
Pharmacogenomics of the response to IFN- in multiple ... · PRIORITY PAPER EVALUATION –...
7
PRIORITY PAPER EVALUATION 10.2217/14622416.9.5.639 © 2008 Future Medicine Ltd ISSN 1462-2416 Pharmacogenomics (2008) 9(5), 639–645 639 part of Pharmacogenomics of the response to IFN-β in multiple sclerosis: ramifications from the first genome-wide screen Koen Vandenbroeck 1† & Carlos Matute 2 † Author for correspondence 1 University of the Basque Country (UPV-EHU), Neurogenomiks Laboratory, Neuroscience Department, 48940 Leioa, Vizcaya, Spain Tel.: +34 944 706 400 ext. 379; E-mail: k.vandenbroeck@ ikerbasque.org 2 University of the Basque Country (UPV-EHU), Neurotek Laboratory, Neuroscience Department, 48940 Leioa, Vizcaya, Spain E-mail: [email protected] Keywords: genome, IFN-β, multiple sclerosis, polymorphism Evaluation of: Byun E, Caillier SJ, Montalban X et al.: Genome-wide pharmacogenomic analysis of the response to interferon-β therapy in multiple sclerosis. Arch. Neurol. 65(3) 337–344 (2008). Specifically, IFN-β is the most widely used disease-modifying therapy for the treatment of multiple sclerosis. The main benefits of the therapy, fewer and less severe relapses as well as delayed disease progression, are seen in only approximately 50% of the patients. Genetic polymorphisms may constitute in-built determinants of individual differences in response to IFN-β. Prior attempts to identify such ‘predictors of response’ were hypothesis-driven in that they were based on preselection of candidate genes associated with Type I interferon pathways. In the present study, the authors performed the first ever nonbiased genome-wide association screen in an attempt to identify response-predictive SNPs. Using a robust four-stage completion strategy coupled to advanced SNP ranking/clustering algorithms, 18 significant SNPs were identified, many of which are located in genes that have never before been linked clearly to Type I interferon biology or therapeutic effects. While this study was not designed per se so as to validate earlier findings, genes arising from previous pharmacogenomic studies were generally not confirmed. This is due to major discrepancies between interstudy sets of used SNPs, but may also reflect differential strategies for ascertainment of response to IFN-β, or simply Type I/II errors. The 100-K SNP screen by Byun et al. hallmarks a new stage of pharmacogenomics research applied to multiple sclerosis treatments. Through the judicious implementation of DNA pooling on SNP microarrays, it vividly demonstrates that informative genome-wide pharmacogenomic screens can be performed at a fraction of the cost of individual microarray genotyping. Although, unquestionably, higher-density SNP screens and further replication studies are needed, this study is instrumental in bringing the concept of personalized medicine a (small) step closer to the multiple sclerosis patient. In addition, it has generated a flurry of novel information of likely importance in furthering our understanding of Type I interferon biology. The results from a recent genome-wide pharmaco- genomic screen have provided a first glimpse into the complex, polygenic nature underlying the clinical response to IFN-β in multiple scle- rosis (MS) [1]. MS is a chronic inflammatory demyelinating disease of the CNS that affects predominantly young female adults (2:3 women:man). IFN-β is the most commonly used disease-modifying treatment and produces significant benefits [2], be it only in a proportion of treated patients (∼50%). The course of MS is largely unpredictable, and, as opposed to other diseases, clear quantitative criteria for manage- ment of both treatment and response to it are lacking. Identification of (a) marker(s) for IFN-β treatment efficacy would allow provision of the treatment to those patients likely to respond to it (or unlikely to experience adverse effects, such as depression or flu-like symptoms), whilst pro- viding an objective rationale for transferring probable nonresponders to a different treatment method (e.g., dacluzimab or natalizumab). Dis- covery of potential response biomarkers is cur- rently undertaken by means of diverse, but complementary, strategies, such as, gene- expression profiling [3–5] and proteomics-driven studies [6,7], or SNP screens [8–13]. Of these approaches, the potential for routine implemen- tation of response-predictive SNPs is particularly attractive (no issues with marker fluctuation over time and according to cell type, cost–effective- ness, minimally invasive to the patient, e.g., DNA collection through mouth swab; by con- trast, proteomics/transcriptomics methods are more likely to depend on accessible specific cell populations and/or body fluids). For now, it is still not known (and will likely remain so for some time) which strategy or combination of strategies will ultimately deliver the most reliable IFN-β response marker(s).
Transcript of Pharmacogenomics of the response to IFN- in multiple ... · PRIORITY PAPER EVALUATION –...

PRIORITY PAPER EVALUATION
Pharmacogenomics of the response to IFN-β in multiple sclerosis: ramifications from the first genome-wide screen
Koen Vandenbroeck1† & Carlos Matute2†Author for correspondence1University of the Basque Country (UPV-EHU), Neurogenomiks Laboratory, Neuroscience Department, 48940 Leioa, Vizcaya, SpainTel.: +34 944 706 400 ext. 379;E-mail: [email protected] of the Basque Country (UPV-EHU), Neurotek Laboratory, Neuroscience Department, 48940 Leioa, Vizcaya, SpainE-mail: [email protected]
part of
Keywords: genome, IFN-β, multiple sclerosis, polymorphism
10.2217/14622416.9.5.639 © 2
Evaluation of: Byun E, Caillier SJ, Montalban X et al.: Genome-wide pharmacogenomic analysis of the response to interferon-β therapy in multiple sclerosis. Arch. Neurol. 65(3) 337–344 (2008). Specifically, IFN-β is the most widely used disease-modifying therapy for the treatment of multiple sclerosis. The main benefits of the therapy, fewer and less severe relapses as well as delayed disease progression, are seen in only approximately 50% of the patients. Genetic polymorphisms may constitute in-built determinants of individual differences in response to IFN-β. Prior attempts to identify such ‘predictors of response’ were hypothesis-driven in that they were based on preselection of candidate genes associated with Type I interferon pathways. In the present study, the authors performed the first ever nonbiased genome-wide association screen in an attempt to identify response-predictive SNPs. Using a robust four-stage completion strategy coupled to advanced SNP ranking/clustering algorithms, 18 significant SNPs were identified, many of which are located in genes that have never before been linked clearly to Type I interferon biology or therapeutic effects. While this study was not designed per se so as to validate earlier findings, genes arising from previous pharmacogenomic studies were generally not confirmed. This is due to major discrepancies between interstudy sets of used SNPs, but may also reflect differential strategies for ascertainment of response to IFN-β, or simply Type I/II errors. The 100-K SNP screen by Byun et al. hallmarks a new stage of pharmacogenomics research applied to multiple sclerosis treatments. Through the judicious implementation of DNA pooling on SNP microarrays, it vividly demonstrates that informative genome-wide pharmacogenomic screens can be performed at a fraction of the cost of individual microarray genotyping. Although, unquestionably, higher-density SNP screens and further replication studies are needed, this study is instrumental in bringing the concept of personalized medicine a (small) step closer to the multiple sclerosis patient. In addition, it has generated a flurry of novel information of likely importance in furthering our understanding of Type I interferon biology.
The results from a recent genome-wide pharmaco-genomic screen have provided a first glimpseinto the complex, polygenic nature underlyingthe clinical response to IFN-β in multiple scle-rosis (MS) [1]. MS is a chronic inflammatorydemyelinating disease of the CNS that affectspredominantly young female adults (2:3women:man). IFN-β is the most commonlyused disease-modifying treatment and producessignificant benefits [2], be it only in a proportionof treated patients (∼50%). The course of MS islargely unpredictable, and, as opposed to otherdiseases, clear quantitative criteria for manage-ment of both treatment and response to it arelacking. Identification of (a) marker(s) for IFN-βtreatment efficacy would allow provision of thetreatment to those patients likely to respond to it(or unlikely to experience adverse effects, suchas depression or flu-like symptoms), whilst pro-viding an objective rationale for transferring
probable nonresponders to a different treatmentmethod (e.g., dacluzimab or natalizumab). Dis-covery of potential response biomarkers is cur-rently undertaken by means of diverse, butcomplementary, strategies, such as, gene-expression profiling [3–5] and proteomics-drivenstudies [6,7], or SNP screens [8–13]. Of theseapproaches, the potential for routine implemen-tation of response-predictive SNPs is particularlyattractive (no issues with marker fluctuation overtime and according to cell type, cost–effective-ness, minimally invasive to the patient, e.g.,DNA collection through mouth swab; by con-trast, proteomics/transcriptomics methods aremore likely to depend on accessible specific cellpopulations and/or body fluids). For now, it isstill not known (and will likely remain so forsome time) which strategy or combination ofstrategies will ultimately deliver the most reliableIFN-β response marker(s).
008 Future Medicine Ltd ISSN 1462-2416 Pharmacogenomics (2008) 9(5), 639–645 639

PRIORITY PAPER EVALUATION – Vandenbroeck & Matute
640
The area of pharmacogenomics researchapplied to MS treatments is, as yet, quite young,and studies focusing on potential SNP bio-markers have been limited to the scrutiny of pre-selected candidate genes thought to beassociated with Type I interferon biology(reviewed in [14]). The study by Byun and col-leagues [1] constitutes a significant departurefrom these earlier studies in that it represents thefirst ever attempt to identify of molecular mark-ers for interferon therapy response heterogeneitythrough a genome-wide screen.
Summary of methods & resultsMultiple sclerosis patients included in the screenwere selected from four collaborating centerslocated in the genetically homogeneous Mediter-ranean basin [1]. This selection was done pur-posely in order to minimize population-basedallelic differences between patient-recruitmentcenters. All patients had clinically definite relaps-ing–remitting MS and were followed up pro-spectively for 2 or more years after onset oftherapy (Rebif®, Betaseron® or Avonex®). Rigidresponse criteria were established so as to includeonly extreme clinical-response phenotypes.Responder and nonresponder DNA pools wereprepared from groups of approximately 20 sub-jects each (total of 206 patients included), andeach DNA sample was included in three differ-ent pools to allow for technical replication onseparate Affymetrix GeneChip® 100K arrays.The screening was performed in four stages:
• Stage 1: microarray SNP typing on DNApools;
• Stage 2: ranking of top candidate SNPs byp-value significance and linkage disequilib-rium clustering, based on the notion that sig-nificant SNPs are likely to segregate inhaplotypes;
• Stage 3: validation of candidate SNPs arisingfrom the pooled-DNA approach throughindividual genotyping;
• Stage 4: increased power for association anal-ysis by inclusion of 81 more response-classifiedsubjects.
Gene ontology analysis of the stage 1 microar-ray data revealed that top-ranked SNPs withdiffering allelic distribution between respondersand nonresponders were more likely to befound in genes coding for ion channels and sig-nal-transduction pathways, for example, gluta-mate and γ-aminobutyric acid receptor genes(discussed below). In stage 2, 35 candidate
SNPs were selected, which were then typedindividually in stage 3. The degree of correla-tion of allele frequencies between the triplicatepools was very high (r2 ≥ 0.99), and pooled fre-quencies closely matched true frequenciesobtained by individual genotyping, attesting tothe success and accuracy of the DNA poolmicroarray strategy. Following stage 4 jointanalysis, significant response-associated SNPswere found in a variety of genes, including theheparan sulfate proteoglycan gene glypican 5and two ECM genes. Interestingly, these genescode for proteins known to be involved in neu-ronal repair and growth or, respectively, to betargeted for degradation by MMPs in conjunc-tion with extravasation of inflammatory leuko-cytes through basement membranes. Inaddition, the authors probed their dataset forSNPs/genes scrutinized in earlier IFN-βpharmacogenomic studies. They generally didnot confirm previously reported associations.
Discussion & implicationsThe study by Byun and colleagues [1] has yieldeda wealth of novel information on genes that arepotentially involved in the manifestation of clin-ical responses to IFN-β in MS. Some of thesegenes were already known to be functionally orbiologically linked to interferon activity. How-ever, this is much less the case for a group ofgenes that surfaced through the gene ontologyanalysis. Intriguingly, the study by Byun et al. [1]
shows top-ranked SNPs that differ betweenresponders and nonresponders in genes encodingseveral members of two major families of neuro-transmitter-gated channels. It is worthwhile toexplore the potential functional implications ofthis finding in a bit more detail.
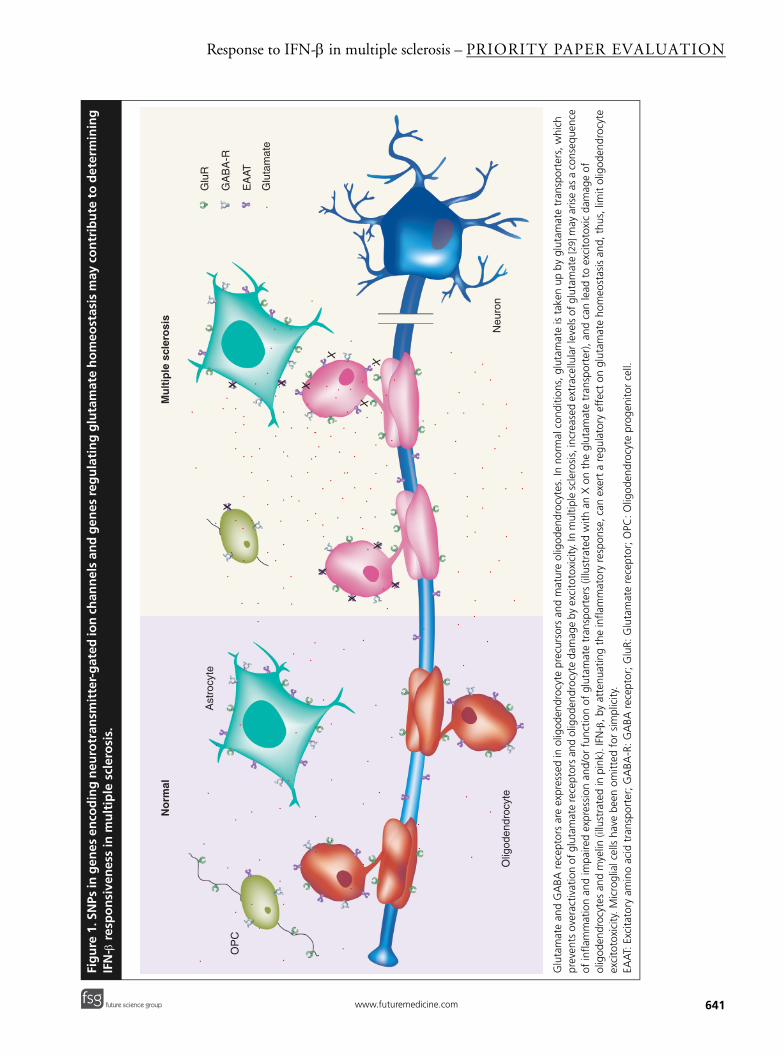
Extracellular neurotransmitter-gated chan-nels are known to be expressed by oligodendro-cytes [15,16]. These types of channel include thoseactivated by glutamate and GABA, which are themain excitatory and inhibitory neurotransmittersof the CNS. The functional significance of thesignals mediated by glutamate and GABA recep-tors are unknown, but it has been suggested thatthey may control oligodendrogenesis and myeli-nation. Strikingly, sustained activation ofglutamate receptors expressed in oligodendrocyteprecursors, as well as mature oligodendrocytes,can cause excitotoxicity, a feature that contributesto the progression of white-matter damage in MSand other pathological conditions (Figure 1)
(reviewed in [16]). In addition, differences betweenresponders and nonresponders to IFN-β in MS
Pharmacogenomics (2008) 9(5) future science groupfuture science group
Response to IFN-β in multiple sclerosis – PRIORITY PAPER EVALUATION
Fig
ure
1. S
NPs
in g
enes
en
cod
ing
neu
rotr
ansm
itte
r-g
ated
ion
ch
ann
els
and
gen
es r
egu
lati
ng
glu
tam
ate
ho
meo
stas
is m
ay c
on
trib
ute
to
det
erm
inin
g
IFN
- β r
esp
on
sive
nes
s in
mu
ltip
le s
cler
osi
s.
Glu
tam
ate
and
GA
BA r
ecep
tors
are
exp
ress
ed in
olig
oden
droc
yte
prec
urso
rs a
nd m
atur
e ol
igod
endr
ocyt
es. I
n no
rmal
con
ditio
ns, g
luta
mat
e is
tak
en u
p by
glu
tam
ate
tran
spor
ters
, whi
ch
prev
ents
ove
ract
ivat
ion
of g
luta
mat
e re
cept
ors
and
olig
oden
droc
yte
dam
age
by e
xcito
toxi
city
. In
mul
tiple
scl
eros
is, i
ncre
ased
ext
race
llula
r lev
els
of g
luta
mat
e [2
9] m
ay a
rise
as a
con
sequ
ence
of
infla
mm
atio
n an
d im
paire
d ex
pres
sion
and
/or
func
tion
of g
luta
mat
e tr
ansp
orte
rs (i
llust
rate
d w
ith a
n X
on
the
glut
amat
e tr
ansp
orte
r), a
nd c
an le
ad t
o ex
cito
toxi
c da
mag
e of
ol
igod
endr
ocyt
es a
nd m
yelin
(illu
stra
ted
in p
ink)
. IFN
-β, b
y at
tenu
atin
g th
e in
flam
mat
ory
resp
onse
, can
exe
rt a
reg
ulat
ory
effe
ct o
n gl
utam
ate
hom
eost
asis
and
, thu
s, li
mit
olig
oden
droc
yte
exci
toto
xici
ty. M
icro
glia
l cel
ls h
ave
been
om
itted
for
sim
plic
ity.
EAA
T: E
xcita
tory
am
ino
acid
tra
nspo
rter
; GA
BA-R
: GA
BA r
ecep
tor;
Glu
R: G
luta
mat
e re
cept
or; O
PC: O
ligod
endr
ocyt
e pr
ogen
itor
cell.
No
rmal
Mu
ltip
le s
cler
osi
s
Olig
oden
droc
yte
OP
C
Ast
rocy
te
Neu
ron
X
XX
X
X
X
X
Glu
R
GA
BA
-R
EA
AT
Glu
tam
ate
..
. .
.. .
.
....
.
..
..
.
.
......
.
...
.
.
.
.
.
.
..
.
.
.
.
.
.
.
..
.
..
.
..
.
...
..
...
.
..
.
.
..
..
.
.
..
.
.
..
.
.
.
.
.
.. .
.. .
..
..
. . ..
.
.
..
. ..
.
.
.
..
. .
.
.
.
..
...
..
.
.
.
.
.
.
.
..
..
.
.
.
..
.
.
..
...
..
..
..
.
.
.
. .
..
.
.
.
.
..
...
..
.
...
..
.
..
.
.
.
.
.
..
.
.
.
X
X
XX
641future science groupfuture science group www.futuremedicine.com

PRIORITY PAPER EVALUATION – Vandenbroeck & Matute
642
included genes associated with glutamate homeo-stasis, such as the excitatory amino acid trans-porter EAAT2 (SLC1A2) and GAD1 [1]. EAAT2 isa critical regulator of extracellular glutamatelevels [17], the function of which is impaired dur-ing primary or secondary CNS inflammation,thus leading to oligodendrocyte excitotoxicity [18].On the other hand, GAD1, also known asGAD67, catalyzes the production of GABA fromglutamate and, as a consequence, regulates theshift from excitation to inhibition in brain cells,including oligodendrocytes. Moreover, GAD isknown to be a major pancreatic β-cell autoanti-gen, recognized by humoral and cellular immunecomponents, which plays an important role in thepathogenesis of autoimmune Type 1 diabetes [19,20].Taken together, the findings described above indi-cate that variability in genes of the glutamate andGABA signaling systems may underlie responsive-ness to IFN-β by shifting the balance betweenelectrical excitation and inhibition of oligo-dendrocytes (Figure 1). Indeed, IFN-β can blockproinflammatory cytokine-induced inhibition ofglutamate uptake and thus regulate tissue excit-ability in cell culture [21]. If this were the case inMS patients, IFN-β-responders may be amenableto the regulation of glutamate- and GABA-relatedgenes by this cytokine.
Approximately 112 candidate genes have beenscrutinized in earlier IFN-β pharmacogenomicsstudies [8–13]. In particular, the IFNAR1 andIFNAR2 genes have received a lot of attention,with the main outcome of only limited, if any,support for a role in response [8,10,11]. In ourgroup, we sequenced the promoter regions of 100genes that contain interferon-stimulated responseelements, hallmarks of Type I interferon induc-ibility, in DNA pools of MS patients respondingand not responding to IFN-β [10]. We detected 54polymorphisms located within or close to theseinterferon-stimulated response elements. Poly-morphisms in 15 genes showed allelic distortionsbetween these pools, and four candidate geneswere retained following individual genotyping in94 responders and 68 nonresponders (MX1,IFNAR1, LMP7 and CTSS, the cathepsin S gene).Byun and colleagues [1] scrutinized their datasetfor SNPs in IFNAR1, IFNAR2, MX1, LMP7 andCTSS, but could not confirm the earlier findingsfor a potential role of these genes (with the caveatthat the analyzed SNPs were different from thosestudied before owing to design limitations of theAffymetrix 100K microarray, which is enriched forSNPs clustering around, rather than within, genes,and thus oversamples the intergenic areas [1]).
However, CTSS was equally identified as a candi-date modifier of the response to glatiramer acetatetherapy in MS [22]. Cathepsin S is a lysosomal cys-tein protease that may be involved in the pro-teolytic processing of human myelin basic protein,leading to release of immunodominant epitopes[23]. Generally, cystein proteases are known to beactive in regulating antigen presentation by bothMHC class II and CD1D [24], and are, as such, ofobvious relevance to the etiopathogenesis ofautoimmune disorders.
The only gene simultaneously, althoughweakly, arising as a candidate response modifierfrom both the Byun [1] and Cunningham [10]
studies is the interferon-inducible dsRNA-dependent protein kinase (PRKR). A promoterPRKR CCGn repeat emerged in the pooled DNAstage [10], while an SNP in the neighboring regionshowed up significantly in the study by Byun et al.[1]. Several reasons may account for this generallack of overlap. It is of relevance to have a closerlook at the differences and similarities in thedesigns of both of these studies, as these maypoint to some of the major challenges to beaddressed if future MS pharmacogenomics studiesare to deliver. First, the criteria for the definitionof (non)response differ between both studies(Table 1). Therefore, as the (non)responder stratain each of both studies do not represent the samephenotypic trait, egression of de facto shared phar-macogenomic determinants may have beenobscured. Second, Byun and colleagues [1]
recruited patients from the Mediterranean region,while the Cunningham study [10] focused onNorthern Irish patients. Well-documented differ-ences in northern versus southern European pop-ulation substructure [25], and in the north–southprevalence [26] and, potentially, genetics of MS [27],together incite the question whether Europeanpopulation substructure may impact upon thedistribution or epiphany of genetic IFN-βresponse determinants. Indeed, subethnic differ-ences in the pharmacogenetics of important poly-morphisms have been documented. For example,within a group of asthma patients of Hispanicdescent, a strong association was found betweengreater bronchodilator drug responsiveness andthe β2 adrenergic receptor Arg16 genotype inPuerto Ricans, which was absent in Mexicans [28].Third, as already indicated above, the microarraysused by Byun et al. [1] did not contain the samepromoter SNPs as those analyzed in our study [10].Thus, the response-associated genes of the Cun-ningham study [10] may have been missed if thecontributing SNPs happened not to be part of a
Pharmacogenomics (2008) 9(5) future science groupfuture science group

Response to IFN-β in multiple sclerosis – PRIORITY PAPER EVALUATION
future science groupfuture science group
Table 1. Clinical IFN-
Study Respon
Byun et al. (2008)
No relapNo incre
Cunningham et al. (2005)
RelapseNo sustaor ≥0.5
*In both studies, patients wfirst 6–9 months of treatmEDSS: Expanded disability s
haplotype tagged in the Byun study [1]. Fourth,data collection and analysis in each study wasbased on a limited number of response-classifiedpatients (<300 and <200 for [1] and [10], respec-tively). Given the lack of correction for multiplecomparisons in both studies, some of the associa-tions found may simply represent false positives,while the power to detect effects conferred byweak response modifiers or SNPs with low minorallele frequency may have been lacking.
Future perspectiveDeciphering the pharmacogenomics of IFN-βtreatment in MS has proved to be a challengeimposed upon the molecular biology, neuro-immunology and biocomputing research com-munities. Not only is MS a complex genetictrait, but the manifestation of clinical benefits inresponse to IFN-β seems to be dependent ongenetic variation in multiple gene loci [1,10,14].Thus, low penetrance, gene–gene and/orgene–environment interactions, as well as lim-ited statistical power may, together, jeopardizethe systematic identification of treatment-response genes. In addition, there is considerablevariation in the range of clinical and paraclini-cal criteria (expanded disability status scale,MRI, relapse frequency, neutralizing antibodiesand so on) currently used to denote the MSpatients’ response status (see, for example, [3]).In the absence of an objective and easilyquantifiable response (bio)marker, standardiza-tion of response criteria, based on internationalconsensual definitions, will be of crucial impor-tance in order to facilitate meaningful perform-ance and comparison of MS drug treatmentpharmacogenomics studies. Current IFN-βpharmacogenomics studies typically includeonly those patients either responding (very) wellor not (at all) to the therapy, and patients with anintermediate-response profile are excluded fromthe screen. Implementation of ‘extreme’ responsecriteria is likely to maximize the separation
distance of response phenotypes, and hopefully,of underlying genetic factors, but necessarilyimplies that the proportion of patients that canbe included in the study is small. The study byByun and colleagues exemplifies this approach:in total, only 285 response-classified patientscould be recruited from four collaboratingcenters [1]. Therefore, well-organized intercenterand international consortia are the sine qua nonof sufficiently powered MS pharmacogenomicsstudies. A more pronounced commitment ofpharmaceutical companies to make samples oftheir clinical trials available to pharmacogenomicstudies is crucial. Only on the condition that theabove requirements are met is it likely thatpharmacogenomics research in MS will yielddata that can be validated in a purposeful man-ner and that, ultimately, may be of translationalrelevance. In the framework of such improvedmise en scène, the use of 500K–1000K SNParrays logically will provide better coverage of thegenome at higher density and, in conjunctionwith adequate bioinformatics approaches andextensive clinical and demographic data, willallow for exhaustive analysis of gene interactions.In conclusion, the study by Byun and colleagues[1] embodies a significant step forward in ourunderstanding of not only the genetics of IFN-βresponse heterogeneity, but also of both the limi-tations inherent to the current settings of MSpharmacogenomics research and the challengesthat lie ahead.
Financial & competing interests disclosure The authors have no relevant affiliations or financialinvolvement with any organization or entity with a finan-cial interest in or financial conflict with the subject matteror materials discussed in the manuscript. This includesemployment, consultancies, honoraria, stock ownership oroptions, expert testimony, grants or patents received orpending or royalties.
No writing assistance was utilized in the production ofthis manuscript.
β responder and nonresponder phenotypes*.
der Nonresponder Ref.
ses andase in EDSS
At least two relapses orAn increase in EDSS of at least 1 point
[1]
rate reduced by a third and ined progression in EDSS (≥1 point if baseline EDSS <5.5
points if baseline EDSS ≥5.5)
Relapse rate is the same or increases [10]
ere followed for 2 years after initiation of treatment. In Cunningham et al. [10] , response status was not judged in the ent, the rationale being that treatment may not be effective until after this period.tatus scale.
643www.futuremedicine.com
PRIORITY PAPER EVALUATION – Vandenbroeck & Matute
BibliographyPapers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.1. Byun E, Caillier SJ, Montalban X et al.:
Genome-wide pharmacogenomic analysis of the response to interferon β therapy in multiple sclerosis. Arch. Neurol. 65(3), 337–344 (2008).
2. The IFNB Multiple Sclerosis Study Group: Interferon β-1b is effective in relapsing–remitting multiple sclerosis. Neurology 43(4), 655–661 (1993).
3. Goertsches R, Serrano-Fernández P, Moeller S, Koczan D, Zettl UK: Multiple sclerosis therapy monitoring based on gene expression. Curr. Pharm. Des. 12(29), 3761–3779 (2006).
•• Excellent discussion of the challenges relating to the identification and use of multiple sclerosis (MS) drug-response markers.
4. Lindberg RL, Kappos L: Transcriptional profiling of multiple sclerosis: towards improved diagnosis and treatment. Expert Rev. Mol. Diagn. 6(6), 843–855 (2006).
5. Fernald GH, Knott S, Pachner A et al.: Genome-wide network analysis reveals the global properties of IFN-β immediate transcriptional effects in humans. J. Immunol. 178(8), 5076–5085 (2007).
• Innovative study investigating gene regulation by IFN-β using network analysis. Discussion of the relevance of this approach for personalized medicine.
6. Alexander JS, Minagar A, Harper M, Robinson-Jackson S, Jennings M, Smith SJ: Proteomic analysis of human cerebral endothelial cells activated by multiple sclerosis serum and IFNβ-1b. J. Mol. Neurosci. 32(3), 169–178 (2007).
7. Kantor AB, Deng J, Waubant E et al.: Identification of short-term pharmacodynamic effects of interferon-β-1a in multiple sclerosis subjects with broad-based phenotypic profiling. J. Neuroimmunol. 188(1–2), 103–116 (2007).
8. Sriram U, Barcellos LF, Villoslada P et al.: Pharmacogenomic analysis of interferon receptor polymorphisms in multiple sclerosis. Genes Immun. 4(2), 147–152 (2003).
9. Fernández O, Fernández V, Mayorga C et al.: HLA class II and response to interferon-β in multiple sclerosis. Acta Neurol. Scand. 112(6), 391–394 (2005).
10. Cunningham S, Graham C, Hutchinson M et al.: Pharmacogenomics of responsiveness to interferon IFN-β treatment in multiple sclerosis: a genetic screen of 100 type I interferon-inducible genes. Clin. Pharmacol. Ther. 78(6), 635–646 (2005).
• Analysis of the pharmacogenomics of IFN-β in MS based on the identification of polymorphisms in 100 Type I interferon-inducible genes. Four genes emerged as potential response modifiers: IFNAR1, MX1, LMP7 and CTSS.
11. Leyva L, Fernández O, Fedetz M et al.: IFNAR1 and IFNAR2 polymorphisms confer susceptibility to multiple sclerosis but not to interferon-β treatment response. J. Neuroimmunol. 163(1–2), 165–171 (2005).
12. Martínez A, de las Heras V, Mas Fontao A et al.: An IFNG polymorphism is associated with interferon-β response in Spanish MS patients. J. Neuroimmunol. 173(1–2), 196–199 (2006).
13. Weinstock-Guttman B, Tamaño-Blanco M, Bhasi K, Zivadinov R, Ramanathan M: Pharmacogenetics of MXA SNPs in interferon-β treated multiple sclerosis patients. J. Neuroimmunol. 182(1–2), 236–239 (2007).
14. O’Doherty C, Villoslada P, Vandenbroeck K: Pharmacogenomics of type I interferon therapy: a survey of response-modifying genes. Cytokine Growth Factor Rev. 18(3–4), 211–222 (2007).
• Review of genes potentially involved in bringing about response to type I interferon therapy (IFN-β in MS and IFN-α in hepatitis).
15. Kettenmann H, Steinhauser C: Receptors for neurotransmitters and hormones. In: Neuroglia (2nd Edition). Kettenmann H, Ransom BR (Eds.). Oxford University Press, Oxford, UK, 131–145 (2005).
16. Matute C, Domercq M, Sánchez-Gómez MV: Glutamate-mediated glial injury: mechanisms and clinical importance. Glia 53(2), 212–224 (2006).
Executive summary
Study design
• DNA samples from a total of 206 IFN-β-response-classified multiple sclerosis (MS) patients from four collaborating centers were screened using the Affymetrix 100K GeneChip® Array (DNA-pooling approach).
Analyses performed
• Stage 1: microarray SNP typing on DNA pools. • Stage 2: ranking of top candidate SNPs by p-value significance and linkage disequilibrium clustering.• Stage 3: validation of candidate SNPs arising from the pooled DNA approach through individual genotyping. • Stage 4: increased power for association analysis by inclusion of 81 more response-classified subjects.
Results
• Stage 1: Gene ontology analysis revealed response-associated top-ranked SNPs in genes coding for ion channels and signal transduction pathways.
• Stage 2: 35 candidate SNPs selected and typed individually. • Stage 3: successful validation of association between response and candidate SNPs. • Stage 4: joint analysis yields 18 significant SNPs.
Implications & perspectives
• Response to IFN-β in MS behaves as a complex genetic trait.• Several genes and gene families arising from the study were not clearly known before to be functionally linked to
interferon activity.• Little overlap with earlier studies potentially owing to differences in selected SNPs and study design.• Increased power and consensus on drug response in MS is needed to facilitate further studies.
644 Pharmacogenomics (2008) 9(5) future science groupfuture science group

Response to IFN-β in multiple sclerosis – PRIORITY PAPER EVALUATION
• Summarizes current knowledge on the role of glutamate signaling in glial cell death. This is of relevance to the findings by Byun and colleagues [1], who identified neurotransmitter-gated channel genes as one of the main gene ontology groups associated with response to IFN-β.
17. Danbolt NC: Glutamate uptake. Prog. Neurobiol. 65(1), 1–105 (2001).
18. Domercq M, Sánchez-Gómez MV, Sherwin C, Etxebarria E, Fern R, Matute C: System xc- and glutamate transporter inhibition mediates microglial toxicity to oligodendrocytes. J. Immunol. 178(10), 6549–6556 (2007).
19. Tisch R, Yang XD, Singer SM, Liblau RS, Fugger L, McDevitt HO: Immune response to glutamic acid decarboxylase correlates with insulitis in non-obese diabetic mice. Nature 366(6450), 72–75 (1993).
20. Zekzer D, Wong FS, Ayalon O et al.: GAD-reactive CD4+ Th1 cells induce diabetes in NOD/SCID mice. J. Clin. Invest. 101(1), 68–73 (1998).
21. Hu S, Sheng WS, Ehrlich LC, Peterson PK, Chao CC: Cytokine effects on glutamate uptake by human astrocytes. Neuroimmunomodulation 7(3), 153–159 (2000).
22. Grossman I, Avidan N, Singer C et al.: Pharmacogenetics of glatiramer acetate therapy for multiple sclerosis reveals drug-responsive markers. Pharmacogenet. Genomics 17(8), 657–666 (2007).
23. Beck H, Schwarz G, Schröter CJ et al.: Cathepsin S and an asparagine-specific endoprotease dominate the proteolytic processing of human myelin basic protein in vitro. Eur. J. Immunol. 31(12), 3726–3736 (2001).
24. Honey K, Rudensky AY: Lysosomal cysteine proteases regulate antigen presentation. Nat. Rev. Immunol. 3(6), 472–482 (2003).
25. Seldin MF, Shigeta R, Villoslada P et al.: European population substructure: clustering of northern and southern populations. PLoS Genet. 2(9), E143 (2006).
26. McDonell GV, Hawkins SA: Multiple sclerosis in Northern Ireland: a historical and global perspective. Ulster Med. J. 69(2), 97–105 (2000).
27. Rosati G: The prevalence of multiple sclerosis in the world: an update. Neurol. Sci. 22(2), 117–139 (2001).
28. Choudhry A, Ung N, Avila PC et al.: Pharmacogenetic differences in response to albuterol between Puerto Ricans and Mexicans with asthma. Am. J. Respir. Crit. Care Med. 171(6), 563–570 (2005).
29. Srinivasan R, Sailasuta N, Hurd R, Nelson S, Pelletier D: Evidence of elevated glutamate in multiple sclerosis using magnetic resonance spectroscopy at 3 T. Brain 128(Pt 5), 1016–1025 (2005).
645future science groupfuture science group www.futuremedicine.com


















