
Characteristics of the T-dependent α(1 → 6) glucosyl (dextran) antibody response induced in mice...
4
916 C. Moreno, C. Hale, R. Hewett and D. Cussell Eur. J. Immunol. 1979.9: 916-919 Carlos Moreno, Christine Hale, Characteristics of the T-dependent a( 1-6) glucosyl (dextran) antibody response induced in mice with Rosemary Hewett and Deborah Cussell Department of Experimental isomaltohexaose coupled to chicken y-globulin Immunobiology, The Wellcome Research Laboratories, Beckenham Isomaltohexaose flavazole coupled to chicken y-globulin (IM6-CGG) induced T cell- dependent anti-a( 1+6) dextran-specific IgM and IgG responses in CBA, BALBlc and A strain mice. The IgG responses were of restricted heterogeneity as judged by isoelectric focusing, and belonged mostly to the IgGl subclass with a minor IgG3 component in the case of BALB/c and CBA mice. All four subclasses of IgG were produced in A strain mice. In contrast, native dextran B 512 induces exclusively T cell-independent IgM responses of the same specificity for all 3 strains. All BALBlc mice immunized with different doses of IM6-CGG either in Freund’s adjuvant or with Al(OH)3 plus Bordetella pertussis showed the same spectrotypes by isoelectric focusing. Sera obtained at different times after immunization of BALBic mice failed to show spectrotype variations. Late, but not early, bleedings from CBA mice showed a tendency towards more uniform isoelectric focusing patterns. 1 Introduction Detailed studies on the correlation between structure and function of antibodies are generally complicated by the heterogeneity of the immune response. Antigenic determi- nants capable of eliciting restricted responses are therefore of great help, and in this context the anti-a(1+3) dextran response of BALB/c mice has proved rewarding for the follow- ing reasons. (a) It is restricted in the sense that in an individual mouse only a few clones are stimulated [l, 21; (b) it is uniform in the sense that individual mice differ very little from each other in terms of spectrotype and idiotyp- ic determinants [I, 31; and (c) close correlation exists between the N-terminal amino acid sequences of the antibody induced by immunization and the corresponding myelomas showing specificity for a(1+3) dextran [4]. Since mice also respond to the a( 1+6)-linked glucosyl deter- minant of dextran [5], we have compared the responses to this epitope in BALB/c and other strains of mice, to evaluate the extent to which the a(l+6) and a(l+3) responses resemble one another. To evaluate T-dependent IgG responses by isoelectric focusing (IEF) pattern and changes in them accord- ing to mouse strains and immunization schedule, we have used an isomaltohexaose (IM 6)- protein conjugate instead of dex- tran as immunogen. An oligosaccharide hapten coupled to a protein carrier can induce antibodies that cross-react with the polysaccharide from which the oligosaccharide was obtained [6]. The coupling of oligosaccharides derived from purified dextrans which behave as thymus-independent antigens in the mouse [7, 81 should stimulate a T-dependent response with [I 24731 Correspondence: Carlos Moreno, Department of Experimental Immunobiology, Wellcome Research Laboratories, Beckenham, Kent BR3 3BS, GB Abbreviations: ABC: Antigen-ill riding capacity CGG: Chicken y- globulin IEF Isoelectric focu\ing IM6: Isomaltohexaose IM6- CGG: Isomaltohexaose flavazole azo-derivative conjugated with CGG PBS: Phosphate-buffered saline, pH 7.2 PFC: Plaque-form- ing cells antibodies cross-reacting with native dextran, provided the coupling procedure has preserved the immunodominant com- ponent of the hapten. Rabbit anti-a( 1+6) dextran antibodies have been obtained by coupling oligosaccharides of the isomaltosyl series to serum albumin using a mixed anhydride reaction [9] and, more recently, the flavazole derivatives of the same oligosaccharides [lo]. The advantage of both methods is that they couple only the reducing end of the oligosaccharide leaving the nonreducing, commonly immunodominant end intact [ll-131. We have used the 1-(m-aminophenyl) flavazole derivative of IM6 coupled to chicken y-globulin (IM6-CGG) [lo] and examined the nature of the murine immune response with special reference to antibody subclass and spectrotype characterizations. 2 Materials and methods 2.1 Materials Inbred strains BALB/c and CBA-T6T6 were obtained from our own colony. Strain A mice were purchased from Olac Ltd., Bicester, GB. Mice were 8-10 weeks old at the onset of the experiments. Rabbit anti-mouse Ig antisera were obtained from Miles Laboratories, Rehovot, Israel. Anti-Thy-1.2 anti- serum was a generous gift from Dr. R. Bomford, The Well- come Research Laboratories, Beckenham. A preparation of IM6-CGG which contained 17 isomaltohexaose residues/ molecule of CGG was kindly provided by Dr. N. A. Mitchi- son, Department of Zoology, University College, London. Almost linear a(l-+6)-linked native dextran B 512 has been described previously [5]. Fluorescein isothiocyanate (F1TC)- treated dextran T-20 (8 FITC residues/103 glycosyl units) was purchased from Pharmacia, Uppsala, Sweden. Iodination of this polysaccharide was done by the chloramine-T method ~41. 2.2 Plaque-forming cells (PFC) Determination of a( 1+6)-specific direct PFC, using sheep erythrocytes sensitized with stearoyl-dextran (T-70), was done as described previously [5, 151. 0 Verlag Chemie, GmbH, D-6940 Weinheim, 1979 0014-2980/79/1111-0916$02.50/0
-
Upload
carlos-moreno -
Category
Documents
-
view
214 -
download
2
Transcript of Characteristics of the T-dependent α(1 → 6) glucosyl (dextran) antibody response induced in mice...

916 C. Moreno, C. Hale, R. Hewett and D. Cussell Eur. J . Immunol. 1979.9: 916-919
Carlos Moreno, Christine Hale, Characteristics of the T-dependent a( 1-6) glucosyl (dextran) antibody response induced in mice with Rosemary Hewett and Deborah Cussell
Department of Experimental isomaltohexaose coupled to chicken y-globulin Immunobiology, The Wellcome Research Laboratories, Beckenham
Isomaltohexaose flavazole coupled to chicken y-globulin (IM6-CGG) induced T cell- dependent anti-a( 1+6) dextran-specific IgM and IgG responses in CBA, BALBlc and A strain mice. The IgG responses were of restricted heterogeneity as judged by isoelectric focusing, and belonged mostly to the IgGl subclass with a minor IgG3 component in the case of BALB/c and CBA mice. All four subclasses of IgG were produced in A strain mice. In contrast, native dextran B 512 induces exclusively T cell-independent IgM responses of the same specificity for all 3 strains. All BALBlc mice immunized with different doses of IM6-CGG either in Freund’s adjuvant or with Al(OH)3 plus Bordetella pertussis showed the same spectrotypes by isoelectric focusing. Sera obtained at different times after immunization of BALBic mice failed to show spectrotype variations. Late, but not early, bleedings from CBA mice showed a tendency towards more uniform isoelectric focusing patterns.
1 Introduction
Detailed studies on the correlation between structure and function of antibodies are generally complicated by the heterogeneity of the immune response. Antigenic determi- nants capable of eliciting restricted responses are therefore of great help, and in this context the anti-a(1+3) dextran response of BALB/c mice has proved rewarding for the follow- ing reasons. (a) It is restricted in the sense that in an individual mouse only
a few clones are stimulated [l, 21; (b) it is uniform in the sense that individual mice differ very
little from each other in terms of spectrotype and idiotyp- ic determinants [I, 31; and
(c) close correlation exists between the N-terminal amino acid sequences of the antibody induced by immunization and the corresponding myelomas showing specificity for a(1+3) dextran [4].
Since mice also respond to the a( 1+6)-linked glucosyl deter- minant of dextran [5], we have compared the responses to this epitope in BALB/c and other strains of mice, to evaluate the extent to which the a(l+6) and a(l+3) responses resemble one another. To evaluate T-dependent IgG responses by isoelectric focusing (IEF) pattern and changes in them accord- ing to mouse strains and immunization schedule, we have used an isomaltohexaose (IM 6)- protein conjugate instead of dex- tran as immunogen. An oligosaccharide hapten coupled to a protein carrier can induce antibodies that cross-react with the polysaccharide from which the oligosaccharide was obtained [6]. The coupling of oligosaccharides derived from purified dextrans which behave as thymus-independent antigens in the mouse [7, 81 should stimulate a T-dependent response with
[I 24731
Correspondence: Carlos Moreno, Department of Experimental Immunobiology, Wellcome Research Laboratories, Beckenham, Kent BR3 3BS, GB
Abbreviations: ABC: Antigen-ill riding capacity CGG: Chicken y- globulin IEF Isoelectric focu\ing IM6: Isomaltohexaose IM6- CGG: Isomaltohexaose flavazole azo-derivative conjugated with CGG PBS: Phosphate-buffered saline, pH 7.2 PFC: Plaque-form- ing cells
antibodies cross-reacting with native dextran, provided the coupling procedure has preserved the immunodominant com- ponent of the hapten. Rabbit anti-a( 1+6) dextran antibodies have been obtained by coupling oligosaccharides of the isomaltosyl series to serum albumin using a mixed anhydride reaction [9] and, more recently, the flavazole derivatives of the same oligosaccharides [lo]. The advantage of both methods is that they couple only the reducing end of the oligosaccharide leaving the nonreducing, commonly immunodominant end intact [ll-131. We have used the 1-(m-aminophenyl) flavazole derivative of IM6 coupled to chicken y-globulin (IM6-CGG) [lo] and examined the nature of the murine immune response with special reference to antibody subclass and spectrotype characterizations.
2 Materials and methods
2.1 Materials
Inbred strains BALB/c and CBA-T6T6 were obtained from our own colony. Strain A mice were purchased from Olac Ltd., Bicester, GB. Mice were 8-10 weeks old at the onset of the experiments. Rabbit anti-mouse Ig antisera were obtained from Miles Laboratories, Rehovot, Israel. Anti-Thy-1.2 anti- serum was a generous gift from Dr. R. Bomford, The Well- come Research Laboratories, Beckenham. A preparation of IM6-CGG which contained 17 isomaltohexaose residues/ molecule of CGG was kindly provided by Dr. N. A. Mitchi- son, Department of Zoology, University College, London. Almost linear a(l-+6)-linked native dextran B 512 has been described previously [5]. Fluorescein isothiocyanate (F1TC)- treated dextran T-20 (8 FITC residues/103 glycosyl units) was purchased from Pharmacia, Uppsala, Sweden. Iodination of this polysaccharide was done by the chloramine-T method ~ 4 1 .
2.2 Plaque-forming cells (PFC)
Determination of a( 1+6)-specific direct PFC, using sheep erythrocytes sensitized with stearoyl-dextran (T-70), was done as described previously [5, 151.
0 Verlag Chemie, GmbH, D-6940 Weinheim, 1979 0014-2980/79/1111-0916$02.50/0
Eur. J. Immunol. 1979. 9: 916-919 T-dependent anti-dextran response in mice 917
2.3 Antigen-binding capacity (ABC)
Dextran-binding capacity was estimated by incubating 25 pl of serum diluted 1 : 20 in phosphate-buffered saline (PBS) with 25 pl '251-labeled dextran (1.86 ygiml) for 1 day at 4°C. The IgG was then precipitated with a dilution of anti-mouse Ig found to be optimal by direct immunoprecipitation with mouse IgG. The mixture was left for 4 days at 4°C and the precipi- tates counted. ABC is expressed as pg of dextran bound/ml of serum after subtracting the amount of radioactivity unspecifi- cally bound by dextran-absorbed mouse sera. A binding capacity of 10 pl antigedml of serum was found experimen- tally to correspond to approximately 150 pg of precipitating antibodyiml.
2.4 IEF
The IEF technique described by Awdeh et al. [16] and Wil- liamson [17] was modified for this system. Ampholine, pH 3.5-10 (LKB, Bromma, Sweden), was used in 5% polyacryl- amide gels layered on 21 x 16 cm glass plates coated with gelatin (Ilford, Essex, GB). The samples were run for 18 h at 300 V per plate at 4"C, and the last 3 h at 500 V. After elec- trophoresis, the gel was immersed in 18% Na2S04 for 1 h and then transferred to 18% Na2S04 containing 1 pglml lz5I- labeled dextran (= lo6 cpmiml) and incubated for 2 h. After this incubation, the gel was kept for 1 h in a solution of 0.1% glutaraldehyde in 18% Na2S04 and then washed with tap water. After 15 min in 30% ethanol, the gel was dried and exposed to X-ray film (Osray M 3 Agfa-Gevaert, Belgium).
2.5 Radioimmunodiffusion and radioimmunoelectrophoresis
Microscope slides were prepared with 3 ml of 1% agarose in PBS. The centre well contained anti-mouse Ig-specific anti- serum mixed with radioiodinated dextran ( lo6 cpmiml). The outside wells were filled with the test serum serially diluted with normal mouse serum. After 2 days at room temperature, the slides were washed, dried, exposed to X-ray film and
stained. After immunoelectrophoresis of the test serum, the plates were developed with 150 pl of anti-mouse Ig subclass- specific antisera to which '2'I-labeled dextran (1 pgiml = lo6 cpm/ml) was added. Slides were incubated at room temper- ature for 2 days, washed, dried and exposed to X-ray film.
2.6 Immunization procedures
Three basic schemes were used. (a) Purified dextran B 512 was given in a single intravenous (i.v.) dose resulting in a T-inde- pendent IgM response. (b) IM6-CGG, emulsified 1 : 1 in com- plete Freund's adjuvant was given subcutaneously (s.c.) (0.2 ml), followed 7 days later by a second intraperitoneal (i.p.) injection of the same antigen in incomplete Freund's adjuvant. A third i.v. injection of soluble antigen was given on day 14. (c) As for scheme (b), omitting Freund's adjuvant. Instead, the first and second doses of antigen were given with 1 mg of AI(OH)3 and injected i.p. with Bordetella pertussis vaccine ( lo9 organismsidose, Burroughs Wellcome). The third dose of antigen was given i.v. without adjuvant, as in scheme (b).
3 Results
3.1 Immune response of different strains
BALB/c, CBA and A mice were immunized as described in Sect. 2.6 using IM6-CGG in Freund's adjuvant or dextran B512. Five days after the last injection, serum antibody levels (IgG and IgM) and direct PFC (IgM) specific for a(1-6) glucosyl dextran epitopes were determined (Table 1). All three strains responded well to dextran B 512 synthesizing IgM antibodies, but only low ABC was detected in the serum. These antibodies were later found to be only IgM by radioim- munodiffusion and radioimmunoelectrophoresis. On the other hand, IM6-CGG induced strong IgM and IgG responses. Although only data pertaining to 10 pg IM6-CGG are shown, these values did not differ significantly from those obtained
Table 1. Comparison of anti-dextran responses induced by immunization with IM6-CGG and dextran B512")
BALB/c IM 6-CGG Dextran
None
Dextran None
Dextran None
CBA IM 6-CGG
A IM 6-CGG
65 OOO (1.09) 3 1 000 ( I S6)
120 ( I . 15) 18700 (1.43) 37 600 ( I .28)
340(1.52)
15 800 (1.34) 9500 (1.24)
320 (1.20)
a) Immunization schemes are described in Sect. 2.6. The values correspond to the 5th day after i.v. injection of the immunogen and are expressed as geometric means of 5 mice/group with standard error in parenthesis. The data given correspond to results obtained with 10 pg IM6-CGG and 1 pg dextran B 512.
b) pg dextran boundhl of serum. c) Qualitative test by radioimmunoelectrophoresis of sera selected at random. Data in parenthesis indicate that these antibodies were not found
in all sera examined.
918 C. Moreno, C. Hale, R. Hewett and D. Cussell Eur. J. Immunol. 1979.9: 916-919
subclasses. While some A mice produced anti-a( 1-6) glucosyl of all IgG subclasses (although only trace amounts of IgG*b), others showed IgG3 antibodies only. The predominant light chain for BALB/c antibodies was found to be lambda.
3.2 IEF
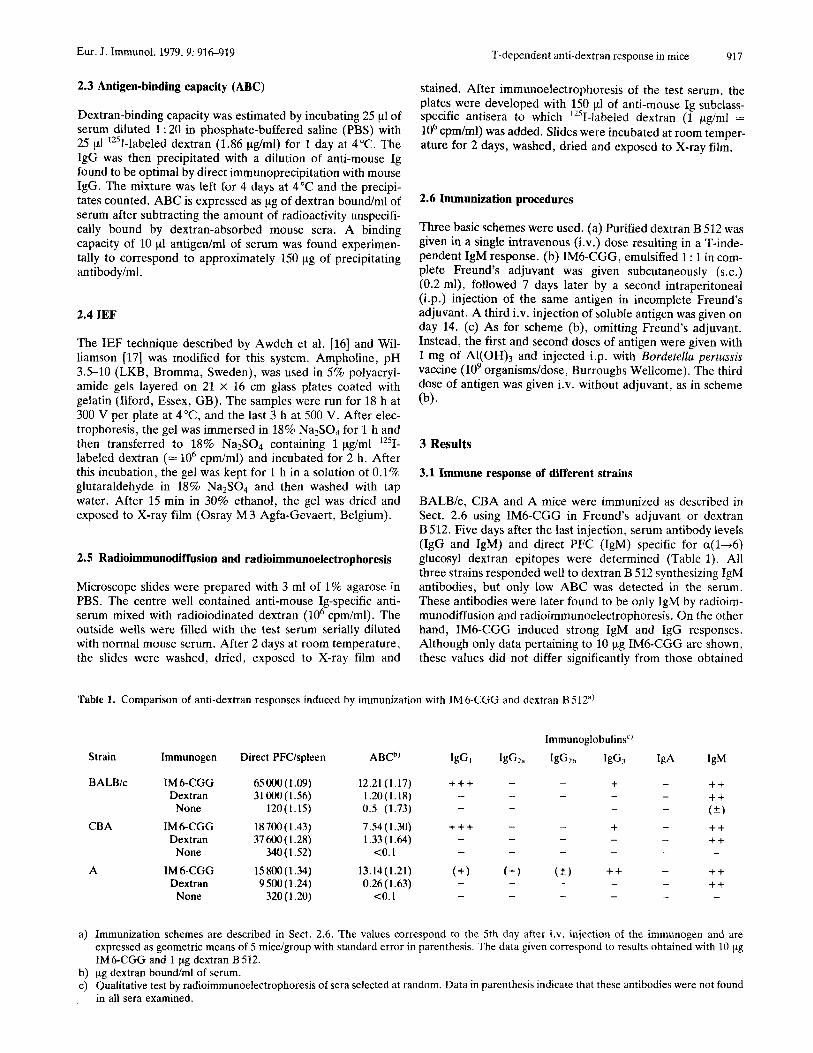
The IEF patterns of the 6-day responses (day 20 after priming) to 10 pg of IM6-CGG in Freund’s adjuvant for strains A, BALB/c and CBA are presented in Fig. 1. While the patterns for all three strains are relatively restricted, only in BALB/c mice is it uniform and constant in all animals tested. Apart from quantitative differences, the serum spectrotypes are practically indistinguishable. Strain A mice present a very restricted response with only occasional sharing of spectro- types between sera, while sometimes two sera are almost identical ( e g . nos. 3 and 4 in Fig. 1). Not a single mouse among 45 immunized with dextran B512 showed a positive band by IEF.
Figure 1. IEF pattern of sera from mice immunized with 10 pg IM6- CGG in Freund’s adjuvant. Strain A, day 20: 1 to 4; BALBk, day 20: 5 to 8; CBA, day 20: 9 to 12 and CBA, day 36: 13 to 15. Plates were developed with ‘ZSI-labeled dextran.
3.3 The response to IMQ-CGG is T-dependent
lI2L73.2) Days After Prirmng
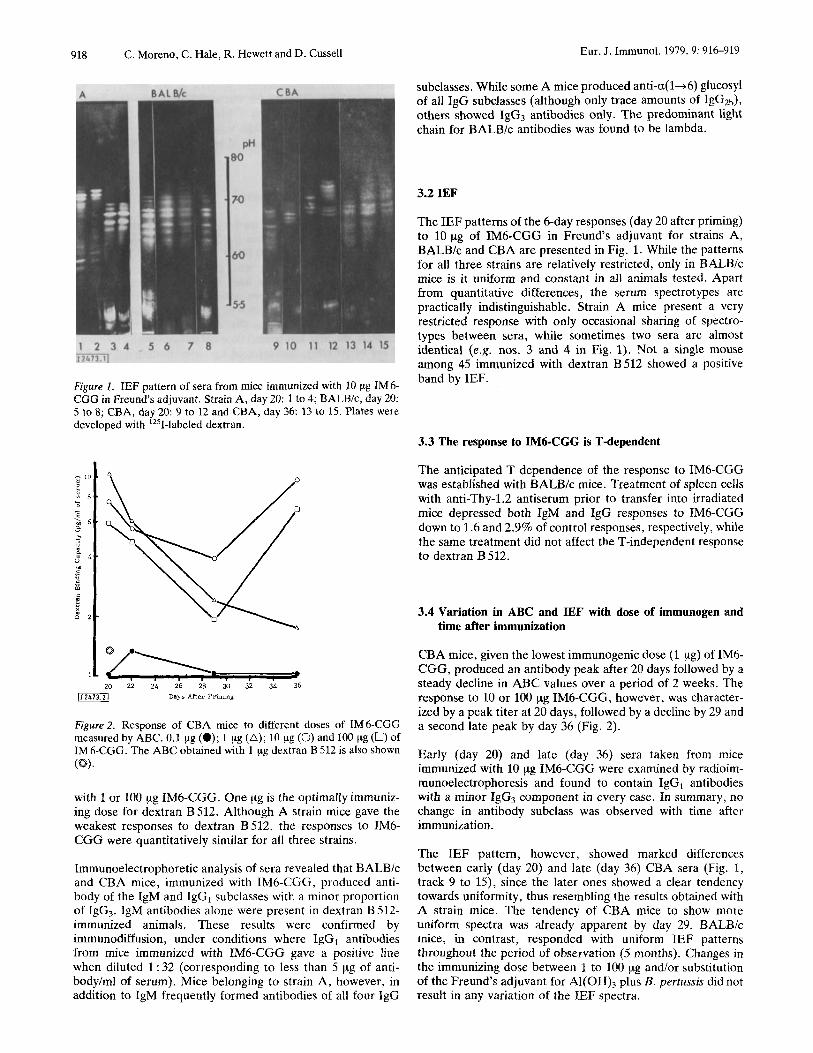
Figure 2. Response of CBA mice to different doses of IM6-CGG measured by ABC. 0.1 pg (0); 1 pg (A); 10 pg (0) and 100 pg (0) of IM 6-CGG. The ABC obtained with 1 pg dextran B 512 is also shown IS).
with 1 or 100 pg IM6-CGG. One pg is the optimally immuniz- ing dose for dextran B 512. Although A strain mice gave the weakest responses to dextran B512, the responses to IM6- CGG were quantitatively similar for all three strains.
Immunoelectrophoretic analysis of sera revealed that BALB/c and CBA mice, immunized with IM6-CGG, produced anti- body of the IgM and IgG, subclasses with a minor proportion of IgG,. IgM antibodies alone were present in dextran B 512- immunized animals. These results were confirmed by immunodiffusion, under conditions where IgGl antibodies from mice immunized with IM6-CGG gave a positive line when diluted 1 : 32 (corresponding to less than 5 pg of anti- body/ml of serum). Mice belonging to strain A, however, in addition to IgM frequently formed antibodies of all four IgG
The anticipated T dependence of the response to IM6-CGG was established with BALBk mice. Treatment of spleen cells with anti-Thy-1.2 antiserum prior to transfer into irradiated mice depressed both IgM and IgG responses to IM6-CGG down to 1.6 and 2.9% of control responses, respectively, while the same treatment did not affect the T-independent response to dextran B512.
3.4 Variation in ABC and IEF with dose of immunogen and time after immunization
CBA mice, given the lowest immunogenic dose (1 pg) of IM6- CGG, produced an antibody peak after 20 days followed by a steady decline in ABC values over a period of 2 weeks. The response to 10 or 100 pg IM6-CGG, however, was character- ized by a peak titer at 20 days, followed by a decline by 29 and a second late peak by day 36 (Fig. 2 ) .
Early (day 20) and late (day 36) sera taken from mice immunized with 10 pg IM6-CGG were examined by radioim- munoelectrophoresis and found to contain IgG, antibodies with a minor lgG3 component in every case. In summary, no change in antibody subclass was observed with time after immunization.
The IEF pattern, however, showed marked differences between early (day 20) and late (day 36) CBA sera (Fig. 1, track 9 to 15), since the later ones showed a clear tendency towards uniformity, thus resembling the results obtained with A strain mice. The tendency of CBA mice to show more uniform spectra was already apparent by day 29. BALB/c mice, in contrast, responded with uniform IEF patterns throughout the period of observation (5 months). Changes in the immunizing dose between 1 to 100 pg and/or substitution of the Freund’s adjuvant for AI(OH)3 plus B. pertussis did not result in any variation of the IEF spectra.

Eur. J. Immunol. 1979.9: 916919 T-dependent anti-dextran response in mice 919
3.5 Immunization with purified dextran in Freund’s adjuvant
Although T-independent responses are enhanced little or not at all by adjuvants ( e .g . P. Del Guercio [18]), we wished to exclude the possibility that failure to demonstrate IgG anti- bodies after immunization with pure dextran could be due to a lack of adjuvant. To test this, BALB/c mice were immunized with doses of 0.1 to 100 pg of dextran B 512 in Freund’s adju- vant following the schemes described for IM6-CGG. Although some of the sera obtained gave low (but detectable) ABC, no mice showed ABC values higher than controls receiving adju- vant alone. Furthermore, the antisera with the highest ABC failed to show any bands in IEF and gave no IgG lines by radioimmunodiffusion, although IgM antibodies were clearly detected by the latter method. Observations were made from 4 to 14 days after rechallenge (day 20 and 30 after priming).
4 Discussion
The striking uniformity in the response of BALBlc mice to IM6-CGG specific for the a(l-+6) dextran determinants, parallels the results obtained by Hansburg et al. [l] for the a(1+3) dextran epitope in the same strains. No occasional unique bands were found, even when animals were immunized with various doses in conjunction with different adjuvants and observed for prolonged periods. These results do not necessar- ily imply that all inherited clones specific for the a(l+6) deter- minant are represented by these simple spectrotypes; they could reflect only a fraction of them, but the rest are not ex- pressed under the conditions used for immunization. How- ever, they indicate a property of the immune system of BALB/ c mice that distinguishes it from other strains, since homogeneity in the immune response is a regular feature in BALBlc mice [19, 201. In contrast, although CBA and A mice under the same immunization procedure gave restricted responses (in the case of CBA belonging also to the same IgGl subclass with a minor IgG3 component), the spectrotypes detected were not uniform. It is interesting that BALBic IgG responses specific for a( 1+3) belong to the IgG3 subclass [21].
Our failure to detect anti-dextran antibodies belonging to any of the IgG subclasses, in the sera of mice immunized with pure dextran (with or without adjuvant), could be the result of a poor sensitivity in our methods. Nevertheless, a quantitative analysis of the results shows that we are able to detect IgG antibodies at a concentration equivalent to 5% of the average response to IM6-CGG by radioimmunodiffusion. A conserva- tive conclusion from our results would be that dextran B 512, with or without adjuvant, is, at best, a very poor inducer of IgG antibodies in all 3 strains studied. (Analysis of the low IgM ABC activity in mice immunized with dextran B512 is complicated by the fact that unprimed mice usually showed values ranging between 0.1-0.9 pg of antigen binding/ml of serum.)
The results obtained with CBA and A strain mice deserve an additional comment. Their IgM and IgG responses to IM6- CGG are quantitatively in the same range although CBA was classified as a “high” and A strain as a “low” responder by Fernandez and Moller [22] when studying the immune response to the a(1-6) glucosyl determinant of dextran. This discrepancy cannot be explained easily, and any genetic or functional interpretation of the data available must await
further clarification of the point. However, our results clearly indicate that A strain mice cannot lack V genes specific for a(l-+6) determinants, as proposed by Fernandez and Moller P I .
CBA mice showed restricted, although not uniform, early responses which became more uniform later post immuniza- tion. These late responses belong also predominantly to the IgG1 subclass, and possibly mean that antibody-producing clones are being selected on the basis of antibody affinity. Analysis of the idiotypic determinants present in myeloma proteins and in the different populations of antibodies induced by IM6-CGG, combined with amino acid sequence in their variable chains would be a very useful complement to the information available for other haptenic systems [20], in par- ticular the a(1-+3) dextran system of BALB/c mice.
The authors wish to thank Drs. R. Smith, J . Ivanyi and J . G . Howard for critical review of the manuscript.
Received April 2, 1979; in revised form June 28, 1979.
5 References
1 Hansburg, D., Briles, D. E. and Davie, J. M., J . Immunol. 1976. 117: 569.
2 Hansburg, D., Briles, D. E. and Davie, J. M., J. Immunol. 1977. 119: 1406.
3 Blomberg, B., Gelecker, W. R. and Weigert, M., Science 1972. 177: 178.
4 Garstad, P., Hubert, J., Hunkapiller, M., Goetze, A., Schilling, J., Black, B. , Eaton, B., Richards, J., Weigert, M. and Hood, L., Eur. J . Immunol. 1978. 8: 497.
5 Howard, J. G., Vicari, G. and Courtenay, B. M., Immunology 1975. 29: 585.
6 Goebel, W. F., Avery, 0. T. and Babers, F. H., J . Exp. Med. 1934. 60: 599.
7 Basten, A. and Howard, J.G., Contemp. Top. Immunobiol. 1973. 2: 265.
8 Klaus, G. G. B., Howard, J. G. andFeldmann, M., Br. Med. Bull. 1976. 32: 141.
9 Arakatsu, Y., Ashwell, G. and Kabat, E. A,, J . Immunol. 1966. 97: 858.
10 Himmelspach, K., Westphal, 0. and Teichmann, B., Eur. J . Imrnunol. 1971. 1: 106.
11 Cisar, J., Kabat, E. A., Liao, J. and Potter, M., J . Exp. Med. 1974. 139: 159.
12 Cisar, J., Kabat, E. A., Dorner, M. M. and Liao, J., J . Exp. Med. 1975. 142: 435.
13 Kabat, E. A., Structural Concepts in lmmunology and Immunochembtry, 2nd Edition, Holt, Rinehart and Winston, New York 1976.
14 Hunter, W. M. and Greenwood, F. C . , Nature 1962. 194: 495. 15 Svensson, S. , Vicari, G. and Wilkinson, S . , 1. Immunol. Methods.
16 Awdeh, Z . L., Williamson, A. R. and Askonas, B. A., Nature
17 Williamson, A. R., Eur. J . Immunol. 1971. 1: 390. 18 Del Guercio, P., Nature-New Biol. 1972. 2.38: 213. 19 Weigert, M. and Potter, M., Immunogenefics 1977. 4: 401. 20 Weigert, M. and Riblet, R., Springer Seminars in lmmunobiology
21 Perlmutter, R. M., Hansburg, D., Briles, D. E., Nicolotti, R. A.
22 Fernandez, C. and Moller, G., J . Exp. Med. 1977. 146: 1663.
1976. 9: 315.
1968. 219: 66.
1978. I: 133.
and Davie, J. M., J . Immunol. 1978. 121: 566.

![METHODOLOGY Open Access Development and … · amination, and then separated by polyacrylamide gel electrophoresis [11]. ... of APTS labelled hydrolysed dextran and β-1,4-xylo oligosaccharides](https://static.fdocument.org/doc/165x107/5adeff457f8b9ab4688b939a/methodology-open-access-development-and-and-then-separated-by-polyacrylamide.jpg)
















