Molecular cloning and expression of IRAK-4, IL-17 and I-κB genes in Haliotis rufescens challenged...
7
Short sequence report Molecular cloning and expression of IRAK-4, IL-17 and I-kB genes in Haliotis rufescens challenged with Vibrio anguillarum Valentina Valenzuela-Muñoz, Cristian Gallardo-Escárate * Laboratory of Biotechnology and Aquatic Genomics, Interdisciplinary Center for Aquaculture Research (INCAR), University of Concepción, P.O. Box 160-C, Concepción, Chile article info Article history: Received 20 September 2013 Received in revised form 26 November 2013 Accepted 18 December 2013 Available online 4 January 2014 Keywords: Haliotis rufescens I-kB IRAK-4 IL-17 Vibrio anguillarum abstract The candidate genes interleukin-1 receptor associated kinase 4 (IRAK-4), Interleukin 17 (IL-17) and In- hibitor of NF-kB (I-kB) were cloned and evaluated in Californian abalone (Haliotis rufescens) hemocytes in response to Vibrio anguillarum. Molecular characterization evidenced that HrI-kB has a full cDNA sequence of 3027 bp with an encoding region of 401 amino acids (aa), HrIRAK-4 comprised 1969 bp that encoded for 516 aa, and Hr-IL17 had a full sequence of 806 bp encoding for 165 aa. qPCR analysis showed the higher constitutive expression level of Hr-IL17 in hemocytes; meanwhile Hr-IkB and Hr-IRAK4 gene expression levels were higher in gills and mantle. The assessment of gene expression in hemocytes after infection with V. anguillarum evidences the immune responses of Hr-IkB, Hr-IRAK4, and Hr-IL17 and their relationships through the NF-kB signaling pathway. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction The innate immune response in mollusks is the first line of defense against the continuous challenge of infection in marine environmental [3]. It triggers diverse humoral and cellular ac- tivities via signal transduction pathways, which are highly conserved in the animal kingdom [4]. Signal transduction in- volves the binding of extracellular signaling molecular and li- gands to cell-surface receptors that trigger the events inside the cell. It occurs when pathogen-associated molecular patterns (PAMPs), activates a cell surface receptors such as Toll-like re- ceptor (TLR) signal pathway, which represents a key feature of innate immune response to pathogen infection. In turn, TLR al- ters intracellular molecules creating a response that activates the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) to control the inflammatory response through cytokines and chemokines [9,15]. The pathway controlled by NF-kB also controls a series of biological processes like development, apoptosis, growth and cellular proliferation. NF-kB is composed by Rel-homology Domain (RHD) DNA binding sites which interact with I-kB to keep NF-kB inactivated, masking its nuclear localization sequence [1]. In the presence of stress-inducing agents, Ik-B is phosphorylated by I-kB kinases (IKK), generating proteolytic degradation which in turn, permits the translocation of NF-kB to the nucleus and the transcription of a series of genes involved in the immune response [2,23]. In mollusks such as Haliotis discus discus [15], Pinctada fucata [25], Metetrix metetrix [24], Crassostrea gigas [11], Argopecten irradians [12], and Rudi- tapes philippinarum [10], a close functional relationship has been observed with an increase of I-kB transcript levels in response to bacteria, suggesting its role within the innate immune system of marine invertebrates. Another activator protein of NF-kB is the IL-1R-associated kinase (IRAK), member of the serine/threonine kinases family whose kinase domains have a high level of homology between different taxa and which have a death domain (DD) on the extreme of their N-terminal. In vertebrates, the adaptive immune response is linked to the actions of IRAK-2 [13]. However, in in- vertebrates, IRAK-4 has been observed to have a key role within the innate immune response pathway through the interaction with the TRL2 receptor, which recognizes lipoproteins, and the TLR4 receptor, which recognizes lipopolysaccharides (LPS) [6,19,21]. In mollusks, IRAK-4 has only been described in C. gigas and Haliotis diversicolor [5], and an up-regulation of this gene was shown in organisms exposed to pathogens. Moreover, IRAK- 4 is a mediator for the translation of the interleukin (IL) family, of which, the interleukin 17 (IL-17) family has been shown to be mediators for the inflammatory response through the activation of NF-kB [4,7]. IL-17 is a family with six members that form a * Corresponding author. E-mail address: crisgallardo@oceanografia.udec.cl (C. Gallardo-Escárate). Contents lists available at ScienceDirect Fish & Shellfish Immunology journal homepage: www.elsevier.com/locate/fsi 1050-4648/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.fsi.2013.12.015 Fish & Shellfish Immunology 36 (2014) 503e509
Transcript of Molecular cloning and expression of IRAK-4, IL-17 and I-κB genes in Haliotis rufescens challenged...

lable at ScienceDirect
Fish & Shellfish Immunology 36 (2014) 503e509
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
Short sequence report
Molecular cloning and expression of IRAK-4, IL-17 and I-kB genesin Haliotis rufescens challenged with Vibrio anguillarum
Valentina Valenzuela-Muñoz, Cristian Gallardo-Escárate*
Laboratory of Biotechnology and Aquatic Genomics, Interdisciplinary Center for Aquaculture Research (INCAR), University of Concepción, P.O. Box 160-C,Concepción, Chile
a r t i c l e i n f o
Article history:Received 20 September 2013Received in revised form26 November 2013Accepted 18 December 2013Available online 4 January 2014
Keywords:Haliotis rufescensI-kBIRAK-4IL-17Vibrio anguillarum
* Corresponding author.E-mail address: [email protected]
1050-4648/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.fsi.2013.12.015
a b s t r a c t
The candidate genes interleukin-1 receptor associated kinase 4 (IRAK-4), Interleukin 17 (IL-17) and In-hibitor of NF-kB (I-kB) were cloned and evaluated in Californian abalone (Haliotis rufescens) hemocytes inresponse to Vibrio anguillarum. Molecular characterization evidenced that HrI-kB has a full cDNAsequence of 3027 bp with an encoding region of 401 amino acids (aa), HrIRAK-4 comprised 1969 bp thatencoded for 516 aa, and Hr-IL17 had a full sequence of 806 bp encoding for 165 aa. qPCR analysis showedthe higher constitutive expression level of Hr-IL17 in hemocytes; meanwhile Hr-IkB and Hr-IRAK4 geneexpression levels were higher in gills and mantle. The assessment of gene expression in hemocytes afterinfection with V. anguillarum evidences the immune responses of Hr-IkB, Hr-IRAK4, and Hr-IL17 and theirrelationships through the NF-kB signaling pathway.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The innate immune response in mollusks is the first line ofdefense against the continuous challenge of infection in marineenvironmental [3]. It triggers diverse humoral and cellular ac-tivities via signal transduction pathways, which are highlyconserved in the animal kingdom [4]. Signal transduction in-volves the binding of extracellular signaling molecular and li-gands to cell-surface receptors that trigger the events inside thecell. It occurs when pathogen-associated molecular patterns(PAMPs), activates a cell surface receptors such as Toll-like re-ceptor (TLR) signal pathway, which represents a key feature ofinnate immune response to pathogen infection. In turn, TLR al-ters intracellular molecules creating a response that activates thenuclear factor kappa-light-chain-enhancer of activated B cells(NF-kB) to control the inflammatory response through cytokinesand chemokines [9,15]. The pathway controlled by NF-kB alsocontrols a series of biological processes like development,apoptosis, growth and cellular proliferation. NF-kB is composedby Rel-homology Domain (RHD) DNA binding sites whichinteract with I-kB to keep NF-kB inactivated, masking its nuclearlocalization sequence [1]. In the presence of stress-inducingagents, Ik-B is phosphorylated by I-kB kinases (IKK), generating
(C. Gallardo-Escárate).
All rights reserved.
proteolytic degradation which in turn, permits the translocationof NF-kB to the nucleus and the transcription of a series of genesinvolved in the immune response [2,23]. In mollusks such asHaliotis discus discus [15], Pinctada fucata [25], Metetrix metetrix[24], Crassostrea gigas [11], Argopecten irradians [12], and Rudi-tapes philippinarum [10], a close functional relationship has beenobserved with an increase of I-kB transcript levels in response tobacteria, suggesting its role within the innate immune system ofmarine invertebrates.
Another activator protein of NF-kB is the IL-1R-associatedkinase (IRAK), member of the serine/threonine kinases familywhose kinase domains have a high level of homology betweendifferent taxa and which have a death domain (DD) on theextreme of their N-terminal. In vertebrates, the adaptive immuneresponse is linked to the actions of IRAK-2 [13]. However, in in-vertebrates, IRAK-4 has been observed to have a key role withinthe innate immune response pathway through the interactionwith the TRL2 receptor, which recognizes lipoproteins, and theTLR4 receptor, which recognizes lipopolysaccharides (LPS)[6,19,21]. In mollusks, IRAK-4 has only been described in C. gigasand Haliotis diversicolor [5], and an up-regulation of this genewas shown in organisms exposed to pathogens. Moreover, IRAK-4 is a mediator for the translation of the interleukin (IL) family, ofwhich, the interleukin 17 (IL-17) family has been shown to bemediators for the inflammatory response through the activationof NF-kB [4,7]. IL-17 is a family with six members that form a

V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509504
strong relation with other cytokines, stimulating macrophagesand transcription factors [7]. Herein, there is little information inmarine invertebrates about the structure and function of IL-17,and there are currently only two studies that report sequencesfor P. fucata and C. gigas [16,22]. The objective of this study was tocharacterize the genes I-kB, IRAK-4, and IL-17 and to evaluate therelationship between these genes in red abalone Haliotis rufes-cens hemocytes challenged with Vibrio anguillarum, causativeagent of vibriosis, a deadly hemorrhagic septicemic diseaseaffecting various marine organisms.
2. Materials and methods
2.1. Experimental design
Adult red abalones (body length 60 � 5 mm, weight22.5 � 4.0 g) were purchased from a commercial farm located innorthern Chile. All abalones were left undisturbed for 2 weeks inpolyethylene tanks, each containing 20 animals in 200 L aeratedseawater at 14 � 1 �C, and fed daily with fresh kelp. Then thirtyspecimens were used to characterize the mRNA of I-kB, IRAK-4and IL-17, and they were also challenged against V. anguillarum.For the challenge, V. anguillarum were grown in LuriaeBertani(LB) and incubated at 20 �C for 24e48 h, once the concentrationof 3.2 � 108 cell/mL was achieved, challenged group was injectedwith 200 mL of bacteria directly in muscle, while the controlgroup was injected with 100 mL LB medium without bacteria.Subsequently, tissue samples (mantle, gills, hemocytes, andmuscle) from four abalones were collected at 0, 3, 6, 12, 24, 32and 48 h post-injection, fixed in RNAlater� RNA StabilizationReagent (Ambion, USA) and stored at �80 �C for subsequent RNAextraction.
2.2. IL-17, IRAK-4 and I-kB gene isolation
Partial sequences of IL-17, IRAK4 and I-kB genes were identi-fied from the 454 pyrosequencing database of H. rufescens,available at the Dryad Digital Repository under the access http://dx.doi.org/10.5061/dryad.dh5qs [20]. These sequences wereused as a template for primer design with the Primer3 tool [17](Table 1) included in the Geneious Pro software [3]. For gene
Table 1Sequence of oligonucleotide primers used.
Primer Sequence 50e30
Hr_IkB_5RACE_outer GAATTTGGCTCAACCATCGCGCAGTHr_IkB_3RACE_outer TTGACAGCTGTACATACGGTGGGCTHr_IkB_5RACE_inner CCTCCGTCTTCGCCGAGGAGTTATHr_IkB_3RACE_inner GTCCTGGCTGCAGGAAGAAAGTTCGHr_IkB_F ACTGCGCGATGGTTGAGCCAAATTHr_IkB_R CGAACTTTCTTCCTGCAGCCAGGACHr_IL17_5RACE_outer GCATGGGGATATGTGGTCATCHr_IL17_3RACE_outer CCTACCGTTATCTATGAGGCCHr_IL17_5RACE_inner GGTCATCACTTGTCCGGAAGTAACGHr_IL17_3RACE_inner CGTTATCTATGAGGCCAGGTGCHr_IRAK4_5RACE_outer GTCTAAAATTGCACGTGTGGHr_IRAK4_3RACE_outer GAACGGGGGCAACATGTACACAGACHr_IRAK4_5RACE_inner GCCTCGATCCTCGATAATACHr_IRAK4_3RACE_inner ACAATGGTTGTGATCGGCACGTCAGHr_IkB_qF CCAAAAATTATGAAGGCCATACCTGHr_IkB_qR AGTCTCAGCAGCATAATGAAGAAHr_IL17_qF GTACTTCTTAAGATCCGTTCTCCAHr_IL17_qR CAATGTATGTCCATCCTCTTCATAGHr_IRAK4_qF CATACGTAAGATCATACTTTGGCACHr_IRAK4_qR GAACAGGCTACAATTACTCCTAAAGHr_EF_F GGAGGGTCAAACCCGAGAACHr_EF_R CCGATGATGAGTTGCTGCTTCACA
amplification, total RNA was isolated using the TRI reagent(Invitrogen, Carlsbad, CA, USA) protocol. The purity was deter-mined (ratio A260/A280) with a Nanodrop ND1000 spectro-photometer (Thermo Fisher Scientific, Copenhagen, USA), andthe integrity was determined by agarose gel under denaturantconditions. From a 200 ng/ml of total RNA, cDNA was synthetizedusing the RevertAid H Minus First Strand cDNA Synthesis Kit(Thermo Scientific, Glen Burnie, Maryland, USA). PCR analysiswas performed using 1 mL of cDNA, 10 mM of each primer(Table 1), 1.5 mM MgCl2, and 0.06 U taq DNA polymerase(Thermo Scientific, Maryland, USA). PCR was performed in 35cycles of denaturation at 94 �C for 30 s, annealing at 60 �C for30 s, and extension at 72 �C for 45 s. The PCR product wasobserved by electrophoresis on 1% agarose gel and sequenced inthe ABI 3730xl sequencer (Applied Biosystems, CA, USA). Thesequence was analyzed using the Geneious 5.1.6 software [3].The resulting partial sequences, termed Hr-IL17, Hr-IKAR4 andHrI-kB, were used for designing specific new primers (Table 1)for the amplification of the 30 and 50 UTR ends through theSMARTer� RACE cDNA Amplification Kit (Clontech) according tothe manufacturer’s instructions. The fragments obtained for the50 and 30 UTR ends were cloned in the TOPO TA Cloning Kit(Invitrogen�, Life Technologies, Carlsbad, CA, USA) and trans-formed into Escherichia coli JM109 electrocompetent bacteria inLB/amp/IPTG/Xgal plates overnight at 37 �C. The positive cloneswere selected and purified to obtain plasmid by using theE.Z.N.A� Plasmid DNA Mini Kit II (Omega Bio-tek, Doraville, GA,USA). The plasmids obtained were sequenced in both directions,and their sequences were then assembled using the softwareGeneious 5.1.3 [3].
2.3. Amino acid sequences analysis
Protein alignments were conducted using MUSCLE, and phylo-genetic trees were constructed using the neighbor-joining methodwith 1000 bootstrap repetitions. Both analyses were carried out inGeneious 5.1.6 [3]. For the I-kB sequence, ankyrin repeats and thepotential PEST domain were predicted using the methodologyproposed by Yang et al. [24].
2.4. Expression profiles of Hr-IL17, Hr-IRAK4 and Hr-IkB
We evaluated transcripts levels in different tissues from fourindividuals as biological replicates. Herein, for each tissue(mantle, muscle, gills, hemocytes and digestive glands) threetechnical replicates were used. For hemocytes, a pool obtainedfrom the four evaluated individuals for each time was used forevaluating gene expressions after the challenge withV. anguillarum. The primers for qPCR reactions were designedfrom the sequences obtained from Hr-IL17, Hr-IRAK4 and Hr-IkB(Table 1). The qPCR runs were performed with StepOnePlus�(Applied Biosystems, Life Technologies, USA) using the compar-ative DCt method. Each reaction was conducted with a volume of10 mL using the Maxima� SYBR Green/ROX qPCR Master Mix(Thermo Scientific, USA). The amplification conditions were asfollows: 95 �C for 10 min, 40 cycles at 95 �C for 30 s, 60 �C for30 s, and 72 �C for 30 s. Three putative housekeeping genes(HKG), Elongation factor alpha, b-tubulin and b-tubulin were sta-tistically analyzed by NormFinder algorithm to assess theirtranscriptional expression stability. Here, Elongation factor alphawas selected as HKG for gene normalization. Statistical analyseswere performed using the software Statistical 8.0. The data ob-tained were analyzed through the KruskaleWallis test withStatistica software (Version 7.0, StatSoft, Inc.). Statistically sig-nificant differences were accepted with a p < 0.05.

V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509 505
3. Results
3.1. Molecular characterization of Hr-IL17, Hr-IKAR4 and Hr-IkBgenes
The sequences for the three characterized genes were obtainedby overlapping partial sequences identified in the 454 EST-database[20] and the complete open reading frames and UTRs were ob-tained by RACE-PCR. The full sequences for Hr-IL17, Hr-IRAK4 andHr-IkB genes were deposited into the NCBI database under GenBankAccession No. KF357963, KF357964, KF357965.
The complete cDNA sequence for Hr-IL17 had 806 bp with anopen reading frame (ORF) of 495 bp encoding for 165 amino acids,while the 50UTR region was 108 bp and the 30UTR was 203 bp(Fig. 1A). The amino acid sequence had a predicted molecularweight of 18.7 kDa and an isoelectric point of 9.19. In the C-terminalregion of IL-17 five conserved cysteine residues were identified(Fig. 1 A).
The Hr-IRAK-4 sequence showed a full cDNA sequence of1969 bp with 50UTR and 30UTR regions of 52 and 369 bp, respec-tively. The ORF region had 1548 bp encoding for 516 amino acids.The predicted molecular weight was 57.9 kDa with an isoelectricpoint of 4.98. For IRAK-4, a death domain (DD) between aminoacids 15 and 119, a protein kinase ATP-binding region, a Serine/
Fig. 1. Full-length cDNA sequence and predicted amino acid sequence for HrIL-17. (A) SchemPCR, and the complete amino acid sequence. In orange: amino acid peptide signal; red: six coresidues. (For interpretation of the references to color in this figure legend, the reader is re
Threonin protein kinase active site, and a prokaryotic membranelipoprotein lipid attachment site were identified (Fig. 2A).
TheHr-IkB gene had a full cDNA sequence of 3027 bp, containing166 and 1658 bp of 50UTR and 30UTR regions, respectively. The ORFshowed 1203 bp encoding a polypeptide of 401 amino acids(Fig. 3A), with a predicted molecular weight of 44.6 kDa and anisoelectric point of 4.72. Here, two lysine residues (K-44 and K-45)that have a role in phosphorylation signaling for I-kB degradationwere identified at the N-terminal end and a DS59GIFS63 conservedregion. The C-terminal region evidenced six ankyrin repeats and aPEST motif (Fig. 3A).
3.2. Sequence analysis
The amino acid sequence obtained from the translation of theORF of IL-17 presented a 23.4% identity to other sequencesdescribed in vertebrates and invertebrates species (Fig. 1B). Herein,this sequence is the first sequence reported for IL-17 in gastropodsspecies. After performing a phylogenetic analysis through theneighbor-joining method, the sequence of Hr-IL17 was groupedwith the other two sequences of IL-17 reported for C. gigas andP. fucata (Fig. S1). From a multiple sequence alignment of Hr-IRAK4,40.0% of identity was observed to sequences reported for verte-brates and invertebrates species, and 82.2% with H. diversicolor
atic representation of complete HrIL-17 for H. rufescens sequences obtained by RACE-nserved Cysteines. (B) Multiple alignment of HrIL-17, in black the conserved amino acidferred to the web version of this article.)

Fig. 2. Full-length cDNA sequence and predicted amino acid sequence for HrIRAK-4. (A) Schematic representation of complete HrIRAK-4 for H. rufescens sequences obtained byRACE-PCR and the complete amino acid sequence. In green: protein kinase ATP-binding region; red: serine/threonine protein kinase actives site; orange: prokaryotic membranelipoprotein lipid attachment site. (B) Multiple alignment of HrIRAK-4, in black the conserved amino acid residues. (For interpretation of the references to color in this figure legend,the reader is referred to the web version of this article.)
V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509506
(Fig. 2B). Further, the phylogenetic analysis for IRAK-4 evidences aMollusca clade composed by sequences described for C. gigas andclosely related to H. diversicolor (Fig. S2). With respect to Inhibitorof NF-kB, this has 34.5% identity to other reported sequences(Fig. 3B), as well as a high conservation in anykrin repeats, phos-phorylation site, and tEST domain. On other hand, we observedfrom the multiple alignment an identity of 95.7% between the I-kBsequences forH. rufescens reported in this studywith the sequencespublished in GenBank for H. discus discus. From the phylogenicanalysis performed, it was possible to identify two importantclades, one that groups vertebrates and that had a subdivisiondiving the I-kBa and I-kBb isoforms, and the clade that groups Hr-IkB with others described for invertebrates (Fig. S3).
3.3. Expression analysis
Through qPCR, the transcript levels for the three characterizedgenes were analyzed in different tissues including the mantle,muscle, gills, and hemocytes. The relative expression was normal-ized using Elongation factor alpha as a housekeeping gene due itsstability value of 0.113. Higher expression levels of Hr-IL17 wereevidenced in hemocytes (Fig. S4), while higher expression for theHr-IRAK4 and Hr-IkB genes was observed in both tissues of mantle
and gills (Fig. S4). From this analysis, the response of Hr-IRAK4, Hr-IL17 and Hr-IkB challenged with V. anguillarum in abalone hemo-cytes was evaluated. ForHr-IL17 an overregulationwas registered at12 hpi (Fig. 4A), which was followed by an increase in the expres-sion of IRAK-4 expressed at 24 hpi (Fig. 4B). Likewise, I-kB showed ahigh transcripts level at the start of the challenge but declined until24 hpi, at which point an overregulation of the genewas once againobserved (Fig. 4C). These results evidence that after bacterialinfection, I-kB is modulated at different times by Hr-IL17 and Hr-IRAK4, triggering an increase of transcription expression from12 hpi to 48 hpi.
4. Discussion
Due to the limited genomic information for non-model aquaticspecies, studies involving genes relating to the innate immuneresponse in marine gastropods, and specifically in abalones, arepractically nonexistent. Thus, the present study characterizes IRAK-4, IL-17 and I-kB for the first time in abalone hemocytes challengedto a bacterial infection. The translated sequence of I-kB presentsdomains and sites characteristic of the I-kB gene family. Amongthese characteristic sites are the ankyrin repeats which have beenidentified in a large number of proteins through proteineprotein

Fig. 3. Full-length cDNA sequence and predicted amino acid sequence for HrI-kB. (A) Schematic representation of complete HrI-kB for H. rufescens sequences obtained by RACE-PCRand the complete amino acid sequence. In orange: phosphorylation sites; red: six ankyrin repeats; green: PEST domain. (B) Multiple alignment of HrI-kB, in black the conservedamino acid residues, and ankyrin conserved region. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509 507
interactions, leading to the proposal that these repeats have therole of maintaining the bond to NF-kB in I-kB [1,23]. In the sequencecharacterized for H. rufescens, six of these repeats were identified,which was equal to the number of ankyrin repeats found inH. discus discus [15], P. fucata [25], A. irradians [12], R. philippinarum[10], and C. gigas [26]. This is in contrast toMeretrix meretrix,whereonly five of these repeats were identified [24]. In the N-terminalextreme of I-kB, a DS59GIFS63 degradation motif is present which isphosphorylated when the cell receives stimuli from the extracel-lular environment [18]. Also, in the C-terminal a PEST domain wasidentified with a 100% homology to H. discus discus, and it is highlyconserved compared to other species of vertebrates and in-vertebrates [15,24]. In mollusks, there is little informationregarding IRAK-4. The IRAK-4 characterized for H. rufescens presentsa site in the N-terminal extreme homologous to the death domain(DD) of vertebrates, which in vertebrates facilitates the interactionbetween IRAK-4 and MyD88 [6], activating the inflammatoryresponse signaling pathway [13]. A kinase domain (KD) was alsoidentified for IRAK-4 that is responsible for the production of cy-tokines and chemokines activated by TIR [21]. In regards to IL-17,this protein has been described mainly in relation to the inflam-matory response for marine invertebrates, as with the regulation of
NF-kB. However, this has not yet been characterized in gastropods.The present study characterized the complete mRNA sequence ofIL-17 for H. rufescens, and this presents five characteristic cysteineresidues and a high homology with IL-17D described for verte-brates similar to those characterized for C. gigas [16] and P. fucata[22].
NF-kB in vertebrates has been shown to be closely related to 150genes, which are involved in different biological functions such asimmune response, cellular growth, stress response, developmentand cellular differentiation, among others [8,14]. Thus, it is possibleto infer the tissues involved in the innate immune response andthose principally activated by NF-kB. For example, the expression ofI-kB has been described in the hemocytes of C. gigas [26] and M.meretrix [24]. An up-regulation of IRAK-4 in hemocytes has alsobeen observed in Penaeus monodon [21]. The current study on thered abalone showed that Hr-IL17 was the candidate gene with thehighest transcript levels in hemocytes. Additionally, the resultsobtained from the evaluation of the immune response of Hr-IRAK4,Hr-IL17 and Hr-IkB in abalone hemocytes challenged withV. anguillarum evidence an interaction among the exposition timesand the regulation of these genes. A decrease in the expressionlevels of HrI-kBwas observed until 12 hpi, which was reversed from
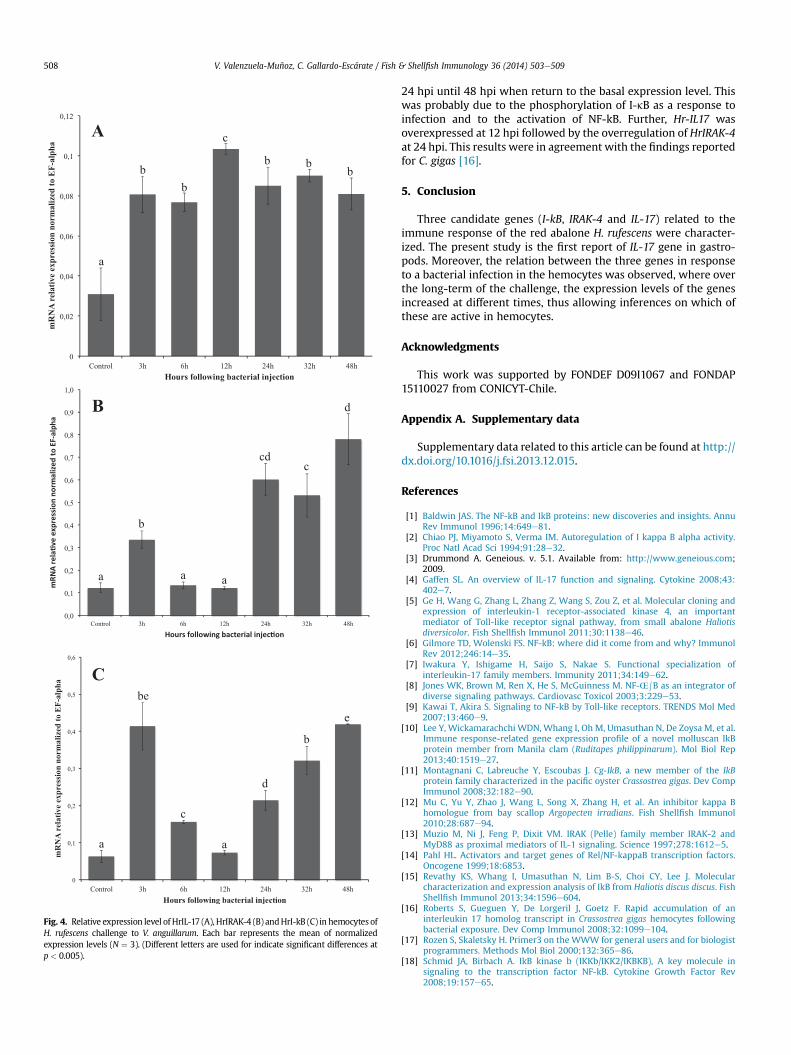
Fig. 4. Relative expression level of HrIL-17 (A), HrIRAK-4 (B) andHrI-kB (C) in hemocytes ofH. rufescens challenge to V. anguillarum. Each bar represents the mean of normalizedexpression levels (N ¼ 3). (Different letters are used for indicate significant differences atp< 0.005).
V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509508
24 hpi until 48 hpi when return to the basal expression level. Thiswas probably due to the phosphorylation of I-kB as a response toinfection and to the activation of NF-kB. Further, Hr-IL17 wasoverexpressed at 12 hpi followed by the overregulation of HrIRAK-4at 24 hpi. This results were in agreement with the findings reportedfor C. gigas [16].
5. Conclusion
Three candidate genes (I-kB, IRAK-4 and IL-17) related to theimmune response of the red abalone H. rufescens were character-ized. The present study is the first report of IL-17 gene in gastro-pods. Moreover, the relation between the three genes in responseto a bacterial infection in the hemocytes was observed, where overthe long-term of the challenge, the expression levels of the genesincreased at different times, thus allowing inferences on which ofthese are active in hemocytes.
Acknowledgments
This work was supported by FONDEF D09I1067 and FONDAP15110027 from CONICYT-Chile.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.fsi.2013.12.015.
References
[1] Baldwin JAS. The NF-kB and IkB proteins: new discoveries and insights. AnnuRev Immunol 1996;14:649e81.
[2] Chiao PJ, Miyamoto S, Verma IM. Autoregulation of I kappa B alpha activity.Proc Natl Acad Sci 1994;91:28e32.
[3] Drummond A. Geneious. v. 5.1. Available from: http://www.geneious.com;2009.
[4] Gaffen SL. An overview of IL-17 function and signaling. Cytokine 2008;43:402e7.
[5] Ge H, Wang G, Zhang L, Zhang Z, Wang S, Zou Z, et al. Molecular cloning andexpression of interleukin-1 receptor-associated kinase 4, an importantmediator of Toll-like receptor signal pathway, from small abalone Haliotisdiversicolor. Fish Shellfish Immunol 2011;30:1138e46.
[6] Gilmore TD, Wolenski FS. NF-kB: where did it come from and why? ImmunolRev 2012;246:14e35.
[7] Iwakura Y, Ishigame H, Saijo S, Nakae S. Functional specialization ofinterleukin-17 family members. Immunity 2011;34:149e62.
[8] Jones WK, Brown M, Ren X, He S, McGuinness M. NF-Œ!B as an integrator ofdiverse signaling pathways. Cardiovasc Toxicol 2003;3:229e53.
[9] Kawai T, Akira S. Signaling to NF-kB by Toll-like receptors. TRENDS Mol Med2007;13:460e9.
[10] Lee Y, Wickamarachchi WDN, Whang I, Oh M, Umasuthan N, De Zoysa M, et al.Immune response-related gene expression profile of a novel molluscan IkBprotein member from Manila clam (Ruditapes philippinarum). Mol Biol Rep2013;40:1519e27.
[11] Montagnani C, Labreuche Y, Escoubas J. Cg-IkB, a new member of the IkBprotein family characterized in the pacific oyster Crassostrea gigas. Dev CompImmunol 2008;32:182e90.
[12] Mu C, Yu Y, Zhao J, Wang L, Song X, Zhang H, et al. An inhibitor kappa Bhomologue from bay scallop Argopecten irradians. Fish Shellfish Immunol2010;28:687e94.
[13] Muzio M, Ni J, Feng P, Dixit VM. IRAK (Pelle) family member IRAK-2 andMyD88 as proximal mediators of IL-1 signaling. Science 1997;278:1612e5.
[14] Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors.Oncogene 1999;18:6853.
[15] Revathy KS, Whang I, Umasuthan N, Lim B-S, Choi CY, Lee J. Molecularcharacterization and expression analysis of IkB from Haliotis discus discus. FishShellfish Immunol 2013;34:1596e604.
[16] Roberts S, Gueguen Y, De Lorgeril J, Goetz F. Rapid accumulation of aninterleukin 17 homolog transcript in Crassostrea gigas hemocytes followingbacterial exposure. Dev Comp Immunol 2008;32:1099e104.
[17] Rozen S, Skaletsky H. Primer3 on the WWW for general users and for biologistprogrammers. Methods Mol Biol 2000;132:365e86.
[18] Schmid JA, Birbach A. IkB kinase b (IKKb/IKK2/IKBKB), A key molecule insignaling to the transcription factor NF-kB. Cytokine Growth Factor Rev2008;19:157e65.

V. Valenzuela-Muñoz, C. Gallardo-Escárate / Fish & Shellfish Immunology 36 (2014) 503e509 509
[19] Suzuki N, Saito T. IRAK-4 a shared NF-kB activator in innate and acquiredimmunity. Trends Immunol 2006;27:566e72.
[20] Valenzuela-Muñoz V, Bueno-Ibarra MA, Escárate CG. Characterization of thetranscriptomes of Haliotis rufescens reproductive tissues. Aquac Res 2012.http://dx.doi.org/10.1111/are.12044.
[21] Watthanasurorot A, Söderhäll K, Jiravanichpaisal P. A mammalian likeinterleukin-1 receptor-associated kinase 4 (IRAK-4), a TIR signaling mediatorin intestinal innate immunity of black tiger shrimp (Penaeus monodon). Bio-chem Biophys Res Commun 2012;417:623e9.
[22] Wu S-Z, Huang X-D, Li Q, He M-X. Interleukin-17 in pearl oyster (Pinctadafucata): molecular cloning and functional characterization. Fish ShellfishImmunol 2013;34:1050e6.
[23] Yamamoto M, Takeda K. Role of nuclear IkB proteins in the regulation of hostimmune responses. J Infect Chemother 2008;14:265e9.
[24] Yang Q, Yang Z, Li H. Molecular characterization and expression analysis of aninhibitor of NF-kB (IkB) from Asiatic hard clamMeretrix meretrix. Fish ShellfishImmunol 2011;31:485e90.
[25] Zhang D, Jiang S, Qiu L, Su T, Wu K, Li Y, et al. Molecular characterization andexpression analysis of the IkB gene from pearl oyster Pinctada fucata. FishShellfish Immunol 2009;26:84e90.
[26] Zhang Y, He X, Yu Z. Two homologues of inhibitor of NF-kappa B (I-kB) areinvolved in the immune defense of the Pacific oyster, Crassostrea gigas. FishShellfish Immunol 2011;30:1354e61.