Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by...
4
British Joiirnal of Haematologg. 1993, 85, 352-355 Haemoglobin Manukau J?67[E11] Val-+Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia KEITH C. FAY, STEPHEN 0. BRENNAN,* JACK M. COSTELLO, HOWARD C. POTTER,* DAVID A. WILLIAMSON,* RONALD J. TRENT,? P A U L A. OCKELFORD AND D. Ross BOSWELL* Departments of Haematology and Paediatrics, Auckland Hospital, Auckland, New Zealand, *Department of Clinical Biochemistry, Christchurch Hospital, Christchurch, New Zealand, and ?Department of Molecular Genetics, Royal Prince Alfred Hospital, Camperdown, NSW, Australia Received 75 March 1993; accepted for publication 23 April 1993 Summary. Haemoglobin Manukau (p67 Val-Gly) is a novel haemoglobin variant presenting in two brothers as non- spherocytic haemolytic anaemia which became transfusion dependent by 6 months of age. The severity of clinical expression seems to be modulated by coexisting alpha thalassaemia: the severely affected children have a normal complement of alpha globin genes with an unusual genotype ( -G?~/cICICI~'~), while their father, who carries the abnormal gene with minimal symptoms, has homozygous CI + thalassae- mia ( - a3 7/ - a39 Another unusual feature of this case is the association of the 867 Val-Gly mutation with modification of p141 Leu to a residue (believed to be hydroxyleucine) that is not detected by standard amino acid analysis. This finding offers an explanation for the previous report of an association of another mutation at this site (Hb Sydney 867 Val+Ala) with Hb Coventry (deletion of 8141 Leu). It is well recognized that the expression of beta-thalassaemia major can be ameliorated by associated alpha-thalassaemia, and conversely, that the presence of a triplicated alpha globin gene cluster may convert beta-thalassaemia trait to thalas- saemia intermedia. We present here a new beta-globin mutation (Hb Manukau) expressed as severe haemolytic anaemia in family members with a normal complement of alpha globin genes, but minimally expressed in a member with two-gene alpha thalassaemia. This beta-globin mutation is also associated with Hb Coventry (apparent deletion of p141 Leu). Hb Coventry was first described in association with Hb Sydney (867 Val- Ala) (Casey et al, 1978) and was thought to represent a third beta- globin gene with deletion of B141 Leu. It has also been found in association with Hb Atlanta (p75 Leu-*Pro) (Brennan et al, 1992), and was recognized in that case to result from a post-translational modification of Leu 141 to a residue with a mass 16 D greater. This residue, believed to be hydroxyleu- cine, is not detected by standard amino acid analysis and sequencing methods. The hydroxyleucine modification of Leu 141 offers a more parsimonious explanation than the Correspondence: Dr K. C. Fay. Department of Haematology. Auck- land Hospital, Auckland. New Zealand. postulated third gene of Casey et a1 (1 9 78) for their findings in this and the Hb Sydney case. METHODS Haematological investigations followed standard procedures (Dacie & Lewis, 1975) and standard techniques were used for measurement of HbA2 and HbF, and for stability tests (Carrel1 & Kay, 1972). Alpha globin gene mapping was performed as previously described (Old, 1983; Trend et al, 1984). Globin from isopropanol precipitates was digested with trypsin and mapped on Whatman 3MM paper (Brennan et al, 1981). After detection with fluorescamine, peptides were eluted and subjected to amino acid analysis and to liquid secondary ion mass spectrometry in a VG70-250s double focusing magnetic sector instrument equipped with a cae- sium ion gun (Brennan et aI. 1992). PCR method. A 600 bp fragment from the human jl globin gene was amplified by PCR using primers 1 and 2 in a standard 100 pl reaction volume (Saiki et RI. 1988). Follow- ing ethanol precipitation and washing to remove unincorpor- ated primers, the DNA was dissolved in 40 jtl of HLO. 1 p1 of this was used as template for a single stranded re-amplifica- tion in a 100 pl reaction using only primer 2. Following phenol/chloroform extraction and ethanol precipitation, the 352
-
Upload
keith-c-fay -
Category
Documents
-
view
213 -
download
0
Transcript of Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by...
![Page 1: Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia](https://reader035.fdocument.org/reader035/viewer/2022080402/575081021a28abf34f8bdd80/html5/thumbnails/1.jpg)
British Joiirnal of Haematologg. 1993 , 85, 352-355
Haemoglobin Manukau J?67[E11] Val-+Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia
KEITH C. FAY, STEPHEN 0. BRENNAN,* JACK M. COSTELLO, HOWARD C. POTTER,* DAVID A. WILLIAMSON,* RONALD J . TRENT,? PAUL A. OCKELFORD A N D D. Ross BOSWELL* Departments of Haematology and Paediatrics, Auckland Hospital, Auckland, New Zealand, *Department of Clinical Biochemistry, Christchurch Hospital, Christchurch, New Zealand, and ?Department of Molecular Genetics, Royal Prince Alfred Hospital, Camperdown, N S W , Australia
Received 75 March 1 9 9 3 ; accepted for publication 23 April 1 9 9 3
Summary. Haemoglobin Manukau (p67 Val-Gly) is a novel haemoglobin variant presenting in two brothers as non- spherocytic haemolytic anaemia which became transfusion dependent by 6 months of age. The severity of clinical expression seems to be modulated by coexisting alpha thalassaemia: the severely affected children have a normal complement of alpha globin genes with an unusual genotype ( - G ? ~ / c I C I C I ~ ' ~ ) , while their father, who carries the abnormal
gene with minimal symptoms, has homozygous CI + thalassae- mia ( - a3 7/ - a 3 9 Another unusual feature of this case is the association of the 867 Val-Gly mutation with modification of p141 Leu to a residue (believed to be hydroxyleucine) that is not detected by standard amino acid analysis. This finding offers an explanation for the previous report of an association of another mutation at this site (Hb Sydney 867 Val+Ala) with Hb Coventry (deletion of 8141 Leu).
It is well recognized that the expression of beta-thalassaemia major can be ameliorated by associated alpha-thalassaemia, and conversely, that the presence of a triplicated alpha globin gene cluster may convert beta-thalassaemia trait to thalas- saemia intermedia. We present here a new beta-globin mutation (Hb Manukau) expressed as severe haemolytic anaemia in family members with a normal complement of alpha globin genes, but minimally expressed in a member with two-gene alpha thalassaemia.
This beta-globin mutation is also associated with Hb Coventry (apparent deletion of p141 Leu). Hb Coventry was first described in association with Hb Sydney (867 Val- Ala) (Casey et al, 1978) and was thought to represent a third beta- globin gene with deletion of B141 Leu. It has also been found in association with Hb Atlanta (p75 Leu-*Pro) (Brennan et al, 1992), and was recognized in that case to result from a post-translational modification of Leu 141 to a residue with a mass 16 D greater. This residue, believed to be hydroxyleu- cine, is not detected by standard amino acid analysis and sequencing methods. The hydroxyleucine modification of Leu 141 offers a more parsimonious explanation than the
Correspondence: Dr K. C. Fay. Department of Haematology. Auck- land Hospital, Auckland. New Zealand.
postulated third gene of Casey et a1 (1 9 78) for their findings in this and the Hb Sydney case.
METHODS
Haematological investigations followed standard procedures (Dacie & Lewis, 1975) and standard techniques were used for measurement of HbA2 and HbF, and for stability tests (Carrel1 & Kay, 1972). Alpha globin gene mapping was performed as previously described (Old, 1983; Trend et al, 1984).
Globin from isopropanol precipitates was digested with trypsin and mapped on Whatman 3MM paper (Brennan et al, 1981). After detection with fluorescamine, peptides were eluted and subjected to amino acid analysis and to liquid secondary ion mass spectrometry in a VG70-250s double focusing magnetic sector instrument equipped with a cae- sium ion gun (Brennan et aI. 1992).
PCR method. A 600 bp fragment from the human jl globin gene was amplified by PCR using primers 1 and 2 in a standard 100 pl reaction volume (Saiki et R I . 1988). Follow- ing ethanol precipitation and washing to remove unincorpor- ated primers, the DNA was dissolved in 40 j t l of HLO. 1 p1 of this was used as template for a single stranded re-amplifica- tion in a 100 pl reaction using only primer 2. Following phenol/chloroform extraction and ethanol precipitation, the
352
![Page 2: Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia](https://reader035.fdocument.org/reader035/viewer/2022080402/575081021a28abf34f8bdd80/html5/thumbnails/2.jpg)
Haemoglobin Manukau P67[E1 I] ValbGly 3 5 3 Table I.
Father. Mother, Propositus, Brother, 20 years 20s 18 months 7 months
Hb (g/dl) 13.6 11.6 Transfused 7.3 Reticulocytes 4 3.2 22 17.8 Haptoglobin (g/l) (normal 0.7-3.8) 0.2 1.72 Absent Absent Bilirubin (pmol/l) (normal 2-20) 40 Normal Heinz bodies Rare Absent Absent Absent HbH Rare Absent Absent Absent HbF (%) (normal < 1.0) 0.9 0.09 3 .1 1 5 HbAz (%) (normal 1.6-2.9) 4.4 2.1 4.1 3.9 Isopropanol stability Weak at L O min Neg. POS. at 20 min Pos. at 20 min*
* Age 5 years.
pellet was dissolved in 20 111 of HrO, and 7 pl was used as template for a standard I5S sequencing reaction using primer 3 and Sequenase version 2.0.
Primer 1 Primer 2 Primer 3
GTA CGG CTG TCA TCA CTT AGA CCT CA TGC AGC TTC TCA CAG TGC AGC TCA CT AGG CAC TGA CTC TCT CTG C
CASE HISTORY AND RESULTS
Patient. The male propositus was born on 18 August 1982 at term (birth weight 3.49 kg) to a Niuean mother and New Zealand Maori father. Moderate neonatal jaundice resolved following phototherapy and he was discharged from hospital on the fourth day of life. Physical examination at 1 month of age was unremarkable, but the full blood count revealed a normocytic normochromic anaemia with blood haemo- globin concentration 8.7 g/dl. Marked anisopoikilocytosis. coarse basophilic stippling and normoblastaemia (6 NRBC/ 100 WBC) were present in the blood film, and the reticulocyte count was increased at 166 x 10y/l (6.2%). HbF was 57.7% consistent with age. Supravital staining did not reveal inclusion bodies, despite prolonged incubation with 0.574 Rhodanile blue, and haemoglobin H bodies were not detected. Plasma haptoglobin concentration was unmeasur- ably low.
The clinical course was characterized by progressive worsening of the haemolytic anaemia so that at the age of 6 months the blood haemoglobin concentration was 4.8 g/dl. Screening tests for defects of enzymes of the Embden- Meyerhof and hexose monophosphate pathways, and 5’ nucleotidase were all normal. The sucrose lysis test and direct and indirect Coombs tests were negative. The isopropanol stability test was weakly positive at 20 min at a time when HbF was 17.3%. and repeat of this test when the propositus was 9 months old (HbF 4.2%) again yielded a positive result at 20 min. The presence of a variant haemoglobin was further suggested by an abnormal heat denaturation test performed when the propositus was 3 years of age (3.6% precipitate at 1 h, normal less than 0.9%).
Electrophoresis of the haemolysate on cellulose acetate (pH 8.3) and citrate agar (pH 6.0) failed to show an abnormal
haemoglobin, and isoelectric focusing was normal in the presence and absence of 8 M urea. No abnormalities were detected when tryptic digests of isopropanol-precipitated globin were mapped by reverse phase HPLC. Globin chain synthesis studies using a 50 min incubation with 3H leucine showed balanced globin chain synthesis with an alpha-beta chain ratio of 0.95.
Transfusion was undertaken to maintain the pre-trans- fusion blood haemoglobin concentration above 5.9 g/dl. This was initially achieved with transfusions three or four times per year, but by the age of 7 years monthly transfusions were required. Biochemical evidence of iron overload without end- organ dysfunction was documented prior to commencing iron chelation therapy with desferrioxamine five times weekly at age 6 years. General developmental milestones and growth remained satisfactory, and none of the bony features associated with severe thalassaemic syndromes were mani- fested. Cyanosis was not observed at any time. Progressive splenic enlargement (8 cm below coastal margin) and increasing transfusion requirements prompted splenectomy at 7f years of age, and following this the transfusion requirement was markedly reduced.
A full brother of the propositus, born 1 year later, followed a virtually identical clinical course leading to splenectomy at age 6;. Two other siblings, 6 and 10 years younger, have no symptoms. The family studies are summarized in Table I.
Gene niupping studies. Alpha globin gene mapping (Table 11) showed the father of the propositus to be homozygous for the --x’ ‘ gene deletion common amongst Polynesian peoples.
Table 11.
u-globiu Mean cell gene map volume (fl)
Father - u 3 7 / - d 1 3 7 72 Mother ud1/uuu3 ‘ 84 Propositus -u3’7/ud1u3 ‘ 82 Brother -u317/uuu3 83*
*Normal (71-87 fl) at 3 months.
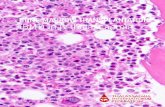
![Page 3: Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia](https://reader035.fdocument.org/reader035/viewer/2022080402/575081021a28abf34f8bdd80/html5/thumbnails/3.jpg)
354 Keith C. Fay et a1
Pig 1. Tryptic peptide map of isopropanol-precipitated Hb Manukau showing more hydrophilic forms of peptides PA9 and PA8-9 (there is minor co- precipitation of HbA). Also present is the ~ C o v c n t r y 1 4 peptide, a minor comparison to PA14.
The mother had a triplicated alpha gene (aaa3 7, on one chromosome, another variation recognized in Polynesians and particularly in Niueans (Trent et aI, 1985; Luan et al, 1985). As expected, both of the sons have inherited a -a3 thalassaemic gene from their father. Both have also inherited the triplicated alpha globin gene cluster from their mother, giving each of them a full complement of alpha genes. This accounts for the normal MCV in the two boys.
After splenectomy, further investigations of the propositus by Rhodamine blue staining showed numerous large Heinz body inclusions, and the isopropanol stability test was strongly positive with marked flocculation at 5 min. Because of the likelihood of obtaining larger amounts of abnormal globin than were previously available, investigation for haemoglobinopathy was repeated. Triton urea electrophore- sis was normal. Two-dimensional tryptic peptide maps of isopropanol-precipitated globin showed that peptide pA9 was replaced by a more hydrophilic peptide p 9 (Fig 1). Mutation in 89 was confirmed by the observation of a new f8-9 peptide. Amino acid analysis of the new p9 peptide showed that it had a composition consistent with the replacement of the single valine residue by glycine. Sequence determination by Edman degradation with identification of PTH derivatives by HPLC confirmed that the new N-terminal residue was glycine, indicating mutation of p67 Val-Gly.
The peptide maps of isopropanol precipitates also showed a companion peptide to pA14. Amino acid analysis of this peptide showed that the single leucine residue was missing, and the peptide was therefore 8Covent ry14 which was pre- viously described in association with Hb Sydney p67 Val-Ala (Casey et al. 1978). Further analysis 0 f ~ ~ ~ ~ ~ ~ ~ ~ ~ 1 4 in the present case by liquid secondary ion mass spectrometry indicated a mass of 11 66 D (expected mass for pA14 11 50 D). This result is consistent with replacement of p141 Leu by a novel amino acid of mass 129 D, probably hydroxyleucine (Brennan et al, 1992).
Investigation of the father showed occasional Heinz bodies in peripheral blood and a weakly positive isopropanol stability test. However, globin mapping studies failed to detect an abnormal peptide.
Sequencing of polymerase chain reaction amplified p globin DNA demonstrated the presence of a G as well as the normal T at the second nucleotide of the ph7 codon. This indicates a GTG(Val)-+GGG(Gly) mutation and confirms that the father is heterozygous for Hb Manukau. The same nucleotide substitution was detected in both sons.
DISCUSSION This report describes a novel haemoglobin variant Hb Manukau (pronounced manookow) which behaves as an unstable haemoglohin producing severe transfusion-depen- dent Heinz-body haemolytic anaemia, and in this respect its clinical expression parallels that of Hb Bristol (p67 Val-Asp) (Cathie, 1952: Steadman et al, 1970). Two other mutations at this site have quite different expressions: Hb Milwaukee (p67 Val-Glu) (Pisciotha et al, 1959) gives rise to congenital mathaemoglobinaemia, and Hb Sydney (867 Val- Ala) (Raik et al. 1967) produces mild Heinz-body haemolytic anaemia.
The sidechain of p67 Val makes important contacts with the haem plate (Perutz et al, 1965) and the residue is phylogenetically highly conserved. The spectrum of clinical presentations seen with substitutions at this position is consistent with current understanding of the importance of the packing of interior hydrophobic sidechains. In Hb Sydney, the substitution of alanine for valine results in loss of two methyl groups and produces some instability in the protein. The substitution of glycine for valine at this site in Hb Manukau involves loss of three methyl groups, and as expected this produces greater instability and a more severe clinical syndrome.
Although Hb Manukau is clinically similar to Hb Bristol, the latter derives its instability from the substitution of a small charged residue into the hydrophobic haem pocket (Stead- man et al, 1970). In Hb Milwaukee (p67 Val+Glu) substitu- tion of a longer charged sidechain allows formation of a salt bridge with the ferrous ion of haem, oxidizing it to the ferric state and accounting for the clinical finding of congenital methaemoglobinaemia (Perutz & Lehmann, 1968).
![Page 4: Haemoglobin Manukau β67[E11] Val→Gly: transfusion-dependent haemolytic anaemia ameliorated by coexisting alpha thalassaemia](https://reader035.fdocument.org/reader035/viewer/2022080402/575081021a28abf34f8bdd80/html5/thumbnails/4.jpg)
Haemoglobin Manukau p6 7[E7 11 Val +Gly 3 5 5 The difficulty in detecting Hb Manukau is explained by two
features: the lack of charge change that prevented its demonstration by electrophoretic techniques: but, more importantly, the moderate instability with rapid degradation of the abnormal haemoglobin in vivo that led to its low concentration in peripheral blood prior to splenectomy. This latter feature is seen also in the hyper-unstable variant Hb Cagliari (/I60 Val-iGlu) (Podda et al, 1991) where rapid degradation results in a normal electrophoretic pattern prior to splenectomy. However. with Hb Cagliari, the gross instability leads to measurable imbalance in apparent ratio of a1pha:beta chain synthesis in vitro. Hb Manukau is less unstable, and the 50 min incubation chain ratio was normal.
Despite the highly variable clinical expression of the different unstable haemoglobins, they tend to run true within the same kindred (Lehmann. 1986; Dacie, 1988). Investiga- tion of the father failed to demonstrate direct evidence of the abnormal globin, but gene sequencing demonstrated that he has the Hb Manukau gene. It is surprising, therefore, that the propositus and his brother have severe haemolytic anaemia while their father has mild fully-compensated haemolytic anaemia. This raises the question of the significance of the associated alpha globin gene abnormalities and the possibi- lity that these may modulate the clinical expression of the unstable haemoglobin.
The deletion of two alpha globin genes in the father would be expected to result in competition between normal and Manukau beta chains for a limited pool of alpha chains. If the abnormal beta globin competes less effectively, it would be left to form highly unstable p4 tetramers which would be destroyed proteolytically during erythropoiesis. The red cells released into circulation would be deficient in haemoglobin. consistent with the alpha thalassaemia. but the haemoglobin they contain would be predominantly HbA and they would not be duly susceptible to haemolysis. Preferential destruc- tion of P-Manukau during erythropoiesis would account for failure to demonstrate it in the father’s peripheral blood.
Although the propositus and his brother have inherited a thalassaemic alpha globin gene from their father, they have received a triplicated alpha gene cluster from their mother giving each of them a normal complement of alpha globin genes. This results in synthesis of normal quantities of alpha globin and there would be no competition between normal and abnormal beta chains. More Hb Manukau would be formed and persist into the mature red cells, resulting in a more severe haemolytic process than that seen in their father.
REFERENCES Brennan. S.O., Shaw. J., Allen, J. &George, P.M. (1992) b141 Leu is
not deleted in the unstable haemoglobin Atlanta-Coventry but
replaced by a novel amino acid of mass 129 daltons. British Journal of Haematology, 81, 99-103.
Brennan. S.O., Wells, R.M.. Smith, H. & Carrell, R.W. (1981) Hemoglobin Brisbane p68 Leu-tHis. A new high oxygen affinity variant. Hemoglobin, 5, 32 5-3 3 5.
Carrell, R. & Kay, R. (1972) A simple method for the detection of unstable haemoglobins. British Journal of Haematology, 23, 61 5.
Casey, R.. Kynoch. P.A.M., Lang. A.. Lehmann, H.. Nozari. G. & Shinton. N.K. (1978) Double heterozygosity for two unstable haemoglobins. Hb Sydney (867 Val-tAla) and Hb Coventry (p141 Leu deleted). British Journal of Haematology. 38, 196-209.
Cathie, I.A.B. (1952) Apparent idiopathic Heinz body anaemia. Great Orniond St Journal. 2. 43.
Dacie, J.V. (1988) The Haemolytic Anaemias, 3rd edn, Vol. 2, Part 2. Churchill Livingstone, Edinburgh.
Dacie. J.V. & Lewis. S.M. (1975) Practical Haematology. 5th edn. Churchill Livingstone, Edinburgh.
Lehmann. H. (1986) Human hemoglobin variants. Hemoglobin: Molecular, Genetic and Clinical Aspecfs (eds Bum and Forget), pp. 380-45 1. Saunders, Philadelphia.
Luan Ng. Li-Injo, Pawson. I.G. & Solai, A. (1985) High frequency of triplicated globin loci and low frequency of thalassaemia in Polynesian Samoans. Human Genetics, 70. 1 16-1 18.
Old, J.M. (1983) Gene analysis. Methods in Haematology. The Thalassaemias. Vol. 6 (ed. by D. J. Weatherall). Churchill Liv- ingstone. Edinburgh.
Perutz. M.F., Kendrew & Watson, H.C. (1965) Structure and function of haemoglobin 11. Journal of Molecular Biology, 13, 669.
Perutz. M.F. & Lehmann, H. (1968) Molecular pathology of human haemoglobin. Nature, 219, 902.
Pisciotha, A.V., Ebbe, S.N. & H i m J.E. (19 59) Clinical and laboratory features of two variants of methemoglobin in disease. Journal of Laboratory and Clinical Medicine, 54, 73.
Podda, A,, Galanello. R.. Maccioni. L.. Melis. M.A., Rosatelli. C.. Persell. L. & Cao, A. (1991) Hemoglobin Cagliari (60 [E4] Val +Glu). A novel unstable thalassemic hemoglobinopathy.
Raik, E.. Hunter, E.G. & Linsay. D.A. (1967) Compensated hereditary haemolytic disease resulting from an unstable haemoglobin fraction. Medical Journal of Australia, i, 955.
Saiki, A.K., Gelfand. D.H., Stoffel, S.. Scharf, S.J., Higuchi, R.. Horn, G.T., Mullis, K.B. & Erlich, H.A. (1988) Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Seience, 239, 487-491.
Steadman. J.H., Yates, A. & Huehns. E.R. (1970) Idiopathic Heinz- body anaemia: Hb Bristol(p67[E11] Val to Asp). British Journal of Haematology, 18, 435.
Trent, R.J., Brock, P.E., Yakas, J., Trent, L.M. & Kronenberg, H. (1 984) Diagnosis of alpha thalassemia in the newborn. Cord blood survey utilizing gene mapping. Pathology, 16, 16-21.
Trent, R.J., Mickelson. K.N.P.. Wilkinson, T. etal (1985) Globin gene rearrangements in Polynesians not associated with malaria. American Journal of Hematology. 18, 431-433.
Blood, 77, 371-375.