![OPEN ACCESS International Journal of Molecular Sciences...hair growth [2].Platelet-derived growth factor (PDGF) isoforms reportedlyinduce and maintain theanagen phase of the murine](https://static.fdocument.org/doc/165x107/60f85444d7faee31306fdb0e/open-access-international-journal-of-molecular-sciences-hair-growth-2platelet-derived.jpg)
Melphalan, alone or conjugated to an FSH-β peptide, kills murine testicular cells in vitro and...
8
Melphalan, alone or conjugated to an FSH-b peptide, kills murine testicular cells in vitro and transiently suppresses murine spermatogenesis in vivo John K. Amory a , SungWoo Hong b , Xiaozhong Yu c , Charles H. Muller d , Elaine Faustman b , Alex Goldstein e, * a Department of Internal Medicine, University of Washington, Seattle, Washington, USA b Department of Environmental and Occupational Health Sciences, University of Washington, Seattle, Washington, USA c Department of Environmental Health Science, University of Georgia, Athens, Georgia, USA d Department of Urology, University of Washington, Seattle, Washington, USA e Focused Scientific Inc., Newcastle, Washington, USA article info Article history: Received 10 September 2013 Received in revised form 14 March 2014 Accepted 14 March 2014 Keywords: Gonadotoxicant Sterilization Male animal contraception Spermatogenesis abstract New approaches to sterilizing male animals are needed to control captive and wild animal populations. We sought to develop a nonsurgical method of permanent sterilization for male animals by administering the gonadotoxicant melphalan conjugated to peptides derived from the b-chain of FSHb. We hypothesized that conjugating melphalan to FSHb peptides would magnify the gonadotoxic effects of melphalan while minimizing systemic toxicity. The ability of conjugates of melphalan and FSHb peptides to kill murine testicular cells was first tested in vitro in a three-dimensional testicular cell coculture system. In this system, melphalan caused considerable cell death as measured both by increases in lactate dehydrogenase concentrations in the culture supernatant and direct visualization of the cultures. Of the conjugates tested, melphalan conjugated to a 20-amino acid peptide derived from human FSHb consisting of amino acids 33 to 53 (FSHb (33–53)-melphalan) was very potent, with cell cytotoxicity and lactate dehydrogenase release roughly one-half that of melphalan. The effects of melphalan and FSHb (33–53)-melphalan on spermatogenesis were then tested in vivo in mature C56Bl/6 male mice. Four weeks after intraperitoneal injection, all mice treated with either FSHb (33–53)-melphalan or melphalan had approximately 75% reductions in testicular spermatid counts compared with control animals. Testicular histology revealed significant reduction in mature spermatids and spermatocytes in most tubules. However, 12 weeks after the injection, testicular spermatid counts and histology were similar to controls, except in one animal receiving FSHb (33–53)-melphalan that had no apparent spermatogenesis. We conclude that melphalan and FSHb (33–53)-melphalan are potent gonadotoxicants in male mice resulting in marked suppression of spermatogenesis 4 weeks after a single intraperi- toneal injection. However, this effect is transient in most mice as spermatogenesis is similar to control animals 12 weeks after drug administration. Melphalan or FSHb (33–53)-melphalan may be useful for the temporary control of fertility in male animals, but additional research will be needed to develop a single dose method of permanent sterilization for male animals. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction The most common method of sterilizing male animals relies on removal of the testes, which requires skilled * Corresponding author. Tel.:/fax: þ1 (206) 221 0927. E-mail address: [email protected] (J.K. Amory). Contents lists available at ScienceDirect Theriogenology journal homepage: www.theriojournal.com 0093-691X/$ – see front matter Ó 2014 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.theriogenology.2014.03.014 Theriogenology xxx (2014) 1–8
Transcript of Melphalan, alone or conjugated to an FSH-β peptide, kills murine testicular cells in vitro and...

ilable at ScienceDirect
Theriogenology xxx (2014) 1–8
Contents lists ava
Theriogenology
journal homepage: www.ther io journal .com
Melphalan, alone or conjugated to an FSH-b peptide, killsmurine testicular cells in vitro and transiently suppressesmurine spermatogenesis in vivo
John K. Amory a, SungWoo Hong b, Xiaozhong Yu c, Charles H. Muller d,Elaine Faustman b, Alex Goldstein e,*
aDepartment of Internal Medicine, University of Washington, Seattle, Washington, USAbDepartment of Environmental and Occupational Health Sciences, University of Washington, Seattle, Washington, USAcDepartment of Environmental Health Science, University of Georgia, Athens, Georgia, USAdDepartment of Urology, University of Washington, Seattle, Washington, USAe Focused Scientific Inc., Newcastle, Washington, USA
a r t i c l e i n f o
Article history:Received 10 September 2013Received in revised form 14 March 2014Accepted 14 March 2014
Keywords:GonadotoxicantSterilizationMale animal contraceptionSpermatogenesis
* Corresponding author. Tel.:/fax: þ1 (206) 221 0E-mail address: [email protected] (J.K. Amory).
0093-691X/$ – see front matter � 2014 Elsevier Inchttp://dx.doi.org/10.1016/j.theriogenology.2014.03.01
a b s t r a c t
New approaches to sterilizing male animals are needed to control captive and wild animalpopulations. We sought to develop a nonsurgical method of permanent sterilization for maleanimals by administering the gonadotoxicantmelphalan conjugated to peptides derived fromthe b-chain of FSHb. We hypothesized that conjugating melphalan to FSHb peptides wouldmagnify the gonadotoxic effects of melphalanwhile minimizing systemic toxicity. The abilityof conjugates of melphalan and FSHb peptides to kill murine testicular cells was first testedin vitro in a three-dimensional testicular cell coculture system. In this system, melphalancaused considerable cell death as measured both by increases in lactate dehydrogenaseconcentrations in the culture supernatant and direct visualization of the cultures. Of theconjugates tested, melphalan conjugated to a 20-amino acid peptide derived from humanFSHb consisting of amino acids 33 to 53 (FSHb (33–53)-melphalan) was very potent, with cellcytotoxicity and lactate dehydrogenase release roughly one-half that ofmelphalan. The effectsof melphalan and FSHb (33–53)-melphalan on spermatogenesis were then tested in vivo inmature C56Bl/6 male mice. Four weeks after intraperitoneal injection, all mice treated witheither FSHb (33–53)-melphalan ormelphalan had approximately 75% reductions in testicularspermatid counts compared with control animals. Testicular histology revealed significantreduction in mature spermatids and spermatocytes in most tubules. However, 12 weeks afterthe injection, testicular spermatid counts and histologywere similar to controls, except in oneanimal receiving FSHb (33–53)-melphalan that had no apparent spermatogenesis. Weconclude that melphalan and FSHb (33–53)-melphalan are potent gonadotoxicants in malemice resulting in marked suppression of spermatogenesis 4 weeks after a single intraperi-toneal injection. However, this effect is transient inmostmice as spermatogenesis is similar tocontrol animals 12 weeks after drug administration. Melphalan or FSHb (33–53)-melphalanmay be useful for the temporary control of fertility in male animals, but additional researchwill be needed to develop a single dose method of permanent sterilization for male animals.
� 2014 Elsevier Inc. All rights reserved.
927.
. All rights reserved.4
1. Introduction
The most common method of sterilizing male animalsrelies on removal of the testes, which requires skilled

Table 1Chemical composition of melphalan and FSHb-melphalan conjugates.
Conjugate Peptide withmelphalan (X)placement
N-terminalmodifications
C-terminalmodifications
Molecularweight(Daltons)
Melphalan d d d 305C X33–53 Amine Acid 2815D 33–53X Amine Acid 2815E X33–53 Acetate Amide 2856F 33–53X Acetate Amide 2856G 81–95X Acetate Amide 1866H X81–95 Amine Acid 1826I 81–95X Amine Acid 1826K 33–54(X) Acetate Amide 2984M 33–47X Acetate Amide 2163N 33–50X Acetate Amide 2521O 33–53XXX Acetate Amide 3431Q 33–52X Acetate Amide 2709R 35–53X Acetate Amide 2592S 37–53X Acetate Amide 2320T 33–53XX Acetate Amide 3143
Aminoacid
Sequence
33–53 Ac-Tyr-Thr-Arg-Asp-Leu-Val-Tyr-Lys-Asp-Pro-Ala-Arg-Pro-Lys-Ile-Gln-Lys-Thr-Ser-Thr-Phe-NH2
81–95 Ac-Gln-Ser-His-Ser-Gly-Lys-Ser-Asp-Ser-Asp-Ser-Thr-Asp-Ser-Thr-NH2
The X represents the position of amolecule of melphalan in the conjugatesrelative to the amino acids and the numbers represent the amino acids inthe sequence of the human FSHb molecule, shown using the standardthree-letter abbreviations.
J.K. Amory et al. / Theriogenology xxx (2014) 1–82
personnel and can cause significant postprocedural pain [1].Removal of the testes also deprives the animal of testos-terone, which may lead to health and behavioral problems[2], and increase the risk of prostate cancer [3–5] or osteo-porosis [2,6] later in life. An alternative approach to malesterilization is the injection of a solution of calcium chloride[7] or zinc gluconate [8] directly into the testes. These in-jections are very effective from a contraceptive standpoint;however, severe injection site reactions requiring surgerycan occur in up to 4% of recipients [9], with others experi-encing scrotal ulceration or dermatitis [10]. Therefore, thereis a need for novel methods of permanent sterilization formale animals. Ideally, such a method would be inexpensiveand administered with a single injection.
Several investigators have examined the potential ofgonadotoxicants for the purposes of male sterilization. Forexample, ketoconazole [11], alpha-chlorohydrin [12], andemblein [13] have been tested in animals. However, thesuppression of spermatogenesis was incomplete, and sys-temic toxicity was limited [14]. We sought to test a novelapproach to male sterilization using the potent gonadotox-icant melphalan. Melphalan, an alkylating agent, is widelyused in the treatment of lymphomas and multiple myelomain both humans and animals. Infertility is frequently seenafter administration of melphalan [15,16], and melphalan isfairly unique among gonadotoxicants in that it kills bothspermatogonial stemcells anddividinggermcells invivo [17].Melphalan has been used extensively in animals for thetreatment of malignancies [18,19], and its pharmacokineticsand marrow toxicity are well described [20,21]. Despite itswidespread use in animals, the effect of melphalan on sper-matogenesis in animals has not beenwell studied.We soughtto: (1) determine if melphalan could be used to induce ste-rility, and (2) investigate if we could facilitate melphalaneffect and minimize any toxicity by targeting the melphalanspecifically to the testes via conjugation to peptides derivedfrom the b-chain of human FSH-b.
FSH is a protein hormone produced in the anterior pi-tuitary. FSH binds to the FSH receptor (FSHr) on Sertoli cellswithin the seminiferous tubules of the testes [22], stimu-lating them to nurture the developing germ cells. The FSHachain is not required for binding to the FSHr, but the FSHbchain specifically binds to the FSHr with high affinity and isrequired for receptor activation [23]. Because of its highexpression in the gonads and very low levels of expressionin other tissues, FSHr provides an elegant targeting mech-anism for drug delivery to the gonads. Indeed, FSHb hasbeen shown to be an effective gonad-specific drug deliveryvehicle for experimental forms of reversible contraceptionin mice. In one such study, the contraceptive efficacy ofadjudin was increased 10,000-fold by conjugation to FSHb[24]. In another example, FSHb conjugated to a peptide thatinterrupted the integrity of the blood–testes barrier causedsignificant loss of germ cells and a decrease in fertility [25].
Use of the entire FSHbmolecule for targeting, however, isimpractical for widespread use because of the expense ofeither the chemical synthesis or recombinant production ofthe entire FSHb chain, which is 112 amino acids in length.Fortunately, FSHb-derived peptides that bind to the FSHrwith high affinity have been described [26–30]. The mostattractive candidates for drug delivery are peptides based on
the amino acids in positions 33 to 53 and 81 to 95 of FSHb[27,28]. Each of these peptides sequences has good in vitroFSHreceptor-binding inhibition and demonstrated in vivoefficacy. Furthermore, there is good recognition of theseconserved peptides by the FSHr across species [27,31]. Pep-tides with terminalmodifications and substitutions of serinefor cysteine increase FSHr binding, as measured by compet-itive inhibition studies [26,30]. These peptides have beenused to target drugs to the gonadal tissue. Both peptidesspecifically targeted ovarian cancer cells with nanoparticles[32,33]. Therefore, we hypothesized that the conjugation ofFSHb-derived peptides to the gonadotoxicant melphalanwould allow for sterilization of male animals while mini-mizing the side effects induced by high-dose melphalanadministered alone. We tested melphalan and FSHb-melphalan conjugates in vitro and in vivo for their ability tokill mouse testicular cells and thereby induce sterility.
2. Material and methods
2.1. Melphalan and FSHb-melphalan conjugate preparation
Melphalan was obtained commercially (Sigma, St. Louis,MO, USA). Two different FSHb-derived peptide sequenceresidues b33 to 53 and b81 to 95 were employed as the car-riers for melphalan. The melphalan was conjugated to eitherthe amino or carboxy terminus of each peptide, with orwithout the presence of terminal modifications and additionor deletion of terminal amino acids. The molecular weightsand FSHb-amino acid composition of the 16 conjugates testedare listed in Table 1. FSHb-melphalan conjugates were ob-tained from RS synthesis (Louisville, KY, USA), Biomatik

J.K. Amory et al. / Theriogenology xxx (2014) 1–8 3
(Wilmington, DE, USA), or SynBioSci (Livermore, CA, USA) andwere prepared using standard Fmoc-based solid-phase pep-tide synthesis followed by High pressure liquid chromotog-raphy purification until greater than 95% purity. Preparationswere free of detectable unconjugated melphalan.
2.2. In vitro cytotoxicity assay and imaging
Melphalan and FSHb-melphalan conjugates were evalu-ated for cytotoxicity using a lactate dehydrogenase (LDH)release assay in a three-dimensional in vitro testicularcoculture model [34]. In brief, the testes of male SpragueDawley rat pups (Harlan, Hayward, CA, USA) were dissectedfrom 5-day-old pups, decapsulated under a dissection mi-croscope, and the seminiferous cords/tubules were pooledand digested in a solution of 0.1% collagenase (WorthingtonBiochemical Corporation, Lakewood, NJ, USA), 0.1% hyal-uronidase (Sigma), DNAse I (0.001 mg/mL; Sigma), andEagle’s minimal essential medium (Life Technologies Inc.,Gaithersburg, MD, USA) for 20 minutes at 37 �C [35,36]. Cellmixtures underwent two rounds of further digestion with0.1% collagenase andDNAse I for 20minutes at 37 �C, before afinal digestion performed by the addition of 0.05% trypsin-EDTA for 3 minutes. Cells were filtered through a 100-mmnylon mesh cell strainer (BD Biosciences, Franklin Lakes,NJ, USA) to yield single cell suspension, which consistsprimarily of Sertoli cells, Leydig cells, and type-A spermato-gonia. Cells were resuspended in hormone-free and serum-free media and plated in 35-mm Primaria (BD Biosciences)dishes at a density of 0.8 � 106 cells/mL in serum-freemedium. An extracellular matrix Matrigel overlay (BD Bio-sciences) was applied directly to the medium, 30 minutesafter seeding at a final concentration of 150-mg/mL (30 mL ofMatrigel/35mmdish)medium. Cells were then placed into a37 �C atmosphere in a humidified chamber with 95%/5% air/CO2 and allowed to acclimate for 48 hours before chemicaltreatment. Melphalan and FSHb-melphalan conjugates weretested atfinal cell culture concentrations of 1, 5, and 20 mmol.For the treatment, culture media was mixed with the stocksolution for each of the constructs or melphalan. The con-structs were prepared in PBS. For the solution of melphalan,3.3 mg of melphalan was suspended in 1 mL ethanol, fol-lowed by an addition of 750 mL DMSO (with vortexing), andthen diluted with 750 mL normal saline to a final volume of2.5 mL. DMSO was added to untreated control cultures. Ateach time point, 50 mL of media was harvested from theculture plates into 96-well plates and used for released LDHcytotoxicity assay. Lactate dehydrogenase is normally acytosolic enzyme, but it is released on cell lysis, and is,therefore, a useful marker of cytotoxicity. Lactate dehydro-genase was measured 24, 48, and 72 hours after treatmentusing CytoTox 96nonradioactive cytotoxicity assay (PromegaCorporation, Madison, WI, USA). The assay results werecalculated relative to the percentage of the maximum LDHreleasedata,whichwasgenerated bya setof threemaximumLDH release controls for each respective time point. In addi-tion, photomicrographs of the cultures were obtained after24, 48, and 72 hours of culture. Finally, cultures were stainedfor anti-Mullerian hormone (AMH), a marker of Sertoli cellsusing anti-AMH primary goat antibody (Santa Cruz Biotech-nology, Dallas, TX, USA) and an Alexa Fluor 488 rabbit
antigoat Immunoglobulin G (Life Technologies, Grand Island,NY, USA) and visualized usingfluorescentmicroscope (NikonInstruments, Melville, NY, USA) and Nuance Imaging Sys-tems, (PerkinElmer, Waltham, MA, USA) after 24, 48, and72 hours of culture.
2.3. The effect of melphalan and FSHb (33–53)-melphalanconjugate on spermatogenesis
Based on the results from the in vitro testing, the FSHb(33–53)-melphalan conjugate (“conjugate F”) was selectedfor testing in male C57/Bl6 mice. A total of 24 male mice,aged 16weeks,were used for this experiment. Sixmice eachwere injected intraperitoneally with: group 1, 7.5 mg/kgmelphalan once on Day 0; group 2, 71mg/kg FSHb (33–53)-melphalan conjugate (F) on Days 0, 3, 5, and 8 (four in-jections); group three, FSHb (33–53)-melphalan conjugate(F) 284 mg/kg once on Day 0; or group four, sterile saline(negative control) once on Day 0. The dose was based on anearlier pilot study. The repeated dose group (group 2) wasincluded to determine if a longer exposure might be supe-rior to the single large dose (group 3). Threemice from eachgroup were sacrificed 4 weeks after the injection(s) fordocumentation of an acute effect on spermatid counts andtesticular histology, and the remaining three mice in eachgroup were sacrificed 12 weeks after the injection(s) todetermine if the effects observed at 4weeks persisted. Bloodwas sent for completeblood counts and routine chemistries.For testicular spermatid counts, one testis was individuallyweighed and homogenized in 0.1 M sodium phosphatebuffer, pH 7.4, containing 0.1% Triton X-100, using an all-glass Kontes 15 mL homogenizer (eight strokes). A total of10 mL of the homogenate was counted using a Neubauerphase contrast hemocytometer. Total homogenization-resistant spermatids per gram and per organ were calcu-lated by correcting for the number of squares counted,dilution, volume, and weight, as described previously[37,38]. In addition, a portion of one testiswasfixed in Bouinsolution and stained with hematoxylin–periodic acid Schiffreagent for morphologic analysis. All procedures involvingthe use of animals and animal tissues were approved by theUniversity of Washington’s Institutional Animal Use andCare Committee before being performed.
2.4. Statistical analysis
The results of the cytotoxicity assay are presented in adescriptive fashion. Comparisons of the testicular weights,spermatid counts, and daily sperm production in betweentreatmentswere performed byaKruskal–Wallis ANOVAwithaWilcoxon rank-sumpost hoc test. Analyseswere performedusing STATA version 10 (StataCorp, College Station, TX, USA).An alpha of less than 0.05 was considered significant.
3. Results
3.1. In vitro cytotoxicity testing of melphalan and FSHb-melphalan conjugates
The results of the cytotoxicity testing of melphalan andthe FSHb-melphalan conjugates (Table 1), as demonstrated
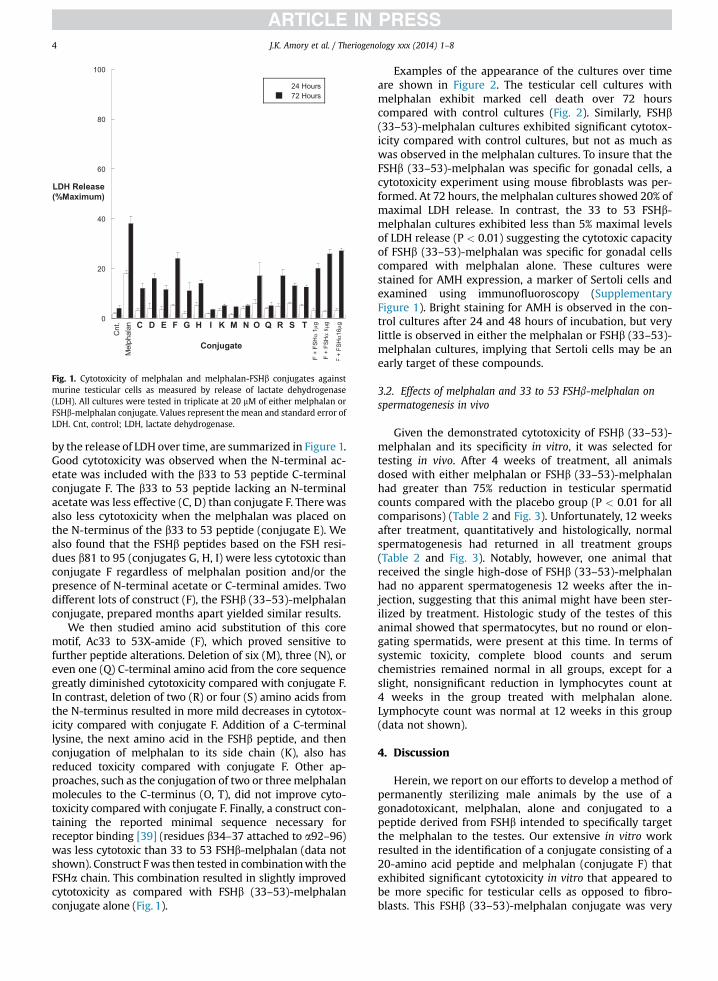
Fig. 1. Cytotoxicity of melphalan and melphalan-FSHb conjugates againstmurine testicular cells as measured by release of lactate dehydrogenase(LDH). All cultures were tested in triplicate at 20 mM of either melphalan orFSHb-melphalan conjugate. Values represent the mean and standard error ofLDH. Cnt, control; LDH, lactate dehydrogenase.
J.K. Amory et al. / Theriogenology xxx (2014) 1–84
by the release of LDH over time, are summarized in Figure 1.Good cytotoxicity was observed when the N-terminal ac-etate was included with the b33 to 53 peptide C-terminalconjugate F. The b33 to 53 peptide lacking an N-terminalacetate was less effective (C, D) than conjugate F. There wasalso less cytotoxicity when the melphalan was placed onthe N-terminus of the b33 to 53 peptide (conjugate E). Wealso found that the FSHb peptides based on the FSH resi-dues b81 to 95 (conjugates G, H, I) were less cytotoxic thanconjugate F regardless of melphalan position and/or thepresence of N-terminal acetate or C-terminal amides. Twodifferent lots of construct (F), the FSHb (33–53)-melphalanconjugate, prepared months apart yielded similar results.
We then studied amino acid substitution of this coremotif, Ac33 to 53X-amide (F), which proved sensitive tofurther peptide alterations. Deletion of six (M), three (N), oreven one (Q) C-terminal amino acid from the core sequencegreatly diminished cytotoxicity compared with conjugate F.In contrast, deletion of two (R) or four (S) amino acids fromthe N-terminus resulted in more mild decreases in cytotox-icity compared with conjugate F. Addition of a C-terminallysine, the next amino acid in the FSHb peptide, and thenconjugation of melphalan to its side chain (K), also hasreduced toxicity compared with conjugate F. Other ap-proaches, such as the conjugation of two or threemelphalanmolecules to the C-terminus (O, T), did not improve cyto-toxicity compared with conjugate F. Finally, a construct con-taining the reported minimal sequence necessary forreceptor binding [39] (residues b34–37 attached to a92–96)was less cytotoxic than 33 to 53 FSHb-melphalan (data notshown). Construct Fwas then tested in combinationwith theFSHa chain. This combination resulted in slightly improvedcytotoxicity as compared with FSHb (33–53)-melphalanconjugate alone (Fig. 1).
Examples of the appearance of the cultures over timeare shown in Figure 2. The testicular cell cultures withmelphalan exhibit marked cell death over 72 hourscompared with control cultures (Fig. 2). Similarly, FSHb(33–53)-melphalan cultures exhibited significant cytotox-icity compared with control cultures, but not as much aswas observed in the melphalan cultures. To insure that theFSHb (33–53)-melphalan was specific for gonadal cells, acytotoxicity experiment using mouse fibroblasts was per-formed. At 72 hours, the melphalan cultures showed 20% ofmaximal LDH release. In contrast, the 33 to 53 FSHb-melphalan cultures exhibited less than 5% maximal levelsof LDH release (P < 0.01) suggesting the cytotoxic capacityof FSHb (33–53)-melphalan was specific for gonadal cellscompared with melphalan alone. These cultures werestained for AMH expression, a marker of Sertoli cells andexamined using immunofluoroscopy (SupplementaryFigure 1). Bright staining for AMH is observed in the con-trol cultures after 24 and 48 hours of incubation, but verylittle is observed in either the melphalan or FSHb (33–53)-melphalan cultures, implying that Sertoli cells may be anearly target of these compounds.
3.2. Effects of melphalan and 33 to 53 FSHb-melphalan onspermatogenesis in vivo
Given the demonstrated cytotoxicity of FSHb (33–53)-melphalan and its specificity in vitro, it was selected fortesting in vivo. After 4 weeks of treatment, all animalsdosed with either melphalan or FSHb (33–53)-melphalanhad greater than 75% reduction in testicular spermatidcounts compared with the placebo group (P < 0.01 for allcomparisons) (Table 2 and Fig. 3). Unfortunately, 12 weeksafter treatment, quantitatively and histologically, normalspermatogenesis had returned in all treatment groups(Table 2 and Fig. 3). Notably, however, one animal thatreceived the single high-dose of FSHb (33–53)-melphalanhad no apparent spermatogenesis 12 weeks after the in-jection, suggesting that this animal might have been ster-ilized by treatment. Histologic study of the testes of thisanimal showed that spermatocytes, but no round or elon-gating spermatids, were present at this time. In terms ofsystemic toxicity, complete blood counts and serumchemistries remained normal in all groups, except for aslight, nonsignificant reduction in lymphocytes count at4 weeks in the group treated with melphalan alone.Lymphocyte count was normal at 12 weeks in this group(data not shown).
4. Discussion
Herein, we report on our efforts to develop a method ofpermanently sterilizing male animals by the use of agonadotoxicant, melphalan, alone and conjugated to apeptide derived from FSHb intended to specifically targetthe melphalan to the testes. Our extensive in vitro workresulted in the identification of a conjugate consisting of a20-amino acid peptide and melphalan (conjugate F) thatexhibited significant cytotoxicity in vitro that appeared tobe more specific for testicular cells as opposed to fibro-blasts. This FSHb (33–53)-melphalan conjugate was very
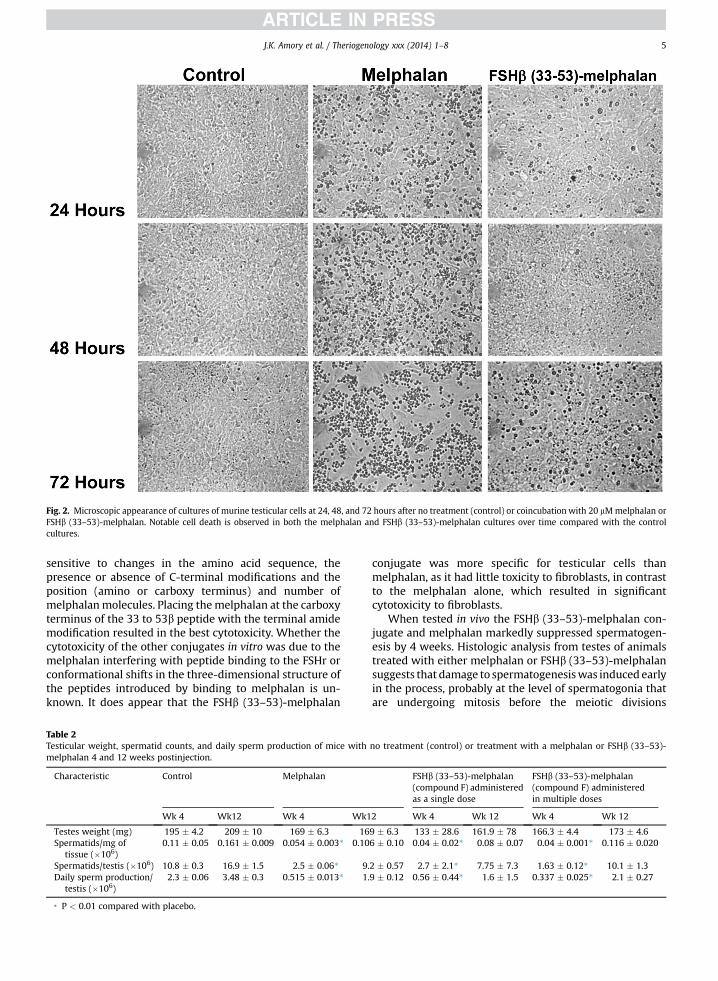
Fig. 2. Microscopic appearance of cultures of murine testicular cells at 24, 48, and 72 hours after no treatment (control) or coincubation with 20 mMmelphalan orFSHb (33–53)-melphalan. Notable cell death is observed in both the melphalan and FSHb (33–53)-melphalan cultures over time compared with the controlcultures.
J.K. Amory et al. / Theriogenology xxx (2014) 1–8 5
sensitive to changes in the amino acid sequence, thepresence or absence of C-terminal modifications and theposition (amino or carboxy terminus) and number ofmelphalanmolecules. Placing themelphalan at the carboxyterminus of the 33 to 53b peptide with the terminal amidemodification resulted in the best cytotoxicity. Whether thecytotoxicity of the other conjugates in vitro was due to themelphalan interfering with peptide binding to the FSHr orconformational shifts in the three-dimensional structure ofthe peptides introduced by binding to melphalan is un-known. It does appear that the FSHb (33–53)-melphalan
Table 2Testicular weight, spermatid counts, and daily sperm production of mice withmelphalan 4 and 12 weeks postinjection.
Characteristic Control Melphalan
Wk 4 Wk12 Wk 4 Wk1
Testes weight (mg) 195 � 4.2 209 � 10 169 � 6.3 16Spermatids/mg of
tissue (�106)0.11 � 0.05 0.161 � 0.009 0.054 � 0.003* 0.10
Spermatids/testis (�106) 10.8 � 0.3 16.9 � 1.5 2.5 � 0.06* 9.Daily sperm production/
testis (�106)2.3 � 0.06 3.48 � 0.3 0.515 � 0.013* 1.
* P < 0.01 compared with placebo.
conjugate was more specific for testicular cells thanmelphalan, as it had little toxicity to fibroblasts, in contrastto the melphalan alone, which resulted in significantcytotoxicity to fibroblasts.
When tested in vivo the FSHb (33–53)-melphalan con-jugate and melphalan markedly suppressed spermatogen-esis by 4 weeks. Histologic analysis from testes of animalstreated with either melphalan or FSHb (33–53)-melphalansuggests that damage to spermatogenesiswas induced earlyin the process, probably at the level of spermatogonia thatare undergoing mitosis before the meiotic divisions
no treatment (control) or treatment with a melphalan or FSHb (33–53)-
FSHb (33–53)-melphalan(compound F) administeredas a single dose
FSHb (33–53)-melphalan(compound F) administeredin multiple doses
2 Wk 4 Wk 12 Wk 4 Wk 12
9 � 6.3 133 � 28.6 161.9 � 78 166.3 � 4.4 173 � 4.66 � 0.10 0.04 � 0.02* 0.08 � 0.07 0.04 � 0.001* 0.116 � 0.020
2 � 0.57 2.7 � 2.1* 7.75 � 7.3 1.63 � 0.12* 10.1 � 1.39 � 0.12 0.56 � 0.44* 1.6 � 1.5 0.337 � 0.025* 2.1 � 0.27

Fig. 3. Murine testis histology. (A) No treatment (control); (B) treatment with a single dose of 33 to 53 FSHb-melphalan; (C and D) treatment with a single dose ofmelphalan. Marked loss of spermatocytes and spermatids is observed at 4 weeks in both the melphalan and 33 to 53 FSHb-melphalan groups. In panel B, a tubulesection with almost complete absence of germ cells and one with few germ cells are shown; some other tubules had more germ cells including condensedspermatids, similar to panels C and D. In panels C and D, complete and quantitatively deficient areas of spermatogenesis are seen. Bouin fixative and Hema-toxylin–Periodic acid Schiff reagent stain. Bars represent 20 microns.
J.K. Amory et al. / Theriogenology xxx (2014) 1–86
necessary to make mature spermatozoa. This would makesense, given the known mechanisms of action of nitrogenmustard alkylating agents that target cells undergoingmitosis [40]. Likely, the postmitotic germ cells are unaf-fected and complete spermatogenesis, leaving only Sertolicells and undifferentiating spermatogonia in many of thetubules 4 weeks after treatment. These unaffected sper-matogonia then begin dividing and appear to successfullyrepopulate many of the tubules 12 weeks after treatment.However, some areas remain depleted even after 12 weeks,with only Sertoli cells present on histologic examination,implying thatmore severe injury to the spermatogoniamayhave occurred in these tubules. However, most of the tu-bules appear to have recovered 12 weeks after the injectionof melphalan or the FSHb (33–53)-melphalan. This suggeststhat most of the undifferentiated spermatogonia seem to beresistant to theeffects of themelphalan, and these cells forma “reservoir” of stem cells that can repopulate the seminif-erous tubules after injury. This finding has significant im-plications for efforts to develop a nonsurgical method ofsterilizing animals, as it suggests that all spermatogoniamust be destroyed to effectively sterilize an animal andprevent recovery of spermatogenesis.Whether a largerdoseof the FSHb (33–53)-melphalan would accomplish this asobserved in one of the mice will be the subject of futurestudy. To date, only certain types of chemotherapy (e.g.,busulfan), pesticides, and ionizing radiation have beendescribed tohave this effect. Busulfan is of some interest as itis one of the most potent gonadotoxicants known and isroutinely used to sterilize mice [41]. Unfortunately, thechemical structure of busulfan did not allow for linkage toFSHb (33–53), and it is too toxicwhen administered alone in
terms of bone marrow and neurologic toxicity to beconsidered for sterilizing larger captive and wild animals[42–44]. Indeed, melphalan is unique among gonadotox-icants in its ability to be conjugated to FSHb (33–53), likelydue to the fact that it is an amino acid derivative. Theseapproaches make them impractical for nonsurgical sterili-zation. A better understanding of the mechanisms of spermstem cell self-renewalmay be required before a nonsurgicalapproach to male sterilization can be safe and practical.
The correlation between the in vitro and observed in vivoeffects on spermatogenesis validates the utility of the three-dimensional testicular coculture system for detectingcompounds that are likely to have deleterious effects onspermatogenesis in vivo. This system is relatively inexpen-sive and sensitive and has been used to determine the toxiceffects of a variety of compounds [34,35]. It must be noted,however, that this approach to screening compoundsin vitro has a limitation in that the behavior of Sertoli cellsin vitro may be different from their behavior in vivo. Sertolicells in culture can proliferate [41], and therefore, may bemore sensitive to the effects of melphalan compared withSertoli cells in vivo, which are nonreplicating. With thesecaveats, future work directed at developing novel com-pounds for male sterility could use this system to screencandidate molecules before in vivo testing.
One limitation of our study is that we cannot determineif the reduction in spermatogenesis observed in the ani-mals treated with FSHb (33–53)-melphalan was due tospecific targeting of the conjugate to the testes as opposedto serving as a “carrier” for the melphalan, which was thencleaved from the peptide after in vivo administration. Intheory, release of the melphalan before binding of FSHb

J.K. Amory et al. / Theriogenology xxx (2014) 1–8 7
(33–53)-melphalan to the FSHr could account for thetesticular toxicity similar to that observed in themelphalan-treated animals. However, we believe this isunlikely due to the remarkable resistance of the fibroblaststo FSHb (33–53)-melphalan compared with melphalanalone. Future studies using labeled FSHb (33–53)-melphalan will be required to understand whether its ef-fect on spermatogenesis is secondary to specific targeting.More broadly, it is uncertain if melphalan needs to bereleased from its peptide carrier to function [42]. Priorwork on melphalan–cytotoxin conjugates suggests that C-terminal conjugation of a peptide with the melphalanamine does not alter the efficacy of the melphalan implyingthat release from the FSHb-derived peptide may not benecessary to damage the germ cells [43]. In theory, FSHb(33–53)-melphalan should share the same biological fate asFSH, which localizes to the plasma membrane of FSHr-bearing cells before accumulating in the endosomalcompartment before degradation in the lysosomes [44].This should then liberate the melphalan into the cytoplasmof the Sertoli cell. However, germ cell toxicity was moreevident histologically than Sertoli cell death. Therefore,either the abovemechanism is incorrect, or the Sertoli cells,which are nondividing in the adult were not susceptible tothe effects of the melphalan. In this case, the toxicity of themelphalan to the germ cells could either occur fromtransfer of the melphalan directly to the germ cells fromSertoli cells, or by passive diffusion within the basalcompartment of the testis. Understanding this process willrequire additional studies using fluorescent-labeled FSHb(33–53)-melphalan to better define its cellular trafficking.
Notably, no severe systemic toxicity was observed fromeither treatment except for a transient nonsignificantdecrease in the white blood cell concentration in the ani-mals receiving melphalan alone. Bone marrow toxicitywith melphalan with larger doses is well described, and isdose limited in the treatment of hematological cancers.Given the small sample size in our study, we likely lackedthe power to detect a significant difference in this endpoint. Moreover, repeated dosing of melphalan may haveresulted in a larger effect on hematopoiesis.
In summary, we have found that melphalan and FSHb(33–53)-melphalan are potent gonadotoxicants in vitro andin vivo. Both effectively but not completely suppress sper-matogenesis for 4 weeks after a single administration.However, spermatogenesis mostly recovers within 12weeksof treatment in most animals. This approach to sterilizinganimals requires refinement to become more uniformlyeffective. In addition, a greater understanding of the biologyof sperm stem cell self-renewal would be very helpful in ef-forts to develop a nonsurgicalmethod of animal sterilization.
Acknowledgments
The authors would like to thank Erin Pagel MA forassistance with the sperm analysis and David W. AmoryMD, PhD for thoughtful review of the article. This work wasfunded by the University of Washington Royalty ResearchFund, and in part, by the Eunice Kennedy Shriver NationalInstitute of Child Health and Human Development throughcooperative agreement U54 HD-42454 as part of the
Cooperative Contraceptive Research Centers Program andfunds from Focused Scientific, Inc. as well as Grants 1 U01FD004242 and CEEH: 5 P30 ES007033 from the US FDA.
Competing interests
The authors have nothing to disclose.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.theriogenology.2014.03.014.
References
[1] Katzler M, Wood A. Non-surgical methods of contraception andsterilization. Theriogenology 2006;66:514–25.
[2] Salmeri KR, Bloomberg MS, Scruggs SL, Shille V. Gonadectomy inimmature dogs: effects on skeletal, physical and behavioral devel-opment. J Am Vet Med Assoc 1991;198:1193–203.
[3] Bell FW, Klausner JS, Hayden DW, Feeney DA, Johnston SD. Clinicaland pathologic features of prostatic adenocarcinoma in sexuallyintact and castrated dogs: 31 cases (1970-1987). J Am Vet MedAssoc 1991;199:1623–30.
[4] Obradovich J, Walsaw R, Goulland E. The influence of castration onthe development of prostatic carcinoma in the dog, 43 cases (1978-1985). J Vet Intern Med 1987;1:183–7.
[5] Johnston SD, Kamolpatana K, Root-Kustritz MV, Johnston GR.Prostatic disorders in the dog. Anim Reprod Sci 2000;60-61:405–15.
[6] Fukuda S, Lida H. Effects of orchidectomy on bone metabolism inbeagle dogs. J Vet Med Sci 2000;62:69–73.
[7] Jana K, Samanta PK. Sterilization of male stray dogs with a singleintratesticular injection of calcium chloride: a dose-dependentstudy. Contraception 2007;75:390–400.
[8] Oliverira EC, Moura MR, Silva Jr VA, Peixoto CA, Saraiva KL, de Sa MJ,et al. Intratesticular injection of a zinc-based solution as a contra-ceptive for dogs. Theriogenology 2007;68:137–45.
[9] Levy JK, Crawford PC, Appel LD, Clifford EL. Comparison of intra-testicular injection of zinc gluconate versus surgical castration tosterilize male dogs. Am J Vet Res 2008;69:140–3.
[10] Tepsumethanaon V, Wilde H, Hemachudha T. Intratesticular injec-tion of a balanced zinc solution for permanent sterilization of dogs. JMed Assoc Thai 2005;88:686–9.
[11] Vickery BH, Burns J, Zaneveld LJ, Goodpasture JC, Bergstrom K.Orally administered ketoconazole rapidly appears in seminalplasma and suppresses sperm motility. Adv Contracept 1985;1:341–53.
[12] Dixit VP, Lohiya NK, Arya M. Reversible changes in the testes andepididymides of dogs treated with alpha-chlorohydrin. Acta BiolMed Ger 1975;34:1851–6.
[13] Dixit VP, Bhargava SK. Reversible contraception like activity ofembelin in male dogs (Canis indicus Linn). Andrologia 1983;15:486–94.
[14] Hieberg JK, Svejgaard E. Toxic hepatitis during ketoconazole treat-ment. Br Med J 1981;283:825–6.
[15] Howell SJ, Shalet SM. Spermatogenesis after cancer treatment:damage and recovery. J Natl Cancer Inst Monogr 2005;34:12–7.
[16] Clark ST, Radford JA, Crowther D, Swindell R, Shalet SM. Gonadalfunction following chemotherapy for Hodgkin’s disease: acomparative study of MVPP and a seven-drug hybrid regimen. Eur JCancer 1991;27:1389–92.
[17] Russell LB, Hunsicker PR, Shelby MD. Melphalan, a second chemicalfor which specific-locus mutation induction in the mouse ismaximum in early spermatids. Mutat Res 1992;282:151–8.
[18] Mellor PJ, Polton GA, Brearly M. Solitary plasmacytoma ofbone in two successfully treated cats. J Feline Med Surg 2007;9:72–7.
[19] Fujino Y, Sawamura S, Kurakawa N. Treatment of chronic lympho-cytic leukemia in three dogs with melphalan and prednisolone. JSmall Anim Pract 2004;45:298–303.
[20] Furner RL, Brown RK, Duncan G. Pharmacokinetics of the absorp-tion, distribution and elimination of melphalan in the dog. CancerTreat Rep 1977;61:1637–46.

J.K. Amory et al. / Theriogenology xxx (2014) 1–88
[21] Page RL, Macy DW, Thrall DE. Unexpected toxicity associated withuse of body surface area for dosing melphalan in the dog. Cancer Res1988;48:288–90.
[22] Heckert LL, Griswold MD. Expression of follicle-stimulating hor-mone receptor mRNA in rat testes and Sertoli cells. Mol Endocrinol1991;5:670–7.
[23] Fan QR, Hendrickson WA. Structure of human follicle-stimulatinghormone in complex with its receptor. Nature 2005;433:269–77.
[24] Mruk DD, Wong C-H, Silvestrini B, Cheng CY. A male contraceptivetargeting germ cell adhesion. Nat Med 2006;12:1323–8.
[25] Wong CH, Mruk DD, Lee WM, Cheng CY. Targeted and reversibledisruption of the blood-testis barrier by and FSH mutant-occludinpeptide conjugate. FASEB J 2007;21:438–48.
[26] Reichert LE. Synthetic peptides corresponding to segments of theFSH-beta subunit have in vivo biologic effects in male and femalemice: a novel and surprising finding. Recent Res Devel Endocrinol2000;1:197–208.
[27] Santa Coloma TA, Dattatreyamurty B, Reichert Jr LE. A syntheticpeptide corresponding to human FSH beta-subunit 33-53 binds tothe FSH receptor, stimulates basal estradiol biosynthesis, and is apartial antagonist of FSH. Biochemistry 1990;29:1194–200.
[28] Santa Coloma TA, Reichert Jr LE. Identification of a follicle-stimulating hormone receptor-binding region in hFSH-beta (81-95) using synthetic peptides. J Biol Chem 1990;265:5037–42.
[29] Grasso P, Rozhavskaya M, Reichart Jr LE. In vivo effects of humanfollicle-stimulating hormone-related synthetic peptide hFSH-beta(81-95) and its subdomain hFSH-beta-(90-95) on the mouseestrous cycle. Biol Reprod 1998;58:821–5.
[30] Santa-Coloma TA, Crabb JW, Reichert Jr LE. Serine analogues ofhFSH-beta-(33-53) and hFSH-beta-(81-95) inhibit hFSH binding toreceptor. Biochem Biophys Res Commun 1992;184:1273–9.
[31] Reichert Jr LE, Dattatreyamurty B. The follicle-stimulating hormone(FSH) receptor in testis: interaction with FSH, mechanism of signaltransduction, and properties of the purified receptor. Biol Reprod1989;40:13–26.
[32] Zhang X, Chen J, Kang Y, Hong S, Zheng Y, Sun H, et al. Targetedpaclitaxel nanoparticles modified with follicle-stimulating hormoneb 81-95 peptide show effective antitumor activity against ovariancarcinoma. Int J Pharm 2013;453:498–505.
[33] Zhang XY, Chen J, Zheng YF, Gao XL, Kang Y, Jiu JC, et al. Fol-licle-stimulating hormone peptide can facilitate paclitaxelnanoparticles to target ovarian carcinoma in vivo. Cancer Res2009;69:6506–14.
[34] Wegner S, Hong S, Yu X, Faustman EM. Preparation of rodent testisco-cultures. Curr Protoc Toxicol 2013 Chapter 16, 60–192.
[35] Hadley MA, Byers SW, Suarez-Quian CA, Kleinman HK, Dym M.Extracellular matrix regulates Sertoli cell differentiation, testicularcord formation, and germ cell development in vitro. J Cell Biol 1985;101:1511–22.
[36] Orth J, McGuinness M, Qiu J, Jester Jr WF, Li L. Use of in vitro systemsto study male germ cell development in neonatal rats. Ther-iogenology 1998;49:431–9.
[37] Amann RP. Detection of alterations in testicular and epididymalfunction in laboratory animals. Env Health Perspect 1986;70:149–58.
[38] Petersen PM, Givercman A, Pakkenberg B. Stereological methods asefficient and unbiased tools to quantitated structures in the testis.Scan J Work Environ Health 1999;25:31–3.
[39] Hage-van Noort M, Puijk WC, Plasman HH, Kuperus D,Schaaper WM. Synthetic peptides based upon a three-dimensionalmodel for the receptor recognition site of follicle-stimulating hor-mone exhibit antagonistic or agonistic activity at low concentra-tions. Proc Natl Acad Sci U S A 1992;89:3922–6.
[40] Samuels BL, Bitran JD. High-dose intravenous melphalan: a review. JClin Oncol 1995;13:1786–99.
[41] Wang DZ, Zhou XH, Yuan YL, Zheng XM. Optimal dose of busulfanfor depleting testicular germ cells of recipient mice before sper-matogonial transplantation. Asian J Androl 2010;12:263–70.
[42] Abkowitz JL, Lindenberger ML, Persik M, Newton MA, Guttop P.Behavior of feline hematopoietic stem cells years after busulfanexposure. Blood 1993;82:2096–103.
[43] Deeg HJ, Schuler U, Shulman H, Ehrsam M, Renner U, Yu C, et al.Myeloablation by intravenous busulfan and hematopoietic recon-stitution with autologous marrow in a canine model. Biol BloodMarrow Transplant 1999;5:316–21.
[44] Tagirov M, Golovan. The effect of busulfan treatment on endoge-nous spermatogonial stem cells in immature roosters. Poult Sci2012;91:1680–5.

![Case Report Initial Biological Evaluations of [18F]KS-7-51 to … · 2020. 9. 22. · and initial biological evaluations of [18F]KS-7-51, a p-fluoroethoxy phenyl derivative in a murine](https://static.fdocument.org/doc/165x107/601e58f23cdaba46814221b9/case-report-initial-biological-evaluations-of-18fks-7-51-to-2020-9-22-and.jpg)