
Clonación y caracterización de la β-glucanasa LicA...
28
Facultat de Biologia Departament de Microbiologia Memoria Practicum Máster en Biotecnología Molecular Clonación y caracterización de la β-glucanasa LicA de Paenibacillus barcinonensis Liliana Cerda Mejía Tutor: Francisco I. Javier Pastor Blasco Barcelona, 2012
Transcript of Clonación y caracterización de la β-glucanasa LicA...

Facultat de Biologia
Departament de Microbiologia
Memoria Practicum
Máster en Biotecnología Molecular
Clonación y caracterización de la β-glucanasa LicA de
Paenibacillus barcinonensis
Liliana Cerda Mejía
Tutor: Francisco I. Javier Pastor Blasco
Barcelona, 2012

ii
Clonación y caracterización de la β-glucanasa LicA de
Paenibacillus barcinonensis
Barcelona, Julio 2012
Visto Bueno del Director Memoria de Practicum del Máster
En Biotecnología Molecular
Dr. Francisco I. Javier Pastor Blasco Liliana Cerda Mejía
Estudios de Máster financiados por una beca del Gobierno de Ecuador (SENESCYT Convocatoria
Abierta 2011).

iii
Índice de contenido
INTRODUCCIÓN .................................................................................................................................. 1
Liquenasas (endo-β-1,3-1,4-glucanasa)............................................................................................... 1
Paenibacillus barcinonensis ................................................................................................................. 2
OBJETIVOS ............................................................................................................................................ 3
MATERIALES Y MÉTODOS ............................................................................................................... 4
Microorganismos y condiciones de cultivo ......................................................................................... 4
Técnicas generales de Biología Molecular .......................................................................................... 4
Electroforesis de ADN en geles de agarosa ........................................................................................ 4
Secuenciación de ADN plasmídico con kit PE BigDye: ..................................................................... 5
Transformación y clonación en Escherichia coli ................................................................................. 5
Obtención de extractos celulares ......................................................................................................... 6
Análisis proteico por electroforesis en geles de poliacrilamida con dodecil sulfato sódico (SDS-
PAGE) ................................................................................................................................................. 7
Purificación de Proteínas por cromatografía de afinidad (AKTA FPLC) ........................................... 7
Cuantificación de las proteínas por el Método de Bradford ................................................................ 8
Determinación de actividad enzimática............................................................................................... 8
Determinación de pH óptimo y temperatura óptima ........................................................................... 9
Determinación de los productos de reacción mediante cromatografía en capa fina .......................... 10
RESULTADOS ..................................................................................................................................... 11
Amplificación y secuenciación de genes homólogos a liquenasas en Paenibacillus barcinonensis y
Paenibacillus illinoisensis.................................................................................................................. 11
Clonación y expresión de LicA de Paenibacillus barcinonensis en Escherichia coli. ....................... 15
Actividad específica sobre diferentes sustratos ................................................................................. 16
Determinación de la Temperatura Óptima ........................................................................................ 17
Determinación de pH óptimo ............................................................................................................ 17
Determinación de la Termoestabilidad .............................................................................................. 18
Análisis de los productos de hidrolisis por cromatografía en capa fina ............................................ 19
DISCUSIÓN .......................................................................................................................................... 20
CONCLUSIONES ................................................................................................................................ 22
BIBLIOGRAFÍA ................................................................................................................................... 23

1
INTRODUCCIÓN
Los procesos productivos en la industria alimentaria están sometidos a un desarrollo continuo
para mejorar las tecnologías de producción con el fin de reducir los costos y la generación de
residuos contaminantes, y para mejorar los productos obtenidos. (Mc Genaro Alberto Gómez.
2012).
Los β-1,4-1,3-glucanos son carbohidratos complejos formados por unidades de glucosa unidas
por enlaces β-1,3 y β-1,4. Son polímeros lineales de hasta 1200 residuos β-D-glucosa que
muestran variación en la proporción enlaces glicosídicos β-1,3 y β-1,4 (25-30% β-1,3). Se
encuentran en las paredes celulares de cereales como la avena y la cebada. El contenido de β-
glucanos en cebada para cervecería puede afectar negativamente al proceso de fabricación ya
que éstos aportan viscosidad indeseable a los mostos cerveceros. Los β-glucanos junto con los
pentosanos presentes en la cebada forman gomas que son solubles en agua caliente y causan
problemas durante la filtración previa a la fermentación del mosto. También se les ha
asociado a problemas de turbidez y el empobrecimiento del extracto cervecero. (Jin Lai-Yu et
al. 2005)
En la fabricación de cerveza la tecnología enzimática juega un papel importante. La
aplicación de β-glucanasas exógenas es especialmente interesante ya que mejora tanto las
cualidades de la cerveza como la eficiencia de producción de la misma. (Sánchez et al. 2005)
Las β-glucanasas tienen también aplicación en la fabricación alimentos para animales de
granja ya que su adición a los piensos de aves de corral mejora su digestibilidad. (Jirawan.
2006)
Liquenasas (endo-β-1,3-1,4-glucanasa)
La depolimerización de los β-1,3-1,4-glucanos (glucanos mixtos) está catalizada por 3 tipos
de glicosil hidrolasas que difieren en la especificidad de sustrato: endoglucanasas (EC
3.2.1.4), específicas para enlaces β-1,4; laminarinasas (EC 3.2.1.39), específicas para enlaces
β-1,3; y liquenasas (EC 3.2.1.73). Estas últimas enzimas también denominadas endo-β-1,3-
1,4-glucanasas son las más activas sobre los glucanos mixtos. Hidrolizan los enlaces

2
glucosídicos β-1,4 adyacentes a los enlaces β-1,3 en β-1,3-1,4 glucanos, tales como el glucano
de cebada o el liquenano, pero no presentan actividad sobre los enlaces glucosídicos β-1,4 en
los auténticos β-(1,4)-glucanos como la celulosa y la carboximetil celulosa. (Planas. 2000)
La mayoría de liquenasas tienen un origen bacteriano y pertenecen a la familia 16 de las
glicosil hidrolasas. (Picart et al. 2012). Contrariamente a las endo-β-1,4-glucanasas
(celulasas), que se encuentran ampliamente distribuidas en distintos organismos, las
liquenasas se encuentran en su mayoría restringidas a plantas y algunas bacterias. (Jirawan.
2006)
Paenibacillus barcinonensis
Paenibacillus barcinonensis es una nueva especie bacteriana aislada de suelos de arrozales
del delta del Ebro (Blanco et al. 1993; Sánchez et al, 2005), lugar rico en material vegetal en
descomposición y por lo tanto adecuado para aislar microrganismos degradadores de
polisacáridos. Esta cepa fue aislada por el grupo de investigación con anterioridad y
seleccionada por su elevada actividad xilanolítica, celulolítica, y en general por la diversidad
de enzimas degradadores de carbohidratos que produce.
En base a los conocimientos ya generados por el grupo de investigación en cuanto a
caracterización y manipulación de enzimas, y ante la necesidad de la industria de contar con
nuevos elementos enzimáticos que permitan mejorar los actuales procesos productivos, se
justifica la necesidad de buscar nuevas enzimas con potencial de aplicación industrial, además
de la caracterización de los mecanismos moleculares de acción que permitan dilucidar los
posibles usos que pueda tener las nuevas enzimas.

3
OBJETIVOS
Los objetivos de este estudio son:
1. Clonación y expresión de una endo-β-1,3-1,4-glucanasa de Paenibacillus
barcinonensis.
2. Caracterización de endo-β-1,3-1,4-glucanasa de Paenibacillus barcinonensis.

4
MATERIALES Y MÉTODOS
Microorganismos y condiciones de cultivo
La liquenasa estudiada en el presente trabajo proviene de la cepa Paenibacillus barcinonensis
(Blanco et al. 1993; Sánchez et al, 2005) aislada a partir de suelo de arrozal de Delta del Ebro.
Para su crecimiento se utilizó medio LB para el cultivo en medio líquido y agar nutritivo para
el cultivo en medio sólido.
Para los trabajos de biología molecular se utilizaron las cepas Escherichia coli DH5α, y E.
coli BL21 Star (DE3). Se cultivaron en medio LB (Sambrook et al. 1989). Cuando fue
requerido, los cultivos se suplementaron con ampicilina (LB-Amp) a una concentración final
de 100 g/ml. Para la preparación de medio sólido se adicionó agar a una concentración final
de 1.5 % (p/v).
Técnicas generales de Biología Molecular
Se realizaron los procedimientos, según las técnicas rutinarias descritas en el Handbook de
Laboratorio (Sambrook et al. 1989) o las instrucciones de los kit comerciales utilizados.
La extracción de ADN genómico se realizó mediante el Genomic DNA extraction Kit, de
Biotools. La extracción de ADN plasmídico con la metodología descrita en el manual de
Illustra™ plasmad prep. Mini Spin Kit de GE™ Health Care.
Electroforesis de ADN en geles de agarosa
Las preparaciones de ADN se analizaron mediante la técnica de electroforesis en geles de
agarosa. Los geles de 0.8% de agarosa se prepararon en tampón TAE 1X suplementados con
1l de Nancy520. Las muestras de ADN se mezclaron con tampón de carga (0.25 % de Azul
de Bromofenol, 40 % de Sacarosa). Las bandas se visualizaron por la fluorescencia emitida al
irradiar con luz ultravioleta.

5
Secuenciación de ADN plasmídico con kit PE BigDye:
Se utilizó la secuenciación automática, empleando para ellos el kit de secuenciación Big Dye
de los Servicios científicos técnicos de la Universidad de Barcelona.
PCR de secuenciación:
- 4 l de terminator ready reaction mix.
- 1 l de cebador forward y backward
- 2 l de molde de ADN
- Agua bidestilada estéril hasta 10 l.
Programa de PCR de secuenciación:
- Hold inicial: 94 ºC 3 min. Permite asegurarse de que se desnaturaliza el molde y se
rompen los puentes de hidrógeno internos de los cebadores.
- Desnaturalización: 96 ºC 10 seg.
- Hibridación: la temperatura de hibridación depende de la Tm (temperatura de fusión)
de los cebadores, y el tiempo de hibridación es de 5 seg.
- Síntesis: 60 ºC 4 min. Con 25 ciclos, y el final a 4ºC.
Análisis de Secuencias:
Se realizaron búsquedas de homología tanto de secuencias de nucleótidos como de
aminoácidos usando el programa Blast del NCBI (Altschul et al., 1997).
Transformación y clonación en Escherichia coli
La ligación y transformación en Escherichia coli se realizó con el kit Champion pET TOPO
® (Invitrogen) siguiendo las instrucciones del fabricante. Los productos de PCR frescos,
purificados y cuantificados se introdujeron en Escherichia coli. Para ello se mezclaron los
productos de PCR (0.5 a 4l) con 1l de solución salina, 1l de Vector TOPO y agua estéril
hasta un volumen final de 5l. Mezclar suavemente e incubar 1 h a temperatura ambiente.
Colocar en hielo y transferir a un vial de células competentes de E. coli One Shot ® TOP10
previamente incubadas en hielo. Incubar en hielo durante 10 min. Dar un choque térmico

6
durante 30 seg a 42ºC sin agitación y transferir inmediatamente el vial a hielo. Añadir 250l
de medio S.O.C a temperatura ambiente, e incubar a 37ºC durante una h a 350 rpm.
Transcurrido este tiempo sembrar 100l en una placa LB/amp100 e incubar toda la noche a
37ºC.
Para transformar E. coli en BL21 Star ™ (DE3) One Shot ®, se sigue el siguiente protocolo:
- Descongelar en hielo, un vial de células BL21 Star ™ (DE3) One Shot ® para la
transformación.
- Añadir 5-10 ng de ADN plásmidico en cada vial de BL21 Star ™ (DE3) One Shot ® y
mezclar las células agitando suavemente con la punta de la pipeta. No mezclar
pipeteando de arriba y hacia abajo.
- Incubar en hielo durante 30 min.
- Choque térmico de las células durante 30 seg. a 42 °C sin agitación.
- Inmediatamente transferir los tubos a hielo.
- Añadir 250 l de medio S.O.C a temperatura ambiente.
- Incubar a 37 °C durante una h con agitación (200 rpm).
- Añadir la reacción de transformación completa a 10 ml de LB con ampicilina 100
g/ml
- Cultivar durante la noche a 37 °C con agitación.
Obtención de extractos celulares
Las cepas recombinantes se cultivaron en LB ampicilina 100 g/ml a 37 ºC. Cuando se
alcanzó una DO600 de aproximadamente 0.5-0.8 (fase exponencial intermedia) se añadió IPTG
a una concentración final de 0,5mM. Tras incubación durante 16h las células bacterianas se
recuperaron por centrifugación a 5000 rpm durante 20 minutos a 4ºC.
Cuando el cultivo celular era de un volumen pequeño, la lisis celular se realizaba por
sonicación, con pulsos de 2 min, a una frecuencia de 0,9 seg y 90 vatios de potencia
(Sonicador Labsonic1510, B. Braun). Cuando el volumen de la suspensión celular era grande,
las células se lisaban por French Press (French Pressure Cell Press, SLM AMINCO),
sometiendo la muestra por 3 ocasiones a una presión de 1000 psi. Los lisados obtenidos se

7
centrifugaron durante 30 minutos a 18.000 rpm y se recogieron los sobrenadantes,
correspondientes a los extractos solubles. Todas las muestras, se conservaron a 4ºC para su
análisis o bien se congelaron a -20ºC para análisis posteriores.
Análisis proteico por electroforesis en geles de poliacrilamida con dodecil sulfato sódico
(SDS-PAGE)
Se procedió según el protocolo diseñado por Laemmli (1970), que consta de los siguientes
pasos
Mezclar las muestras tanto de sobrenadante como de pellet con tampón de carga, de
composición Tris-HCl 0,0625 M (pH 6,8), SDS al 2%, glicerol al 10%, 2-mercaptoetanol al
5% y azul de bromofenol al 0,001%. Incubar 10 min a 50ºC.
Para el análisis de las proteínas totales se utilizaron geles de poliacrilamida al 12%. Las
electroforesis se desarrollaron a 100 V, hasta que el frente de migración alcanzó el borde
inferior del gel. Los geles se tiñeron 30 min con una solución de azul de Coomassie al 0.2%,
metanol al 50% y ácido acético al 7%. Se destiñeron luego mediante lavados sucesivos con
una solución de metanol al 20% y ácido acético al 7%.
Para la detección de actividad liquenasa en geles de proteínas se utilizaron geles habituales
que contenían β-glucano de cebada al 0,2%, añadido antes de polimerizarlos. Una vez
desarrollada la electroforesis los geles se lavaron con Triton X-100 al 2,5% durante 30min,
tampón fosfato pH 6,5 durante 30min, y se incubaron a 55 ºC durante 15 min. Los geles se
tiñeron a continuación con Rojo Congo al 0,1% y se lavaron con NaCl 1M hasta que las
bandas de actividad liquenasa se visualizaron. Los geles se sumergieron a continuación en
ácido acético al 5%, en los que se volvieron de color azul oscuro, y se fotografiaron.
Purificación de Proteínas por cromatografía de afinidad (AKTA FPLC)
La enzima con cola de histidinas en su extremo C-terminal, se purifico mediante
cromatografía de afinidad a columnas de níquel (His-Trap columns, Amersham-Pharmacia)
en un equipo AKTA-FPLC.

8
Al trabajar con proteínas solubles, la purificación se realizó bajo condiciones no
desnaturalizantes. Estas condiciones nativas permiten una alta unión inespecífica a las
columnas de níquel, para reducirla se añadió imidazol a baja concentración (10mM) en el
tampón de lavado. Se inyectaron 2ml de extractos celulares de E. coli
BL21(DE3)/pET101LicA en tampón de lavado a las columnas de níquel, previamente
equilibradas con 10 volúmenes de tampón de lavado. Tras la inyección. Las proteínas
retenidas se eluyeron de la columna con un gradiente lineal del tampón de elución y las
fracciones positivas recogidas se analizaron mediante SDS-PAGE.
Cuantificación de las proteínas por el Método de Bradford
Se prepararon diferentes diluciones de albumina de suero bovina (BSA) entre 0,1 y 1 mg/ml,
y de la solución a cuantificar en un volumen de 15l. Cada muestra, tanto la estándar de BSA
como la muestra analizarse, se preparan por triplicado. Se añadieron 150 l de colorante Bio-
Rad Protein Assay (Bio-Rad), se mezcló y se midió la absorbancia a 595nm.
Determinación de actividad enzimática
La actividad de la liquenasa (endo-β-1,3-1,4-glucanasa) se determinó por valoración de
azúcares reductores mediante el ensayo de Nelson-Somogyi (Spiro, 1966).
Cada una de las mezclas de reacción contenía 33 μl de la muestra a valorar, 33 μl de tampón
fosfato sódico 150 mM pH 6,5 y 33 μl de sustrato (1,5%). Las mezclas de reacción se
prepararon en hielo y fueron incubadas a continuación 15 min a 55 ºC. Luego se enfriaron en
hielo durante 5 min y se agregó 150 l de agua bidestilada. Se añade 250l del reactivo
alcalino de cobre, el cual se prepara mezclando las soluciones I y II en relación 4:1
inmediatamente antes de su utilización.
Preparación solución I: 15 g de tartrato sódico potásico y 30 g de Na2CO3 se disuelven en 300
ml de agua; se añade 20 g de NaHCO3 y una solución de Na2SO4 de 180 g en 500 ml de agua
y se afora a 1 l.
Preparación solución II: 5g de CuSO4·5H2O y 45 g de Na2SO4 se disolvieron en 250 ml de
agua.

9
Las muestras se incuban a 100ºC por 10 min en baño de agua hirviendo y se enfrían después
en hielo. Se añade 250l de solución de arsenomolibdato mezclado antes de su utilización con
H2SO4 1,5N en una relación 1:2. Después se agita y se agrega 750 μl de agua. Finalmente se
centrifugan las muestras 5 min a 4ºC y se mide la absorbancia a 520 nm de los sobrenadantes.
Cada reacción se realiza por triplicado y se lee frente a un blanco en que la enzima se agrega a
los tubos después de añadir el reactivo de cobre.
Preparación de la solución de arsenomolibdato: Se disuelve 25 g de (NH4)6Mo7O24·4H2O en
450 ml de agua, se añade 21 ml de H2SO4 96%, agitar, y luego añadir 3 g de Na2HAsO4·7H2O
disueltos en 25 ml de agua. El reactivo se incuba durante 24-48 h a 37 ºC y se guarda en una
botella opaca.
La unidad enzimática se definió como la cantidad de enzima que cataliza la liberación de un
μmol de azúcar reductor por min en las condiciones anteriormente descritas.
La especificidad de sustrato se determino realizando el ensayo de actividad sobre diferentes
tipos de sustratos con el método antes descrito. Los sustratos fueron xilano de madera de
abedul, xilano de espelta de avena, arabinoxilano de trigo, xilano de madera de haya, 4-O-
metilglucoroxilano, liquenano, laminarina, β-glucano de cebada, pectina, ácido
poligalacturónico, almidón, Avicel y carboximetil celulosa, y se valoro la actividad
enzimática sobre sustrato.
Determinación de pH óptimo y temperatura óptima
El pH óptimo de la actividad liquenasa se determinó valorando dicha actividad a diferentes
pH entre 3,0 y 10,0 en intervalos de 0,5 y 1 unidades.
Las soluciones tampón utilizadas fueron:
Soluciones de Britton Bristol para pH entre 3 y 10
Fosfato sódico para pH de 6,5.
Para la determinación de la temperatura optima se realizaron valoraciones de actividad
liquenasa por el método habitual, aplicando diferentes temperaturas de incubación en el rango
de 20 – 70ºC, con intervalos de 5 y 10ºC.

10
La estabilidad de la actividad liquenasa a diferentes temperaturas se determinó incubando la
muestra a diferentes temperaturas, en el rango entre 45 y 65ºC, durante diferentes intervalos
de tiempo al pH optimo, valorándose a continuación la actividad enzimática residual por el
método habitual.
Determinación de los productos de reacción mediante cromatografía en capa fina
La cromatografía en capa fina TLC (Thin layer chromatography) se utilizó para analizar los
productos de hidrolisis generados por la actividad de la enzima LicA sobre diferentes
sustratos glucanos (liquenano y β–glucano de cebada).
Para ello, se incubó 76 ng de enzima purificada con sustratos al 1,5% en tampón fosfato
150mM pH 6,5 a 55ºC. A los intervalos de tiempo establecidos se extrajeron alícuotas, se
centrifugaron a 14000 rpm durante 3 min y los sobrenadantes fueron congelados. Muestras de
10 l de los sobrenadantes se aplicaron en placas de sílica gel (Kieselgel 20 F254 20x20 cm,
Merck), en paralelo con una mezcla de azucares patrón: glucosa, celobiosa, celotriosa,
celotetraosa y celopentosa (Sigma), (20 g de cada uno). La fase móvil consistió en una
mezcla de cloroformo, ácido acético y agua (30:60:10 v/v/v). Se realizaron dos ascensos del
eluyente y, a continuación, se secaron las placas y se rociaron con una solución de H2SO4 al
5% en etanol. Finalmente, las placas se revelaron calentándolas con un decapador hasta
observar la aparición de las manchas correspondientes a los productos de degradación.

11
RESULTADOS
El grupo de investigación en el que se ha realizado el trabajo estudia la degradación de
polisacáridos por la nueva especie Paenibacillus barcinonensis, cepa aislada a partir de suelo
de arrozal del Delta del Ebro. En resultados previos, el grupo ha caracterizado y clonado
varias xilanasas. Uno de los clones obtenidos contiene un gen con un marco abierto de lectura
que hipotéticamente codifica una proteína con homología a liquenasas. El objetivo del
presente trabajo ha sido clonar y verificar la expresión de dicha proteína y caracterizar su
actividad enzimática.
Amplificación y secuenciación de genes homólogos a liquenasas en Paenibacillus
barcinonensis y Paenibacillus illinoisensis
La secuencia del gen de la hipotética liquenasa, permitió el diseño de cebadores específicos
para la amplificación por PCR del gen codificante.
La secuencia de los cebadores es la siguiente: Forward Liquenasa: 5’-
CACCATGATGAAGAGGAAAACAT-3’, Backward TOPO Liquenasa: 5’-
GTTGCTCGTATATTTCACCCAGT-3’
Se utilizó como molde el ADN genómico Paenibacillus barcinonencis, así como de otras
cepas de la colección del Grupo de Investigación seleccionadas previamente debido a su gran
poder degradador de polisacáridos. Las cepas fueron Paenibacillus illinoisensis y
Paenibacillus flavisporus, cepas previamente aisladas de suelos áridos de São João do Carini,
Paraíba, ubicado en el Noreste de Brasil.
Con los cebadores específicos para la enzima de Paenibacillus barcinonencis, no solo se
logro amplificar el fragmento correspondiente de esta cepa. También se consiguió amplificar
un fragmento de ADN de la cepa de Paenibacillus illinoisensis, mientras que no se obtuvo
amplificado cuando se utilizó como molde Paenibacillus flavisporus (Fig. 1).

12
Fig. 1. Amplificación de ADN por PCR. M, Marcador de Peso Molecular; 1, Paenibacillus barcinonensis; 2,
Paenibacillus flavisporus; 3, Fragmento de Paenibacillus illinoisensis.
La secuenciación del fragmento amplificado del genoma de Paenibacillus illinoisensis mostró
que contenía un marco abierto de lectura de 695 pb que codificaba una proteína de 231
aminoácidos (Fig. 2). La secuencia de aminoácidos deducida muestra una región N-terminal
de 32 aminoácidos con las características de un péptido señal. El peso molecular teórico de la
proteína es 26,07 kDa y el punto isoélectrico (pI) de la proteína es 6,41.
ATG ATG AAG AGG AAA ACA TGG TAC ACA TTG ATT GTT TCG GGA GTG
1 Met Met Lys Arg Lys Thr Trp Tyr Thr Leu Ile Val Val Ser Gly 15
GTT TCT TTA TTT TTT TCG GTA AGC GCT TTC GCG GGG TAT GTG TTC
16 Val Val Ser Leu Phe Phe Ser Val Ser Ala Phe Ala Gly Tyr Val 30
TGG GAG CCG TTA ACC TAC TTC AAT TCT AGT ACG TGG CAA AAG GCA
31 Phe Trp Glu Pro Leu Thr Tyr Phe Asn Ser Ser Thr Trp Gln Lys 45
GAT GGA TAC TCC AAC GGC TCC ATG TTT AAC TGT ACT TGG CGG GCG
46 Ala Asp Gly Tyr Ser Asn Gly Ser Met Phe Asn Cys Thr Trp Arg 60
AAC AAT GTC AAT TTT ACA AGT GAT GGT AAA CTG CGA CTG GGT TTG
61 Ala Asn Asn Val Asn Phe Thr Ser Asp Gly Lys Leu Arg Leu Gly 75
ACT AGT TCG GCA TAC AAC AAA TTT GAT TGC GGA GAA TAC CGA ACA
76 Leu Thr Ser Ser Ala Tyr Asn Lys Phe Asp Cys Gly Glu Tyr Arg 90
ACC AAC AGT TAT GGA TAT GGA CTT TAC GAA GTT AGC ATG AAG CCT
91 Thr Thr Asn Ser Tyr Gly Tyr Gly Leu Tyr Glu Val Ser Met Lys 105
GCT AAA AAT ACA GGG ATT GTA TCC TCT TTT TTC ACC TAT ACC GGT
106 Pro Ala Lys Asn Thr Gly Ile Val Ser Ser Phe Phe Thr Tyr Thr 120
CCT TCT GAT GGC ACA CAA TGG GAT GAG ATT GAT ATC GAG TTC CTT
121 Gly Pro Ser Asp Gly Thr Gln Trp Asp Glu Ile Asp Ile Glu Phe 135
GGT AAA GAC ACA ACG AAA GTA CAA TTC AAC TAT TAT ACA AAC GGC
136 Leu Gly Lys Asp Thr Thr Lys Val Gln Phe Asn Tyr Tyr Thr Asn 150
1 2 3 M
10000
750 500
250
3000
4000
6000
2000
1500
1000

13
GTT GGA AAT CAT GAG AAG ATC GTA AAT CTG GGC TTC GAT GCG TCC
151 Gly Val Gly Asn His Glu Lys Ile Val Asn Leu Gly Phe Asp Ala 165
CAG GGA TTC CAC ACG TAT GCG TTT GAT TGG CAA CCA GGT CAC ATT
166 Ser Gln Gly Phe His Thr Tyr Ala Phe Asp Trp Gln Pro Gly His 180
AAG TGG TAC GTC GAT GGC GTG CTG AAA CAT ACA GCT ACA ACC AAT
181 Ile Lys Tyr Val Asp Gly Val Leu Lys His Thr Ala Thr Thr Asn 195
ATT CCG AAA ACA CCA GGT AAA ATT ATG ATG AAC TTG TGG AAC GGA
196 Ile Pro Lys Thr Pro Gly Lys Ile Met Met Asn Leu Trp Asn Gly 210
ACC GGC GTT GAC AGC TGG TTA GGG GCA TAT AAT GGA GCT AAT CCG
211 Thr Gly Val Asp Ser Trp Leu Gly Ala Tyr Asn Gly Ala Asn Pro 225
TTA TAC GCA GAA TAT GAC UGA
226 Leu Tyr Ala Glu Tyr Asp Stop
Fig. 2. Secuencia de nucleótidos y secuencia deducida de aminoácidos del gen de Paenibacillus illinoisensis.
El fragmento de ADN amplificado de Paenibacillus barcinonensis fue secuenciado para
verificación. Contiene un marco de abierto de lectura de 717 pb que codifica una proteína de
239 aminoácidos (Fig. 3). La secuencia de aminoácidos deducida muestra una región N-
terminal de 25 aminoácidos con las características de un péptido señal (Fig. 4). El peso
molecular teórico de la proteína es de 25,83 kDa y el punto isoélectrico (pI) de la proteína es
6,71.
ATG ATG AAG AGG AAA ACA TGG TTC ACA TTA ATG GTA TCG GGA GTG
1 Met Met Lys Arg Lys Thr Trp Phe Trp Leu Met Val Ser Gly Val 15
GTT TCA TTA TTT ATT TCA GTA AGC GCT TTC GCA GGG TAT GTA TTT
16 Val Ser Leu Phe Ile Ser Val Ser Ala Phe Ala Gly Tyr Val Phe 30
TGG GAG CCG CTA ACG TAT CAC AAT CCA AGC ACA TGG CAG AAG GCA
31 Trp Glu Pro Leu Thr Tyr His Asn Pro Ser Thr Trp Gln Lys Ala 45
GAT GGG TAC TCC AAC GGC TCG ATG TTC AAT TGT ACA TGG CGA GCG
46 Asp Gly Tyr Ser Asn Gly Ser Met Phe Asn Cys Thr Trp Arg Ala 60
AAC AAT GCT AAT TTT ACT AAT GAC GGT AAG CTG CGT TTG AGT CTG
61 Asn Asn Ala Asn Phe Thr Asn Asp Gly Lys Leu Arg Leu Ser Leu 75
ACA AGT CCT TCA AAC AAC AAG TTT GAT TGT GGT GAG TAT CGG TCA
76 Thr Ser Phe Ser Asn Asn Lys Phe Asp Cys Gly Glu Tyr Arg Ser 90
ACG AAT ACT TAC GGG TAT GGT CTG TAT GAG GTC AGC ATG AAA CCT
91 Thr Asn Thr Lys Gly Tyr Gly Leu Tyr Glu Val Ser Met Lys Pro 105
GCT AAA AAC ACA GGA ATT GTA TCC TCC TTC TTT ACG TAT ACG GGA
106 Ala Lys Asn Thr Gly Ile Val Ser Ser Phe Phe Thr Tyr Thr Gly 120
CCA GCC CAT GGC ACA CAA TGG GAT GAG ATT GAT ATT GAG TTT CTT
121 Pro Ala His gly Thr Gln Trp Asp Glu Ile Asp Ile Glu Phe Leu 135
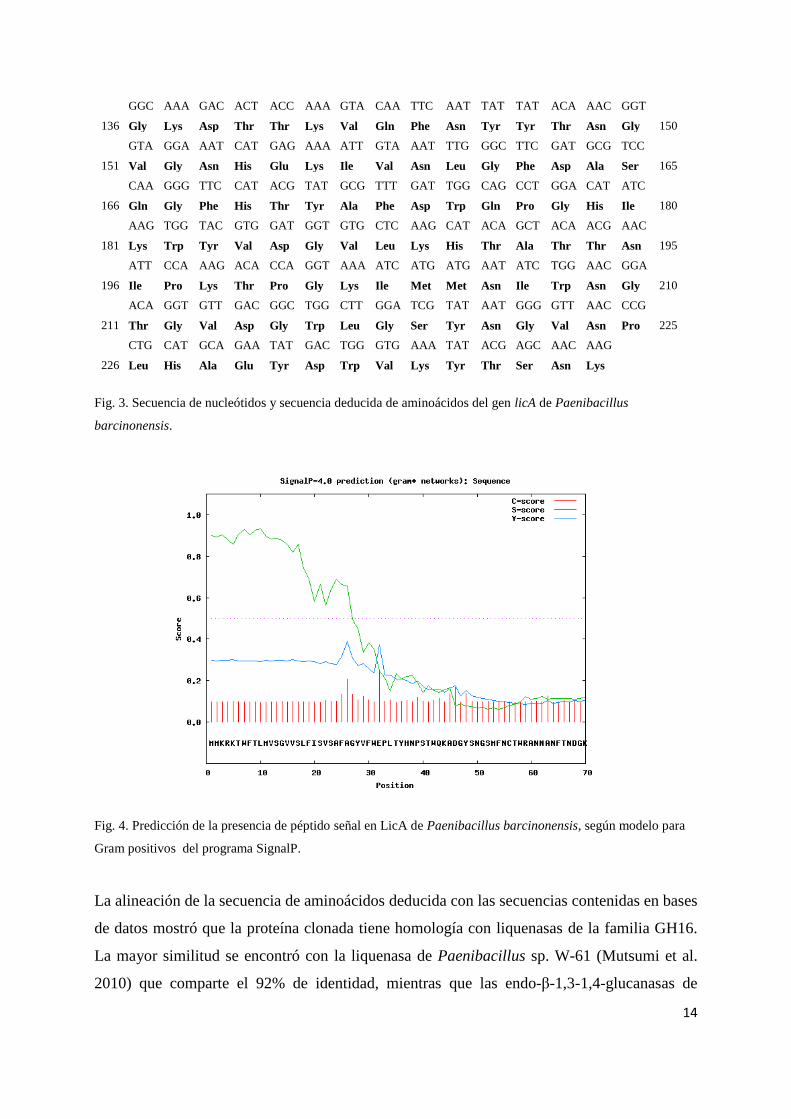
14
GGC AAA GAC ACT ACC AAA GTA CAA TTC AAT TAT TAT ACA AAC GGT
136 Gly Lys Asp Thr Thr Lys Val Gln Phe Asn Tyr Tyr Thr Asn Gly 150
GTA GGA AAT CAT GAG AAA ATT GTA AAT TTG GGC TTC GAT GCG TCC
151 Val Gly Asn His Glu Lys Ile Val Asn Leu Gly Phe Asp Ala Ser 165
CAA GGG TTC CAT ACG TAT GCG TTT GAT TGG CAG CCT GGA CAT ATC
166 Gln Gly Phe His Thr Tyr Ala Phe Asp Trp Gln Pro Gly His Ile 180
AAG TGG TAC GTG GAT GGT GTG CTC AAG CAT ACA GCT ACA ACG AAC
181 Lys Trp Tyr Val Asp Gly Val Leu Lys His Thr Ala Thr Thr Asn 195
ATT CCA AAG ACA CCA GGT AAA ATC ATG ATG AAT ATC TGG AAC GGA
196 Ile Pro Lys Thr Pro Gly Lys Ile Met Met Asn Ile Trp Asn Gly 210
ACA GGT GTT GAC GGC TGG CTT GGA TCG TAT AAT GGG GTT AAC CCG
211 Thr Gly Val Asp Gly Trp Leu Gly Ser Tyr Asn Gly Val Asn Pro 225
CTG CAT GCA GAA TAT GAC TGG GTG AAA TAT ACG AGC AAC AAG
226 Leu His Ala Glu Tyr Asp Trp Val Lys Tyr Thr Ser Asn Lys
Fig. 3. Secuencia de nucleótidos y secuencia deducida de aminoácidos del gen licA de Paenibacillus
barcinonensis.
Fig. 4. Predicción de la presencia de péptido señal en LicA de Paenibacillus barcinonensis, según modelo para
Gram positivos del programa SignalP.
La alineación de la secuencia de aminoácidos deducida con las secuencias contenidas en bases
de datos mostró que la proteína clonada tiene homología con liquenasas de la familia GH16.
La mayor similitud se encontró con la liquenasa de Paenibacillus sp. W-61 (Mutsumi et al.
2010) que comparte el 92% de identidad, mientras que las endo-β-1,3-1,4-glucanasas de

15
Paenibacillus sp. F- 40 (Yang et al. 2006) y Paenibacillus polymyxa (Gosalbes et al. 2006)
muestran una identidad de 89% con la enzima clonada. Los residuos catalíticos teóricos son:
Glu89 y Glu85, estos fueron encontrados en la posición de las regiones conservadas de las
GH16. (Pekka et al. 2009). La enzima clonada se denominó LicA.
La comparación con el programa BLAST NCBI de las enzimas clonadas de Paenibacillus
barcinonensis y Paenibaacillus illinoisensis, mostraron un 83% de identidad
Clonación y expresión de LicA de Paenibacillus barcinonensis en Escherichia coli.
El fragmento amplificado de ADN de Paenibacillus barcinonensis se clonó en el vector
pET101-TOPO utilizando como huésped E. coli BL21(DE3) dando lugar a la cepa
recombinante E. coli BL21(DE3)/pET101licA. Dicha construcción se diseñó para añadir una
cola de histidinas al C-terminal de la enzima clonada. Análisis por SDS-PAGE de extractos
celulares del clon recombinante mostraron un buen nivel de expresión de la proteína en la
fracción soluble de los mismos, comprobándose que la proteína no se expresaba en cuerpos de
inclusión. Por este motivo la cepa construida se utilizó para producir y caracterizar la enzima.
Los ensayos de expresión de la proteína recombinante llevados a cabo determinaron que las
condiciones óptimas para la expresión de la proteína son inducción con 0,5 mM de IPTG
durante 16 horas a 30ºC.
Cultivos de la cepa en 500 mL de LB con ampicilina 100 g/ml fueron recogidos por
centrifugación, las células se lisaron por French Press, y los extractos celulares obtenidos se
utilizaron para purificar la liquenasa. La liquenasa fue purificada mediante cromatografía de
afinidad a una columna His-trap y se analizó mediante SDS-PAGE y zimografía (Fig. 5). La
proteína mostró un peso molecular aparente de 27 kDa, de acuerdo con el peso molecular
deducido de la secuencia de nucleótidos.

16
Fig. 5. Análisis electroforético por SDS–PAGE y zimografía de LicA purificada. M, marcador de peso
molecular; 1, tinción proteica; 2, zimograma
Actividad específica sobre diferentes sustratos
La actividad hidrolítica de la enzima recombinante se determinó sobre varios sustratos. La
enzima mostró actividad únicamente sobre liquenano (118,37 U/mg) y β–glucano de cebada
(197,86 U/mg), lo que nos indica que es una autentica liquenasa, ya que no degrada celulosa
ni otros polisacáridos (Tabla 1).
Tabla 1. Especificidad de Sustrato de LicA
Sustratos
Actividad (U/mgProt)
Xilano de madera de haya ND
Xilano de espelta de avena ND
Arabinoxilano de trigo ND
Xilano de madera de abedul ND
4-O-metil glucuronoxilano ND
Liquenano 118,36
β-glucano de cebada 197,85
Pectina ND
Almidón ND
Avicel ND
Ácido Poligalacturonico ND
Carboximetil Celulosa ND
Laminarina ND
ND: No detectada
200
50
30
25
kDa
M 1 2
20
85
60
40
17
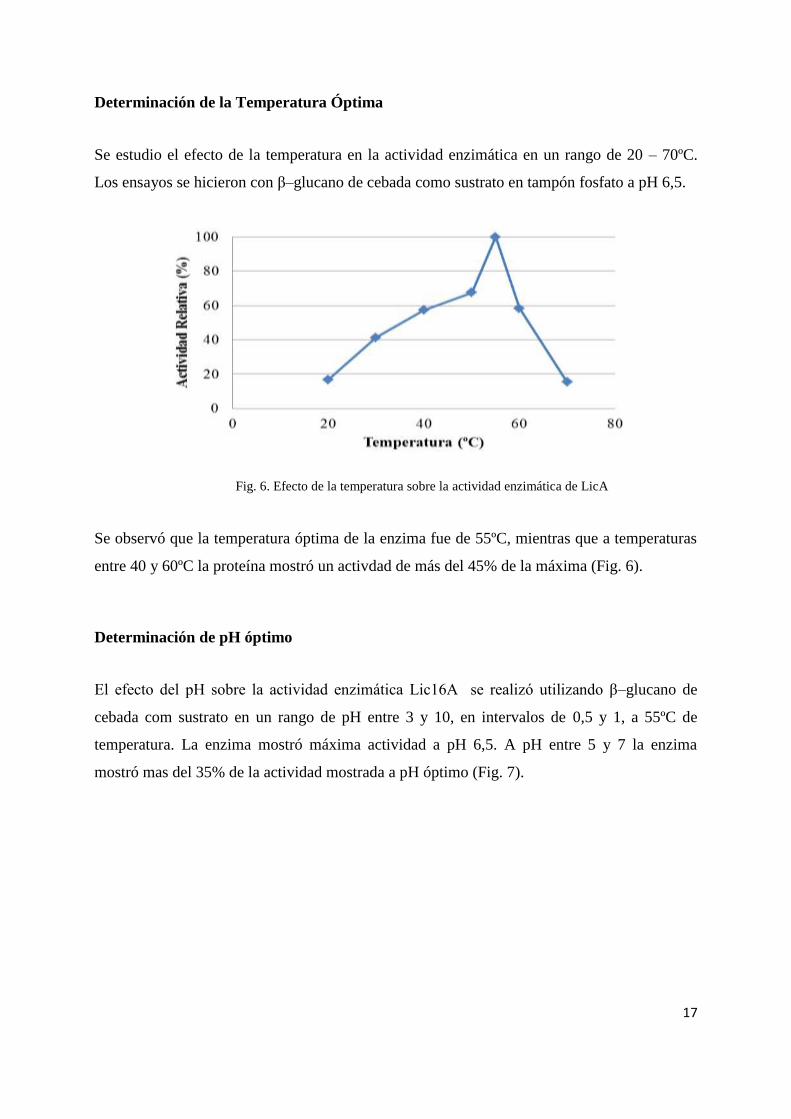
Determinación de la Temperatura Óptima
Se estudio el efecto de la temperatura en la actividad enzimática en un rango de 20 – 70ºC.
Los ensayos se hicieron con β–glucano de cebada como sustrato en tampón fosfato a pH 6,5.
Fig. 6. Efecto de la temperatura sobre la actividad enzimática de LicA
Se observó que la temperatura óptima de la enzima fue de 55ºC, mientras que a temperaturas
entre 40 y 60ºC la proteína mostró un activdad de más del 45% de la máxima (Fig. 6).
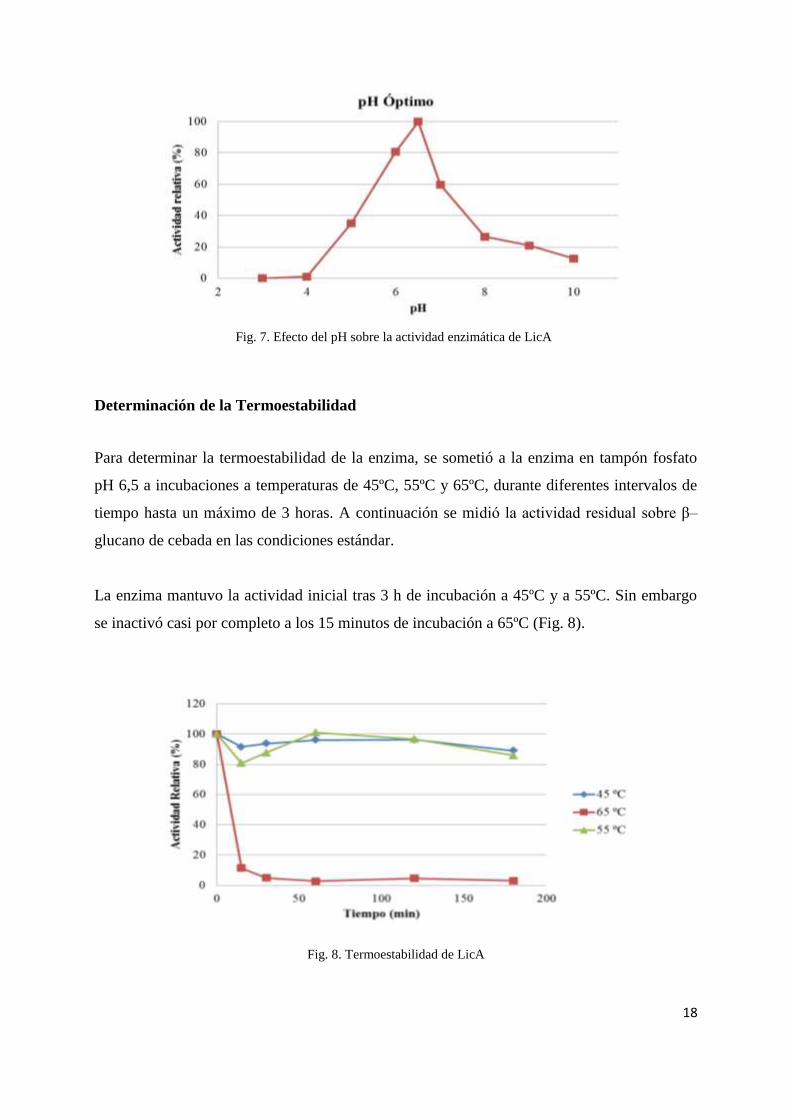
Determinación de pH óptimo
El efecto del pH sobre la actividad enzimática Lic16A se realizó utilizando β–glucano de
cebada com sustrato en un rango de pH entre 3 y 10, en intervalos de 0,5 y 1, a 55ºC de
temperatura. La enzima mostró máxima actividad a pH 6,5. A pH entre 5 y 7 la enzima
mostró mas del 35% de la actividad mostrada a pH óptimo (Fig. 7).
18
Fig. 7. Efecto del pH sobre la actividad enzimática de LicA
Determinación de la Termoestabilidad
Para determinar la termoestabilidad de la enzima, se sometió a la enzima en tampón fosfato
pH 6,5 a incubaciones a temperaturas de 45ºC, 55ºC y 65ºC, durante diferentes intervalos de
tiempo hasta un máximo de 3 horas. A continuación se midió la actividad residual sobre β–
glucano de cebada en las condiciones estándar.
La enzima mantuvo la actividad inicial tras 3 h de incubación a 45ºC y a 55ºC. Sin embargo
se inactivó casi por completo a los 15 minutos de incubación a 65ºC (Fig. 8).
Fig. 8. Termoestabilidad de LicA

19
Análisis de los productos de hidrolisis por cromatografía en capa fina
Los productos de hidrólisis de la enzima sobre de β–glucano de cebada y liquenano se
analizaron mediante cromatografía en capa fina (TLC). Los principales productos de
degradación del liquenano fueron mayoritariamente celotriosa y celopentosa (Fig. 9). Un
patrón de hidrólisis más complejo se obtuvo a partir de β–glucano de cebada, que fue
degradado a una mezcla de productos que van desde los oligosacáridos de tamaño largos hasta
productos de mayor movilidad como la celotriosa, incluyendo productos de movilidad
intermedia entre celotriosa y celotetraosa, y entre celotetraosa y celopentaosa sugiriendo que
estos productos de hidrólisis contienen enlaces β-1,3 conjuntamente con enlaces β-1,4.
Fig. 9. Productos de hidrolisis de liquenano y β-glucano de cebada. LicA (0,076ug) se incubó a 55ºC y pH 6,5
con β–glucano de cebada o liquenano al 1,5% y se analizaron muestras a distintos tiempos de incubación. 1:
tiempo 0 de incubación, 2: 15 min, 3: 60 min, 4: 24 h. M: marcador de degradacion de oligosacaridos. G1:
glucosa, G2: celobiosa, G3: celotriosa, G4: celotetraosa; G5: celopentosa.
Liquenano β – glucano de cebada
G1
G2
G3
G4
G5
M 1 3 4 2 1 3 4 2 M

20
DISCUSIÓN
Se ha clonado, expresado y purificado un nueva enzima degradadora de polisacáridos de
Paenibacillus barcinonensis. Los resultados de especificidad de sustrato de la nueva enzima,
que se ha denominado LicA indican que esta enzima es una endo-β-1,3-1,4-glucanasa o
liquenasa (EC 3.2.1.73) ya que presenta elevada actividad sobre β–glucano de cebada y
liquenano, mientras que no muestra actividad sobre celulosa (CMC, Avicel), laminarina, ni
otros polisacáridos. El hecho de que la enzima clonada solo muestra actividad sobre glucanos
con enlaces mixtos β-1,3-1,4 y no sobre glucanos β-1,4 (celulosa) ni sobre glucanos β-1,3
(laminarina) indica claramente que es un liquenasa.
Los análisis de secuencia revelaron que LicA de Paenibacillus barcinonensis es una enzima
de dominio único con alta homología con liquenasas de la familia 16 de glicosil hidrolasas
(GH16). LicA mostró elevada homología de secuencia con la liquenasa de Paenibacillus sp.
W-61 (Mutsumi et al. 2010) y las endo-β-1,3-1,4-glucanasas de Paenibacillus sp. F- 40 (Yang
et al. 2006) y Paenibacillus polymyxa (Gosalbes et al. 1991). LicA presenta características
bioquímicas (especificidad de sustrato, pH y temperatura óptimos) similares a la liquenasa de
Paenibacillus sp. W-61, mientras que las características bioquímicas de las otras dos enzimas
homólogas citadas no se han descrito.
LicA muestra mayor actividad sobre β-glucanos de cebada que sobre liquenano,
probablemente debido a la diferencia en la proporción de enlaces glicosídicos β-1,3; β-1,4 y
su distribución. Tanto el liquenano como el β-glucano de cebada están formados
principalmente por unidades celotriosil y celotetraosil, unidas por enlaces glicosídicos β-1,3,
pero la proporción de los enlaces β-1,3 y β-1,4 es 1:2 para el liquenano y 1:2,3-2,5 para el β-
glucano de cebada. Es posible que el mayor número de enlaces glucosídicos β-1,4 en el β-
glucano de cebada sea causante de la mayor actividad del enzima clonada (Picart et al. 2012).
El modo de acción enzimática se estudió mediante cromatografía en capa fina (TLC)
utilizando liquenano y β-glucano de cebada como sustratos. Como resultado se obtuvo una
mezcla de productos, incluyendo entre los mayoritarios tetra y pentasacáridos, que migraron
en las placas cromatográficas ligeramente por delante de los marcadores de celotetraosa y
celopentaosa. La movilidad de estos productos liberados por LicA indica que contiene enlaces

21
β-1,3, hecho que sugiere que la enzima puede cortar en enlaces β-1,4 adyacentes a enlaces β-
1,3. Esto indica su elevada actividad específica sobre β-glucano de cebada y liquenano.
(Jirawan. 2006)
En cuanto a la termoestabilidad de la enzima, se mostró que la enzima es estable a 55ºC y pH
6,5 como mínimo 3 horas, inactivándose rápidamente a 65ºC. En base a esto, la enzima no
plantearía problemas a la hora de su mantenimiento y almacenamiento. Sin embargo dada su
baja actividad y estabilidad a temperaturas altas, la enzima no soportaría las altas
temperaturas de algunos procesos industriales, de manera que para su utilización en este tipo
de procesos biotecnológicos podría ser interesante someterla a ensayos de evolución dirigida
para incrementar su termoestabilidad. Recientemente se ha descrito una liquenasa fúngica de
Rhizomucor miehei (Yanbin et al. 2012) que al contrario que LicA muestra alta actividad a
65ºC y a pH superiores a 8,0, características que permiten su uso en procesos industriales que
se realizan en condiciones extremas.
La alta actividad de LicA sobre β–glucano de cebada posibilita su ensayo de aplicación en la
industria cervecera, donde las β-1,3-1,4-glucanasas se aplican en el proceso de clarificación
para facilitar dicho proceso y aumentar el rendimiento (Sánchez et al. 2003). Sin embargo, la
explotación comercial de la liquenasa requiere mejorar la producción de la enzima
recombinante para minimizar los costos de producción a precios competitivos. El aislamiento
del gen codificante de la misma posibilita su clonación en huéspedes diferentes de las cepas
de E. coli utilizadas en el presente trabajo, que permitan su expresión y/o secreción
aumentada con el fin de poder producirla industrialmente para evaluar su aplicación en la
industria cervecera y ponerlo a prueba en otras aplicaciones biotecnológicas.

22
CONCLUSIONES
- Se ha identificado, clonado y caracterizado la liquenasa LicA de Paenibacillus
barcinonensis.
- LicA es una enzima homóloga a las liquenasas de la familia 16 de glicosil hidrolasas,
presentando alta identidad con liquenasas bacterianas
- La liquenasa LicA presenta máxima actividad sobre β-glucano de cebada
- La condiciones óptimas de actividad de LicA son 55ºC y pH 6,5
- Los principales productos de hidrólisis producidos por la enzima a partir del β–
glucano de cebada y del liquenano son oligosacáridos cortos, incluyendo
oligosacáridos con enlaces mixtos β-1,3-1,4

23
BIBLIOGRAFÍA
Blanco A., Diaz P., Zueco J., Parascandola P. and Pastor, F.I.J. (1999) A multidomain
xylanase from Bacillus sp. with a region homologous to thermostabilizing domains of
thermophilic enzymes. Microbiology 145, 2163-2171.
Blanco A. and Pastor F.I.J. (1993). Characterization of cellulase-free xylanases from the
newly isolated Bacillus sp. strain BP-23. Can. J. Microbiol. 39:1162–1166.
Blanco A., Vidal T., Colom J.F. and Pastor F.I. (1995) Purification and properties of
xylanase A from alkali-tolerant Bacillus sp. strain BP-23. Appl Environ Microbiol.
Dec;61(12):4468-70.
Brunswick Pamela, Manners David and Rogerstark J. (1990) Degradation of Isolated
Barley Endosperm Cell Walls by Purified Endo-(1--+ 3)(1--+4)-p-n-glucanasesand Malt
Extracts. Journal of Cereal Science 7: 153-168
Da Teng . Jian-hua Wang . Ying Fan . Ya-lin Yang. Zi-gang Tian . Jin Luo . Guan-pin
Yang . Fan Zhang (2006) Cloning of β-1,3-1,4-glucanase gene from Bacillus licheniformis
EGW039 (CGMCC 0635) and its expression in Escherichia coli BL21 (DE3). Appl Microbiol
Biotechnol 72: 705–712
Gallardo O., Diaz P. and Pastor F.I.J. (2003) Characterization of a Paenibacillus cell-
associated xylanase with high activity on aryl-xylosides: a new subclass of family 10
xylanases. Appl Microbiol Biotechnol. May;61(3):226-33.
Gosalbes M., Pérez – González J., Gonzáles R. and Navarro. (1991) Two β – Glycanase
Genes are Clustered in Bacillus polymyxa: Molecular Cloning, Expression, and Sequence
Alalysis of Genes Encoding a Xylanasa and an Endo – β – (1,3) – (1,4) – Glucanase. J.
Bacteriology. P. 7705 – 7710.
Jirawan Apiraksakorn (2006) Characterization of β-1,3-1,4-glucanase from Bacillus subtilis
GN156 for silage fermentation. Kasetsart University

24
Mutsumi Fukuda, Seiji Watanabe, Shigeki Yoshida, Hiroya Itoh, Yoshifumi Itoh,
Yoshiyuki Kamio, and Jun Kaneko (2010) Cell Surface Xylanases of the Glycoside
Hydrolase Family 10 Are Essential for Xylan Utilization by Paenibacillus sp. W-61 as
Generators of Xylo-Oligosaccharide Inducers for the Xylanase Genes. Journal of
bacteriology, p. 2210–2219
Planas Antoni (2000) Bacterial 1,3-1,4-L-glucanases: structure, function and protein
engineering. Biochimica et Biophysica Acta 1543 361-382
Pekka Mark, Martin J. Baumann, Jens M. Eklo, Fredrika Gullfot, Gurvan Michel, Asa
M. Kallas, Tuula T. Teeri, Harry Brumer, and Mirjam Czjzek. (2009) Analysis of
nasturtium TmNXG1 complexes by crystallography and molecular dynamics provides
detailed insight into substrate recognition by family GH16 xyloglucan endo-transglycosylases
and endo-hydrolases. Proteins; 75:820–836.
Picart Pere, Goedegebuur Frits, Díaz Pilar, and Pastor F. I. Javier (2012) Expression of
novel b-glucanase Cel12A from Stachybotrys atra in bacterial and fungal hosts. fungal
biology 116: 443 – 451.
Rodríguez V. Pérez R., (2002) Nuevo método para la producción de liquenasas en bacterias
aislando los genes directamente de muestras naturales. Organismo de la Propiedad Intelectual.
Sánchez, M., Fritze, D., Spröer, C., Tindall, B., Schumann, P., Koppenstedt, R.M., díaz,
P. and Pastor, F.I.J. (2003) Paenibacillus barcinonensis sp. Nov., a polysaccharide
hydrolyzing bacterium isolated from a rice field in Ebro river`s delta.
Shaoqing Y., Yan Q., Zhengqiang J., Guangsen F. And Lan W. (2008) Biochemical
Characterization of a Novel Thermostable _-1,3-1,4-Glucanase (Lichenase) from
Paecilomyces thermophile. J. Agricultural and Food Chemistry 56: 5345-5351.

25
Stephen d. Baird,Douglas a. Johnson,' and Verner l. Seligy (1990) Molecular Cloning,
Expression, and Characterization of Endo-3-1,4-Glucanase Genes from Bacillus polymyxa
and Bacillus circulans J. Bacteriology. P. 1576 – 1586
Takumi Takeda & Machiko Takahashi & Tsugumi Nakanishi-Masuno & Yuki Nakano
& Hiromasa Saitoh & Akiko Hirabuchi & Shizuko Fujisawa & Ryohei Terauchi (2010)
Characterization of endo-1,3–1,4-β-glucanases in GH family 12 from Magnaporthe oryzae.
Appl Microbiol Biotechnol 88:1113–1123
Valenzuela Susana v., Díaz Pilar, and Pastor F. I. Javier (2010) Recombinant Expression
of an Alkali Stable GH10 Xylanase from Paenibacillus barcinonensis. J. Agric. Food Chem.
2010, 58, 4814–4818
Yanbin T., Shaoqing Y., Qiaojuan Y., Peng Z., Jian C., and Zhengqiang J., (2012)
Purification and Characterization of a Novel β-1,3−1,4-Glucanase (Lichenase) from
Thermophilic Rhizomucor miehei with High Specific Activity and Its Gene Sequence J.
Agricultural and Food Chemistry 60: 2354-2361
Yang, P., Yao, B., Wang, Y., Yang, H. and Shi, P. (2006) Paenibacillus sp. endo-glucanase
gene












