IL-2 and IL-12 Alter NK Cell Responsiveness to IFN-γ ... · PDF fileIL-2 and IL-12 Alter...
10
Click here to load reader
Transcript of IL-2 and IL-12 Alter NK Cell Responsiveness to IFN-γ ... · PDF fileIL-2 and IL-12 Alter...

of May 16, 2018.This information is current as
Down-Regulating CXCR3 Expression-Inducible Protein 10 byγto IFN-
IL-2 and IL-12 Alter NK Cell Responsiveness
YoungBlanca, Della A. Reynolds, John R. Ortaldo and Howard A. Deborah L. Hodge, William B. Schill, Ji Ming Wang, Isaac
http://www.jimmunol.org/content/168/12/6090doi: 10.4049/jimmunol.168.12.6090
2002; 168:6090-6098; ;J Immunol
Referenceshttp://www.jimmunol.org/content/168/12/6090.full#ref-list-1
, 24 of which you can access for free at: cites 37 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

IL-2 and IL-12 Alter NK Cell Responsiveness to IFN-�-InducibleProtein 10 by Down-Regulating CXCR3 Expression1
Deborah L. Hodge,* William B. Schill,‡ Ji Ming Wang,† Isaac Blanca,* Della A. Reynolds,*John R. Ortaldo,* and Howard A. Young2*
Cytokine treatment of NK cells results in alterations in multiple cellular responses that include cytotoxicity, cytokine production,proliferation, and chemotaxis. To understand the molecular mechanisms underlying these responses, microarray analysis wasperformed and the resulting gene expression patterns were compared between unstimulated, IL-2, IL-2 plus IL-12, and IL-2 plusIL-18-stimulated NK92 cells. RNase protection assays and RT-PCR confirmed microarray predictions for changes in mRNAexpression for nine genes involved in cell cycle progression, signal transduction, transcriptional activation, and chemotaxis.Multiprobe RNase protection assay also detected changes in the expression of CCR2 mRNA, a gene that was not imprinted on themicroarray. We subsequently expanded our search for other chemokine receptor genes absent from the microarray and found anIL-2- and IL-12-dependent decrease in CXCR3 receptor mRNA expression in NK92 cells. A detailed analysis of CXCR3 expres-sion in primary NK cells revealed that an IL-2 and an IL-12 together significantly decreased the CXCR3 receptor mRNA andreceptor surface expression by 6 and 24 h of treatment, respectively. This decrease in receptor expression was associated with asignificant reduction in chemotaxis in the presence of IFN-�-inducible protein-10. The decline in CXCR3 mRNA was due totranscriptional and posttranscriptional mechanisms as the addition of actinomycin D to IL-2- and IL-12-treated NK92 slightlyaltered the half-life of the CXCR3 mRNA. Collectively, these data suggest that IL-2 and IL-12 directly affect NK cell migratoryability by rapid and direct down-regulation of chemokine receptor mRNA expression. The Journal of Immunology, 2002, 168:6090–6098.
N atural killer (NK) cells are large granular lymphocytesthat play an important role in the defense against virallyinfected or malignant cells (1). Their activity can be
characterized as nonadaptive and independent of MHC restriction(1, 2). A variety of NK cell functions including cytotoxicity, pro-liferation, chemotaxis, and cytokine production are modulated byregulatory cytokines including IFN-��, IL-2, IL-12, IL-18, IL-10,and TNF (reviewed in Refs. 3 and 4). Because cytokines inducesuch a broad range of effects in NK cells, the potential for alter-ations in gene expression in stimulated cells is very great. To de-termine which genes are regulated in response to cytokine stimu-lation, our laboratory has used cDNA microarray technology toexamine gene expression in NK cells. Microarray technology isvery useful because it allows for large-scale examination of geneexpression. Additionally, this technology has proved useful inidentifying physiologically relevant gene expression patterns ineukaryotic systems such as yeast (5) and fibroblasts (6) as well as
predicting patterns of gene expression in tumor cells (7, 8). Toexamine gene expression in response to cytokine stimulation, ahuman NK cell line, NK92, was stimulated with IL-2 alone or incombination with IL-12 or IL-18. These cytokines were chosenbecause of their ability to induce NK cell responses; however, littleis known about the repertoire of genes that are activated by thesecytokines. Microarray analysis of gene expression in NK92 cellsidentified a variety of genes whose mRNA expression patternschange in response to cytokine stimulation. The genes encodingthe mRNAs are not specific to any one pathway; however, changesin cytokine, chemokine, and chemokine receptor gene mRNAswere prevalent. Our mRNA studies on chemokine receptor geneexpression were extended to cell surface analysis of receptor den-sities in cytokine-treated primary NK cells. Using FACS analysis,we observed a significant decrease in CXCR3 receptor expressionin NK cells treated for 24 h with IL-2 and IL-12 alone or in com-bination. Recently, alterations in chemokine receptor expressionwere reported in IL-2-stimulated NK cells (9); however, the cellswere cultured in IL-2 for 8–10 days. In contrast, our data demon-strate that cytokines can modify chemokine receptor functionwithin hours, thus supporting a model whereby cytokines, in par-ticular IL-2 and IL-12, regulate chemokine receptor expression ina direct, rapid, and novel manner.
Materials and MethodsNK cell preparation
PBMC were isolated from buffy coats of healthy donors (obtained from theNational Institutes of Health Blood Bank, Bethesda, MD) after centrifu-gation on a lymphocyte separation medium. Cells were washed twice withDulbecco’s PBS and suspended in RPMI 1640 medium supplemented with2 mM L-glutamine, 100 IU/ml penicillin, 50 �g/ml streptomycin, and 10%FCS. Adherent cells were removed by incubation in plastic flasks for 1 hat 37°C. Nonadherent cells were recovered by gently washing with warmedmedium and were further purified by incubating on nylon wool columns for
Laboratories of *Experimental Immunology and †Molecular Immunoregulation, Cen-ter for Cancer Research, National Cancer Institute, Frederick, MD 21702; and ‡Na-tional Fish Health Research Laboratory, U.S. Geological Survey-Leetown ScienceCenter, Kearneysville, WV 25430
Received for publication September 14, 2001. Accepted for publication April 5, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This project has been funded in whole or in part with Federal funds from the Na-tional Cancer Institute, National Institutes of Health under Contract No. N01-CO-12400. The publisher or recipient acknowledges the right of the U. S. Government toretain a nonexclusive, royalty-free license in and to copyright covering the article. Thecontent of this publication does not necessarily reflect the views or policies of theDepartment of Health and Human Services, nor does mention of trade names, com-mercial products, or organizations imply endorsement by the U. S. Government.2 Address correspondence and reprint requests to Dr. Howard A. Young, NationalCancer Institute, Building 560, Room 31-23, Frederick, MD 21702-1201. E-mailaddress: [email protected]
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

1 h at 37°C. The nylon-nonadherent cells (mostly T cells and NK cells)were eluted with prewarmed RPMI-640 medium and fractionated on aseven-step Percoll gradient as previously described (10). The NK cell-enriched low-density fraction-2 (40–60% NK cells) was further depletedof remaining T lymphocytes and monocytes by negative selection withanti-CD3 and anti-CD14 mAbs. Briefly, the cells were labeled for 30 minon ice with biotinylated anti-CD3 and anti-CD14 Abs. After removing theunbound Abs by washing with cold PBS plus 1% BSA, the cells wereincubated 15 min with streptavidin microbeads (Miltenyi Biotec, Oslo,Norway) and the positive cells (CD3� and CD14�) were removed with amagnetic column (MACS; Miltenyi Biotec). Purified NK cell populationswere �95% CD56�/CD5� cells as determined by two-color flow cytom-etry analysis (FACSort; BD Biosciences, San Jose, CA) with anti-CD56 PEand anti-CD5 FITC (BD Biosciences).
Cell culture
NK92 cells were maintained in RPMI 1640 medium (BioWhittaker, Walk-ersville, MD), supplemented with 10% FCS, 2 mM L-glutamine, 100 U/mlpenicillin, 100 �g/ml streptomycin, 200 U/ml recombinant human IL-2,and 10 ng/ml recombinant human IL-15. Cells were cultured at a densityof 5 � 105/ml in a 37°C incubator with 5% CO2. For all experiments, cellswere grown at a density of 1 � 106/ml in medium lacking IL-2 and IL-15for 12 h before cytokine stimulation.
Freshly isolated primary NK cells were placed into warm RPMI 1640medium, supplemented with 10% FCS, 2 mM L-glutamine, 100 U/ml pen-icillin, and 100 �g/ml streptomycin and immediately treated with 100 U/mlIL-2, 10 U/ml IL-12, or 10 ng/ml IL-18, alone or in combination.
Poly(A)� RNA preparation, microarrays, and data analysis
Approximately 100 � 106 NK92 cells were stimulated for 3 h with 100U/ml IL-2 alone or in combination with 10 U/ml IL-12 or 10 ng/ml IL-18.Poly(A)� RNA was isolated according to the manufacturer’s protocol us-ing a FastTrack 2.0 mRNA isolation kit (Invitrogen, Carlsbad, CA). IncyteGenomics (St. Louis, MO) performed the cDNA generation, hybridization,and data collection. Alterations in gene expression were evaluated by re-verse transcription of poly(A)� RNAs in the presence of Cy3 or Cy5 flu-orescent labeling dyes followed by hybridization to UniGEM human Vmicroarray chips (Incyte Genomics). Each chip contains a total of 7075elements of which 6794 are unique genes/clusters. These unique genes/clusters can be further defined as 4610 annotated and 2184 unannotatedsequences. Subsets of genes were selected based on differential Cy3/Cy5expression ratios that were ��2� in response to any treatment. Differentialexpression of representative selected genes was confirmed by RT-PCRand/or RNase protection assay (RPA).3
RNA isolation, relative quantitative RT-PCR, RPA
To confirm microarray predictions, �10 � 106 NK92 cells were restedovernight without IL-2 and IL-15. The cells were either nonstimulated orstimulated with IL-2 (100 U/ml), IL-12 (10 U/ml), and IL-18 (10 ng/ml)alone or in combination for 3 h. Total RNA was isolated from the untreatedand cytokine-treated cells using the TRIzol (Life Technologies, Bethesda,MD) extraction procedure. For relative quantitative RT-PCR, �5 �g ofRNA was reverse transcribed using a Thermoscript RT-PCR system (LifeTechnologies). The cDNA template was generated according to the man-ufacturer’s protocol with random hexamers as primers for initiation ofreverse transcription. Multiplex PCR was performed using Platinum PCRSupermix (Life Technologies) and primers specific for the human 18SrRNA gene and also the gene of interest. The amplification efficiency of the18S rRNA gene was modulated using an 18S Competitimer technology(Ambion, Austin, TX). This allowed the 18S gene to be expressed in thesame linear range as the gene of interest when amplified. The amount ofPCR product for the gene of interest could then be compared with theamount of 18S product to estimate the amount of variation betweensamples.
The multiprobe RPAs were performed according to the manufacturer’sdirections (BD PharMingen, San Diego, CA) with the following modifi-cations: RNase inactivation and precipitation was performed using a mastermixture containing 200 �l Ambion RNase inactivation reagent, 50 �l eth-anol, 5 �g yeast tRNA, and 1 �l Ambion GycoBlue coprecipitate per RNAsample. After adding the individual RNase-treated samples to 250 �l of theinactivation/precipitation mixture, the samples were mixed well, placed at
�70°C for 30 min, and subjected to centrifugation at 14,000 rpm for 15min in a room temperature microcentrifuge. The supernatants were de-canted, a sterile cotton swap was used to remove excess liquid, and thepellet was resuspended in 3 �l of BD PharMingen sample buffer. Eachmultiprobe RNase assay included probes specific for ribosomal L32 andGAPDH mRNAs to assure equal amounts of input mRNA in each assayand to control for lane to lane variation during PAGE.
For guanylate binding protein 1 (GBP-1)-1 and Src homology 2 domain-containing leukocyte protein of 76 kD (SLP-76) mRNA analysis, RPAprobes were synthesized from plasmid DNA templates using T7 RNApolymerase and [�-33P]UTP in an in vitro transcription reaction. The newlysynthesized riboprobes were loaded onto a 6% denaturing polyacrylamidegel and full-length probes were excised and eluted from the gel by over-night incubation at 37°C in gel elution buffer (supplied in RPA II kit;Ambion). For each RPA, �100 � 104 cpm and 50 � 104 cpm of thegene-specific and 18S rRNA probes were used, respectively. RPAs forGBP-1 and SLP-76 were performed according to the manufacturer’s pro-tocol using a RPA II RNase protection assay kit (Ambion) with 10 �g ofinput RNA per reaction. For all multiprobe and single-probe RPAs, pro-tected RNA products were separated by size on a 6% denaturing poly-acrylamide gel. Gels were dried under vacuum at 80°C for �2 h andexposed to either x-ray film or a PhosphorImager screen for 16 to 24 h.Film images were scanned and signal intensities were quantitated usingTotalLab image analysis software (Phoretix, Newcastle Upon Tyne, U.K.).Phosphorimages were generated and quantitated using PhosphorImager SIanalysis and ImageQuant software (Molecular Dynamics, Sunnyvale, CA).
mRNA half-life measurement
Rested NK92 cells were treated with IL-2 (100 U/ml) and IL-12 (10 U/ml)alone or in combination in the presence of actinomycin D (5 �g/ml). TotalRNA was isolated at multiple times and CXCR3 mRNA expression wasmeasured by multiprobe RPA using [�-33P]UTP-labeled riboprobes tran-scribed from a hCR-6 template (BD PharMingen). The protected mRNAfragments were size separated on a 6% polyacrylamide gel under denatur-ing conditions. The RPA gels were dried and exposed to a PhosphorImagerscreen for 12–24 h. Images were visualized and quantitated using Phos-phorImager SI analysis and ImageQuant software (Molecular Dynamics).
Flow cytometry analysis
To examine CXCR expression, primary NK cells were stained with FITC-conjugated CXCR3 or CXCR4 mAbs (R&D Systems, Minneapolis, MN)and PE-conjugated CD56 mAb (BD Biosciences). For CCR expression,NK cells were stained with PE-labeled CCR1 or CCR2 mAbs (R&D Sys-tems) and FITC-conjugated CD16 mAb (BD Biosciences). Cells were alsostained using isotype controls FITC-conjugated IgG1 and PE-conjugatedIgG2A Abs (BD Biosciences). All cells were analyzed using a FACSortflow cytometer (BD Biosciences).
Chemotaxis assay
NK cell migration was assessed with 48-well microchemotaxis chambers(NeuroProbe, Cabin John, MD). The chemoattractants were placed in thewells of the lower compartment. The cells (at 3 � 106/ml) were placed inthe wells of the upper compartment. A 5-�m pore-sized polycarbonatefilter precoated with fibronectin separated the two compartments. Afterincubation at 37°C for 3 h, the filter was removed, stained with DiffQuik,and the cells migrated across the filter were counted under microscopy withthe samples coded. The cell migration was tested in triplicates and thechemotaxis index represents the fold-increase of cell migration in responseto chemokines over medium control. The statistical significance was ex-amined with the Student t test.
ResultsCytokine-induced alterations in gene expression
Microarray experiments were performed to examine changes ingene expression patterns in NK cells in response to cytokine ac-tivation. In one experiment, changes in mRNA expression in rest-ing cells were compared with those observed in IL-2-activated NKcells. In a second experiment, gene expression patterns were com-pared in NK cells under two different stimulatory conditions. Forthis, mRNA from IL-2 plus IL-12-stimulated cells was comparedwith mRNA from IL-2 plus IL-18-stimulated cells. In all experi-ments, cells were stimulated for 3 h and poly(A)� mRNA wasisolated and evaluated by microarray analysis.
3 Abbreviations used in this paper: RPA, RNase protection assay; GBP-1, guanylatebinding protein 1; SLP-76, Src homology 2 domain-containing leukocyte protein of76 kD; MIP, macrophage-inflammatory protein; MX1, myxovirus resistance 1; PI3K,phosphoinositide-3-kinase; IP-10, IFN-�-inducible protein-10.
6091The Journal of Immunology
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Stimulation of NK cells by IL-2 resulted in identification of atotal of 65 genes with greater than 2-fold changes in expression(Table I). Of these, the majority of the genes (n � 49) were up-regulated by IL-2. The identified genes were involved in a varietyof cellular processes that included transcriptional activation, inter-mediary metabolism, signal transduction, and cell cycle regulation.Microarray analysis of mRNA from IL-2 plus IL-12 and IL-2 plusIL-18-stimulated NK92 cells did not reveal a large of amount ofdifferential regulation. Of the 17 genes identified, most (n � 10)were up-regulated by IL-2 plus IL-12 as compared with IL-2 plusIL-18 (Table II). We were surprised at the overall lack of differ-ential gene expression between cells treated with IL-12 and IL-18.Many cytokines bind receptors that have common subunit chainsthat are shared with other receptors. The commonality betweenreceptors may attribute to the activation of similar signaling path-ways by different cytokines. IL-12 and IL-18 receptors, however,are structurally very different and do not share common receptorchains. Thus, their cellular target genes would potentially be verydifferent; however, the converse is true. These data suggest thatIL-12 and IL-18 signaling pathways converge at the nuclear levelto activate similar subsets of genes.
Predicted activation of genes confirmed by relative quantitativeRT-PCR and RPA
Genes were selected from the microarray results for further anal-ysis based on their predicted expression change or their potential toaffect NK cell biology. These genes were directly tested forchanges in mRNA expression by RPA or relative quantitative RT-PCR. Multiprobe and single-probe RPA were performed on RNAisolated from unstimulated NK92 cells as well as cells treated withIL-2 alone or in combination with IL-12 or IL-18. Predictedchanges in gene expression in three primary families of genes wereconfirmed by multiprobe RPA. These families represent genes thatcode for cytokine, chemokine, and chemokine receptors.
To ensure that our cytokine treatments properly induced knowncellular functions, an IFN-�-specific probe was used as a positivecontrol in RPA and RT-PCR analyses. Changes in IFN-� mRNAexpression are directly linked to cytokine stimulation (11–13) andthus, are useful measurements of cytokine-induced changes ingene expression. Consistent with previous reports (11, 14, 15), theRPA result demonstrated cytokine-induced increases in IFN-�mRNA by IL-2 alone (Fig. 1). Moreover, the expected synergybetween IL-2 and IL-12 or IL-18 on IFN-� mRNA expression wasobserved. These data demonstrated that NK92 cells responded tocytokine treatment in an expected manner thus providing assur-ance that the microarray results accurately predicted changes inmRNA expression of other genes.
For examining changes in IFN-� mRNA, we used a multiprobetemplate (hCK1) which also contains an IL-10-specific probe. Thiswas useful because our microarray results predicted a greater than2-fold increase in IL-10 mRNA in response to IL-2. Consistentwith previous reports (16, 17, 18), RPA analysis demonstrated thatIL-10 gene expression was altered by cytokine stimulation of NKcells (Fig. 1). Interestingly, the patterns of IL-10 and IFN-� mRNAexpression were parallel with respect to IL-2 alone and in combi-nation with IL-12 or IL-18. We do not know why these two an-tagonistic cytokines are coexpressed in a similar fashion in NKcells; however, it is possible that the IL-10 may act as a negativeregulator to shutdown IFN-� expression. This is supported by areport that demonstrates that IL-10 secretion from Th2 cells acts toinhibit IFN-� production in both Th1 cells and NK cells (19).
We next used RPA analysis to examine chemokine receptormRNA expression in NK92 cells. Using a CCR-specific multi-probe RPA, we confirmed the microarray prediction for up-regu-
lation of the chemokine receptor CCR1 by IL-2 (Fig. 1). Addi-tionally, RPA analysis demonstrated a slight up-regulation ofCCR2 mRNA expression by IL-2 plus IL-12. This gene was notimprinted on the microarray and was examined only as a result ofour usage of the multiprobe RPA. As a result of this observation,we extended our study to other chemokine receptor genes thatwere absent on the microarray. Using a CXCR-specific RPA tem-plate, we found that CXCR3 mRNA expression was down-regu-lated 13% by IL-2. The combination of IL-2 with IL-12 or IL-18further enhanced this effect with decreases of 38 and 56%, respec-tively (Fig. 1). In contrast, CXCR4 mRNA expression was up-regulated �1.2-fold by all cytokine treatments (Fig. 1), suggestingthat effects on this gene were IL-2-specific.
Although our microarray data did not contain information per-taining to chemokines, changes in cytokine and chemokine recep-tor mRNA expression lead us to examine changes in these impor-tant chemotactic-inducing proteins. Consistent with a previousreport (20), multiprobe RPA analysis demonstrated an IL-2 induc-tion of macrophage-inflammatory protein (MIP)-1� mRNA. Theinability of IL-12 to further enhance the MIP-1� mRNA expres-sion indicated that IL-2 was the primary inducer of this mRNA.This was further supported by only a slight enhancement inMIP-1� mRNA expression in IL-2 plus IL-18-treated NK92 cells.In contrast to MIP-1�, MIP-1� mRNA expression was not affectedby IL-2. However, IL-18 in combination with IL-2 did up-regulateexpression of this chemokine.
We next examined mRNA expression of GBP-1 and myxovirusresistance 1 (MX1) genes. Both the GBP-1 and MX1 genes code forIFN-inducible GTP-binding proteins that protect cells against viralinfection (21–23). Because NK cells are primary defenders againstviral invaders, we speculated that these proteins might provideantiviral protection to NK cells during virus encounters. Our mi-croarray results suggested that IL-2 should specifically up-regulatemRNA expression of both the GBP-1 and MX1 genes. RPA probeswere designed and used for RPA analysis of these mRNAs. Con-sistent with the microarray results, we found that IL-2 up-regulatedexpression of the GBP-1 mRNA in NK92 cells (Fig. 2). This ex-pression was specific to IL-2 in that costimulation with IL-12 orIL-18 had little or no effect on GBP-1 mRNA expression. UnlikeGBP-1, MX1 mRNA expression did not correlate with the mi-croarray results. RPA analysis revealed that NK92 cells did ex-press MX1 mRNA; however, the expression was constitutive andcytokine-independent (Fig. 2). The lack of inducibility of the MX1gene demonstrates that microarray results, like those from any sin-gle method, can be incorrect, thus necessitating independent con-formation of gene expression changes by alternative methods.
The microarray results also predicted that cytokine treatments ofNK cells could alter mRNA expression of genes that code forsignal transduction proteins. Two such genes were the p85 subunitof phoshoinositide-3-kinase (PI3K) and the lymphocyte cytosolicprotein 2/SLP-76. RPA analysis of PI3K mRNA expression dem-onstrated an IL-2-dependent decrease in PI3K mRNA (Fig. 3). TheIL-2 effect was enhanced by costimulation with IL-12 and IL-18,thus implicating these cytokines as coregulators of PI3K mRNAexpression. In contrast, SLP-76 mRNA expression was signifi-cantly increased by IL-2 (Fig. 2). IL-2 appeared to be the primaryregulator of SLP-76 mRNA in that no increase in mRNA expres-sion was observed in the IL-2 plus IL-12-treated NK cells. Addi-tionally, costimulation by IL-2 and IL-18 only slightly up-regu-lated the SLP-76 mRNA expression over that observed with IL-2alone.
Relative-quantitative RT-PCR corroborated the predictedchanges in the expression of five genes in response to IL-2 (Fig. 3).Of these, pim-1, cyclin D2, and c-myc genes code for proteins that
6092 CYTOKINE-INDUCED CHANGES IN NK CELL GENE EXPRESSION
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Table I. Analysis of gene expression in nonstimulated and IL-2–stimulated NK92 cells
Gene Name GenBank Accession Number Nonstimulated Signal IL-2–Stimulated Signal Fold Change
Genes down–regulated by IL-2EST AI380052 17,963 6,861 �2.62AHNAK nucleoprotein (desmoyokin) M80899 5,022 2,085 �2.41Homo sapiens nuclear Ag H731-like protein U96628 4,810 2,287 �2.1IL-9R M84747 3,902 1,415 �2.76V-yes-1 Yamaguchi sarcoma viral–related oncogene
homologM16038 2,562 1,233 �2.08
IL-16 (lymphocyte chemoattractant factor) AI652705 2,540 1,235 �2.06DB1 AA195154 2,492 1,050 �2.37Conserved gene amplified in osteosarcoma AF022231 2,162 1,043 �2.075� fragment, not mapped (Incyte PD: 194162) N/Aa 1,668 713 �2.34PI3K, regulatory subunit, polypeptide 1 (p85 �) M61906 1,275 398 �3.2Fucosidase, �-L-1 M80815 1,122 318 �3.53Human EV12 protein gene M55267 800 394 �2.03EST, highly similar to HMG box containing protein 1 H86395 729 364 �2B cell translocation gene 1, anti-proliferative AI925293 654 234 �2.79EST AI436055 370 159 �2.33Sortilin-related receptor, L(DLR class) Y08110 265 126 �2.1Genes up-regulated by IL-2Chemokine (C-C motif) receptor 1 D10925 4,456 22,840 5.13EST, moderately similar to heat shock protein HSP 90-� AL040289 7,070 15,715 2.22Human pim-2 protooncogene homolog pim-2h AI634033 4,062 13,916 3.43V-myc avian myelocytomatosis viral oncogene homolog V00568 4,229 13,642 3.23Human serine protease gene J02907 5,694 12,758 2.24Lactate dehydrogenase B AL044172 4,993 12,238 2.45Solute carrier family 7 (cationic amino acid transporter,
y� system), member 7AJ130718 2,242 11,686 5.21
Differentiated embryo chondrocyte expressed gene 1 AB004066 3,309 9,317 2.82Lymphocyte cytosolic protein 2 U20158 4,184 9,299 2.22IL-10 M57627 3,373 7,861 2.33Pim-1 oncogene M54915 1,533 7,829 5.11GBP 1, interferon-inducible M55542 3,229 7,274 2.25Cyclin D2 D13639 1,918 6,180 3.22CSF 1 (macrophage) M11296 2,691 5,753 2.14Perforin 1 (preforming protein) AI076019 2,618 5,736 2.19Tryptophanyl-tRNA synthetase X59892 2,247 5,730 2.55Ornithine decarboxylase 1 M16650 1,806 5,380 2.98Putative translation initiation factor (Incyte PD: 58399) N/A 2,478 5,344 2.16MX1 (influenza), homolog of murine (IFN-inducible
protein p78)M30817 1,645 5,275 3.21
IFN regulatory factor 1 X14454 2,385 5,254 2.2FLN29 gene product AB007447 1,854 5,040 2.72Dyskeratosis congenita 1, dyskerin U59151 2,068 4,873 2.36Solute carrier family 1 (neutral amino acid transporter),
member 5U53347 2,031 4,818 2.37
5� not mapped fragment (Incyte PD: 2498667) N/A 1,456 4,625 3.18EST AI127628 1,839 4,381 2.38Dual specificity phosphatase 5 U15932 1,735 4,286 2.47Paired basic amino acid cleaving enzyme (furin,
membrane associated receptor protein)X17094 1,450 3,341 2.3
EST AI188513 1,165 3,335 2.86EST AA559093 695 3,260 4.69Nucleolar phosphoprotein p130 Z34289 1,116 2,929 2.62Human ras-like protein AA459547 1,109 2,914 2.63Pleckstrin homology, Sec7 and coiled/coil domains,
binding proteinAI538459 745 2,546 3.42
Topoisomerase (DNA) I U07806 911 2,138 2.35Flotillin 1 AA618098 850 1,915 2.25Protein tyrosine phosphatase type IVA, member 1 U48296 912 1,904 2.09Carcinoembryonic Ag gene family member 6 X52378 654 1,847 2.82B cell CLL/lymphoma 2 M14745 789 1,691 2.14Serine/threonine kinase 17b (apoptosis-inducing) AA725600 783 1,580 2.02Bystin-like L36720 665 1,437 2.16H. sapiens mRNA expressed in osteoblast F12860 459 1,393 3.03Pregnancy specific �-1-glycoprotein 7 (Incyte PD:
64457)N/A 551 1,389 2.52
Methionine adenosyltransferase II, � F07456 592 1,342 2.27Glutamyl aminopeptidase (aminopeptidase A) L12468 535 1,231 2.3Cytochrome P450, 51 (lanosterol 14-�-demethylase) U23942 601 1,227 2.04Low density lipoprotein receptor (familial
hypercholesterolemia)L00352 559 1,121 2.01
Fatty-acid-coenzyme A ligase, very long-chain 1 D88308 460 1,012 2.2H. sapiens DKFZp564L176 (from clone
DKFZp564L176)D64110 353 862 2.44
Regulator of G-protein signaling 16 U70426 327 778 2.38IL 6 signal transducer (gp 130, oncostatin M receptor) M57230 348 709 2.04
a N/A represents genes that do not have GenBank accession numbers. The Incyte PD identification numbers are provided. Expression of genes listed in italics was evaluatedby RPA or semi-quantitative RT-PCR analysis.
6093The Journal of Immunology
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

are involved in cell cycle progression. Additionally, pim-1 andc-myc have been reported to act synergistically to prevent apopto-sis (24) and thus may play a novel role in protecting NK cells fromstress-induced cellular death.
The NK92 cell line is a good model for studying changes incytokine gene expression; however, it is cytokine-dependent andmay not entirely reflect changes in primary NK cells. To addressthis concern, we compared alterations in cell cycle gene expressionin primary and NK92 cells. Overnight depletion of IL-2 fromNK92 cells might result in changes in cell cycle gene expressionthat could be dramatically different from primary NK cells thathave not been IL-2-depleted in culture.
The results from RT-PCR and RPA analysis of RNA fromNK92 and primary NK cells, respectively, demonstrated that bothcell types had large increases in cyclin D2 mRNA expression after3 h of stimulation by all cytokines (Figs. 3 and 4A). Interestingly,the primary NK cells exhibited a more robust response to IL-2 andIL-18 than did the NK92 cells. Moreover, cyclin D3 mRNA ex-pression was slightly up-regulated in primary NK cells. This effectwas not seen in NK92 cells (data not shown).
We continued our study of cell cycle-related genes by examin-ing c-myc and pim-1 mRNA expression in primary NK cells. RT-PCR analysis revealed that all cytokine treatments could inducec-myc and pim-1 mRNA levels in primary NK cells (Fig. 4B).These observations were in good agreement with RT-PCR resultsfrom NK92 cells. Collectively, these data indicate that regulationof NK92 cells is similar in many ways to primary NK cells withregard to IL-2, IL-2 plus IL-12, and IL-2 plus IL-18 treatments.Thus, NK92 cells appear to be a reliable model to predict globalchanges in gene expression in primary NK cells.
Cytokine-induced changes in chemokine receptor density onprimary NK cells
RPA analysis demonstrated that IL-2 alone and in combinationwith IL-12 or IL-18 had a large impact on cytokine, chemokine,and chemokine receptor expression in NK92 cells. To determine
whether these changes were functional in primary NK cells, weexamined the potential for cytokine-induced alterations in chemo-kine receptor expression. Recent studies have demonstrated thepresence of chemokine receptors on NK cells (9, 25–27); however,no studies have addressed alterations in these receptors in responseto short-term activation by IL-2 alone and in combination withIL-12 or IL-18.
To examine changes in chemokine receptor density, freshly iso-lated NK cells that were greater than 95% CD56�/CD5� by FACSanalysis, were placed into culture and stimulated with IL-2, IL-12,and IL-18 alone. Additionally, cells were stimulated with IL-2 incombination with IL-12 or IL-18. After 24 h of cytokine stimula-tion, alterations in CCR1, CCR2, CXCR3, and CXCR4 densitieswere evaluated by FACS analysis. Expression of CCR1 and CCR2was virtually absent in all treatments (Table III). The lack of CCR1expression was in direct contrast to CCR1 mRNA expression inNK92 cells. We considered the likelihood that there were differ-ences in the expression of this receptor between the NK92 cell lineand primary cells; however, it has been recently reported that pri-mary NK cells express CCR1 mRNA although surface expressionof the receptor is undetectable (9).
Examination of CXCR3 surface expression revealed dramaticdecreases in receptor density in response to cytokine treatment. Incells treated with IL-2 and IL-12 alone, there was a 28 and 26%reduction in CXCR3 expression, respectively (Table III and Fig.5). The combination of these cytokines was synergistic with a 68%decrease in receptor density. IL-18 alone or in combination withIL-2 had no effect on CXCR3 expression. There was generalagreement between CXCR3 expression in primary NK cells andmRNA expression in NK92 cells with regard to IL-2 and IL-12regulation of this receptor. In contrast, RPA analysis of CXCR3mRNA expression in NK92 cells predicted that the maximum de-crease in receptor density should result from IL-2 plus IL-18 treat-ment. This prediction was not substantiated in primary NK cellsand may reflect differences in the manner by which primary NKcells and NK92 respond to IL-2 plus IL-18 with respect to RNA
Table II. Analysis of gene expression in IL-2 � IL-12 and IL-2 � IL-18–treated NK92 cells
Gene NameGenBank Accession
NumberIL-2 � IL-12–Stimulated
SignalIL-2 � IL-18–Stimulated
SignalFold
Change
Genes down–regulated by IL-2 � IL-18 ascompared to IL-2 � IL-12
Ornithine decarboxylase 1 M16650 2,372 1,134 �2.09Peptidylprolyl isomerase E (cyclophilin E) AF042386 810 320 �2.53GTP-binding protein overexpressed in skeletal
muscleU10550 719 134 �5.37
PGER2 (subtype EP2) U19487 477 225 �2.12EST highly similar to DNAJ protein homolog
MTJ1 (M. musculus)AI337322 236 115 �2.05
EST AI890347 145 68 �2.13Hyaluronan-mediated motility receptor (RHAMM) U29343 128 58 �2.21EST R55801 122 55 �2.22EST AA868888 113 37 �3.05V-maf musculoaponeurotic fibrosarcoma (avian)
oncogene family, protein GAF059195 108 46 �2.35
Genes up-regulated by IL-2 � IL-18 as comparedto IL-2 � IL-12
Apoptosis inhibitor 2 U37546 724 2,466 3.41NF of � light polypeptide gene enhancer in B cell
inhibitor, �M69043 546 1,752 3.21
Discoidin domain receptor family, member 1 Z29093 663 1,334 2.01Carboxypeptidase D D85390 427 1,007 2.36Regulator of G-protein signaling 16 U70426 327 674 2.06Hypoxia-inducible factor 1, � subunit (basic helix-
loop-helix transcription factor)U22431 239 536 2.24
Apoptosis inhibitor 1 U37547 181 477 2.64
6094 CYTOKINE-INDUCED CHANGES IN NK CELL GENE EXPRESSION
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

and protein expression of CXCR3. Finally, we examined changesin CXCR4 receptor expression. Our RPA results suggested thatthere should be cytokine-independent expression of CXCR4 in NKcells. Flow cytometry analysis demonstrated that �25% of pri-mary NK cells expressed CXCR4 (Table III). Moreover, receptordensity remained unchanged with respect to cytokine stimulation.
Alterations in chemotaxis in cytokine-treated primary NK cells
The decrease in CXCR3 density in IL-2- and IL-12-treated NKcells predicted that these cells would have decreased migratorycapacity in the presence of CXCR3-specific chemoattractants. Totest this assumption, freshly isolated primary NK cells were treatedwith IL-2 and IL-12 alone or in combination for 24 h. Cells wereplaced into microchemotaxis chambers and their migration poten-tial was evaluated in the presence of CXCR3 ligand, IFN-�-induc-ible protein (IP)-10. At IP-10 concentrations of 100 ng/ml orgreater, we observed a significant reduction in chemotaxis in theIL-2- and IL-12-treated cells (Fig. 6). Furthermore, the combina-tion of IL-2 and IL-12 resulted in a substantial decrease in migra-tion that was greater than that observed in cells treated with thecytokines alone (Fig. 6). Collectively, these data demonstrate thatIL-2 and IL-12 can directly alter the chemotactic function of NKcells through changes in chemokine receptor mRNA and proteinexpression.
Down-regulation of CXCR3 mRNA expression
Decreases in CXCR3 density are evident by 24 h after IL-2 plusIL-12 treatment. Therefore, to understand the molecular mecha-nism responsible for these changes, we chose to examine the ki-netic pattern of CXCR3 mRNA expression at times immediatelyfollowing cytokine addition to NK cells. For these studies, primary
human NK cells were isolated and changes in CXCR3 mRNAwere monitored for up to 12 h following IL-2 plus IL-12 addition.In all donors tested, CXCR3 mRNA expression remained rela-tively unchanged for at least 3 h following cytokine treatment (Fig.7, A and B). However, by 6 h of IL-2 plus IL-12 treatment, therewas a slight 20% reduction in CXCR3 mRNA accumulation. TheCXCR3 mRNA continued to decrease rapidly with time until atotal 80% drop in expression was seen after 12 h of cytokine treat-ment. Interestingly, we found that IL-2 plus IL-12 did not nega-tively affect all chemokine receptors. The fractalkine receptor,CX3CR1 is expressed on NK cells and is involved in NK cell-mediated endothelial cell injury (28). In contrast to CXCR3, IL-2plus IL-12 greatly enhanced CX3CR1 mRNA expression (Fig. 7A).By 3 h of IL-2 plus IL-12 treatment, CX3CR1 mRNA increased3-fold. The increase in mRNA expression continued to rise with a4-fold increase after 6 h of cytokine treatment. Moreover, thisincrease in mRNA was sustained throughout the remainder of thetime course.
To understand whether CXCR3 mRNA regulation was entirelydue to transcriptional changes, we determined the half-life of theCXCR3 mRNA in the presence and absence of the transcriptionalinhibitor, actinomycin D. For these experiments, NK92 cells weretreated with IL-2 plus IL-12 � actinomycin D and changes inCXCR3 mRNA were measured at various times posttreatment. AsIL-2 plus IL-12 reduces the quantity of CXCR3 mRNA, theamount of CXCR3 mRNA measured at all times was expressed asa percentage of that observed at the initial time of cytokine addi-tion. In the absence of actinomycin D, NK92 cells responded toIL-2 plus IL-12 with the expected reduction in CXCR3 mRNA(Fig. 8, A and B). Under these conditions, the CXCR3 mRNAhalf-life was estimated to be 1.5 h (Fig. 8B). The addition of ac-tinomycin D slightly increased the half-life of the CXCR3 mRNA
FIGURE 1. Multiprobe RNase protection analysis of gene expression inNK92 cells. NK92 cells were rested overnight in RPMI medium withoutIL-2 and IL-15. Cells were either untreated (NS) or stimulated with IL-2(100 U/ml) alone or in combination with IL-12 (10 U/ml) and IL-18 (10ng/ml) for 3 h. Total RNA was isolated as described in Materials andMethods and �10 �g of input RNA was used per hybridization. All RPAswere performed by hybridizing RNA to hCK1, hCK5, hCR5, hCR6, orhAPO5 multiprobe templates (BD PharMingen). The exposure time to filmvaried for individual RPAs and depended on the strength of the signal ofthe target genes. Differences in exposure are reflected by differences inGAPDH signal intensities. The data are representative of two independentexperiments.
FIGURE 2. Single-probe RPA analysis of mRNA expression in NKcells. All probes were in vitro synthesized from gene specific sequencesand gel purified before hybridization. The RPAs were performed accordingto manufacturer specifications using a RPA II kit (Ambion). The gels wereexposed to film for a minimum of 24 h; however, exposure times varied forindividual RPAs and depended on the strength of the signal of the targetgenes. The data are representative of two independent experiments.
6095The Journal of Immunology
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

from 1.5 to 2 h (Fig. 8B). This slight increase in the mRNA half-life was surprising in that mRNA stabilization, in the absence ofother factors, should lead to an overall increase in mRNA accu-mulation. However, we found that IL-2 plus IL-12 act collectivelyto decrease CXCR3 expression. Although this small increase inCXCR3 mRNA half-life suggests that posttranscriptional effectsmay contribute to the regulation of CXCR3 expression, it shouldbe noted that the overall increase in mRNA half-life is small andthat the rate of CXCR3 mRNA decay is similar in the absence orpresence of actinomycin D. Thus, the overall increase in CXCR3
mRNA half-life may be insignificant and other as yet unknownfactors may contribute to the down-regulation of CXCR3expression.
DiscussionNK cells play a pivotal role in protecting the body against infec-tious agents and cancers (1, 29, 30). Thus, cytokine-induced vari-ations in gene expression in NK cells may have profound effects onNK cell biology and immune response. In this study, we haveexamined global changes in gene expression in response to IL-2alone and in combination with IL-12 or IL-18 and found that 82genes changed at least 2-fold in response to cytokine treatment.Collectively, alterations in mRNA expression were associated withgenes whose protein products are involved with multiple cellularfunctions such as, cell cycle progression, signal transduction, tran-scriptional activation, and mRNA processing. However, the ma-jority of genes that responded to cytokine treatment code for che-mokines, chemokine receptors, and cytokines.
Of the altered chemokine receptor genes identified, we chose tofurther investigate the effect of IL-2 and IL-12 on CXCR3 expres-sion. Cellular distribution of CXCR3 is mainly localized to acti-vated T cells, eosinophils, basophils, and NK cells but control ofthis expression is poorly understood. Alterations in CXCR3 recep-tor and mRNA expression have been studied in IL-2-treated T andNK cells (9, 31); however, cells were incubated in the presence ofIL-2 for many days before mRNA analysis was made. This dem-onstrated the long-term effects of IL-2, but did not address rapidshort-term effects of cytokines on CXCR3 expression.
FIGURE 4. RPA and RT-PCR analysis of cell cycle-associated genes inprimary human NK cells. Primary NK cells were stimulated with IL-2 (100U/ml) alone or in combination with IL-12 (10 U/ml) or IL-18 (10 ng/ml)for 3 h. Total mRNA was isolated from the cells and mRNA expressionwas evaluated. A, mRNA expression of cyclins D2 and D3 were examinedby RPA using a hCYC-1 multiprobe. GAPDH was included as a control forsample-to-sample variation. B, Relative-quantitative RT-PCR analysis c-myc and Pim-1 expression. 18S ribosomal RNA was used as an internalcontrol in this assay. These results are representative of assays performedon RNA from two independent donors.
Table III. FACS analysis of cytokine-induced changes in surfacechemokine receptor expression in primary NK cellsa
ChemokineReceptor NS IL-2 IL-12 IL-18
IL-2�
IL-12
IL-2�
IL-18
CCR1 0 0 0 0 0 0CCR2 0 0 0 0 0 0CXCR3 81 58 60 83 26 55CXCR4 25 28 26 27 26 29
a These data represent one of two separate experiments. For both experiments,freshly isolated NK cells were from individual donors.
FIGURE 3. Relative-quantitative RT-PCR analy-sis of alterations in gene expression in NK92 cells.Approximately 5 �g of RNA was reverse transcribedusing a Thermoscript RT-PCR system (Life Technol-ogies). The resulting cDNA template was used formultiplex PCR with gene-specific and human 18SrRNA gene primers. The final PCR products weresize-separated by agarose gel electrophoresis and vi-sualized using ethidium bromide staining. The mi-croarray values for the NS and IL-2-treated samplesare listed to the left of each gene tested. No values aregiven for the IFN-� gene as it was not represented onthe microarray chip. However, relative-quantitativeRT-PCR analysis was performed on the IFN-� geneas a positive control for cytokine-induced changes ingene expression.
6096 CYTOKINE-INDUCED CHANGES IN NK CELL GENE EXPRESSION
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Our data show that IL-12 and IL-2 alone and in combinationeffectively down-regulate CXCR3 cell surface expression in NKcells. Furthermore, alterations in CXCR3 mRNA precede changesin protein expression suggesting that changes in CXCR3 surfaceexpression are due to rapid changes in mRNA accumulation andare not due to receptor internalization. Initially, we observed anIL-2 plus IL-12 mediated down-regulation of CXCR3 mRNA ex-pression in NK92 cells. This observation was extended to primaryhuman NK cells. The kinetics of IL-2 plus IL-12 down-regulationof CXCR3 mRNA in primary NK cells demonstrated that CXCR3mRNA expression was relatively constant after 3 h of cytokinetreatment; however, by 6 h, an �25% decrease in mRNA accu-mulation was seen. At times following, CXCR3 mRNA continuedto decline. The decline in CXCR3 mRNA was also observed inNK92 cells but was more rapid with a significant decline visual-ized by 3 h of IL-2 plus IL-12 treatment. The difference in decay
rates between NK92 and primary NK cells did not affect the over-all outcome of CXCR3 down-regulation and may reflect differ-ences in cultured and primary cells.
To closely examine the mechanism of CXCR3 mRNA regula-tion, we conducted CXCR3 mRNA half-life studies with actino-mycin D to determine whether regulation was entirely due to tran-scriptional effects. We expected that the transcriptional blocker,actinomycin D, would either have no effect or would shorten thehalf-life of CXCR3 mRNA as an increase in CXCR3 mRNA half-life would reflect a mRNA stabilization event that should ulti-mately lead to an increase in mRNA accumulation. Surprisingly,we found that concurrent addition of actinomycin D, IL-2, andIL-12 to NK92 cells slightly increased the half-life of the CXCR3
FIGURE 6. Effect of IL-2 and IL-12 on chemotaxis of primary NKcells. Freshly isolated primary NK cells were stimulated with 100 U/mlIL-2 and 10 U/ml IL-12, alone or in combination for 24 h. The NK cellswere placed into the upper compartment of a microchemotaxis chamberwhile the chemoattractant, IP-10, was placed into the lower compartmentat the various concentrations indicated. The amount of chemotactic activitywas evaluated following a 4-h incubation. All results are expressed as themean � SD for triplicate determinations. Significant changes in migrationas compared with medium alone are indicated by an asterisk (�, p 0.05).The data are from one representative experiment of two performed.
FIGURE 7. Kinetics of CXCR3 mRNA accumulation in primary hu-man NK cells. A, Freshly isolated NK cells were treated with IL-2 (100U/ml) plus IL-12 (10 U/ml) for the times indicated. Total mRNA wasisolated and CXCR3 and CX3CR1 mRNAs were measured by RPA anal-ysis using a hCR6 multiprobe. GAPDH was included as a control for sam-ple-to-sample variation. A 24-h exposure is shown for all mRNAs. A 12-hexposure (exp) of GAPDH mRNA is included to demonstrate sample andgel loading variations. B, Graphical representation of the percent change inCXCR3 mRNA shown in A. The amount of CXCR3 mRNA at time 0 wasarbitrarily assigned as 100%. This experiment is representative of threeindependent assays performed on three separate donors.
FIGURE 8. Half-life of CXCR3 mRNA in IL-2 plus IL-12-treatedNK92 cells. A, NK92 cells were rested overnight without IL-2 and IL-15and then treated with IL-2 (100 U/ml), IL-12 (10 U/ml) � actinomycin D(5 �g/ml) for the times indicated. Total mRNA was isolated from the cellsand a multiprobe RPA analysis was performed with a hCR-6 probe set.GAPDH was included as an internal control for sample-to-sample varia-tion. The gel was exposed to a PhosphorImager screen for 24 h. B, Graph-ical representation of the quantitation performed by ImageQuant analysisof the image shown in A. The amount of CXCR3 mRNA at time 0 wasarbitrarily set to 100%. The experiment is one of three separate assays, allwith similar results.
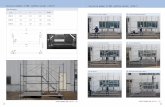
FIGURE 5. Inhibition of CXCR3 expression by IL-2 and IL-12. Freshlyisolated primary NK cells were stimulated with 100 U/ml IL-2, 10 U/mlIL-12, or 10 ng/ml IL-18, alone or in combination for 24 h and CXCR3expression was compared with that observed in nontreated (NT) cells byflow cytometry analysis. A, Receptor expression in cells treated with IL-2and IL-12 alone and in combination. B, Receptor expression in cells treatedwith IL-2 and IL-18 alone and in combination. In both A and B, cntl rep-resents isotype control Ig staining. The data are from one representativeexperiment of two performed. Individual donors were used as the source ofNK cells for each experiment.
6097The Journal of Immunology
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

mRNA suggesting that these cytokines have a stabilizing effect onthe CXCR3 mRNA. These data demonstrate that IL-2 plus IL-12control of CXCR3 mRNA expression is not entirely at the tran-scriptional level and that as yet unknown posttranscriptional ef-fectors must negate any stabilization effect of IL-2 plus IL-12 onCXCR3 mRNA so that accumulation is ultimately reduced.
Overall, our data suggest that IL-2 and IL-12 alone or in com-bination have the ability to modulate immune function by alteringCXCR3 receptor expression on NK cells. This is particularly im-portant because NK cells are primary cellular defenders againstviral infections and viral infections induce cellular release of po-tent cytokines and chemokines. The chemokine MIP-1� is a potentinducer of NK cell chemotaxis (32, 33) and has been shown toinduce NK cell migration to livers of murine CMV-infected mice(34). Once in the liver, NK cells produce IFN-� in an IL-12-de-pendent fashion that in turn up-regulates the expression of CXCR3receptor ligands monocyte interferon-�-inducible protein, IFN-�-inducible T-cell � chemoattractant, and IP-10 (34–37). Concurrentwith the IL-12 up-regulation of IFN-�, it is possible that IL-12alone or in combination with IL-2 down-regulates CXCR3 expres-sion on NK cells thus reducing NK cell chemotactic responsive-ness to CXCR3 ligands. As monocyte interferon-�-inducible pro-tein, IFN-�-inducible T-cell � chemoattractant, and IP-10 canpromote the migration of activated Th1 lymphocytes to inflamma-tory sites, these chemokines may then initiate a secondary T cell-mediated immune response. This model illustrates how IL-12alone and in combination with IL-2 may trigger both primary andsecondary immune responses, thus profoundly affecting NK cellbiology and immune responses.
AcknowledgmentsWe thank William Bere and Stephanie Krebs for their assistance with pri-mary NK cell isolation and RPAs. We are also indebted to Wanghua Gongfor her assistance with the chemotaxis assays.
References1. Trinchieri, G. 1989. Biology of natural killer cells. Adv. Immunol. 47:187.2. Herberman, R. B., and J. R. Ortaldo. 1981. Natural killer cells: their roles in
defenses against disease. Science 214:24.3. Biron, C. A., K. B. Nguyen, G. C. Pien, L. P. Cousens, and T. P. Salazar-Mather.
1999. Natural killer cells in antiviral defense: function and regulation by innatecytokines. Annu. Rev. Immunol. 17:189.
4. Maghazachi, A. A. 2000. Chemokines, G proteins and natural killer cells. Arch.Immunol. Ther. Exp. 48:65.
5. DeRisi, J. L., V. R. Iyer, and P. O. Brown. 1997. Exploring the metabolic andgenetic control of gene expression on a genomic scale. Science 278:680.
6. Iyer, V. R., M. B. Eisen, D. T. Ross, G. Schuler, T. Moore, J. C. Lee, J. M. Trent,L. M. Staudt, J. Hudson, Jr., M. S. Boguski, et al. 1999. The transcriptionalprogram in the response of human fibroblasts to serum. Science 283:83.
7. Ross, D. T., U. Scherf, M. B. Eisen, C. M. Perou, C. Rees, P. Spellman, V. Iyer,S. S. Jeffrey, M. Van de Rijn, M. Waltham. 2000. Systematic variation in geneexpression patterns in human cancer cell lines. Nat. Genet. 24:227.
8. Perou, C. M., S. S. Jeffrey, M. Van de Rijn, C. A. Rees, M. B. Eisen, D. T. Ross,A. Pergamenschikov, C. F. Williams, S. X. Zhu, J. C. Lee, et al. 1999. Distinctivegene expression patterns in human mammary epithelial cells and breast cancers.Proc. Natl. Acad. Sci. USA 96:9212.
9. Inngjerdingen, M., B. Damaj, and A. A. Maghazachi. 2001. Expression and reg-ulation of chemokine receptors in human natural killer cells. Blood 97:367.
10. Timonen, T., J. R. Ortaldo, and R. B. Herberman. 1981. Characteristics of humanlarge granular lymphocytes and relationship to natural killer and K cells. J. Exp.Med. 153:569.
11. Chan, S. H., M. Kobayashi, D. Santoli, B. Perussia, and G. Trinchieri. 1992.Mechanisms of IFN-� induction by natural killer cell stimulatory factor (NKSF/IL-12): role of transcription and mRNA stability in the synergistic interactionbetween NKSF and IL-2. J. Immunol. 148:92.
12. Chan, S. H., B. Perussia, J. W. Gupta, M. Kobayashi, M. Pospisil, H. A. Young,S. F. Wolf, D. Young, S. C. Clark, and G. Trinchieri. 1991. Induction of inter-feron � production by natural killer cell stimulatory factor: characterization of theresponder cells and synergy with other inducers. J. Exp. Med. 173:869.
13. Ahn, H. J., S. Maruo, M. Tomura, J. Mu, T. Hamaoka, K. Nakanishi, S. Clark,M. Kurimoto, H. Okamura, and H. Fujiwara. 1997. A mechanism underlyingsynergy between IL-12 and IFN-�-inducing factor in enhanced production ofIFN-�. J. Immunol. 159:2125.
14. Ye, J., J. R. Ortaldo, K. Conlon, R. Winkler-Pickett, and H. A. Young. 1995.Cellular and molecular mechanisms of IFN-� production induced by IL- 2 andIL-12 in a human NK cell line. J. Leukocyte Biol. 58:225.
15. Bohn, E., A. Sing, R. Zumbihl, C. Bielfeldt, H. Okamura, M. Kurimoto, J. Heese-mann, and I. B. Autenrieth. 1998. IL-18 (IFN-�-inducing factor) regulates earlycytokine production in, and promotes resolution of, bacterial infection in mice.J. Immunol. 160:299.
16. Mehrotra, P. T., R. P. Donnelly, S. Wong, H. Kanegane, A. Geremew,H. S. Mostowski, K. Furuke, J. P. Siegel, and E. T. Bloom. 1998. Production ofIL-10 by human natural killer cells stimulated with IL-2 and/or IL-12. J. Immu-nol. 160:2637.
17. Lauwerys, B. R., N. Garot, J. C. Renauld, and F. A. Houssiau. 2000. Cytokineproduction and killer activity of NK/T-NK cells derived with IL-2, IL-15, or thecombination of IL-12 and IL-18. J. Immunol. 165:1847.
18. D’Andrea, A., M. Aste-Amezaga, N. M. Valiante, X. Ma, M. Kubin, andG. Trinchieri. 1993. Interleukin 10 (IL-10) inhibits human lymphocyte interferon�-production by suppressing natural killer cell stimulatory factor/IL-12 synthesisin accessory cells. J. Exp. Med. 178:1041.
19. Oliva, A., A. L. Kinter, M. Vaccarezza, A. Rubbert, A. Catanzaro, S. Moir,J. Monaco, L. Ehler, S. Mizell, R. Jackson, et al. 1998. Natural killer cells fromhuman immunodeficiency virus (HIV)-infected individuals are an importantsource of CC-chemokines and suppress HIV-1 entry and replication in vitro.J. Clin. Invest. 102:223.
20. Chieux, V., W. Chehadeh, J. Harvey, O. Haller, P. Wattre, and D. Hober. 2001.Inhibition of coxsackievirus B4 replication in stably transfected cells expressinghuman MxA protein. Virology 283:84.
21. Staeheli, P., O. Haller, W. Boll, J. Lindenmann, and C. Weissmann. 1986. Mxprotein: constitutive expression in 3T3 cells transformed with cloned Mx cDNAconfers selective resistance to influenza virus. Cell 44:147.
22. Anderson, S. L., J. M. Carton, J. Lou, L. Xing, and B. Y. Rubin. 1999. Interferon-induced guanylate binding protein-1 (GBP-1) mediates an antiviral effect againstvesicular stomatitis virus and encephalomyocarditis virus. Virology 256:8.
23. Shirogane, T., T. Fukada, J. M. Muller, D. T. Shima, M. Hibi, and T. Hirano.1999. Synergistic roles for Pim-1 and c-Myc in STAT3-mediated cell cycle pro-gression and antiapoptosis. Immunity 11:709.
24. Qin, S., J. B. Rottman, P. Myers, N. Kassam, M. Weinblatt, M. Loetscher,A. E. Koch, B. Moser, and C. R. Mackay. 1998. The chemokine receptorsCXCR3 and CCR5 mark subsets of T cells associated with certain inflammatoryreactions. J. Clin. Invest. 101:746.
25. Inngjerdingen, M., B. Damaj, and A. A. Maghazachi. 2000. Human NK cellsexpress CC chemokine receptors 4 and 8 and respond to thymus and activation-regulated chemokine, macrophage-derived chemokine, and I-309. J. Immunol.164:4048.
26. Campbell, J. J., S. Qin, D. Unutmaz, D. Soler, K. E. Murphy, M. R. Hodge,L. Wu, and E. C. Butcher. 2001. Unique subpopulations of CD56� NK and NK-Tperipheral blood lymphocytes identified by chemokine receptor expression rep-ertoire. J. Immunol. 166:6477.
27. Yoneda, O., T. Imai, S. Goda, H. Inoue, A. Yamauchi, T. Okazaki, H. Imai,O. Yoshie, E. T. Bloom, N. Domae, and H. Umehara. 2000. Fractalkine-mediatedendothelial cell injury by NK cells. J. Immunol. 164:4055.
28. Biron, C. A. 1997. Activation and function of natural killer cell responses duringviral infections. Curr. Opin. Immunol. 9:24.
29. Scott, P., and G. Trinchieri. 1995. The role of natural killer cells in host-parasiteinteractions. Curr. Opin. Immunol. 7:34.
30. Loetscher, M., P. Loetscher, N. Brass, E. Meese, and B. Moser. 1998. Lympho-cyte-specific chemokine receptor CXCR3: regulation, chemokine binding andgene localization. Eur. J. Immunol. 28:3696.
31. Maghazachi, A. A., A. al Aoukaty, and T. J. Schall. 1994. C-C chemokinesinduce the chemotaxis of NK and IL-2-activated NK cells: role for G proteins.J. Immunol. 153:4969.
32. Taub, D. D., T. J. Sayers, C. R. Carter, and J. R. Ortaldo. 1995. � and � che-mokines induce NK cell migration and enhance NK-mediated cytolysis. J. Im-munol. 155:3877.
33. Salazar-Mather, T. P., J. S. Orange, and C. A. Biron. 1998. Early murine cyto-megalovirus (MCMV) infection induces liver natural killer (NK) cell inflamma-tion and protection through macrophage inflammatory protein 1� (MIP-1�)-de-pendent pathways. J. Exp. Med. 187:1.
34. Wright, T. M., and J. M. Farber. 1991. 5� regulatory region of a novel cytokinegene mediates selective activation by interferon �. J. Exp. Med. 173:417.
35. Farber, J. M. 1990. A macrophage mRNA selectively induced by �-interferonencodes a member of the platelet factor 4 family of cytokines. Proc. Natl. Acad.Sci. USA 87:5238.
36. Cole, K. E., C. A. Strick, T. J. Paradis, K. T. Ogborne, M. Loetscher,R. P. Gladue, W. Lin, J. G. Boyd, B. Moser, D. E. Wood, et al. 1998. Interferon-inducible T cell � chemoattractant (I-TAC): a novel non-ELR CXC chemokinewith potent activity on activated T cells through selective high affinity binding toCXCR3. J. Exp. Med. 187:2009.
37. Farber, J. M. 1997. Mig and IP-10: CXC chemokines that target lymphocytes.J. Leukocyte Biol. 61:246.
6098 CYTOKINE-INDUCED CHANGES IN NK CELL GENE EXPRESSION
by guest on May 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from