
SAR and ISR - Iowa State Universityswhitham/WWW/plp692/1017_sar_ISR...PR (Pathogenesis-Related)...
81
SAR and ISR
Transcript of SAR and ISR - Iowa State Universityswhitham/WWW/plp692/1017_sar_ISR...PR (Pathogenesis-Related)...

SAR and ISR
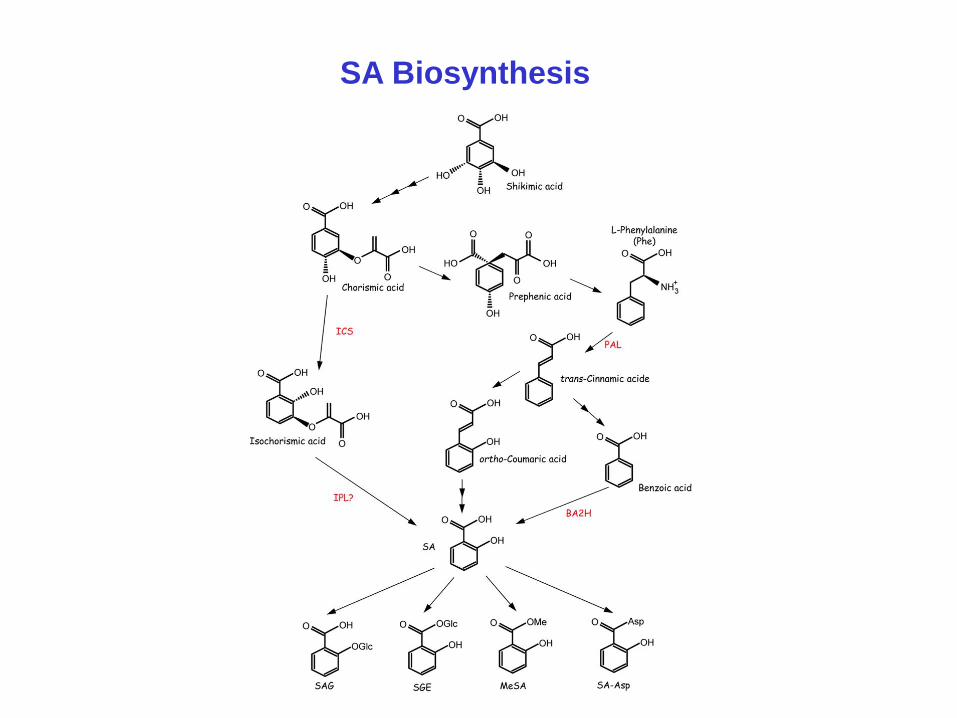
SA Biosynthesis
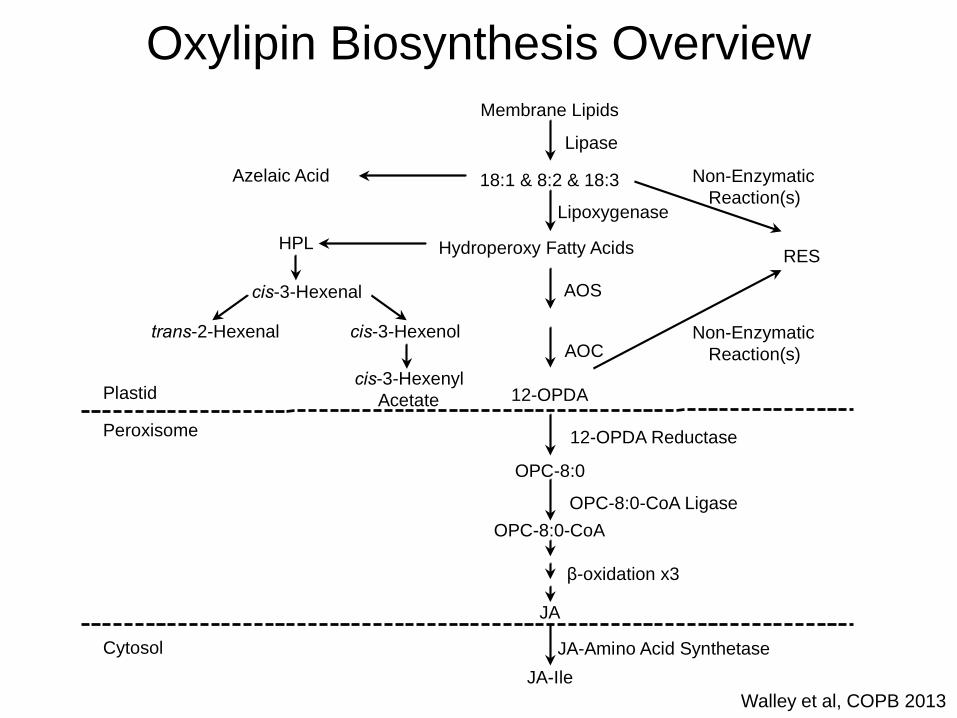
Walley et al, COPB 2013
18:1 & 8:2 & 18:3
Membrane Lipids
Lipase
Lipoxygenase
Hydroperoxy Fatty Acids
AOS
AOC
12-OPDA
12-OPDA Reductase
OPC-8:0
OPC-8:0-CoA Ligase
OPC-8:0-CoA
β-oxidation x3
JA
JA-Amino Acid Synthetase
JA-Ile
Azelaic Acid
RES
Non-Enzymatic
Reaction(s)
Non-Enzymatic
Reaction(s)
Plastid
Peroxisome
Cytosol
HPL
cis-3-Hexenal
cis-3-Hexenol
cis-3-Hexenyl
Acetate
trans-2-Hexenal
Oxylipin Biosynthesis Overview
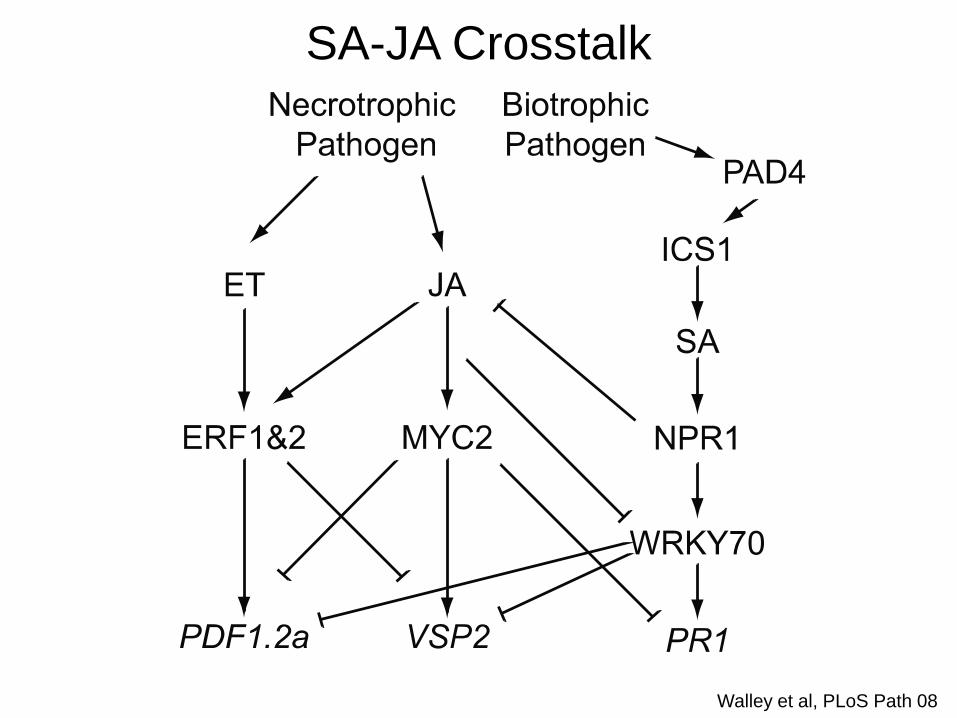
Walley et al, PLoS Path 08
SA-JA Crosstalk

Systemic acquired resistance (SAR)
Background on SAR
Molecular changes associated with SAR
Role of SA and other signaling molecules
Role of NPR1 (non-expresser of pathogenesis related genes 1)
Roles of TGA and WRKY transcription factors
Role of chromatin modification in SAR
What do we know about SAR?
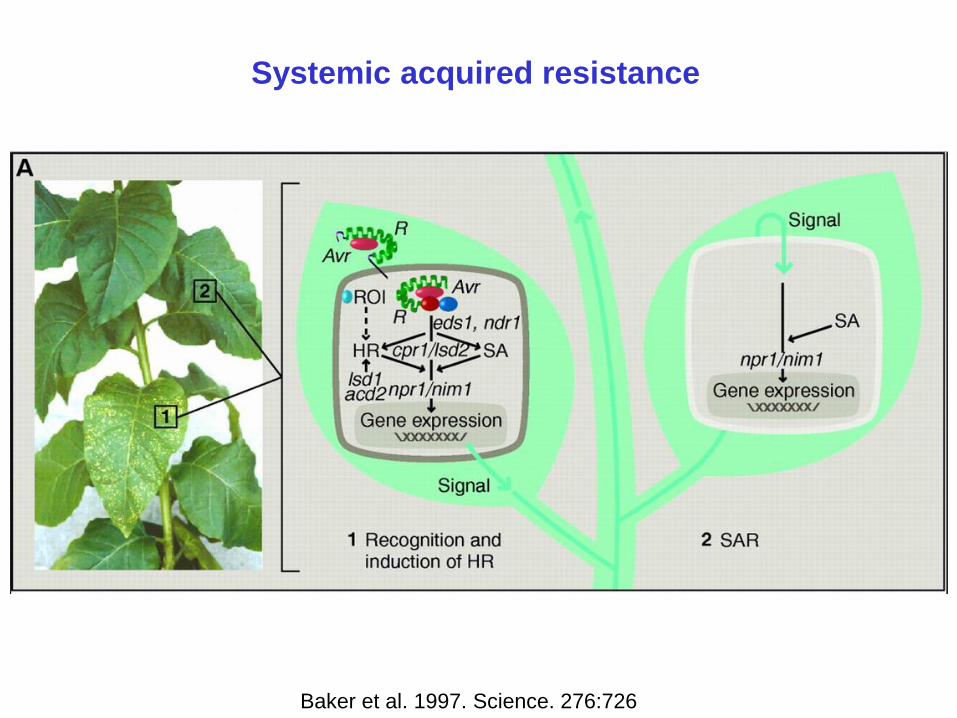
Systemic acquired resistance
Baker et al. 1997. Science. 276:726

Systemic acquired resistance
Systemic acquired resistance was first recognized as a significant phenomenon in the early 1900s.
At that time, it was recognized that infection of plants with “necrotizing” pathogens (causing HR)
often results in enhanced resistance to subsequent infections by a variety of fungal, bacterial and
viral pathogens. This physiological immunity was termed systemic acquired resistance (SAR).
SAR confers a broad spectrum type of resistance
SAR is effective against some but not all pathogens:
Tobacco: Phytophthora parasitica, Cercospora nicotianae, Peronospora tabacina
Tobacco mosaic virus, Tobacco necrosis virus,
Pseudomonas syringae pv. tabaci, Erwinia carotovora
Not effective against: Botrytis cinerea or Alternaria alternata
Arabidopsis: Phytophthora parasitica
Turnip crinkle virus
Pseudomonas syringae pv. tomato DC3000

Systemic acquired resistance was associated with the coordinated induction of a set of SAR genes
encoding proteins known as Pathogenesis-related (PR) proteins (Van Loon and Gianinazzi (early 1970s).
(1979) White found that acetyl salicylic acid application sufficient to induce PR gene expression
and enhanced resistance to tobacco mosaic virus in tobacco plant. Discovery came out the
interest in developing chemical control methods for viral infection. After that several groups went
on to show that salicylic acid application on tobacco leaves mimics pathogen induced expression
of PR genes and pathogen resistance in treated tissues.
(1990) Two groups one led by Klessig and Raskin and another led by Metraux
found that salicylic acid accumulates in cucumber and tobacco plants prior to pathogen infection, but
before the onset of resistance.
The work by these and many others led to the hypothesis that salicylic acid (SA) is the endogenous
signal molecule that is required for the induction of systemic acquired resistance.
(1993/1994) The group headed by Ryals made tobacco plants that could not accumulate SA and found
that these plants were defective in their ability to develop systemic acquired resistance.
This work demonstrated a central role for SA in establishing systemic acquired resistance.
The group also demonstrated that these tobacco plants were defective in their ability to
accumulate PR proteins.
(1997) Cloning of NPR1, a key regulator of SAR
Some key events in understanding regulation of SAR

PR (Pathogenesis-Related) proteins
Proteins secreted predominantly into intercellular
spaces in response to wounding or infection.
Soluble at pH 3
Basic homologs also found (in vacuole).
Proteinase resistant (but not proteinase inhibitors).
Some are developmentally expressed as part of
normal plant development in absence of wound or
infection (e.g. flowering).
PR proteins first identified as major proteins induced by necrotizing pathogens
(pathogens that induced the hypersensitive response)
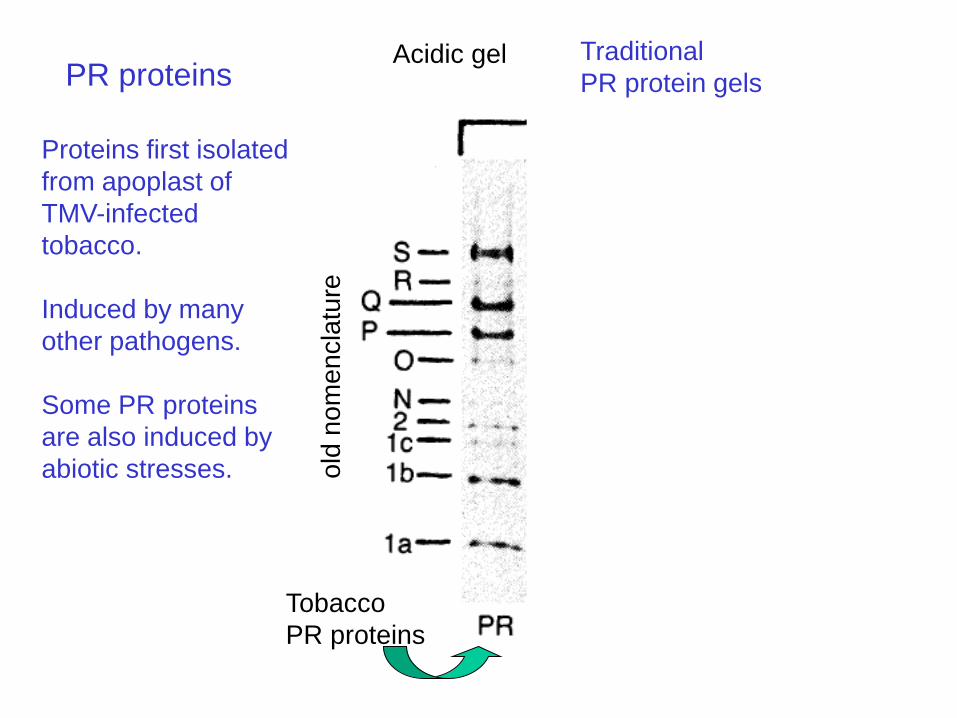
Acidic gel Basic gel
Tobacco
PR proteins
PR proteins
Proteins first isolated
from apoplast of
TMV-infected
tobacco.
Induced by many
other pathogens.
Some PR proteins
are also induced by
abiotic stresses.
Traditional
PR protein gels
old
no
me
ncla
ture
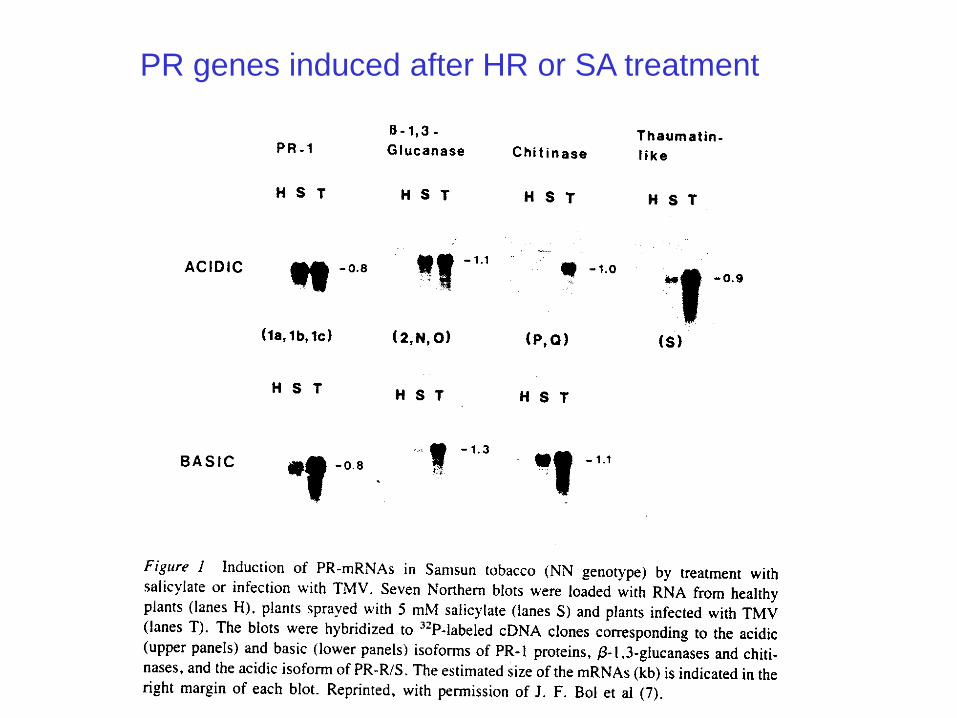
PR genes induced after HR or SA treatment
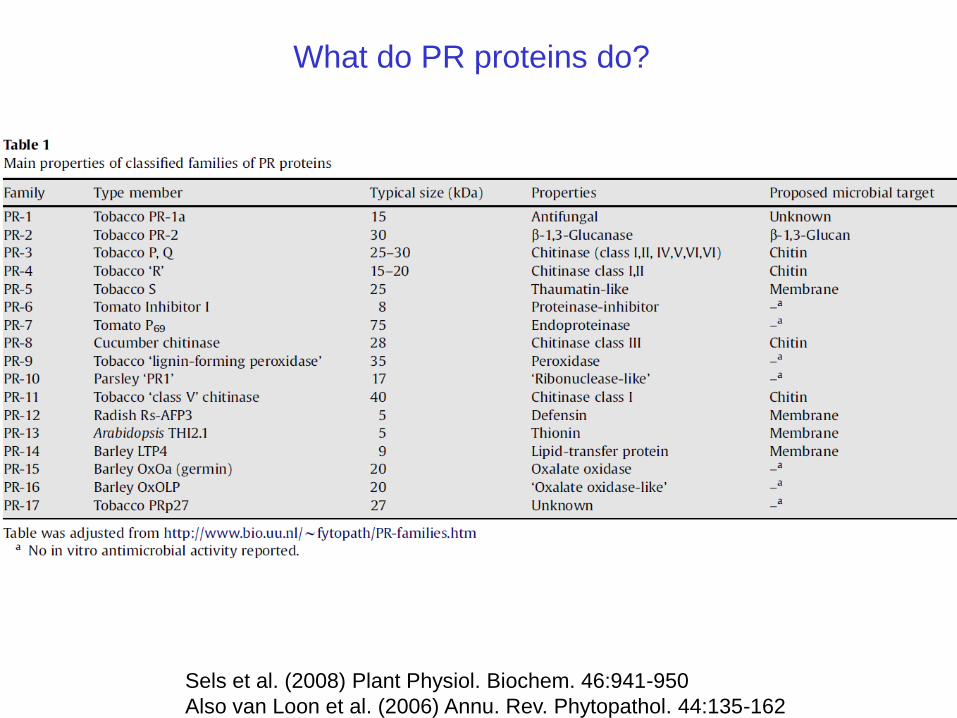
What do PR proteins do?
Sels et al. (2008) Plant Physiol. Biochem. 46:941-950
Also van Loon et al. (2006) Annu. Rev. Phytopathol. 44:135-162
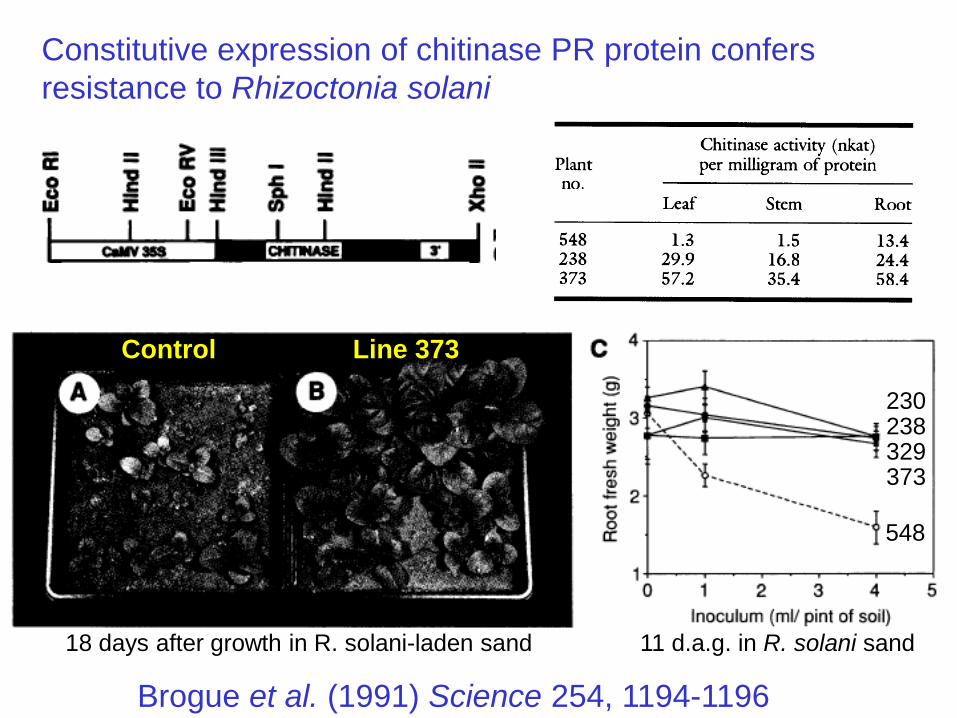
Constitutive expression of chitinase PR protein confers
resistance to Rhizoctonia solani
Brogue et al. (1991) Science 254, 1194-1196
Control Line 373
230238329373
548
18 days after growth in R. solani-laden sand 11 d.a.g. in R. solani sand

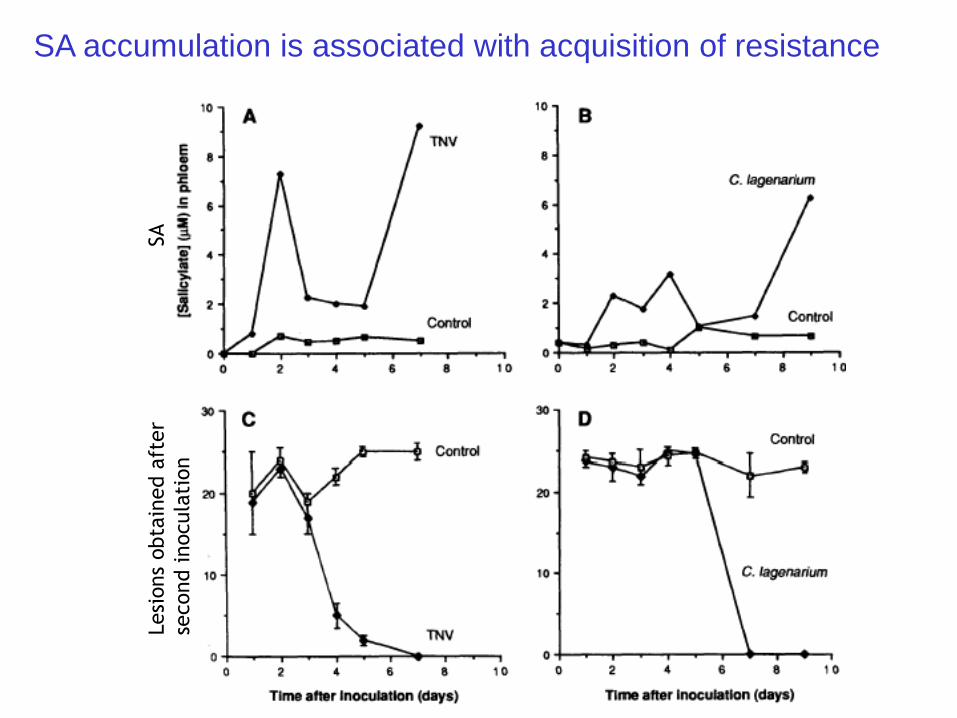
Application of salicylic acid mimics SAR
(1979) White found that the application of aspirin, salicylic acid, and benzoic acid
resulted in enhanced resistance to TMV. Used 3 tobacco cultivars that contained
the N resistance gene that confers HR to TMV. Found > 90% reduction in lesion
number in treated leaves versus water control.
Lesi
ons
obta
ined a
fter
second inocula
tion
SA
SA accumulation is associated with acquisition of resistance

Science (1993) 261, 754-756.
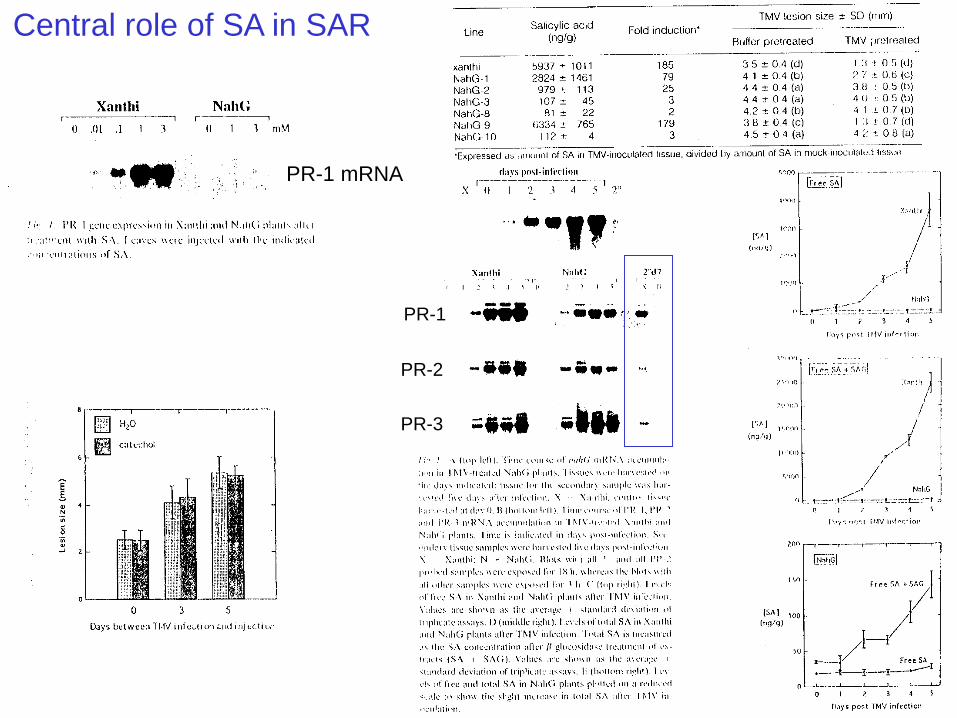
Central role of SA in SAR
PR-1
PR-2
PR-3
PR-1 mRNA
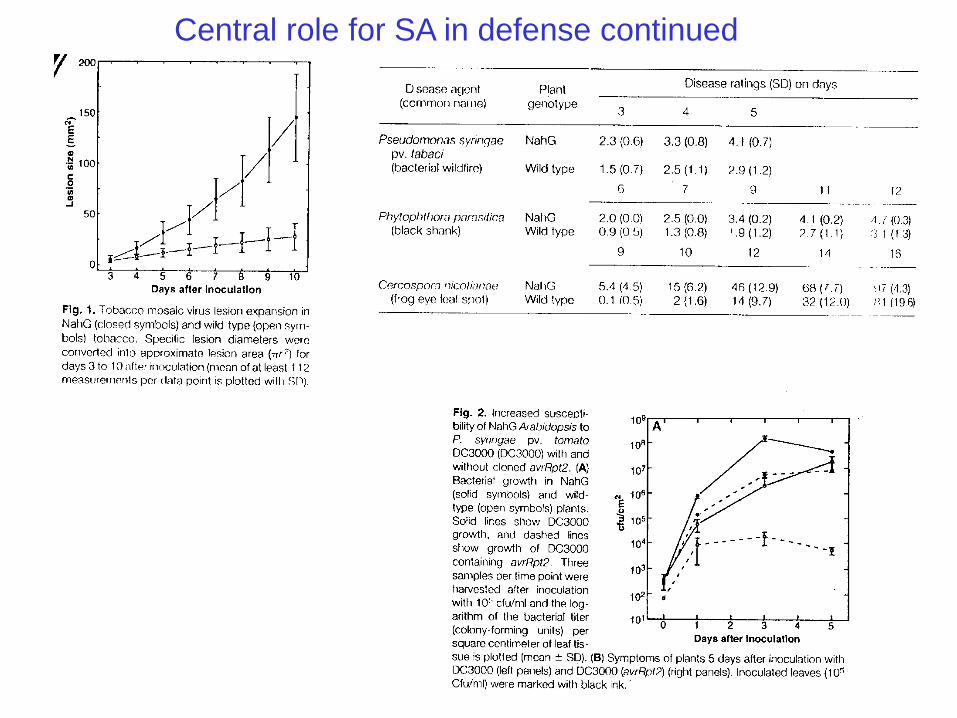
Central role for SA in defense continued
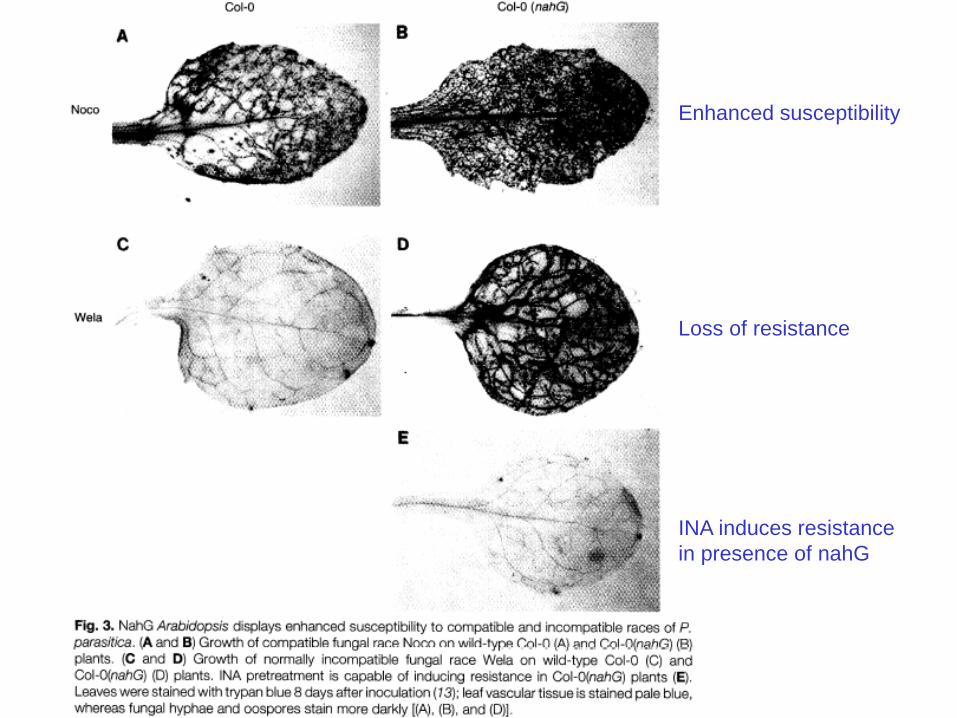
Enhanced susceptibility
Loss of resistance
INA induces resistance
in presence of nahG
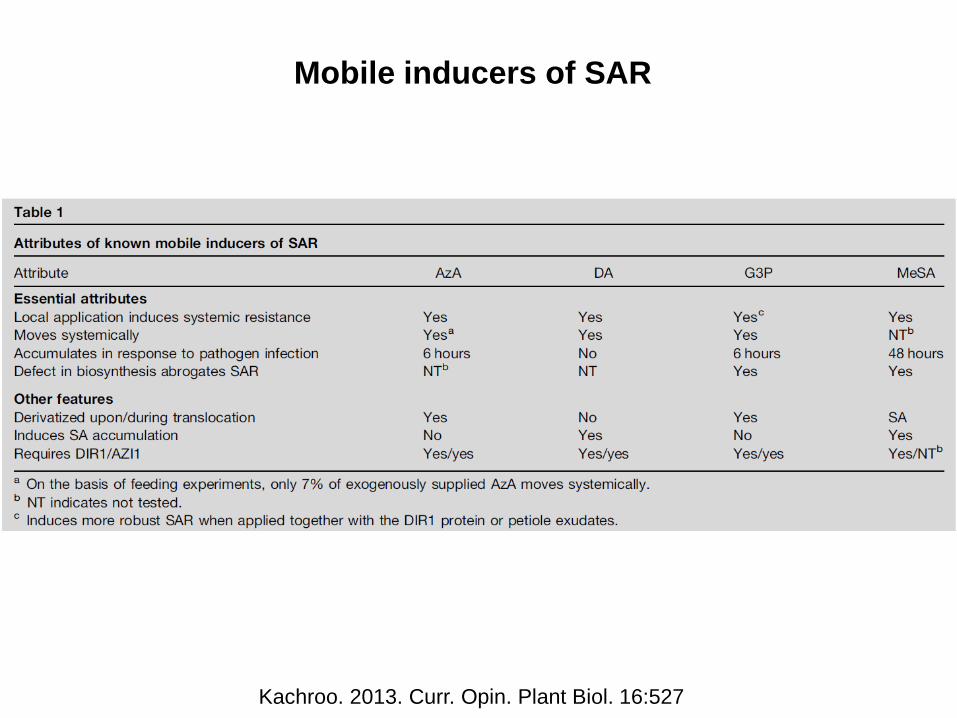
Mobile inducers of SAR
Kachroo. 2013. Curr. Opin. Plant Biol. 16:527
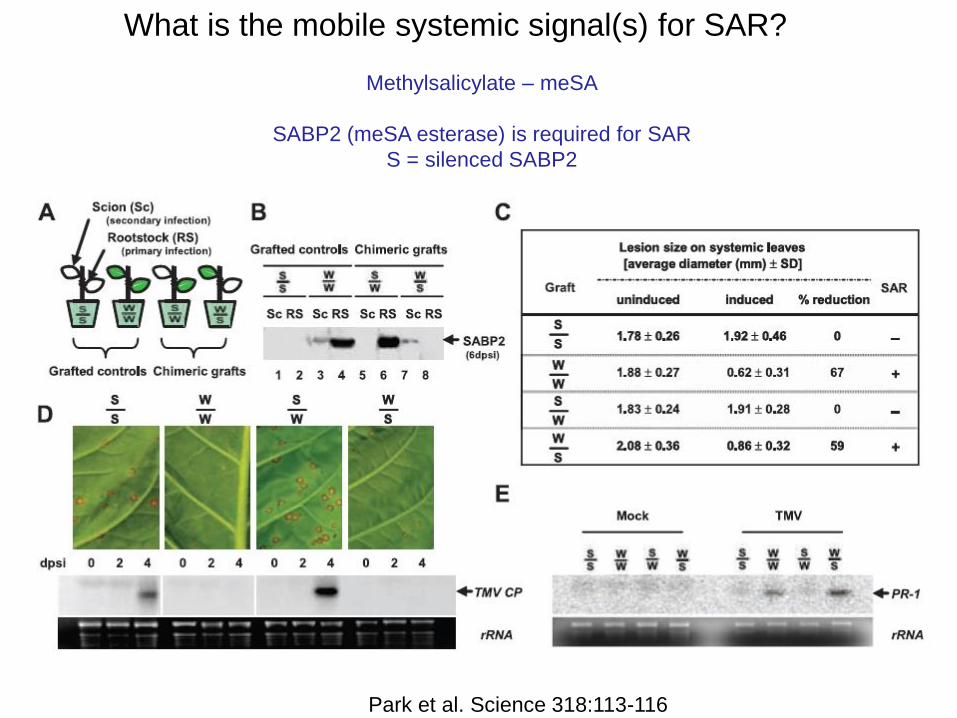
What is the mobile systemic signal(s) for SAR?
Methylsalicylate – meSA
SABP2 (meSA esterase) is required for SAR
S = silenced SABP2
Park et al. Science 318:113-116

What is the mobile systemic signal(s) for SAR?
Methylsalicylate – meSA
SAMT1 (SA methyltransferase) is required for SAR
Park et al. Science 318:113-116
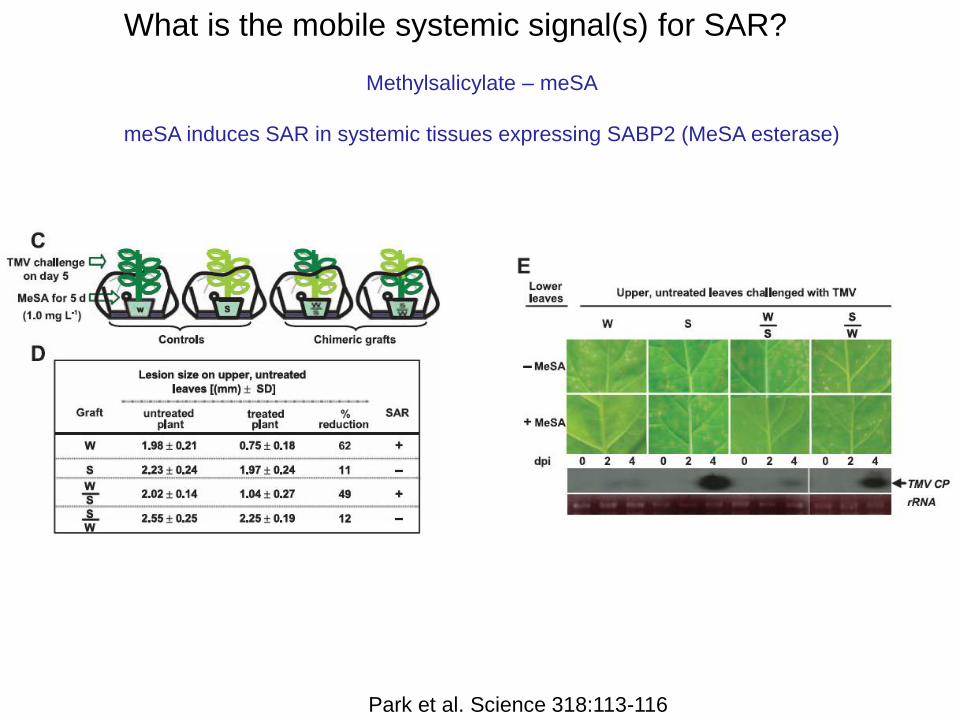
What is the mobile systemic signal(s) for SAR?
Methylsalicylate – meSA
meSA induces SAR in systemic tissues expressing SABP2 (MeSA esterase)
Park et al. Science 318:113-116
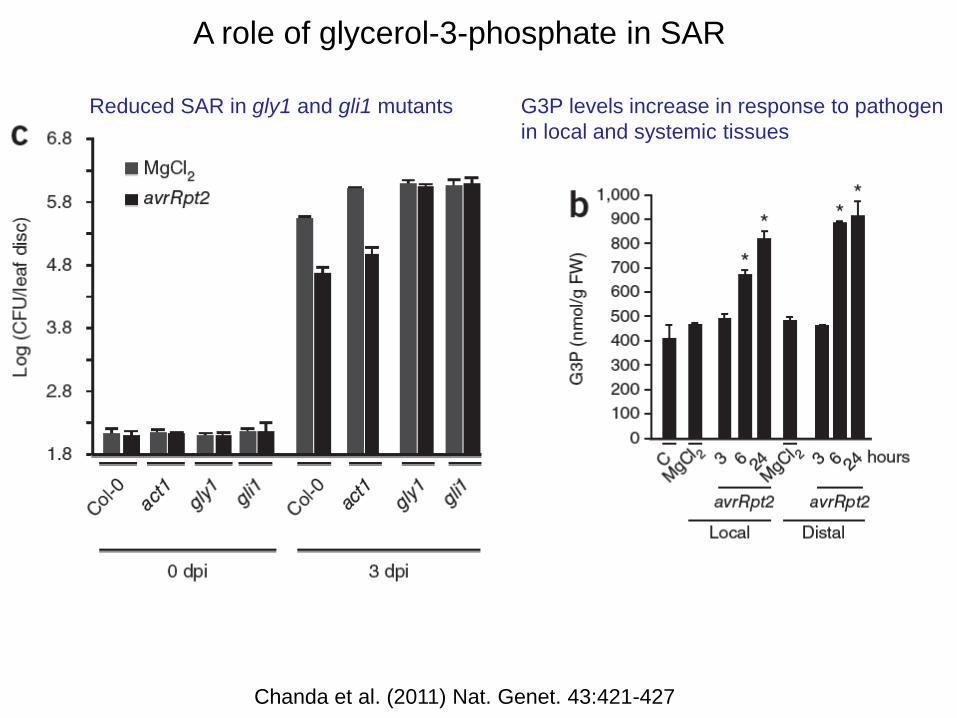
A role of glycerol-3-phosphate in SAR
Reduced SAR in gly1 and gli1 mutants G3P levels increase in response to pathogen
in local and systemic tissues
Chanda et al. (2011) Nat. Genet. 43:421-427
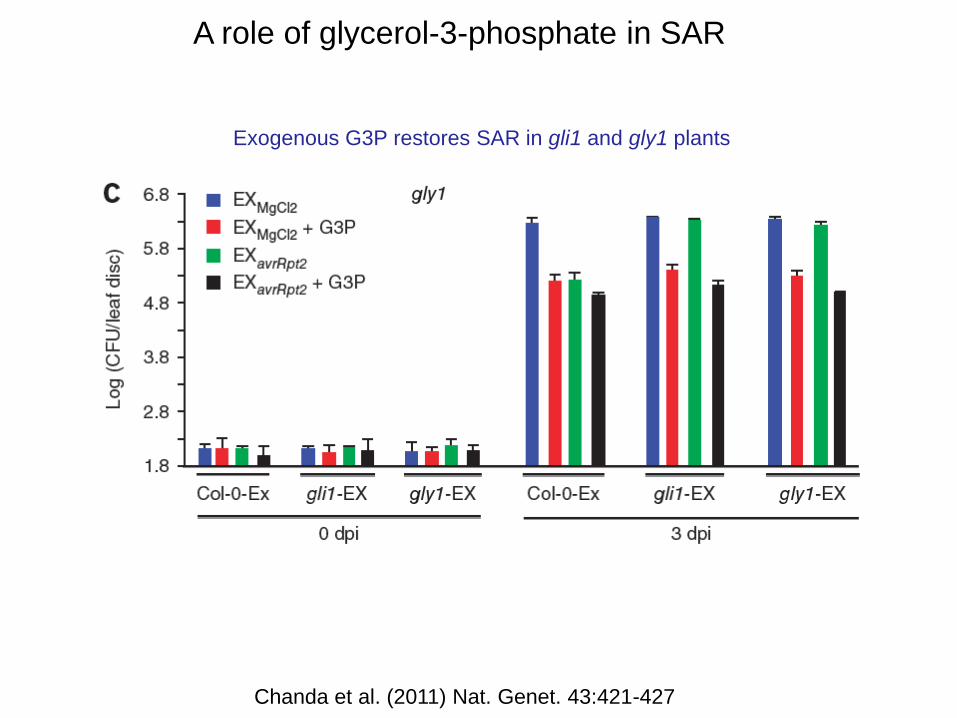
Exogenous G3P restores SAR in gli1 and gly1 plants
A role of glycerol-3-phosphate in SAR
Chanda et al. (2011) Nat. Genet. 43:421-427
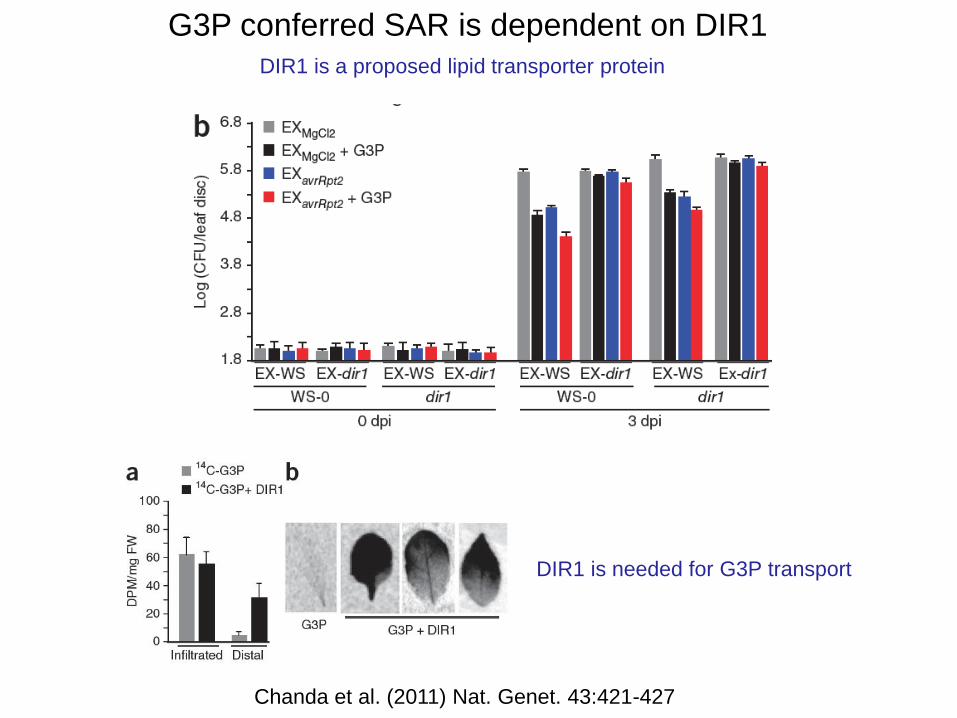
G3P conferred SAR is dependent on DIR1
DIR1 is a proposed lipid transporter protein
Chanda et al. (2011) Nat. Genet. 43:421-427
DIR1 is needed for G3P transport
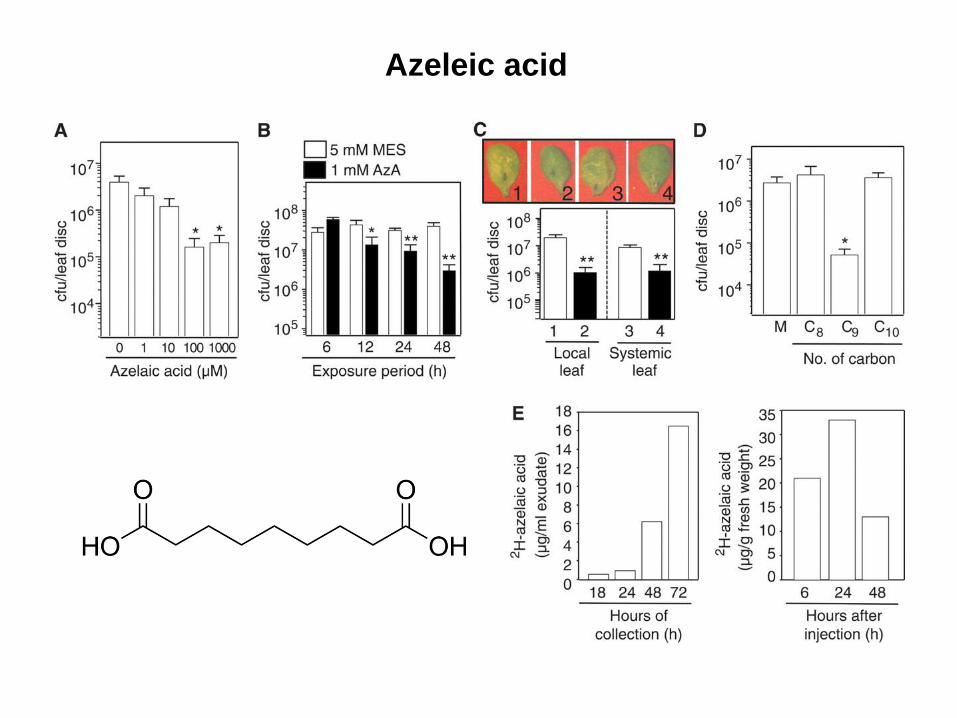
Azeleic acid
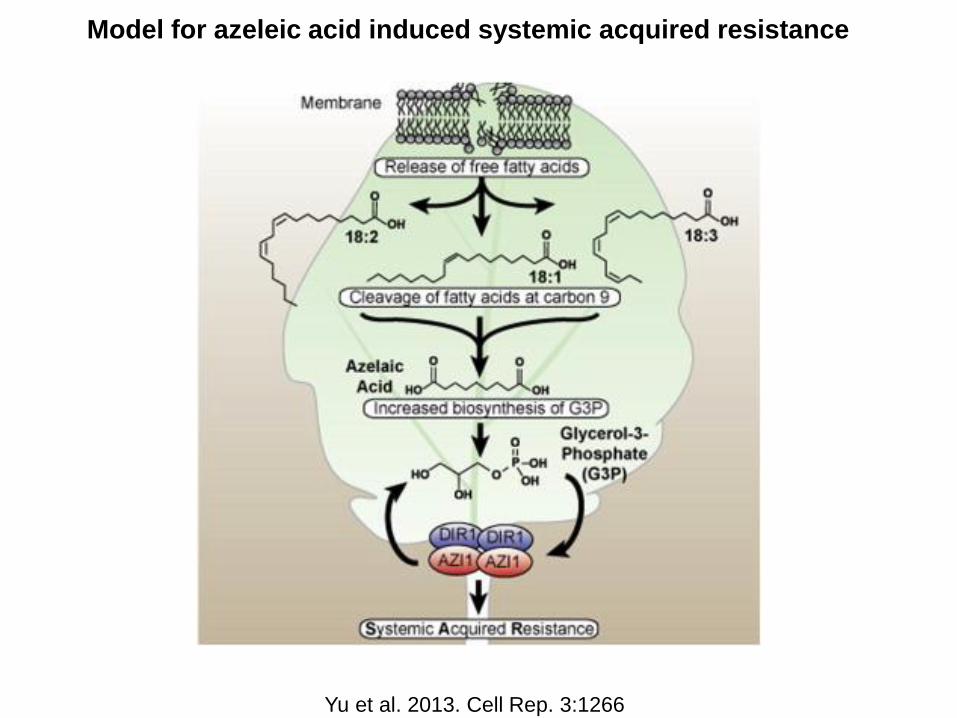
Model for azeleic acid induced systemic acquired resistance
Yu et al. 2013. Cell Rep. 3:1266
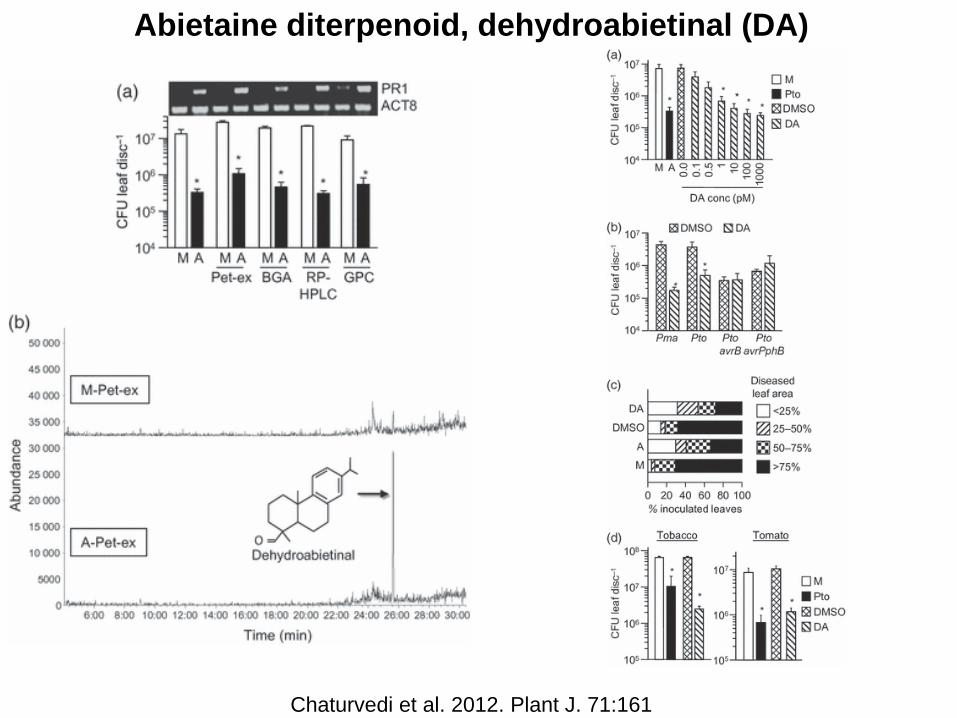
Abietaine diterpenoid, dehydroabietinal (DA)
Chaturvedi et al. 2012. Plant J. 71:161
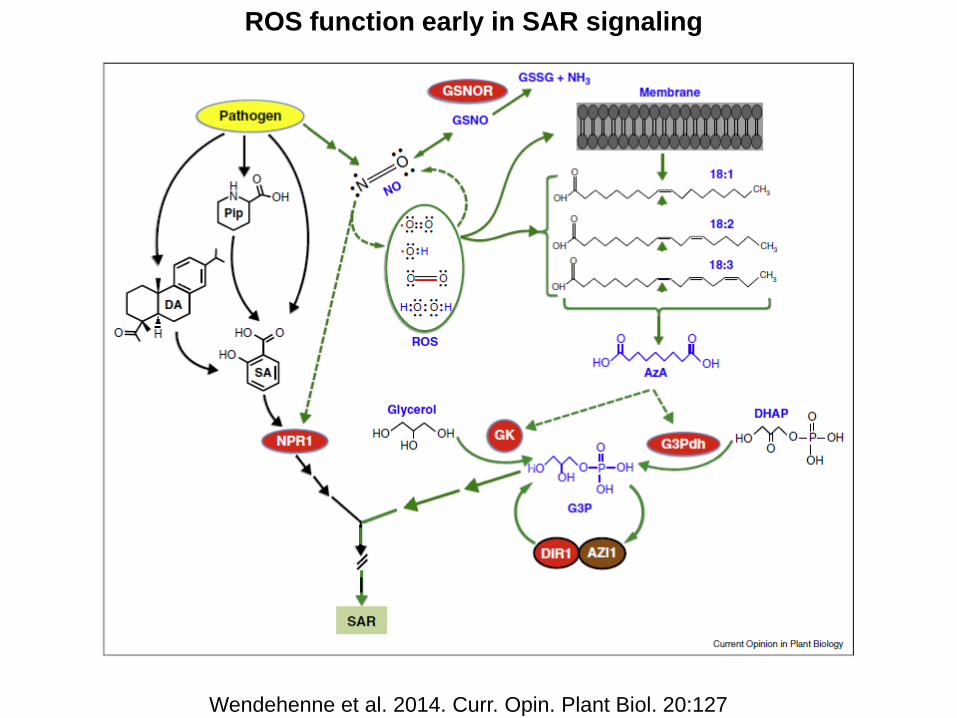
ROS function early in SAR signaling
Wendehenne et al. 2014. Curr. Opin. Plant Biol. 20:127
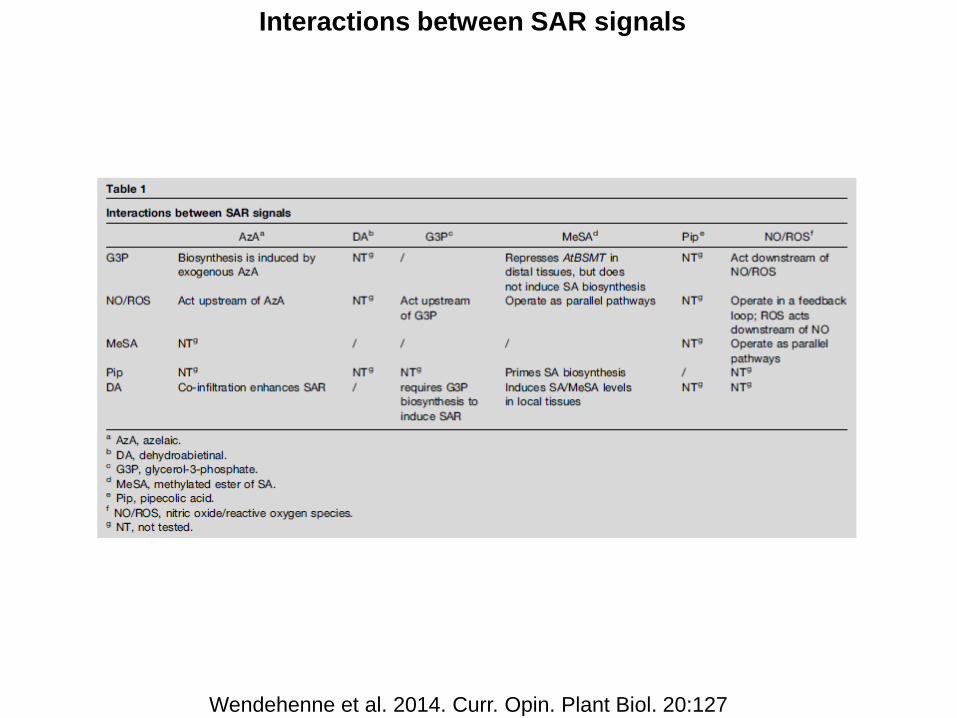
Interactions between SAR signals
Wendehenne et al. 2014. Curr. Opin. Plant Biol. 20:127

Signaling steps between SA and PR protein expression
and disease resistance.
The Arabidopsis NPR1 Gene That Controls
Systemic Acquired Resistance Encodes
a Novel Protein Containing Ankyrin RepeatsHui Cao, Jane Glazebrook, Joseph D. Clarke,
Sigrid Volko, and Xinnian Dong (1997) Cell 88, 57–63,
Previously: Linked a PR protein promoter called BGL2 to GUS.
Screened thousands of mutant transgenic BGL2-GUS plants for
ABSENCE of GUS activity induced by SA treatment.
Using standard Arabidopsis genetic mapping methods, identified a
single mutant gene, npr1. Phenotype:
Complete absence of GUS activity in response to SA
Absence of PR-1, PR-5, BGL2 expression in response to SA
Is now susceptible to Peronospora parasitica and to
Pseudomonas syringae pv maculicola (Psm).
Cao et al. (1997) Cell 88, 57–63,
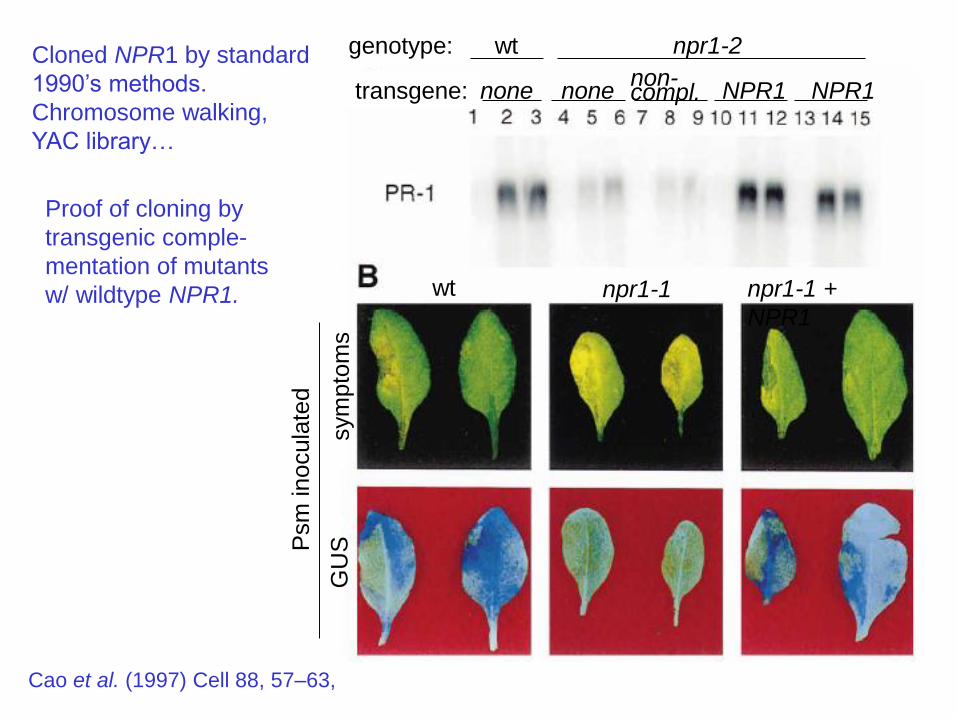
Cloned NPR1 by standard
1990’s methods.
Chromosome walking,
YAC library…
Proof of cloning by
transgenic comple-
mentation of mutants
w/ wildtype NPR1.
wt
nonenon-compl. NPR1
npr1-2
NPR1
genotype:
transgene: none
wt npr1-1 npr1-1 +
NPR1P
sm
inocula
ted
GU
Ssym
pto
ms
Cao et al. (1997) Cell 88, 57–63,

Cao et al. (1997) Cell 88, 57–63,
NPR1 has ankyrin repeats
Ankyrin repeats are
in lots of different
proteins.
Involved in protein-
protein interactions.
Especially in proteins
that control trans-
cription.
In NF-kB and I-kB in
mammals. Induced
by many pathogens,
stresses…
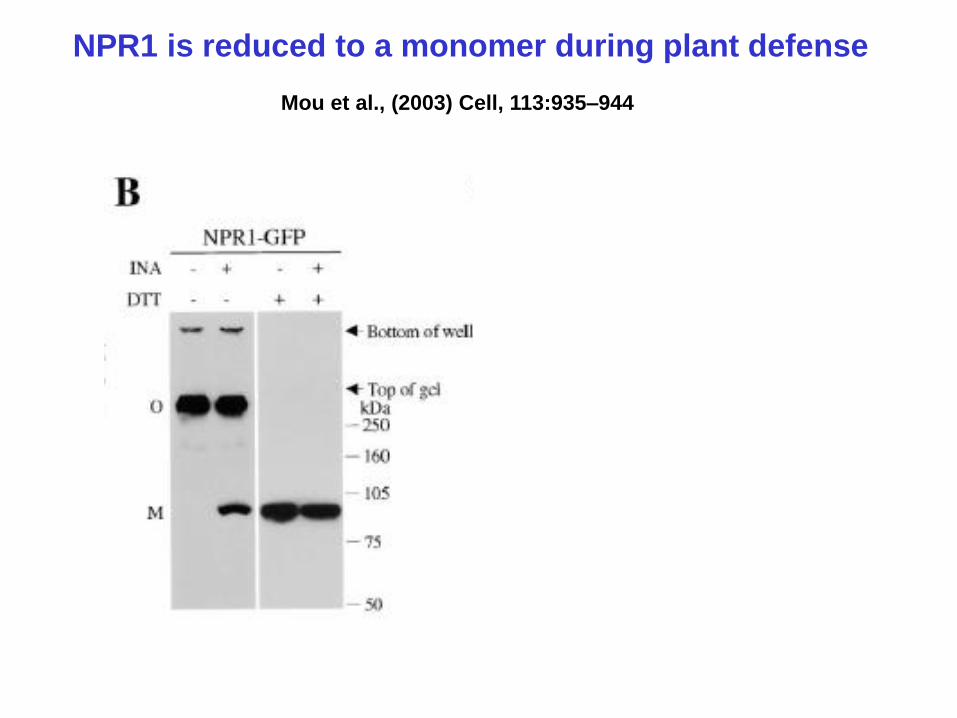
NPR1 is reduced to a monomer during plant defense
Mou et al., (2003) Cell, 113:935–944
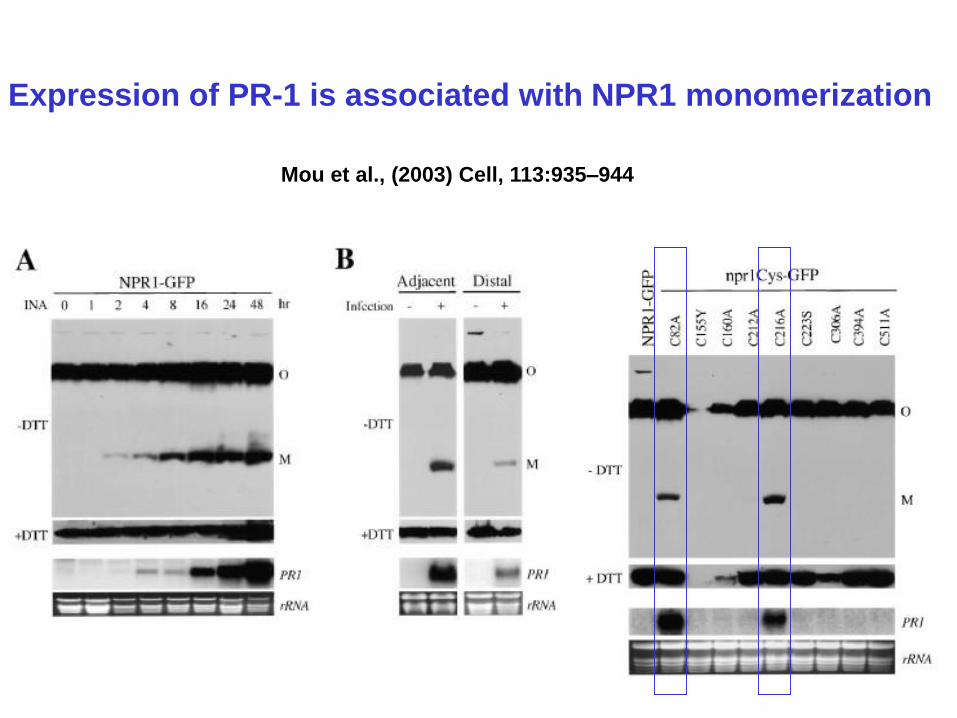
Expression of PR-1 is associated with NPR1 monomerization
Mou et al., (2003) Cell, 113:935–944
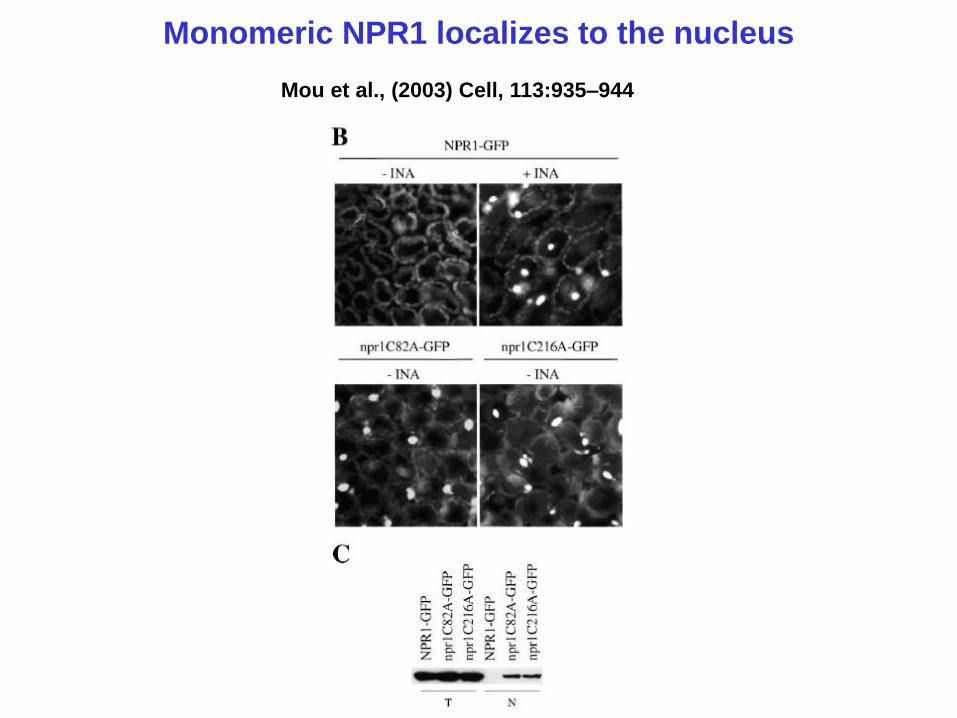
Monomeric NPR1 localizes to the nucleus
Mou et al., (2003) Cell, 113:935–944

Zhou et al. (2000) MPMI 13:191–202
NPR1 interacts with TGA transcription factors in yeast two-hybrid assays
TGA transcription factors bind to TGACG sequences found in the promoter of genes
such as PR-1
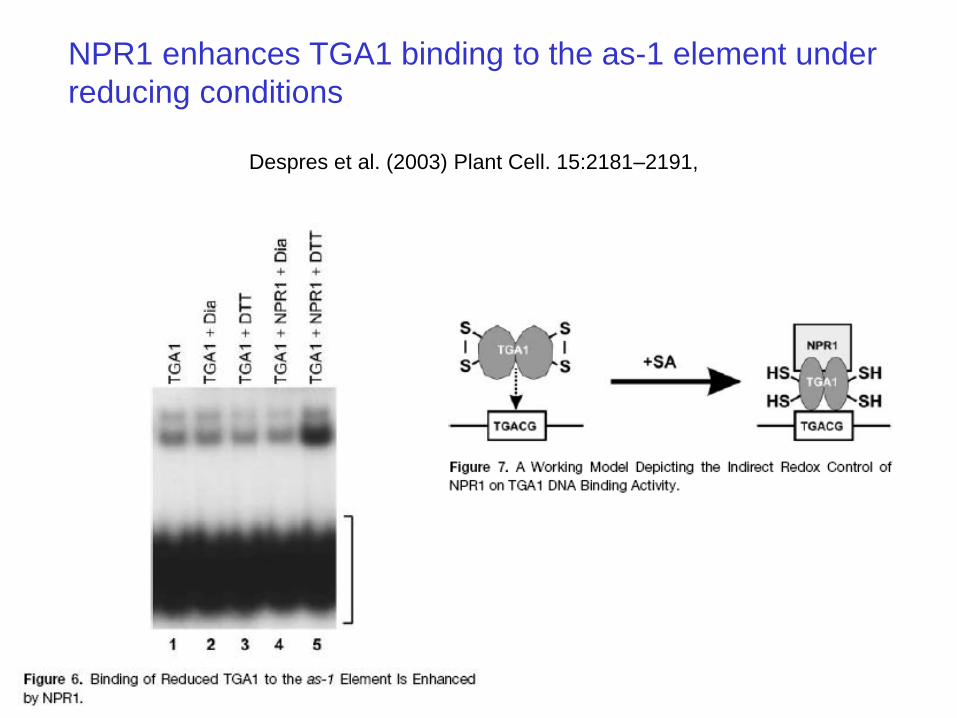
NPR1 enhances TGA1 binding to the as-1 element under
reducing conditions
Despres et al. (2003) Plant Cell. 15:2181–2191,

Recognize the motif: (T)(T)TGAC(C/T).
Have the conserved WRKYGQK at N-terminal end.
Have a novel zinc-finger-like motif.
Bind DNA via divalent cation (probably zinc).
Approx. 100 members of WRKY family in Arabidopsis.
NPR1 has 3 WRKY motifs in its promoter:
TTGACTTGACTTGGCTCTGCTCGTCAA
WRKY transcription factors: another key group of transcription
factors that regulate plant defense responses
The WRKY superfamily of plant transcription factors
Thomas Eulgem, Paul J. Rushton, Silke Robatzek and Imre E. Somssich
(2000) Trends Plant Sci 5, 199-205.

Conserved amino acids
in WRKY proteins of
Arabidopsis (red).
Putative Zn-finger ligands
are highlighted in black.
Eulgem et al. (2000) TIPS 5, 199-205.
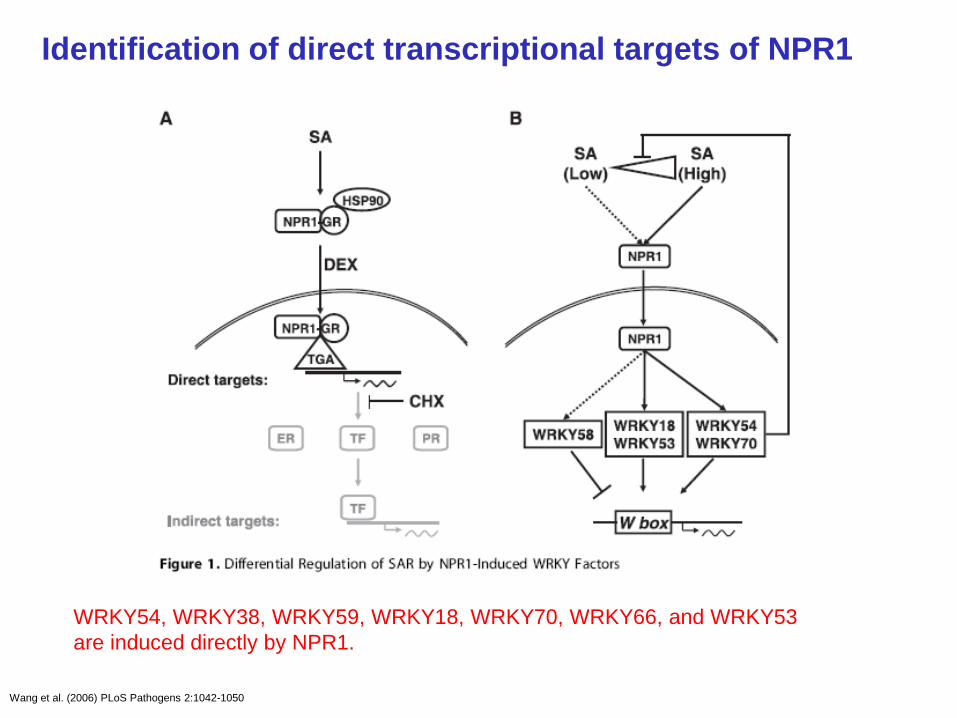
Wang et al. (2006) PLoS Pathogens 2:1042-1050
Identification of direct transcriptional targets of NPR1
WRKY54, WRKY38, WRKY59, WRKY18, WRKY70, WRKY66, and WRKY53
are induced directly by NPR1.
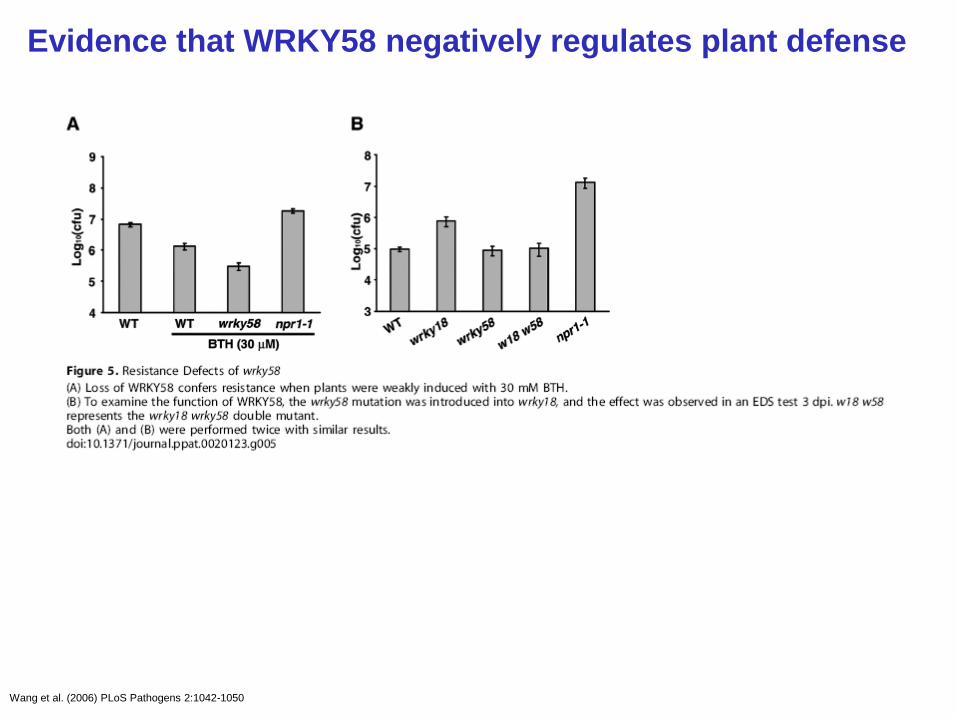
Evidence that WRKY58 negatively regulates plant defense
Wang et al. (2006) PLoS Pathogens 2:1042-1050
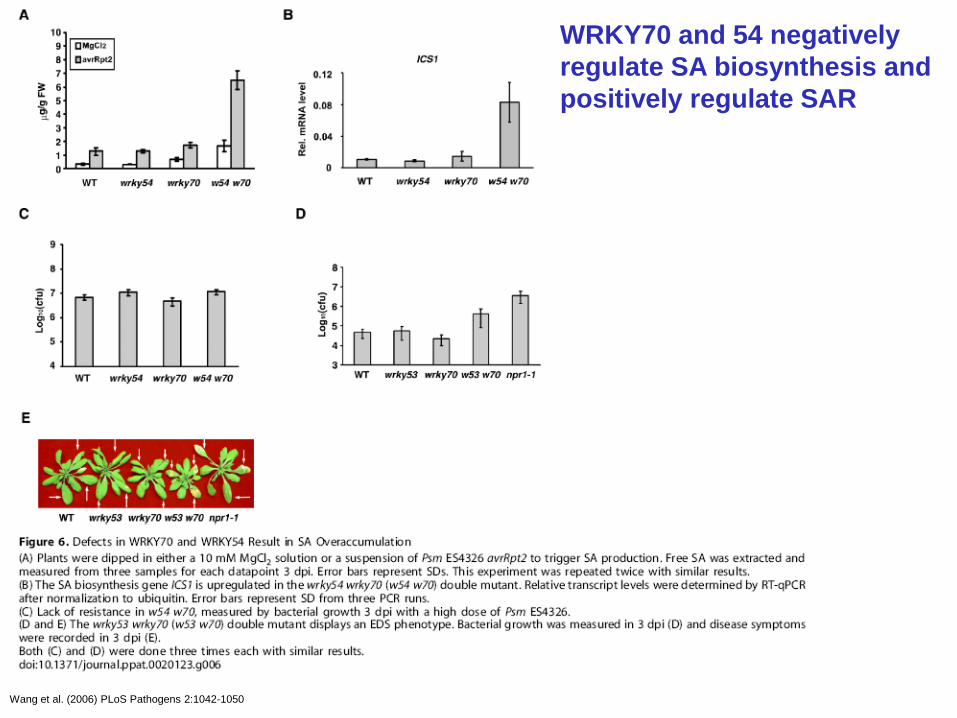
WRKY70 and 54 negatively
regulate SA biosynthesis and
positively regulate SAR
Wang et al. (2006) PLoS Pathogens 2:1042-1050

Identification and Cloning of a Negative Regulator
of Systemic Acquired Resistance, SNI1,
through a Screen for Suppressors of npr1-1Xin Li, Yuelin Zhang, Joseph D. Clarke, Yan Li,†
and Xinnian Dong* Cell, 98, 329–339, 1999.
Screened for EMS mutants of npr1-1 plants containing BGL2-GUS
reporter.
Look for plants that turn blue in response to INA (SA analog)
like NPR1 wild type plants.
But which, of course still harbor the npr1-1 mutation.
Found 11 loci that gave increased GUS, out of 7000 plants screened.
Li et al., Cell 98, 329–339.
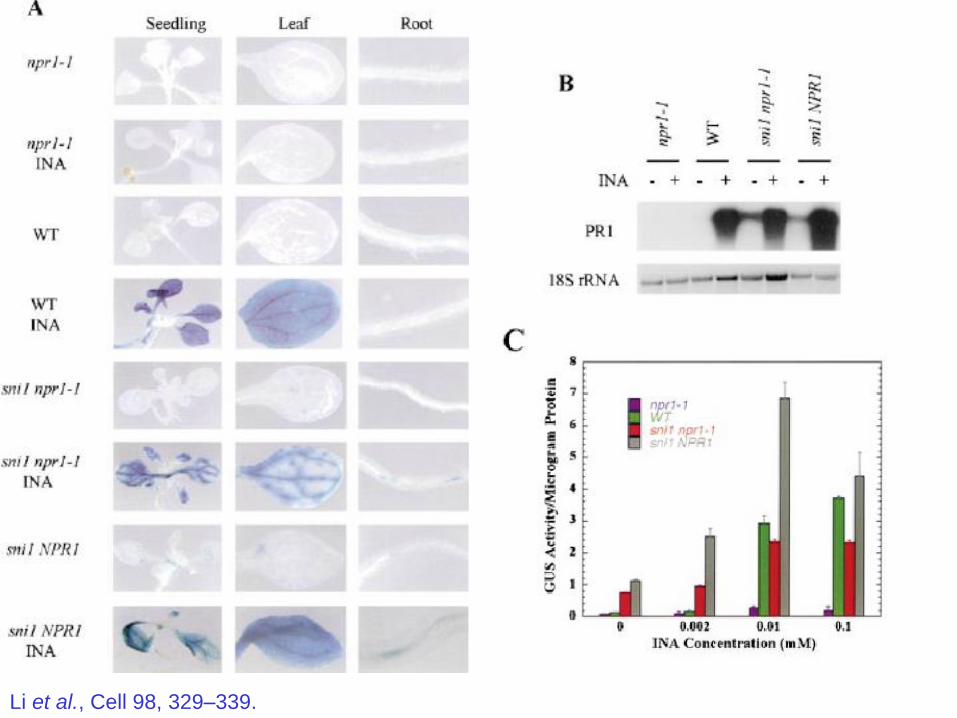
Li et al., Cell 98, 329–339.
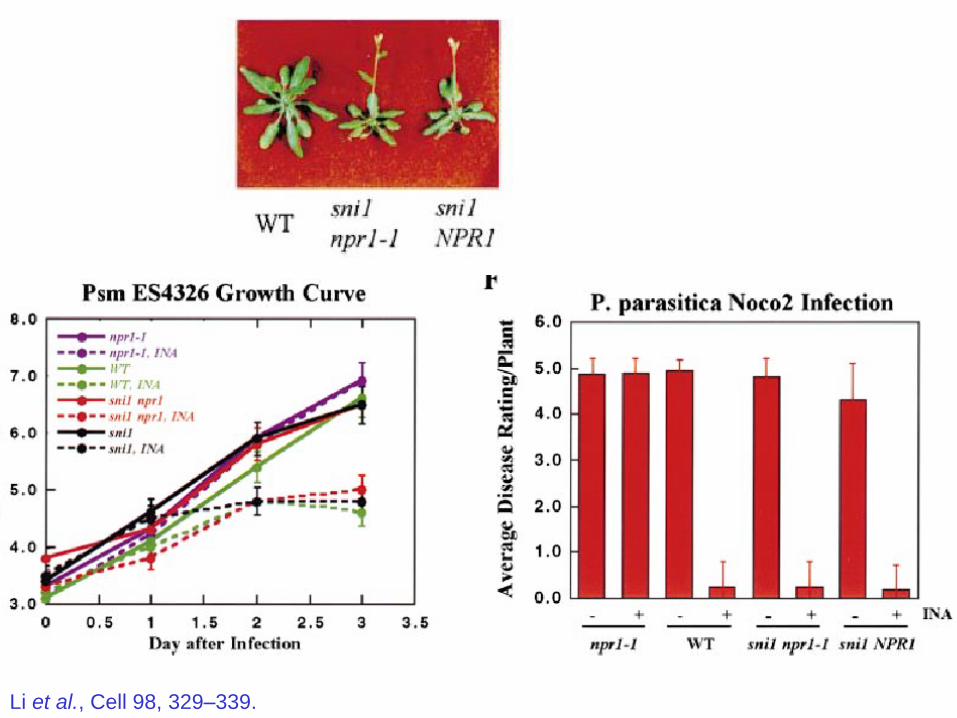
Li et al., Cell 98, 329–339.

SNI1 is similar to mouse Retinoblastoma (Rb). Rb is a tumor suppressor
that represses function of E2F transcription factor
Li et al., Cell 98, 329–339.
93-95% of genes upregulated in sni1 mutants are induced by BTH
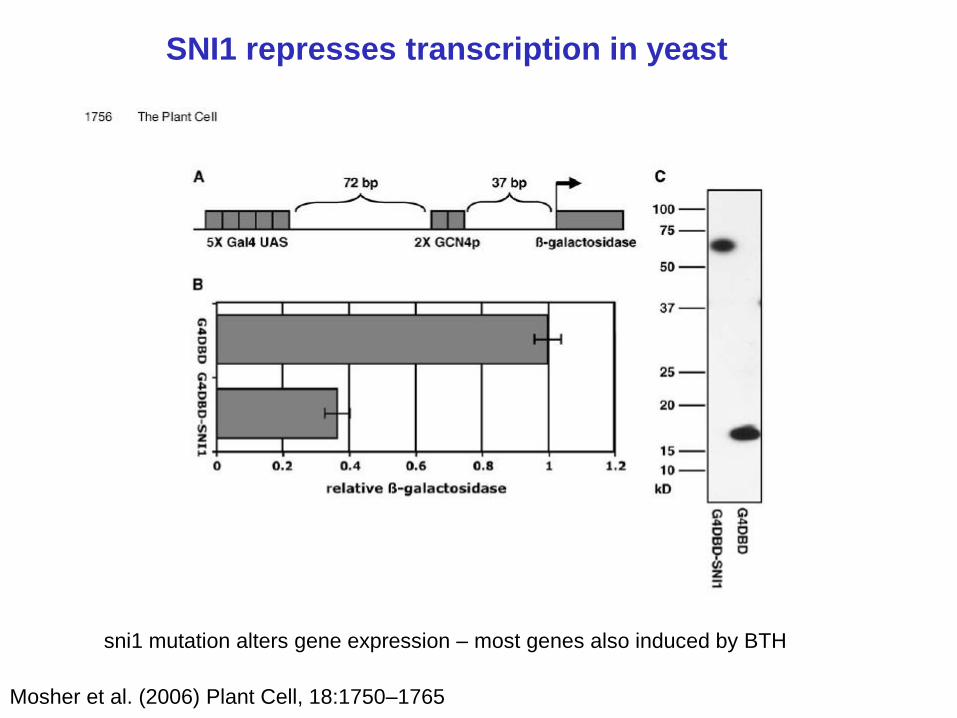
SNI1 represses transcription in yeast
Mosher et al. (2006) Plant Cell, 18:1750–1765
sni1 mutation alters gene expression – most genes also induced by BTH
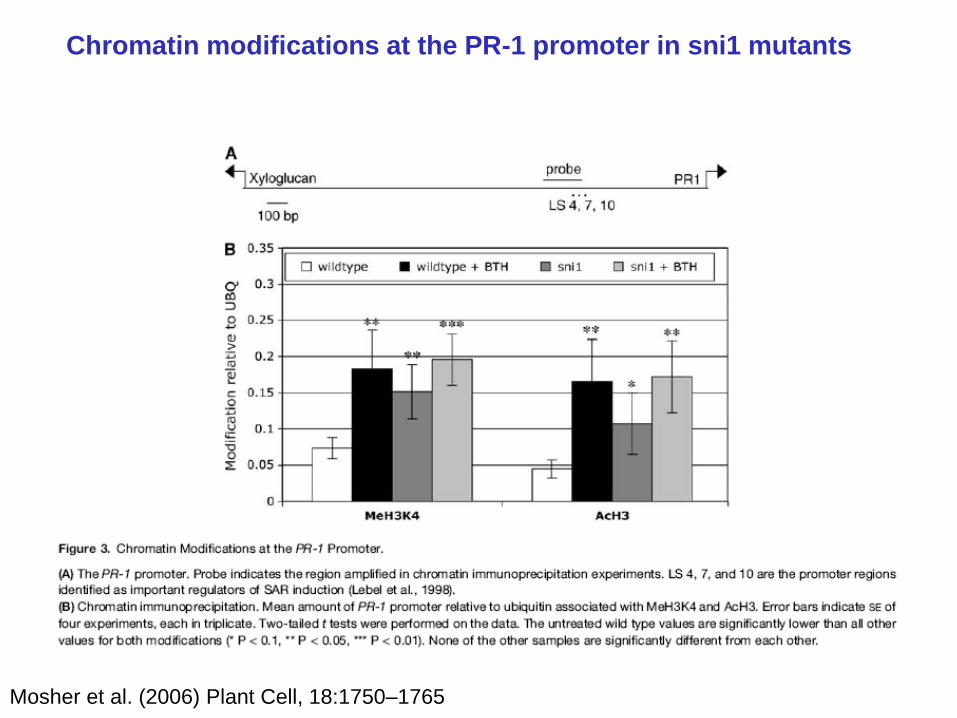
Chromatin modifications at the PR-1 promoter in sni1 mutants
Mosher et al. (2006) Plant Cell, 18:1750–1765
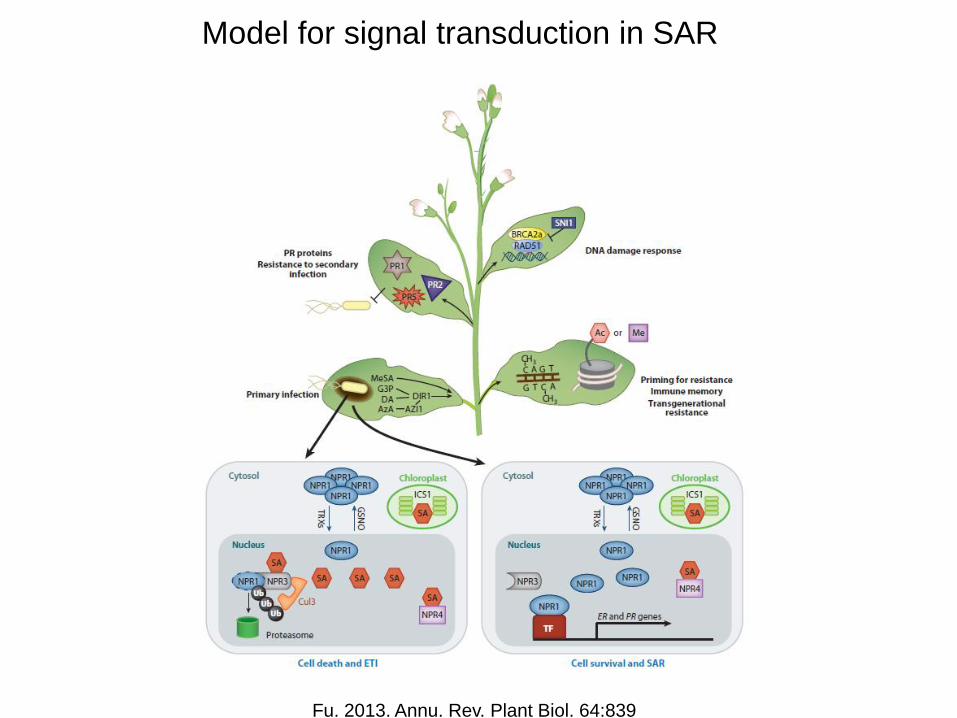
Model for signal transduction in SAR
Fu. 2013. Annu. Rev. Plant Biol. 64:839

Induced Systemic Resistance
(ISR)
- Plant responses to plant growth promoting rhizobacteria
- Herbivore induced resistance

Pieterse et al. (2014) Annu. Rev. Phytopathol. 52:347-75
Biologically induced systemic resistance
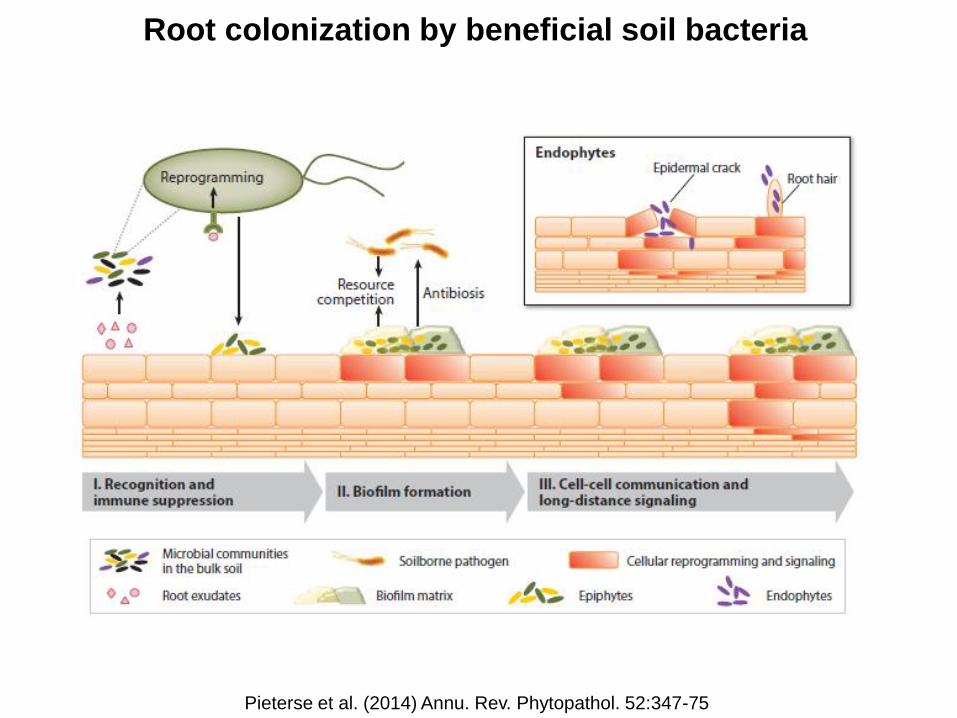
Root colonization by beneficial soil bacteria
Pieterse et al. (2014) Annu. Rev. Phytopathol. 52:347-75
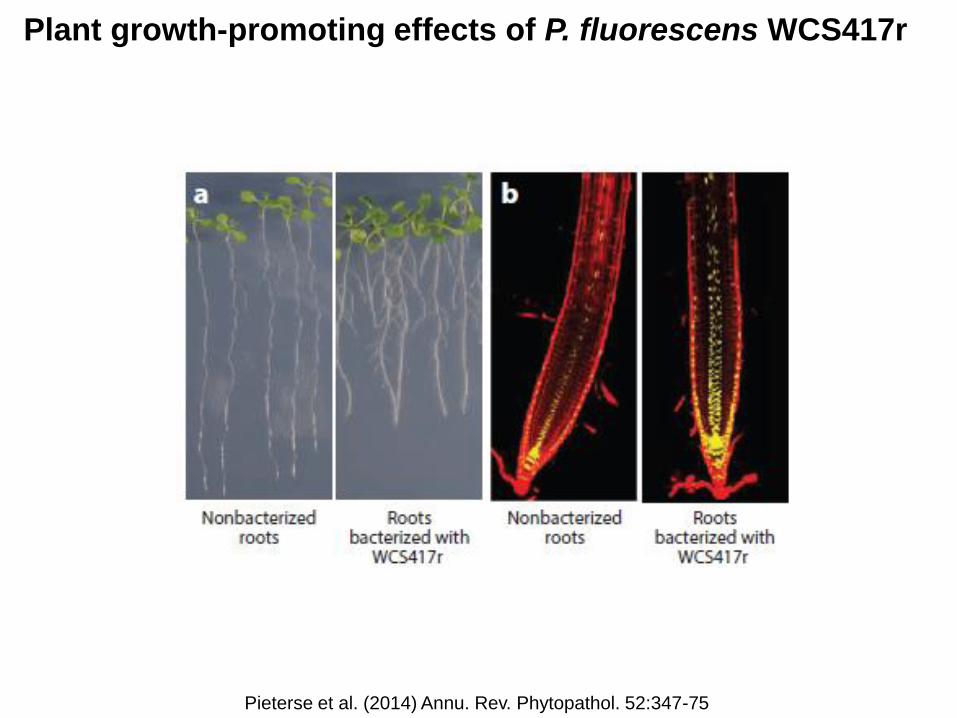
Plant growth-promoting effects of P. fluorescens WCS417r
Pieterse et al. (2014) Annu. Rev. Phytopathol. 52:347-75

Systemic protection against Cucumber mosaic virus
Kloepper. 2004. Phytopathology. 94:1259-1266
Bacillus pumulis strain SE34Nonbacterized
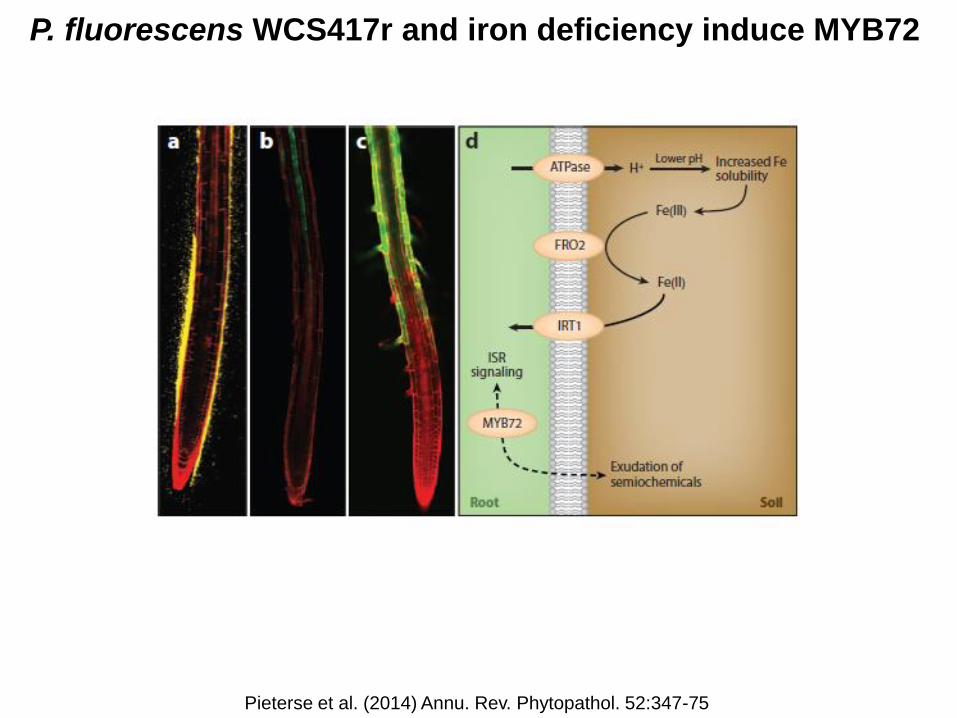
P. fluorescens WCS417r and iron deficiency induce MYB72
Pieterse et al. (2014) Annu. Rev. Phytopathol. 52:347-75

Properties of PGPR
- Stimulate growth
• N fixation
• Increase solubility of limiting nutrients (siderophores)
• Stimulate nutrient delivery and uptake
• Production of phytohormones
• Modulation of plant development (e.g. reduce ethylene
enhances root growth)
- Plant-mediated disease suppression
• Non-pathogens antagonize pathogens (competition,
antibiotics, lytic enzymes)
• Activating plant to better defend itself (ISR)
• Induced resistance observed on spatially separated
parts of same plant

The nature of systemically induced resistance in plants
(A) Characteristics of induced systemic resistance
• The defensive capacity of the plant is enhanced through microbial stimulation or
similar stresses
• The enhanced defensive capacity is expressed systemically throughout the
plant
• Induced systemic resistance is active against fungi, bacteria, viruses and,
sometimes, nematodes and insects
• Once induced, systemic resistance is maintained for prolonged periods
(B) Mechanisms of induced systemic resistance
• Developmental, escape: linked to growth promotion
• Physiological, tolerance: reduced symptom expression
• Environmental: associated with microbial antagonism in the rhizosphere;
altered plant-insect interactions
• Biochemical, resistance: induction of cell wall reinforcement,
• Induction of phytoalexins
• Induction of pathogenesis-related proteins
• ‘Priming’ of defence responses (resistance)
From Van Loon (2007) Eur. J. Plant Pathol. 119:243-254

ISR potentiates plant defense responses
Fusarium wilt of carnation and radish
Biocontrol by P. fluorescens WCS358
• Iron competition important: sid- mutant not effective
Biocontrol by P. fluorescens WCS417
•Twice as effective
•Sid-mutant still 100% effective
•Worked when WCS417 and fusarium were spatially separated on the
plant*
•WCS417 did not trigger phytoalexin accumulation*
•WCS417 treated plants produced more phytoalexin in response to
Fusarium*
*Van Peer et al (1991) Phytopathol 81:728-734
Root rot in beanAccelerated and potentiated papilla formation
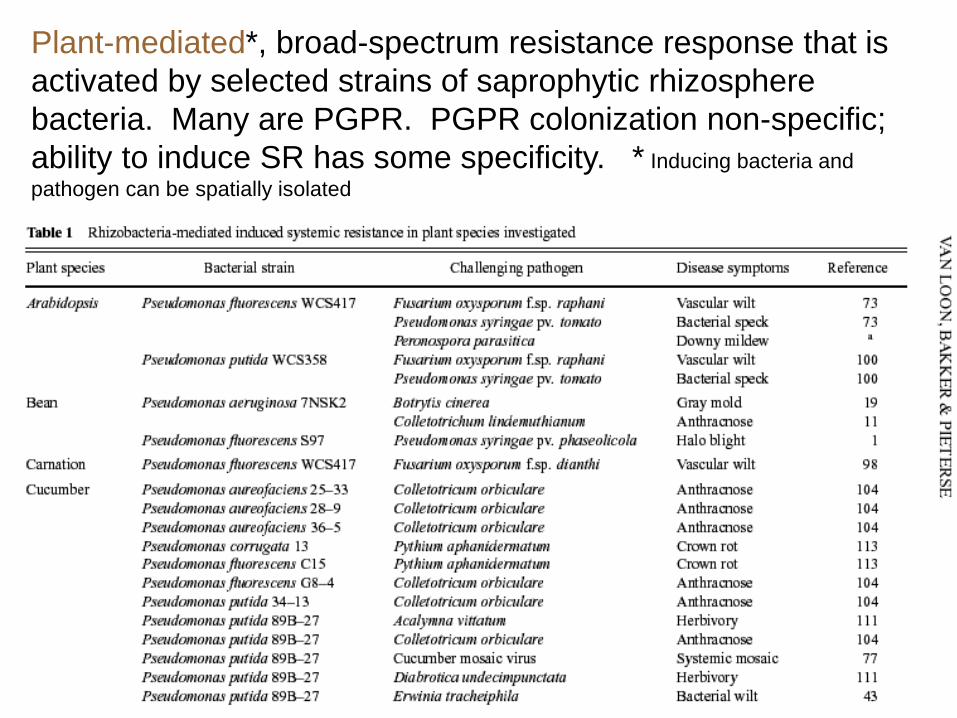
Plant-mediated*, broad-spectrum resistance response that is
activated by selected strains of saprophytic rhizosphere
bacteria. Many are PGPR. PGPR colonization non-specific;
ability to induce SR has some specificity. * Inducing bacteria and
pathogen can be spatially isolated


• ISR potentiates plant defense responses
• ISR is SA-independent
• ISR is independent of PR gene activation
• ISR requires JA and C2H4 response pathways
• ISR not associated with JA- and C2H4-responsive gene
activation
• ISR primes plant for enhanced C2H4 production?
• Summary: compare and contrast ISR and SAR
• Not covered in class:
1) bacterial determinants of ISR
2) field application of ISR
Summary of ISR molecular properties
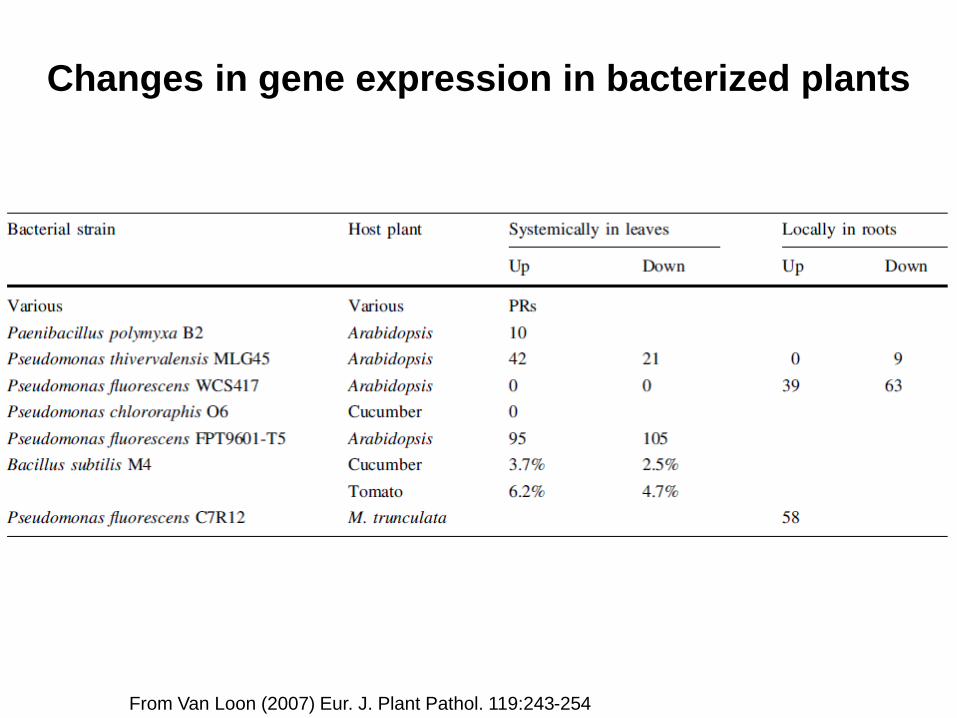
From Van Loon (2007) Eur. J. Plant Pathol. 119:243-254
Changes in gene expression in bacterized plants
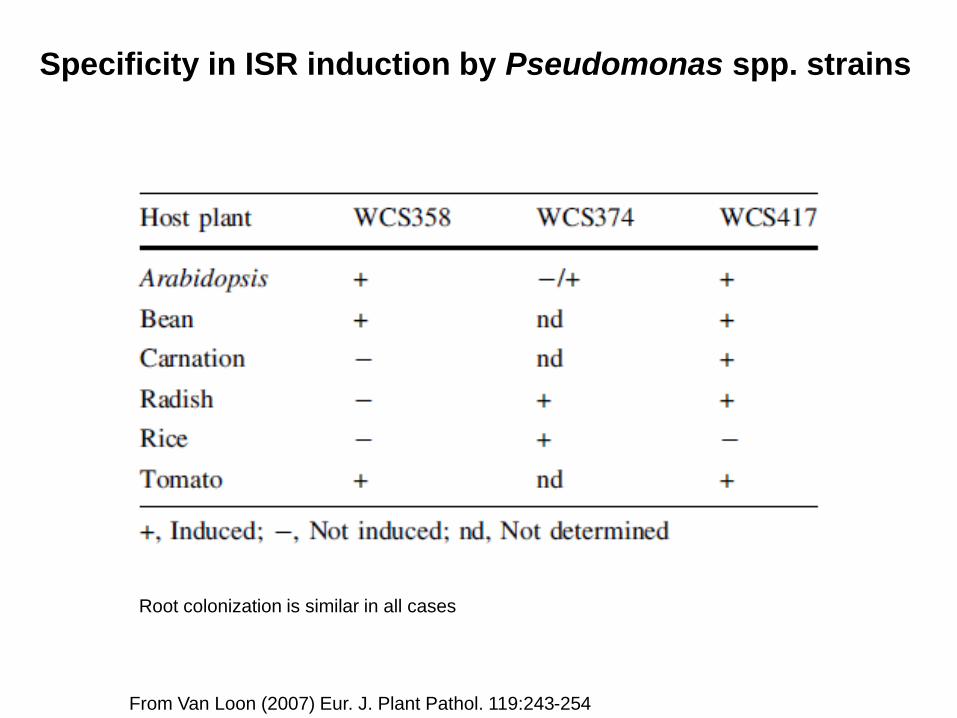
Specificity in ISR induction by Pseudomonas spp. strains
From Van Loon (2007) Eur. J. Plant Pathol. 119:243-254
Root colonization is similar in all cases

Model system – Arabidopsis/Pseudomonas fluorescens WCS417r
For Ps
Control
WCS417r
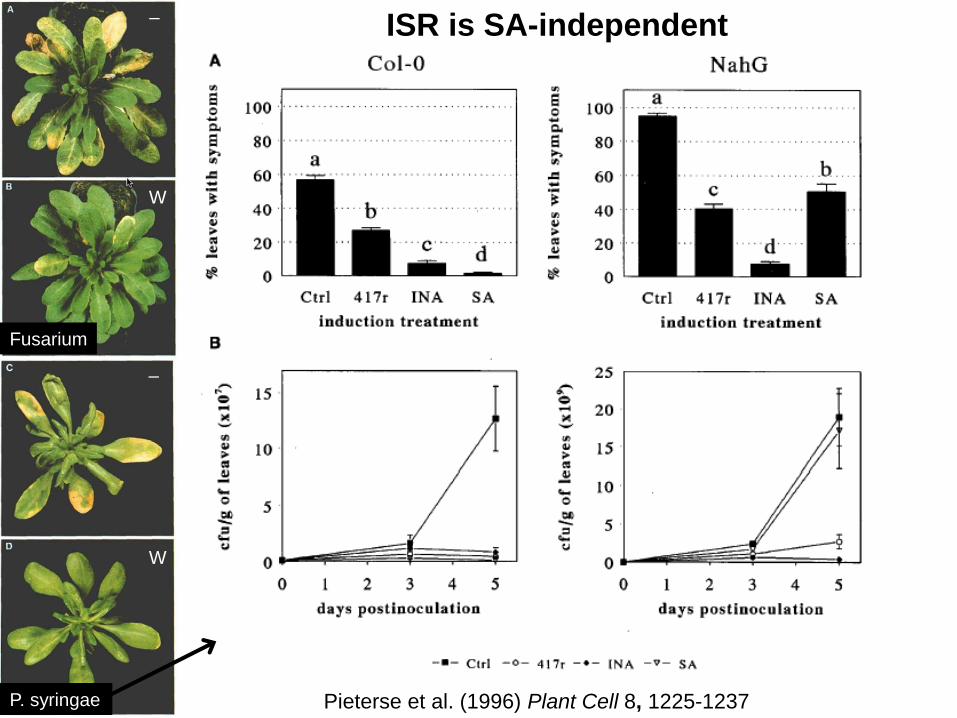
ISR is SA-independent
Pieterse et al. (1996) Plant Cell 8, 1225-1237
Fusarium
P. syringae
–
W
–
W
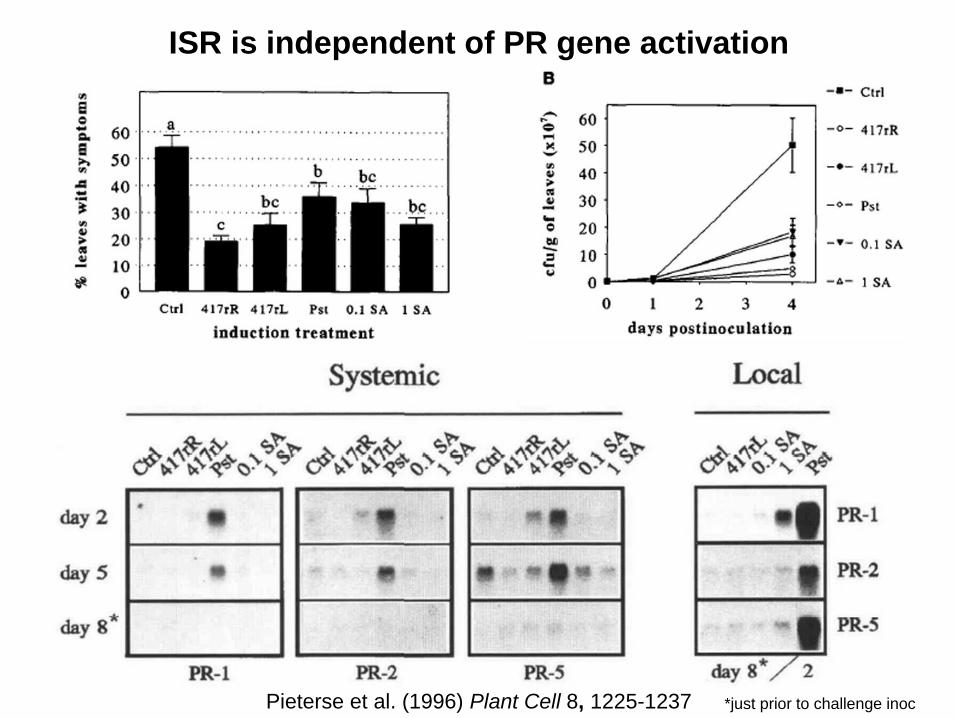
ISR is independent of PR gene activation
Pieterse et al. (1996) Plant Cell 8, 1225-1237 *just prior to challenge inoc
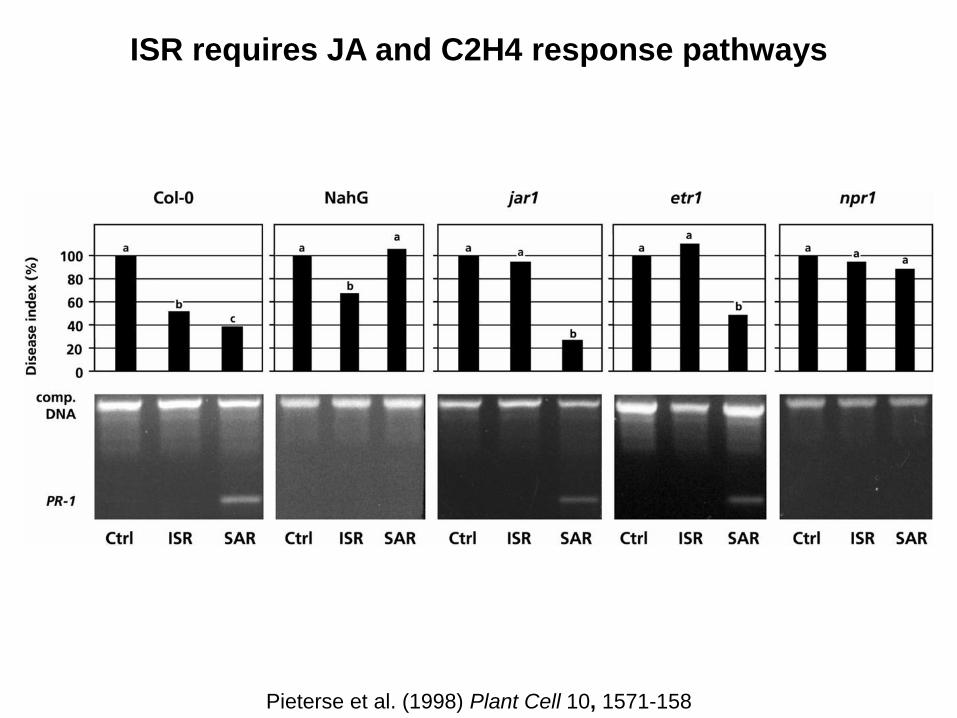
ISR requires JA and C2H4 response pathways
Pieterse et al. (1998) Plant Cell 10, 1571-158
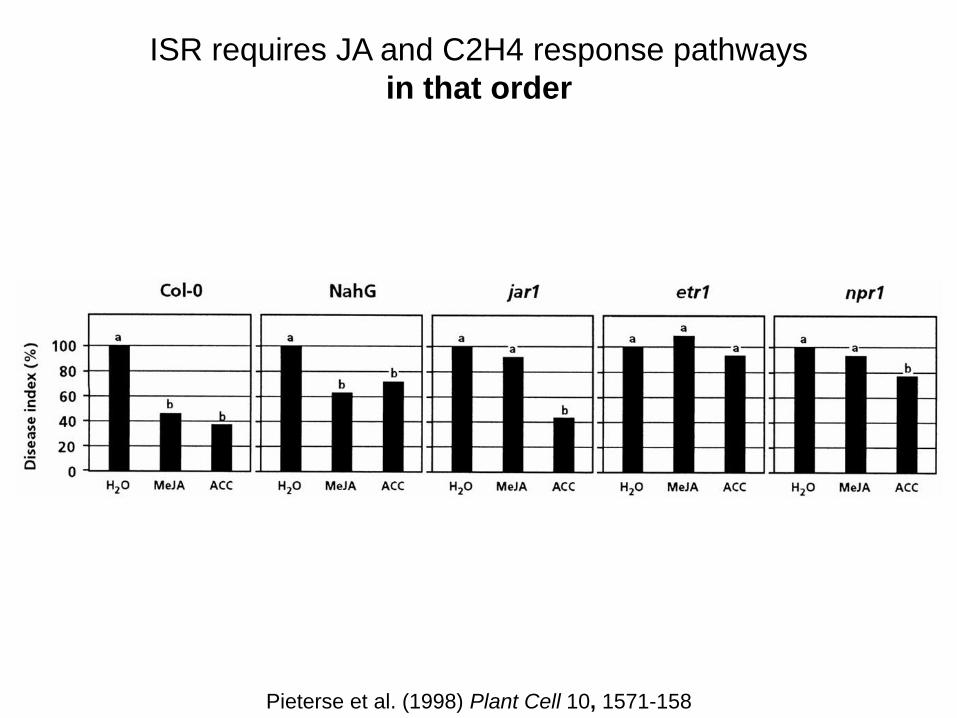
ISR requires JA and C2H4 response pathways
in that order
Pieterse et al. (1998) Plant Cell 10, 1571-158
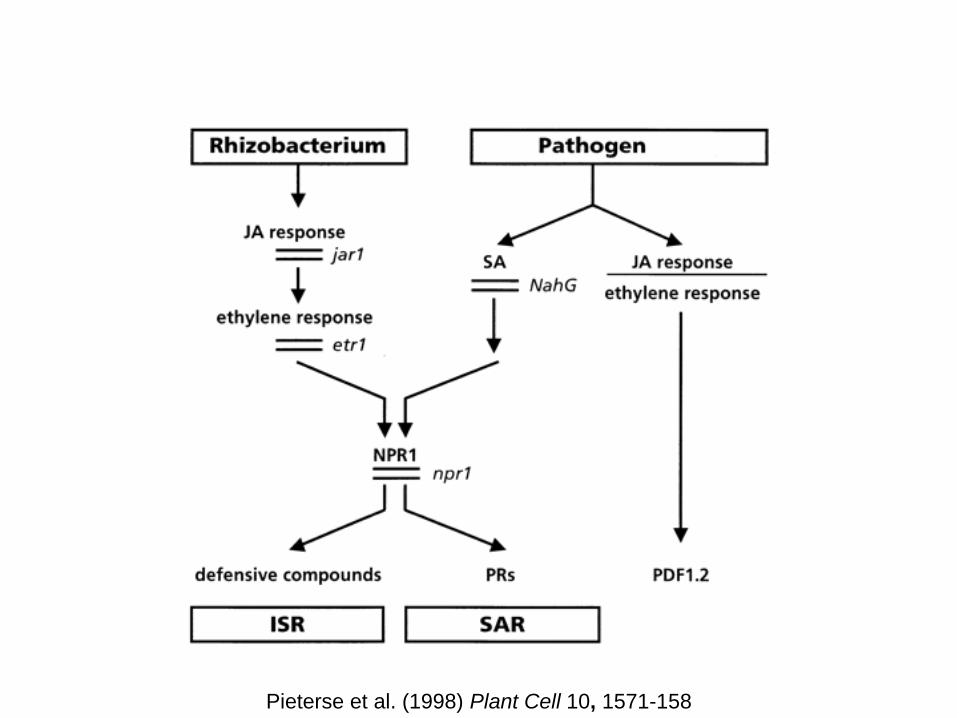
Pieterse et al. (1998) Plant Cell 10, 1571-158

ISR not associated with JA- and C2H4-responsive gene
activation
Pieterse et al. (1998) Plant Cell 10, 1571-158
But expression of this (and
not others) is potentiated in
plants undergoing ISR Van Wees et al. (1999) Plant Mol Biol
41:537-549
So ISR is associated with
potentiation of a specific
set of JA-responsive
genes?
Also, increased sensitivity
rather than increased
production of JA and
C2H4(see also Pieterse et al. 2000 Physiol.
Mol. Plant Pathol. 57:123-134)

ISR primes plant for enhanced C2H4 production?
ISR plants do not show increased levels of C2H4, or JA,
but
ISR activated plants convert more ACC to C2H4
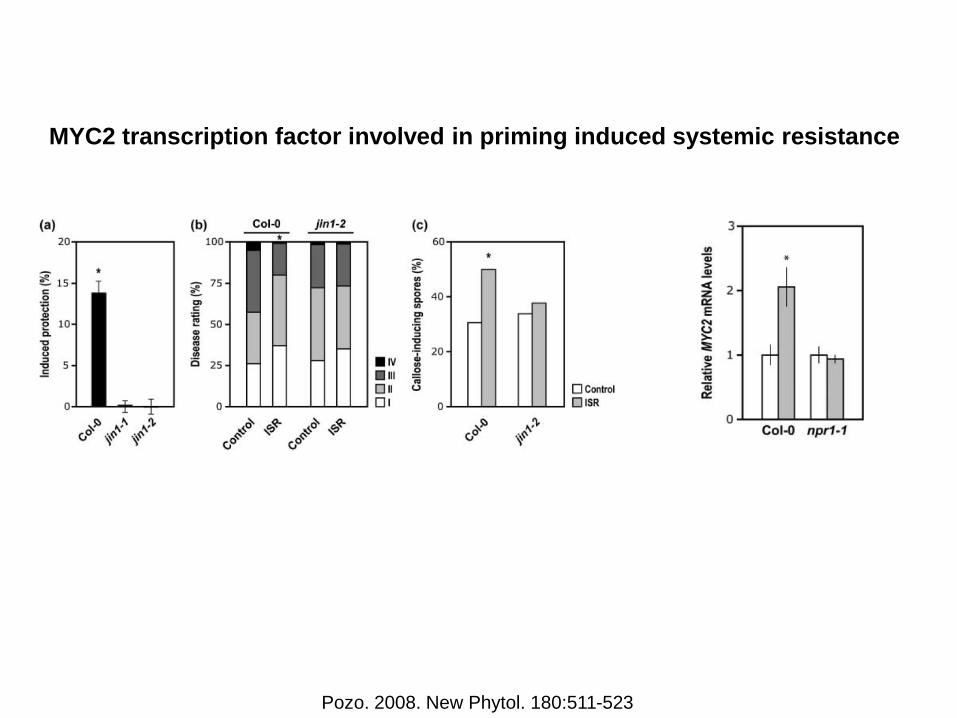
MYC2 transcription factor involved in priming induced systemic resistance
Pozo. 2008. New Phytol. 180:511-523
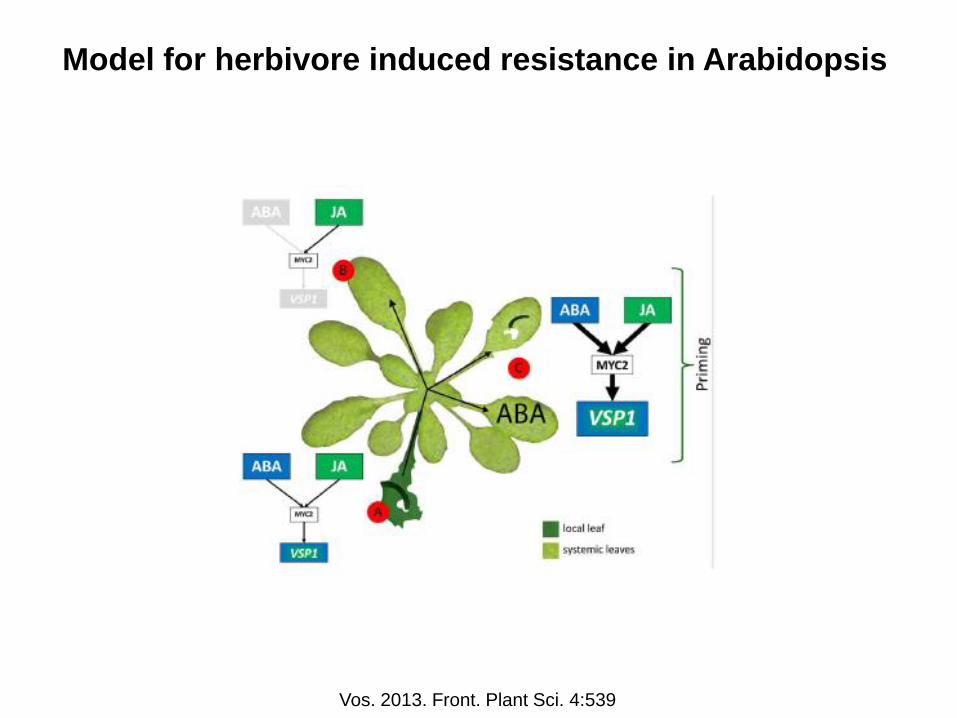
Model for herbivore induced resistance in Arabidopsis
Vos. 2013. Front. Plant Sci. 4:539
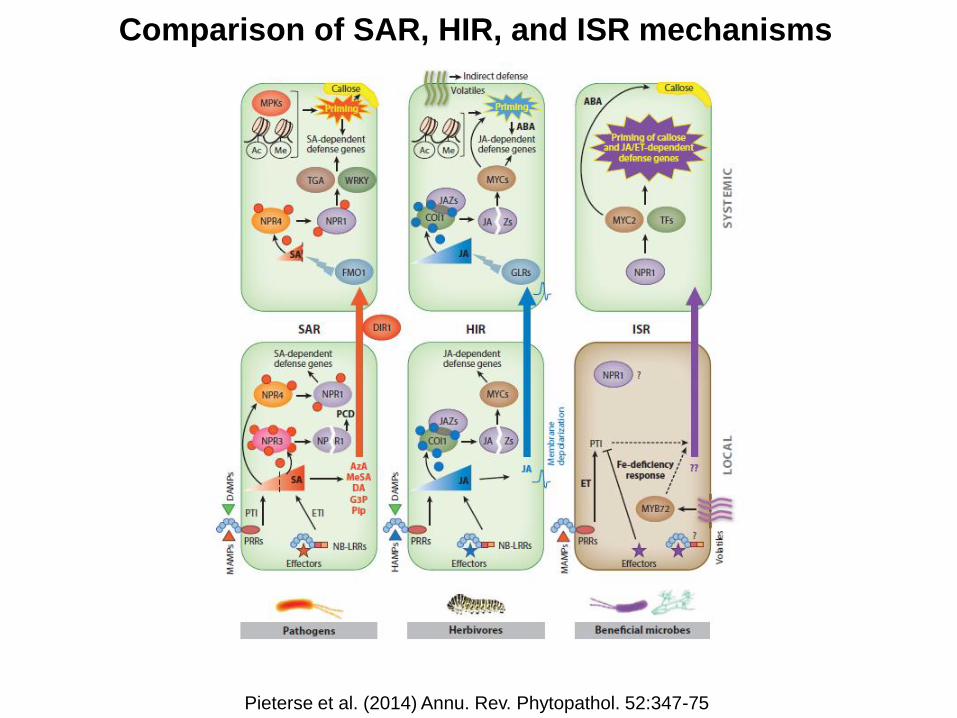
Comparison of SAR, HIR, and ISR mechanisms
Pieterse et al. (2014) Annu. Rev. Phytopathol. 52:347-75

Discussion
1) why is it important to study ISR?
2) do differences between ISR, HIR, SAR matter?
3) selective advantage to the plant and practical
significance?
4) what role does root microbiome play in immunity
5) how might ISR be triggered?
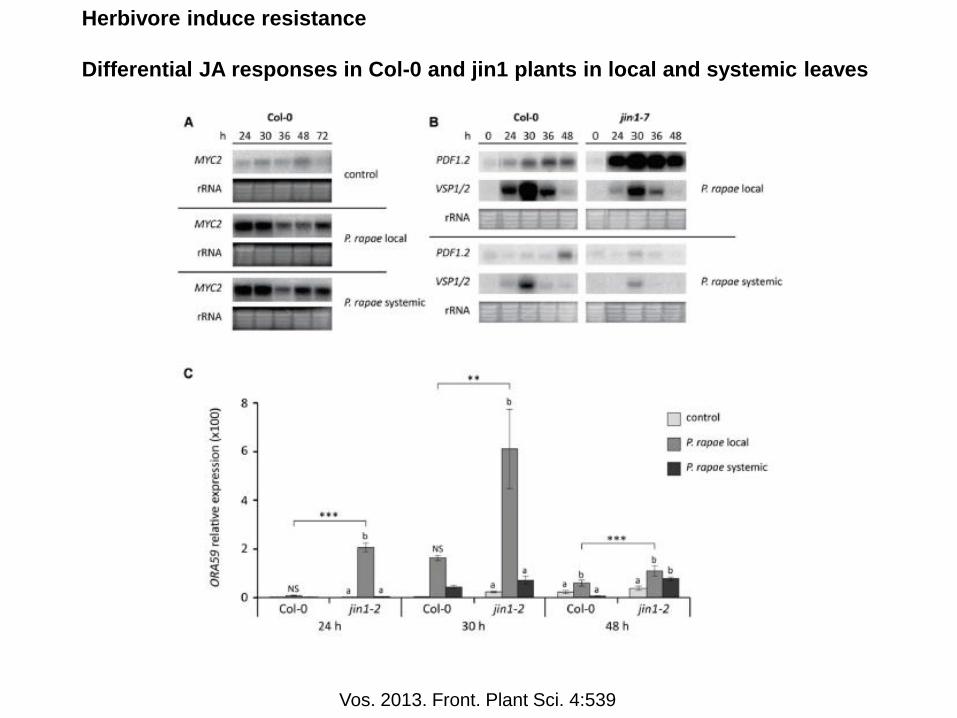
Herbivore induce resistance
Differential JA responses in Col-0 and jin1 plants in local and systemic leaves
Vos. 2013. Front. Plant Sci. 4:539
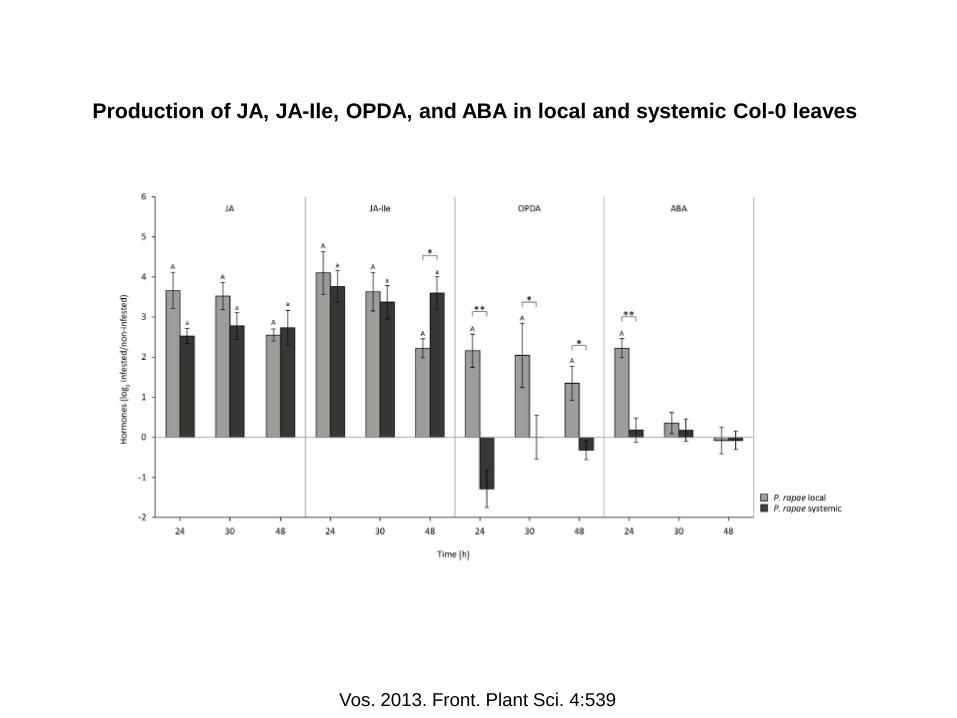
Production of JA, JA-Ile, OPDA, and ABA in local and systemic Col-0 leaves
Vos. 2013. Front. Plant Sci. 4:539
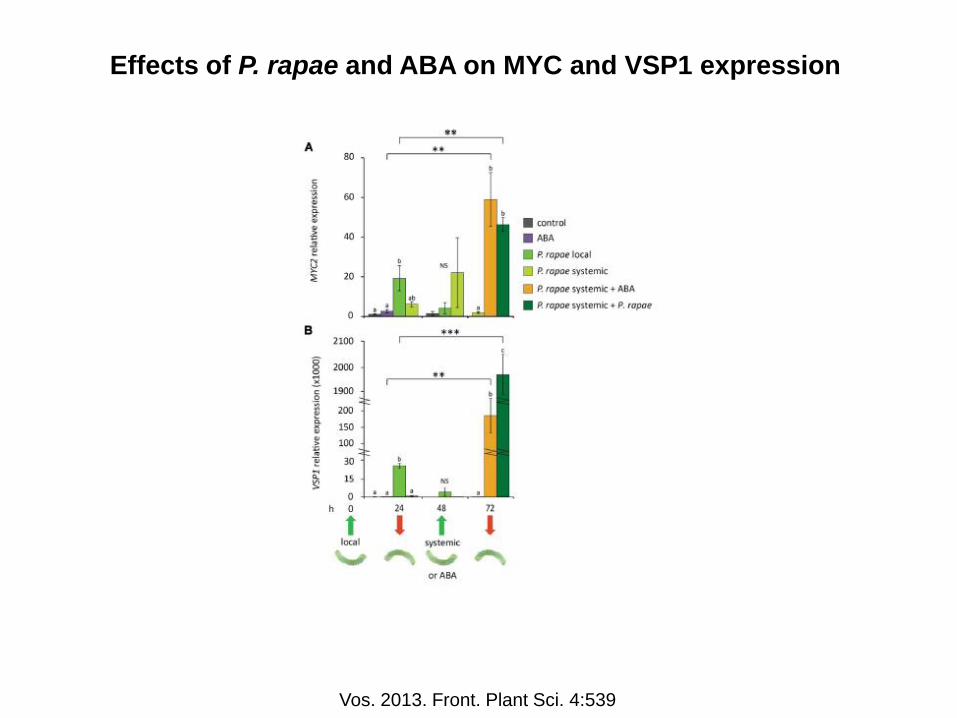
Effects of P. rapae and ABA on MYC and VSP1 expression
Vos. 2013. Front. Plant Sci. 4:539
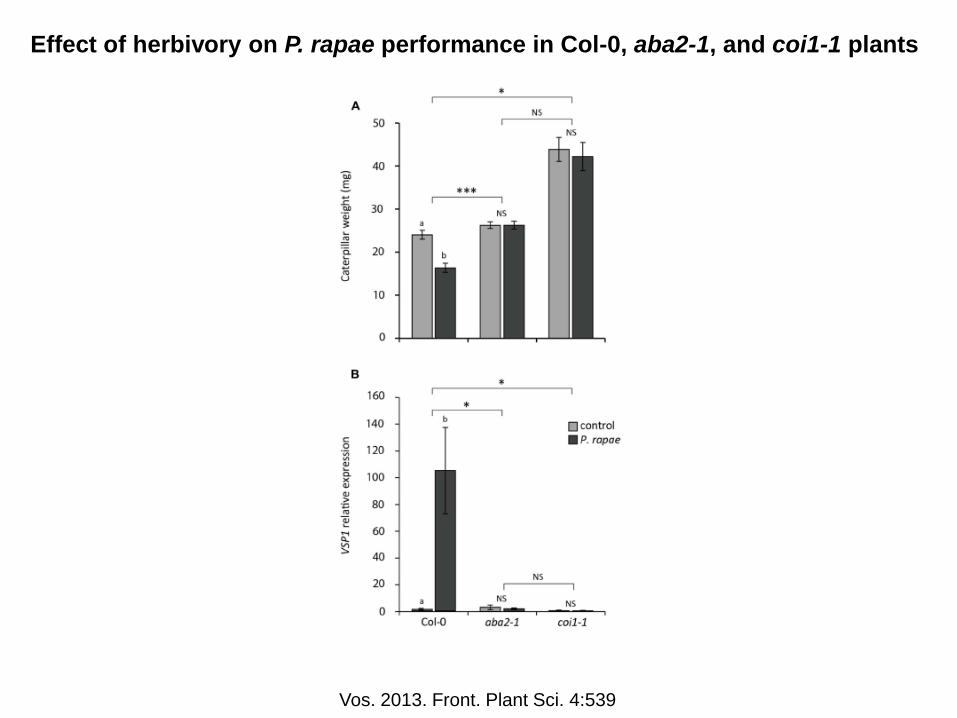
Effect of herbivory on P. rapae performance in Col-0, aba2-1, and coi1-1 plants
Vos. 2013. Front. Plant Sci. 4:539


















