
Downregulation of phorbol 12-myristate 13-acetate–induced tumor necrosis factor–alpha and...
8
Downregulation of Phorbol 12-Myristate 13-Acetate-Induced Tumor Necrosis Factor -Alpha and Interleukin- 10 Production and Gene Expression in Human Monocytic Cells by Human Alpha-Fetoprotein WE1 WANG AND ELLIOT hPERT We previously identified a specific receptor for alpha- fetoprotein (AFP) on human monocytes. Although AFP alters many immune cell functions, the effect of AFP on monocyte cytokine production is unknown. Because tumor necrosis factor-alpha (TNF-a) and interleukin-lj3 (IL-lp)are important cytokines in immunoregulation, we investigatedwhetherAFP could modulate TNF-a and IL-lP production in U937, a human monocytic cell line. Our results showed that U937 cells secreted TNF-a and IL-lP in response to either phorbyll2-myristate13-ace- tate (PMA) or WN-y + LPS. In contrast,AFP significantly suppressed PMA-induced TNF-a and nPlP production by U937 cells in a time and dose dependent fashion. Pre- treatment of U937 cells with AFP resulted in maximal inhibition of PMA-stimulated TNF-a and IL-lP produc- tion by 58% and 67%, respectively. AFP also inhibited interferon-y plus lipopolysaccharide (IFN-y + LPS) -in- duced TNF-a and ILlp production. Furthermore,North- ern blot analysis showed that AFP suppressed PMA-me- diated TNF-a and IL-1p messenger RNA (mRNA) expression. PMA-induced prostaglandin E2 (PGE2) pro- duction by U937 cells was enhanced by AFP. Pretreat- ment with indomethacin,a cyclooxygenaseinhibitor,re- versed AFP-inhibited TNF-a production by 78%. Thus, we conclude that AFP downregulates TNF-a and IL-lP production via a PGE2-dependentmechanism. (HEPA- TOLOGY 199% 22:92 1-928.) Alpha-fetoprotein (AFP) is a 70-kd a-globulin glyco- protein normally synthesized and secreted in fetal life by the yolk sac and liver' but is expressed only in low levels in adult sera.' Other than in normal pregnancy, Abbreviations: AFP, alpha-fetoprotein; TNF-a, tumor necrosis factor- alpha; Il-IB, interleukin-lp; PMA, phorbol 12-myristate 13-acetate; LPS, lipo- polysaccharide; ELISA, enzyme-linked immunosorbent assay; PGEz, prosta- glandin EQ; mRNA, messenger Rh'A Ig, immunoglobulin. From the Department of Medicine (Gastroenterology Division), and Lady Davis Institute for Medical Research, Sir Mortimer B. Davis-Jewish General Hospital, and the Department of Medicine, McGill University, Montreal, Que- bec, Canada. Received July 12, 1994; accepted April 20, 1995. Address reprint requests to: Elliot Alpert, MD, Department of Medicine SMBD-Jewish General Hospital, and McGill University, 3755 Cote St, Cath- erine Rd, Montreal, Quebec H3T 1E2, Canada. Copyright 0 1995 by the American Association for the Study of Liver Diseases. 0270-9139/95/2203-0032$3.00/0 the increase of serum AFP level in adult life usually signifies disease. Serum levels of AFP can be signifi- cantly increased in patients with hepatocellular carci- noma~,~ germ cell tumors, and occasionally gastric and pancreatic cancer^.^ Numerous studies have suggested that immunoregula- tion may be one of the biological functions of AFP. AFP has been reported to suppress a variety of immune cell functions, including inhibiting constitutive and lympho- kine-induced macrophage Ia antigen expression, antigen presentation and phagocyto~is:-~ suppressing lympho- cyte proliferation?' inducing suppressor T cells," and decreasing natural killer cell activity." In addition, AFP has been postulated to contribute to the immunologic hyporesponsiveness of the fetus and neonate, as well as protecting the fetus from rejection by the mother.12,13 These studies suggest that AFP may be an immunosup- pressive molecule promoting cancer growth and fetal sur- vival. We have recently identified that human monocytes possess the specific membrane-associated receptors for AFP.14 Monocytedmacrophages have been implicated in playing vital roles in host defense against viral and micro- bial infections and tumor growth. Accumulating evidence has shown that monocytedmacrophages contribute sig- nificantly to immune responses by modulating both cellu- lar and humoral immunity. This immunoregulation may be via the synthesis and release of immunoregulatory cytokines, which have important autocrine and paracrine effects, including the stimulation of further cytokine re- lease and regulation of macrophage and other immune cell function^.'^,^^ Among these secretory products from monocytedmacrophages, tumor necrosis factor-alpha (TNF-a)and interleukin-10 (IL-1p) are the principal me- diators that participate in immune responses and inflam- matory processes.15 Although the effects of AFP on immune cell functions have been intensively studied, it is not known whether AFP could modulate monocyte/macrophage cytokine production. The present study was designed to define the effects of human AFP on TNF-a and IL-10 produc- tion using a human monocytic cell line U937. Our re- sults showed that human AFP downregulated phorbol 12-myristate 13-acetate (PMA)-induced TNF-a and IL-1p production in human monocytic cells. 92 1
Transcript of Downregulation of phorbol 12-myristate 13-acetate–induced tumor necrosis factor–alpha and...

Downregulation of Phorbol 12-Myristate 13-Acetate-Induced Tumor Necrosis Factor -Alpha and Interleukin- 10
Production and Gene Expression in Human Monocytic Cells by Human Alpha-Fetoprotein
WE1 WANG AND ELLIOT h P E R T
We previously identified a specific receptor for alpha- fetoprotein (AFP) on human monocytes. Although AFP alters many immune cell functions, the effect of AFP on monocyte cytokine production is unknown. Because tumor necrosis factor-alpha (TNF-a) and interleukin-lj3 (IL-lp) are important cytokines in immunoregulation, we investigated whether AFP could modulate TNF-a and IL-lP production in U937, a human monocytic cell line. Our results showed that U937 cells secreted TNF-a and IL-lP in response to either phorbyll2-myristate 13-ace- tate (PMA) or WN-y + LPS. In contrast, AFP significantly suppressed PMA-induced TNF-a and nPlP production by U937 cells in a time and dose dependent fashion. Pre- treatment of U937 cells with AFP resulted in maximal inhibition of PMA-stimulated TNF-a and IL-lP produc- tion by 58% and 67%, respectively. AFP also inhibited interferon-y plus lipopolysaccharide (IFN-y + LPS) -in- duced TNF-a and ILlp production. Furthermore, North- ern blot analysis showed that AFP suppressed PMA-me- diated TNF-a and IL-1p messenger RNA (mRNA) expression. PMA-induced prostaglandin E2 (PGE2) pro- duction by U937 cells was enhanced by AFP. Pretreat- ment with indomethacin, a cyclooxygenase inhibitor, re- versed AFP-inhibited TNF-a production by 78%. Thus, we conclude that AFP downregulates TNF-a and IL-lP production via a PGE2-dependent mechanism. (HEPA- TOLOGY 199% 22:92 1-928.)
Alpha-fetoprotein (AFP) is a 70-kd a-globulin glyco- protein normally synthesized and secreted in fetal life by the yolk sac and liver' but is expressed only in low levels in adult sera.' Other than in normal pregnancy,
Abbreviations: AFP, alpha-fetoprotein; TNF-a, tumor necrosis factor- alpha; Il-IB, interleukin-lp; PMA, phorbol 12-myristate 13-acetate; LPS, lipo- polysaccharide; ELISA, enzyme-linked immunosorbent assay; PGEz, prosta- glandin EQ; mRNA, messenger Rh'A Ig, immunoglobulin.
From the Department of Medicine (Gastroenterology Division), and Lady Davis Institute for Medical Research, Sir Mortimer B. Davis-Jewish General Hospital, and the Department of Medicine, McGill University, Montreal, Que- bec, Canada.
Received July 12, 1994; accepted April 20, 1995. Address reprint requests to: Elliot Alpert, MD, Department of Medicine
SMBD-Jewish General Hospital, and McGill University, 3755 Cote St, Cath- erine Rd, Montreal, Quebec H3T 1E2, Canada.
Copyright 0 1995 by the American Association for the Study of Liver Diseases.
0270-9139/95/2203-0032$3.00/0
the increase of serum AFP level in adult life usually signifies disease. Serum levels of AFP can be signifi- cantly increased in patients with hepatocellular carci- n o m a ~ , ~ germ cell tumors, and occasionally gastric and pancreatic cancer^.^
Numerous studies have suggested that immunoregula- tion may be one of the biological functions of AFP. AFP has been reported to suppress a variety of immune cell functions, including inhibiting constitutive and lympho- kine-induced macrophage Ia antigen expression, antigen presentation and phagocyto~is:-~ suppressing lympho- cyte proliferation?' inducing suppressor T cells," and decreasing natural killer cell activity." In addition, AFP has been postulated to contribute to the immunologic hyporesponsiveness of the fetus and neonate, as well as protecting the fetus from rejection by the mother.12,13 These studies suggest that AFP may be an immunosup- pressive molecule promoting cancer growth and fetal sur- vival. We have recently identified that human monocytes possess the specific membrane-associated receptors for AFP.14 Monocytedmacrophages have been implicated in playing vital roles in host defense against viral and micro- bial infections and tumor growth. Accumulating evidence has shown that monocytedmacrophages contribute sig- nificantly to immune responses by modulating both cellu- lar and humoral immunity. This immunoregulation may be via the synthesis and release of immunoregulatory cytokines, which have important autocrine and paracrine effects, including the stimulation of further cytokine re- lease and regulation of macrophage and other immune cell function^.'^,^^ Among these secretory products from monocytedmacrophages, tumor necrosis factor-alpha (TNF-a) and interleukin-10 (IL-1p) are the principal me- diators that participate in immune responses and inflam- matory processes.15
Although the effects of AFP on immune cell functions have been intensively studied, it is not known whether AFP could modulate monocyte/macrophage cytokine production. The present study was designed to define the effects of human AFP on TNF-a and IL-10 produc- tion using a human monocytic cell line U937. Our re- sults showed that human AFP downregulated phorbol 12-myristate 13-acetate (PMA)-induced TNF-a and IL-1p production in human monocytic cells.
92 1

922 WANG AND ALPERT HEPATOLOGY September 1995
MATERIALS AND METHODS
Reagents. RPMI 1640 with L-glutamine and low-endotoxin fetal calf serum were purchased from Gibco/BRL Life Techno- logies (Burlington, Ontario, Canada). PMA, LPS, and indo- methacin were obtained from Sigma Chemical Co. (St. Louis, MO). Recombinant human IFN--y was purchased from En- zyme (Cambridge, MA). [a-"'P dCTP1 was obtained from ICN Biomedicals (St. Laurent, Quebec, Canada). All other chemi- cals were of reagent grade.
Preparation of U P . Human AFP was purified as described previously"' from human amniotic fluid by immunoaffinity column chromatography and Sephadex G200 (Pharmacia LKB Biotechnology, Piscataway, NJ) column separation. Pu- rified AFP showed a single band on sodium dodecylsulfate- polyacrylamide gel electrophoresis, a single peak on p Bonda- pak C18 (Millipore Corp., Milford, MA) high-pressure liquid chromatography column, and no reaction to anti-human se- rum. At the concentration of 10 pg/mL, AFP contained unde- tectable (<0.01 endotoxin U/mL) LPS contamination by Lim- ulus Amebocyte Lysate Assay with kit from Sigma Co.
Cell Culture and Induction of Cytokine Release. U937 hu- man monocytic cells (American Type Culture Collections, Rockville, MD) were grown in complete 1640 medium supple- mented with penicillin (100 U/mL), streptomycin (100 U/mL), HEPES (10 mmol/L), and 10% heat-inactivated low-endo- toxin fetal calf serum a t a 37°C humidified atmosphere with 5% carbon dioxide. TNF-(1 and IL-lD production by U937 cells was induced by incubation o f cells with PMA or IFN--y and lipopolysaccharide (LPS). To determine the effects of AFP on PMA-induced 'I'NF-a and IL-1p production, U937 cells were pretreated with AFP in the concentration as indi- cated in the figures for 4 hours and then incubated with PMA (10 ng/mL) for 2, 24, 48, and 72 hours. Cell-free conditioned supernatants were collected and tested for TNF-a and IL-lP activities either fresh or after storage at -80°C.
Measurement of TNF-a and ZL-ZP Production. TNF-a and IL-1p levels in the conditioned medium were determined by commercially available enzyme-linked immunosorbent assay (EIJSA) kits (R & D systems, Minneapolis, MN). Briefly, the frozen conditioned cell supernatants were thawed at room temperature and added to wells of rigid flat-bottom microtiter plates coated with a murine monoclonal antibody against hu- man TNF-a or human IL-lP. After incubation and washing of the wells, horseradish peroxidase-conjugated anti-TNF- LY antibody or anti-IL-10 antibody was added to the test wells. After a second incubation, the excess horseradish per- oxidase-conjugated antibody was removed by washing. The horseradish peroxidase substrate was then added, and the color intensity measured with an ELISA reader.
Radioimmunoassay for Prostaglandin El. Cell-free condi- tioned media were extracted by passing through C2 ethyl Amprep columns ( h e r s h a m Canada Ltd., Oakville Ontario, Canada) following the manufacturer's protocol. The extracts were evaporated to dryness under a n N2 stream. Extracts were tested for prostaglandin E2 (PGE2) with [3H] radioim- munoassay kit (Advanced Magnetics Inc., Cambridge, MA).
RNA Extraction and Northern Blot Analysis. After incuba- tion, U937 cells were washed twice with serum-free medium and total RNA was extracted with RNAzol (CinnaA3iotecx Laboratories International Inc., Friendswood, TX), which is a modification of the guanidinium-phenol chloroform method." Total RNA samples (20 pg) were denatured for 1 hour at 50°C in the presence of'glyoxal, and 1 pg of ethidium bromide was added to each sample. After electrophoresis in a 1% agarose gel, RNA was blotted by capillary transfer to Hybond-N mem-
brane (Amersham Canada Ltd.). The membrane bolts were prehybridized for 5 hours at 42°C in 5 0 8 formamide, 0.1% sodium dodecylsulfate, 5~ SSPE ( l x SSPE is 0.18 m o m of NaCl, 10 mmol/L of NaP04 , and 1 mmol/L of EDTA [pH 7.711, 50x Denhardt's (1% polyvinylpyrodilone and 1% boric acid solution), and 0.25 mg/mL denatured salmon sperm DNA. Hybridization was preformed at 42°C for 12 to 18 hours with probes purified from agarose and nick-translated in the pres- ence of 125 pCi of "P-dCTP (ICN Biomedicals). After hybrid- ization, membranes were washed in 0.5x SSC ( l x SSC is 0.15 mol/L of NaCl plus 0.015 mol/L of sodium citrate) at 20°C for 20 minutes, in 0 . 5 ~ SSC a t 55°C for 30 minutes, and 0 . 1 ~ SSC a t 65°C for 15 minutes. Autoradiography was performed at -80°C with an intensifying screen, using Kodak XAR-5 films (Picker International Canada Inc., St. Laurent, Quebec, Canada). Bacillus suhtilis SMS 118 containing plas- mid with human IL-10 insert, Escherichia coli MM294 con- taining plasmid with human TNF-a insert, and E coli XL1- Blue containing phagemid with human P-actin insert were obtained from American Type Culture Collections. TNF-a in- sert (1.1 kb) was removed with EcoRI. IL-lP (0.7 kb) was excised with EcoRI + PstI double digest. Actin (1.1 kb) insert was removed with EcoRI. Blots were reprobed with actin as an internal control for the quantity of total messenger RNA (mRNA) in each line. Densitometry analysis was performed with a Laster densitometer (UltrosScan XL; LKB, Phamacia, Sweden). The quantification of mRNA levels was obtained by calculating the ratio between measured optical density of TNF-a and IL-IP and actin mRNA.
Statistical Analysis. Data were analyzed by the Student's t-test or one-way ANOVA. A value of P < .05 is regarded as statistically significant.
RESULTS Induction of TNF-a and IL-1P Production by U937 Cells
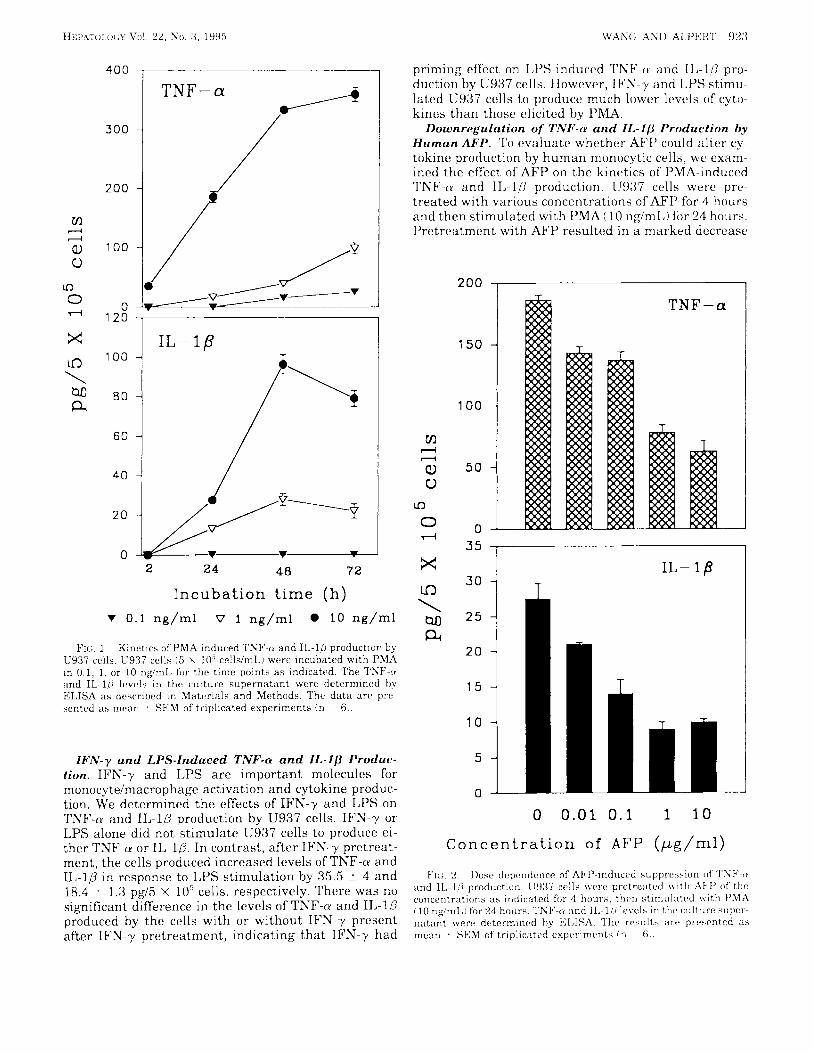
PMA-Induced TNF-a and IL-lP Production. The dif- ferent lines of U937 cells exhibited slightly different properties with respect to induction of HLA class I1 by IFN-y or induction of IL-1 by LPS.18 First, we deter- mined the induction of TNF-a and IL-1P production by U937 cells with PMA. Unstimulated U937 cells did not release detectable levels of TNF-a and IL-10. In con- trast, stimulation of the cells with PMA resulted in a rapid increase in the levels of both cytokines. The kinet- ics of induction of TNF-a and IL-10 was shown in Fig. 1. TNF-a level was detectable at 2 hours after addition of PMA (10 ng/mL), and a consistent increase was ob- served that reached a plateau at 72 hours. PMA (1 ng/ mL) induced a low detectable level of TNF-a at 24 hours after stimulation, whereas 0.1 ng/mL of PMA stimu- lated TNF-a secretion at 48 hours. These results showed a time- and dose-dependent fashion of PMA- induced TNF-cr production.
On the other hand, a dose- and time-response pattern was also observed in the induction of IL-10 by PMA. U937 cells did not release IL-1P when incubated with 0.1 ng/mL of PMA. A detectable level of IL-10 was mea- sured at 24 hours, and a maximal release was found at 48 hours and decreased after 72 hours after addition of PMA (1 and 10 ng/mL). However, the level of IL-10 produced by U937 cells in response to PMA was much less than that of TNF-a.
400
3 0 0
2 0 0
100
0 120
2 24 40 72
Incubat ion t ime (h) v 0.1 ng/ml v 1 ng/ml 0 10 ng/ml
FIC:. 1. Kinetics of PMA-induced Th-F-rr and II,-l[I production by C937 cells. U937 cells (5 x 10‘ cellsimL) werc incubated with I’MA in 0.1, 1. or 10 ng/mI, for the time points a s indicated. The TNF-rr and IL-lLI l ~ v e l s in t h r culture supernatant were detrrnmincd hy ELISA as described in Matvrials and Methods. The da ta arc’ pre- sented as mPan . SEM of triplicated experiments (11 61.
IFN-y and LPS-Induced TNF-a and IL-lS Produc- tion. IFN-y and LPS are important molecules for monocyte/macrophage activation and cytokine produc- tion. We determined the effects of IFN-y and LPS on TNF-a and I L l P production by U937 cells. IFN-y or LPS alone did not stimulate U937 cells to produce ei- ther TNF-a or IL-10. In contrast, after IFN-y pretreat- ment, the cells produced increased levels of TNF-cr and I L l p in response to LPS stimulation by 35.5 ? 4 and 18.4 ‘ 1.3 pgi5 x 10’’ cells. respectively. There was no significant difference in the levels of TNF-cu and I L l f i produced by the cells with o r without IFN-y present after IFN-y pretreatment, indicating tha t IFN-y had
priming effect on LI’S-inducctd ‘I”I%u and 11,- 1/1 pro- duction by U937 cells. IIowever, I F S - y and LPS stimu- lated C937 cells t o produce much lower levels of cyto- kines than those elicited by I’MA.
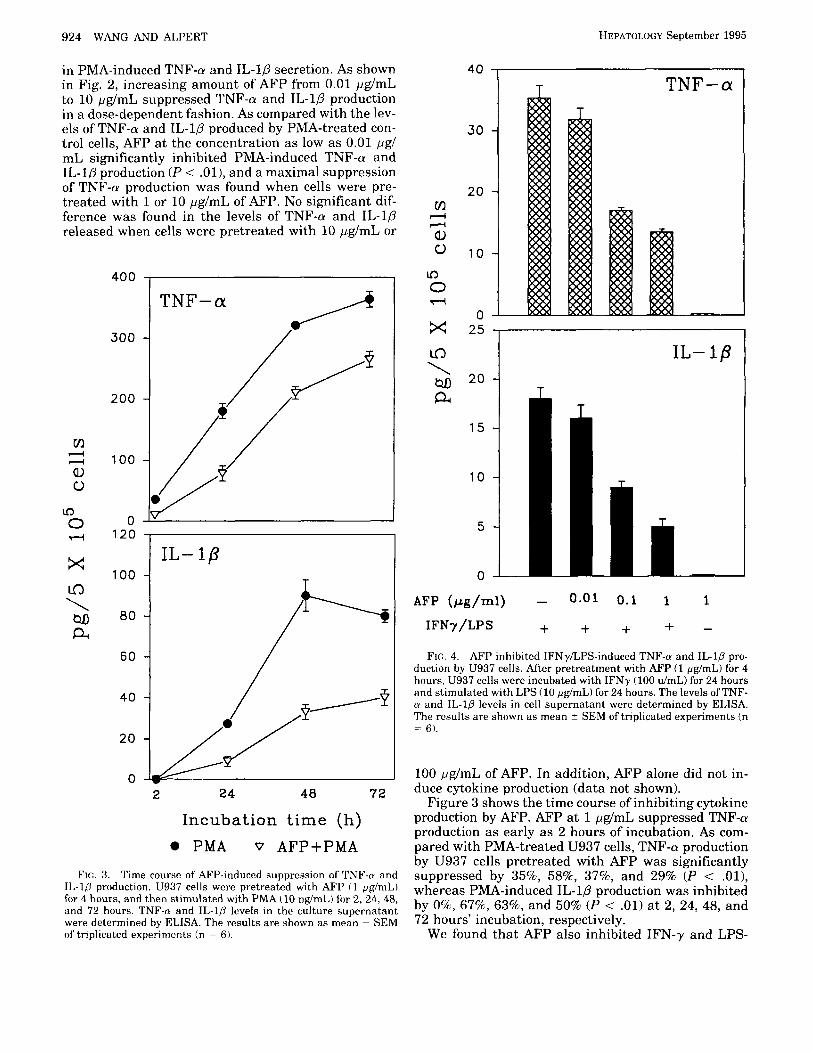
Downregulation of TNF-LU and I L - l s Production by Human AFP. To evaluate whether MI’ could alter cy- tokine production by human monocytic cells, we exam- ined the effect of‘ AFP on the kinetics of‘ I’MA-induced ‘I’NF-tu and II,-lfi production. U937 cells were pre- treated with various concentrations of AFI’ for 4 hours and then stimulated with PMA (10 ng/mL) for 24 hours. Pretreatment with AFP resulted in a marked decrease
2 0 0
150
100
5 0
0
T NF-a
IL- 1p 3 0
2 5 - 2 0 -
1 5 -
10 -
5 -
0
0 0.01 0.1 1 10
Concent ra t ion of AFP ( p g / m l )
924 WANG AND ALPERT
30 -
20 -
10 -
0 -
HEPATOLOGY September 1995
15 -
10 -
5 -
0
in PMA-induced TNF-n and IL-10 secretion. As shown in Fig. 2, increasing amount of AFP from 0.01 pg/mL to 10 pg/mL suppressed TNF-a and IL-10 production in a dose-dependent fashion. As compared with the lev- els of TNF-a and IL-10 produced by PMA-treated con- trol cells, AFP a t the concentration as low as 0.01 pg/ mL significantly inhibited PMA-induced TNF-a and IL-10 production ( P < . O l ) , and a maximal suppression of TNF-n production was found when cells were pre- treated with 1 or 10 pg/mL of AFP. No significant dif- ference was found in the levels of TNF-a and IL-1p released when cells were pretreated with 10 pg/mL or
400
300
200
100
0 120
100
80
60
40
20
0
~
IL- 18 T
2 24 40 7 2
Incubation time (h) 0 PMA v AFP+PMA
FIG. 3. Time course of' AFP-induced suppression of TNF-n and Il,-lfl production. U937 cells were pretreated with AFP ( 1 pg/mL) for 4 hours, and then stimulated with PMA (10 ng/mL) for 2, 24, 48, and 72 hours. TNF-n and IL-10 lcvels in the culture supernatant were determined by ELISA. The results a re shown as mean -C SEM of' triplicated experiments (n = 6).
40 T TNF-cx
25 , I IL- 1p
20-i T
AFP (pg/ml) - 0.01 0.1 1 1
IFNy/LPS + + + + - FIG. 4. AFP inhibited IFNyLPS-induced TNF-u and IL-1fl pro-
duction by U937 cells. After pretreatment with AFP (1 pg/mL) for 4 hours, U937 cells were incubated with IFNy (100 d m L ) for 24 hours and stimulated with LPS (10 pg/mL) for 24 hours. The levels ofTNF- (r and IL-1fl levels in cell supernatant were determined by ELISA. The results are shown as mean z SEM of triplicated experiments (n = 6) .
100 pg/mL of AFP. In addition, AFP alone did not in- duce cytokine production (data not shown).
Figure 3 shows the time course of inhibiting cytokine production by AFP. AFP at 1 pg/mL suppressed TNF-a production as early as 2 hours of incubation. As com- pared with PMA-treated U937 cells, TNF-a production by U937 cells pretreated with AFP was significantly suppressed by 35%, 58%, 3796, and 29% (P < . O l ) , whereas PMA-induced IL-10 production was inhibited by O%, 67%, 63%, and 50% ( P < .01) at 2, 24, 48, and 72 hours' incubation, respectively.
We found that AFP also inhibited IFN-y and LPS-
HEPATOLOGY Vol. 22, No. 3, 1995
ki 100 - 0 ..-I 8 0 -
60 - Q 40 - 20 -
0
-c,
v1
WANG AND ALPERT 925
I L - l P
induced TNF-a and IL-1P production (Fig. 4). However, differently from PMA, AFP at the concentration of 0.01 pg/mL did not suppress IFN-y and LPS-induced TNF-a and IL-1P production (P > .05), whereas AFP at 0.1 and 1 pg/mL significantly decreased TNF-a production by 52% and 62% (P < .Ol), respectively. IL-1P produc- tion was also inhibited by 50% and 72% (P < . O l ) , respectively, as compared with the levels of TNF-a and IL-10 produced by PMA-treated cells.
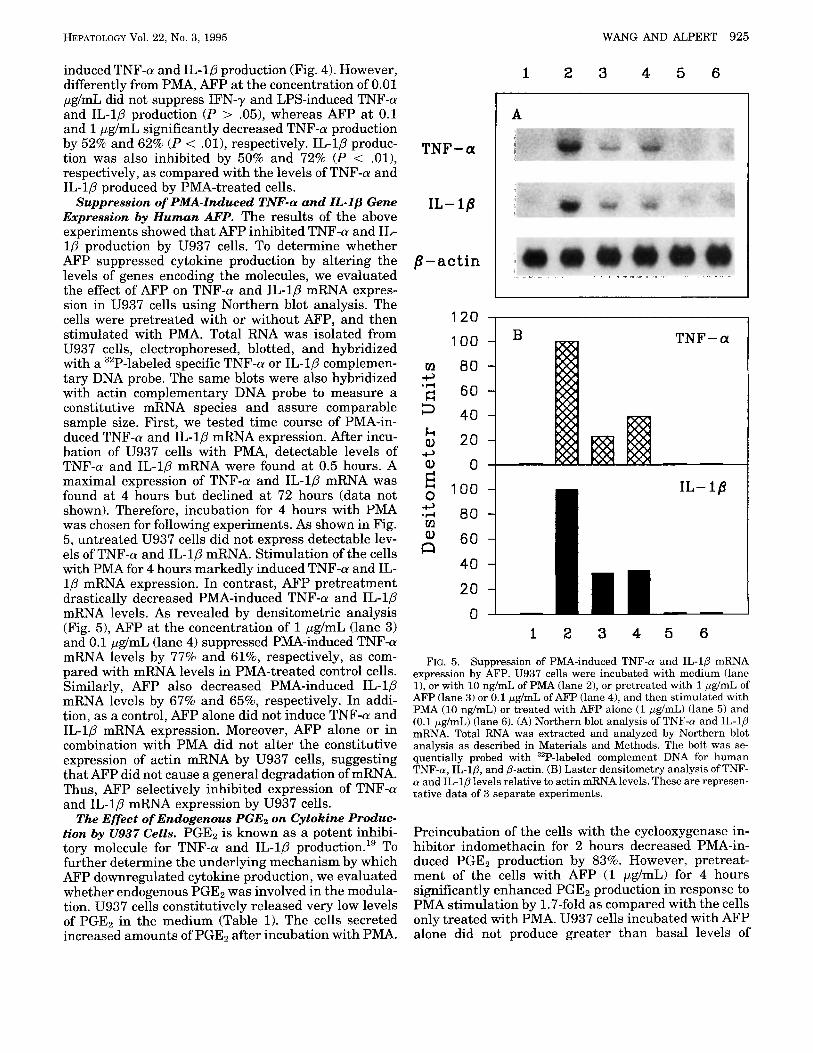
Suppression of PMA-Induced TNF-CY and IL-lp Gene Expression by Human AFP. The results of the above experiments showed that AFP inhibited TNF-a and IL- 1P production by U937 cells. To determine whether AFP suppressed cytokine production by altering the levels of genes encoding the molecules, we evaluated the effect of AFP on TNF-a and IL-10 mRNA expres- sion in U937 cells using Northern blot analysis. The cells were pretreated with or without AFP, and then stimulated with PMA. Total RNA was isolated from U937 cells, electrophoresed, blotted, and hybridized with a 32P-labeled specific TNF-a or IL-1P complemen- tary DNA probe. The same blots were also hybridized with actin complementary DNA probe to measure a constitutive mRNA species and assure comparable sample size. First, we tested time course of PMA-in- duced TNF-a and IL-1P mRNA expression. After incu- bation of U937 cells with PMA, detectable levels of TNF-a and IL-1p mRNA were found at 0.5 hours. A maximal expression of TNF-a and IL-1P mRNA was found at 4 hours but declined at 72 hours (data not shown). Therefore, incubation for 4 hours with PMA was chosen for following experiments. As shown in Fig. 5 , untreated U937 cells did not express detectable lev- els of TNF-a and IL-1p mRNA. Stimulation of the cells with PMA for 4 hours markedly induced TNF-a and IL- l p mRNA expression. In contrast, AFP pretreatment drastically decreased PMA-induced TNF-a and IL-1p mRNA levels. As revealed by densitometric analysis (Fig. 51, AFP at the concentration of 1 pg/mL (lane 3) and 0.1 pg/mL (lane 4) suppressed PMA-induced TNF-a mRNA levels by 77% and 61%, respectively, as com- pared with mRNA levels in PMA-treated control cells. Similarly, AFP also decreased PMA-induced IL-1P mRNA levels by 67% and 65%, respectively. In addi- tion, as a control, AFP alone did not induce TNF-a and IL-1p mRNA expression. Moreover, AFP alone or in combination with PMA did not alter the constitutive expression of actin mRNA by U937 cells, suggesting that AFP did not cause a general degradation of mRNA. Thus, AFP selectively inhibited expression of TNF-a and IL-1P mRNA expression by U937 cells.
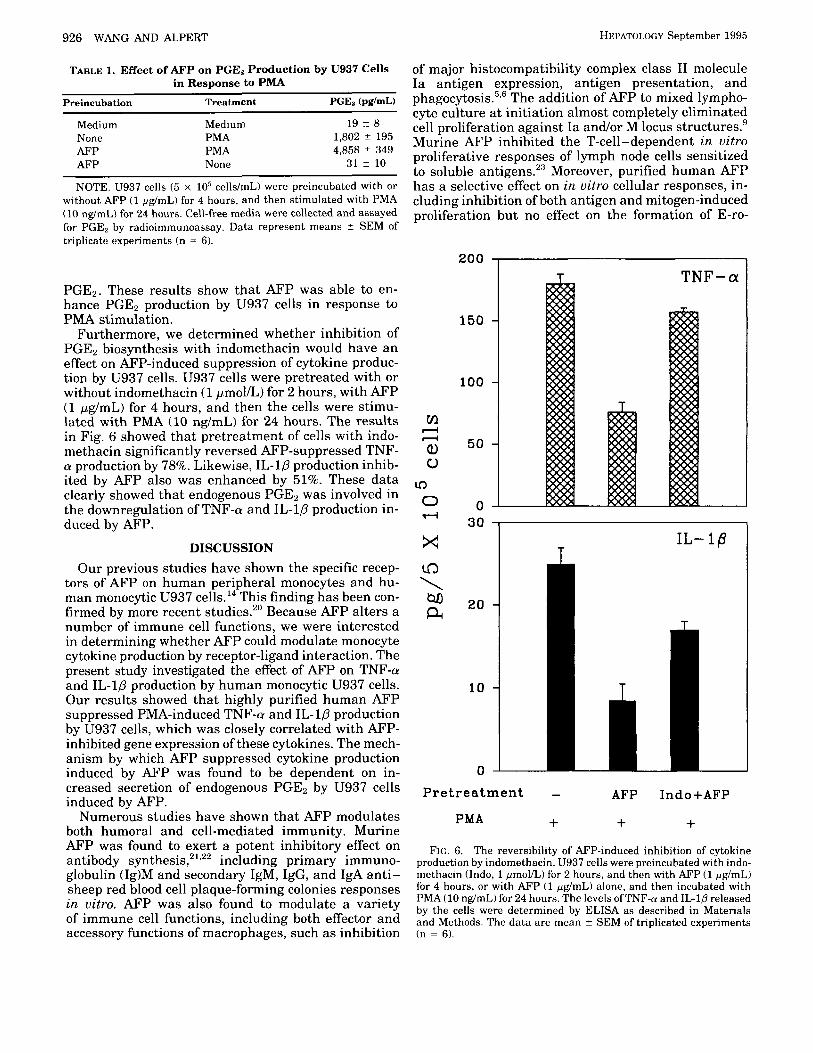
The Effect of Endogenous PGEp on Cytokine Produc- tion by U937 Cells. PGEz is known as a potent inhibi- tory molecule for TNF-a and IL-1p production." To further determine the underlying mechanism by which AFP downregulated cytokine production, we evaluated whether endogenous PGEz was involved in the modula- tion. U937 cells constitutively released very low levels of PGEz in the medium (Table 1). The cells secreted increased amounts of PGEz after incubation with PMA.
1 2 3 4 5 6
I A TNF-a
IL- 18
,8 - actin
120 I
100
40 20
0
B T N F - a
d I
1 2 3 4 5 6
FIG. 5. Suppression of PMA-induced TNF-a and IL-lP mRNA expression by AFP. U937 cells were incubated with medium (lane l), or with 10 ng/mL of PMA (lane 2), or pretreated with 1 pg/mL of AFP (lane 3) or 0.1 pg/mL of AFP (lane 4), and then stimulated with PMA (10 ng/mL) or treated with AFP alone (1 pg/mL) (lane 5) and (0.1 pg/mL) (lane 6). (A) Northern blot analysis of TNF-a and IL-1p mRNA. Total RNA was extracted and analyzed by Northern blot analysis as described in Materials and Methods. The bolt was se- quentially probed with 32P-labeled complement DNA for human TNF-a, IL-lD, and P-actin. (B) Laster densitometry analysis of TNF- a and IL-1p levels relative to actin mRNA levels. These are represen- tative data of 3 separate experiments.
Preincubation of the cells with the cyclooxygenase in- hibitor indomethacin for 2 hours decreased PMA-in- duced PGEz production by 83%. However, pretreat- ment of the cells with AFP (1 pg/mL) for 4 hours significantly enhanced PGE2 production in response to PMA stimulation by 1.7-fold as compared with the cells only treated with PMA. U937 cells incubated with AFP alone did not produce greater than basal levels of
926 WANG AND ALPERT
150 -
100 -
50 -
0 -
HEIWTOLOGY September 1995
20 -
10 -
0 -
TABLE 1. Effect of AFP on PGE2 Production by U937 Cells in Response to PMA
Preincubation Treatment PGEa (pg/mW
Medium Medium 19 -c 8 None PMA 1,802 2 195 AFP PMA 4,858 z 349 AFP None 31 z 10
NOTE. U937 cells (5 x lo5 cells/mL) were preincubated with or without AFP (1 pg/mL) for 4 hours, and then stimulated with PMA (10 ng/mL) for 24 hours. Cell-free media were collected and assayed for PGE, by radioimmunoassay. Data represent means z SEM of triplicate experiments (n = 6).
PGE2. These results show that AFP was able to en- hance PGE2 production by U937 cells in response to PMA stimulation.
Furthermore, we determined whether inhibition of PGEz biosynthesis with indomethacin would have an effect on AFP-induced suppression of cytokine produc- tion by U937 cells. U937 cells were pretreated with or without indomethacin (1 pmol/L) for 2 hours, with AFP (1 pg/mL) for 4 hours, and then the cells were stimu- lated with PMA (10 ng/mL) for 24 hours. The results in Fig. 6 showed that pretreatment of cells with indo- methacin significantly reversed AFP-suppressed TNF- a production by 78%. Likewise, IL-10 production inhib- ited by AFP also was enhanced by 51%. These data clearly showed that endogenous PGEz was involved in the downregulation of TNF-a and IL-1P production in- duced by AFP.
DISCUSSION Our previous studies have shown the specific recep-
tors of AFP on human peripheral monocytes and hu- man monocytic U937 cells.14 This finding has been con- firmed by more recent studies.2" Because AFP alters a number of immune cell functions, we were interested in determining whether AFP could modulate monocyte cytokine production by receptor-ligand interaction. The present study investigated the effect of AFP on TNF-u and IL-1P production by human monocytic U937 cells. Our results showed that highly purified human AFP suppressed PMA-induced TNF-a and IL- l p production by U937 cells, which was closely correlated with AFP- inhibited gene expression of these cytokines. The mech- anism by which AFP suppressed cytokine production induced by AFP was found to be dependent on in- creased secretion of endogenous PGE2 by U937 cells induced by AFP.
Numerous studies have shown that AFP modulates both humoral and cell-mediated immunity. Murine AFP was found to exert a potent inhibitory effect on antibody synthesis,2'-22 including primary immuno- globulin (1g)M and secondary IgM, IgG, and IgA anti- sheep red blood cell plaque-forming colonies responses in uitro. AFP was also found to modulate a variety of immune cell functions, including both effector and accessory functions of macrophages, such as inhibition
of major histocompatibility complex class I1 molecule Ia antigen expression, antigen presentation, and phagocytosis."' The addition of AFP to mixed lympho- cyte culture at initiation almost completely eliminated cell proliferation against Ia andor M locus structures.9 Murine AFP inhibited the T-cell-dependent in uitro proliferative responses of lymph node cells sensitized to soluble antigens."$ Moreover, purified human AFP has a selective effect on in uitro ccllular responses, in- cluding inhibition of both antigen and mitogen-induced proliferation but no effect on the formation of E-ro-
200 , r TNF-a
30 I
T IL-16
T
T
LL Pretreatment - AFP Indo+AFP
+ + + PMA
FIG. 6. The reversibility of AFP-induced inhibition of cytokine production by indomethacin. U937 cells were preincubated with indo- methacin (Indo, 1 pmollL) for 2 hours, and then with AFP (1 pg/mL) for 4 hours, or with AFP (1 pg/mL) alone, and then incubated with PMA (10 ng/mL) for 24 hours. The levels ofTNF-cr and IL-10 released by the cells were determined by ELISA as described in Materials and Methods. The da ta are mean z SEM of triplicated experiments (n = 6).

HEPATOLOGY Vol. 22, No. 3, 1995 WANG AND ALPERT 927
settes by T cells, antigen-induced MIF production, or concanavalin A-induced lymphocyte cytotoxicity.8 In addition, Gershwin et a124 showed that AFP adminis- tered in uiuo accelerated tumor growth, increased mor- tality, and lowered the threshold dose necessary to in- duce tumors by Maloney sarcoma virus, implying that AFP may function as an immunoregulatory agent in uiuo during oncogenetic development. Thus, it appears that AFP is an important immunomodulating mole- cule.
Although the earlier studies cited previously showed that immunosuppression is a property of AFP, the re- sults of the present investigation are the first to show that human AFP downregulates human monocytic cell TNF-a and IL-1p production and gene expression. TNF-a and IL-1p play important roles in inflammation and immunoregulation and share a large number of biological properties. Both directly mediate multiple aspects of inflammation and immune responses, in- cluding fever,25 hepatic acute phase protein synthe- sis,26,27 endothelial cell procoagulant activity,28 and the activation of T lymphocytes and antitumor a c t i ~ i t y . ~ ~ ~ ~ ~ TNF-a is a potent immunoregulator influencing growth, differentiation, and function of a broad range of cells. TNF-a has multiple stimulatory activities in activated T cells, including increasing the proliferation response to antigen, increasing IL-1 receptor expres- sion and response to IL-2 as a proliferative stimulus, and in the induction of IFN-y p r o d ~ c t i o n . ~ ~ TNF-a also has stimulatory effect on human B lymphocyte prolifer- ation and Ig secretion.32 Alone or synergistically with IFN-y, TNF-a exerts a pleiotropic effect on macrophage activity by stimulating both accessory and effector functions, such as stimulating IL-1 production, increas- ing the expression of Ia antigen on a murine macro- phage cell line,33 and enhancing macrophage antitumor cytotoxic In addition, TNF-a is a potent me- diator for macrophage cytotoxicity. Depending on the type of target cell and on the presence of metabolic inhibitors, TNF-a may mediate apoptotic or necrotic cell l y ~ i s . ~ ~ As TNF-a and IL-1p are critical mediators in immune responses, our study suggests that down- regulation of cytokine production by AFP may contrib- ute to immunosuppression induced by AFP.
PGE2 is a well-known immunomodulator that has multiple effects on the immune system.36 Recent stud- ies have indicated that PGE2 inhibits macrophage-de- rived TNF-a and IL-1 p r o d u ~ t i o n . ~ ~ , ~ ~ , ~ ~ It h as been found that human monocytic U937 cells are able to metabolize arachidonic acid and release PGE2 upon ti mu la ti on.^' PGE2 posttranscriptionally inhibited IL-1 release by U937 cells by increasing intracellular cyclic adenosine monopho~phate.~' The present study showed that AFP enhanced PGE2 secretion by U937 cells in response to PMA stimulation. Pretreatment of U937 cells with indomethacin significantly reversed AFP-suppressed TNF-(r and IL-lP production, sug- gesting that PGE2 was involved in the downregulation of cytokine production induced by AFP.
U937 cells differentiate along the monocyte lineage
after treatment with different agents, including phor- bol esters,41 l ~ m p h o k i n e , ~ ~ I F N - Y , ~ ~ and vitamin D de- r i v a t i v e ~ . ~ ~ Incubation of U937 cells with phorbol ester PMA results in cessation of proliferation, significant alteration in the morphology and the functions of the cells characterized by adherence, the presence of Fc receptors, nonspecific esterase staining, ability to phagocytize latex beads, secreting lysozyme, antibody- dependent and nonspecific tumor killing activity, and expression of pro to oncogene^.^^,^^^^^ Our results showed that PMA induced U937 cells to express TNF-a and IL-1p gene expression and cytokine production, sug- gesting that the differentiation of U937 cells was asso- ciated with the alteration of cytokine production. Addi- tionally, IFN-y, a major macrophage-activating factor, enhances oxidative metabolism and antimicrobial ac- tivity of m o n ~ c y t e s . ~ ~ Recombinant human IFN-y en- ables U937 cells to be stimulated for oxygen produc- t i ~ n . ~ ~ In our experiments, IFN-y alone did not stimulate U937 cells to produce TNF-a and IL-1p. However, IFN-y was able to prime U937 cells to pro- duce cytokines in response to LPS. It is well known that LPS induces cytokine production including TNF-a and IL-10. However, our results showed that LPS was unable to stimulate cytokine production by U937 cells, suggesting that U937 cells were distinct from macro- phage at the stage of maturation or differentiation.
In conclusion, the present study provides the evi- dence that human AFP downregulates PMA-induced TNF-a and IL-1p production and gene expression by a PGE2-dependent mechanism. Our ongoing studies are aimed at determining the effect of AFP on intracellular signal transduction pathways.
Acknowledgment: We thank Lina Ricci for assis- tance in the preparation of the manuscript.
REFERENCES 1. Alpert E. Human alpha-fetoprotein (AFP). In: Crandall B, Brazio
RM, eds. UCLA forum in medical sciences. New York: Academic,
2. Ball D, Rose E, Alpert E. Alpha-fetoprotein levels in normal adults. The American Journal of the Medical Sciences.
3. Adinolfi A, Adinolfi M, Lessof MH. Alpha-fetoprotein during de- velopment and in disease. J. Med. Genet. 1975; 12:138-151.
4. Alpert E. Alpha-1-fetoprotein. In: Sherlock P, Zamcheck N, eds. Clinics in Gastroenterology. London: Saunders, 1976539-644.
5. Lu CY, Changelian PS, Unanue ER. a-fetoprotein inhibits mac- rophage expression of Ia antigens. J Immunol 1984; 132:1722- 1727.
6. Crainie MA, Semeluk K, Lee C, Wegmann T. Regulation of con- stitutive and lymphokine-induced Ia expression by murine a- fetoprotein. Cell Immunol 1989; 118:41-52.
7. Olinescu A, Laky M, Popescu DE, Dumitrescu A, Ganea D. The effect of alpha-fetoprotein on the immune response. 111. Diminu- tion of the phagocytosis capacity of macrophages cultured in vitro in the presence of mouse amniotic fluid or alpha-fetopro- tein. Scand J Immunol 1978;8:397-401.
8. Alpert E, Dienstag JL, Sepersky S, Littman B, Rocklin R. Immu- nosuppressive characteristics of human AFP: effect on tests of cell mediated immunity and induction of human suppressor cells. Immunol Commun 1978;7:163-185.
9. Peck AB, Murgita RA, Wigzell H. Cellular and genetic restric- tions in the immunoregulatory activity of alpha-fetoprotein. I.
1978119-26.
1992; 303: 157-159.

928 WANG AND ALPERT HEPATOLO(:Y Septcmber 1995
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
Selective inhibition of anti-Ia-associated proliferative reactions. J Exp Med 1978; 147:667-683. Murgita RA. Goidl EA, Kontiainen S, Wigzell H. u-Fetoprotein induces suppressor T cells in oitro. Nature 1977;267:257-259. Cohen HL, O m A, Gronvik KO, Gidlund M, Wigzell II , Murgita RA. Suppression by alpha-fetoprotein of murine natural killer cell activity stimulated in vitro and in oiuo by interferon and interleukin 2. Scand tJ Immunol 1986;23:211-223. Yachnin S. Demonstration of the inhibitory effect of human alpha-fetoprotein on in citro transformation of human lympho- cytes. Proc Natl Acad Sci U S A 1976;73:2857-2861. Van Oers NSC, Cohen BL, Murgita KA. Isolation and character- ization of a distinct immunoregulatory isoform of alpha-fetopro- tein produccd by the normal fetus. J Exp Med 1989;170:811- 825. Suzuki Y, Zeng CQY, Alpert E. Isolation and partial character- ization of‘ a specific alpha-fetoprotein receptor in human mono- cytes. J Clin Invest 1992;90:1530-1536. Rurchctt SK, Weaver WM, Westall JA, Larsen A, Kronheim S, Wilson CH. Regulation of tumor necrosis factor/cachcctin and II,-l sccretion in human mononuclear phagocytes. J Immunol
Trede NS, Geha RS, Chatila T. Transcriptional activation of IL- 10 and tumor necrosis factor-n genes by MHC class I1 ligands. ?J Immunol 1.991; 146:2:310-2315. Chomczynski P, Sacchi N. Single-step methods of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 1987; 162:156-159. Fenton MJ, Vermeulen WM, Clark BL), Webb AC, Auron PE. Human pro-1L-10 gene expression in monocytic cells is regulated by two distinct pathways. J Immunol 1988; 140:2267-2273. Kunkel SL, Scales WF, Spcngler K, Spengler M, Spengler M, Larrick J . Dynamics and regulation of murine tumor nccrosis factor-a (TNF), interleukin-ln ~ 1 L - l ~ ) . and intcrleukin-Lj (IL- 10) gene expression by arachidonate metabolites. In: Pawanda MC, ed. Monokines and other non-lymphocytic cytokincs. New York: Liss, 1988:61-72. Esteban C, Trojan J, Macho A, Mishal A, Lafarge-Frayssinc CH, Uriel J. Activation of an alpha-fetoprotcinheceptor pathwmy in human normal and malignant peripheral blood mononuclear cells. Leukemia 1993;7:1807-1816. Murgita RA, Tomasi TB. Suppression of the immune response by alpha-fetoprotein. I. The effect of mouse alpha-Vetoprotein on the primary and secondary antibody response. J Exp Med
Zimmerman EF, Voorting-Hawking M, Michael JG. Immuno- suppression by mouse sialylatcd alpha-fetoprotein. Nature 1977; 265:354. Suzuki K, Tomasi TR, J r . Inhibition of antigen-induced lymph node cell proliferation by murinc amniotic fluid and its compo- nents. Immunology 1979;38:539-545. Gershwin ME, Castles J J , Ahmed A, Makishima R. The influ- ence of tr-fetoprotcin on Maloney sarcoma virus oncogenesis: evi- dence for generation of antigen nonspecific supprctssor T cells. J Immunol 1978; 121 2292-2298. Dinarello CA, Cannon JG, WolK SM, Rernheim HA, Beutler R , Ccrami A, Figari IS, et al. Tumor nccrosis factor (cachectin) is an endogenous pyrogen and induces production of interleukin 1.
Ramadori G, Sipe JL), Dinarello CA, Mizel SR, Colten H. Prc- translational modulation of acute phase hepatic protein synthc- sis by murine recornhinant intcrleukin-1 (IL-1) and purified hu- man 1L-1. J Exp Med 1985; 162:930-942. Perimutter L)H, Dinarello CA, I’unsal PH, Colter HK. Cachectid tumor necrosis factor r e p h t e s hepatic acute-phase gene exprcs- sion. J Clin Invest 1986;78:1349-1354.
1988; 140:3473-3481.
1975; 141x269-286.
J EXP Med 1986; 163: 1433-1450.
28. Bevilacqua MP, Pober JS, Majeau GR, Fiers W, Cotra RS, Gim- bronc MA ?Jr. Recombinant tumor necrosis factor induces proco- agulant activity in cultured human vascular endothelium: char- acterization and comparison with the actions of interleukin-I. Proc Natl Acad Sci U S A 1986;83:4533-4537.
29. Feinman R, Henriksen-DeStefano D, Tsujimoto M, Vilcek J. Tu- mor necrosis factor is an important mediator oftumor cell killing by human monocytes. J Immunol 1987; 138:635-640.
30. Ruggiero V, Latham K, Haglioni C. Cytostatic and cytotoxic ac- tivity of tumor necrosis factor in human cancer cells. J Immunol
31. Scheurich P, Thomas R, Ucer M. Immunoreylatory activity of recombinant human tumor necrosis factor (TNF 1-u: induction of TNF receptors on human T cells and TNP-tr mediated enhance- ment of T cell responses. J Immunol 1987;138:1786-1790.
32. Yokota S, Geppert TD, Lipsky PE. Enhancement of antigen- and mitogen-induced human T lymphocyte proliferation by tumor necrosis factor-u. J Immunol 1988; 14053 1-536.
33. Scott KL), Oppenheim J J . Macrophagc-derived mediator: in- terleukin 1, tumor necrosis factor, interleukin 6, interferon and related cytokines. In: Paul WE, ed. Fundamental Immunology. New York: Raven, 1989;639-661.
34. Chen L, Suzuki Y, Whelock EF. Interferon-y synergizes with tumor necrosis factor and with interleukin-1 and requires the presencc of both monokiries to induce antitumor cytotoxic activ- ity in macrophages. J Immunol 1987; 139:4096-4101.
35. Laster SM, Wood JG, Gooding GR. Tumor necrosis factor can induce both apoptic and necrotic forms of cell lysis. J Immunol
36. Phipps RP, Stein HS, Hoper RL. A new view of prostaglandin E regulation of the immune rcsponse. Immunol Today 1991; 12:
37. Kunkol ST, Chensue SW, Phan SH. Prostaglandins a s endoge- nous mediators of interleukin-1 production. J Immunol
38. Bonta IL, Hen-Efraim S. Involvement of inflammatory mediator in macrophage antitumor activity. J Lcukoc Biol 1993;54:613- 626.
39. Koehler L, Hass R, Wessel K, DcWitt DI,, Kaever V. Resch K, Goppelt-Struebc M. Altered arachidonic acid metabolism during differentiation of the human monoblastoid cell line U937. Rio- chim Riophys Acta 1990; 1042:395-403.
40. Knudsen P, Dinarello CA, Strom TR. Prostaglandins posttran- scriptionally inhihit monocytc expression of interleukin 1 activ- ity by increasing intracellular cyclic adenosine monophosphate. J Immunol 1986; 137:3189-3194.
41. Hass It, Fannkuche I), Kharbanda H, Gunji H, Meyw G , Hart- mcnn A, Hidaka H , et al. Protein kinase C activation and pro- tooncogene expression in differentiatiodretrodiffercntiation of human U937 leukemia cc!lls. Cell Growth Differ 1991;2:541-548.
42. Harris P, Ralph P. Human leukemic models of myelomonocytic dcvelopment: a review ofthe HL-60 and U937 cell lines. cJ Leukoc Hiol 1985;37:407-422.
43. Dodd RC, Cohen MS, Newman SL, Gray TK. Vitamin D metabo- lites change the phenotype of monoblastic U937 cells. Proc Natl Acad Sci U S A 1983;80:7538-7545.
44. Ralph PM, Moore AS, Nilsson K. Lysozyme synthcsis by estab- lished human and murine histiocytic lymphoma cell lines. J Exp Med 1976; 143:1528-1533.
45. Sztcin IMR. Steeg PS, Johnson HAM, Oppcnheim J J . Regulation of human peripheral blood monocyte DK antigen cxpression in uilro by lymphokines and recombinant interferon. J Clin Invest
46. Roux-Lombard P, Cruchaud A, Dayer ,J. Effect of interferon-y and l t r , 25-Dihydroxyvitamin D3 on superoxide anion, prosta- glandins E2, and mononuclear cell factor production by U937 cells. (:ell Immunol 1986;97:286-296.
1987; 138:27 11-2717.
1988; 14 1 :2629-2685.
271-276.
1986; 136x186-192.
1984; 73x556465.


















