11 and 15 to a level of 50.1 _+ 1 .I % which is maintained on day 17.
DISCUSSION
Sa-Dihydrotestosterone at a low dosage (25 pg/day) is able to restore the secre- tory acid phosphatase by day 11 on polyacrylamide gels, although the specific activity is elevated at 10.20 + 0.8 pmoles/h/mg protein on day 11, and the per cent inhibition by tartrate rises to 50.5 + 1 .O% on day 15. These results are indistin- guishable from those found when 3P-androstanediol is administered at a dosage of 2 mg/day. Thus administration of Sa-dihydrotestosterone at a dosage of 25 pg/day mimics the effects of 3/3-androstanediol at a dosage of 2 mg/day on prostatic acid phosphatase. We previously suggested that 3P-androstanediol has a rapid and unique effect on rat prostatic acid phosphatase (Tenniswood et al., 1978b), and others have suggested that 3P-androstanediol may control the secretory processes of the rat prostate (Lasnitzki, 1970). However, in view of the present results we now sug-

228 M.P. Tenniswood, P.P. Abrahams, C.E. Bird, A.F. Clark
gest that the action of 3P-androstanediol is more probably exerted through periph- eral conversion to 5ol-dihydrotestosterone. Support for this suggestion comes from the finding that [3H]So.-dihydrotesto~terone is accumulated in the rat prostate in small amounts after both contant infusion of [3H]3/3-androstanedio1 (Tenniswood
et al., 1980) and single injection of the same steroid (Krieg et al., 1975). In the studies from our laboratory (Tenniswood et al., 1980) the levels of [3H]S~-dihy- drotestosterone in the prostate (906 A 204 dpm/g for normal animals vs. 6025 dpm/g for animals 24 h after castration) are higher than those in plasma (421 + 116 dpm/ml and 14 19 + 3 18 dpm/ml, respectively) 3 h after initiation of constant infu- sion of [3H]3/3-androstanedio1; the corresponding figures for [3H]3/3-androstanedio1 in the prostate were 3278 2 468 dpm/mg for normal animals and 3176 it 959 dpm/g 24 h after castration and in the plasma were 13 666 2 850 dpm/ml and 16 432 +_ 1138 dpm/ml, respectively. While these studies clearly established the fact that small amounts of %-dihydrotestosterone were formed from 3/3-androstanediol infused into the tail vein, one cannot utilize the exact conversion rates for the studies in this paper as 3P-androstanediol was administered as the dipropionate intraperitoneally and the kinetic studies were performed only 24 h after castration and not at later time intervals. Other workers have examined the response of the secretory acid phosphatase on polyacrylamide gels to different dosages of So-dihy- drotestosterone (Rennie et al., 1978). H owever since their lowest dosage of 5ol-dihydrotestosterone was 100 pg/day, they were unable to see any abnormal effects on prostatic acid phosphatase; they did observe the more rapid return of the secretory enzyme on polyacrylamide gel electrophoresis when low dosages of 5a-dihydrotestosterone were administered.
Although 3flandrostanediol and Sa-dihydrotestosterone at dosages of 2 mg/day and 25 pg/day respectively produce marked changes in the characteristics of pros-
tatic acid phosphatase neither of the steroids produces any alteration in the OW/BW ratio at these dosages. However the changes in the enzyme characteristics are not solely due to the inability of these 2 steroid dosages to cause growth of the prostate since neither the 50 pg/day nor the 100 pg/day dosages of 5ol-dihydrotestosterone produced any change in the OW/BW ratio even though they do lead to normal char- acteristics of acid phosphatase activity.
This suggests that a second effect is also present. It appears that if the dosage of 5a-dihydrotestosterone is sufficient to cause proliferation, the prostate grows and later differentiates. This accounts for the delayed appearance of secretory acid phosphatase in rats treated with higher doses of Sa-dihydrotestosterone (Tennis- wood et al., 1978a; Rennie et al., 1978). On the other hand if the dosage of 5cu-dihydrotestosterone is not sufficient to cause growth, then androgen-stimulated prostatic function occurs immediately. Thus the secretory acid phosphatase reap- pears very rapidly in rats treated with 100 or 50 pg/day of 5a-dihydrotestosterone. Only when Sol-dihydrotestosterone falls to very low levels (as in the rats treated with 25 pg/day Sol-dihydrotestosterone or 2 mg/day 3/3-androstanediol) is the prostate unable to maintain all the androgen-dependent processes, and as a result

Androgenic control of rat prostatic acid phosphate 229
the abnormal characteristics of acid phosphatase (as manifest by the per cent inhi- bition by tartrate) appear. We have previously suggested that the changes in these characteristics, in particular the per cent inhibition by tartrate, could be due to changes in the glycosylation of the acid phosphatase protein (Tenniswood et al., 1976). Thus the increase in the per cent inhibition by tartrate may be due to the altered function of an androgen-dependent glycosylating enzyme, which is unable to function at very low androgen levels.
ACKNOWLEDGEMENTS
This research was supported by grants from the National Cancer Institute of Canada and the Medical Research Council of Canada (MT-2338 and MT-18.52).
REFERENCES
Baulieu, E.-E. (1975) Mol. Cell. Endocrinol. 7,157-174.
Baulieu, E.-E., Le Goascogne, C., Groyer, A., FeyelCabanes, T., and Robe], P. (1975) Vitamins
Hormones 33, l-38.
Heyns, W., and De Moor, P. (1974) Biochim. Biophys. Acta 358,1-13.
Jackson, A.C., Tenniswood, M., Bird, C.E., and Clark, A.F. (1977) Invest. Urol. 14, 351-355.
Krieg, M., Horst, H.J., and Sterba, M.-L. (1975) J. Endocrinol. 64,529-538.
Lasnitzki, I. (1970) in: Some Aspects of the Aetiology and Biochemistry of Prostatic Cancer,
(Eds. K. Griffiths and C.G. Pierrepoint) Third Tenovus Workshop, Alpha, Omega, Alpha,
Cardiff, pp. 68-73.
Lesser, B., and Bruchovsky, N. (1973) Biochim. Biophys. Acta 308,426-437.
Levy, C., Marchut, M., Baulieu, E.-E., and Robe& P. (1974) Steroids 23,291-300.
Lisboa, B.P., and Gustafsson, J.-A. (1970) Eur. J. Biochem. 14,556-559.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., and Randall, F.J. (1951) J. Biol. Chem. 193,265- 275.
Miller, G.L. (1959) Anal. Chem. 31,94.
Rennie, P.S., Bruchovsky, N., and Hook, S.L. (1978) J. Steroid Biochem. 9,585-593.
Robel, P., Lasnitzki, I., and Baulieu, E.-E. (1971) Biochimie 53,81-97.
Tenniswood, M., Bird, C.E., and Clark, A.F. (1976) Can. J. Biochem. 54. 350-357.
Tenniswood, M., Abrahams, P., Bird, C.E., and Clark, A.F. (1978a) J. Endocrinol. 77, 301- 308.
Tenniswood, M., Abrahams, P., Bird, C.E., and Clark, A.F. (1978b) J. Endocrinol. 79,9-16. Tenniswood, M., Bird, C.E., and Clark, A.F. (1980) J. Steroid Biochem., in press,
Wiebe, J.P. (1976) Endocrinology 98,505-513.











![ARegenerativeAntioxidantProtocolof VitaminEand α ...downloads.hindawi.com/journals/ecam/2011/120801.pdf · plications [2–4]. Rats fed a high fructose diet mimic the progression](https://static.fdocument.org/doc/165x107/5f0acf087e708231d42d71f7/aregenerativeantioxidantprotocolof-vitamineand-plications-2a4-rats-fed.jpg)





![[FeFe]‐Hydrogenase Mimic Employing κ2‐C,N‐Pyridine ... · DOI: 10.1002/ejic.201900405 Full Paper Proton Reduction Catalysts [FeFe]-Hydrogenase Mimic Employing κ2-C,N-Pyridine](https://static.fdocument.org/doc/165x107/60cf254691c2d1101b09b0e4/fefeahydrogenase-mimic-employing-2acnapyridine-doi-101002ejic201900405.jpg)
