Cell Mechanosensitivity is Enabled by the LINC...
50
Gunes Uzer InspireME Seminar Series Boise State University November 6 th 2015 Cell Mechanosensitivity is Enabled by the LINC Nuclear Complex
-
Upload
nguyencong -
Category
Documents
-
view
216 -
download
0
Transcript of Cell Mechanosensitivity is Enabled by the LINC...

Gunes Uzer
InspireME Seminar SeriesBoise State University
November 6th 2015
Cell Mechanosensitivity is Enabled by the LINC Nuclear Complex
β
α
ECM
CellMembrane
Focal Adhesion
Cytoskeleton
Lamin Chromatin
NucleusNuclear Envelope
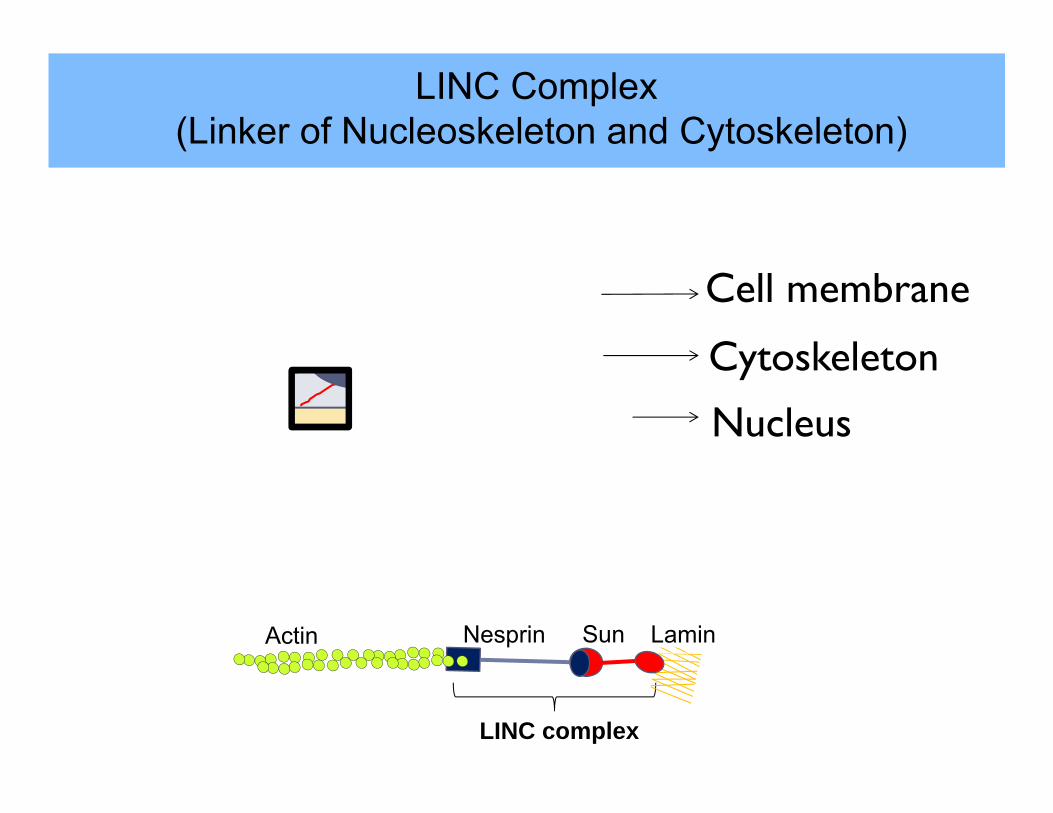
Nesprin
Sun
Cell membrane
Cytoskeleton
Nucleus
LINC complex
LaminSunNesprinActin
LINC Complex(Linker of Nucleoskeleton and Cytoskeleton)

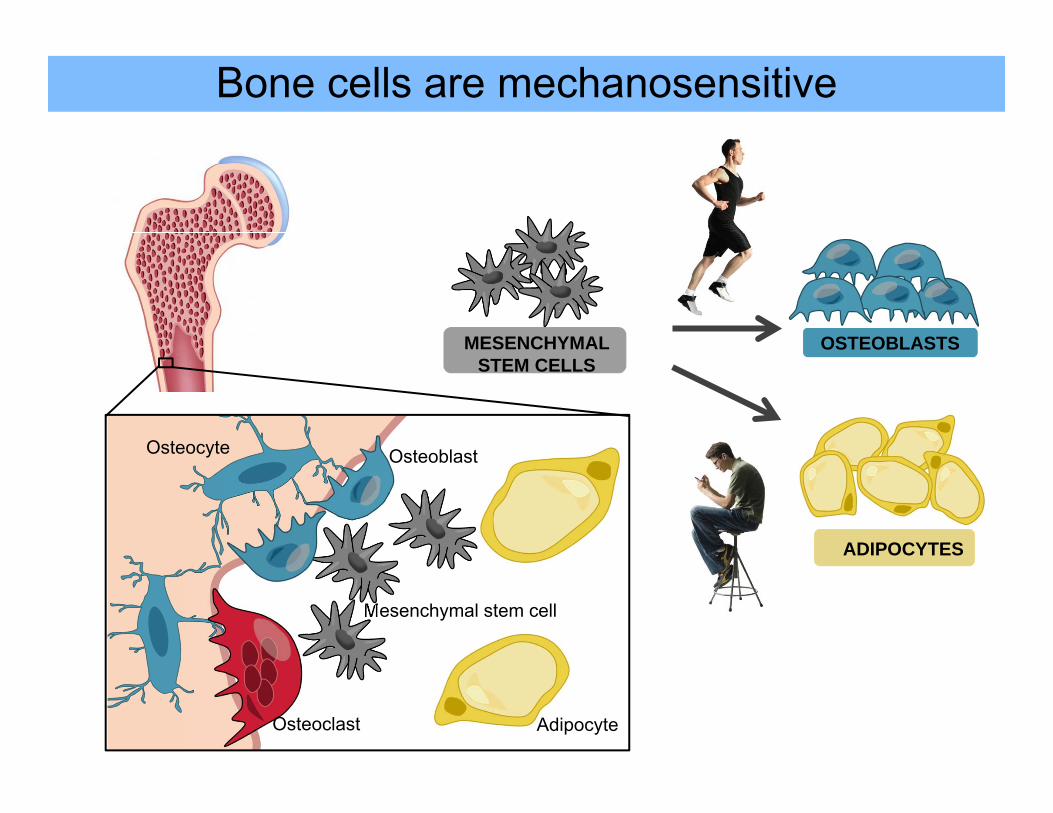
Mechanical signals regulate bone mass
Bone will adapt to the loads under which it is placed1
Playing arm of tennis players have higher bone mass2
During microgravity, astronauts can lose up to 2% hip density per month3
1. Wolff, J. SpringerVerlag,1986 Berlin. 2.Huddleston, A. L. et al. JAMA(1980) 244(10): p.1107., 2.Lang, T.et al. JBMR,2004 19(6): p.1006.
OSTEOBLASTS
ADIPOCYTES
Bone cells are mechanosensitive
MESENCHYMAL STEM CELLS
Osteocyte
Mesenchymal stem cell
Adipocyte
Osteoblast
Osteoclast
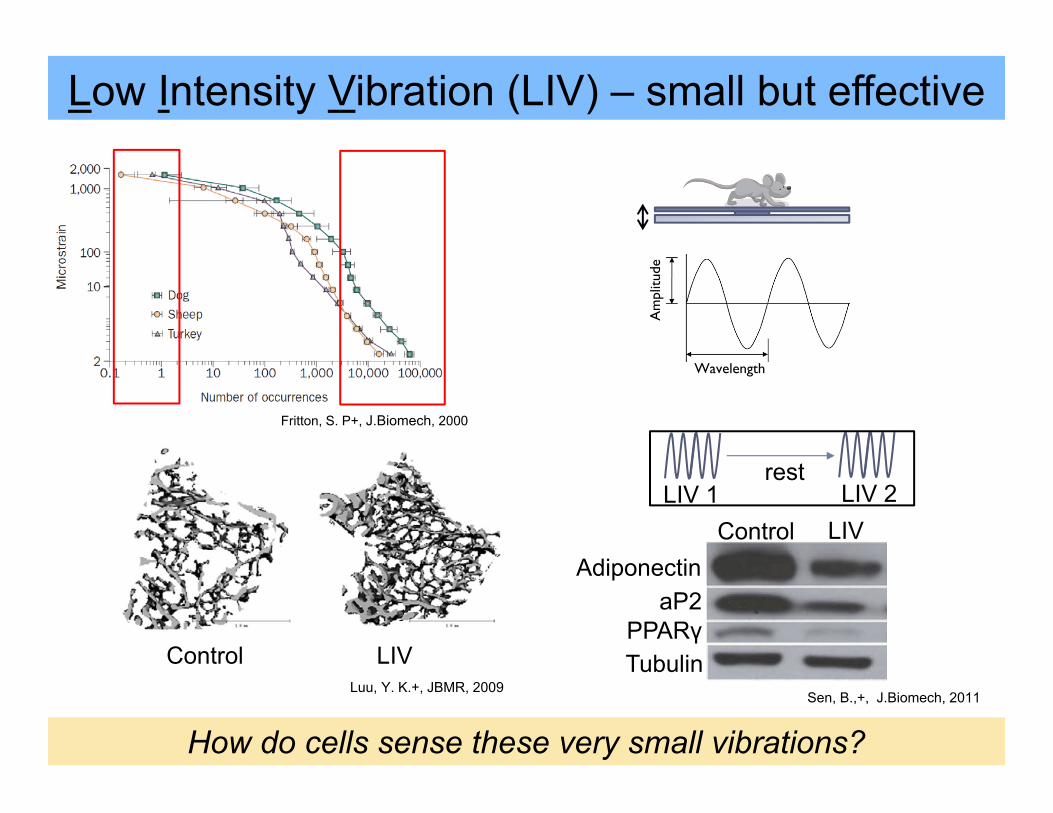
Low Intensity Vibration (LIV) – small but effective
Control LIV
How do cells sense these very small vibrations?
LIV 1 LIV 2rest
Luu, Y. K.+, JBMR, 2009Sen, B.,+, J.Biomech, 2011
Fritton, S. P+, J.Biomech, 2000
AdiponectinLIVControl
aP2PPARγTubulin
Am
plitu
de
Wavelength
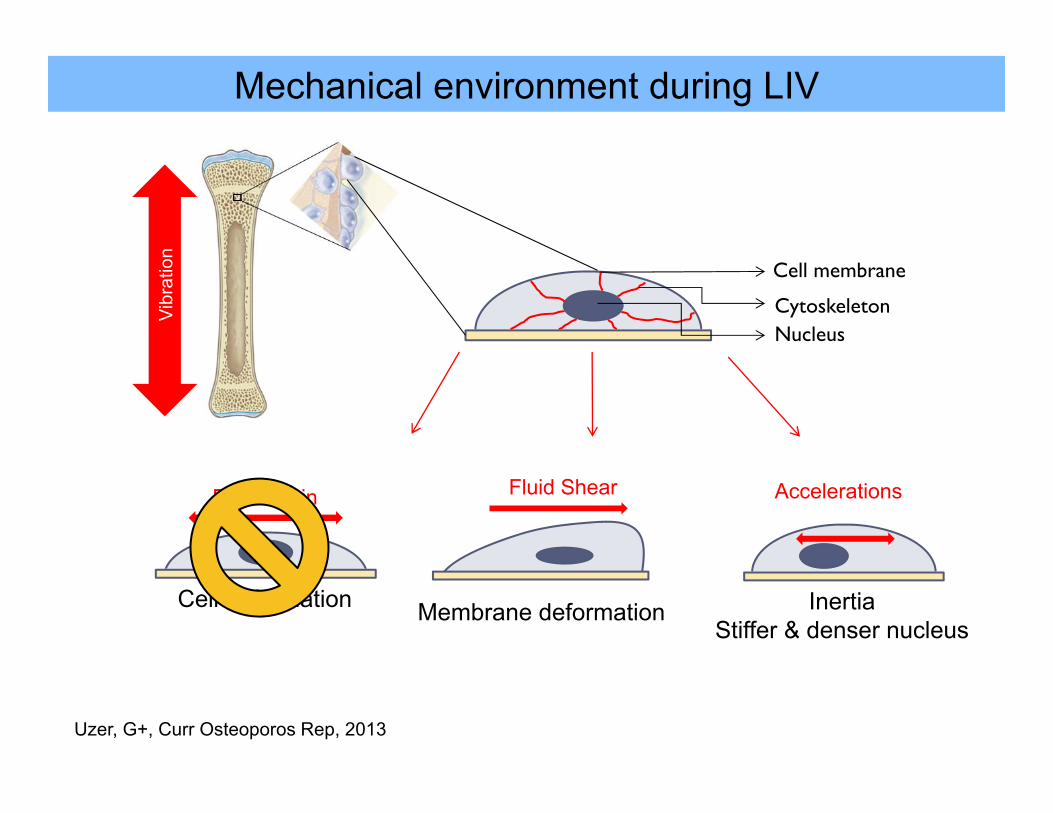
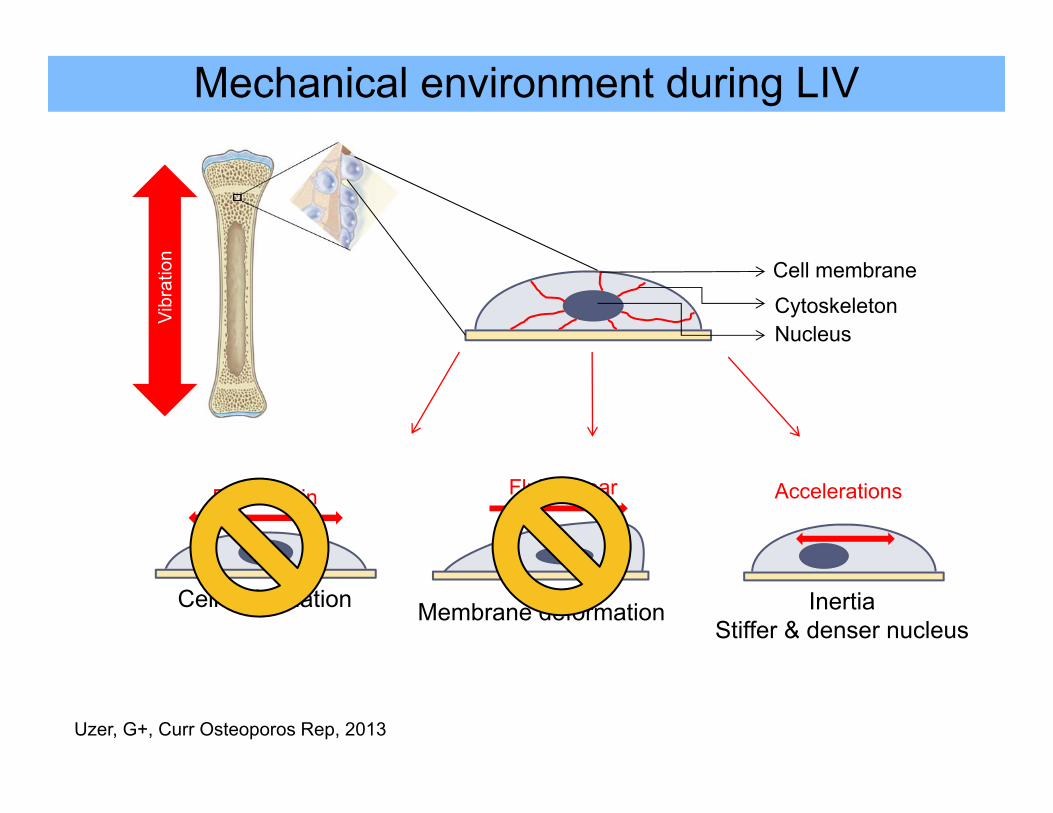
Mechanical environment during LIV
Cell membrane
CytoskeletonNucleus
Vibr
atio
n
Fluid Shear
Membrane deformation
Accelerations
InertiaStiffer & denser nucleus
ECM strain
Cell deformation
Uzer, G+, Curr Osteoporos Rep, 2013
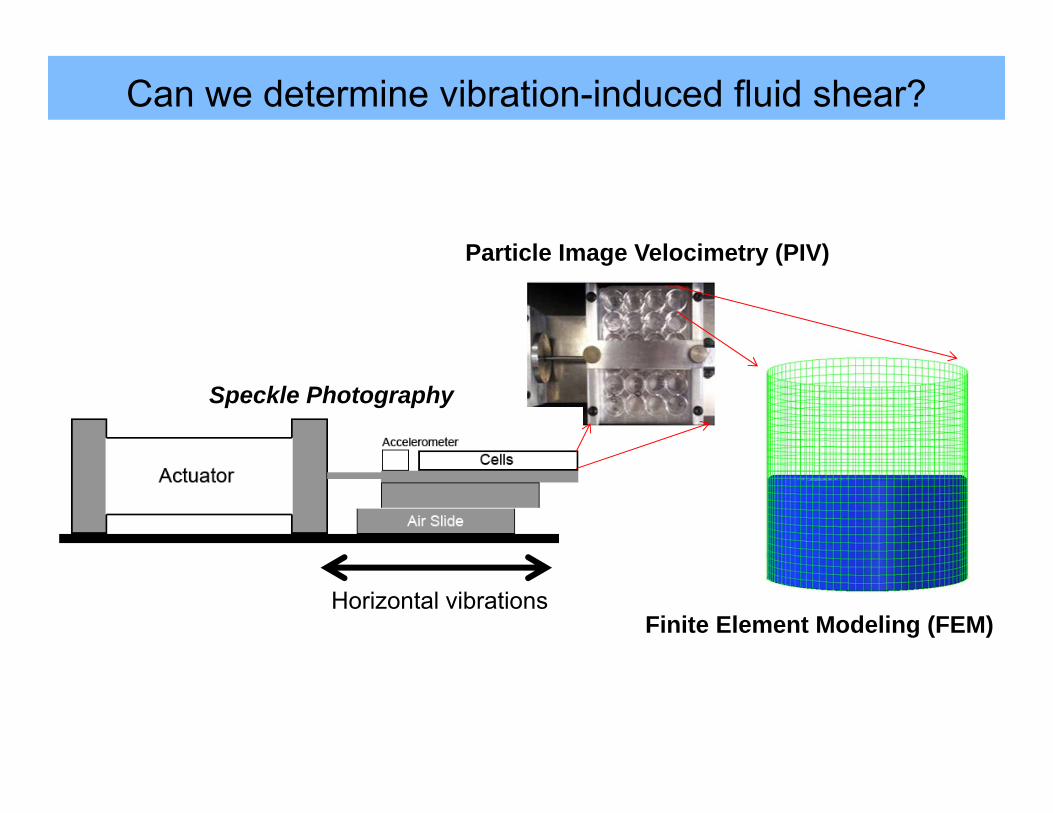
Can we determine vibration-induced fluid shear?
Horizontal vibrations
Speckle Photography
Particle Image Velocimetry (PIV)
Finite Element Modeling (FEM)
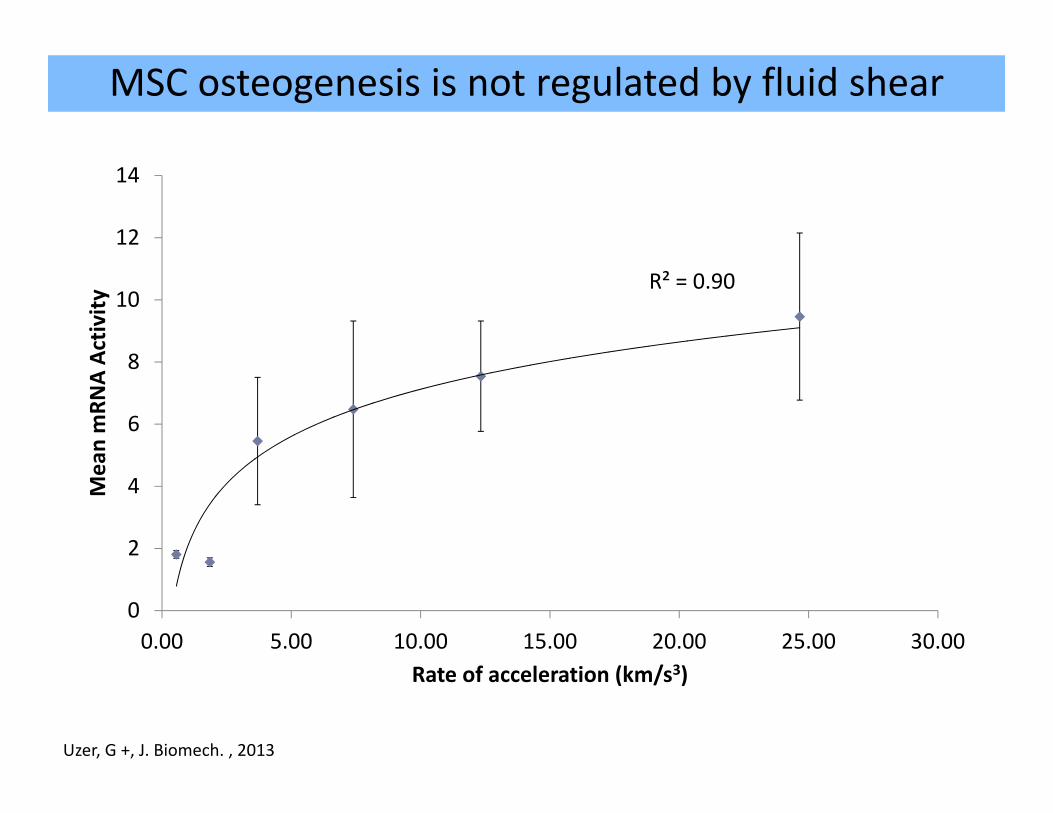
MSC osteogenesis is not regulated by fluid shear
0
0.5
1
1.5
2
2.5
3
Min
eral
izat
ion
(Aliz
arin
Red
, mM
)
**
†¥
*†
Control
LIV
R² = 0.90
0
2
4
6
8
10
12
14
0.00 5.00 10.00 15.00 20.00 25.00 30.00
Mean mRN
A Ac
tivity
Rate of acceleration (km/s3)
Uzer, G +, J. Biomech. , 2013
Mechanical environment during LIV
Cell membrane
CytoskeletonNucleus
Vibr
atio
n
Fluid Shear
Membrane deformation
Accelerations
InertiaStiffer & denser nucleus
ECM strain
Cell deformation
Uzer, G+, Curr Osteoporos Rep, 2013
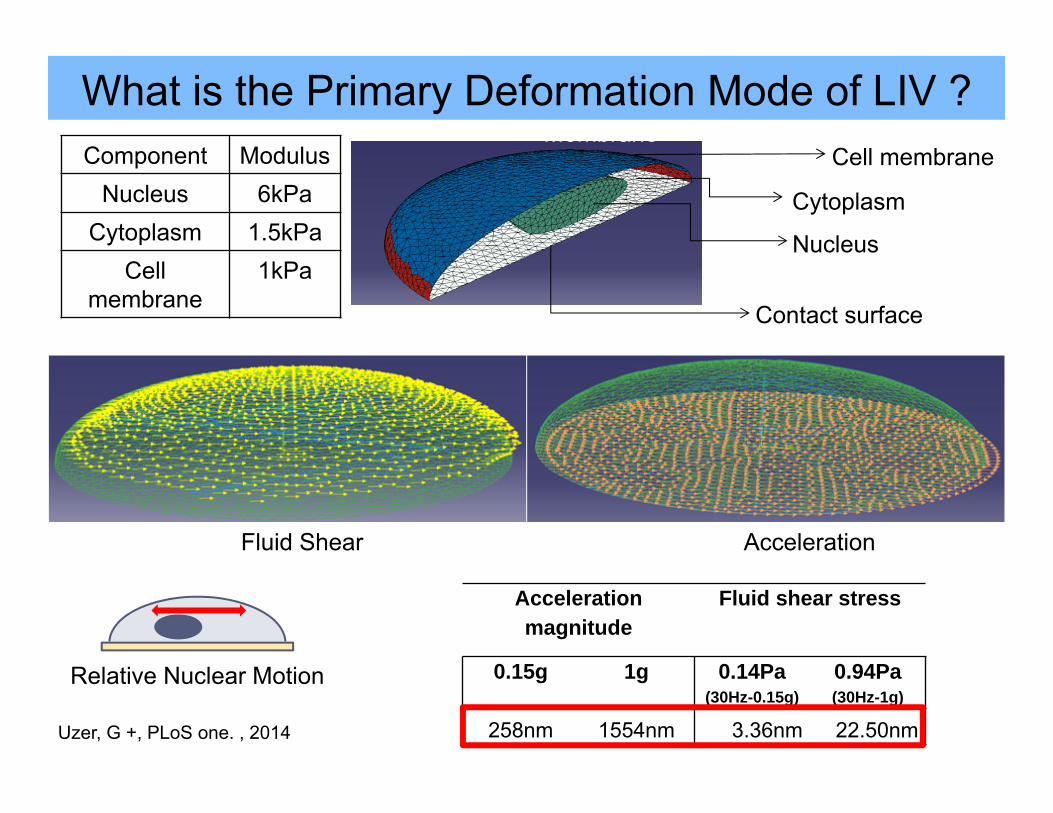
What is the Primary Deformation Mode of LIV ?
Nucleus
Cytoplasm
Membrane
Fluid Shear Acceleration
Nucleus
Cytoplasm
MembraneCell membrane
Cytoplasm
Nucleus
Contact surface
Component ModulusNucleus 6kPa
Cytoplasm 1.5kPaCell
membrane1kPa
Relative Nuclear Motion
Acceleration magnitude
Fluid shear stress
0.15g 1g 0.14Pa(30Hz-0.15g)
0.94Pa(30Hz-1g)
258nm 1554nm 3.36nm 22.50nmUzer, G +, PLoS one. , 2014
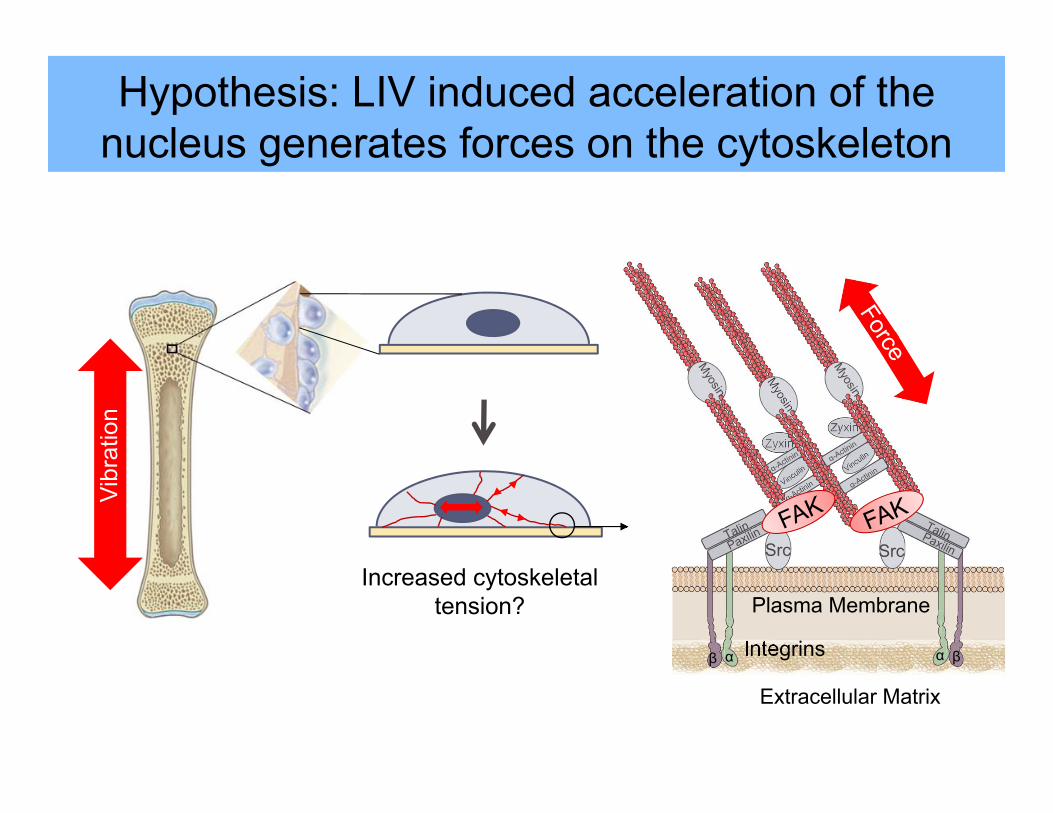
Increased cytoskeletal tension?
Hypothesis: LIV induced acceleration of the nucleus generates forces on the cytoskeleton
Vibr
atio
n
Integrins
Extracellular Matrix
αβ
Src
Zyxin
α β
Src
Zyxin
Plasma Membrane
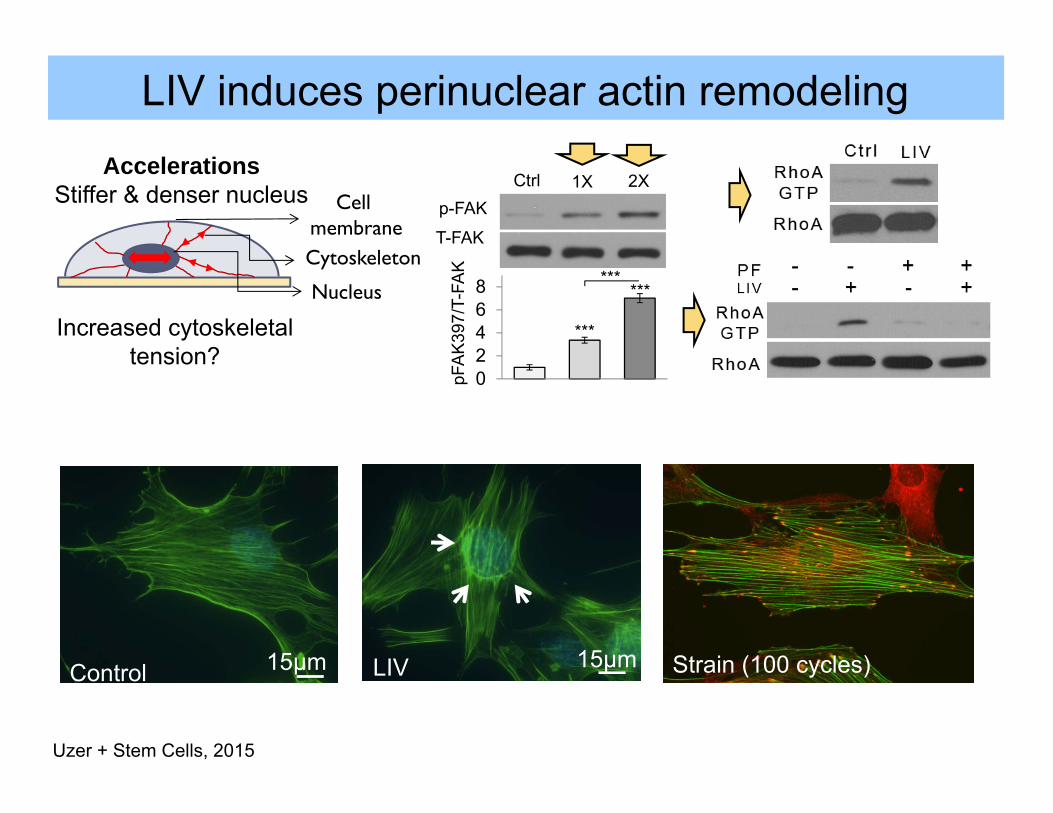
35μm LIV induces perinuclear actin remodeling
15μmLIV0
20
40
60
Ctrl LMS
¥
LIVPer
inuc
lear
actin
form
atio
n (%
)
02468
Ctrl LIV 1XLIV 2X
***
***
***
pFA
K39
7/T-
FAK
1X 2XCtrlp-FAK
T-FAK
15μmControl
Uzer + Stem Cells, 2015
Increased cytoskeletal tension?
AccelerationsStiffer & denser nucleus Cell
membraneCytoskeleton
Nucleus
Strain (100 cycles)
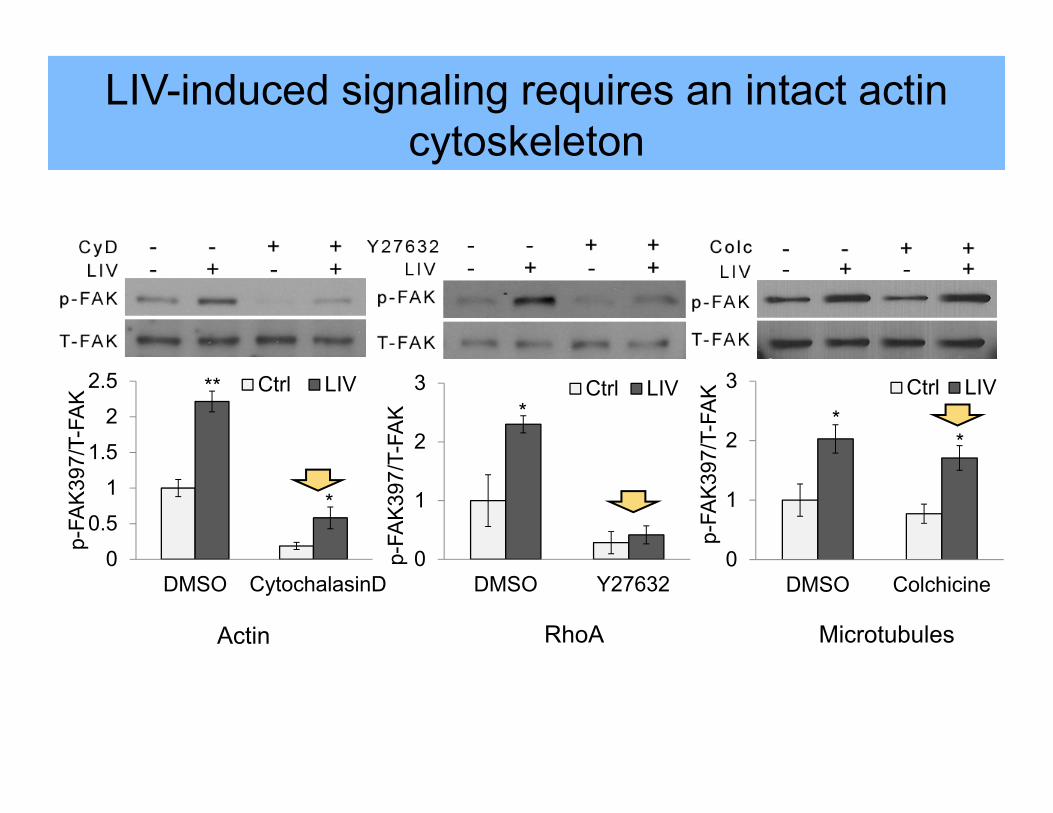
0
1
2
3
DMSO Y2732
p-FA
K39
7/T-
FAK
Ctrl LIV*
Y27632
RhoAActin
LIV-induced signaling requires an intact actin cytoskeleton
35μm
0
1
2
3
DMSO Colchicine
p-FA
K39
7/T-
FAK Ctrl LIV
**
Microtubules
0
0.5
1
1.5
2
2.5
DMSO Cytocalasin D
p-FA
K39
7/T-
FAK Ctrl LIV**
*
CytochalasinD
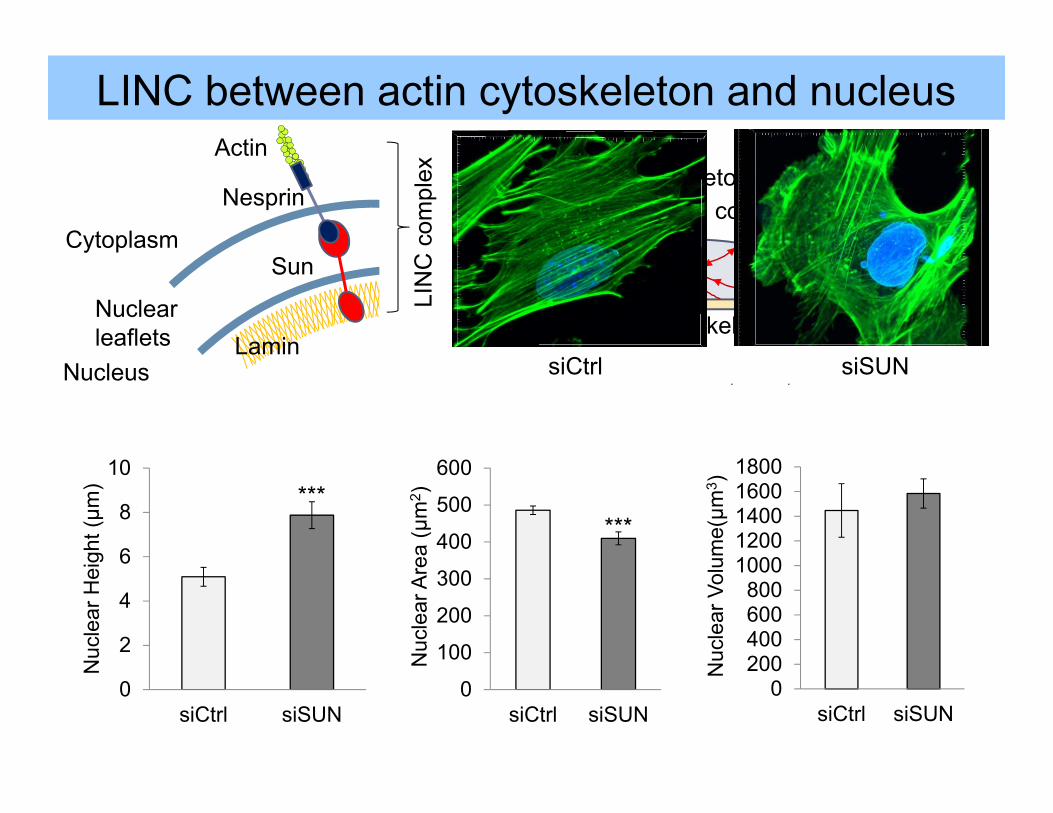
Nuclearleaflets Lamin
Nucleus
Cytoplasm
Nesprin
Sun
Actin
LINC between actin cytoskeleton and nucleus
Linker of Nucleoskeleton and Cytoskeleton Provides mechanical coupling
0
1
2
siCtrl siSUN
Ctrl LIV**
pFA
K39
7/T-
FAK
LIV signals require
a LINCednucleus
LIN
C c
ompl
ex
G. Uzer +Stem Cells, 2015
Increased cytoskeletal tension?
0
1
2
Ctrl DNKASH
Ctrl LIV¥pF
AK
397/
T-FA
K
siCtrl siSUN
0
2
4
6
8
10
siCtrl siSUN
Nuc
lear
Hei
ght (μm
)
0
100
200
300
400
500
600
siCtrl siSUN
Nuc
lear
Are
a (μ
m2 )
0200400600800
10001200140016001800
siCtrl siSUNN
ucle
ar V
olum
e(μm
3 )
******
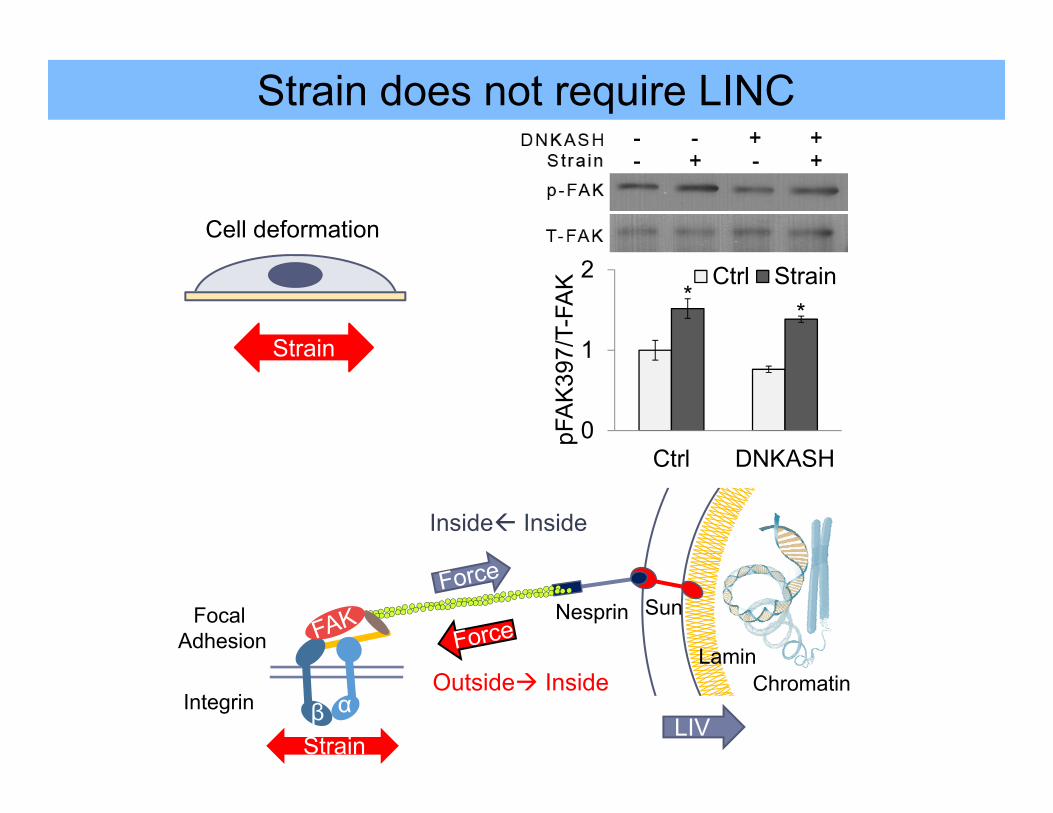
Strain does not require LINC
β α
0
1
2
Ctrl DNKASH
pFA
K39
7/T-
FAK Ctrl Strain
* *
Strain
Outside Inside
Inside Inside
ChromatinIntegrin
Focal Adhesion
Lamin
Nesprin Sun
StrainLIV
Cell deformation
Strain
Nuclearleaflets
TCF/LEF
Nuclear Pore Complex
S
GSK-3βAktβ
WT
Progeria
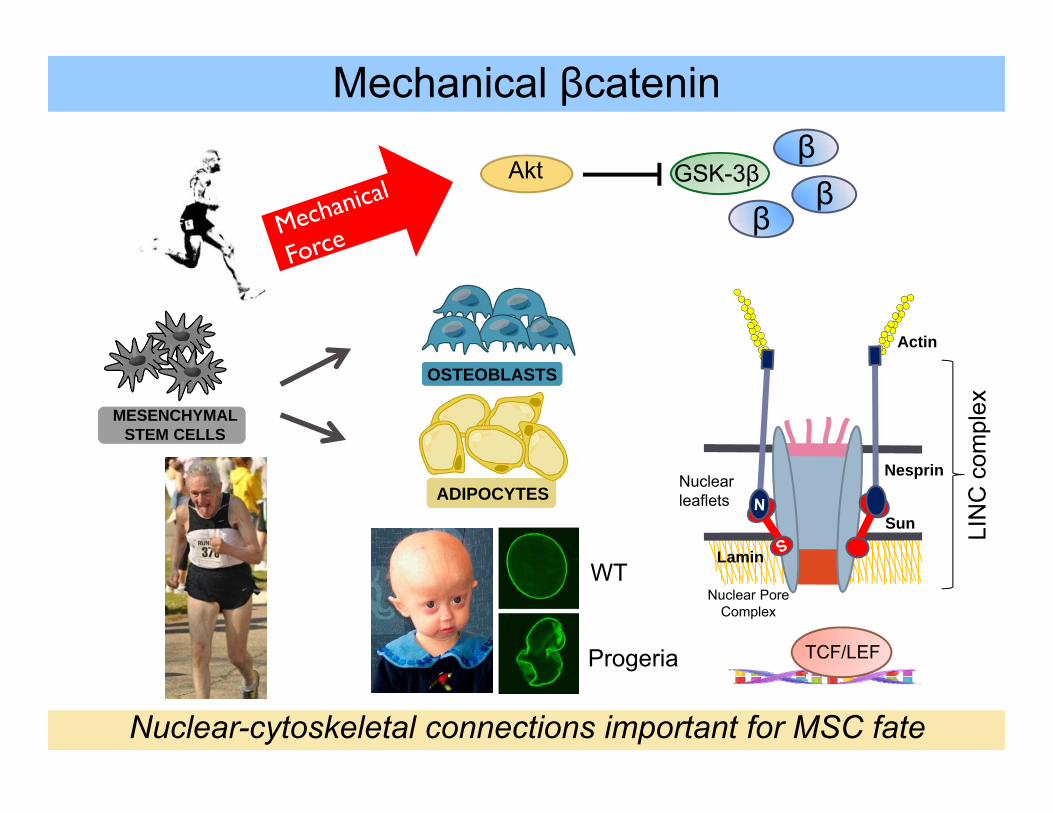
Mechanical βcatenin
MESENCHYMAL STEM CELLS
OSTEOBLASTS
ADIPOCYTES
β
Actin
Nuclear-cytoskeletal connections important for MSC fate
LIN
C c
ompl
ex
Lamin
Sun
Nesprin
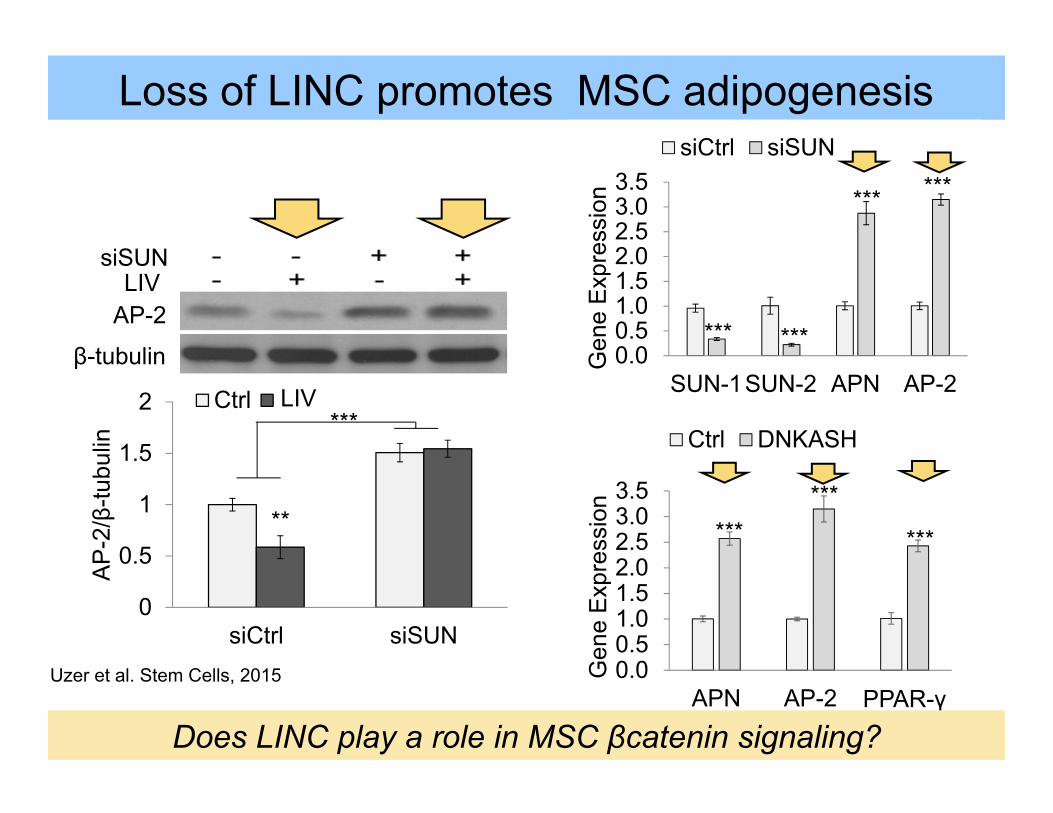
Loss of LINC promotes MSC adipogenesis
0
0.5
1
1.5
2
siCtrl siSUN
AP
-2/β
-tubu
lin
Ctrl LMS***
**
LIV
siSUNLIV
AP-2
β-tubulin
0.00.51.01.52.02.53.03.5
APN AP-2 PPAR-γ
Gen
e E
xpre
ssio
n
Ctrl DNKASH
******
***
0.00.51.01.52.02.53.03.5
SUN-1SUN-2 APN AP-2
Gen
e E
xpre
ssio
n
siCtrl siSUN******
******
Does LINC play a role in MSC βcatenin signaling?
Uzer et al. Stem Cells, 2015
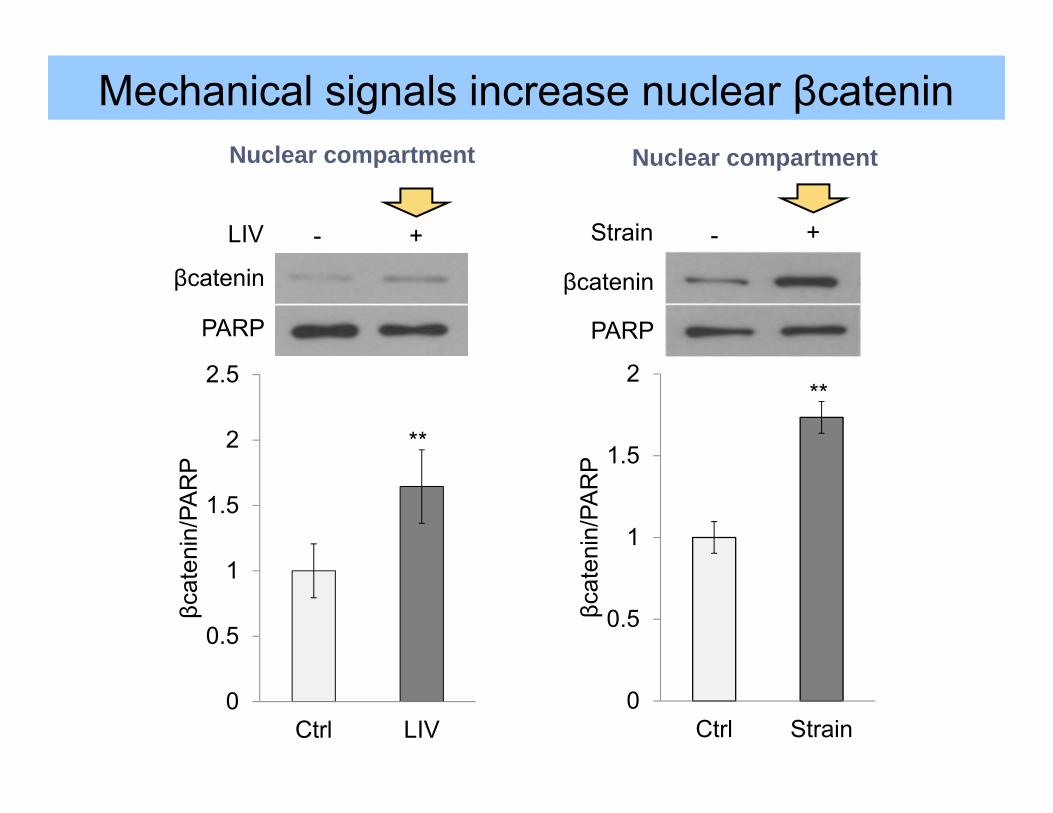
Mechanical signals increase nuclear βcatenin
0
0.5
1
1.5
2
2.5
Ctrl LIV
βcat
enin
/PA
RP
βcatenin
LIV - +
PARP
**
0
0.5
1
1.5
2
Ctrl Strain
βcat
enin
/PA
RP
βcatenin
Strain - +
PARP
**
Nuclear compartment Nuclear compartment
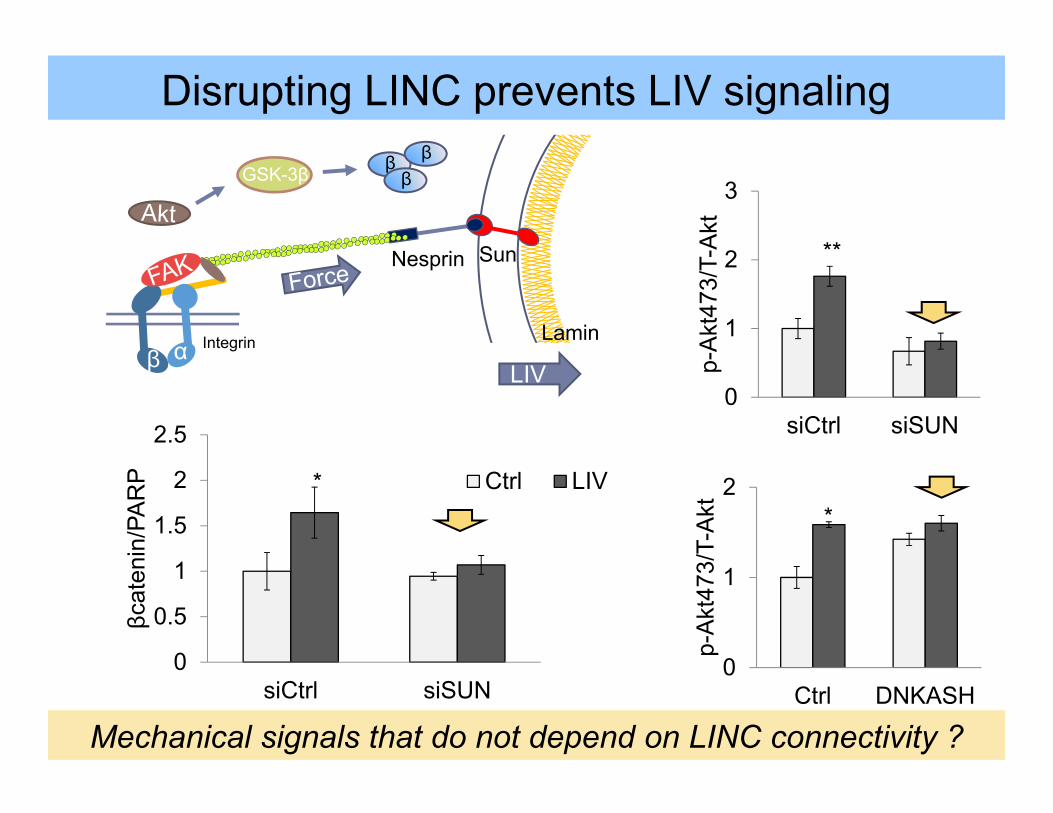
β α Integrin Lamin
Nesprin Sun
StrainLIV
0
1
2
Ctrl DNKASHp-
Akt
473/
T-A
kt *
0
1
2
3
siCtrl siSUN
p-A
kt47
3/T-
Akt
**
Disrupting LINC prevents LIV signaling
GSK-3β β ββ
0
0.5
1
1.5
2
2.5
siCtrl siSUN
βcat
enin
/PA
RP Ctrl LIV*
Mechanical signals that do not depend on LINC connectivity ?
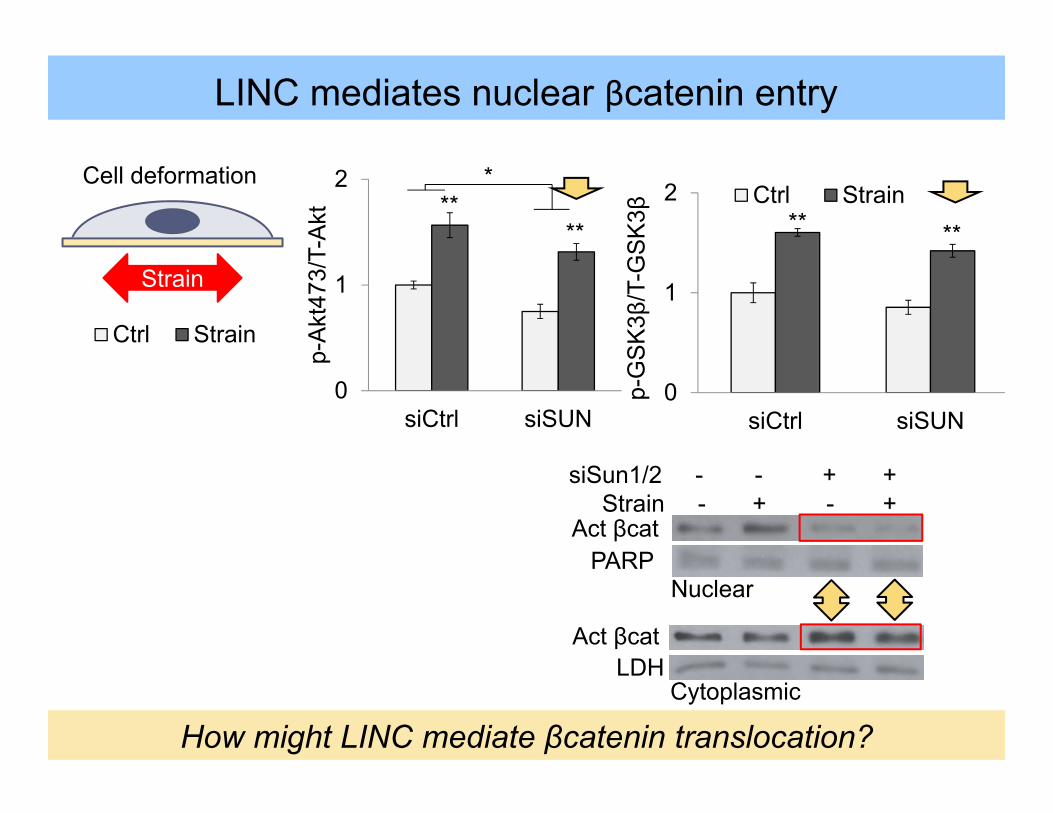
Cell deformation
LDHAct βcat
siSun1/2 - - + +Strain - + - +
Act βcatPARP
0
1
2
siCtrl siSUN
Ctrl Strain
p-G
SK
3β/T
-GS
K3β ** **
LINC mediates nuclear βcatenin entry
Strain
Nuclear
Cytoplasmic
How might LINC mediate βcatenin translocation?
β
β
β
Nucleusβ
Controlβ
β
β
Nucleusβ
siSUN0
1
2
siCtrl siSUN
p-A
kt47
3/T-
Akt
Ctrl Strain
** **
*
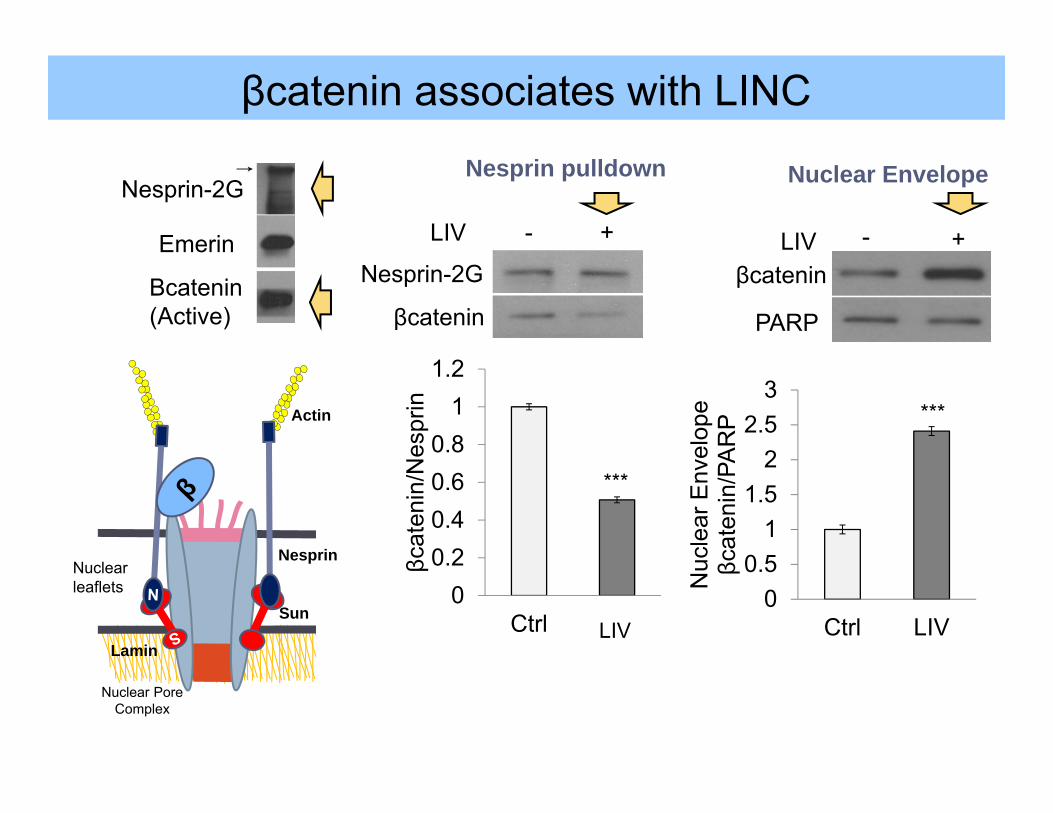
Nuclearleaflets
Nuclear Pore Complex
S
Actin
Lamin
Sun
Nesprin
S
Nesprin-2G
Emerin
Βcatenin(Active)
00.20.40.60.8
11.2
Ctrl Strain
βcat
enin
/Nes
prin
***
LIV
Nesprin-2G
βcatenin
LIV - +
βcateninLIV - +
00.5
11.5
22.5
3
Ctrl LIV
Nuc
lear
Env
elop
eβc
aten
in/P
AR
P
PARP
***
βcatenin associates with LINC
Nesprin pulldown Nuclear Envelope
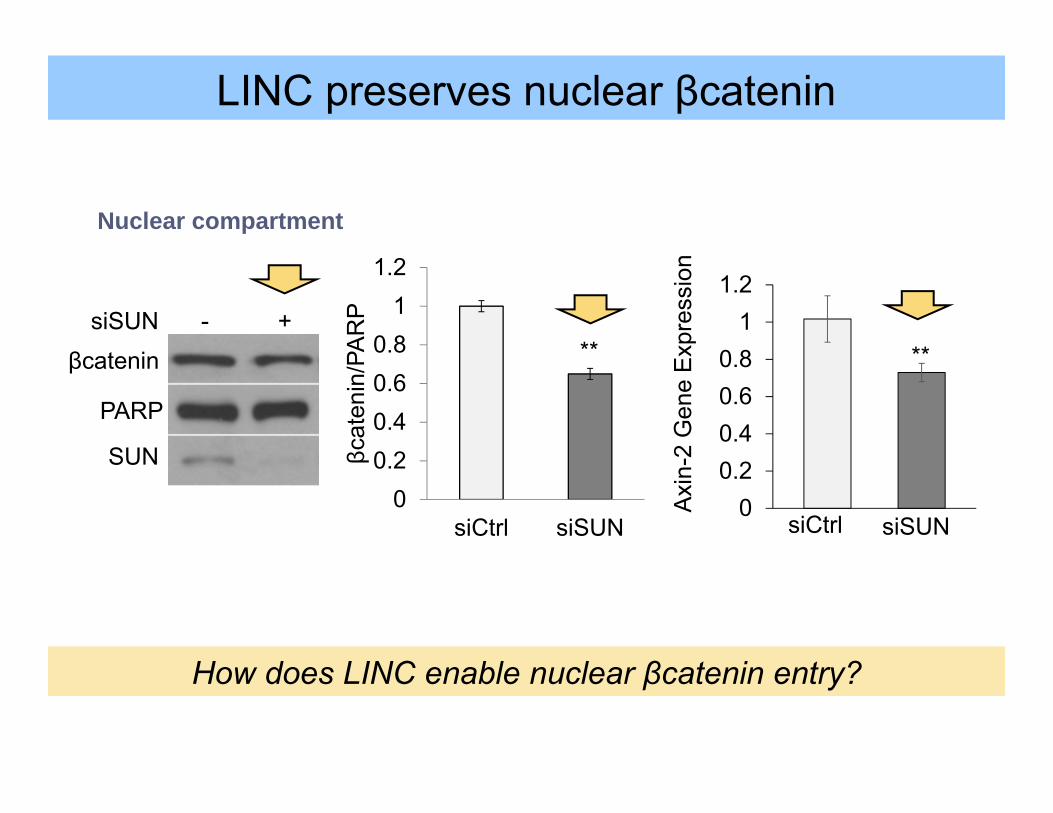
00.20.40.60.8
11.2
Axi
n-2
Gen
e E
xpre
ssio
n
siCtrl siSUN
**
How does LINC enable nuclear βcatenin entry?
LINC preserves nuclear βcatenin
00.20.40.60.8
11.2
siCtrl siSUN
βcat
enin
/PA
RP
**
PARP
SUN
siSUN - +βcatenin
Nuclear compartment
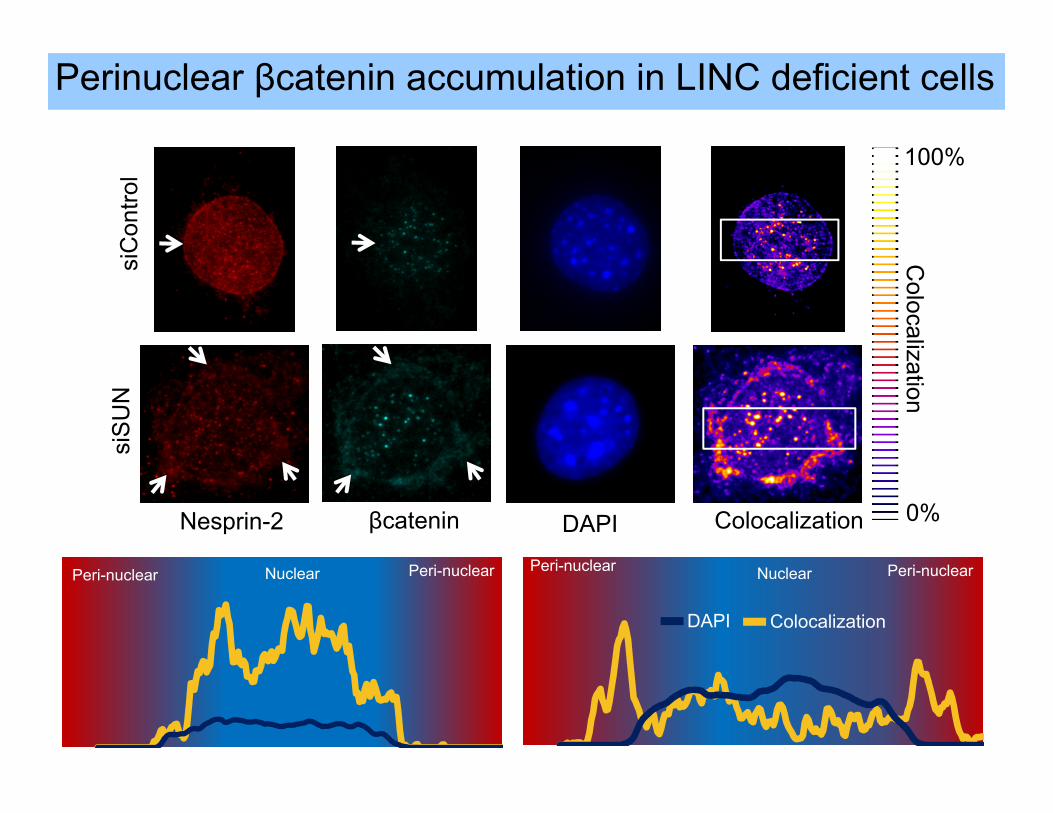
Peri-nuclear Peri-nuclear Nuclear
DAPI Colocalization
Peri-nuclear Peri-nuclear Nuclear
siC
ontro
lsi
SU
N
Nesprin-2 βcatenin
**
DAPI Colocalization
100%
0%
Colocalization
Perinuclear βcatenin accumulation in LINC deficient cells
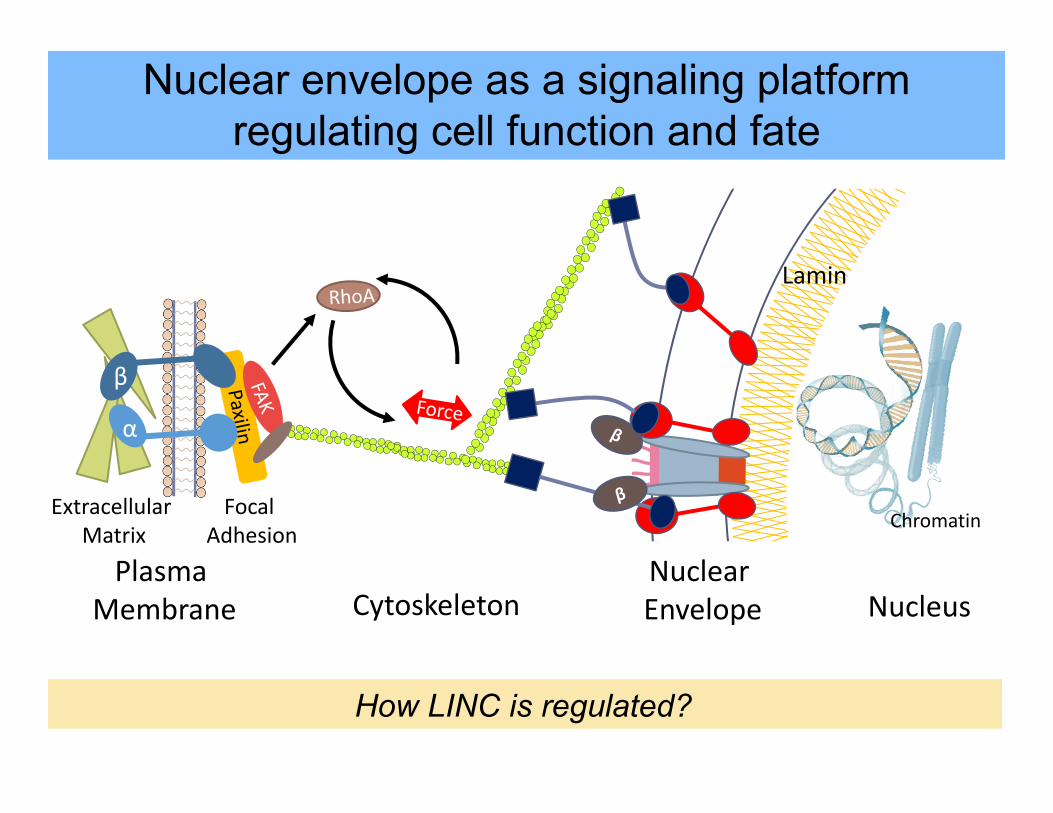
Lamin
Chromatin
Plasma Membrane
β
α
Focal Adhesion
Extracellular Matrix
CytoskeletonNuclear Envelope Nucleus
Nuclear envelope as a signaling platformregulating cell function and fate
How LINC is regulated?
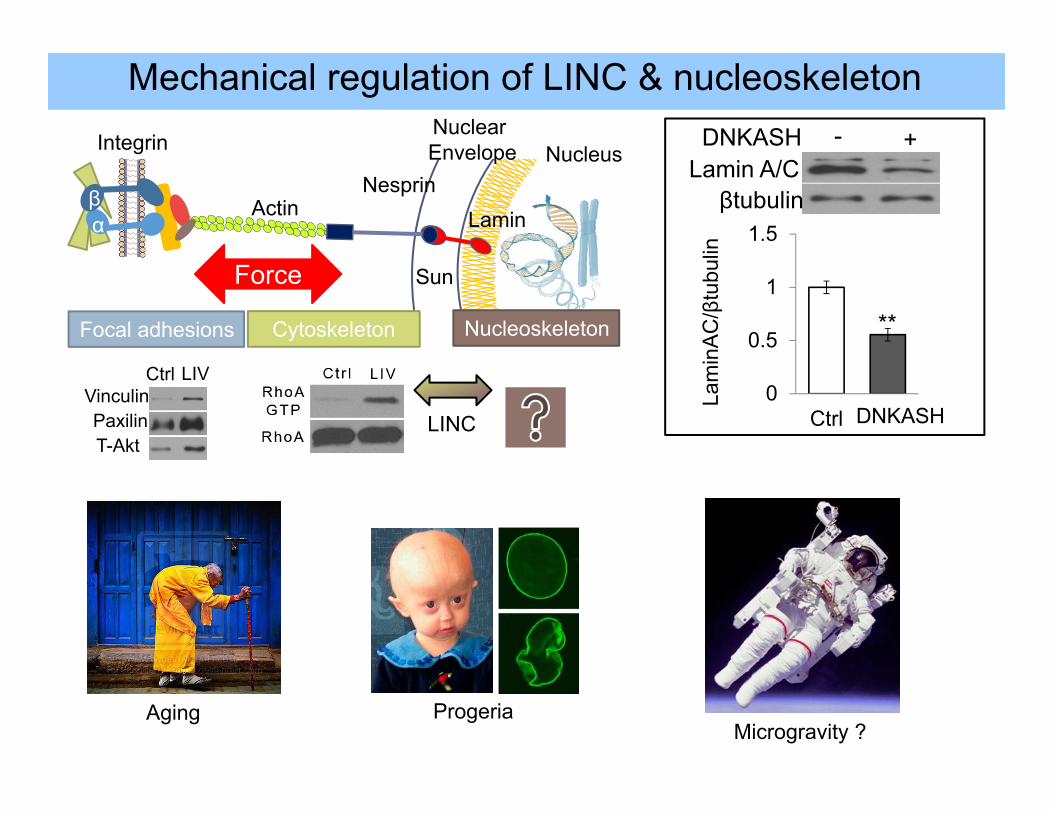
βα
Integrin
Lamin
NucleusNuclear Envelope
Nesprin
Sun
Ctrl LIVVinculinPaxilinT-Akt
Focal adhesions Cytoskeleton Nucleoskeleton
Actin
LINC
Force
βtubulin
DNKASHLamin A/C
- +
0
0.5
1
1.5
Ctrl DNKASH
**
Lam
inA
C/β
tubu
lin
Mechanical regulation of LINC & nucleoskeleton
Aging ProgeriaMicrogravity ?
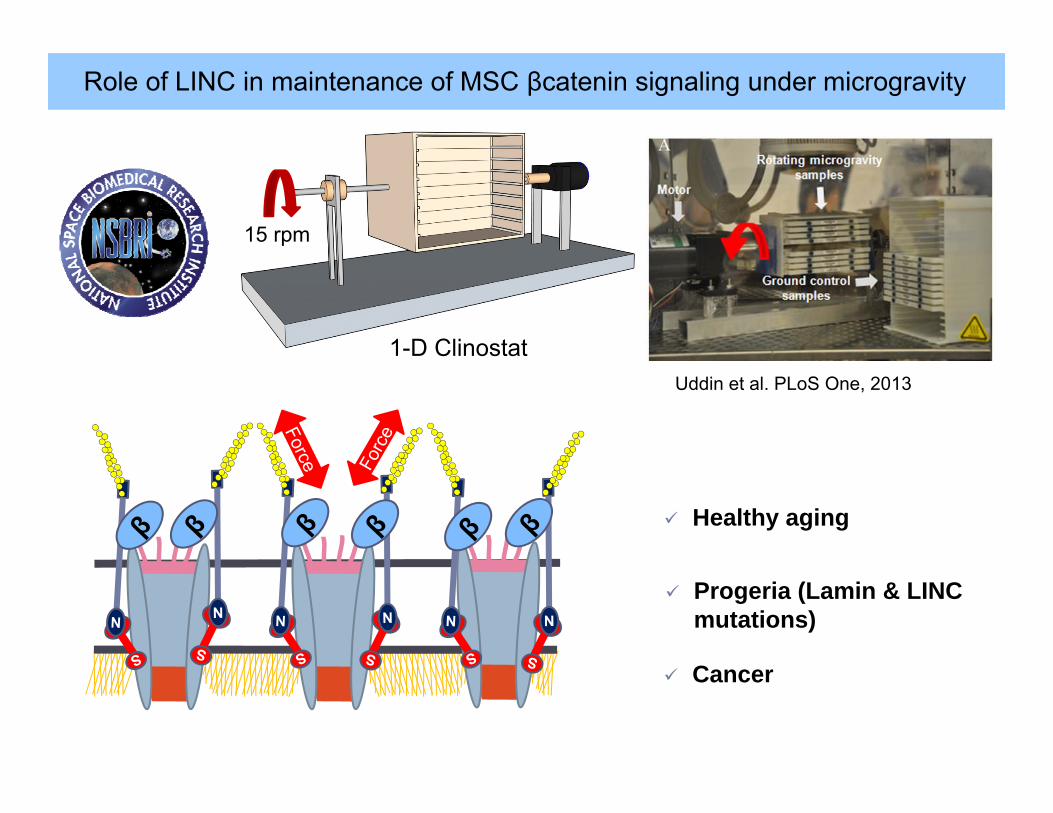
Role of LINC in maintenance of MSC βcatenin signaling under microgravity
15 rpm
1-D ClinostatUddin et al. PLoS One, 2013
S
Healthy aging
Progeria (Lamin & LINC mutations)
Cancer

Acknowledgements
Janet Rubin , MD
Maya Styner, MD
Buer Sen, MD
Zhihui Xie, MD
Guniz Bas Uzer
Stefan Judex , PhD
Clinton Rubin , PhD
Yi-Xian Qin, PhD
Aditi Nivedita Senthilnathan
Kaushik Puranam
Sophia Kim
Junaid Qureshi
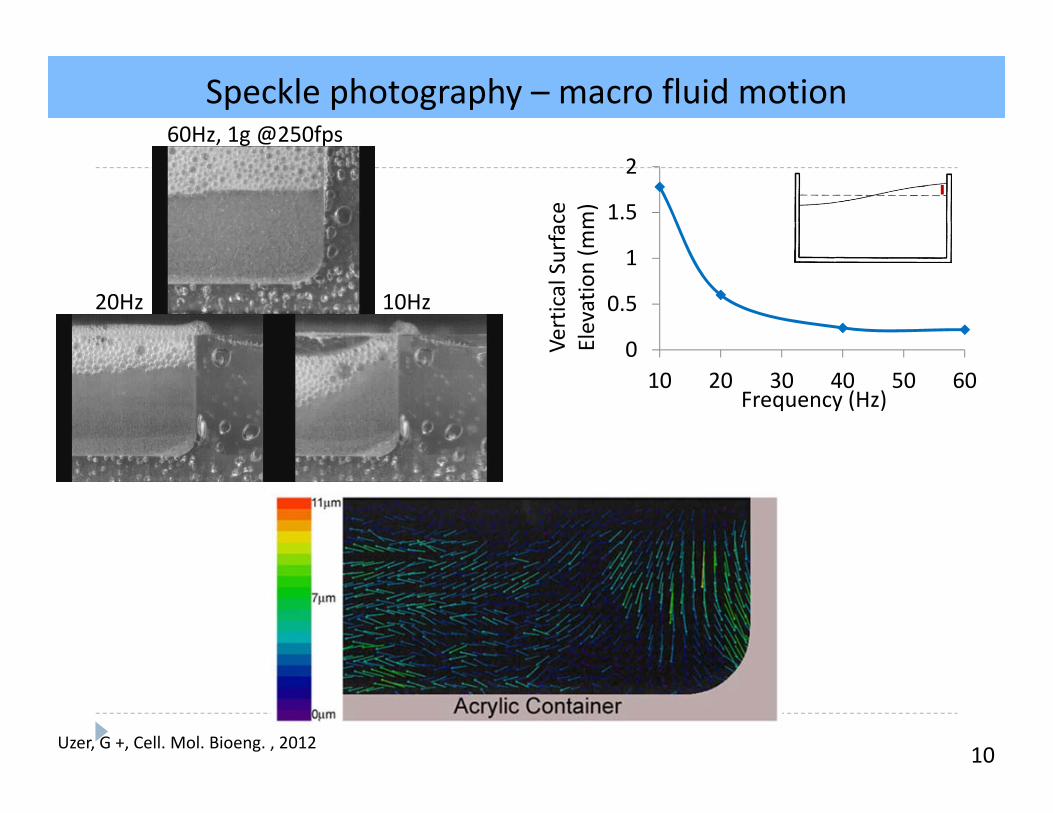
Speckle photography – macro fluid motion
10
A
B
0
0.5
1
1.5
2
10 20 30 40 50 60
Vertical Surface
Elevation (m
m)
Frequency (Hz)
60Hz, 1g @250fps
10Hz20Hz
Uzer, G +, Cell. Mol. Bioeng. , 2012
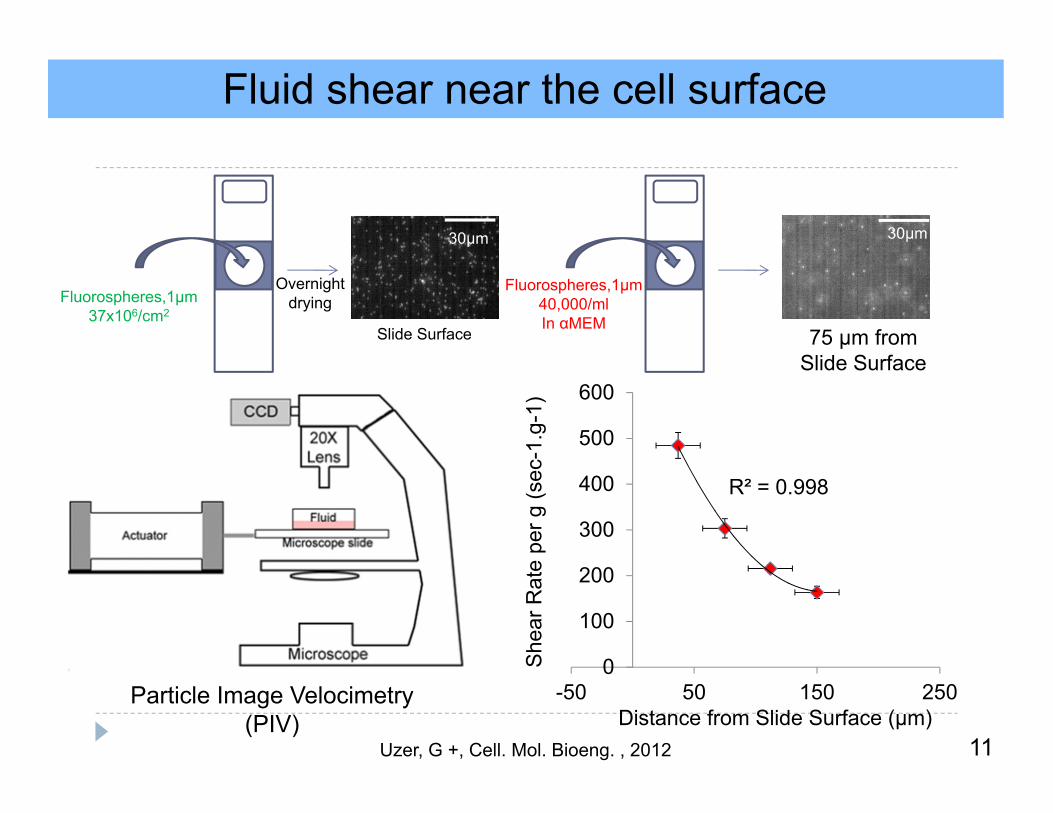
Fluid shear near the cell surface
Particle Image Velocimetry(PIV)
11
30µm30µm
OvernightdryingFluorospheres,1µm
37x106/cm2
Fluorospheres,1µm40,000/mlIn αMEM
Slide Surface 75 µm fromSlide Surface
R² = 0.998
0
100
200
300
400
500
600
-50 50 150 250
She
ar R
ate
per g
(sec
-1.g
-1)
Distance from Slide Surface (µm)Uzer, G +, Cell. Mol. Bioeng. , 2012
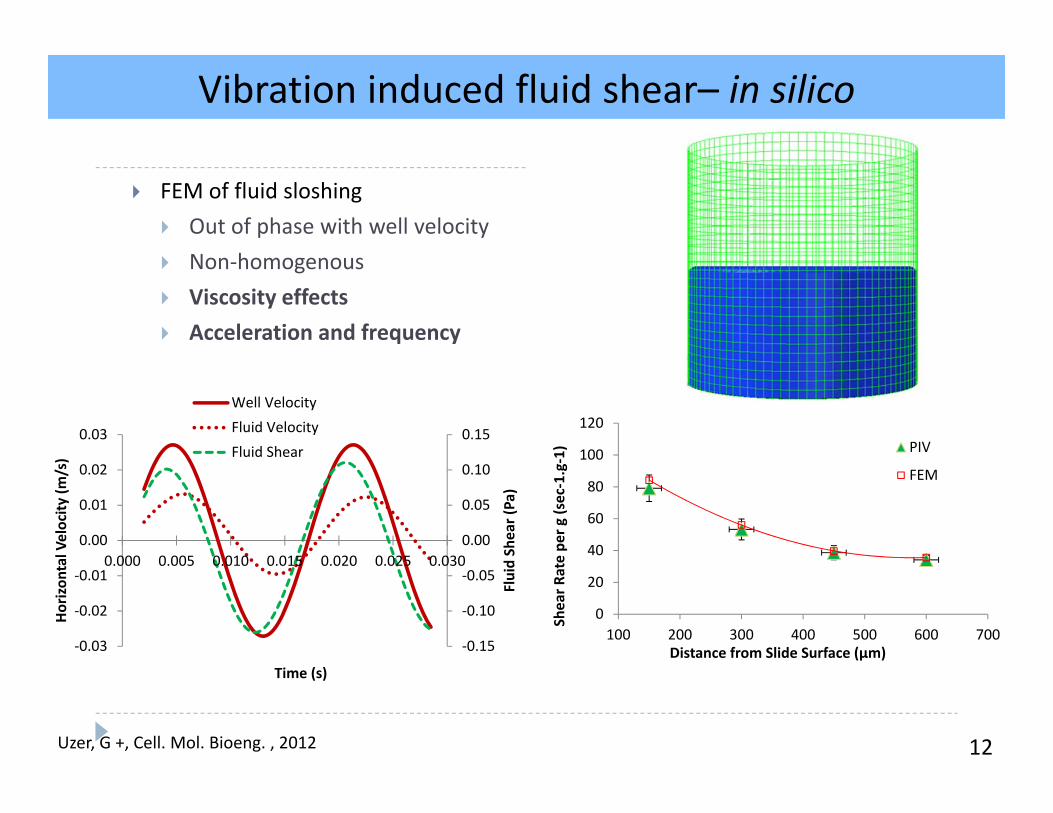
12
FEM of fluid sloshing Out of phase with well velocity Non‐homogenous Viscosity effects Acceleration and frequency
‐0.15
‐0.10
‐0.05
0.00
0.05
0.10
0.15
‐0.03
‐0.02
‐0.01
0.00
0.01
0.02
0.03
0.000 0.005 0.010 0.015 0.020 0.025 0.030
Fluid Shear (Pa)
Horizon
tal Velocity
(m/s)
Time (s)
Well VelocityFluid VelocityFluid Shear
Velocity (m
m/s)
5mm
14mm
A
0
20
40
60
80
100
120
100 200 300 400 500 600 700
Shear R
ate pe
r g (sec‐1.g‐1)
Distance from Slide Surface (µm)
PIV
FEM
Vibration induced fluid shear– in silico
Uzer, G +, Cell. Mol. Bioeng. , 2012
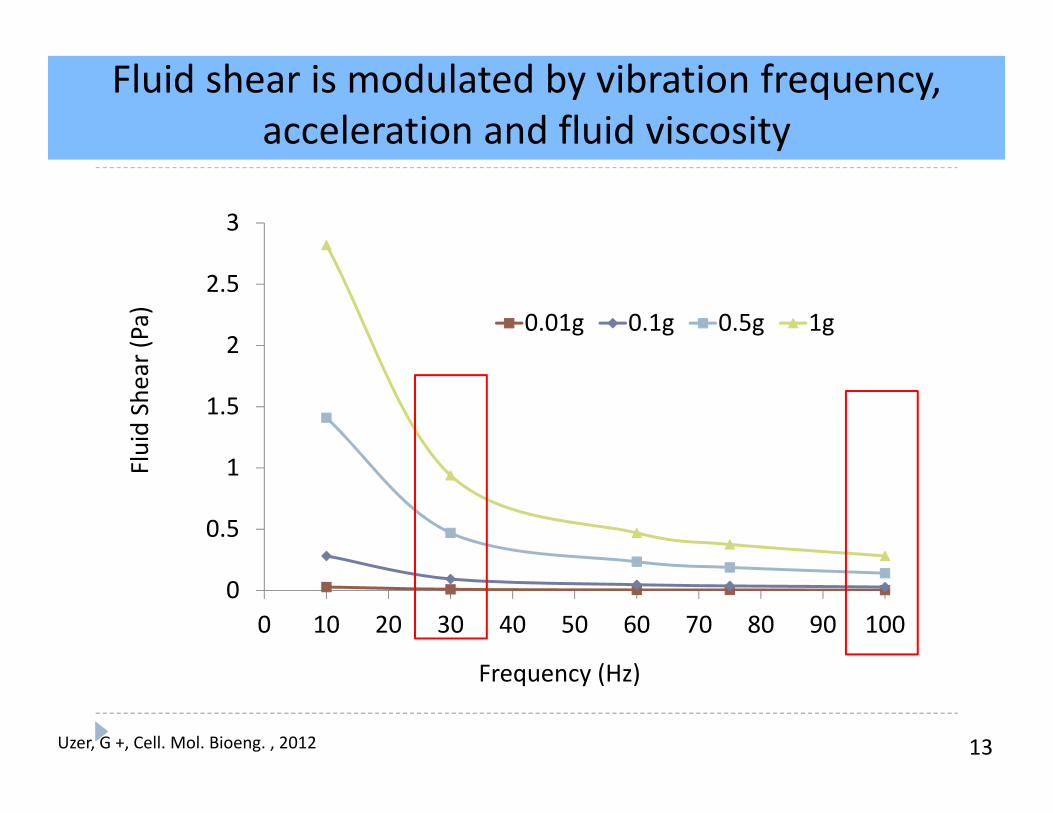
13
Fluid shear is modulated by vibration frequency, acceleration and fluid viscosity
0
0.5
1
1.5
2
2.5
3
0 10 20 30 40 50 60 70 80 90 100
Fluid Shear (Pa)
Frequency (Hz)
0.01g 0.1g 0.5g 1g
Uzer, G +, Cell. Mol. Bioeng. , 2012
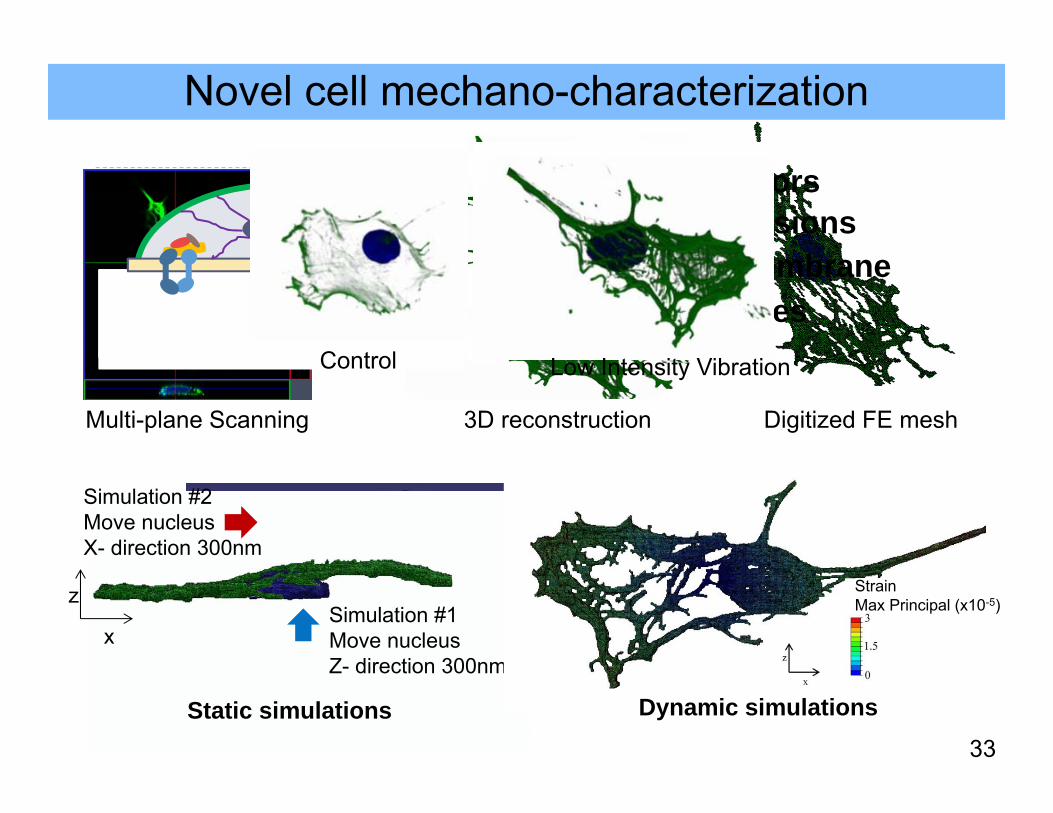
Novel cell mechano-characterization
Multi-plane Scanning 3D reconstruction Digitized FE mesh
Cell receptors Focal adhesions Plasma membrane Microtubules
Nucleus
Cytoplasm
MembraneCell membrane
Cytoplasm (not shown)
Nucleus
33
0
3500
7000
10500
14000
Maximum Principal Strain (x10-3)
0
3
9
12
15
Num
ber o
f Ele
men
ts (x
103 )
0.6
0.9
1.2
1.5
1.8
2.1
2.4
2.7
3.0
3.3
3.6
3.9
4.2
4.5
4.8
Simulation #2Move nucleus X- direction 300nm
Simulation #1Move nucleus Z- direction 300nm
x
z
Static simulations
StrainMax Principal (x10-5)
Dynamic simulations
Control Low Intensity Vibration
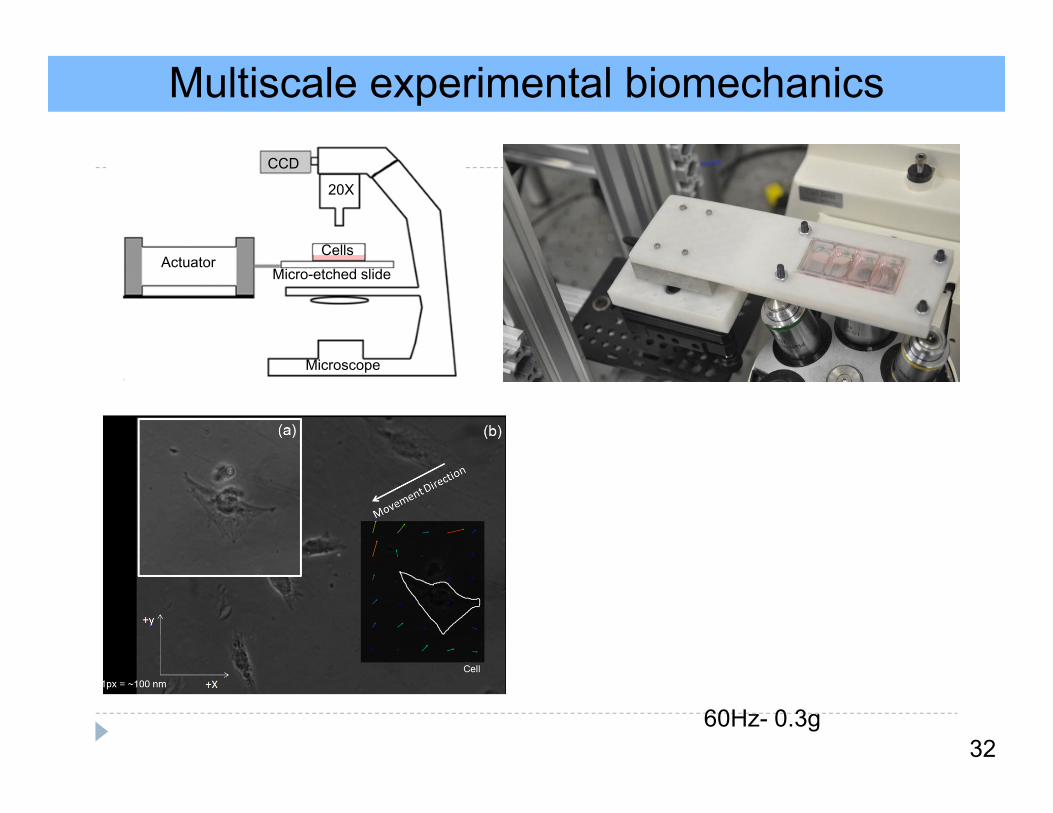
Multiscale experimental biomechanicsV
ibra
tion
Dire
ctio
n
32
20X
CCD
ActuatorMicro-etched slide
Cells
Microscope
60Hz- 0.3g
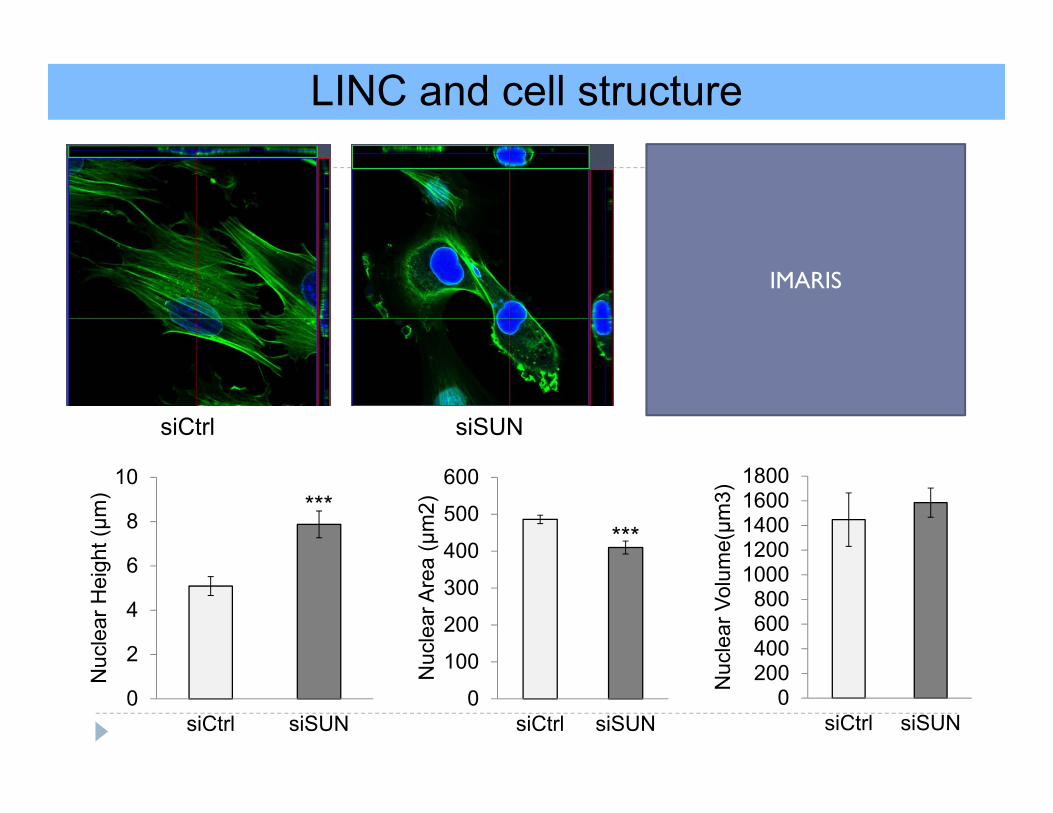
LINC and cell structure
0
2
4
6
8
10
siCtrl siSUN
Nuc
lear
Hei
ght (μm
)
0
100
200
300
400
500
600
siCtrl siSUN
Nuc
lear
Are
a (μ
m2)
0200400600800
10001200140016001800
siCtrl siSUNN
ucle
ar V
olum
e(μm
3)
******
siCtrl siSUN
IMARIS
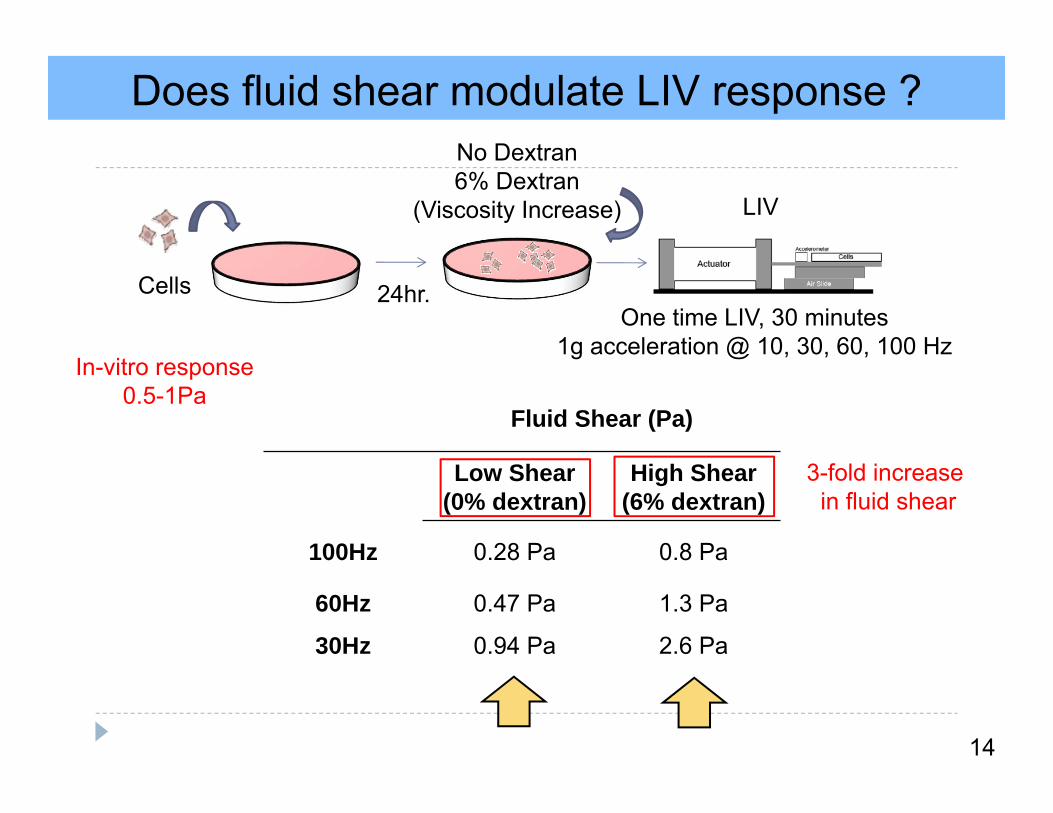
Does fluid shear modulate LIV response ?
14
24hr.Cells
No Dextran6% Dextran
(Viscosity Increase)
One time LIV, 30 minutes1g acceleration @ 10, 30, 60, 100 Hz
Fluid Shear (Pa)
Low Shear (0% dextran)
High Shear (6% dextran)
100Hz 0.28 Pa 0.8 Pa
60Hz 0.47 Pa 1.3 Pa
30Hz 0.94 Pa 2.6 Pa
LIV
In-vitro response0.5-1Pa
3-fold increase in fluid shear
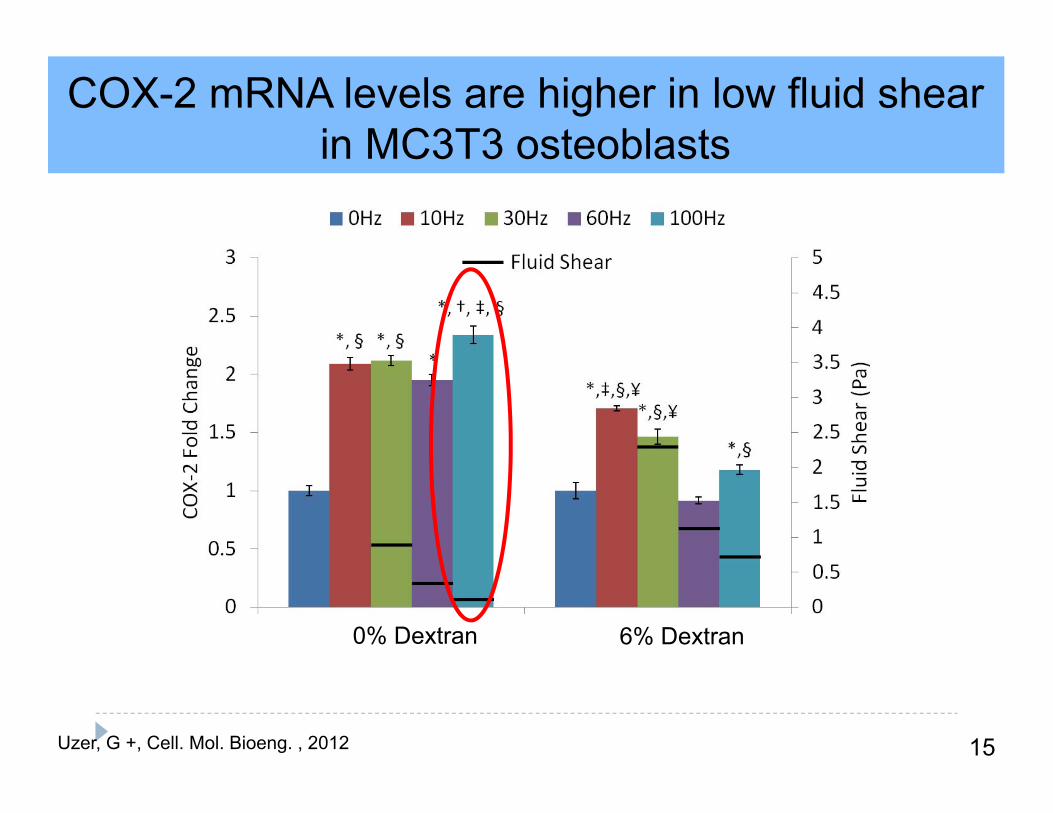
COX-2 mRNA levels are higher in low fluid shear in MC3T3 osteoblasts
15Uzer, G +, Cell. Mol. Bioeng. , 2012
6% Dextran0% Dextran
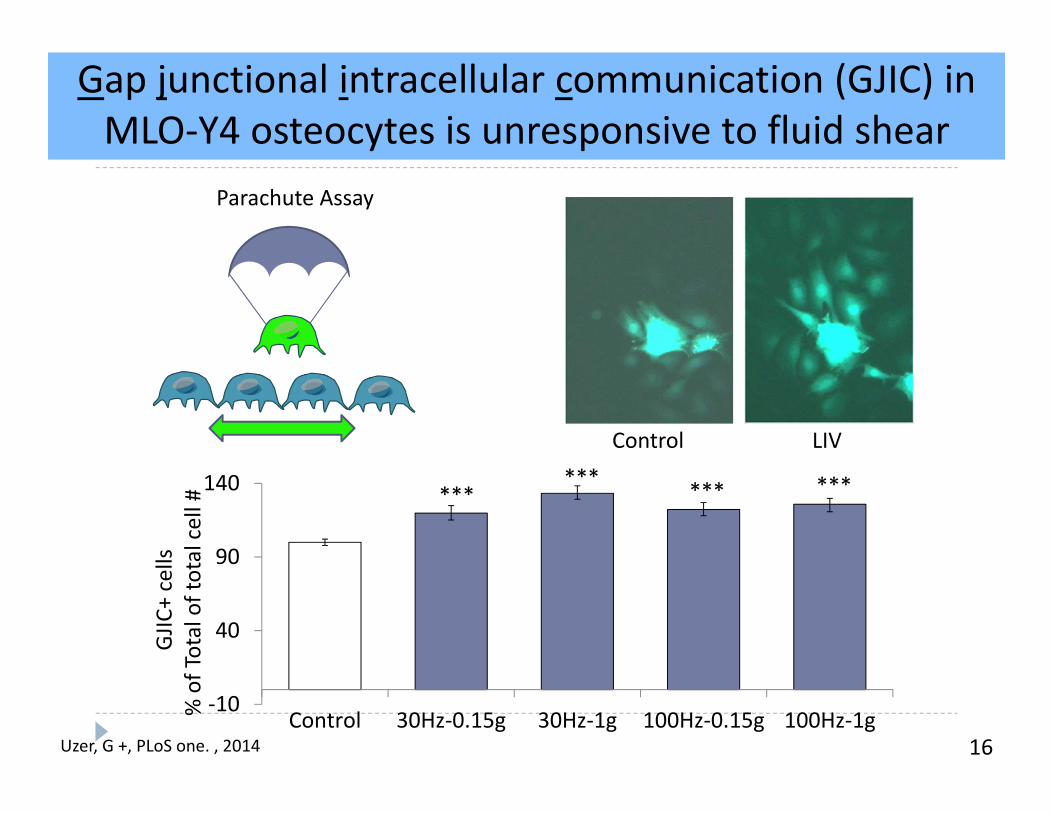
Gap junctional intracellular communication (GJIC) in MLO‐Y4 osteocytes is unresponsive to fluid shear
16
Control LIV
Uzer, G +, PLoS one. , 2014
‐10
40
90
140
Control 30Hz‐0.15g 30Hz‐1g 100Hz‐0.15g 100Hz‐1g
****** *** ***
GJIC
+ cells
% of Total of total cell #
Parachute Assay
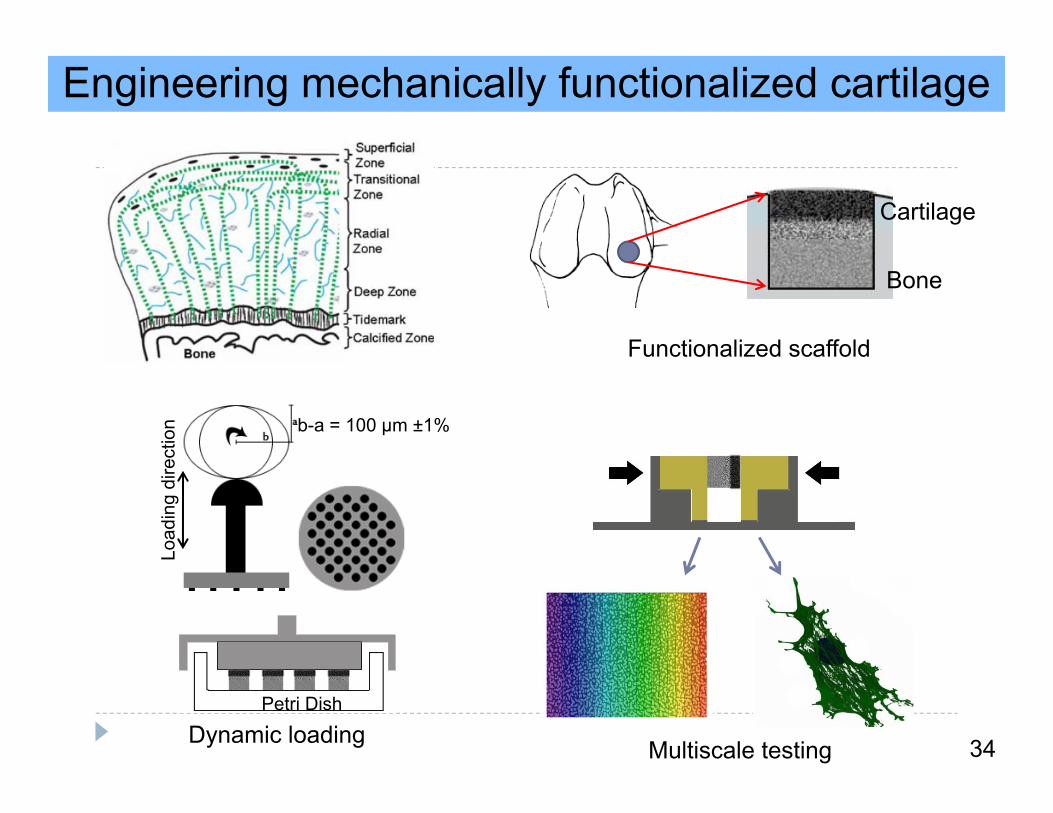
Engineering mechanically functionalized cartilage
Cartilage
Bone
Functionalized scaffold
Load
ing
dire
ctio
n b-a = 100 μm ±1%
Petri Dish
Dynamic loading Multiscale testing 34
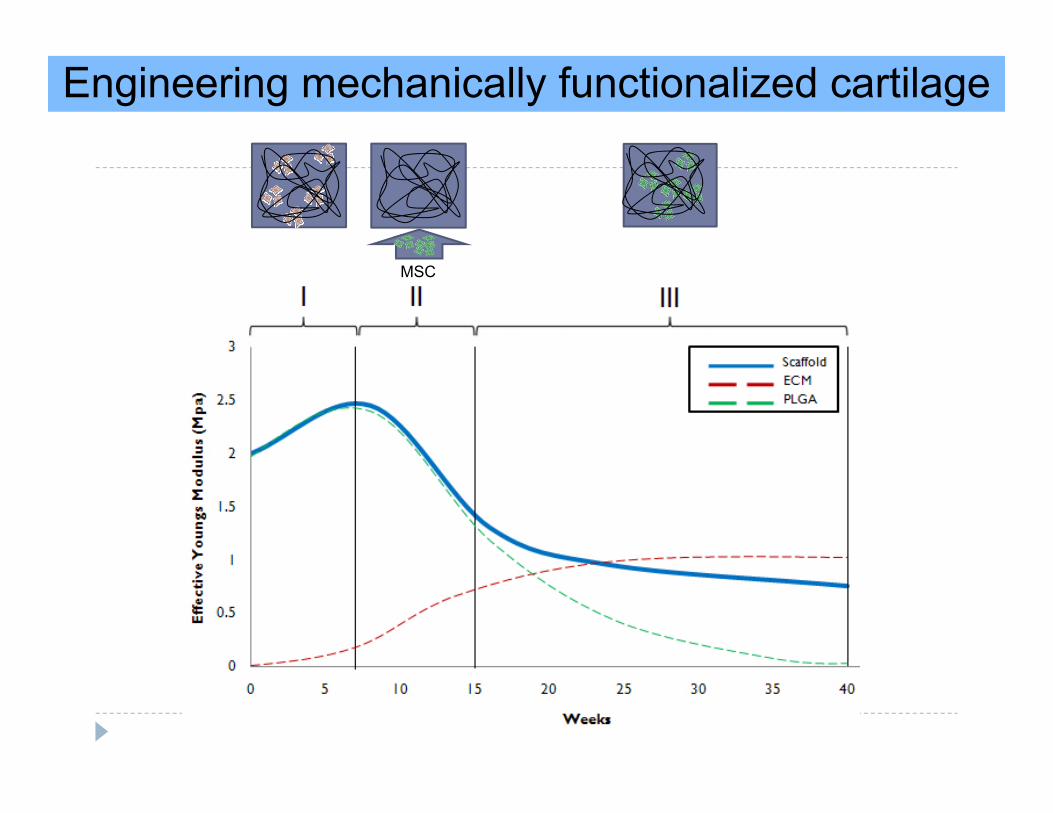
Engineering mechanically functionalized cartilage
MSC
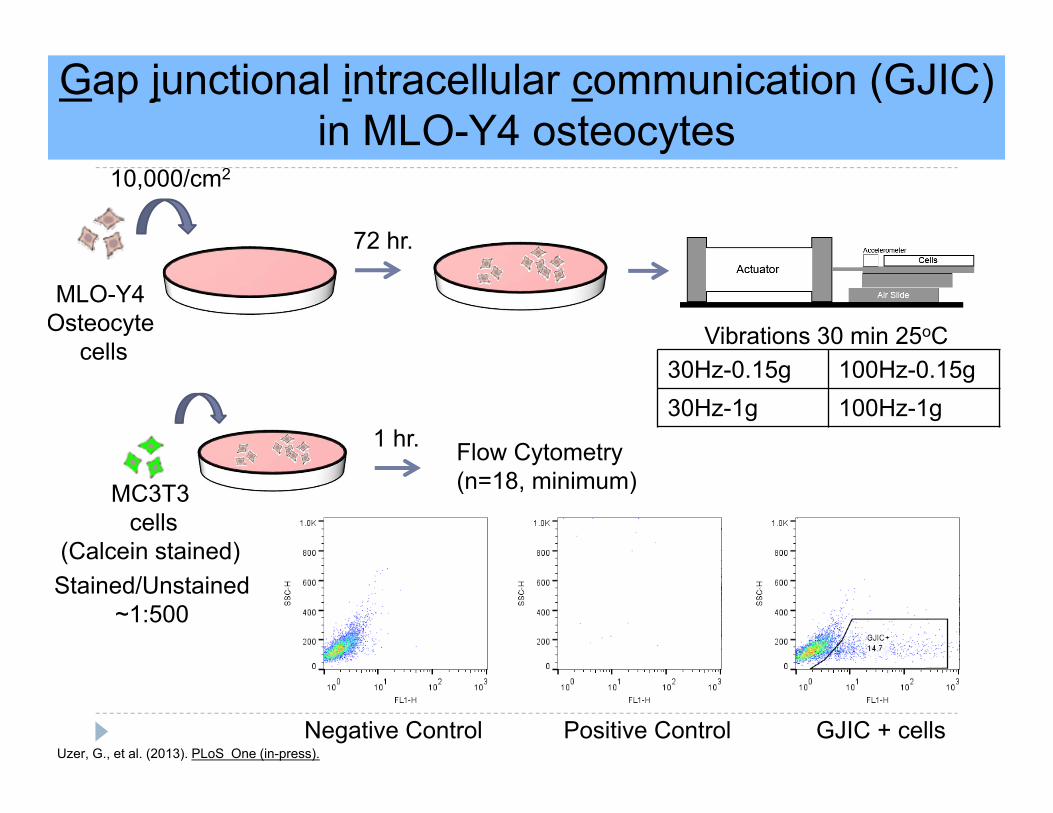
Gap junctional intracellular communication (GJIC) in MLO-Y4 osteocytes
MC3T3cells
(Calcein stained)Stained/Unstained
~1:500
MLO-Y4Osteocyte
cells
72 hr.
Vibrations 30 min 25oC
Flow Cytometry(n=18, minimum)
Negative Control Positive Control GJIC + cells
10,000/cm2
30Hz-0.15g 100Hz-0.15g30Hz-1g 100Hz-1g
Uzer, G., et al. (2013). PLoS One (in-press).
1 hr.
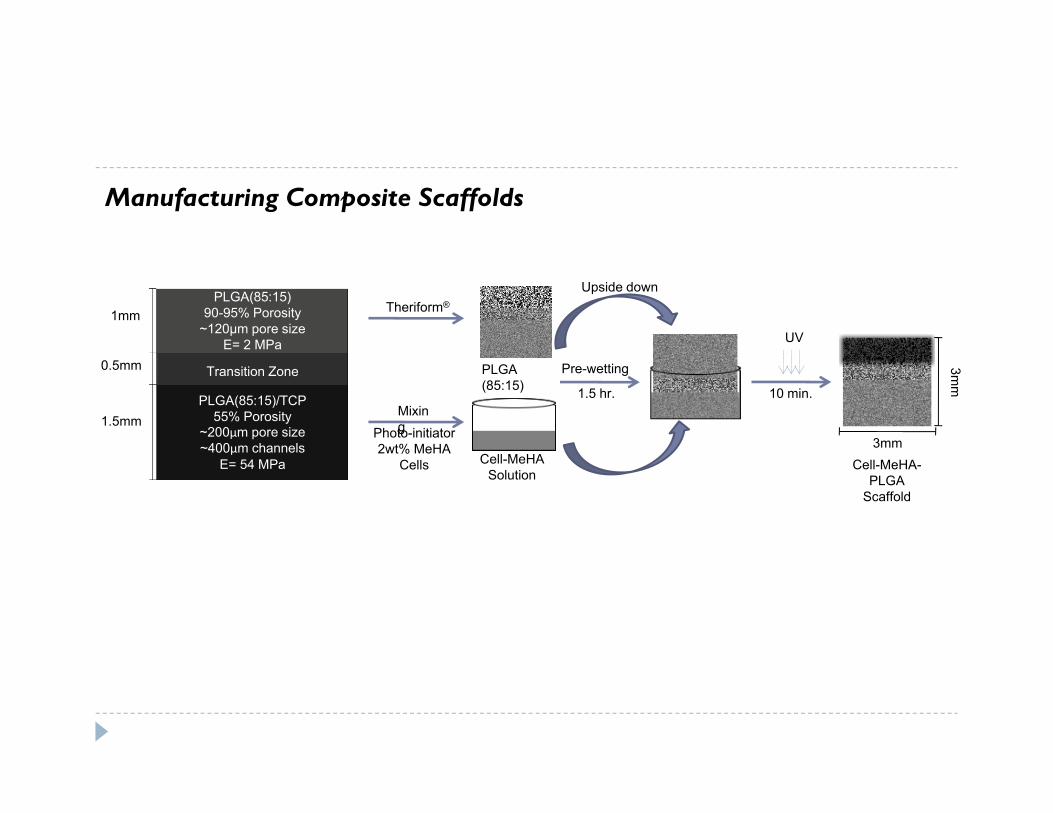
Manufacturing Composite Scaffolds
PLGA (85:15)
Cell-MeHASolution
Theriform®
Pre-wetting
Photo-initiator2wt% MeHA
Cells
UV
3mm
3mm
1.5 hr. 10 min.
PLGA(85:15)90-95% Porosity
~120μm pore sizeE= 2 MPa
1mm
PLGA(85:15)/TCP55% Porosity
~200μm pore size~400μm channels
E= 54 MPa
Transition Zone
1.5mm
0.5mm
Mixing
Cell-MeHA-PLGA
Scaffold
Upside down

Dynamic Compression Loading (DCL) Loading regime. Displacement control. 1Hz, 4hr a day. 10% Strain.
Sterile technique. Loading inside an incubator. 5% CO2, 37oC
Up to 70 scaffolds.
Load
ing
dire
ctio
n
b-a = 100 μm ±1%
Petri Dish
1. Stem Cells. 2004;22(3):313-23.
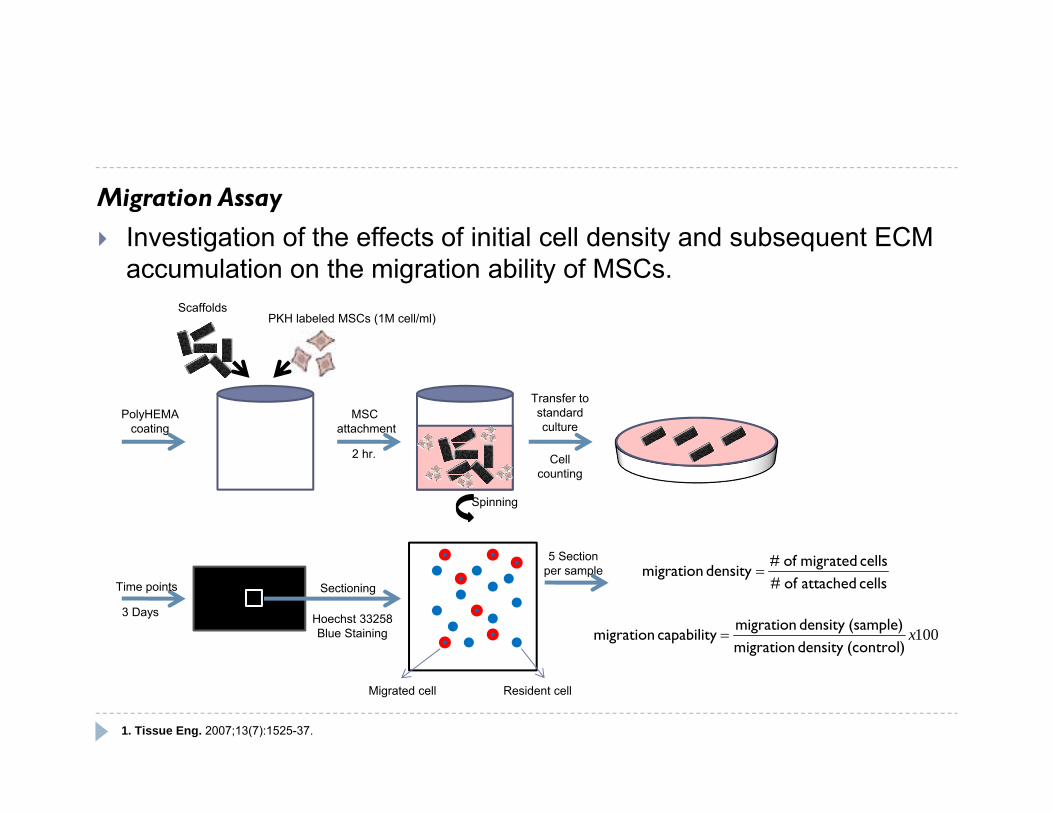
Migration Assay Investigation of the effects of initial cell density and subsequent ECM
accumulation on the migration ability of MSCs.
PolyHEMAcoating
ScaffoldsPKH labeled MSCs (1M cell/ml)
MSC attachment
2 hr.
Spinning
Transfer to standardculture
Hoechst 33258Blue Staining
SectioningTime points
3 Days
Cell counting
cells attached of #cells migrated of #
density migration
100x(control)density migration(sample)density migration
capability migration
Migrated cell Resident cell
5 Sectionper sample
1. Tissue Eng. 2007;13(7):1525-37.
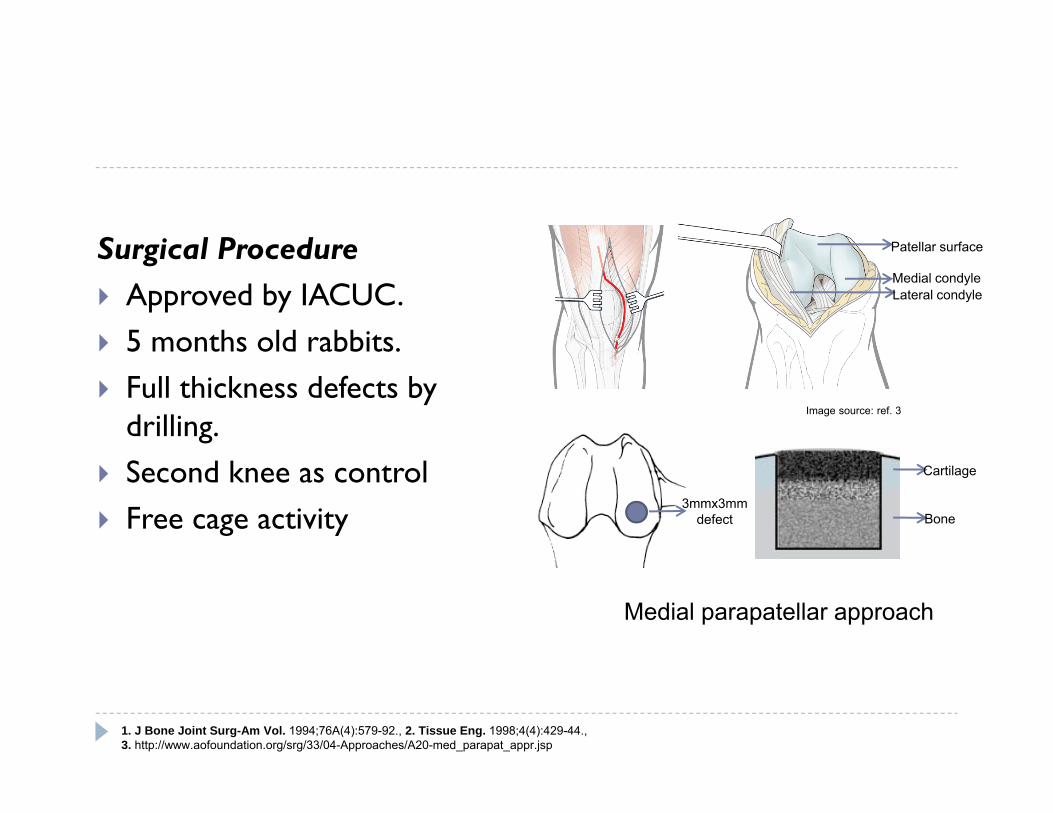
Surgical Procedure Approved by IACUC. 5 months old rabbits. Full thickness defects by
drilling. Second knee as control Free cage activity
1. J Bone Joint Surg-Am Vol. 1994;76A(4):579-92., 2. Tissue Eng. 1998;4(4):429-44., 3. http://www.aofoundation.org/srg/33/04-Approaches/A20-med_parapat_appr.jsp
3mmx3mmdefect
Patellar surface
Medial condyleLateral condyle
Cartilage
Bone
Medial parapatellar approach
Image source: ref. 3
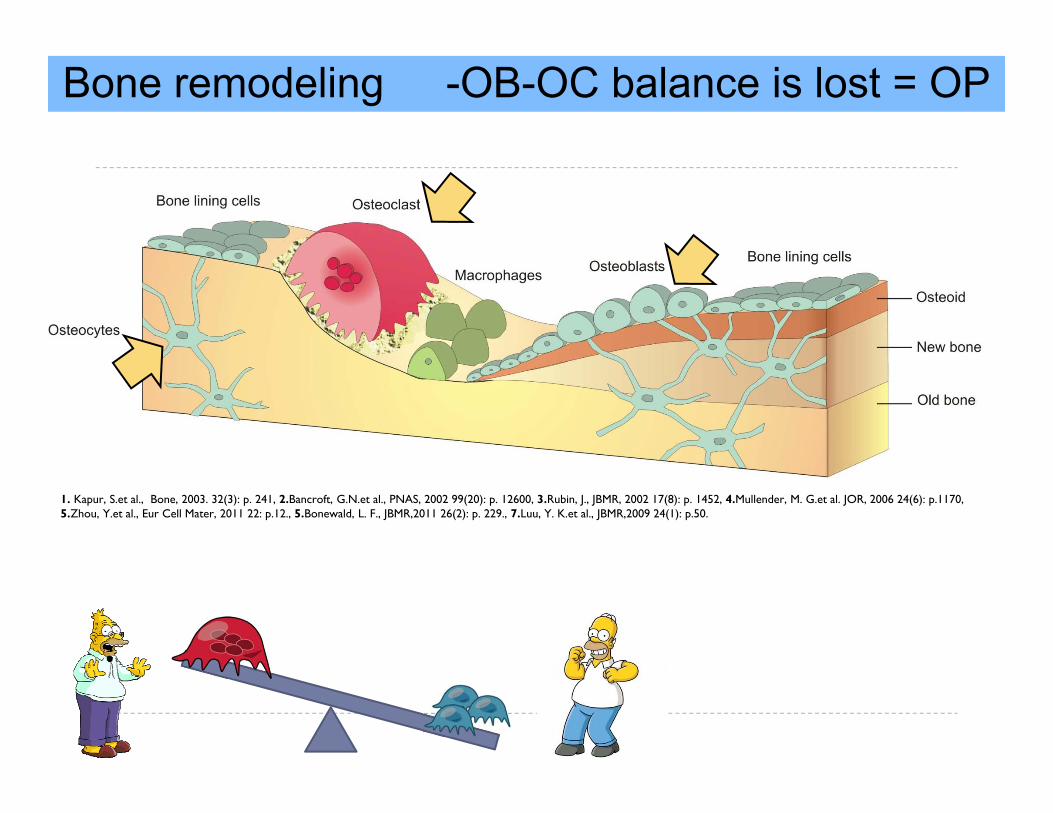
Bone remodeling -OB-OC balance is lost = OP
1. Kapur, S.et al., Bone, 2003. 32(3): p. 241, 2.Bancroft, G.N.et al., PNAS, 2002 99(20): p. 12600, 3.Rubin, J., JBMR, 2002 17(8): p. 1452, 4.Mullender, M. G.et al. JOR, 2006 24(6): p.1170, 5.Zhou, Y.et al., Eur Cell Mater, 2011 22: p.12., 5.Bonewald, L. F., JBMR,2011 26(2): p. 229., 7.Luu, Y. K.et al., JBMR,2009 24(1): p.50.
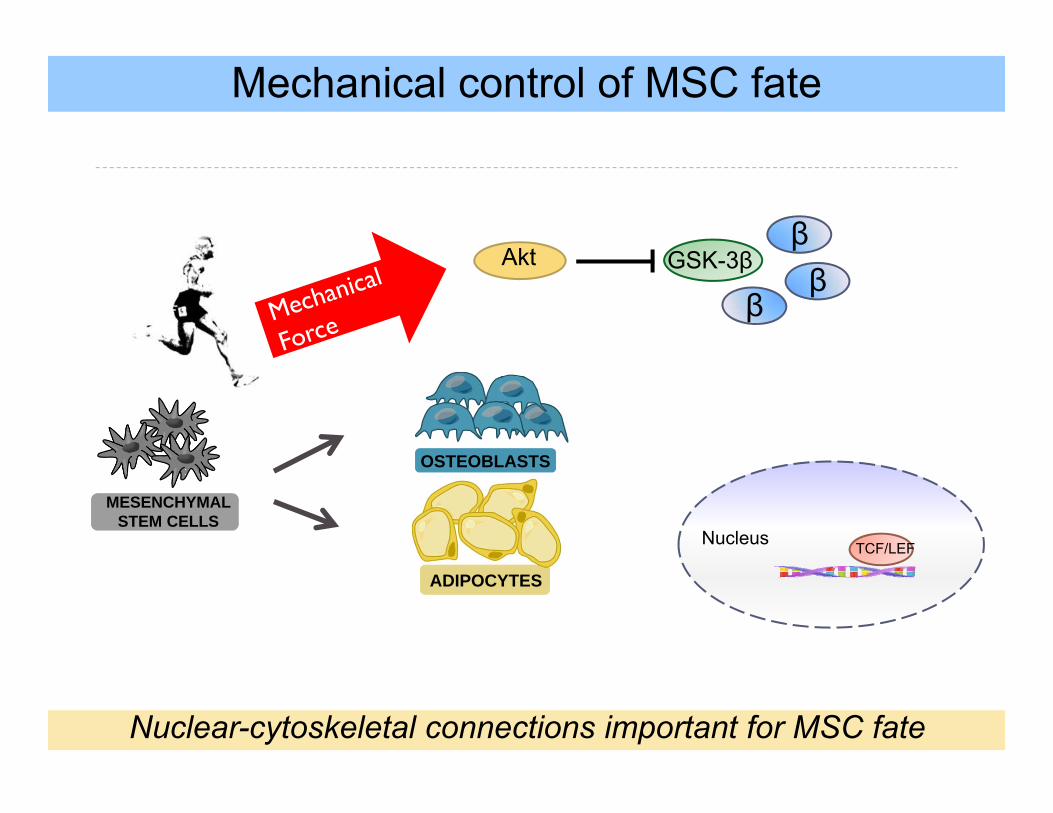
S
GSK-3βAktβ
Mechanical control of MSC fate
MESENCHYMAL STEM CELLS
OSTEOBLASTS
ADIPOCYTES
β
Nuclear-cytoskeletal connections important for MSC fate
TCF/LEFNucleus

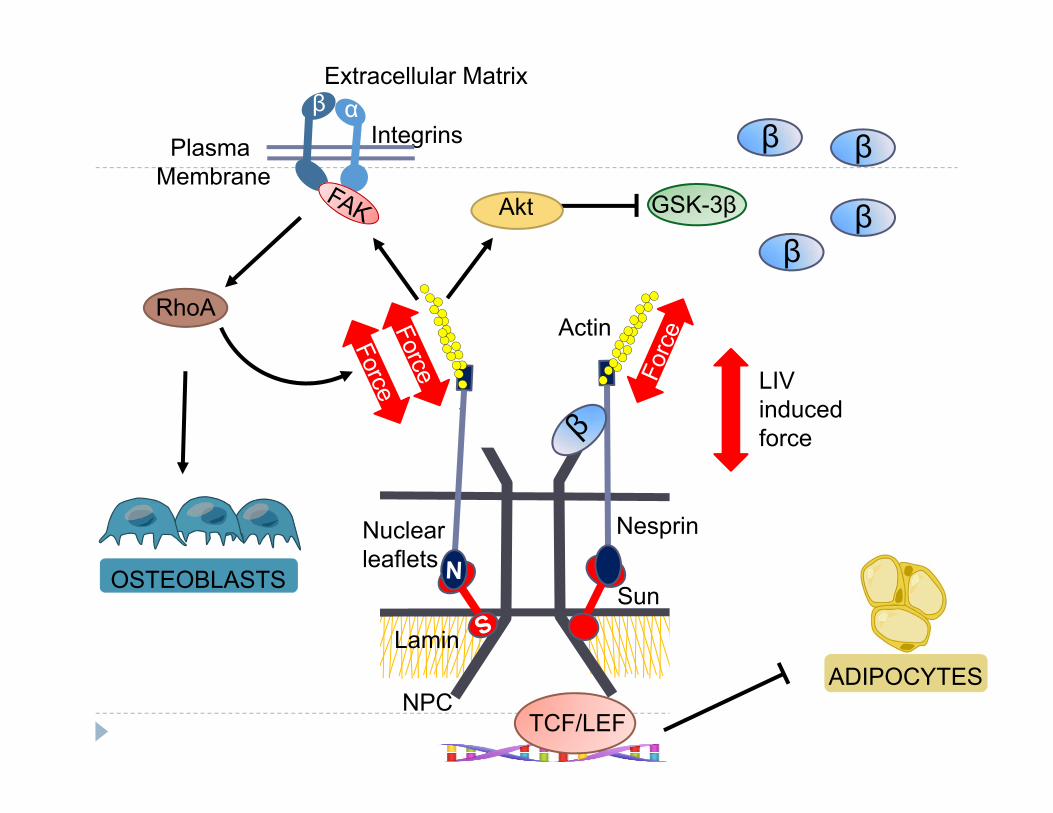
S
NPC
Lamin
Nuclearleaflets
Actin
Nesprin
Sun
Plasma Membrane
Integrins
Extracellular Matrixβ α
GSK-3β
STCF/LEF
β
Akt
β
ββ
LIV induced force
RhoA
ADIPOCYTES
OSTEOBLASTS

Actin
Force
Mechanical regulation of LINC & nucleoskeleton
15 rpm
1-D ClinostatUddin et al. PLoS One, 2013