
BAB II KAJIAN PUSTAKA 2.1 Selulosamedia.unpad.ac.id/thesis/230110/2009/230110090080_2… · ·...
22
6 BAB II KAJIAN PUSTAKA 2.1 Selulosa Selulosa adalah polimer glukosa yang berbentuk rantai linier dan dihubungkan oleh ikatan β-1,4 glikosidik. Struktur yang linier menyebabkan selulosa bersifat kristalin dan tidak mudah larut. Selulosa tidak mudah didegradasi secara kimia maupun mekanis. Di alam, biasanya selulosa berasosiasi dengan polisakarida lain seperti hemiselulosa atau lignin membentuk kerangka utama dinding sel tumbuhan (Holtzapple et.al 2003). Unit penyusun (building block) selulosa adalah selobiosa karena unit keterulangan dalam molekul selulosa adalah 2 unit gula (D-glukosa). Selulosa adalah senyawa yang tidak larut di dalam air dan ditemukan pada dinding sel tumbuhan terutama pada tangkai, batang, dahan, dan semua bagian berkayu dari jaringan tumbuhan. Selulosa merupakan polisakarida struktural yang berfungsi untuk memberikan perlindungan, bentuk, dan penyangga terhadap sel, dan jaringan (Lehninger 1993) (Gambar 1). Selulosa tidak pernah ditemukan dalam keadaan murni di alam, tetapi selalu berasosiasi dengan polisakarida lain seperti lignin, pectin, hemiselulosa, dan xilan (Goyskor dan Eriksen 1980 dalam Fitriani 2003). Kebanyakan selulosa berasosiasi dengan lignin sehingga sering disebut sebagai lignoselulosa. Selulosa, hemiselulosa dan lignin dihasilkan dari proses fotosintesis. Di dalam tumbuhan molekul selulosa tersusun dalam bentuk fibril yang terdiri atas beberapa molekul paralel yang dihubungkan oleh ikatan glikosidik sehingga sulit diuraikan (Goyskor dan Eriksen 1980 dalam Fitriani 2003). Komponen-komponen tersebut dapat diuraikan oleh aktifitas mikroorganisme. Beberapa mikroorganisme mampu menghidrolisis selulosa untuk digunakan sebagai sumber energi, seperti bakteri dan fungi (Sukumaran et.al 2005).
Transcript of BAB II KAJIAN PUSTAKA 2.1 Selulosamedia.unpad.ac.id/thesis/230110/2009/230110090080_2… · ·...

6
BAB II
KAJIAN PUSTAKA
2.1 Selulosa
Selulosa adalah polimer glukosa yang berbentuk rantai linier dan
dihubungkan oleh ikatan β-1,4 glikosidik. Struktur yang linier menyebabkan
selulosa bersifat kristalin dan tidak mudah larut. Selulosa tidak mudah didegradasi
secara kimia maupun mekanis. Di alam, biasanya selulosa berasosiasi dengan
polisakarida lain seperti hemiselulosa atau lignin membentuk kerangka utama
dinding sel tumbuhan (Holtzapple et.al 2003).
Unit penyusun (building block) selulosa adalah selobiosa karena unit
keterulangan dalam molekul selulosa adalah 2 unit gula (D-glukosa). Selulosa
adalah senyawa yang tidak larut di dalam air dan ditemukan pada dinding sel
tumbuhan terutama pada tangkai, batang, dahan, dan semua bagian berkayu dari
jaringan tumbuhan. Selulosa merupakan polisakarida struktural yang berfungsi
untuk memberikan perlindungan, bentuk, dan penyangga terhadap sel, dan
jaringan (Lehninger 1993) (Gambar 1).
Selulosa tidak pernah ditemukan dalam keadaan murni di alam, tetapi
selalu berasosiasi dengan polisakarida lain seperti lignin, pectin, hemiselulosa,
dan xilan (Goyskor dan Eriksen 1980 dalam Fitriani 2003). Kebanyakan selulosa
berasosiasi dengan lignin sehingga sering disebut sebagai lignoselulosa. Selulosa,
hemiselulosa dan lignin dihasilkan dari proses fotosintesis. Di dalam tumbuhan
molekul selulosa tersusun dalam bentuk fibril yang terdiri atas beberapa molekul
paralel yang dihubungkan oleh ikatan glikosidik sehingga sulit diuraikan
(Goyskor dan Eriksen 1980 dalam Fitriani 2003). Komponen-komponen tersebut
dapat diuraikan oleh aktifitas mikroorganisme. Beberapa mikroorganisme mampu
menghidrolisis selulosa untuk digunakan sebagai sumber energi, seperti bakteri
dan fungi (Sukumaran et.al 2005).

7
Gambar 1. Struktur Kimia Selulosa
(Sumber : Lehninger 1993).
Rantai selulosa terdiri dari satuan glukosa anhidrida yang saling berikatan
melalui atom karbon pertama dan ke empat. Ikatan yang terjadi adalah ikatan ß-
1,4-glikosidik. Secara alamiah molekul-molekul selulosa tersusun dalam bentuk
fibril-fibril yang terdiri dari beberapa molekul selulosa yang dihubungkan dengan
ikatan glikosidik. Fibril-fibril ini membentuk struktur kristal yang dibungkus oleh
lignin. Komposisi kimia dan struktur yang demikian membuat kebanyakan bahan
yang mengandung selulosa bersifat kuat dan keras. Sifat kuat dan keras yang
dimiliki oleh sebagian besar bahan berselulosa membuat bahan tersebut tahan
terhadap peruraian secara enzimatik. Secara alamiah peruraian selulosa
berlangsung sangat lambat (Fan et al., 1982).
Berdasarkan derajat polimerisasi dan kelarutan dalam senyawa natrium
hidroksida (NaOH) 17,5%, selulosa dapat dibedakan atas tiga jenis yaitu :
1. Selulosa α (Alpha Cellulose) adalah selulosa berantai panjang, tidak larut
dalam larutan NaOH 17,5% atau larutan basa kuat dengan derajat polimerisasi
600 - 1500. Selulosa α dipakai sebagai penduga dan atau penentu tingkat
kemurnian selulosa. Selulosa α merupakan kualitas selulosa yang paling
tinggi (murni). Selulosa α > 92% memenuhi syarat untuk digunakan
sebagai bahan baku utama pembuatan propelan dan atau bahan peledak,
sedangkan selulosa kualitas dibawahnya digunakan sebagai bahan baku
pada industri kertas dan industri sandang/kain. Semakin tinggi kadar alfa
selulosa, maka semakin baik mutu bahannya (Nuringtyas 2010) (Gambar
2).
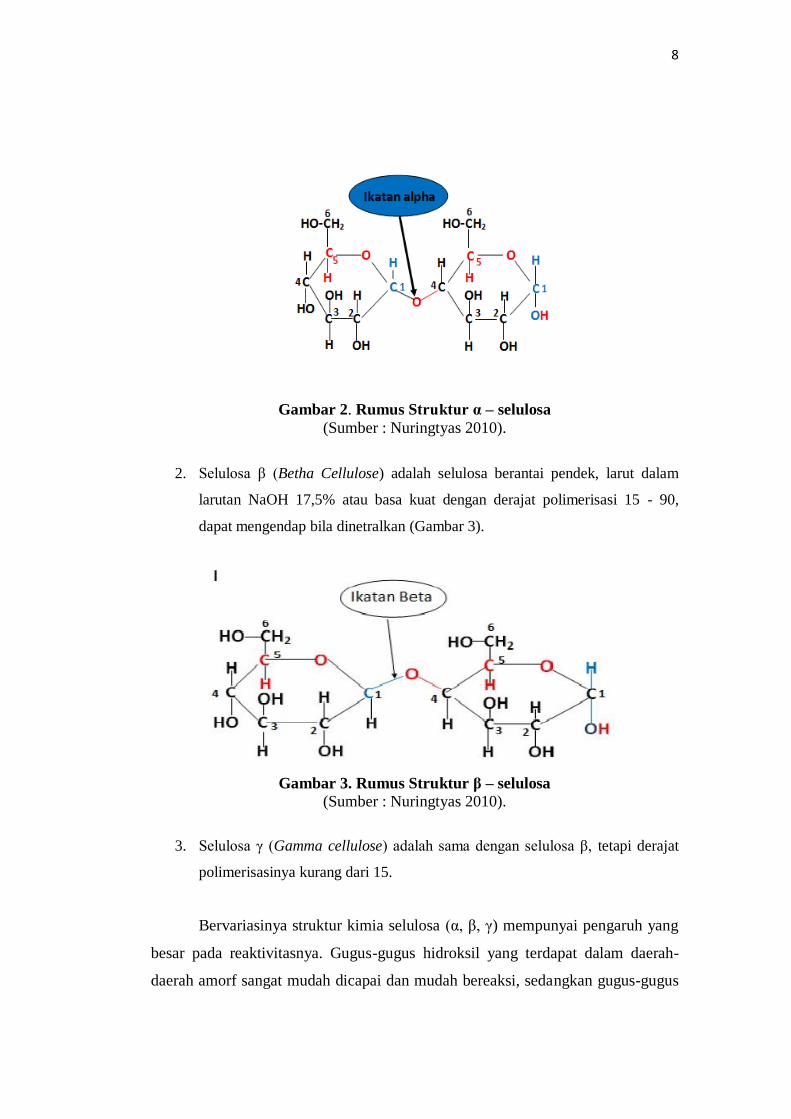
8
Gambar 2. Rumus Struktur α – selulosa
(Sumber : Nuringtyas 2010).
2. Selulosa β (Betha Cellulose) adalah selulosa berantai pendek, larut dalam
larutan NaOH 17,5% atau basa kuat dengan derajat polimerisasi 15 - 90,
dapat mengendap bila dinetralkan (Gambar 3).
Gambar 3. Rumus Struktur β – selulosa (Sumber : Nuringtyas 2010).
3. Selulosa γ (Gamma cellulose) adalah sama dengan selulosa β, tetapi derajat
polimerisasinya kurang dari 15.
Bervariasinya struktur kimia selulosa (α, β, γ) mempunyai pengaruh yang
besar pada reaktivitasnya. Gugus-gugus hidroksil yang terdapat dalam daerah-
daerah amorf sangat mudah dicapai dan mudah bereaksi, sedangkan gugus-gugus

9
hidroksil yang terdapat dalam daerah-daerah kristalin dengan berkas yang rapat
dan ikatan antar rantai yang kuat mungkin tidak dapat dicapai sama sekali.
Pembengkakan awal selulosa diperlukan baik dalam eterifikasi (alkali) maupun
dalam esterfikasi (asam) (Sjőstrőm 1995).
Selulosa memiliki struktur yang unik karena kecenderungannya
membentuk ikatan hidrogen yang kuat. Ikatan hidrogen intramolekular terbentuk
antara: (1) gugus hidroksil C3 pada unit glukosa dan atom O cincin piranosa yang
terdapat pada unit glukosa terdekat, (2) gugus hidroksil pada C2 dan atom O pada
C6 unit glukosa tetangganya. Ikatan hidrogen antarmolekul terbentuk antara
gugus hidroksil C6 dan atom O pada C3 di sepanjang sumbu b (Gambar 4).
Dengan adanya ikatan hidrogen serta gaya van der Waals yang terbentuk, maka
struktur selulosa dapat tersusun secara teratur dan membentuk daerah kristalin. Di
samping itu, juga terbentuk rangkaian struktur yang tidak tersusun secara teratur
yang akan membentuk daerah nonkristalin atau amorf. Semakin tinggi packing
density-nya maka selulosa akan berbentuk kristal, sedangkan semakin rendah
packing density maka selulosa akan berbentuk amorf. Derajat kristalinitas selulosa
dipengaruhi oleh sumber dan perlakuan yang diberikan. Rantai-rantai selulosa
akan bergabung menjadi satu kesatuan membentuk mikrofibril, bagian kristalin
akan bergabung dengan bagian nonkristalin. Mikrofibril-mikrofibril akan
bergabung membentuk fibril, selanjutnya gabungan fibril akan membentuk serat
(Klemm 1998). (Gambar 5).

10
Gambar 4. Ikatan Hidrogen Intra dan Antar Rantai Selulosa
(Sumber : Klemm 1998).
Gambar 5. Model Fibril Struktur Supramolekul Selulosa (Sumber : Klemm 1998).
Selulosa dapat dikonversi menjadi produk-produk bernilai ekonomis yang
lebih tinggi seperti glukosa dan etanol dengan jalan menghidrolisis selulosa
dengan bantuan selulase sebagai biokatalisator atau dengan hidrolisis secara

11
asam/basa (Ariestaningtyas 1991). Selulosa terdapat pada semua tanaman dari
pohon tingkat tinggi hingga organisme primitif seperti rumput laut, flagelata, dan
bakteria (Fengel and Wegener 1995). Luthfy (1988) menyebutkan bahwa rumput
laut jenis Eucheuma sp ternyata mengandung kadar abu 19,92 %, protein 2,80 %,
lemak 1,78 %, serat kasar 7,02 % dan mengandung karbohidrat yang cukup tinggi
yaitu sekitar 68,48 %. Kandungan karbohidrat yang tinggi menjadikan makroalga
jenis Eucheuma sp berpotensi sebagai bahan dasar pembuatan bioetanol.
Makroalga jenis Sargassum sp mulai dikembangkan sebagai bahan
pembuatan etanol. Salah satu negara yang mengembangkannya adalah Jepang.
Untuk mengembangkan pembuatan bioetanol berbahan dasar Sargassum sp
Jepang membuat proyek bernama Ocean Sunrise Project yang bertujuan untuk
memproduksi bioetanol dari rumput laut Sargassum horneri (Maulana dan
Wibowo 2011).
2.2 Enzim Selulase
Enzim adalah katalis biologis/ senyawa yang mempercepat proses reaksi
tanpa ikut bereaksi dalam suatu reaksi kimia organik. Seluruh enzim adalah
protein. Molekul awal yang disebut substrat akan dipercepat perubahannya
menjadi molekul lain yang disebut produk. Jenis produk yang akan dihasilkan
bergantung pada suatu kondisi/ zat yang disebut promoter. Enzim bekerja dengan
cara bereaksi dengan molekul substrat untuk menghasilkan senyawa intermediat
melalui suatu reaksi kimia organik yang membutuhkan energi aktivasi lebih
rendah, sehingga percepatan reaksi kimia terjadi karena reaksi kimia dengan
energi aktivasi lebih tinggi, membutuhkan waktu lebih lama (Palmer 1995).
Salah satu jenis enzim yang dapat menghidrolisis ikatan β(1-4) pada
selulosa adalah enzim selulase. Enzim selulase merupakan enzim yang memegang
peranan penting dalam proses biokonversi limbah-limbah organik berselulosa
menjadi glukosa, protein sel tunggal, makanan ternak, etanol dan lain-lain (Chalal
1983).
Selulase merupakan enzim ekstraseluler yang terdiri atas kompleks endo-
β-1,4-glukonase (CMCase, Cx selulase endoselulase, atau carboxymethyl

12
cellulase) , kompleks ekso-β-1,4-glukonase (aviselase, selobiohidrolase, C1
selulase), dan β-1,4-glukosidase atau selobiase (Meryandini dkk., 2009).
Enzim selulase atau enzim yang dikenal dengan nama sistematik β-1,4
glukan-4-glukano hidrolase adalah enzim yang dapat menghidrolisis selulosa
dengan memutus ikatan glikosidik β-1,4 dalam selulosa, selodektrin, selobiosa,
dan turunan selulosa lainnya menjadi gula sederhana atau glukosa. Sistem
pemecahan selulosa menjadi glukosa terdiri atas tiga jenis enzim selulase yaitu
endo-β-1,4-glukanase, ekso-β-1,4-glukanase, dan β-glukosidase. (Silva et al.
2005). Proses pemecahan selulosa oleh enzim selulase ditunjukkan pada (Gambar
6).
Gambar 6. Skema Tahap-Tahap Pemecahan Selulosa
(Sumber : Nugraha 2006)
Gambar 6 memperlihatkan tahap-tahap pemecahan selulosa oleh kompleks
enzim selulase (endoglukanase, eksoglukanase, dan β-glukosidase). Tahap

13
pertama, enzim endoglukonase menyerang daerah amorf dari selulosa secara acak
dan membentuk makin banyak ujung-ujung nonpereduksi yang memudahkan
kerja eksoglukonase. Enzim eksoglukonase selanjutnya menghidrolisis daerah
kristal dari selulosa dengan membebaskan dua unit glukosa. Kerja sama kedua
enzim ini menghasilkan unit-unit sakarida yang lebih kecil yang selanjutnya
dihidrolisis oleh β-glukosidae menghasilkan glukosa (Nugraha 2006).
Menurut Enari (1983) (Tabel 1) serta Prescott and Dunns (1981) (Gambar
7) mengelompokkan enzim utama selulase berdasarkan spesifikasi substrat
masing-masing enzim yaitu :
1. Endo-β-1,4-glukanase (β-1,4-D-glukan-4-glukanohidrolase, EC 3.2.1.4)
menghidrolisis ikatan glikosidik β-1,4 secara acak. Enzim ini dapat bereaksi
dengan selulosa kristal tetapi kurang aktif. Enzim ini secara umum dikenal
sebagai CMC-ase atau selulase Cx.
2. β-1,4-D-glukan selobiohidrolase (EC.3.2.1.91) atau secara umum dikenal
dengan selulase C1, memutus ujung rantai selulosa non pereduksi dan
membebaskan selobiosa.
3. β-1,4-D-glukan glukohidrolase (EC.3.2.1.74) memutus ujung rantai selulosa
non pereduksi dan membebaskan glukosa. Enzim ini menghidrolisis selulosa yang
telah dilunakkan dengan asam fosfat, selo-oligosakarida dan CMC.
4. β-1,4-glikosidase (β-1,4-D-glukosida glukohidrolase, EC 3.2.1.21)
menghidrolisis selobiosa dan rantai pendek selo-oligosakarida yang menghasilkan
glukosa. Enzim ini tidak dapat memecah selulosa dan selodekstrin.
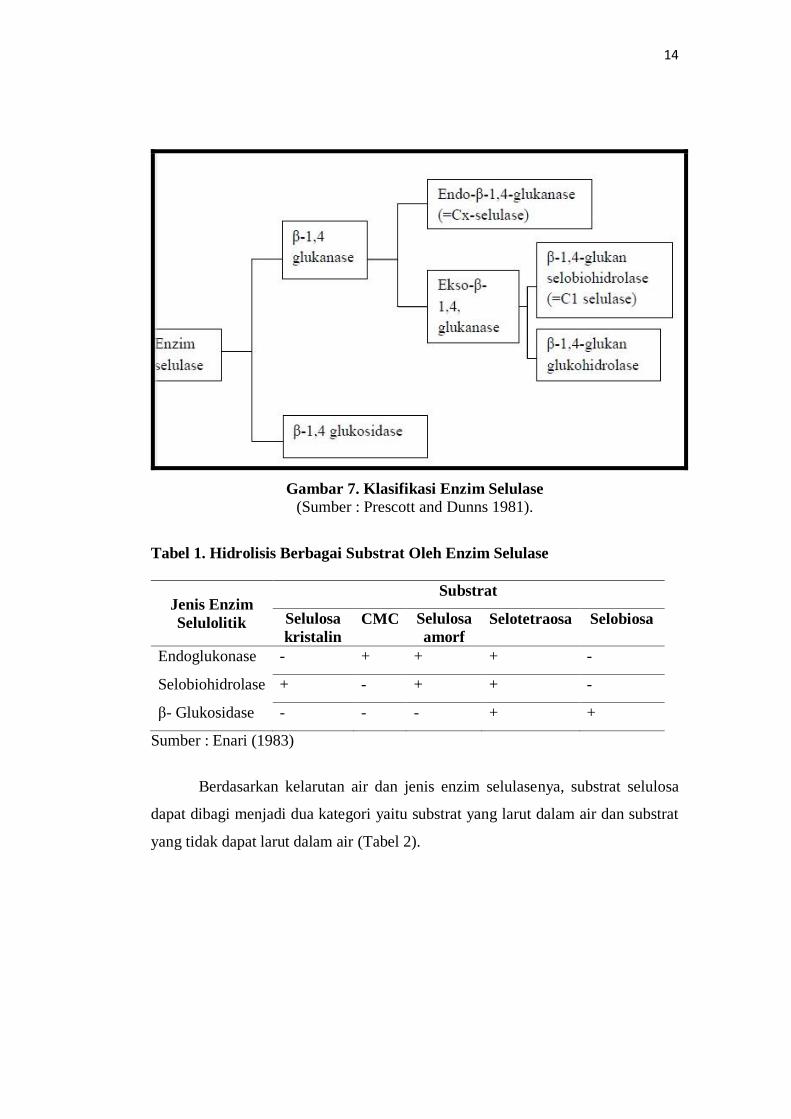
14
Gambar 7. Klasifikasi Enzim Selulase (Sumber : Prescott and Dunns 1981).
Tabel 1. Hidrolisis Berbagai Substrat Oleh Enzim Selulase
Jenis Enzim
Selulolitik
Substrat
Selulosa
kristalin
CMC Selulosa
amorf
Selotetraosa Selobiosa
Endoglukonase - + + + -
Selobiohidrolase + - + + -
β- Glukosidase - - - + +
Sumber : Enari (1983)
Berdasarkan kelarutan air dan jenis enzim selulasenya, substrat selulosa
dapat dibagi menjadi dua kategori yaitu substrat yang larut dalam air dan substrat
yang tidak dapat larut dalam air (Tabel 2).
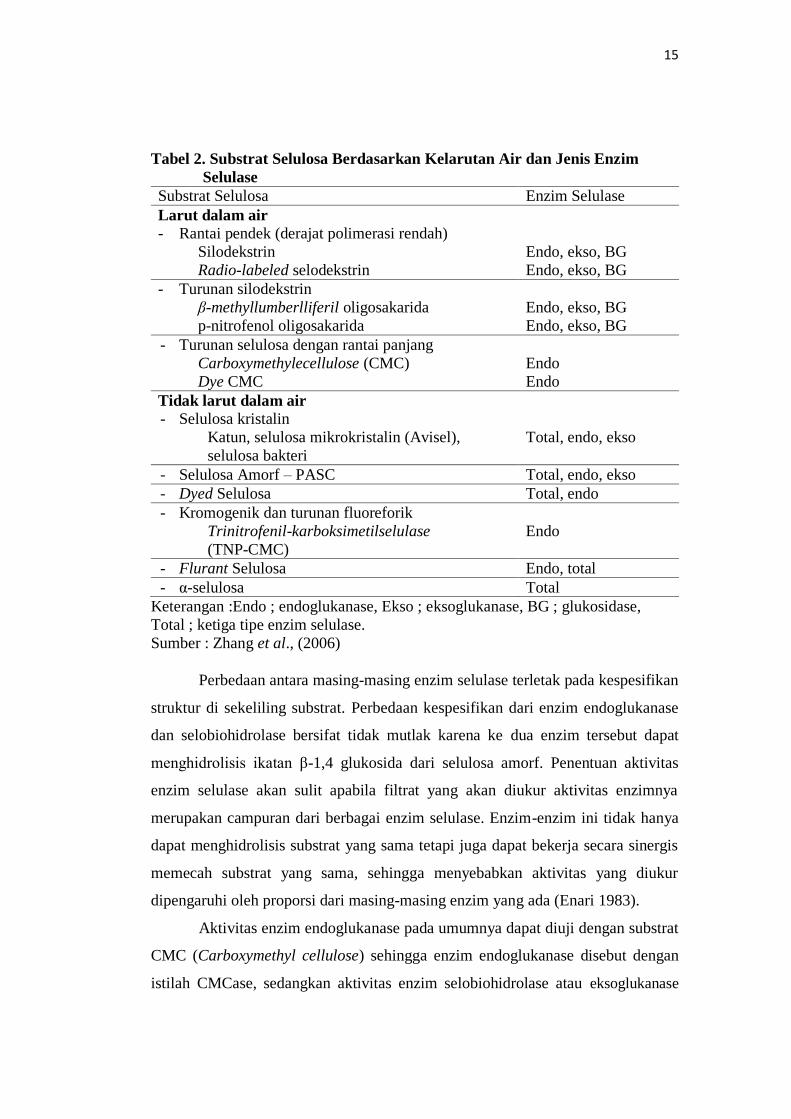
15
Tabel 2. Substrat Selulosa Berdasarkan Kelarutan Air dan Jenis Enzim
Selulase
Substrat Selulosa Enzim Selulase
Larut dalam air
- Rantai pendek (derajat polimerasi rendah)
Silodekstrin Endo, ekso, BG
Radio-labeled selodekstrin Endo, ekso, BG
- Turunan silodekstrin
β-methyllumberlliferil oligosakarida Endo, ekso, BG
p-nitrofenol oligosakarida Endo, ekso, BG
- Turunan selulosa dengan rantai panjang
Carboxymethylecellulose (CMC) Endo
Dye CMC Endo
Tidak larut dalam air
- Selulosa kristalin
Katun, selulosa mikrokristalin (Avisel), Total, endo, ekso
selulosa bakteri
- Selulosa Amorf – PASC Total, endo, ekso
- Dyed Selulosa Total, endo
- Kromogenik dan turunan fluoreforik
Trinitrofenil-karboksimetilselulase Endo
(TNP-CMC)
- Flurant Selulosa Endo, total
- α-selulosa Total
Keterangan :Endo ; endoglukanase, Ekso ; eksoglukanase, BG ; glukosidase,
Total ; ketiga tipe enzim selulase.
Sumber : Zhang et al., (2006)
Perbedaan antara masing-masing enzim selulase terletak pada kespesifikan
struktur di sekeliling substrat. Perbedaan kespesifikan dari enzim endoglukanase
dan selobiohidrolase bersifat tidak mutlak karena ke dua enzim tersebut dapat
menghidrolisis ikatan β-1,4 glukosida dari selulosa amorf. Penentuan aktivitas
enzim selulase akan sulit apabila filtrat yang akan diukur aktivitas enzimnya
merupakan campuran dari berbagai enzim selulase. Enzim-enzim ini tidak hanya
dapat menghidrolisis substrat yang sama tetapi juga dapat bekerja secara sinergis
memecah substrat yang sama, sehingga menyebabkan aktivitas yang diukur
dipengaruhi oleh proporsi dari masing-masing enzim yang ada (Enari 1983).
Aktivitas enzim endoglukanase pada umumnya dapat diuji dengan substrat
CMC (Carboxymethyl cellulose) sehingga enzim endoglukanase disebut dengan
istilah CMCase, sedangkan aktivitas enzim selobiohidrolase atau eksoglukanase

16
seringkali diuji dengan substrat avisel sehingga enzim eksoglukanase disebut dengan
aviselase (Zhang et al., 2006).
Tahapan hidrolisis selulosa tergantung kepada struktur selulosa, interaksi
antara enzim selulase dengan serat selulosa, mekanisme hidrolisis enzim tersebut
di alam dan inhibitor yang terbentuk. Fase adsorbsi dan pembentukan kompleks
enzim substrat adalah fase kritis di dalam hidrolisis selulosa. Glukosa dan
selobiosa adalah inhibitor enzim dalam menghidrolisis selulosa. Selobiosa
menghambat enzim selobiohidrolase dan glukosa menghambat enzim
penghidrolisis selobiosa yaitu β-glukosidase pada kompleks enzim selulase.
Selobiosa mempunyai potensi lebih kuat menjadi inhibitor dibandingkan dengan
glukosa (Coughlan 1985). Laju hidrolisis enzim selulase ditentukan oleh struktur
substrat. Struktur kristal lebih sulit dihidrolisis dibandingkan dengan struktur
amorf maka hidrolisis dilakukan oleh enzim endoselulase atau endoglukanase
(Coughlan 1985).
Faktor yang mempengaruhi aktivitas selulase yaitu adanya senyawa
penghambat berupa ion logam. Penghambatan tersebut dapat dinetralkan dengan
menambahkan sistein sehingga aktivitas enzim dapat berlangsung kembali (Kulp
1975). Beberapa senyawa logam dan senyawa lainnya yang dapat menghambat
aktivitas selulase ialah Hg2+, Ag2+, dan Cu2+ (Deng and Tabatai 1994; Oikawa
et al. 1994), glukanolakton (Kulp 1975), surfaktan, senyawa pengkelat khususnya
Sodium Dodecyl Sulphate (SDS), Ethylene Diamine Tetraacetyc Acid (EDTA)
(Oikawa et al. 1994), laktat dalam konsentrasi agak rendah (Chesson 1987), dan
etanol serta alkohol lainnya (Ooshima et al., 1985). Senyawa penghambat tersebut
dapat menekan seluruh kecepatan hidrolisis dengan menghambat adsorbsi
eksoglukanase dan endoglukanase pada selulosa, dan menghambat aksi sinergis
eksoglukanase dan endoglukanase yang bekerja pada permukaan selulosa.
2.3 Bakteri Laut
Sekitar 80 persen dari spesies bakteri laut telah teridentifikasi dan
diketahui sebagai bakteri kelompok basil gram negatif. Dari pengamatan terhadap
koloni yang ditumbuhkan pada cawan petri maupun pemeriksaan mikroskopis

17
secara langsung dari bahan laut yang diwarnai dengan metode Gram, diperkirakan
sekitar 95 persen dari bakteri laut adalah gram negatif. Hal ini menunjukkan
perbedaan yang cukup besar dibandingkan dengan persentasi bakteri gram negatif
yang ditemukan di perairan tawar dan dua kali jumlah dari gram negatif yang
ditemukan di dalam tanah. Sebagian besar bakteri yang ditemukan di laut
termasuk motil aktif (Zobell 1990).
Bakteri pembentuk spora juga diketahui tidak banyak ditemukan di
lingkungan laut. Bakteri pembentuk spora yang ditemukan di lingkungan laut di
antaranya Bacillus abysseus, B.borborokoites, B.cirroflagellosus, B.filicolonicus,
B.imomarinus, B.submarinus, B.thalassokoites, dan B.epiphyticus. Secara rata-
rata, ukuran bakteri laut lebih kecil dibandingkan bakteri yang terdapat di dalam
susu, air limbah, air tawar, atau tanah .Rata-rata panjang ukuran sel adalah 2
sampai 3 µm dan lebar ukuran sel rata-rata 0,4 - 0,6 µm (Zobell 1990).
Secara umum, bakteri laut tumbuh lebih lambat dan koloninya lebih kecil
dibandingkan kebanyakan mikroorganisme dari tanah, limbah, dan susu. Pada
pengamatan pertumbuhan bakteri, nampak bahwa bakteri terestrial sudah
mencapai pertumbuhan optimum pada usia kultur tujuh hari, sedangkan pada
bakteri laut pada usia kultur 10 hari masih tampak kenaikan pertumbuhan (Zobell
1990).
Ciri-ciri bakteri yang tumbuh di lingkungan perairan, termasuk bakteri laut
adalah bersifat kromogenik, yaitu mampu menghasilkan beragam pigmen.
Distribusi warna adalah sebagai berikut: 31,3 persen kuning, 15,2 persen oranye,
9,9 persen kecoklatan, 7,4 persen neon, 5.4 persen merah atau pink, dan 0,2
persen hijau. Jenis paling umum yang berpendar berwarna kehijauan. Bakteri laut
yang mempunyai pigmen berwarna kuning diantaranya Flavohaclerium
marinotypicum, F.marinovirosum, Fl.okeanokoites, Pseudomonas neritica, Ps.
obscura, Ps.oceanica, Ps.vadosa, Ps.xanthochrus, Vibrio adaptatus,
V.marinoflavus, dan V.marinovulgaris (Zobell 1990).
Bakteri yang hidup di lingkungan laut pada umumnya mampu
mendegradasi hampir semua jenis substrat organik, dan banyak senyawa
anorganik yang diubah oleh aktifitas mikroorganisme laut. Sebagian besar bakteri

18
laut memiliki aktifitas yang cukup tinggi sebagai agen proteolitik, namun sedikit
yang merupakan agen sakarolitik. Semua bakteri heterotrofik laut mampu
menyerap glukosa, hanya 46 dari 60 kultur yang di fermentasi glukosa dengan
pembentukan asam, dan tidak satupun dari mereka menghasilkan "gas" dari
glukosa. Hal ini mungkin karena kurangnya kemampuan fermentasi, tetapi lebih
besar kemungkinan karena efisiensi dari organisme dalam asimilasi bahan
organik. Bila dikultivasi dalam media cair, bakteri laut mampu mengkonversi 30
sampai 35 persen glukosa pada protoplasma atau produk-produk metabolit,
sisanya yang teroksidasi menjadi karbon dioksida dan air sebagai sumber energi
(Zobell 1990).
Bakteri laut dapat dibedakan dari bakteri terestrial berdasarkan
toleransinya terhadap garam. Pada penelitian yang telah dilakukan diketahui
bahwa bakteri laut mampu mentolerir keberadaan garam hingga konsentrasi diatas
100% dalam medium. Kebutuhan salinitas dari bakteri laut terkait dengan adaptasi
fisiologis sel terhadap lingkungan berkadar garam tinggi. Bakteri genus Bacillus
dan Micrococcus yang diisolasi dari lingkungan laut diketahui memiliki tingkat
toleransi salinitas yang lebih tinggi bila dibandingkan dengan genus Pseudomonas
atau Vibrio (Zobell 1990).
2.4 Bakteri Selulolitik
Mikroorganisme yang mampu mendegradasi selulosa dinamakan
mikroorganisme selulolitik. Bakteri yang dapat mendegradasi selulosa disebut
juga bakteri selulolitik. Bakteri selulolitik memiliki kemampuan dalam
menghidrolisis bahan-bahan dari alam yang mengandung selulosa menjadi produk
yang lebih sederhana (Marganingtyas 2011).
Beberapa jenis bakteri yang dapat mendegradasi selulosa adalah
Chaetonium sp, Chytophaga sp, dan Clostridium sp (Rao dalam Nurmayani
2007). Sebagai contoh, dalam beberapa penelitian yang menggunakan bakteri
jenis Clostridium sp untuk produksi beberapa pelarut antara lain dinyatakan Ezeji
et.al (2007) memproduksi aseton, butanol dan etanol dari tepung jagung dengan
kadar 14,28 g/L.

19
“Cellulose-decomposing bacteria” atau bakteri selulolitik merupakan
suatu komunitas bakteri yang hidup pada bahan yang mengandung selulosa di
lingkungan laut yang mempunyai kemampuan untuk mendegradasi selulosa.
Bahan selulosa yang berada di lingkungan laut misalnya limbah kertas, limbah
galangan kapal, kapas, linen, dan selulosa hasil pengendapan organisme laut
berupa sedimen laut (Zobell 1990). Kebanyakan bakteri laut merupakan bakteri
halofil, bakteri ini memerlukan NaCl untuk pertumbuhan optimumnya dan
tumbuh paling baik pada konsentrasi garam 2,5-4,0%, selain itu juga bersifat
fakultatif anaerob yang tumbuh lebih baik pada ketersediaan oksigen. Haigler dan
Weiner (1991) menyatakan bahwa bakteri selulolitik yang sering dijumpai di laut
tropik adalah bakteri yang hidup pada suhu sekitar 350C-40
0C, yaitu genus
Pseudomonas dan Cytophaga. Rheinheimer (1994) dalam Handayani (2002)
menyatakan bahwa jenis-jenis bakteri laut yang berperan dalam dekomposisi
senyawa karbohidrat adalah jenis bakteri selulolitik aerob seperti Pseudomonas sp
dan Bacillus sp serta bakteri selulolitik anaerob seperti Clostridium sp, sedangkan
spesies-spesies bakteri selulolitik dari genus Cytophaga dan Sporocytophaga
paling berperan dalam mendegradasi selulosa.
Bakteri selulolitik dapat mendegradasi molekul komplek pada substrat
tidak larut dalam air dengan menggunakan berbagai enzim melalui berbagai cara
di dalam memutuskan bagian yang berbeda di dalam substrat. Proses perombakan
secara enzimatis terjadi dengan adanya enzim selulase sebagai agen perombak
yang bersifat spesifik untuk menghidrolisis ikatan β-(1,4)-glikosidik, rantai
selulosa dan derivatnya (Ambriyanto 2010).
Hidrolisis sempurna selulosa akan menghasilkan monomer selulosa yaitu
glukosa, sedangkan hidrolisis tidak sempurna akan menghasilkan disakarida dari
selulosa yaitu selobiosa (Fan et.al 1982). Selulosa dapat dihidrolisis menjadi
glukosa dengan menggunakan media air dan dibantu dengan katalis asam atau
enzim. Selanjutnya glukosa yang dihasilkan dapat difermentasi menjadi menjadi
produk fermentasi yang nantinya dapat diolah lagi menjadi bioetanol. Umumnya
degradasi selulosa terjadi pada pH normal (Hatami et al., 2008).

20
2.5 Penapisan Gen dengan Metode PCR
PCR adalah reaksi berantai suatu primer dari urutan (sequence) DNA
dengan bantuan enzym polymerase sehingga terjadi amplifikasi DNA target
secara in vitro. Teknik PCR ditemukan oleh Dr. Kary Mullis pada tahun 1985 dan
mendapatkan hadiah Nobel atas temuannya pada tahun 1993 (Sunarto 2006).
Pada prinsipnya alat PCR merupakan semacam inkubator yang mampu
mengatur/mengubah suhu dengan sangat cepat sesuai dengan setting atau
program yang dimasukkan. Alat PCR sendiri tidak mampu melakukan
amplifikasi DNA, tetapi setting suhu didalamnya merupakan prasyarat bagi
berlangsungnya reaksi amplifikasi. Amplifikasi akan berlangsung bila komponen
– komponennya telah dimasukkan. Adapun komponen yang dibutuhkan pada
proses amplifikasi diantaranya adalah :
a. Deoxyribonucleotide triphosphate (dNTP) yang memberikan energi dan
nukleosida untuk sintesis DNA. dNTP terdiri atas 4 macam sesuai dengan basa
penyusun DNA, yaitu dATP, dCTP, dGTP dan dTTP.
b. DNA polymerase yang akan memanjangkan primer yang telah menempel pada
cetakan. Biasanya digunakan enzym Taq Polymerase yang tahan terhadap
suhu tinggi.
c. Magnesium Chlorida (MgCl2) sebagai kofaktor dalam proses PCR.
d. PCR Buffer yang biasanya terdiri atas bahan-bahan kimia untuk
mengkondisikan reaksi agar berjalan optimum dan menstabilkan enzim DNA
polymerase.
e. Sepasang primer yang terdiri dari primer F (forward) dan R (reverse) yang
akan menentukan urutan (sequence) DNA yang akan disintesa. Primer yang
digunakan bisa satu pasang ( 2 buah) atau 2 pasang (4 buah).
f. Template yaitu DNA target yang diekstraksi dari sampel dan berfungsi sebagai
bahan cetakan DNA yang diperbanyak (amplifikasi) (Sunarto 2006).
Proses amplifikasi DNA dalam mesin PCR terdiri dari tiga proses utama.
Pertama, denaturasi (pemisahan segmen DNA ganda menjadi DNA untai tunggal)
yang berlangsung pada suhu 94o - 96
oC, kemudian yang kedua adalah terjadinya
penempelan primer (annealing) yang berlangsung pada suhu 55o
- 65 o
C dan

21
proses yang ketiga adalah terjadinya pemanjangan dari primer yang telah
menempel (polimerasi) pada suhu 72o - 75
oC (Setiawan 2003).
Menurut Nur’utami (2011) tiga tahap utama kerja PCR dalam satu siklus
yaitu :
a. Denaturation (Pemecahan)
Tahap pemecahan berlangsung pada suhu tinggi, 94o - 96
oC ikatan
hidrogen DNA terputus dari DNA utas ganda menjadi utas tunggal. Biasanya
pada siklus awal PCR, tahap ini dilakukan lebih lama (5 menit) untuk
memastikan semua berkas DNA terpisah. Denaturation menyebabkan DNA tidak
stabil dan siap menjadi template (patokan) bagi primer. Umumnya waktu durasi
yang dibutuhkan yaitu selama 1 sampai 2 menit.
b. Annealing (penempelan)
Primer menempel pada bagian template DNA yang berkomplementer
urutan basanya. Tahap ini dilakukan pada suhu antara 46o - 60
oC. Penempelan ini
bersifat spesifik. Suhu yang tidak tepat menyebabkan tidak terjadinya
penempelan atau primer menempel di sembarang tempat. Umumnya waktu/durasi
yang dibutuhkan untuk tahap ini yaitu selama 1 sampai 2 menit.
c. Extension (pemanjangan)
Suhu untuk proses ini tergantung dari jenis enzim DNA polimerase yang
dipakai. Karena suhu dinaikkan ke dalam kisaran suhu optimum kerja enzim
DNA polimerase. Dengan Taq polimerase, proses ini biasanya dilakukan pada
suhu 70o
- 76 o
C. Pada tahap ini DNA polimerase akan memasangkan dNTP yang
sesuai pada pasangannya (pasangan A adalah T, dan C dengan G, begitu pula
sebaliknya). Enzim akan memperpanjang rantai baru ini hingga ke ujung
(Gambar 8a). Lamanya waktu ekstensi bergantung pada panjang daerah yang
akan diamplifikasi, misalnya adalah 1 menit untuk setiap 1000 basepair (bp).
Selain ketiga proses tersebut umumnya tahap kerja PCR didahului dan
diakhiri oleh tahapan berikut :
a. Pra-denaturasi : Dilakukan selama 1 sampai 9 menit pada awal reaksi
untuk memastikan kesempurnaan denaturasi dan mengaktivasi DNA
polimerase (jenis hot-start/aktif jika dipanaskan terlebih dahulu).

22
b. Final Elongasi : Umumnya dilakukan pada suhu optimum enzim (70o
- 72
oC). Tahap final elongasi dilakukan selama 5 sampai 15 menit untuk
memastikan bahwa setiap utas tunggal yang tersisa sudah diperpanjang secara
sempurna. Proses final elongasi dilakukan setelah PCR terakhir.
Karena hasil sintesa DNA dalam satu siklus dapat berperan sebagai
cetakan (template) pada siklus berikutnya, maka jumlah DNA target menjadi
berlipat dua pada setiap akhir siklus (Gambar 8b), dengan kata lain DNA target
meningkat secara eksponensial, sehingga setelah 30 siklus akan dihasilkan
milyaran (230
) amplifikasi DNA target. DNA target yang telah berlipat ganda
jumlahnya dapat dideteksi dengan elektroforesis gel agarosa. Setelah diwarnai
dengan Ethidium Bromide (EtBr), hasil elektroforesis yang berupa pita DNA
dapat dilihat dengan alat UV trans-iluminator (Contoh Gambar 9) dan foto
dengan kamera Polaroid atau sistem dokumentasi gel yang lain (Sunarto 2006).
a b
Gambar 8. Prinsip dan Cara Kerja PCR
(Sumber : Sunarto 2006)
Tahapan kerja PCR konvensional terdiri dari isolasi DNA/RNA
(ekstraksi), amplifikasi menggunakan mesin thermal cycler, visualisasi DNA
(elektroforesis) menggunakan tangki elektroforesis dan dokumentasi (Astuti
2011).

23
Gambar 9. Elektroforegram Amplifikasi Fragmen Penyandi Kitinase
Bakteri Kitinolitik
(Sumber : Agustin 2013)
Keunggulan metode PCR dapat dilihat dalam hal kecepatan, spesifitas dan
sensitifitasnya untuk mendeteksi mikroorganisme patogen. PCR sangat spesifik
karena mendeteksi mikroorganisme pada tingkat DNA/RNA. DNA/RNA virus
dapat diketahui secara cepat dan sampel yang akan diuji dapat dalam jumlah yang
banyak. Penggunaan teknik PCR juga memiliki beberapa kelemahan antara lain
kemungkinan memperoleh hasil positif maupun negatif yang salah. Hasil positif
yang keliru dapat disebabkan karena adanya kontaminasi oleh kontrol positif
maupun sampel yang lain sebelum proses amplifikasi. Hasil negatif yang salah
dapat disebabkan karena jumlah kopian DNA yang tidak mencukupi dan tingkat
infeksi yang terlalu rendah sehingga pita DNA berpendar redup atau bahkan tidak
berpendar (Sukenda et.al 2009).
2.6 Primer
Primer adalah untai DNA pendek yang terdiri atas beberapa nukleotida,
umumnya 10-25 nukleotida (oligonukleotida). Primer berperan sebagai pemula
pada proses sintesis menggunakan PCR. Primer akan menempel pada DNA target
dan membentuk rangkaian komplementer pada untai DNA yang berlawanan serta

24
mengapit rangkaian sasaran (DNA cetakan). Konsentrasi primer untuk proses
PCR menurut William et al (1990) dalam Ain (2011) berkisar antara 0.1-0.5 μM.
Desain primer merupakan salah satu prasyarat sebelum melakukan proses
pengkopian gen dengan PCR. Primer PCR biasanya berukuran panjang 20-30 bp,
Ujung 5' pada primer menentukan ujung produk PCR yang dihasilkan. Primer
yang ideal memiliki kandungan GC vs. AT yang seimbang (45-55% GC), dan
tanpa untaian satu basa yang terlalu panjang. Dua primer dari pasangan primer
tidak mengandung struktur komplemen lebih dari 2 kb. Jarak amplifikasi atau
sekuens target yang diamplifikasi mempunyai ukuran sekitar 200-400 bp (David
2005).
Menurut Innis and Gelfand (1990), Syarat sebuah primer dalam PCR
adalah :
1. Ukuran primer berkisar antara 17-28 basa
2. Komposisi basa adalah 50-60% (G+C)
3. Tm (suhu leleh) berkisar antara 55-80oC.
4. Primer PCR tidak boleh memiliki ujung (3') berupa G atau C, atau CG atau GC.
Hal ini menghindari terjadinya "breathing" pada ujung dan meningkatkan efisiensi
priming;
5. Tiga atau lebih sekuen C atau G pada ujung 3' primer dapat memicu terjadinya
kesalahan priming pada sekuen yang kaya akan G dan C (karena stabilitas
annealing), sehingga harus dihindari.
6. Ujung 3' primer bukan merupakan komplementer (pasangan basa), karena akan
mengakibatkan terjadinya dimer primer.
7. Primer reverse dan primer forward harus dihindari saling berkomplemen,
sehingga saling dapat membentuk ikatan atau menyatu.
Primer biasanya terdiri dari 10-20 nukleotida dan dirancang berdasarkan
daerah konservatif dalam genom tersebut. Semakin panjang primer, maka
semakin spesifik daerah yang diamplifikasi. Jika suatu kelompok organisme
memang berkerabat dekat, maka primer dapat digunakan untuk mengamplifikasi
daerah tertentu yang sama dalam genom kelompok tersebut. Beberapa faktor

25
seperti konsentrasi DNA contoh, ukuran panjang primer, komposisi basa primer,
konsentrasi ion Mg, dan suhu hibridisasi primer harus dikontrol dengan hati-hati
agar dapat diperoleh pita-pita DNA yang utuh dan baik. Keberhasilan teknik ini
lebih didasarkan kepada kesesuaian primer dan efisiensi dan optimasi proses PCR.
Primer yang tidak spesifik dapat menyebabkan teramplifikasinya daerah lain
dalam genom yang tidak dijadikan sasaran atau sebaliknya tidak ada daerah
genom yang teramplifikasi (Rybicki 1996).
Optimasi PCR juga diperlukan untuk menghasilkan karakter yang
diinginkan. Optimasi ini menyangkut suhu denaturasi dan annealing DNA dalam
mesin PCR. Suhu denaturasi yang rendah dapat menyebabkan belum terbukanya
DNA utas ganda sehingga tidak dimungkinkan terjadinya polimerisasi DNA baru.
Proses penempelan primer pada utas DNA yang sudah terbuka memerlukan suhu
optimum, sebab suhu yang terlalu tinggi dapat menyebabkan amplifikasi tidak
terjadi atau sebaliknya suhu yang terlalu rendah menyebabkan primer menempel
pada sisi lain genom yang bukan sisi homolognya; akibatnya dapat teramplifikasi
banyak daerah tidak spesifik dalam genom tersebut. Suhu penempelan (annealing)
ini ditentukan berdasarkan primer yang digunakan yang dipengaruhi oleh panjang
dan komposisi primer. Suhu penempelan ini sebaiknya sekitar 50C di bawah suhu
leleh. Tm merupakan temperatur yang diperlukan oleh separuh primer dupleks
mengalami disosiasi/lepas ikatan (President's DNA Initiative 2013). Secara umum
suhu leleh (Tm) dihitung dengan rumus Tm = 4(G+C) + 2(A+T)0C (Rybicky
1996), sedangkan suhu annealing (Ta) dapat dihitung dengan rumus Ta = 0.3 x
Tm (primer) + 0.7 Tm (produk) – 25 (President's DNA Initiative 2013)
Primer-primer degenerate adalah campuran oligonukleotida yang
bervariasi dalam hal urutan basa nukleotidanya, tetapi mempunyai panjang atau
jumlah nukleotida yang sama. Campuran oligonukleotida tersebut dapat
digunakan sebagai primer dalam PCR untuk melakukan amplifikasi suatu gen atau
fragmen DNA tertentu. Primer-primer degenerate tersebut berguna dalam PCR
jika ingin mengamplifikasikan suatu fragmen DNA yang belum banyak diketahui
urutan nukleotidanya sehingga primer yang spesifik tidak dapat dibuat. Primer-
primer semacam itu juga berguna jika ingin melakukan amplifikasi suatu gen

26
yang belum diketahui urutan nukleotidanya tetapi gen tersebut termasuk dalam
kelompok famili gen (gene family) tertentu. Dengan menggunakan informasi
mengenai urutan nukleotida gen lain yang termasuk dalam famili yang sama maka
dapat menyintesis primer-primer degenerate untuk mengamplifikasi gen yang
belum diketahui urutannya tersebut. Di samping itu, primer-primer degenerate
juga dapat digunakan untuk melakukan amplifikasi anggota-anggota suatu famili
gen untuk identifikasi atau analisis selanjutnya (Yuwono 2006).
2.7 Karakterisasi In Silico
Karakterisasi In Silico merupakan salah satu kegiatan penggunaan
perangkat bioinformatik untuk mengidentifikasi karakter-karakter pada suatu
sekuen gen, diantaranya domain fungsional, sisi aktif, motif protein, analisis
homologi, konstruksi pohon filogenetik dan sebagainya. Sebuah aspek penting
dari karakterisasi sekuen biologis adalah identifikasi motif dan domain. Ini
merupakan cara penting untuk mengkarakterisasi fungsi protein karena sekuen
protein baru sering tidak signifikan dengan database sekuen yang ada sehingga
sulit untuk diketahui. Dalam hal ini, para ahli biologi dapat mendefinisikan fungsi
protein berdasarkan identifikasi sekuen konsensus pendek yang telah diketahui
fungsinya. Pola sekuen konsensus ini disebut sebagai motif dan/atau domain
(Xiong 2006).
Motif adalah suatu pola sekuen lestari yang berukuran pendek yang
berkaitan dengan perbedaan fungsi dari suatu protein atau DNA. Sebuah motif
yang khas, seperti motif zinc finger (jari-jari zinc) memiliki sepuluh sampai dua
puluh asam amino. Sebuah domain juga merupakan sekuen lestari, yang
didefinisikan sebagai suatu unit fungsional dan struktural yang independen.
Domain biasanya lebih panjang dari motif. Sebuah domain itu terdiri lebih dari 40
residu dan sampai 700 residu, dengan rata-rata panjang 100 residu. Sebuah
domain dapat memiliki suatu bagian motif didalamnya dapat pula tidak
mengandung motif. Contoh domain adalah domain transmembran dan domain
yang berikatan dengan ligan. Motif dan domain secara evolusi lebih lestari
dibandingkan dengan daerah lain pada suatu protein dan memiliki kecenderungan

27
untuk berevolusi melalui penambahan atau pengurangan suatu unit tertentu pada
modul domain tersebut. Identifikasi motif dan domain pada suatu protein
merupakan aspek penting pada proses klasifikasi sekuen protein dan annotasi
fungsional. Dikarenakan aspek divergensi pada evolusi, hubungan fungsional
diantara protein-protein seringkali tidak dapat dibedakan begitu saja melalui
BLAST atau pencarian database FASTA. Selain itu, protein atau enzim terkadang
memiliki fungsi ganda yang tidak dapat sepenuhnya dijelaskan menggunakan
annotasi tunggal melalui pencarian database sekuen sehingga diperlukan
identifikasi motif dan domain untuk mengatasi hal tersebut (Xiong 2006).









![Arab Journal of Nuclear Sciences and Applications · M. DIAA EL-DIN H. FARAG et.al and polysaccharides and high levels of bioactive compounds [6]. Pomegranate peel, being free from](https://static.fdocument.org/doc/165x107/604ffea56941bf0b4c2e87d8/arab-journal-of-nuclear-sciences-and-applications-m-diaa-el-din-h-farag-etal.jpg)
