
TCP Increase/Decrease Behavior with Exp licit Congestion Notification (ECN)

Cell, Vol. 12, 609-617, November 1977, Copyright 0 1977 by MIT
Localization of Serum-Derived a2 Macroglobulin in Cultured Cells and Decrease after Moloney Sarcoma Virus Transformation
ha Pastan, Mark Willingham, Wayne Anderson and Maria Gallo Laboratory of Molecular Biology National Cancer Institute Bethesda, Maryland 20014
Summary
NRK cells and many other cultured fibroblasts were found to contain the protease inhibitor, (Ye macroglobulin (+M). This o#l is present as a result of uptake of cu,M from the calf serum in the culture medium. Some of this auzM is released back into the medium. In radiolabeling experi- ments with ‘%-amino acids, no radioactivity was detected in intracellular or extracellular (YAM. Flu- orescence microscopy of fixed cells using rhoda- mine-labeled antibodies indicated that (YAM is present in vesicular organelles different from pri- mary lysosomes. Fluorescence microscopy of liv- ing cells shows that rhodamine-labeled cwzM (rho- damine-rY,M) is taken up into similar structures. Of the many cell lines examined, Moloney sar- coma virus-transformed cells had the lowest amounts of (YAM. Some of the effects of serum on the behavior of cultured cells could be a conse- quence of inhibition of cellular proteases by CQM.
Introduction
The analysis of the protein composition of trans- formed fibroblastic cells by SDS gel electrophore- sis has yielded interesting differences between normal and transformed cells (Nicolson, 1976; Ya- mada and Pastan, 1976). Two proteins that are decreased in many transformed cells are LETS and collagen (reviewed in Yamada and Pastan, 1976; Adams, et al., 1977). LETS is found mainly on the cell surface, although there is also some released into the medium. Collagen is found mainly in the medium. While investigating the proteins released into the medium by normal cells, we noted that one prominent protein band failed to become la- beled with radioactivity when the cells were incu- bated with j4C-amino acids. This protein had a subunit molecular weight of approximately 185,000 daltons. Since the cultured cells were propagated in a medium containing calf serum, it seemed probable that the nonradioactive band was derived from calf serum. A major serum protein with a subunit molecular weight of approximately 185,000 daltons is (Y* macroglobulin [The molecular weight of the intact molecule is 780,000 daltons (Frenoy, et al., 1977).] In this paper we present evidence that cultured fibroblastic cells contain arpM that is
derived from serum; part of this is released back into the medium. Within the cells, azM is concen- trated in certain vesicular structures. We also re- port that cell lines transformed by Moloney sar- coma virus (MoSV), but not by some other agents, have a markedly diminished content of CQM.
Previously Mosher and Wing (1976) investigated embryonic human lung fibroblasts (line W138) and reported that WI38 cells synthesize apM. In normal rat kidney (NRK) cells, we did not detect CQM syn- thesis. Possible reasons for the difference between the results of Mosher and Wing (1976) and our own appear below.
Results
To investigate the nature of proteins released into the medium by cultured cells, we grew NRK cells in medium containing 10% calf serum, washed them 5 times with serum-free medium and then placed them in serum-free medium for 16 hr. We removed the medium, centrifuged it at 100,000 X g for 30 min to remove formed elements and ana- lyzed it by SDS gel electrophoresis. Figure 1A shows the pattern of the high molecular proteins found on a 5% gel. A prominent band with a molecular weight of about 185,000 daltons is evi- dent. The 185,000 molecular weight protein cannot be an intracellular protein released due to cell lysis because other intracellular proteins were not present. For example, the heavy chain of myosin which has a molecular weight of 200,000 daltons, and constitutes up to 5% of the total protein of NRK cells (Ostlund and Pastan, 1976), would have been very evident if there were significant cell lysis. Cultured human fibroblasts have previously been reported to make (Ye macroglobulin (Mosher and Wing, 1976), a protein with asubunit molecular weight of about 185,000 daltons (Harpel, 1973). Fibroblasts also make and release a form of pro- collagen that has a similar molecular weight (Ya- mada and Weston, 1974). To identify the nature and source of the prominent 185,000 dalton band, we labeled cells for 2 hr with a mixture of 14C- amino acids, washed the cells, and collected the labeled proteins that were released into the me- dium over the ensuing 4 hr and analyzed them by SDS gel electrophoresis and autoradiography. Two dark bands were evident by autoradiography that have molecular weights of 185,000 and 160,000 daltons (Figure 1C). Both are completely destroyed by brief treatment with collagenase (Figure 1D). Under these conditions, collagenase did not hy- drolyze LETS, myosin and other intracellular pro- teins (data not shown). Collagenase has been reported to hydrolyze aZM into two fragments with molecular weights of about 85,000 daltons (Werb
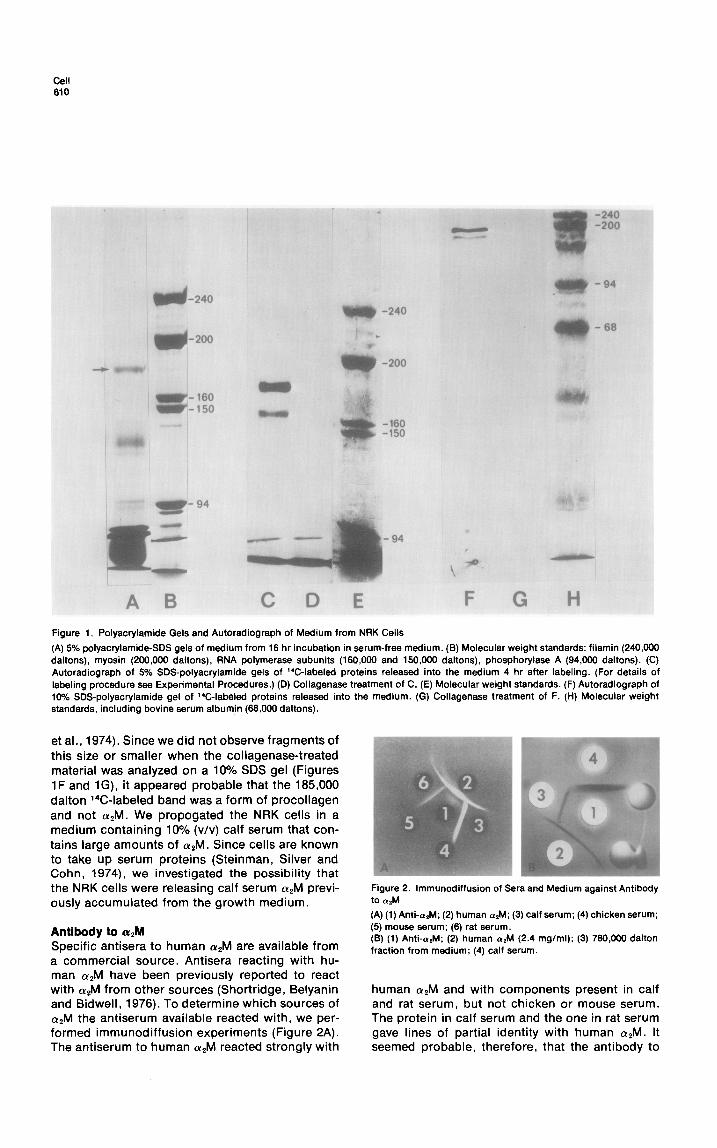
cell 610
Figure 1. Polyacrylamide Gels and Autoradiograph of Medium from NRK Cells
(A) 5% polyacrylamide-SDS gels of medium from 16 hr incubation in serum-free medium. (B) Molecular weight standards: filamin (240,000 daltons), myosin (200,000 daltons), RNA polymerase subunits (160,000 and 150,000 daltons), phosphorylase A (94,000 daltons). (C) Autoradiograph of 5% SDS-polyacrylamide gels of 14C-labeled proteins released into the medium 4 hr after labeling. (For details of labeling procedure see Experimental Procedures.) (D) Collagenase treatment of C. (E) Molecular weight standards. (F) Autoradiograph of 10% SDS-polyacrylamide gel of 14C-labeled proteins released into the medium. (G) Collagenase treatment of F. (H) Molecular weight standards, including bovine serum albumin (66,000 daltons).
et al., 1974). Since we did not observe fragments of this size or smaller when the collagenase-treated material was analyzed on a 10% SDS gel (Figures 1F and lG), it appeared probable that the 185,000 dalton X-labeled band was a form of procollagen and not (YAM. We propogated the NRK cells in a medium containing 10% (v/v) calf serum that con- tains large amounts of aZM. Since cells are known to take up serum proteins (Steinman, Silver and Cohn, 1974), we investigated the possibility that the NRK cells were releasing calf serum OL*M previ- ously accumulated from the growth medium.
Figure 2. lmmunodiffusion of Sera and Medium against Antibody to a&A
(A) (1) Anti-rrtM; (2) human a&f; (3) calf serum: (4) chicken serum:
Antibody to a# Specific antisera to human (Y*M are available from a commercial source. Antisera reacting with hu- man (Y*M have been previously reported to react with CQM from other sources (Shortridge, Belyanin and Bidwell, 1976). To determine which sources of apM the antiserum available reacted with, we per- formed immunodiffusion experiments (Figure 2A). The antiserum to human ol,M reacted strongly with
(5) mouse serum; (6) rat serum. (8) (1) Anti-aeM; (2) human nlM (2.4 mglml); (3) 760,000 dalton fraction from medium; (4) calf serum.
human apM and with components present in calf and rat serum, but not chicken or mouse serum. The protein in calf serum and the one in rat serum gave lines of partial identity with human CQM. It seemed probable, therefore, that the antibody to
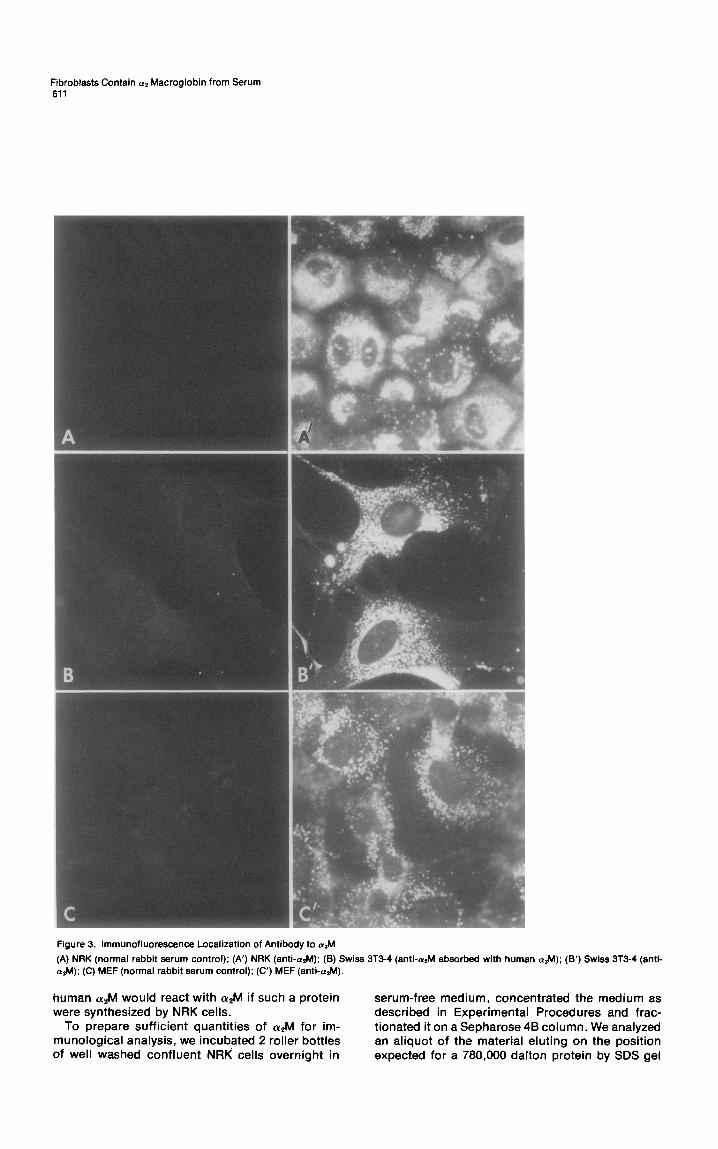
Fibroblasts Contain al Macroglobin from Serum 611
Figure 3. lmmunofluorescence Localization of Antibody to a&
(A) NRK (normal rabbit serum control); (A’) NRK (anti-&l); (6) Swiss 3T3-4 (anti-&I absorbed with human aaM); (6’) Swiss 3T3-4 atM); (C) MEF (normal rabbit serum control); (C’) MEF (anti-a.M).
human azM would react with CQM if such a protein serum-free medium, concentrated the mediun were synthesized by NRK cells. described in Experimental Procedures and f
To prepare sufficient quantities of (Y*M for im- tionated it on a Sepharose 4B column. We anal) munological analysis, we incubated 2 roller bottles an aliquot of the material eluting on the posi of well washed confluent NRK cells overnight in expected for a 780,000 dalton protein by SDS
n as ‘rat- lzed Ition gel
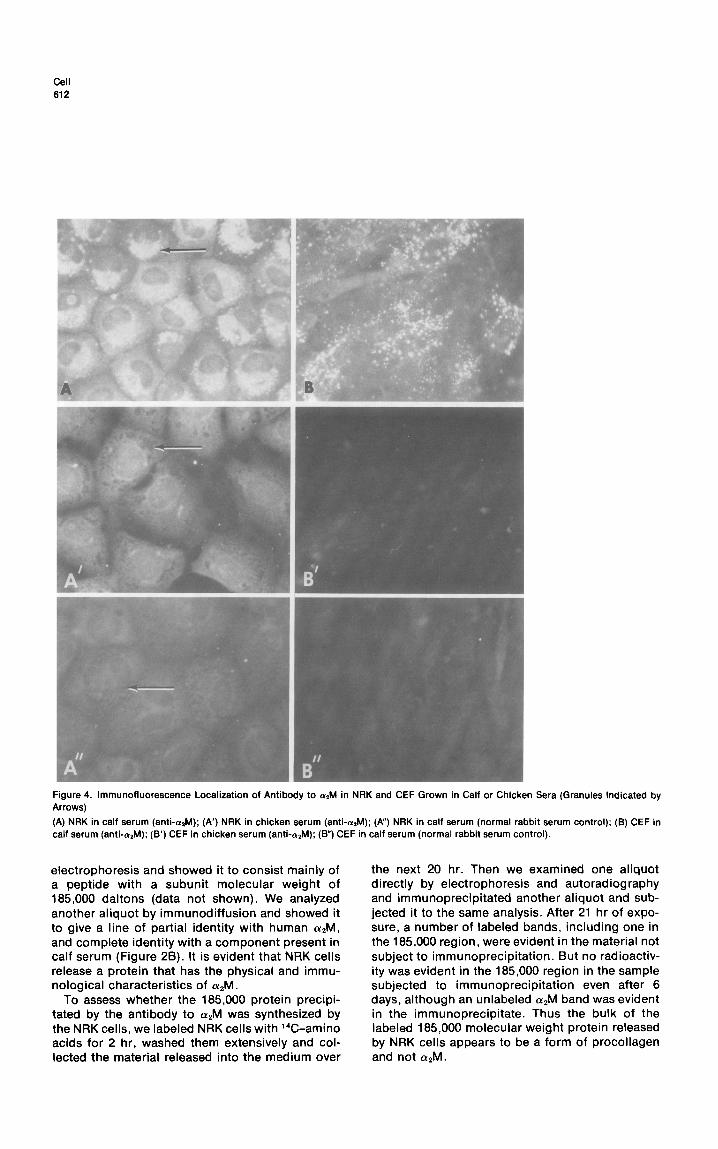
Cell 612
Figure 4. lmmunofluorescence Localization of Antibody to alM in NRK and CEF Grown in Calf or Chicken Sera (Granules Indicated by Arrows)
(A) NRK in calf serum (anti-&l); (A’) NRK in chicken serum (anti-arM); (A”) NRK in calf serum (normal rabbit serum control); (8) CEF in calf serum (anti-&J); (B’) CEF in chicken serum (anti-&l); (B”) CEF in calf serum (normal rabbit serum control).
electrophoresis and showed it to consist mainly of a peptide with a subunit molecular weight of 185,000 daltons (data not shown). We analyzed another aliquot by immunodiffusion and showed it to give a line of partial identity with human (YAM, and complete identity with a component present in calf serum (Figure 28). It is evident that NRK cells release a protein that has the physical and immu- nological characteristics of CX~M.
To assess whether the 185,000 protein precipi- tated by the antibody to a&l was synthesized by the NRK cells, we labeled NRK cells with %-amino acids for 2 hr, washed them extensively and col- lected the material released into the medium over
the next 20 hr. Then we examined one aliquot directly by electrophoresis and autoradiography and immunoprecipitated another aliquot and sub- jected it to the same analysis. After 21 hr of expo- sure, a number of labeled bands, including one in the 185,000 region, were evident in the material not subject to immunoprecipitation. But no radioactiv- ity was evident in the 185,000 region in the sample subjected to immunoprecipitation even after 6 days, although an unlabeled asM band was evident in the immunoprecipitate. Thus the bulk of the labeled 185,000 molecular weight protein released by NRK cells appears to be a form of procollagen and not CQM.
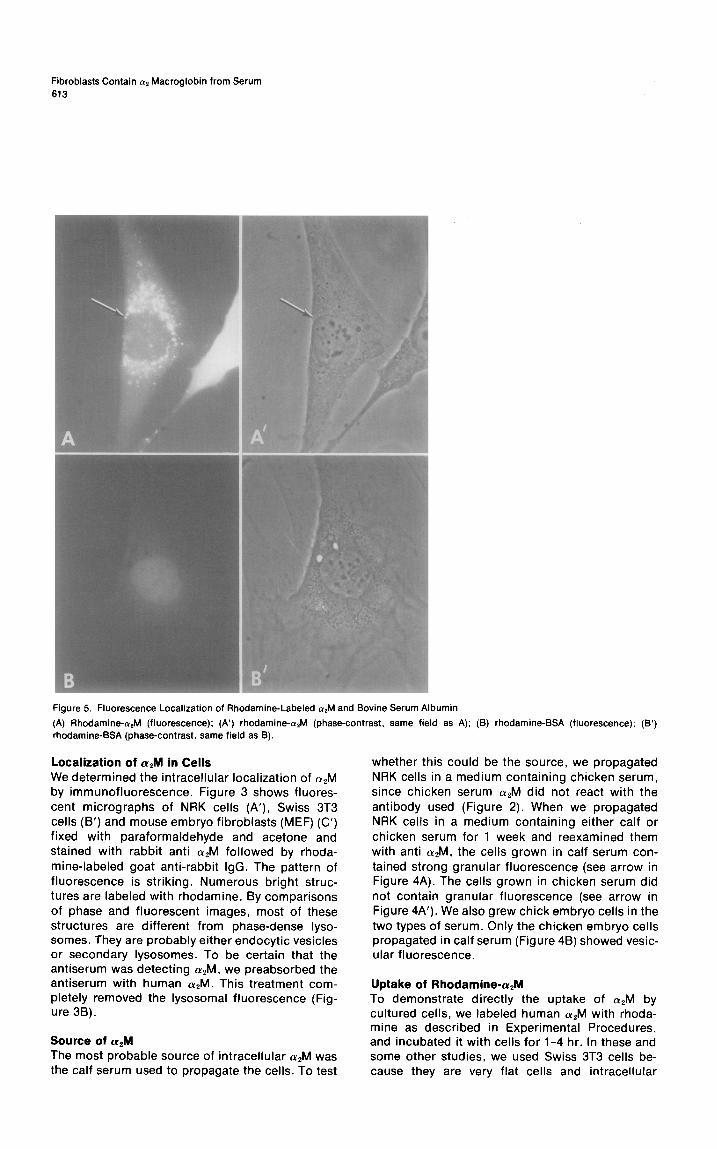
Fibroblasts Contain a2 Macroglobin from Serum 613
Figure 5. Fluorescence Localization of Rhodamine-Labeled &I and Bovine Serum Albumin
(A) Rhodamine-alM (fluorescence): (A’) rhodamine- (phase-contrast, same field as A); (B) rhodamine-BSA (fluorescence); rhodamine-BSA (phase-contrast, same field as 8).
Localization of cw,M in Cells We determined the intracellular localization of (YAM by immunofluorescence. Figure 3 shows fluores- cent micrographs of NRK cells (A’), Swiss 3T3 cells (B’) and mouse embryo fibroblasts (MEF) (C’) fixed with paraformaldehyde and acetone and stained with rabbit anti CQM followed by rhoda- mine-labeled goat anti-rabbit IgG. The pattern of fluorescence is striking. Numerous bright struc- tures are labeled with rhodamine. By comparisons of phase and fluorescent images, most of these structures are different from phase-dense lyso- somes. They are probably either endocytic vesicles or secondary lysosomes. To be certain that the antiserum was detecting apM, we preabsorbed the antiserum with human (YAM. This treatment com- pletely removed the lysosomal fluorescence (Fig- ure 3B).
Source of a# The most probable source of intracellular (Y*M was the calf serum used to propagate the cells. To test
W
whether this could be the source, we propagated NRK cells in a medium containing chicken serum, since chicken serum azM did not react with the antibody used (Figure 2). When we propagated NRK cells in a medium containing either calf or chicken serum for 1 week and reexamined them with anti cwzM, the cells grown in calf serum con- tained strong granular fluorescence (see arrow in Figure 4A). The cells grown in chicken serum did not contain granular fluorescence (see arrow in Figure 4A’). We also grew chick embryo cells in the two types of serum. Only the chicken embryo cells propagated in calf serum (Figure 48) showed vesic- ular fluorescence.
Uptake of Rhodamine-a,M To demonstrate directly the uptake of apM by cultured cells, we labeled human cvzM with rhoda- mine as described in Experimental Procedures, and incubated it with cells for l-4 hr. In these and some other studies, we used Swiss 3T3 cells be- cause they are very flat cells and intracellular
Cell
614
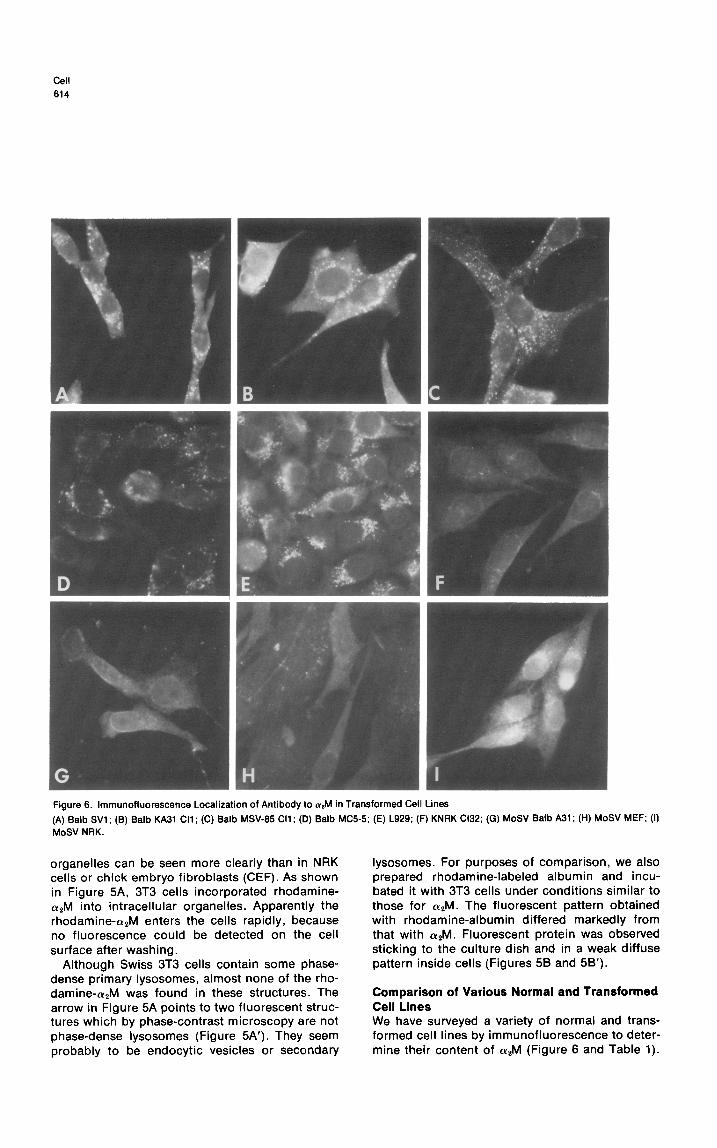
Figure 6. lmmunofluorescence Localization of Antibody to arM in Transformed Cell Lines
(A) Balb WI ; (6) Balb KA31 Cl1 ; (C) Balb MSV-65 Cl1 ; (D) Balb MC55; (E) L929; (F) KNRK Cl32; (G) MoSV Balb A31 ; (H) MoSV MEF; (I) MoSV NRK.
organelles can be seen more clearly than in NRK cells or chick embryo fibroblasts (CEF). As shown in Figure 5A, 3T3 cells incorporated rhodamine- CQM into intracellular organelles. Apparently the rhodamine-cY,M enters the cells rapidly, because no fluorescence could be detected on the cell surface after washing.
Although Swiss 3T3 cells contain some phase- dense primary lysosomes, almost none of the rho- damine-a,M was found in these structures. The arrow in Figure 5A points to two fluorescent struc- tures which by phase-contrast microscopy are not phase-dense lysosomes (Figure 5A’). They seem probably to be endocytic vesicles or secondary
lysosomes. For purposes of comparison, we also prepared rhodamine-labeled albumin and incu- bated it with 3T3 cells under conditions similar to those for CQM. The fluorescent pattern obtained with rhodamine-albumin differed markedly from that with crzM. Fluorescent protein was observed sticking to the culture dish and in a weak diffuse pattern inside cells (Figures 58 and 5B’).
Comparison of Various Normal and Transformed Cell Lines We have surveyed a variety of normal and trans- formed cell lines by immunofluorescence to deter- mine their content of a2M (Figure 6 and Table 1).

Fibroblasts Contain 0% Macroglobin from Serum 615
Table 1. Intensity of Fluorescence of a,M in Normal and Transformed Cells
Intensity of Fluorescence
Cell Class Strong Weak
Normal NRK
Balb 3T3
Swiss 3T3
MEF
CEF
Viral Transformants SV40 Balb (SVl)
Ki Balb (KA31)
RSV-SR CEF
MoSV NRK
MoSV Balb
MoSV MEF
KNRK Cl32
Other Transformants MC5-5
L929
MSV-65 Cl1
When NRK cells, Balb 3T3 cells, or mouse embryo fibroblasts were transformed by the Moloney strain of murine sarcoma virus, they all had a greatly diminished content of a2M. On the other hand, many transformed cell lines, including those trans- formed by SV40, by Schmidt-Ruppin strain of Rous sarcoma virus (SR-RSV), by methylcholanthrene and spontaneously had strong a2M fluorescence. Two cell lines transformed by the Kirsten strain of murine sarcoma virus (KiSV) have been examined so far. One has definitely diminished levels of apM; the other does not.
Discussion
In this paper we have demonstrated that various types of cultured fibroblastic cells contain cuzM that they acquire from the culture medium. We used two approaches to identify where a2M is located within cells. One was to stain the protein by fluorescently labeled antibodies after fixation of the cells. The second was to incubate the cells with rhodamine-labeled a2M and to observe where this material becomes located. Both types of stud- ies yield the same result. The protein is principally found in structures that appear to be secondary lysosomes or endocytic vesicles. Some of this intracellular (Y*M is released when cells are placed in serum-free medium.
Previously, Mosher and Wing (1976) reported that human lung fibroblasts synthesized a2M and released it into the medium. We find that the intracellular CQM evident by fluorescence in many cell types is derived from the serum in which the cells are grown. In studies in which cellular pro- teins of rat fibroblast line NRK were labeled with
‘4C-amino acids, and the proteins released into the medium precipitated with antibody to a2M, we were unable to detect any radioactivity associated with a2M. We have also been unable to detect labeled a2M in cell extracts by immunoprecipita- tion. Thus if NRK cells make azM, the amount must be very small. Since Mosher and Wing (1976) studied QM synthesis in human lung fibroblasts, and we studied rat NRK fibroblasts, the results are not directly comparable. It is possible that the cells differ in the proteins they synthesize. It is also possible that NRK cells make very small amounts of a2M that escaped detection by the methods employed. Another difference between the two studies is in the method of analysis of (YAM. In the studies of Mosher and Wing (1976), WI38 cells were labeled with 35S-L-methionine, and the se- creted proteins were subjected to immunodiffusion in 1% agarose followed by autoradiography. Pro- teases synthesized by the WI38 cells would be labeled with 35S-methionine; they could bind to serum-derived a2M and precipitate in a single com- plex with antibody to apM. Analysis of the immu- noprecipitate by SDS gel electrophoresis would clarify this possibility.
Many substances that are taken up by cells enter lysosomes and are degraded. (Y*M, however, is readily detected in “lysosomal” structures by im- munofluorescence. Perhaps the ability of cvzM to inhibit proteolytic activity accounts for its survival in normal cells.
One class of cells in which (YAM fluorescence was markedly diminished were cells transformed by the Moloney strain of murine sarcoma virus (MoSV). Since this defect was found in cells re- cently transformed by MoSV and in virtually all cells in a MoSV-transformed culture, we believe that the loss of a2M fluorescence is an early conse- quence of transformation by MoSV and not sec- ondary to other factors that affect the behavior of cultured cells. Decreased a2M fluorescence was also observed in one of two clones of cells trans- formed by another murine sarcoma virus, KiSV. Cells transformed by RSV-SR and a variety of other transformed cells, however, had normal amounts of aPM. A more extensive examination of the amount of CY*M in transformed cells is in progress.
At the current time, the reason for the diminished content of a2M in the lysosomes of MoSV trans- formed cells is unclear. By electron microscopy and phase-contrast microscopy, some of these MoSV-transformed cells appear to have normal numbers of lysosomes. Furthermore, we have mea- sured the levels of two lysosomal enzymes, p- galactosidase and P-glucosaminidase, in normal and MoSV-transformed cells, and found no striking differences. Transformed cells often contain and

WI 616
release increased amount of proteases (Reich, Rif- kin and Shaw, 1975). The possibility that MoSV- transformed cells make excessive amounts of a protease that destroys apM warrents investigation. Since proteases appear to be able to stimulate cell growth, and a2M is a protease inhibitor, it is possi- ble that CY~M derived from the culture medium has a role in maintaining normal growth control.
Experimental Procedures
Materlals Purified human ar macroglobulin was a gift from Dr. Y. L. Hao of the American Red Cross. The L~*M was further purified on a Sepharose 6B column (0.9 X 50 cm) in IO mM Tris-Cl (pH 7.6). Purified bacterial collagenase was a gift from Dr. B. Peterkofsky. “C-amino acid mixture (100 Q/ml, 50 mCi/m atom) was pur- chased from New England Nuclear. All chemicals used for SDS- polyacfylamide gels were of electrophoresis purity and were purchased from Bio-Rad. Tetramethyl rhodamine isothiocyanate was a gift from Dr. Yossi Schlessinger (Cornell University). All sera used in the immunoprecipitation procedures were purchased from Miles Research Products.
Growth of Cells NRK cells and NRK cells transformed with the Kirsten strain (KNRK) and Maloney strain (MoSV NRK) of murine sarcoma virus were obtained from Dr. E. Scolnick (National Institutes of Health). Mouse embryo fibroblasts (MEF) were prepared from 14-16 day embryos by the method of Vogt (1969) in the same manner that chick embryo fibroblasts (CEF) are prepared. Moloney MSV-trans- formed MEF cells (MOW MEF) were prepared by treating primary MEF cultures with the medium from an NRK cultured cell line that is a producer of MoSV/MoLV (obtained from Dr. E. Scolnick). Swiss 3T3-4 fibroblasts were obtained from Dr. H. Green (Massa- chusetts Institute of Technology). Balb SV-TP (SVl), KA,,CI 1 and MSV65 Cl 1 were obtained from Dr. E. Scolnick. Balb MC55 and Moloney Balb A31 were obtained from Dr. George Todaro (Na- tional Institutes of Health). L929 cells were obtained from Dr. S. Baron (National Institutes of Health).
All cells (except CEF) were incubated at 37°C in a 95% air/596 CO, humidified atmosphere in Dulbecco-Vogt’s modified Eagle’s medium supplemented with 10% heat-inactivated calf serum (Col- orado Serum) and penicillin-streptomycin (50 units per ml each; Flow Labs). Cells were grown under these conditions on either 9 cm2 or 20 cm* Falcon tissue culture grade plastic dishes. The preparation and culture of CEF has previously been described (Vogt. 1969).
‘X-Amlno ACM Labeling Cells were grown to confluency in 20 cm* dishes. The medium was removed and the plates were rinsed 3 times with amino acid-free, serum-free medium. To each plate were added 3 ml amino acid-free, serum-free medium and IO @Zi ‘T-amino acid mixture. The plates were incubated at 37°C for 2 hr. At the end of the labeling period, the medium was removed, the plates were rinsed 5 times with serum-free medium containing unlabeled amino acids, and 3 ml of the serum-free medium with unlabefed amino acids were added. The cultures were incubated at 37°C. After 4 hr the medium was removed, centrifuged at 100,000 X g for 30 min to remove formed elements and either run directly on SDS gels or handled as described for collagenase treatment or immunoprecipitation.
Collagenase Treatment 1 ml medium from cell samples was dialyzed against 500 ml 10 mM Tris-Cl (pH 7.6) at 4°C overnight, then lyophilized to dryness. The lyophilized sample was taken up in 0.2 ml of 0.2 M
NaCI, 0.05 M Tris-Cl (pH 7.6). 1 ~1 collagenase was added to 50 ~1 of the resolubilized sample [565 pg/ml in 0.05 M Tris-Cl (pH 7.6), 5 mM CaCld or 1 pl buffer (control), 1 ~1 of 25 mM CaCI,. 2 ~1 of 62.5 mM N-ethylmaleimide. The samples were incubated at 37°C for 10 min. The reaction was stopped by the addition of SDS preparatory to running SDS-polyacrylamide gels.
Immunoprecipitatkn Media samples were dialyzed overnight against 400-500 vol of 10 mM Tris-Cl (pH 7.6) at 4°C. All sera to be used were heated at 60°C for 30 min and centrifuged to remove any precipitate, and the supernatant was precipitated with ammonium sulfate to 4@/. final concentration. The ammonium sulfate pallet was resus- pended to original volume in 10 Mm Tris-Cl (pH 7.6) dialyzed against the same buffer. 20 ~1 of rabbit anti-human crtM antibody (2Ct gl of normal rabbit serum for control) were added to 1 ml of dialyzed medium, mixed well and allowed to stand 1 hr at room temperature. 100 @I of goat anti-rabbit IgG serum were added, mixed well and allowed to stand 30 min at room temperature. The resulting suspension was centrifuged 12.006 rpm (17,000 X g) in an SS-34 rotor in a Sorvall centrifuge at 4°C. The pellets were washed 2 times with phosphate-buffered saline. The final pellet was taken up in 100 ~1 of 2% SDS preparatory to SDS- polyacrylamide gel electrophoresis. 25 ~1 of the SDS-pellet sus- pension were run per gel lane.
Rhodamlne Laballng 0.15 ml of 0.5 M Na,CO, (pH 9.5) were added to 1.35 ml of PBS containing 5.6 mg arM. This was transferred to a vial containing 160 pg of dry tetramethyl rhodamine isothiocyanate. The mixture was stirred with a magnetic flea in the dark at 49: for 16 hr and then dialyzed against 1006 ml of PBS overnight to remove most of the free rhodamine. The labeled protein was then applied to a 0.9 X 10 cm G-50 Sephadex column equilibrated with PBS, and 1 ml fractions were collected. The first peak off the column was rhodamine-crrM. The arM fractions were stored frozen in the dark.
Preparatton of a# Released from Cells NRK cells were grown to confluency in glass roller bottles. The medium was removed, and the cells were rinsed with PBS (37°C) followed by two rinses with serum-free medium. The cells were then incubated overnight at 37°C in serum-free medium. The medium was removed, centrifuged to remove cellular debris and lyophilized to dryness. The lyophilized preparation was resus- pended in 20 ml of Buffer A [So mM Tris-Cl (pH 6) 0.1 M NaCI], dialyzed against 4 liters of the same buffer with two changes over 30 hr and centrifuged to remove a precipitate which formed; the supernatant was concentrated to 3 ml with an Amicon PM 30 ultrafilter. The concentrated material was chromatographed on a G-200 Sephadex column (1.5 X 95 cm) equilibrated with Buffer A. The material from the first protein peak (>150,006 daltons) off the column was pooled and concentrated to 2 ml (1.4 mg protein per ml).
Immunofluore8cence Labeling C&s were grown in 9 cm2 tissue culture dishes. After propagation for 46 hr, each dish was washed with phosphate-buffered saline (PBS) and fixed for 5 min in 1% formaldehyde in PBS. followed by 60% acetone in H&I for 5 min at 23°C. After rinsing with PBS, 1 ml of a l/50 dilution of serum was added (either normal rabbit serum or rabbit anti-a,M) and incubated for 10 min. The dish was then rinsed again with PBS, and 1 ml of a I/20 dilution of goat anti-rabbit gammaglobulin coupled to rhodamine isothiocyanate (Cappel Labs) was added and incubated for IO min. A final wash in PBS was followed by mounting in glycerol buffer medium under a circular coverslip. The dishes were directly viewed using epifluorescent or phase-contrast illumination with a Zeiss RA microscope and a 40X oil immersion objective. Photographs were taken using Polaroid 107 film (3000 ASA). lmmunoabsorption was accomplished by incubating 1 ml of l/50 ant&M with 100

Fibroblasts Contain ar Macroglobin from Serum 617
kg of purified human (I*M for 5 min at 23°C. Rhodamine-labeled QM or bovine serum albumin (BSA) (50 pg/ml) was incubated with live cells for 15-45 min in complete medium containing 0.1% serum.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 16 U.S.C. Section 1734 solely to indicate this fact.
Received July 6, 1977; revised August 23, 1977
References
Adams, S. L., Sobel, M. E.. Howard, 8. H., Olden, K., Yamada, K. M., de Crombrugghe, B. and Pastan, I. (1977). Proc. Nat. Acad. Sci. USA, in press.
Frenoy. J. P., Bourrillon, R.. Lippoldt, R. and Edelhoch. l-f. (1977). J. Biol. Chem., in press.
Harpel, P. C. (1973). J. Exp. Med. 738, 508-521.
Mosher, D. F. and Wing, D.A. (1976). J. Exp. Med. 743,462-467.
Nicolson, G. L. (1976). Biochim. Biophys. Acta 458, i-72.
Ostlund, R. E. and Pastan, I. (1976). Biochim. Biophys. Acta 453, 37-47.
Reich, E., Rifkin, D. B. and Shaw, E. (1975). Proteases and Biological Control (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory).
Shortridge, K. F., Belyavin, G. and Bidwell, D. E. (1976). Comp. Biochem. Physiol. 54, 319-321.
Steinman, R. M.. Silver, J. M. and Cohn, 2. A. (1974). J. Cell Biol. 83, 949-969.
Vogt. P. K. (1969). In Fundamental Techniques in Virology. K. Habel and N. Salzman. eds. (New York: Academic Press), pp. 196-211.
Werb, Z., Burleigh, M. C., Barrett, A. J. and Starkey, P. M. (1974). Biochem. J. 139, 359-368.
Yamada, K. M. and Weston, J. A. (1974) Proc. Nat. Acad. Sci. USA 77, 3492-3496.
Yamada, K. and Pastan, I. (1976). Trends Biochem. Sci. 1, 222- 224.
Note Added in Proof
D. F. Mosher (personal communication) has also recently found that cultured fibroblasts take up significant amounts of armacro- globulin from the culture medium.


















