
NNK-induced hamster lung adenocarcinomas over-express β2-adrenergic and EGFR signaling pathways
11
Lung Cancer (2005) 49, 35—45 NNK-induced hamster lung adenocarcinomas over-express 2 -adrenergic and EGFR signaling pathways Hildegard M. Schuller ∗ , Maria Cekanova Experimental Oncology Laboratory, Department of Pathobiology, College of Veterinary Medicine, University of Tennessee, 2407 River Drive, Knoxville, TN 37996, USA Received 13 August 2004; received in revised form 6 December 2004; accepted 13 December 2004 KEYWORDS NNK hamster; 2 -Adrenergic; EGFR Summary Pulmonary adenocarcinma (PAC) is the most common type of human lung cancer. A diagnosis of PAC, history of non-smoking and presence of mutations in the EGFR are predictive factors for responsiveness of lung cancer to EGFR-specific tyrosine kinase inhibitors. Unfortunately, less than 50% of PAC cases demonstrate this mutation-based responsiveness. Our immunohistochemical analysis of NNK-induced PAC in hamsters demonstrates the simultaneous over-expression of a 2 -adrenergic receptor pathway, including PKA, cAMP, CREB and phosphorylated CREB and of an EGFR pathway, including over- expression of EGFR-specific phosporylated tyrosine kinase, Raf-1 and ERK1/2 and their phosphorylated forms. These findings implicate, for the first time, PKA/CREB- mediated signaling in the development and regulation of any type of lung cancer. In light of reports that NNK acts as a -adrenergic agonist and that -blockers inhibit the growth of PAC of Clara cell lineage in the NNK hamster model and in human cancer cell lines from smokers, our current data suggest transactivation of the EGFR pathway via -adrenergic signaling as a novel regulatory mechanism in a subpopu- lation of PACs in smokers. Taken together, these data point to PKA/CREB as novel targets for the development of cancer therapeutics for PAC patients non-responsive to EGFR-specific tyrosine kinase inhibitors. © 2004 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Pulmonary adenocarcinoma (PAC) is the leading type of lung cancer today [1—3] and accounts for * Corresponding author. Tel.: +1 865 974 8217; fax: +1 865 974 5616. E-mail address: [email protected] (H.M. Schuller). about 60% of the family of non-small cell lung cancer (NSCLC). Smoking [2,3] and a high fat diet that increase the systemic levels of arachi- donic acid (AA) are documented risk factors for the development of PAC [4,5]. Moreover, the AA- metabolizing enzymes cyclooxygenase-2 (COX-2) and 5-lipoxygenase (5-LOX) are particularly often over-expressed in this cancer type [6—8]. The epi- 0169-5002/$ — see front matter © 2004 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.lungcan.2004.12.012
Transcript of NNK-induced hamster lung adenocarcinomas over-express β2-adrenergic and EGFR signaling pathways

Lung Cancer (2005) 49, 35—45
NNK-induced hamster lung adenocarcinomasover-express �2-adrenergic and EGFRsignaling pathways
Hildegard M. Schuller∗, Maria Cekanova
Experimental Oncology Laboratory, Department of Pathobiology, College of Veterinary Medicine,University of Tennessee, 2407 River Drive, Knoxville, TN 37996, USA
Received 13 August 2004; received in revised form 6 December 2004; accepted 13 December 2004
KEYWORDSNNK hamster;�2-Adrenergic;EGFR
Summary Pulmonary adenocarcinma (PAC) is the most common type of humanlung cancer. A diagnosis of PAC, history of non-smoking and presence of mutations inthe EGFR are predictive factors for responsiveness of lung cancer to EGFR-specifictyrosine kinase inhibitors. Unfortunately, less than 50% of PAC cases demonstratethis mutation-based responsiveness.Our immunohistochemical analysis of NNK-induced PAC in hamsters demonstrates
the simultaneous over-expression of a �2-adrenergic receptor pathway, includingPKA, cAMP, CREB and phosphorylated CREB and of an EGFR pathway, including over-expression of EGFR-specific phosporylated tyrosine kinase, Raf-1 and ERK1/2 andtheir phosphorylated forms. These findings implicate, for the first time, PKA/CREB-mediated signaling in the development and regulation of any type of lung cancer. Inlight of reports that NNK acts as a �-adrenergic agonist and that �-blockers inhibitthe growth of PAC of Clara cell lineage in the NNK hamster model and in humancancer cell lines from smokers, our current data suggest transactivation of the EGFRpathway via �-adrenergic signaling as a novel regulatory mechanism in a subpopu-lation of PACs in smokers. Taken together, these data point to PKA/CREB as noveltargets for the development of cancer therapeutics for PAC patients non-responsiveto EGFR-specific tyrosine kinase inhibitors.© 2004 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Pulmonary adenocarcinoma (PAC) is the leading
about 60% of the family of non-small cell lungcancer (NSCLC). Smoking [2,3] and a high fatdiet that increase the systemic levels of arachi-
type of lung cancer today [1—3] and accounts for
* Corresponding author. Tel.: +1 865fax: +1 865 974 5616.
E-mail address: [email protected] (H
donic acid (AA) are documented risk factors for
0169-5002/$ — see front matter © 200doi:10.1016/j.lungcan.2004.12.012
974 8217;
.M. Schuller).
the development of PAC [4,5]. Moreover, the AA-metabolizing enzymes cyclooxygenase-2 (COX-2)and 5-lipoxygenase (5-LOX) are particularly oftenover-expressed in this cancer type [6—8]. The epi-
4 Elsevier Ireland Ltd. All rights reserved.

36 H.M. Schuller, M. Cekanova
dermal growth factor receptor (EGFR) and its asso-ciated tyrosine kinases are thought to play an im-portant role in the regulation of cell proliferation,apoptosis and the over-expressed AA-cascade ob-served in PAC [9,10].
Smoking and a high fat diet are also risk factorsfor cardiovascular disease [11—13]. �-Adrenergicreceptors have long been recognized as key reg-ulators of cardiovascular function and antagonistsof these receptors (�-blockers) are widely used forthe treatment of cardiovascular disease [14]. �-Adrenergic receptors also regulate the release of AAin the heart and blood vessels, thus elevating thelevels of AA metabolites that contribute to the de-velopment of atherosclerosis [14]. Because of this,the COX inhibitor aspirin is often used for the treat-ment of acute heart attacks and for the preventionof cardiovascular disease [15].
Because of the similarities between cardiovas-cular disease and PAC, we started to explore a po-tential role of �-adrenergic signaling in the devel-opment and growth regulation of PAC. We foundthat DNA synthesis of cell lines derived from hu-man PAC of Clara cell lineage is stimulated by �-adrenergic agonists that increased the release of
studied. The cAMP/PKA/CREB pathway plays an im-portant role in the development and tumorigenesisof endocrine tissues [29] but has not been impli-cated in the development of other types of cancer.In fact, the growth of cell lines derived from humansmall cell lung carcinoma (SCLC) [30], a neuroblas-toma [31] or from a large cell carcinoma [32] wasinhibited by agents that increase intracellular cAMP.
The current immunohistochemical analysis ofNNK-induced hamster PAC of Clara cell lineage pro-vides first evidence in support of the hypothesisthat pathways signaling via �2-adrenergic recep-tors/PKA/CREB as well as EGFRs/RAF/ERK1/2 mayjointly regulate the growth of this cancer type.
2. Materials and methods
This study was reviewed and approved by theInstitutional Animal Care and Concerns Commit-tee. Male outbred Syrian golden hamsters werepurchased from Charles River (Wilmington, DL) andhoused five per cage in plastic shoebox-type cagesunder standard laboratory conditions. The animalshFntdaibTdetpSa�Psfapif3lGrtcwd
AA [16]. These effects were simulated by forskolin,which activates the �-adrenergic second messengercyclic adenosine 3′5′-monophosphate (cAMP), whilean inhibitor of cAMP blocked these effects [16].Moreover, we showed that the tobacco-specific car-cinogenic nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) simulated these growthstimulating effects in PAC by acting as a high affin-ity agonist for �-adrenergic receptors [16]. NNK alsoincreased apoptosis in vascular endothelial cells via�-adrenergic signalling [17]. Studies in a hamstermodel of NNK-induced PAC of Clara cell lineagedemonstrated that tumor incidence and multiplic-ity were significantly reduced by the �-blocker pro-pranolol or the COX-inhibitor aspirin whereas the�-adrenergic agonist epinephrine or forskolin werestrong promoters of carcinogenesis [18]. Taken to-gether, these data point to an important role of�-adrenergic cAMP-dependent signaling and of theAA-cascade in the genesis and growth regulation ofClara cell-derived PAC.
The predominate signaling pathway activatedby G-protein coupled receptors such as the �-adrenergic receptors includes the activation ofadenylyl cyclase, resulting in an increase in in-tracellular cAMP and activation of protein kinaseA (PKA). In turn, PKA phosporylates the cAMP re-sponse element binding protein (CREB) [19—22].CREB is a nuclear transcription factor whose rolein cardiovascular disease [23—25], neurodegenera-tive disease [26] and stroke [27,28] has been widely
ad free access to food (Purina Rodent Chow, Agrieed, Knoxville, TN) and tap water. Pulmonary ade-ocarcinomas were induced in 10 hamsters by mul-iple subcutaneous injections of NNK as previouslyescribed [18,33]. Five untreated hamsters serveds controls. The animals were killed by isofluorenenhalation. Lung tissues were fixed in 10% neutraluffered formalin and embedded in paraffin.issue sections were subjected to immunostains asescribed below using the primary antibodies andxposure times listed in Table 1. Sections subjectedo identical procedures without exposure to therimary antibodies served as negative controls.ections from untreated hamster pancreas serveds positive controls for antibodies that detect the2-adrenergic receptor, its phosphorylated form,KA, CREB and phosporylated CREB. Deparrafinizedections were rinsed in distilled water, then PBS,ollowed by incubation with 3% H2O2 in 50% MetOHt room temperature for 20min. Incubation torimary antibodies at the concentrations andncubation conditions detailed in Table 1 wasollowed by two rinses in PBS and incubation for0min at room temperature with super sensitiveink biotynilated anti-rabbit immunoglobulin (Bio-enex, San Ramon, CA). This was followed by twoinses with PBS and incubation for 30min at roomemperature with super sensitive label-peroxydaseonjugated streptavidin (BioGenex). The slidesere then rinsed twice with PBS incubated withiaminobenzydine solution (Vector, Burlingame,

NNK-induced adenocarcinomas and EGFR signaling pathways 37
Table 1 Primary antibodies used for immunostains of NNK-induced hamster lung tumors
Primary Antibody Dilution Exposure Vendor
Anti-�2-AR, rabbit polyclonalrecognizing the carboxy terminusof the mouse and rat �2-AR
1:25 Overnight at 4 ◦C Santa Cruz Biotechnology, Santa Cruz, CA
Anti-phospho-�2-AR, rabbitpolyclonal recognizingphosphorylated Ser-355/Ser 356of human �2-AR
1:25 Overnight at 4 ◦C Santa Cruz Biotechnology
Anti-PKARII, mouse monoclonalrecognizing the regulatorysubunit R type II of human, dog,rat, mouse PKA
1:100 20min at 20 ◦C BD Transduction Laboratories, San Jose, CA
Anti-CREB, rabbit polyclonalrecognizing total human, mouse,rat, pig and bovine CREB protein
1: 25 Overnight at 4 ◦C Upstate, Lake Placid, NY
Anti-phospho-CREB, mousemonoclonal recognizing human,mouse and rat CREBphosphorylated at Ser 133 andphosphorylated ATF-1
1:25 Overnight at 4 ◦C Cell Signaling Technology, Beverly, MA
Anti-EGFR, rabbit polyclonalrecognizing total human, mouse,rat EGFR protein
1:50 Overnight at 4 ◦C Cell Signaling Technology
Anti-phospho-EGFR, rabbitpolyclonal recognizing human,mouse, rat and chicken EGFRphosporylated at tyrosine 1173
1:25 Overnight at 4 ◦C Sigma, St Louis, MO
Anti-Raf-1, rabbit polyclonalrecognizing the carboxy terminusof human, mouse and rat Raf-1p74 protein
1:25 Overnight at 4 ◦C Santa Cruz Biotechnology
Anti-p44/42 MAP kinase, rabbitpolyclonal recognizing totalhuman, rat, mouse, hamster andchicken ERK1/2 protein
1:25 Overnight at 4 ◦C Cell Signaling Technology
Anti-phospho-p44/42 MAP kinase,rabbit polyclonal recognizinghuman, mouse, rat, and hamsterERK1/2 phosphorylated atthreonine 202 and tyrosine 204 orrat ERK2 phosphorylated atthreonine 183 or tyrosine 185
1:10 Overnight at 4 ◦C Cell Signaling Technology
Anti-COX-2, rabbit polyclonalrecognizing amino acids 50—111of the amino terminus of human,mouse, and rat COX -2
1:25 Overnight at 4 ◦C Santa Cruz Biotechnology
Anti-5-LOX, rabbit polyclonalrecognizing amino acids 130—149of human, rat, mouse andhamster 5-LOX
1:50 Overnight at 4 ◦C Cayman Chemical, Ann Arbor, MI
Anti-CC10 (H-75), rabbit polyclonalrecognizing amino acids 17—91 ofhuman CC10 Clara cell protein
1:25 Overnight at 4 ◦C Santa Cruz Biotechnology
38 H.M. Schuller, M. Cekanova
CA) for 7min at room temperature, rinsed in PBS,then distilled water and counterstained with hema-toxylin (Richard Allen Scientific, Kalamazoo, MI).Densitometric analysis of immunostains was per-formed with the NIH Scion Image analysis software.Following background subtraction, mean densityof tumor tissue versus normal lung tissue was as-sessed for significant differences by one way ANOVA
followed by Tukey—Kramer multiple comparisontest.
3. Results
All NNK-treated hamsters used in this study hadmultiple lung tumors (three to six per animal). In
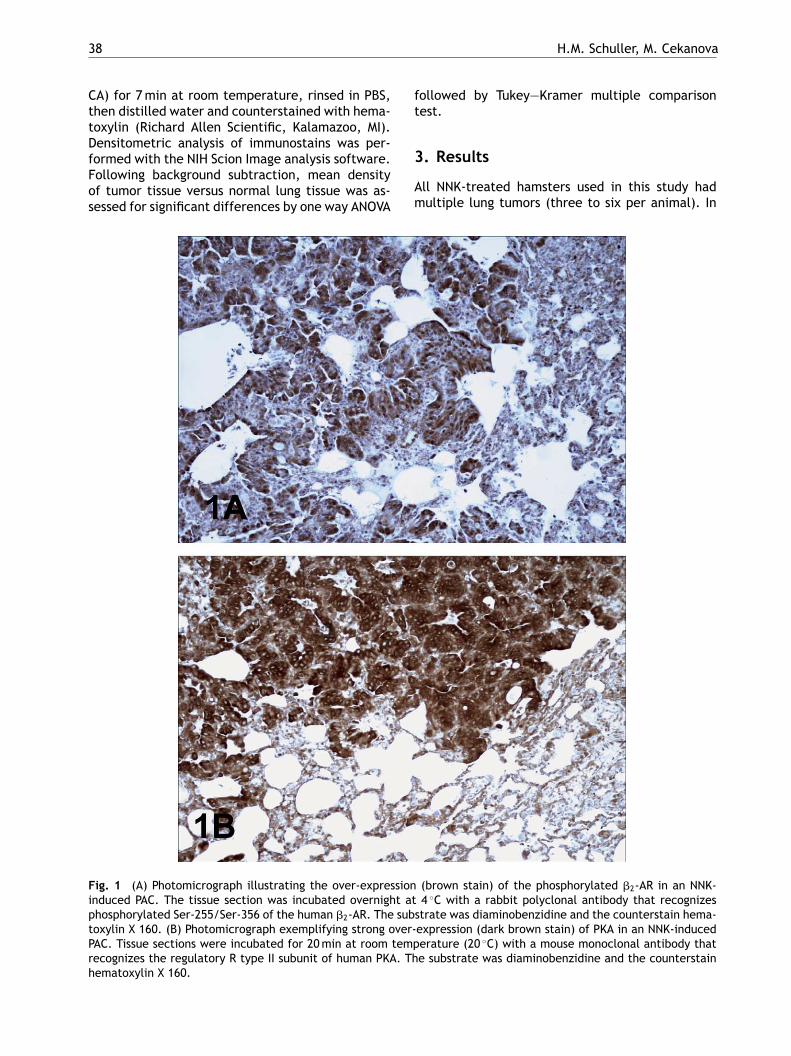
Fig. 1 (A) Photomicrograph illustrating the over-expressioninduced PAC. The tissue section was incubated overnight atphosphorylated Ser-255/Ser-356 of the human �2-AR. The substoxylin X 160. (B) Photomicrograph exemplifying strong over-PAC. Tissue sections were incubated for 20min at room temprecognizes the regulatory R type II subunit of human PKA. Thhematoxylin X 160.
(brown stain) of the phosphorylated �2-AR in an NNK-4 ◦C with a rabbit polyclonal antibody that recognizestrate was diaminobenzidine and the counterstain hema-expression (dark brown stain) of PKA in an NNK-inducederature (20 ◦C) with a mouse monoclonal antibody thate substrate was diaminobenzidine and the counterstain

NNK-induced adenocarcinomas and EGFR signaling pathways 39
Fig. 2 (A) Photomicrograph demonstrating strong over-expression (dark brown stain) of CREB protein in an NNK-induced PAC. The tissue section was incubated overnight at 4 ◦C with a rabbit polyclonal antibody that recognizesthe total human CREB protein. The substrate was diaminobenzidine and the counterstain hematoxylin X 160. (B)Photomicrograph showing strong and highly selective over-expression (dark brown stain) of phosphorylated CREB in anNNK-induced PAC. The tissee section was incubated overnight at 4 ◦C with a mouse monoclonal antibody that recognizeshuman CREB phosphorylated at Ser 133 and human phosphorylated ATF-1. The substrate was diaminobenzidine andthe counterstain hematoxylin X 160.
support of our earlier report [33] that had iden-tified Clara cells of small airway epithelium asthe origin of NNK-induced PAC in hamsters, alllung tumors demonstrated positive immunoreactiv-ity for the Clara cell-specific CC10 protein (datanot shown). Immunoreactivity to �1-adrenergic re-ceptors was detectable but not over-expressed intumor cells compared with surrounding normal air-
way epithelia and alveolar epithelia of NNK-treatedor of untreated control hamsters (data not shown).By contrast, slides incubated with a polyclonal an-tibody that recognizes the carboxy terminus ofthe �2-adrenergic receptor (�2-AR) or with a poly-clonal antibody that recognizes phosphorylation atSer-355/Ser-356 of the �2-AR demonstrated a pro-nounced increase in immunoreactivity of tumor

40 H.M. Schuller, M. Cekanova
Fig. 3 (A) Photomicrograph exhibiting strong and highly selective over-expression (dark brown stain) of phosphorylatedEGFR-specific tyrosine kinase in an NNK-induced PAC. The tissue section was incubated overnight at 4 ◦C with a rabbitpolyclonal antibody that recognizes human, mouse and rat EGFR phosphorylated at tyrosine 1173. The substrate wasdiaminobenzidine and the counterstain hematoxylin X 160. (B) Photomicrograph demonstrating strong and selectiveover-expression (dark brown stain) of Raf-1 in an NNK-induced PAC. The tissue section was incubated overnight at 4 ◦Cwith a rabbit polyclonal antibody that recognizes the carboxy terminus of human Raf-1 p74 protein. The substrate wasdiaminobenzidine and the counterstain hematoxylin X 160.
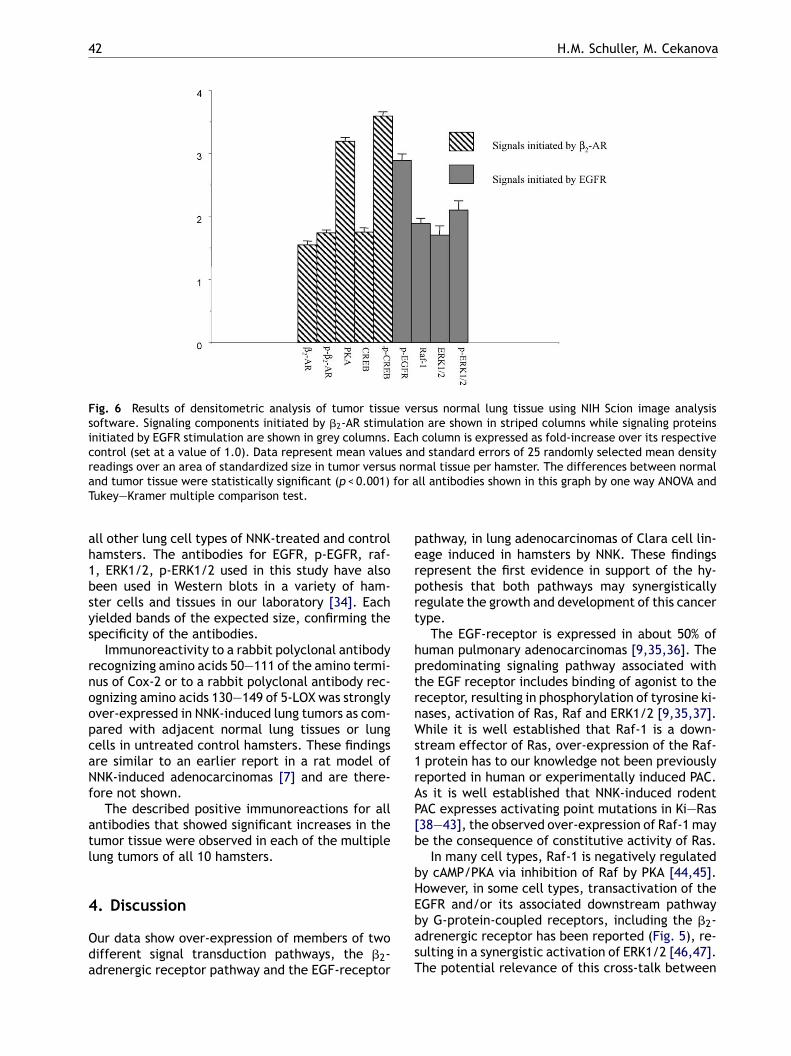
cells (1.6- and 1.74-fold, respectively) as comparedwith adjacent normal airway or alveolar epithelia(Figs. 1a and 5). Similarly, immunoreactivity to amonoclonal antibody that recognizes the regulatorysubunit R type II of PKA was strongly over-expressedin tumor cells of NNK-treated hamsters (3.19-fold;Figs. 1b and 5). Immunoreactivity to an antibody
that recognizes total CREB protein was suggestiveof strong over-expression of the protein in NNK-induced lung tumors (1.75-fold) as compared withthe surrounding lung parenchyma (Figs. 2a and 5).Interestingly, strong positive immunoreactivity toa monoclonal antibody that detects CREB phos-phorylated at Ser 133 and phosphorylated ATF-

NNK-induced adenocarcinomas and EGFR signaling pathways 41
Fig. 4 Photomicrograph illustrating selective over-expression (brown stain) of phosphorylated ERK1/2 in an NNK-induced PAC. The tissue section was incubated overnight at 4 ◦C with a rabbit polyclonal antibody that recognizeshuman ERK1/2 phosphorylated at threonine 202 and tyrosine 204 or rat ERK2 phosphorylated at threonine 183 ortyrosine 185. The substrate was diaminobenzidine and the counterstain hematoxylin X 160.
1was found exclusively in NNK-induced lung tumors(Figs. 2a and 5) whereas all other lung cells inNNK-treated or untreated control animals demon-strated negative reactions with densitometry lev-els at background values under the conditions used.Pancreatic islet cells whose secretion is regulatedby �2-ARs and their associated PKA/CREB pathwayshowed positive immunoreactivity to the �2-AR, p-�2-AR, PKA and CREB, supporting the specificity ofthe immunoreactions observed in the lung tumors.
Fbtm
Exposure of slides to a rabbit polyclonal antibodythat recognizes the total EGFR protein resulted inpositive immunoreactivity in all tumor cells. How-ever, this reaction was not more intense than inother normal appearing lung cells such as airwayor alveolar epithelia of NNK-treated hamsters oruntreated controls (data not shown). By contrast,slides that had been incubated with a rabbit poly-clonal antibody that recognizes EGFR phospory-lated at tyrosine 1173 demonstrated strong (2.89-fold)and selective positive immunoreactivity in thetumor tissue (Figs. 3a and 5) while being at back-ground levels in surrounding normal lung tissue orin the lungs of control hamsters. Similarly, expo-sure of slides to a rabbit polyclonal antibody thatrecognizes the carboxy terminus of Raf-1 p74 pro-tein yielded strong (1.89-fold) and selective posi-tive immunoreactivity in NNK-induced lung tumors(Figs. 3b and 5) whereas expression levels were atbackground levels in normal appearing other lungcells of NNK-treated hamsters or in lung tissuesof untreated controls. Over-expressed immunore-activity to a rabbit polyclonal antibody that de-tects total ERK1/2 protein (1.7-fold) or to a rabbitpolyclonal antibody that recognizes ERK1/2 phos-p2nsi
ig. 5 Diagram illustrating the hypothetical cross-talketween �2-AR and EGFR-initiated regulatory signalransduction pathways in NNK-induced hamster pul-onary adenocarcinomas.
horylated at human threonine 202 and tyrosine04 of ERK or rat ERK2 phosphorylated at threo-ine 183 or tyrosine 185 was strong (2.1-fold) andelective in NNK-induced lung tumors (exemplifiedn Figs. 4 and 5) while being at background levels in
42 H.M. Schuller, M. Cekanova
Fig. 6 Results of densitometric analysis of tumor tissue versus normal lung tissue using NIH Scion image analysissoftware. Signaling components initiated by �2-AR stimulation are shown in striped columns while signaling proteinsinitiated by EGFR stimulation are shown in grey columns. Each column is expressed as fold-increase over its respectivecontrol (set at a value of 1.0). Data represent mean values and standard errors of 25 randomly selected mean densityreadings over an area of standardized size in tumor versus normal tissue per hamster. The differences between normaland tumor tissue were statistically significant (p < 0.001) for all antibodies shown in this graph by one way ANOVA andTukey—Kramer multiple comparison test.
all other lung cell types of NNK-treated and controlhamsters. The antibodies for EGFR, p-EGFR, raf-1, ERK1/2, p-ERK1/2 used in this study have alsobeen used in Western blots in a variety of ham-ster cells and tissues in our laboratory [34]. Eachyielded bands of the expected size, confirming thespecificity of the antibodies.
Immunoreactivity to a rabbit polyclonal antibodyrecognizing amino acids 50—111 of the amino termi-nus of Cox-2 or to a rabbit polyclonal antibody rec-ognizing amino acids 130—149 of 5-LOX was stronglyover-expressed in NNK-induced lung tumors as com-pared with adjacent normal lung tissues or lungcells in untreated control hamsters. These findingsare similar to an earlier report in a rat model ofNNK-induced adenocarcinomas [7] and are there-fore not shown.
The described positive immunoreactions for allantibodies that showed significant increases in thetumor tissue were observed in each of the multiplelung tumors of all 10 hamsters.
4. Discussion
pathway, in lung adenocarcinomas of Clara cell lin-eage induced in hamsters by NNK. These findingsrepresent the first evidence in support of the hy-pothesis that both pathways may synergisticallyregulate the growth and development of this cancertype.
The EGF-receptor is expressed in about 50% ofhuman pulmonary adenocarcinomas [9,35,36]. Thepredominating signaling pathway associated withthe EGF receptor includes binding of agonist to thereceptor, resulting in phosphorylation of tyrosine ki-nases, activation of Ras, Raf and ERK1/2 [9,35,37].While it is well established that Raf-1 is a down-stream effector of Ras, over-expression of the Raf-1 protein has to our knowledge not been previouslyreported in human or experimentally induced PAC.As it is well established that NNK-induced rodentPAC expresses activating point mutations in Ki—Ras[38—43], the observed over-expression of Raf-1 maybe the consequence of constitutive activity of Ras.
In many cell types, Raf-1 is negatively regulatedby cAMP/PKA via inhibition of Raf by PKA [44,45].However, in some cell types, transactivation of theEGFR and/or its associated downstream pathwayby G-protein-coupled receptors, including the �2-asT
Our data show over-expression of members of twodifferent signal transduction pathways, the �2-adrenergic receptor pathway and the EGF-receptor
drenergic receptor has been reported (Fig. 5), re-ulting in a synergistic activation of ERK1/2 [46,47].he potential relevance of this cross-talk between

NNK-induced adenocarcinomas and EGFR signaling pathways 43
G-protein-coupled receptors and the EGF receptorfor the regulation and development of cancer is farfrom understood. Transactivation of EGFR-specifictyrosine kinases and their downstream effectors Rafand ERK1/2 in response to �2-adrenergic receptorstimulation by NNK (Fig. 6) provides a possible ex-planation for our current results and is in line withthe discovery that NNK is a �-adrenergic agonist[16].
It has been recently shown that a family of drugsthat inhibit EGFR-associated tyrosine kinases hadstunning therapeutic effects in a subpopulation ofhuman PAC cases while the majority of these pa-tients did not respond [48,49]. On the other hand,we have shown that the �-blocker propranolol sig-nificantly reduced the development of NNK-inducedPAC in hamsters [18]. We have also shown that NNKor the classic �-adrenergic agonist isoproterenolstimulated DNA synthesis in cell lines derived fromhuman pulmonary adenocarcinomas of Clara celllineage [16]. This effect was significantly inhibitedby propranolol [16], by the tyrosine kinase inhibitorAG1478 [50], as well as by inhibitors of COX and 5-LOX [16]. Moreover, the activator of cAMP, forskolin,had similar stimulatory effects on DNA synthesis,ahtawhttsvrCTdni
CmiooptPPgsPpC
cused on the EGFR pathway as a potential targetfor cancer intervention [48,49,52] while researchrelated to other histological lung cancer types haseven provided evidence for a growth inhibiting roleof cAMP, PKA and CREB [30,32]. In light of our cur-rent data, these findings underline the need for alung cancer diagnosis that goes beyond the clas-sic WHO classification. The recent identification ofEGFR mutations as a predictive factor for responseto EGFR-specific tyrosine kinase inhibitors [52] is afirst step into the right direction. However, there isan urgent need for the development of additionaldiagnostic tools that allow for the detection of sig-nal transduction pathways in control of an individ-ual patient’s lung cancer and/or precancerous le-sions so that custom-tailored cancer interventionmay be successfully applied.
Acknowledgement
Supported by grants RO1CA096128 and RO1CA88809with the National Cancer Institute.
R
response significantly reduced by both, an in-ibitor of cAMP and an inhibitor of EGFR-specificyrosine kinases [50]. By contrast, DNA synthesisnd activity of ERK1/2 of a human PAC cell lineith characteristics of alveolar type II cells was in-ibited by activators of cAMP [50]. Taken together,hese published in vitro and in vivo data in conjunc-ion with our current immunohistochemical analy-is point to transactivation of the EGFR pathwayia �-adrenergic receptor stimulation as a growth-egulating component in a subset of human PAC oflara cell lineage and in NNK-induced hamster PAC.hey also point to PKA/CREB as novel target for theevelopment of effective therapy for PAC patientson-responsive to EGFR-specific tyrosine kinasenhibitors.The tumor-selective over-expression of PKA,REB and phosphorylated CREB in our hamsterodel is a particularly interesting and novel find-ng that warrants immediate study in human PACf smokers and non-smokers. Recent studies inur laboratory have shown that activation of thisathway by agents that increase cAMP stimulateshe proliferation of human cell lines derived fromAC of Clara cell phenotype [51]. The role of theKA/CREB pathway in cardiovascular and neurode-enerative disease as well as stroke has been exten-ively studied [23—28,50]. To our knowledge, theKA/CREB pathway has to this date only been im-licated in the genesis of endocrine tumors [29].ancer research related to PAC has primarily fo-
eferences
[1] Levi F, Franceschi S, La Vecchia C, Randimbison L,Te VC. Lung carcinoma trends by histologic type inVaud and Neuchatel, Switzerland, 1974—1994. Cancer1997;79:906—14.
[2] Kelly A, Blair N, Pechacek TF. Women and smoking: is-sues and opportunities. J Womens Health Gend Based Med2001;10:515—8.
[3] Wynder EL, Muscat JE. The changing epidemiology of smok-ing and lung cancer histology. Environ Health Perspect1995;103(Suppl 8):143—8.
[4] Alavanja MC, Field RW, Sinha R, Brus CP, Shavers VL, FisherEL, et al. Lung cancer risk and red meat consumption amongIowa women. Lung Cancer 2001;34:37—46.
[5] Veierod MB, Laake P, Thelle DS. Dietary fat intake and riskof lung cancer: a prospective study of 51,452 Norwegianmen and women. Eur J Cancer Prev 1997;6:540—9.
[6] Hida T, Yatabe Y, Achiwa H, Muramatsu H, Kozaki K, Naka-mura S, et al. Increased expression of cyclooxygenase 2 oc-curs frequently in human lung cancers, specifically in ade-nocarcinomas. Cancer Res 1998;58:3761—4.
[7] El-Bayoumy K, Iatropoulos M, Amin S, Hoffmann D, Wyn-der EL. Increased expression of cyclooxygenase-2 in ratlung tumors induced by the tobacco-specific nitrosamine 4-(methylnitrosamino)-4-(3-pyridyl)-1-butanone: the impactof a high-fat diet. Cancer Res 1999;59:1400—3.
[8] Hong SH, Avis I, Vos MD, Martinez A, Treston AM, Mul-shine JL. Relationship of arachidonic acid metabolizingenzyme expression in epithelial cancer cell lines to thegrowth effect of selective biochemical inhibitors. CancerRes 1999;59:2223—8.
[9] Lonardo F, Dragnev KH, Freemantle SJ, Ma Y, Memoli N,Sekula D, et al. Evidence for the epidermal growth factorreceptor as a target for lung cancer prevention. Clin CancerRes 2002;8:54—60.

44 H.M. Schuller, M. Cekanova
[10] Maa MC, Leu TH, McCarley DJ, Schatzman RC, Parsons SJ.Potentiation of epidermal growth factor receptor-mediatedoncogenesis by c-Src: implications for the etiology of mul-tiple human cancers. Proc Natl Acad Sci USA 1995;92:6981—5.
[11] Kuller LH. Dietary fat and chronic diseases: epidemiologicoverview. J Am Diet Assoc 1997;97:S9—15.
[12] Eaton L. Smoking habits of ethnic groups lead to higher riskof heart disease. BMJ 2004;328:1397.
[13] Ueshima H, Choudhury SR, Okayama A, Hayakawa T, KitaY, Kadowaki T, et al. Cigarette smoking as a risk factor forstroke death in Japan, NIPPON DATA80. Stroke 2004.
[14] Kaplan JR, Manuck SB. Antiatherogenic effects of �-adrenergic blocking agents: theoretical, experimental,and epidemiologic considerations. Am Heart J 1994;128:1316—28.
[15] Kong DF. Aspirin in cardiovascular disorders: what is the op-timum dose? Am J Cardiovasc Drugs 2004;4:151—8.
[16] Schuller HM, Tithof PK, Williams M, Plummer III H.The tobacco-specific carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone is a �-adrenergic agonist andstimulates DNA synthesis in lung adenocarcinoma via �-adrenergic receptor-mediated release of arachidonic acid.Cancer Res 1999;59:4510—5.
[17] Tithof PK, Elgayyar M, Schuller HM, Barnhill M, An-drews R. 4-(Methylnitrosamino)-1-(3-pyridyl)-1-butanone,a nicotine derivative, induces apoptosis of endothe-lial cells. Am J Physiol Heart Circ Physiol 2001;281:H1946—54.
[18] Schuller HM, Porter B, Riechert A. �-Adrenergic modulation
[28] Gerzanich V, Ivanova S, Simard JM. Early pathophysiologi-cal changes in cerebral vessels predisposing to stroke. ClinHemorheol Microcirc 2003;29:291—4.
[29] Rosenberg D, Groussin L, Jullian E, Perlemoine K, BertagnaX, Bertherat J. Role of the PKA-regulated transcrip-tion factor CREB in development and tumorigenesisof endocrine tissues. Ann N Y Acad Sci 2002;968:65—74.
[30] Shafer SH, Phelps SH, Williams CL. Reduced DNA syn-thesis and cell viability in small cell lung carcinoma bytreatment with cyclic AMP phosphodiesterase inhibitors.Biochem Pharmacol 1998;56:1229—36.
[31] Giorgi M, Leonetti C, Citro G, Augusti-Tocco G. In vitro andin vivo inhibition of SK-N-MC neuroblastoma growth usingcyclic nucleotide phosphodiesterase inhibitors. J Neuroon-col 2001;51:25—31.
[32] Wagner B, Jakobs S, Habermeyer M, Hippe F, Cho-Chung YS,Eisenbrand G, et al. 7-Benzylamino-6-chloro-2-piperazino-4-pyrrolidino-pteridine, a potent inhibitor of cAMP-specificphosphodiesterase, enhancing nuclear protein binding tothe CRE consensus sequence in human tumour cells.Biochem Pharmacol 2002;63:659—68.
[33] Schuller HM, Witschi HP, Nylen E, Joshi PA, Correa E,Becker KL. Pathobiology of lung tumors induced in ham-sters by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanoneand the modulating effect of hyperoxia. Cancer Res1990;50:1960—5.
[34] Jull BA, Plummer III HK, Schuller HM. Nicotinic receptor-mediated activation by the tobacco-specific nitrosamineNNK of a Raf-1/MAP kinase pathway, resulting in phospho-
[
[
[
[
[
[
[
[
[
of NNK-induced lung carcinogenesis in hamsters. J CancerRes Clin Oncol 2000;126:624—30.
[19] Tran TM, Friedman J, Qunaibi E, Baameur F, Moore RH,Clark RB. Characterization of agonist stimulation of cAMP-dependent protein kinase and G protein-coupled recep-tor kinase phosphorylation of the �2-adrenergic recep-tor using phosphoserine-specific antibodies. Mol Pharmacol2004;65:196—206.
[20] Wallukat G. The �-adrenergic receptors. Herz 2002;27:683—90.
[21] Hu LA, Chen W, Premont RT, Cong M, Lefkowitz RJ.G protein-coupled receptor kinase 5 regulates � 1-adrenergic receptor association with PSD-95. J Biol Chem2002;277:1607—13.
[22] Lefkowitz RJ. The superfamily of heptahelical receptors.Nat Cell Biol 2000;2:E133—6.
[23] Muller FU, Boknik P, Knapp J, Linck B, Luss H, Neumann J, etal. Activation and inactivation of cAMP-response element-mediated gene transcription in cardiac myocytes. Cardio-vasc Res 2001;52:95—102.
[24] Pilz RB, Casteel DE. Regulation of gene expression by cyclicGMP. Circ Res 2003;93:1034—46.
[25] Chen Y, Zhuang S, Cassenaer S, Casteel DE, Gudi T, Boss GR,et al. Synergism between calcium and cyclic GMP in cyclicAMP response element-dependent transcriptional regula-tion requires cooperation between CREB and C/EBP-�. MolCell Biol 2003;23:4066—82.
[26] Takeo S, Niimura M, Miyake-Takagi K, Nagakura A, FukatsuT, Ando T, et al. A possible mechanism for improvementby a cognition-enhancer nefiracetam of spatial memoryfunction and cAMP-mediated signal transduction systemin sustained cerebral ischaemia in rats. Br J Pharmacol2003;138:642—54.
[27] Nagakura A, Takagi N, Takeo S. Impairment of cerebralcAMP-mediated signal transduction system and of spatialmemory function after microsphere embolism in rats. Neu-roscience 2002;113:519—28.
rylation of c-myc in human small cell lung carcinoma cellsand pulmonary neuroendocrine cells. J Cancer Res Clin On-col 2001;127:707—17.
35] Scagliotti GV, Selvaggi G, Novello S, Hirsch FR. The biologyof epidermal growth factor receptor in lung cancer. ClinCancer Res 2004;10:s4227—32.
36] Hsieh ET, Shepherd FA, Tsao MS. Co-expression of epider-mal growth factor receptor and transforming growth factor-alpha is independent of ras mutations in lung adenocarci-noma. Lung Cancer 2000;29:151—7.
37] Budillon A, Di Gennaro E, Caraglia M, Barbarulo D, Ab-bruzzese A, Tagliaferri P. 8-Cl-cAMP antagonizes mitogen-activated protein kinase activation and cell growth stim-ulation induced by epidermal growth factor. Br J Cancer1999;81:1134—41.
38] Belinsky SA, Devereux TR, Maronpot RR, Stoner GD, Ander-son MW. Relationship between the formation of promuta-genic adducts and the activation of the K-ras protooncogenein lung tumors from A/J mice treated with nitrosamines.Cancer Res 1989;49:5305—11.
39] Downward J. Targeting RAS signalling pathways in cancertherapy. Nat Rev Cancer 2003;3:11—22.
40] Gao HG, Chen JK, Stewart J, Song B, Rayappa C, Whong WZ,et al. Distribution of p53 and K-ras mutations in human lungcancer tissues. Carcinogenesis 1997;18:473—8.
41] Kovalchuk O, Naumnik W, Serwicka A, Chyczewska E, Niklin-ski J, Chyczewski L. K-ras codon 12 mutations may be de-tected in serum of patients suffering from adeno- and largecell lung carcinoma. A preliminary report. Folia HistochemCytobiol 2001;39(Suppl 2):70—2.
42] Malkinson AM. Molecular comparison of human andmouse pulmonary adenocarcinomas. Exp Lung Res1998;24:541—55.
43] Oreffo VI, Lin HW, Padmanabhan R, Witschi H. K-ras andp53 point mutations in 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced hamster lung tumors. Carcinogenesis1993;14:451—5.

NNK-induced adenocarcinomas and EGFR signaling pathways 45
[44] Pursiheimo JP, Kieksi A, Jalkanen M, Salmivirta M. Proteinkinase A balances the growth factor-induced Ras/ERK sig-naling. FEBS Lett 2002;521:157—64.
[45] Ramstad C, Sundvold V, Johansen HK, Lea T. cAMP-dependent protein kinase (PKA) inhibits T cell activation byphosphorylating ser-43 of raf-1 in the MAPK/ERK pathway.Cell Signal 2000;12:557—63.
[46] Maudsley S, Pierce KL, Zamah AM, Miller WE, Ahn S, DaakaY, et al. The �(2)-adrenergic receptor mediates extracel-lular signal-regulated kinase activation via assembly of amulti-receptor complex with the epidermal growth factorreceptor. J Biol Chem 2000;275:9572—80.
[47] Pierce KL, Luttrell LM, Lefkowitz RJ. New mechanisms inheptahelical receptor signaling to mitogen activated pro-tein kinase cascades. Oncogene 2001;20:1532—9.
[48] Miller VA, Kris MG, Shah N, Patel J, Azzoli C, Comez J, et al.Bronchioloalveolar pathologic subtype and smoking history
predict sensitivity to gefinitib in advanced non-small-celllung cancer. J Clin Oncol 2004;22:1103—9.
[49] Gandara DR, West H, Chansky K, Davies AM, Lau DH, Crow-ley J, et al. Bronchioloalveolar carcinoma: a model for in-vestigating the biology of epidermal growth factor receptorinhibition. Clin Cancer Res 2004;10:4205—9.
[50] Adissu HA, Schuller HM. Antagonistic growth regulation ofcell lines derived from human lung adenocarcinomas ofClara cell and aveolar type II cell lineage: implications forchemoprevention. Int J Oncol 2004;24:1467—72.
[51] Schuller HM, Al-Wadei HA, Laag E. Chemoprevention of lungcancer: novel challenges and opportunities. In: Proceed-ings of the 9th World Conference on Advances in Oncology,Crete, Greece; 2004.
[52] Paez JG, Janne PA, Lee JC, Tracy S, Greulich H, Gabriel S, etal. EGFR mutations in lung cancer: correlation with clinicalresponse to Gefitinib therapy. Science 2004.

![Oridonin protects LPS-induced acute lung injury by ......and acute lung injury (ALI) [1, 2]. Lipopolysaccharide (LPS), from the outer membrane of gram-negative bacteria, has been widely](https://static.fdocument.org/doc/165x107/608e9a4b0654131b49646243/oridonin-protects-lps-induced-acute-lung-injury-by-and-acute-lung-injury.jpg)
















