
Aus dem Institut für Anatomie ΙΙ -...
74
Aus dem Institut für Anatomie ΙΙ: Experimentelle Morphologie (Direktor: Prof. Dr. med. Udo Schumacher) des Zentrums für Experimentelle Medizin des Universitätsklinikums Hamburg-Eppendorf Die Wirkung des Proteasomeninhibitors Bortezomib auf humane Neuroblastomzellen in vitro und in vivo in einem Xenograftmodell Dissertation zur Erlangung des Doktorgrades der Medizin der Medizinischen Fakultät der Universität Hamburg vorgelegt von Christina Haane aus Dorsten Hamburg 2007
Transcript of Aus dem Institut für Anatomie ΙΙ -...

Aus dem Institut für Anatomie ΙΙ:
Experimentelle Morphologie
(Direktor: Prof. Dr. med. Udo Schumacher)
des Zentrums für Experimentelle Medizin
des Universitätsklinikums Hamburg-Eppendorf
Die Wirkung des Proteasomeninhibitors Bortezomib auf
humane Neuroblastomzellen in vitro und in vivo in einem Xenograftmodell
Dissertation
zur Erlangung des Doktorgrades der Medizin
der Medizinischen Fakultät der Universität Hamburg vorgelegt von
Christina Haane
aus Dorsten
Hamburg 2007

Angenommen von der Medizinischen Fakultät
der Universität Hamburg am: 01.12.2008
Veröffentlicht mit Genehmigung der Medizinischen
Fakultät der Universität Hamburg
Prüfungsausschuss, der Vorsitzende: Prof. Dr. U. Schumacher
Prüfungsausschuss: 2. Gutachter: Prof. Dr. H.-E. Laack
Prüfungsausschuss: 3. Gutachterin: PD Dr. S. Klutmann

Meinen Eltern

Abkürzungsverzeichnis
I
Abkürzungsverzeichnis
Abb. Abbildung
Alu Arthrobacter luteus
bzw. beziehungsweise
ca. circa
CASY Cellcounter Analyser System
CLL Chronisch lymphatische Leukämie
CT Computertomographie
°C Grad Celsius
ddH2O double distilled H2O
evtl. eventuell
[18 F]FDG Fluordeoxyglukose
[18F]FLT Fluorothymidin
g Gramm
GPOH Gesellschaft für Pädiatrische Onkologie und Hämatologie
h Stunde
HE Hämatoxylin-Eosin
HPLC High Performance Liquid Chromatography
INSS International Neuroblastoma Staging System
i.v. intravenous
kg Kilogramm
KG Körpergewicht
KV Kilo Volt
nm nanometer
nM nano Mol
Nr. Nummer
n.s. nicht signifikant
min Minuten
MHz Megahertz
MRT Magnetresonanztomographie
µm micrometer
* p < 0,05
** p < 0,01
*** p < 0,001

Abkürzungsverzeichnis
II
PCR Polymerasekettenreaktion
PET Positronenemissionstomographie
s Sekunden
s.c. subcutan
scid Severe combined immune deficiency
SD Standard deviation
SEM Standard error of the mean
Tab. Tabelle
UKE Universitätsklinikum Hamburg-Eppendorf
Voxel Volumenenlement (dreidimensionales Pixel)
z.B. zum Beispiel
z.T. zum Teil

Inhaltsverzeichnis
1
Inhaltsverzeichnis 1
1 Arbeitshypothese und Fragestellung 3
2 Einleitung 4
2.1 Das humane Neuroblastom 4
2.2 Bortezomib 8
3 Material und Methoden 10
3.1 Neuroblastomzelllinien 10
3.2 Bortezomib 10
3.3 Charakterisierung von Bortezomib in vitro 10
3.3.1 Zellproliferationsassay 10
3.3.1.1 Zellkultur 10
3.3.1.2 Detektion von Mykoplasmen in der Zellkultur 11
3.3.1.3 Zellzählung 12
3.3.1.4 Erstellen von Eichkurven 12
3.3.1.5 Zellproliferationsassay mit Zusatz von Bortezomib 13
3.4 Charakterisierung von Bortezomib in vivo 14
3.4.1 Versuchstiere 14
3.4.2 Behandlung der Tiere 15
3.4.3 Histologie 16
3.4.3.1 Primärtumoren 16
3.4.3.1.1 Lichtmikroskopie 16
3.4.3.1.2 Elektronenmikroskopie 17
3.4.3.2 Lungenmetastasierung 17
3.4.4 Disseminierte Tumorzellen im Blut 18
3.4.5 Imaging 19
3.4.5.1 Magnetresonanztomographie 19

Inhaltsverzeichnis
2
3.4.5.2 Positronen-Emissions-Tomographie /
Computer Tomographie 20
3.5 Statistische Auswertung 22
4 Ergebnisse 23
4.1 Wirkung von Bortezomib in vitro 23
4.2 Wirkung von Bortezomib in vivo 28
4.2.1 Primärtumor 28
4.2.1.1 Lichtmikroskopie 29
4.2.1.2 Elektronenmikroskopie 33
4.2.2 Lungenmetastasierung 35
4.2.3 Disseminierte Tumorzellen im Blut 36
4.2.4 Imaging 39
4.2.4.1 MRT 39
4.2.4.2 PET-CT 43
5 Diskussion 47
6 Zusammenfassung 59
7 Literaturverzeichnis 60
Danksagung 67
Lebenslauf 68
Selbstständigkeitserklärung 69

Arbeitshypothese und Fragestellung
3
1 Arbeitshypothese und Fragestellung
Bei Kindern mit der Diagnose Neuroblastom ist die Prognose häufig infaust, so dass ein
großes Interesse für neue Therapieansätze besteht. Besonders bei dem großen Anteil der
Patienten, die sich im Stadium IV befinden, ist die Aussicht auf Heilung äußerst gering. Bei
Einsatz des Proteasomeninhibitors Bortezomib wurden bereits antiproliferative Wirkungen bei
verschiedenen Tumorentitäten nachgewiesen. Daher war das Ziel der vorliegenden Arbeit
folgende Fragen zu klären:
• Hat der Proteasomeninhibitor Bortezomib eine wachstumshemmende
Wirkung auf die humanen Neuroblastomzelllinien in vitro?
• Hat das Medikament Bortezomib auch eine antiproliferative Wirkung in vivo,
wenn humane Neuroblastomzellen in scid – Mäuse transplantiert werden?
• Gibt es Unterschiede zwischen der behandelten Bortezomibgruppe und der
Kontrollgruppe in Hinblick auf Apoptosen und Mitosen der Primärtumoren,
sowie der Anzahl der Spontanmetastasen in den Lungen und der Anzahl der
freien Tumorzellen im Blut?
• Können die Imagingverfahren MRT und PET-CT die humanen
Neuroblastome im scid -Mausmodell charakterisieren?

Einleitung
4
2 Einleitung
2.1 Das Neuroblastom
Das Neuroblastom ist mit 7 - 14% aller kindlichen Tumoren der häufigste extrakranielle solide
Tumor des Kindesalters (Morgenstern et al., 2004). In Deutschland liegt die Zahl der
Neuerkrankungen bei 120 - 130 Fällen pro Jahr. Die Inzidenz liegt bei circa 1,3 Erkrankungen
auf 100.000 Kinder, die sich in einem Alter unter 15 Jahren befinden (Hero und Berthold,
2002; Schwab et al., 2003; Simon, 2005). Das Neuroblastom gehört zu den embryonalen
Tumoren, die charakteristischerweise in 90% der Fälle in einem Alter von unter 5 Jahren
diagnostiziert werden (Schwab et al., 2003). Das mittlere Lebensalter der Patienten liegt bei
unter zwei Jahren und 35% der Kinder erkranken bereits in einem Alter von unter einem Jahr
(Morgenstern et al., 2004).
Es handelt sich um eine maligne Erkrankung, die von primitiven sympathischen neuralen
Vorläufern des Nervengewebes ausgeht (Brodeur, 2003). Histologisch findet man zwei
verschiedene Zelltypen, die Neuroblast / Ganglienzelle und die Schwann-Zellen. Letztere sind
vermutlich nicht neoplastisch, werden aber wahrscheinlich als Stromazelle von den
Tumorzellen rekrutiert (Koletzko, 2003).
Aufgrund seiner Histogenese, kann das Neuroblastom überall dort auftreten, wo
sympathisches Nervengewebe zu finden ist. In erster Linie entstehen Neuroblastome deshalb
im Nebennierenmark. Die übrigen Lokalisationen verteilen sich, vom Retroperitonealraum
ausgehend, in den paraspinalen Ganglien von Thorax, Abdomen oder Pelvis (Brodeur, 2003).
Die Ausbreitung des Neuroblastoms erfolgt durch infiltratives Wachstum und durch
lymphogene und hämatogene Metastasierung. Diese erfolgt zumeist in das Knochenmark, in
Fernlymphknoten und in die Leber, seltener in die Haut, ins Gehirn und in die Lunge (Hero
und Berthold, 2002; Simon, 2005). Das Stadium IV-S nimmt eine Sonderstellung ein. Hierbei
ist das Erkrankungsalter auf das erste Lebensjahr begrenzt, es findet sich ein lokalisierter
Primärtumor mit Metastasen, die sich definitionsgemäß allerdings nur im Knochenmark, in der
Haut oder in der Leber befinden dürfen (Hero und Berthold, 2002).
Die betroffenen Kinder leiden sowohl unter Krankheitszeichen, die direkt auf den Tumor
zurückzuführen sind, als auch unter so genannten Allgemeinsymptomen. Entsprechend der
verschiedenen Ursprungsorte kann ein Neuroblastom durch unterschiedliche Symptome
auffallen wie z. B. ein tastbarer Knoten im Bauchraum, Atembeschwerden, Husten und
verdickte Halslymphknoten oder auch Anzeichen einer Querschnittslähmung durch

Einleitung
5
Vorwachsen des Tumors in Richtung Rückenmark. Als Allgemeinsymptome können Blässe,
Gewichtsverlust, Fieber und Durchfall auftreten (Simon et al., 2004; Simon, 2005).
Der Verdacht auf das Vorliegen eines Neuroblastoms ergibt sich aus den typischen
Beschwerden und dem Befund der körperlichen Untersuchung. Häufig wird ein Neuroblastom
aber auch zufällig im Rahmen einer Routineuntersuchung entdeckt. Die Diagnosesicherung
erfolgt durch weiterführende Untersuchungen: Blut- und Urinuntersuchung auf
Katecholaminmetabolite, feingewebliche Beurteilung eines Biopsats, Röntgenuntersuchung
des Brustkorbs, Ultraschalluntersuchung des Bauchraums, Computer- und
Kernspintomographie und Szintigraphie.
Ein System zum Staging des Neuroblastoms stellt das INSS (=International Neuroblastoma
Staging System) dar. Zusätzlich können verschiedene Parameter wie z.B. der Ferritin-Spiegel
bestimmt werden, um die Staging Analyse zu vervollständigen.
Stadium Kriterien I Lokalisierter Tumor mit makroskopisch kompletter Entfernung, repräsentative
ipsi- und contralaterale Lymphknoten histologisch ohne Tumorbefall. Lediglich unmittelbar am Tumor adhärente, chirurgisch entfernte Lymphknoten dürfen positiv sein. Auch bilaterale Tumoren, die makroskopisch komplett exstirpiert werden können und keinen regionalen Lymphknotenbefall aufweisen
IIa Unilateraler Tumor mit makroskopisch inkompletter Resektion, ipsi- und contralaterale Lymphknoten histologisch negativ
IIb Tumor wie bei IIa, jedoch positive regionale ipsilaterale Lymphknoten III Nichtresektabler unilateraler Tumor infiltriert über die Mittellinie mit oder ohne
Lymphknotenbefall oder Nichtresektabler Mittellinientumor mit bilateraler Ausdehnung durch Infiltration oder Lymphknotenbefall (Das Überschreiten der Mittellinie ist definiert durch infiltratives Erreichen / Überschreiten der Wirbelkante der Gegenseite)
IV Tumoraussaat in Lymphknoten, Knochen, Knochenmark, Leber und andere Organe
IV-S Sonderform, nur bei Säuglingen im 1. Lebensjahr, lokalisierter Primärtumor wie bei Stadium I, IIa oder IIb, Tumoraussaat nur in Leber, Haut und/oder Knochenmark
Tab. 1: Internationale Stadieneinteilung des Neuroblastoms nach INSS (Simon, 2005).
Einleitung
6
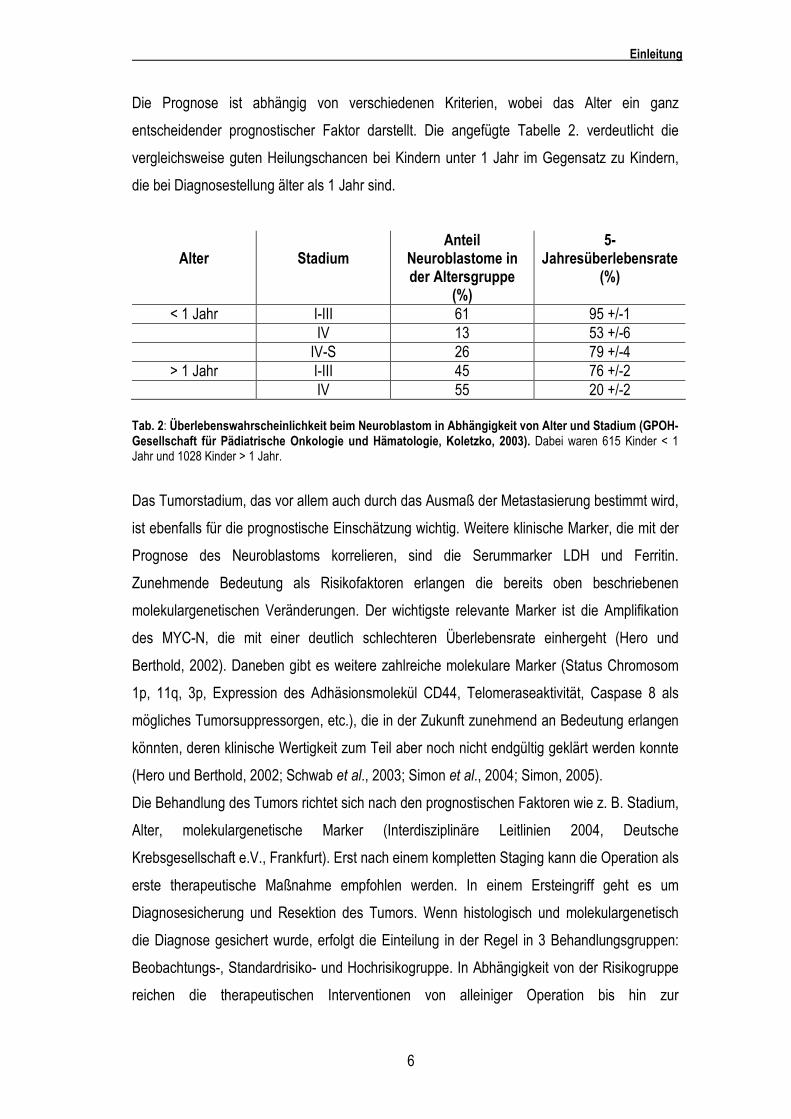
Die Prognose ist abhängig von verschiedenen Kriterien, wobei das Alter ein ganz
entscheidender prognostischer Faktor darstellt. Die angefügte Tabelle 2. verdeutlicht die
vergleichsweise guten Heilungschancen bei Kindern unter 1 Jahr im Gegensatz zu Kindern,
die bei Diagnosestellung älter als 1 Jahr sind.
Alter
Stadium
Anteil Neuroblastome in der Altersgruppe
(%)
5-Jahresüberlebensrate
(%)
< 1 Jahr I-III 61 95 +/-1 IV 13 53 +/-6 IV-S 26 79 +/-4
> 1 Jahr I-III 45 76 +/-2 IV 55 20 +/-2
Tab. 2: Überlebenswahrscheinlichkeit beim Neuroblastom in Abhängigkeit von Alter und Stadium (GPOH- Gesellschaft für Pädiatrische Onkologie und Hämatologie, Koletzko, 2003). Dabei waren 615 Kinder < 1 Jahr und 1028 Kinder > 1 Jahr.
Das Tumorstadium, das vor allem auch durch das Ausmaß der Metastasierung bestimmt wird,
ist ebenfalls für die prognostische Einschätzung wichtig. Weitere klinische Marker, die mit der
Prognose des Neuroblastoms korrelieren, sind die Serummarker LDH und Ferritin.
Zunehmende Bedeutung als Risikofaktoren erlangen die bereits oben beschriebenen
molekulargenetischen Veränderungen. Der wichtigste relevante Marker ist die Amplifikation
des MYC-N, die mit einer deutlich schlechteren Überlebensrate einhergeht (Hero und
Berthold, 2002). Daneben gibt es weitere zahlreiche molekulare Marker (Status Chromosom
1p, 11q, 3p, Expression des Adhäsionsmolekül CD44, Telomeraseaktivität, Caspase 8 als
mögliches Tumorsuppressorgen, etc.), die in der Zukunft zunehmend an Bedeutung erlangen
könnten, deren klinische Wertigkeit zum Teil aber noch nicht endgültig geklärt werden konnte
(Hero und Berthold, 2002; Schwab et al., 2003; Simon et al., 2004; Simon, 2005).
Die Behandlung des Tumors richtet sich nach den prognostischen Faktoren wie z. B. Stadium,
Alter, molekulargenetische Marker (Interdisziplinäre Leitlinien 2004, Deutsche
Krebsgesellschaft e.V., Frankfurt). Erst nach einem kompletten Staging kann die Operation als
erste therapeutische Maßnahme empfohlen werden. In einem Ersteingriff geht es um
Diagnosesicherung und Resektion des Tumors. Wenn histologisch und molekulargenetisch
die Diagnose gesichert wurde, erfolgt die Einteilung in der Regel in 3 Behandlungsgruppen:
Beobachtungs-, Standardrisiko- und Hochrisikogruppe. In Abhängigkeit von der Risikogruppe
reichen die therapeutischen Interventionen von alleiniger Operation bis hin zur

Einleitung
7
Maximaltherapie (Interdisziplinäre Leitlinien: Deutsche Krebsgesellschaft e.V. Frankfurt,
2004).
Das Therapiekonzept Beobachtungsgruppe besteht nach der Operation aus regelmäßigen
Kontrollen und eventuell einer leichten Chemotherapie. Die Patienten mit einem
Standardrisiko bekommen nach der Operation eine Chemotherapie. Je nach Ansprechen im
Therapieverlauf kann eine sekundäre Tumoroperation oder eine Strahlentherapie notwendig
sein. Hingegen werden Patienten mit einem Hochrisiko - Neuroblastom sowohl primär operiert
als auch adjuvant / palliativ mit Chemotherapie und Strahlentherapie behandelt. Je nach
Ansprechen kann eine Second-Look-Operation erfolgen (Simon, 2005; Hero und Berthold,
2002).
Im Rahmen von Tumorstudien erhalten alle Patienten anschließend eine
Hochdosischemotherapie mit autologer Stammzelltransplantation (Simon, 2005). In der Regel
wird die Behandlung der Hochrisikogruppe durch weitere konsolidierende Therapieverfahren
ergänzt, z. B. Immuntherapie mit Antikörpern. Morgenstern et al., (2004) und Brodeur et al.,
(2003) berichteten von einer biologischen Therapie mit cis-Retinsäure, welche gute Resultate
nach autologer Stammzelltransplantation zeigte. Fang et al., (2006) berichteten von H+linked
monocarboxylate transporter (MCT1 / SLC 16A1), die einen Erfolg versprechenden Einfluss
auf das Tumorwachstum von Hochrisikopatienten haben sollen. Simon (2005) wies weiterhin
auf neue Ansätze der Tumorvakzinierung mit dendritischen Zellen hin. All diese
verschiedenen Therapieansätze sind Gegenstand aktueller klinischer Studien.
Die Behandlung von Patienten mit Neuroblastom im Stadium IV-S nimmt eine Sonderstellung
ein, da Tumor und Metastasen oft spontan regredieren. Beim Auftreten von Lebermetastasen
oder Bedrohung durch Tumormassen kann auch hier eine milde Chemotherapie erforderlich
sein (Interdisziplinäre Leitlinien 2004, Deutsche Krebsgesellschaft, e.V. Frankfurt).
Diese Therapieansätze führen aber noch immer nicht zu einem durchschlagenden Erfolg in
der Behandlung des Neuroblastoms. Insbesondere Hochrisikopatienten im Stadium IV haben
eine Fünfjahresüblerlebensrate von 20 ± 2%. Die hohe Mortalität (je nach Stadium bis zu
80%) dieses kindlichen Tumors (Koletzko 2003, Berthold et al., 2005) macht die Dringlichkeit
der Suche nach weiteren Therapieoptionen deutlich. Ein solcher neuer Ansatzpunkt besteht
möglicherweise in der Beeinflussung der Tumorzellen durch den Proteasomeninhibitor
Bortezomib (Velcade ®).

Einleitung
8
2.2 Bortezomib
Bortezomib ist das erste zugelassene Medikament aus der neuen Wirkstoffklasse der
Proteasomeninhibitoren. Bortezomib hemmt spezifisch die Aktivität des ubiquitär
vorkommenden 26S-Proteasoms in Säugetierzellen. Das Proteasom ist ein essentieller
Enzymkomplex, welches Apoptose, Transkription, Zelladhäsion, Angiogenese,
Antigenpräsentation und den Zellzyklus reguliert (Le Blanc, 2002). In vielfachen Studien
wurde gezeigt, dass durch die Inhibition des Proteasoms die Tumorgröße verringert werden
kann. Beispiele hierfür sind das maligne Myelom (Hidshima et al., 2001; Katayoun et al.,
2004), das Mantelzelllymphom (Pham et al., 2003), das nichtkleinzellige Lungenkarzinom
(Ling et al., 2003), das Ovarialkarzinom (Frankel et al., 2000), das Pankreaskarzinom (Shah
et al., 2001; Bold et al., 2001), das Prostatakarzinom (Adams et al., 1999; Frankel et al.,
2000) und bösartige Tumoren im Kopf – Halsbereich (Sunwoo et al. 2001, van Waes et al.,
2005). Neueste Studien von Michaelis et al. (2006), Brignole et al. (2006) und Hamner et al.
(2007) zeigten erstmals antiproliferative Wirkungen auf humane Neuroblastomzelllinien in
vitro und in vivo. Zudem konnte in einer Phase Ι Studie bei pädiatrischen Patienten mit
refraktären soliden Tumoren (inklusive 2 Patienten mit Neuroblastom) gezeigt werden, dass
Bortezomib ein gut tolerables Medikament ist, welches nur eine geringe Toxizität aufweist
(Blaney et al., 2004).
Der molekulare Mechanismus, mit dem Bortezomib die Apoptose der Tumorzellen induziert, ist
weitestgehend unklar. Vermutet wird eine Veränderung der Balance zwischen pro- und
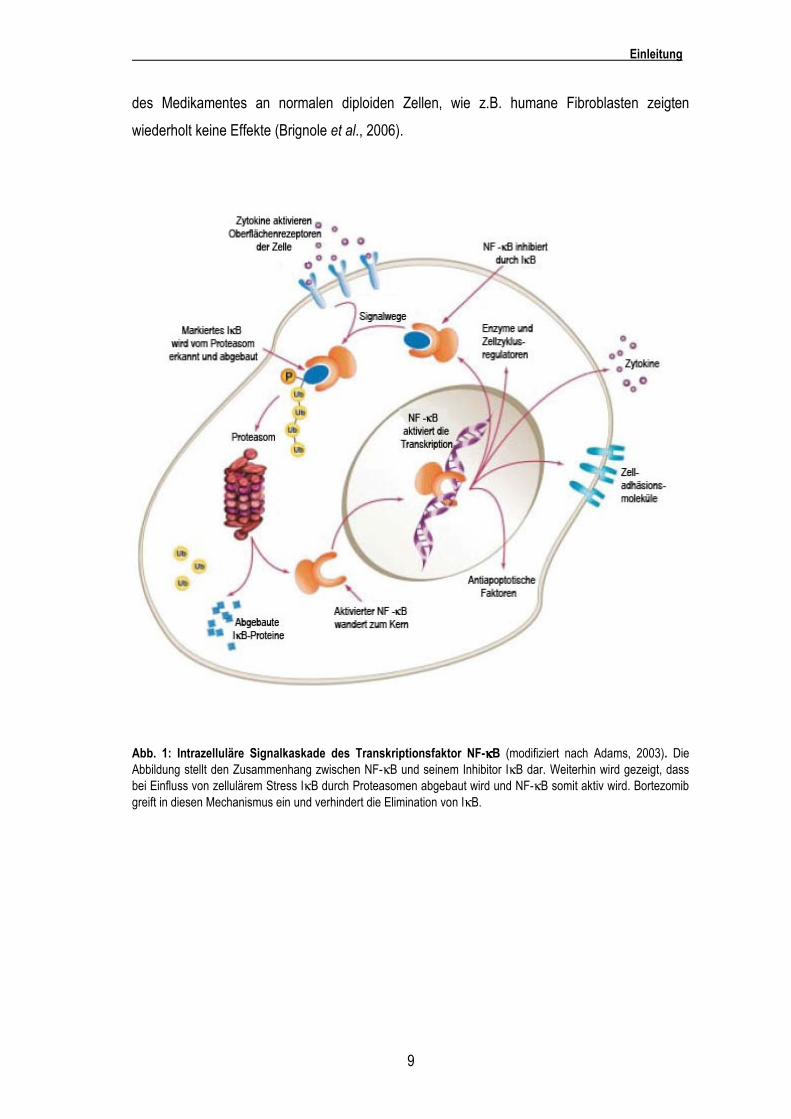
antiapoptotischen Signalen (Adams, 2003). Ein Molekül, das eine zentrale Rolle in der
Vermittlung vieler Effekte des Bortezomibs darstellt, ist der Transkriptionsfaktor NF-κB (Karin et
al., 2002). NF-κB liegt normalerweise im Zytosol im Komplex mit dem Inhibitormolekül IκB vor.
Durch zellulären Stress wird IκB durch die Proteasomen abgebaut, so dass es zur Aktivierung
von NF-κB kommt. Durch Bortezomib kann die Elimination des IκB reduziert werden und
folglich die Aktivität von NF-κB gehemmt werden. Beim Multiplen Myelom konnte weiter
beobachtet werden, dass Bortezomib die IL-6 induzierten Wachstumsstimulationen durch
Verminderung von gp 130 induziert. Dies wiederum führt zu einer vermehrten Aktivität von
Caspase 3 und somit zu einer verstärkten Apoptoseaktivität (Ludwig, 2004).
Weitere Studien haben gezeigt, dass Tumorzellen empfindlicher gegenüber den Effekten des
Bortezomibs sind als normale Zellen. Durch die Reversibilität der Proteasomeninhibition mit
Bortezomib erholten sich periphere mononukleare Blutzellen sehr schnell. Tumorzellen
hingegen waren dazu nicht in der Lage und gingen in Apoptose (Richardson, 2003). Testungen
Einleitung
9
des Medikamentes an normalen diploiden Zellen, wie z.B. humane Fibroblasten zeigten
wiederholt keine Effekte (Brignole et al., 2006).
Abb. 1: Intrazelluläre Signalkaskade des Transkriptionsfaktor NF-κκκκB (modifiziert nach Adams, 2003). Die Abbildung stellt den Zusammenhang zwischen NF-κB und seinem Inhibitor IκB dar. Weiterhin wird gezeigt, dass bei Einfluss von zellulärem Stress IκB durch Proteasomen abgebaut wird und NF-κB somit aktiv wird. Bortezomib greift in diesen Mechanismus ein und verhindert die Elimination von IκB.

Material und Methoden
10
3 Material und Methoden
3.1 Neuroblastomzelllinien
Es wurden sieben verschiedene humane Neuroblastomzelllinien (Kelly, LS, SH-SY5Y, SK-N-
SH, IMR-32, LAN-1 und LAN-5) untersucht. Die Zelllinien Kelly, LAN-1, LAN-5, SK-N-SH und
IMR-32 wurden uns freundlicherweise von Herrn Prof. Dr. Erttmann (Abteilung Pädiatrische
Onkologie, Universitätsklinikum Hamburg-Eppendorf, Deutschland) zur Verfügung gestellt. Die
Zelllinien LS und SH-SY5Y wurden uns freundlicherweise von Herrn Prof. Dr. Hildebrandt
(Abteilung Zelluläre Chemie, Medizinische Hochschule Hannover, Deutschland) zur
Verfügung gestellt.
3.2 Bortezomib (Velcade®)
Das Medikament Bortezomib wurde von der Firma Janssen Pharmaceutica N.V. (Beerse,
Belgien) über die Apotheke des Universitätsklinikums bezogen und als steriles Pulver in einer
Packungsgröße von 3,5 mg geliefert. Aus diesem Pulver wurde dann durch Zugabe von 3,5
ml PBS eine Stammlösung hergestellt, deren Konzentration 1 mg / ml betrug. Gelagert wurde
die Stammlösung dann bei – 20° C im Gefrierschrank.
Bei jedem Versuch wurde diese Stammlösung erneut aufgetaut und in einer
Verdünnungsreihe wurden die zu untersuchenden Konzentrationen hergestellt (siehe 3.3.1.5).
3.3. Charakterisierung von Bortezomib in vitro
3.3.1 Zellproliferationsassays
3.3.1.1 Zellkultur
Die Arbeiten mit den Zelllinien erfolgten unter einer Hera Safe Sicherheitswerkbank (Heraeus
Instruments, Hanau, Deutschland) unter sterilen Bedingungen. In 50 ml Zellkulturfalschen
(Nunclon®, Nunc, Roskilde, Dänemark) erfolgte die Kultivierung der Zelllinien unter
Standardbedingungen (37° C, 100% Luftfeuchtigkeit, 5% CO2 / 95% Luft) in einem Hera Cell
Brutschrank (Heraeus Instruments, Hanau, Deutschland). Als Nährmedium wurde RPMI
Medium (Gibco/Life Technologies, Paisley, Scotland) verwendet, welches zusätzlich mit 10%
hitzeinaktiviertem fetalen Kälberserum (FCS, Gibco®), 2 mM L-Glutamin (Gibco®), sowie 100

Material und Methoden
11
U / ml Penicillin und 100 µg / ml Streptomycin (Gibco®) versetzt wurde. Dieses Medium wird
im Weiteren als Kulturmedium bezeichnet und wurde 2 – 3 mal pro Woche gewechselt. Wenn
die Zellen konfluent gewachsen waren, wurden sie passagiert. Hierzu wurde das alte
Kulturmedium abgesaugt, die Zellen wurden mit 5 ml PBS (Gibco® Dulbecco´s Phosphate-
Buffered Saline, Invitrogen, Carlsbad, USA) gespült, anschließend für fünf Minuten mit je 5 ml
Trypsin-EDTA (Gibco®) versetzt und darauf folgend bei 37° C und 5% CO2 5 Minuten
inkubiert. Nach dem Ablösen der Zellen vom Flaschenboden, erfolgte die Zugabe von 5 ml
Kulturmedium, um das Trypsin zu inaktivieren und so eine Schädigung der Zellen durch das
Trypsin-EDTA zu verhindern. Abschließend wurden die abgelösten Zellen auf neue
Zellkulturflaschen verteilt, die zuvor mit 15 ml Kulturmedium aufgefüllt waren, oder die Zellen
wurden für die Zellproliferationsexperimente eingesetzt. Da für die Standardisierung und
Reproduktion der Experimente eine hohe Zellzahl erforderlich war, wurden vor Beginn aller
Experimente alle sieben Zelllinien über mehrere Wochen in Zellkulturflaschen vermehrt und
ein Suspensionspool von jeder der sieben Zelllinien erstellt. Die Suspensionen wurden fünf
Minuten bei 1500 U / min zentrifugiert und der Überstand abgesaugt. Die entstandenen
Zellpellets wurden mit Gefriermedium (Cryo-safe Ir, c.c. pro GmbH, Neustadt, Deutschland)
resuspendiert und in Einfrierröhrchen (Nunc Cryo Tube Vials, Nalge Nunc International
Naperville, Roskilde, Dänemark) auf identische Aliquots von 1,5 ml verteilt. Über Nacht
wurden die Aliquots auf - 80° C gekühlt und abschließend in flüssigem Stickstoff
kryokonserviert. Für die weiteren beschriebenen Zellproliferationsexperimente wurden jeweils
ein bis zwei Aliquots pro Zelllinie aufgetaut, auf Zellkulturflaschen verteilt und ca. drei bis vier
Wochen kultiviert, bis eine ausreichende Zellzahl herangewachsen war, um die Versuche
durchführen zu können.
3.3.1.2 Detektion von Mykoplasmen-Kontamination in der Zellkultur
Zur Überprüfung der Zellkultur auf Kontamination durch Mykoplasmen wurde das VenorGeM®
Mykoplasmen-Detektionskit (Minerva Biolabs GmbH, Berlin, Deutschland) auf der Basis der
Polymerase-Kettenreaktion eingesetzt. Der Test ist zum Nachweis der typischerweise als
Kontamination in Zellkulturen auftretenden Mykoplasmenspezies geeignet und wurde
entsprechend der Angaben des Herstellers durchgeführt. Bei keiner der Zelllinien konnte eine
Kontamination durch Mykoplasmen nachgewiesen werden.

Material und Methoden
12
3.3.1.3 Zellzählung
Die Bestimmung der Zellzahl wurde mittels des CASY®-Cell Counter Analyser System
(CASY®-Technology, Schärfe System GmbH, Reutlingen, Deutschland) durchgeführt. Dabei
wurden vitale Zellen gezählt. Für die Messung wurden die Zellen in einer speziellen
Elektrolytlösung (CASY®ton) suspendiert und anschließend durch eine Präzisionsmesspore
gesaugt.
3.3.1.4 Erstellen von Eichkurven
In den folgenden Zellproliferationsassays wurde der Einfluss des Proteasomeninhibitors
Bortezomib der Firma Janssen Pharmaceutica N.V., (Beerse, Belgien) mit Hilfe des XTT-
Zellproliferationskit (Roche Molecular Biochemicals, Mannheim, Deutschland) untersucht. Zur
Erstellung der Eichreihen wurde für jede Zelllinie eine Verdünnungsreihe mit sieben (Kelly,
LAN-1, LAN-5, SH-SY5Y, IMR-32, LS) bzw. zehn (SK-N-SH) verschiedenen
Zellkonzentrationen erstellt. Jeweils 80 µl der Zellsuspensionen wurden in die Vertiefungen
einer 96er Mikrotiterplatte (Greiner, Frickenhausen, Deutschland) pipettiert. Als Leerwert für
den ELISA-Reader wurden 80 µl Kulturmedium verwendet. Jeder Ansatz wurde fünffach
getestet. Die Platten wurden im Brutschrank für 24 Stunden unter Standardbedingungen
präinkubiert. Dann wurden 20 µl PBS zu den Testansätzen gegeben, was der Menge der in
den endgültigen Versuchen zugefügten Substanz Bortezomib gelöst in PBS entsprach.
Anschließend wurden die Zellen für 48 Stunden im Brutschrank unter Standardbedingungen
inkubiert. Zur Ermittlung der Zelldichte wurde 72 Stunden nach dem Aussäen der Cell
Proliferation Kit II (Roche Diagnostics GmbH, Mannheim, Deutschland) benutzt. Dafür wurde
XTT labeling reagent und electron coupling reagent in einer Konzentration von 50 : 1
gemischt, was einer XTT Endkonzentration von 0,3 mg / ml entspricht. Von dieser
Konzentration wurden jeweils 50 µl in jede Vertiefung der Mikrotiterplatte gegeben und die
Platten wurden zur weiteren Inkubation in den Brutschrank gestellt. Die Extinktion wurde nach
4, 6 und 24 Stunden bei 450 nm im ELISA Reader Dynatch MR3.13 (Dynex Technologies,
Ashford, UK) gemessen. Für die Zelllinien Kelly, LAN-1, LAN- 5, IMR-32, LS und SK-N-SH
stellten sich 6 Stunden und für SH-SY5Y 24 Stunden als eine optimale Inkubationszeit mit
XTT heraus. Die Zellkonzentrationen, die nach 72 Stunden Wachstum im exponentiellen
Bereich der Eichkurve lagen, wurden zum Aussäen der einzelnen Neuroblastomzellreihen
angewandt. Die jeweiligen Zellzahlen / ml sind in der Tabelle 3 zusammengefasst.
Material und Methoden
13
Zelllinie Kelly LAN-1 LAN-5 LS SK-N-SH SH-SY5Y IMR-32
Zellzahl / ml 150.000 150.000 200.000 70.000 60.000 420.000 700.000
Tab. 3: Zellzahlen / ml der Neuroblastomzelllinien, die ein geeignetes Wachstum über die Versuchsdauer von 72 Stunden gezeigt haben.
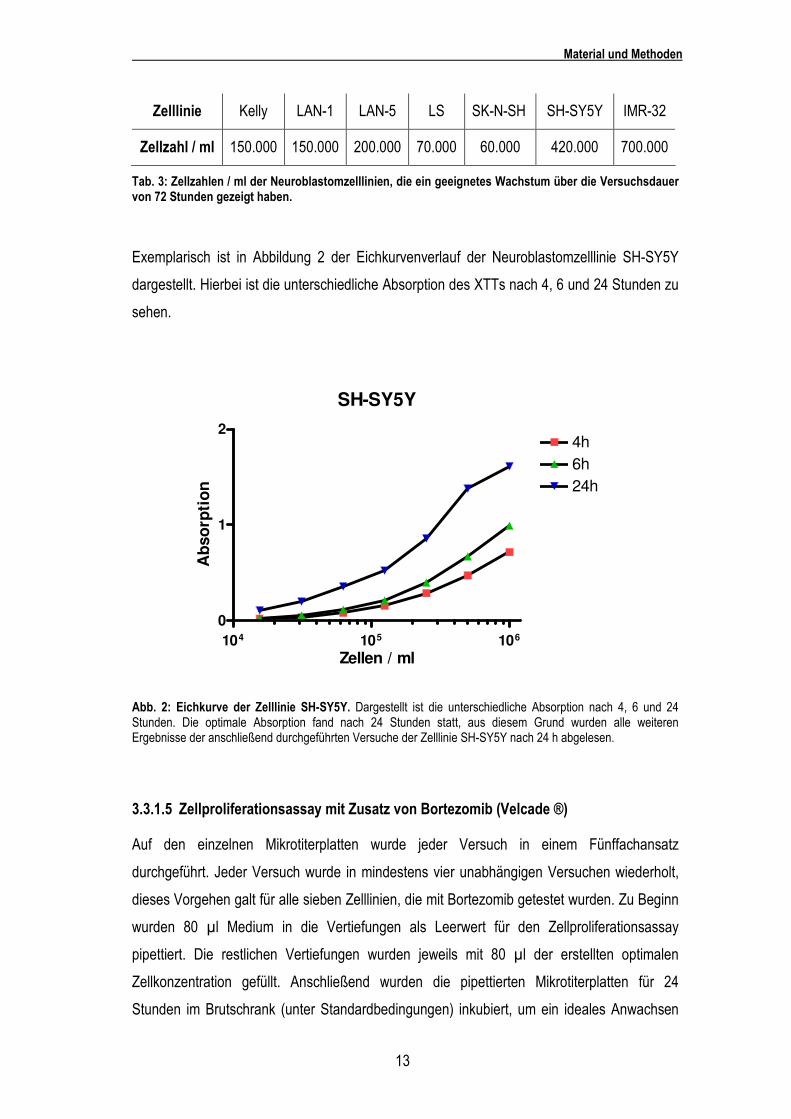
Exemplarisch ist in Abbildung 2 der Eichkurvenverlauf der Neuroblastomzelllinie SH-SY5Y
dargestellt. Hierbei ist die unterschiedliche Absorption des XTTs nach 4, 6 und 24 Stunden zu
sehen.
SH-SY5Y
104 105 106
0
1
24h
6h
24h
Zellen / ml
Ab
so
rpti
on
Abb. 2: Eichkurve der Zelllinie SH-SY5Y. Dargestellt ist die unterschiedliche Absorption nach 4, 6 und 24 Stunden. Die optimale Absorption fand nach 24 Stunden statt, aus diesem Grund wurden alle weiteren Ergebnisse der anschließend durchgeführten Versuche der Zelllinie SH-SY5Y nach 24 h abgelesen.
3.3.1.5 Zellproliferationsassay mit Zusatz von Bortezomib (Velcade ®)
Auf den einzelnen Mikrotiterplatten wurde jeder Versuch in einem Fünffachansatz
durchgeführt. Jeder Versuch wurde in mindestens vier unabhängigen Versuchen wiederholt,
dieses Vorgehen galt für alle sieben Zelllinien, die mit Bortezomib getestet wurden. Zu Beginn
wurden 80 µl Medium in die Vertiefungen als Leerwert für den Zellproliferationsassay
pipettiert. Die restlichen Vertiefungen wurden jeweils mit 80 µl der erstellten optimalen
Zellkonzentration gefüllt. Anschließend wurden die pipettierten Mikrotiterplatten für 24
Stunden im Brutschrank (unter Standardbedingungen) inkubiert, um ein ideales Anwachsen

Material und Methoden
14
der Zellen zu gewährleisten. Die Testlösungen wurden hergestellt, indem die Stammlösung
von Bortezomib (1 mg / ml) mit PBS in einer Verdünnungsreihe (1:10) versetzt wurde.
Insgesamt wurden sechs verschiedene Konzentrationen in einer Verdünnungsreihe von 100
µg / ml bis 0,001 µg / ml des Bortezomib hergestellt. 20 µl dieser Verdünnungen wurden
dann jeweils zu den 80 µl Medium in die Vertiefungen gegeben. Dies entsprach dann einer
Endkonzentration von 20 µg / ml bis 0,0002 µg / ml des Medikamentes. Die Mikrotiterplatte
wurde nach Medikamentengabe für weitere 48 Stunden im Brutschrank (unter
Standardbedingungen) inkubiert.
72 Stunden nach dem Aussäen der Zelllinien, wurde zur Auswertung der Cell Proliferation Kit
II (XTT) (Roche Diagnostics GmbH, Mannheim) verwendet. Von dieser XTT-Lösung wurden
anschließend 50 µl in jede Vertiefung der Mikrotiterplatte gegeben. Bei den Zelllinien Kelly,
LAN-1, LAN-5, SK-N-SH, LS und IMR-32 wurden nach 6 Stunden und bei der Zelllinie SH-
SY5Y nach 24 Stunden bei einer Testfilter-Wellenlänge von 490 nm und einer Referenzfilter-
Wellenlänge von 630 nm die Messungen durchgeführt. Die Ergebnisse der Kontrolle dienten
dann bei der Auswertung des Proliferationsassays als 100% Zellwachstum, zu denen die
übrigen Werte in Relation gesetzt wurden.
3.4 Charakterisierung von Bortezomib in vivo
3.4.1 Severe combined immuno deficiency Mausmodell
3.4.1.1 Versuchstiere
Die Versuche wurden an männlichen und weiblichen severe combined immuno deficiency
(scid) Mäusen der Firma Charles River durchgeführt. Die Tiere sind in der 48. Kalenderwoche
2005 geboren und waren zu Versuchsbeginn 15 Wochen alt. Die Tiere hatten im Mittel ein
Gewicht von 25 g mit einer Standardabweichung von 3,2 g. Die Haltung erfolgte in Filter-Top
Käfigen in Gruppen zu jeweils 5 Tieren bei einer Raumtemperatur von 19 – 21° C und unter
Beibehaltung des normalen Tag-Nacht-Rhythmus (Dunkelphase von 19 – 7 Uhr). Die Tiere
erhielten steriles Wasser und steriles Standard-Einstreufutter (sniff M-Z Alleindiät extrudiert,
sniff Spezialdiäten GmbH, Soest, Deutschland) ad libitum und wurden mindestens 6 – 8 Tage
vor Versuchsbeginn zur Akklimatisierung und Eingewöhnung im Tierstall des
Universitätsklinikums Hamburg–Eppendorf (Haus Süderfeldstraße) gehalten. Der im Rahmen
dieser Arbeit durchgeführte Tierversuch wurde nach § 8 des Tierschutzgesetzes von 1998 der
Bundesrepublik Deutschland unter der Versuchsnummer: 64 / 05 von der Behörde für Umwelt
und Gesundheit in Hamburg genehmigt.

Material und Methoden
15
Während der Studie ist die scid-Maus Nr. 8.3 (Bortezomibgruppe) aus unbekannter Ursache
verstorben. Es gab keine Anzeichen dafür, dass der Tumor oder die Behandlung mit
Bortezomib Einfluss auf den Gesundheitszustand des Tieres genommen hatten.
3.4.2 Behandlung der Tiere
Für den Tierversuch wurde die Zelllinie SK-N-SH ausgewählt, da diese Zelllinie in bereits
durchgeführten Tierversuchen unseres Institutes ein gutes Tumorwachstum und eine hohe
Metastasierungsrate aufwies (Ursula Valentiner, unveröffentlicht).
Aus den in flüssigem Stickstoff gelagerten Zellpoolen wurden je zwei Aliquots der SK-N-SH
Zellen aufgetaut, mindestens zwei Wochen lang kultiviert und anschließend hinsichtlich ihrer
Vitalität geprüft. Nach Absaugen des Kulturmediums und zweifacher Spülung im eisgekühlten
PBS wurden die Zellen mittels Trypsinierung gelöst und bei 1500 U / min fünf Minuten
zentrifugiert. Der Überstand wurde abgesaugt. Die Zellpellets wurden mit jeweils 1 ml
Kulturmedium resuspendiert und alle Zellen der jeweiligen Zelllinie wurden erneut gepoolt.
Nach Herstellung einer 1 : 10 Verdünnung wurde die Zellzahl der Suspension durch das
CASY®-Cell Counter Analyser System (CASY®-Technology, Schärfe System GmbH,
Reutlingen, Deutschland) bestimmt. Die Suspension wurde dann wieder zentrifugiert und die
Zellpellets wurden nach Absaugen des Überstandes mit Kulturmedium ohne FCS
resuspendiert und die Zellzahl wurde auf 5 x 106 Zellen / ml eingestellt. Die Tiere wurden mit
einem Narkosegemisch aus 20% O2 und 80% CO2 anästhesiert und danach wurden jedem
Tier 1 x 106 Zellen gelöst in 200 µl Kulturmedium subkutan zwischen die Schulterblätter
injiziert. Drei Tage nach der Tumorzellinjektion wurden die vorher in Kontrollgruppe und
Bortezomibgruppe eingeteilten Tiere, zwei mal pro Woche mit dem gleichen Verhältnis an
Narkosegemisch betäubt. Danach wurden den 20 Tieren der Behandlungsgruppe 0,2 ml der
Medikamentenlösung (Bortezomib: 1 mg / ml) in die Schwanzvene gespritzt. Dies entspricht
einer Menge von 1 mg / kg KG (Körpergewicht). Die 20 Mäuse der Kontrollgruppe erhielten
0,2 ml PBS intravenös verabreicht. Der Gesundheitszustand der Tiere wurde regelmäßig
kontrolliert. Es konnte keine Veränderung der scid-Mäuse im Verlauf der Behandlung mit dem
Medikament Bortezomib festgestellt werden. Nach 31 Tagen Behandlung (insgesamt 9
Injektionen) wurde bei den Tieren ein Imaging mittels MRT und PET-CT durchgeführt.
Abschließend erfolgte eine Betäubung der scid-Mäuse mit einem Narkosegemisch (0,1 ml / 10
g Körpergewicht) bestehend aus Rompun-Ketanest (0,8 ml Rompun 2% (Bayer Leverkusen,
Deutschland) und 1,2 ml Ketamin Gräub (100 mg / ml; Albrecht, Aulendorf, Deutschland) in 8

Material und Methoden
16
ml 0,9% NaCl). Die Tiere wurden nach Blutentnahme aus dem Augenplexus durch cervicale
Dislokation getötet. Danach wurden die Primärtumoren und die Lungen herauspräpariert. Die
Primärtumoren wurden gewogen und in beschriftete Gitterkörbchen überführt. Zur späteren
Einbettung in Paraffin nach einem automatisierten Standardverfahren wurden sie vorerst für
zwei Tage in 4% gepuffertem Formalin für 48 Stunden fixiert. Die Lungen wurden en bloc in
gepuffertem, mit 0,1 mol Phosphatpuffer (zusammengesetzt aus Natriumdihydrogenphosphat
und Dinatriumphosphat) angesetztem Formalin für 48 Stunden fixiert. Anschließend wurden
sie unter einem Stereo-Mikroskop in 1 mm dicke Scheiben geschnitten. Die Schnittdicke
wurde durch unter der Glasplatte liegendes Millimeterpapier kontrolliert. Die Lungenschnitte
wurden zufällig über einen Objektträger verteilt, in 4% igem Ager eingelegt und mit einem
Deckgläschen zusammengedrückt. Dieser Block wurde dann, wie zuvor beschrieben, in
Paraffin eingebettet. Die verdichteten Agarblöcke waren somit für die Histologie vorbereitet.
3.4.3 Histologie
3.4.3.1 Primärtumor
3.4.3.1.1 Lichtmikroskopie
Von den in Paraffin eingebetteten Primärtumoren wurden mit einem Mikrotom (Mircrom,
Techno Med GmbH, Bielefeld) 5 µm dicke Paraffinschnitte angefertigt. Diese wurden
anschließend in ein Wärmebad überführt und auf einen beschichteten Objektträger (Histo
Bond, Marienfeld, Deutschland) aufgenommen. Danach wurden die Schnitte für 24 Stunden
im Wärmeschrank bei 37° C getrocknet und anschließend mit Hämatoxylin – Eosin (HE)
gefärbt. Dieses wurde nach einem Standardprotokoll in einem Färbeautomaten (Varistain,
Shandon, Frankfurt, Deutschland) durchgeführt. Daraufhin wurden die Objektträger
eingedeckelt (Deckgläser: Engelbrecht, Edermüde, Deutschland; Eukitt Kleber: Kindler
Freiburg, Deutschland). Von den Primärtumorblöcken wurden zusätzlich Schnitte für die
Immunhistologie gewonnen.
Die HE gefärbten Schnitte der Primärtumoren wurden anschließend mittels Lichtmikroskopie
auf die Anzahl ihrer Apoptosen und Mitosen untersucht. Hierzu wurde ein Okular mit
Gitternetz benutzt, welches aus 10 mal 10 Kästchen bestand. Es wurden die vitalen Areale
des Primärtumors bestimmt und anschließend eine Zählung von 100 Zellen vorgenommen,
die in apoptotische Zellen, mitotische Zellen und normale Zellen eingeteilt wurden. Dieses
wurde an vier weiteren Bereichen des vitalen Primärtumorgewebes durchgeführt, so dass

Material und Methoden
17
insgesamt 500 Zellen ausgewertet wurden. Insgesamt wurde diese Zellzählung drei Mal
durchgeführt.
Zusätzlich wurde eine Bestimmung des Nekroseanteils mittels Lichtmikroskopie durchgeführt.
Dieses wurde durch eine prozentuale Einschätzung des vitalen und nekrotischen
Primärtumorgewebes durchgeführt.
3.4.3.1.2 Elektronenmikroskopie
Für die Elektronenmikroskopie wurden etwa 1 x 1 x 1 mm3 Gewebestücke von den Tumoren
der scid-Mäuse in 3,5% Glutaraldehyd und in 0,1 Phosphatpuffer fixiert. Anschließend erfolgte
eine Nachfixierung mit Osmiumtetroxid (1% Osmiumtetroxidin 0,1 M Phosphat-Saccarose-
Puffer) für zwei Stunden und anschließend wurde das Gewebe in einer aufsteigenden
Ethanolreihe (35% - 100%) dehydriert. Nach Spülung in Propylenoxid wurden die
Gewebestücke in Glycidether (Serva, Heidelberg, Deutschland) eingebettet. Der Primärtumor
wurde unter der Stereolupe aufgeblockt. Jetzt konnten 80 - 100 nm ultradünne Schnitte mit
dem Sorvall Porter-Blum MT2-B ULTRA-MICROTOME (Norwalk, Connecticut, USA) von den
Neuroblastomen angefertigt werden. Nach Kontrastierung mit 1% Uranylacetet und Bleicitrat
wurden die Schnitte mit einem Philips CM100 Transmissions-Elektronenmikroskop untersucht.
3.4.3.2 Lungenmetastasierung
Die vorher eingebetteten Lungen wurden ebenfalls in 5 µm dicke Paraffinschnitte geschnitten.
Die Lungen wurden komplett in Serie aufgeschnitten, wobei jeder zehnte Schnitt in ein
Wärmebad überführt und auf einen beschichteten Objektträger (Histo Bond, Marienfeld,
Deutschland) aufgenommen wurde. Von den Lungenblöcken wurden zusätzlich aus der Mitte
heraus jeweils 2 mal 20 Schnitte für die Immunhistologie gewonnen. Nach 24 Stunden im
Wärmeschrank bei 37° C waren die Schnitte getrocknet, so dass sie dann weiter HE gefärbt
wurden. Dieses wurde nach einem Standardprotokoll in einem Färbeautomaten (Varistain,
Shandon, Frankfurt, Deutschland) durchgeführt.
Die HE gefärbten Schnitte der Lungen wurden lichtmikroskopisch bei 100 facher
Vergrößerung betrachtet und die Anzahl der Metastasen gezählt. Dabei wurde mäanderförmig
über den Objektträger gefahren, um alle Flächen des Schnittes zu erfassen und gleichzeitig
eine fehlerhafte Doppelzählung von Lungenmetastasen auszuschließen. Die
Lungenmetastasen stellen sich in der HE-Färbung unter dem Lichtmikroskop deutlich

Material und Methoden
18
abgegrenzt zum umgebenden Lungengewebe (Parenchym und Stroma) dar. Die Schnitte
wurden jeweils zweimal durchgemustert. Die Metastasen der 10 mittleren HE-gefärbten
Serienschnitte wurden unter dem Lichtmikroskop ausgezählt. Die Anzahl der
Lungenmetastasen der mittleren 10 Schnitte einer Lunge wurden addiert und ein Mittelwert für
die Anzahl an Lungenmetastasen pro Schnitt gebildet. Dieser wurde mit der Gesamtzahl der
Schnitte einer Lunge multipliziert. Anschließend wurden von dem so errechneten Wert 20%
subtrahiert. Das so erhaltene Ergebnis gibt die Anzahl an Lungenmetastasen in der gesamten
Lunge wieder (Jojovic und Schumacher, 2000).
3.4.4 Disseminierte Tumorzellen im Blut
Für die Untersuchung der Anzahl der freien Tumorzellen im Blut der scid-Mäuse wurde eine
Real-time Polymerasekettenreaktion (PCR) durchgeführt. Dazu wurde das LightCycler-
System. LightCycler 2.0 der Firma Roche (Mannheim, Deutschland) benutzt.
Zur DNA Extraktion aus dem Blut wurde das QIAampDNA Blood Mini Kit 250 (Qiagen,
Hilden, Deutschland) verwendet. Die Herstellung des LightCycler Master Mixes (LightCycler
Fast Start DNA Master Plus SYBR Green Ι Cat No. 03515885001, Roche, Mannheim,
Deutschland) wurde nach Angaben des Herstellers zubereitet. Dieses QIAampDNA Blood
Mini Kit 250 besteht aus dem Fluoreszenzfarbstoff SYBR Green Ι, einer Taq DNA
Polymerase, einem Taq PCR Puffer, einer Nukleotidmixtur und 1 mmol MgCl2. Für eine
Blutprobe wurde ein 8 µl Arbeitsmastermix hergestellt. Dieser beinhaltete 100 pmol des ALU-
Primers forward (TGGCTCACGC CTGTAATCCC A) und 100 pmol des ALU-Primers reverse
(GCCACTACGC CCGGCTAATT T) (freundlicherweise von Frau Dr. Nehmann zur Verfügung
gestellt). Dazu wurde der LightCycler Master Mix und ddH2O gegeben.
Nachdem die Proben für 10 min bei 95° C erhitzt wurden, erfolgte die Denaturierung für 5 s.
Anschließend erfolgte das Annealing für 5 s bei 67° C und danach die Elongation bei 72° C
für 20 s. Nach diesem Zyklus wurde dann die Messung durchgeführt. Es wurden für jede
Probe 45 Zyklen durchgeführt. Durch die mitlaufenden Eichkurvenproben bestehend aus
sieben Proben von 106 Tumorzellen bis zu 100 Tumorzellen, konnte anschließend der
Rückschluss auf die Anzahl der freien Tumorzellen der scid-Mäuse im Blut hergestellt werden.
Zusätzlich liefen immer zwei Negativproben bestehend aus H2O und scid-Kontroll-DNA mit,
um sicherzustellen, dass die Messungen ordnungsgemäß abgelaufen waren.

Material und Methoden
19
Nach der PCR wurde eine Schmelzkurvenanlayse durchgeführt, anhand derer die Spezifität
der Produkte bestimmt wurde. Jede DNA-Probe wurde mindestens drei Mal analysiert. Die
Datenverarbeitung erfolgte mit der gerätespezifischen LightCycler™ Software 3.5 (Roche,
Mannheim, Deutschland).
3.4.5 Imaging
3.4.5.1 Magnetresonanztomographie
Vier Wochen nach der Inokulation der humanen Neuroblastomzellen wurde bei acht scid-
Mäusen ein Imaging mittels der Magnetresonanztomografie (MRT) durchgeführt. Dabei
stammten jeweils vier scid-Mäuse aus der Kontrollgruppe und vier Tiere aus der
Bortezomibgruppe. Für die Untersuchungen wurden die scid-Mäuse gewogen und mit einem
Gemisch aus Rompun-Ketanest anästhesiert (0,8 ml Rompun 2% (Bayer Leverkusen,
Deutschland) und 1,2 ml Ketamin Gräub (100 mg / ml; Albrecht, Aulendorf, Deutschland) in 8
ml 0,9% NaCl). Es wurden 0,1 ml / 10 g Mauskörpergewicht verabreicht. Nach Erreichen der
Narkosetiefe, wurden die scid-Mäuse auf einer speziellen Kunststofftrage fixiert und diese
dann in der MRT-Kleintierspule gelagert.
Die MRT-Untersuchungen erfolgten mit einem 3-Tesla-Ganzkörpergraphen (Intera Philips,
Best, Niederlande). Es wurde eine splenoidförmige Kleintierspule mit einem
Innendurchmesser von 7 cm als Empfangsspule verwendet (Philips Forschungslabor,
Hamburg). Zur Übersicht wurden T1 gewichtete Gradientenechosequenzen in drei Ebenen
angefertigt (Survey). Anschließend erfolgte die Akquisition hochauflösender T1 gewichteter
Gradientenecho- und T2 gewichteter Turbo-Spinecho-Sequenzen in sagittaler, coronaler und
axialer Schichtführung. Es wurden die folgenden Sequenzparameter verwendet: (T1
Gewichtung) TR 424 ms, TE 4,9 ms, Flip 45°; FOV 90 mm, Matrix 400 x 400, Schichtdicke 1
mm, Schichtabstand 0,1 mm, Zeit 5:57,9 min; (T2 Gewichtung) TR 3438,4 ms; TE 90 ms; Flip
90°; FOV 90 mm; Matrix 400 x 400; Schichtdicke 0.6 mm; Schichtabstand 0,06 mm, Zeit
6:04,5 min. Die gesamte Untersuchungszeit pro Maus betrug circa 30 Minuten.
Material und Methoden
20
Abb. 3: MRT (Tesla-Ganzkörpergraphen) mit Kleintierspule und spezieller Auflage zur Mauslagerung. Das Foto wurde freundlicherweise von Herrn Dr. med. Kersten Peldschus zur Verfügung gestellt.
Die Auswertung der Bilddaten zur Bestimmung der Tumorgrößen erfolgte durch Mitarbeiter
der Forschungsgruppe VOXEL-MAN (Institut für Mathematik und Datenverarbeitung in der
Medizin, UKE, Hamburg). Es wurde ein halbautomatisches Segmentationsverfahren
basierend auf der interaktiven Festlegung von Schwellenwertbereichen (verschiedene
Grauabstufungen des Tumors) und der anschließenden, automatischen Erfassung
zusammenhängender Volumenelemente (Voxel) im Bereich der Tumore angewandt. Die
korrespondierenden Tumorvolumina wurden jeweils aus der Anzahl der segmentierten Voxel
und deren räumlichen Abmessungen berechnet.
3.4.5.2 Positronen-Emissions-Tomographie / Computer Tomographie
Weiterhin wurde ein Imaging mittels Positronen-Emissions-Tomographie (PET) / Computer-
Tomographie (CT) bei 36 Versuchstieren durchgeführt. Die scid-Mäuse wurden in zwei
Gruppen eingeteilt. Jede Gruppe bestand aus scid-Mäusen der Behandlungsgruppe und der
Kontrollgruppe. Als Tracer wurden [18F]FDG und [18F]FLT (ABX Advanced Biochemical
Compounds, Radeberg, Deutschland) benutzt. Diese wurden dann in jeweils 18 Mäuse
appliziert. Je 13 MBq von [18F]FDG und [18F]FLT wurden über die Schwanzvene jeder scid-
Maus verabreicht. Nach 60 min wurden die Tiere durch intraperitoneale Injektion bestehend
aus einem Gemisch aus Rompun-Ketanest (0,8 ml Rompun 2% (Bayer Leverkusen,
Material und Methoden
21
Deutschland) und 1,2 ml Ketamin Gräub (100 mg / ml; Albrecht, Aulendorf, Deutschland) in 8
ml 0,9% NaCl) betäubt. Wie auch schon bei vorherigen Betäubungen wurden 0,1 ml / 10 g
Mauskörpergewicht gespritzt. Nach Erreichen der Narkosetiefe wurden die Tiere auf einem
Gerüst platziert und in das PET-CT gefahren. Dynamische PET Aufnahmen wurden für 45 min
unter Verwendung eines konventionellen Vollringes, Ganzkörper PET durchgeführt (Gemini
GXL 10 PET/CT System, Philips Medical Systems, Deutschland).
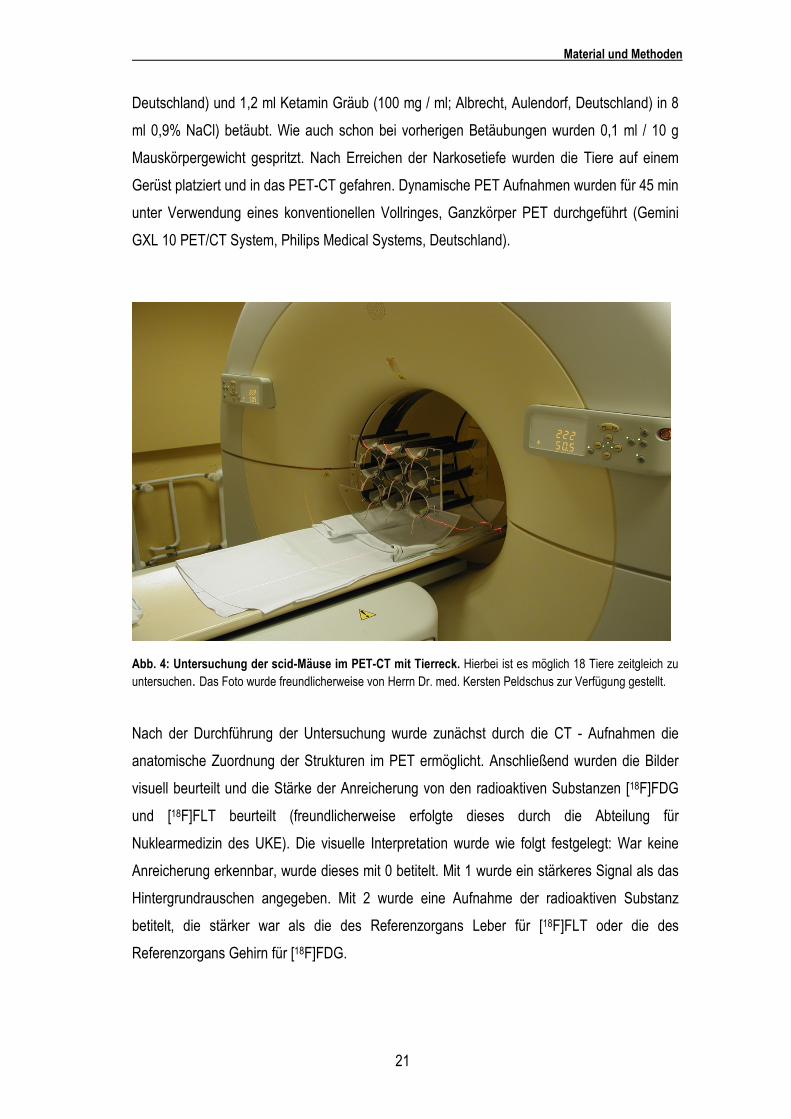
Abb. 4: Untersuchung der scid-Mäuse im PET-CT mit Tierreck. Hierbei ist es möglich 18 Tiere zeitgleich zu untersuchen. Das Foto wurde freundlicherweise von Herrn Dr. med. Kersten Peldschus zur Verfügung gestellt.
Nach der Durchführung der Untersuchung wurde zunächst durch die CT - Aufnahmen die
anatomische Zuordnung der Strukturen im PET ermöglicht. Anschließend wurden die Bilder
visuell beurteilt und die Stärke der Anreicherung von den radioaktiven Substanzen [18F]FDG
und [18F]FLT beurteilt (freundlicherweise erfolgte dieses durch die Abteilung für
Nuklearmedizin des UKE). Die visuelle Interpretation wurde wie folgt festgelegt: War keine
Anreicherung erkennbar, wurde dieses mit 0 betitelt. Mit 1 wurde ein stärkeres Signal als das
Hintergrundrauschen angegeben. Mit 2 wurde eine Aufnahme der radioaktiven Substanz
betitelt, die stärker war als die des Referenzorgans Leber für [18F]FLT oder die des
Referenzorgans Gehirn für [18F]FDG.

Material und Methoden
22
3.5 Statistische Auswertung
Die Ergebnisse der XTT-Tests sind als Mittelwert ± SD von mindestens vier unabhängigen
Versuchen dargestellt. Die Absorption der unbehandelten Kontrollzellen wurde dabei einer
Zellzahl von 100 % gleichgesetzt, die Absorption der behandelten Zellen entsprach dem
prozentualen Anteil der Kontrolle. Mittels des t-Tests (ungepaart) konnte dann für alle oben
genannten Untersuchungen ermittelt werden, ab welcher Konzentration des Bortezomibs eine
signifikante Hemmung im Vergleich zum Kontrollwert vorhanden war.
Primärtumorgewicht, Anzahl an Apoptosen, Mitosen und nekrotischen Arealen in den
Primärtumoren, sowie die Anzahl an disseminierten Tumorzellen im Blut von Kontroll- und
Bortezomibgruppe wurden verglichen, um auch hier die Wirkung von Bortezomib auf das
Neuroblastomzellwachstum in vivo zu untersuchen.
Die Ergebnisse der in vivo Versuche wurden auf ganze Zahlen auf- bzw. abgerundet.
Für alle statistischen Berechnungen und Graphen wurde die Software Graph Pad Prism
(Graph Pad TM, San Diego, USA) verwendet, wobei p-Werte <0,05 als statistisch signifikant
bewertet wurden.
Ergebnisse
23
4 Ergebnisse
4.1 Wirkung von Bortezomib in vitro
Die Wirkung von Bortezomib wurde an humanen Neuroblastomzellen untersucht. Dafür wurde
der Effekt einzelner Konzentrationen von Bortezomib auf die Zelllinien Kelly, SK-N-SH, SH-
SY5Y, LAN-1, LAN-5, IMR-32 und LS getestet. Wenn eine bestimmte Konzentration von
Bortezomib nur bei drei oder weniger Zelllinien eine Wirkung zeigte, wurden statistisch
signifikante Effekte von weniger als 20% als biologisch nicht relevant eingestuft (Valentiner et
al., 2005). Bortezomib verursachte eine dosisabhängige Wachstumsinhibierung bei allen
untersuchten Neuroblastomzelllinien. Die IC50 Werte der sieben Zelllinien reichten von 1,9 ng /
ml (SH-SY5Y) zu 8,4 ng / ml (LAN-1).
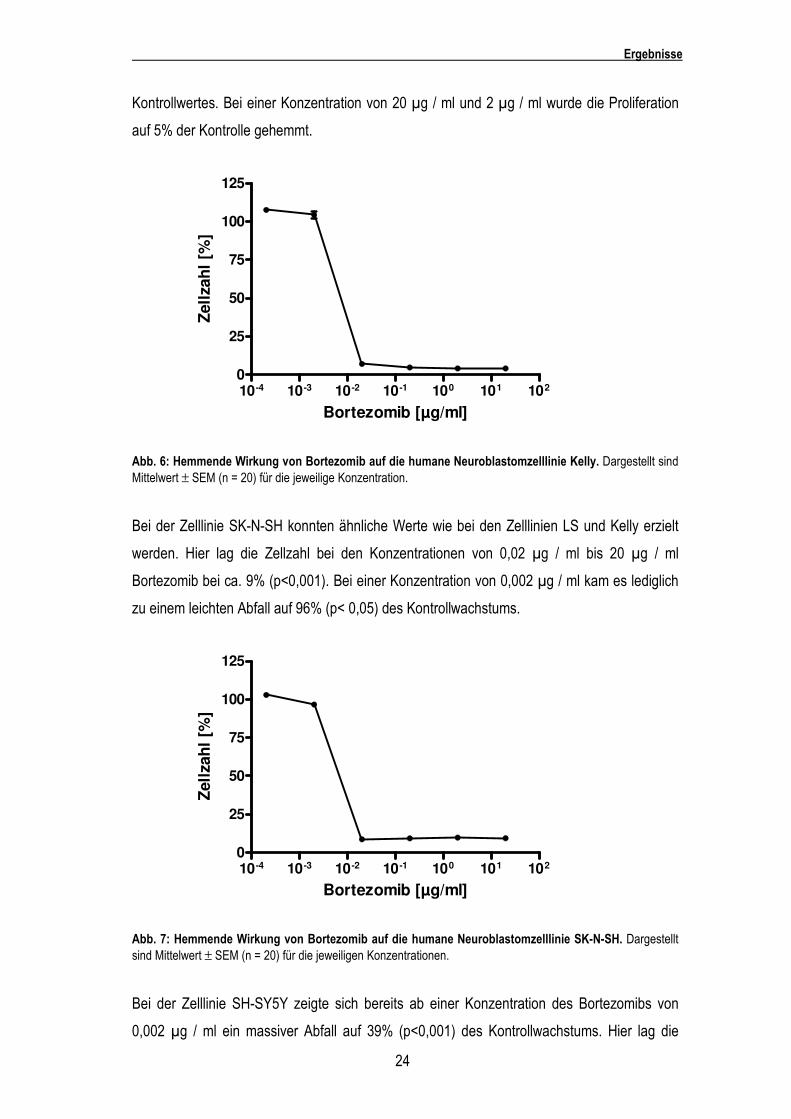
Bei der Zelllinie LS kam es bei einer Konzentration des Bortzomibs von 0,0002 µg / ml und
0,002 µg / ml zu keinerlei Veränderung der Zellproliferation. Erst ab einer Konzentration von
0,02 µg / ml Bortezomiblösung kam es zu einem massiven Abfall der Zellzahl auf 6,9%
(p<0,001) der Kontrolle. Bei einer Konzentration von 2 µg / ml und 20 µg / ml wurde die
Proliferation auf 1,5% der Kontrolle gesenkt.
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 5: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie LS. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweilige Konzentration.
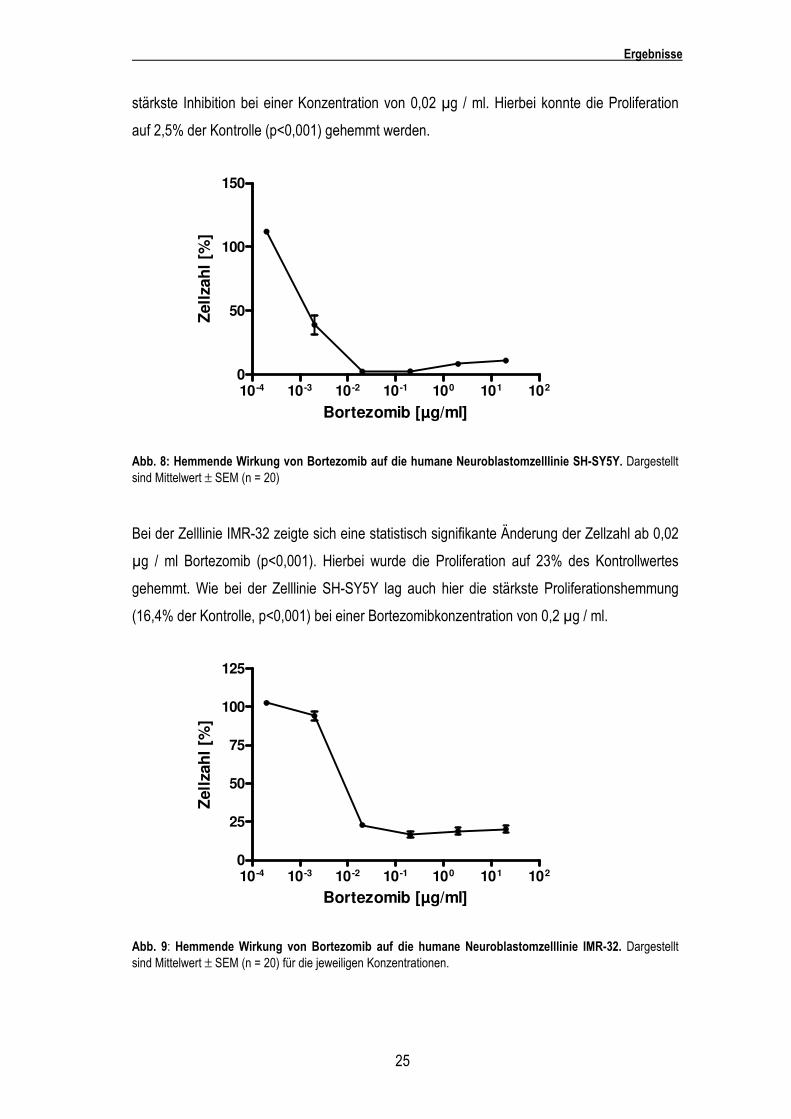
Bei der Zelllinie Kelly setzte die inhibitorische Wirkung ebenfalls erst bei einer Konzentration
von 0,02 µg / ml Bortezomib ein. Die Zellzahl erreichte nur noch 7,8% (p<0,001) des
Ergebnisse
24
Kontrollwertes. Bei einer Konzentration von 20 µg / ml und 2 µg / ml wurde die Proliferation
auf 5% der Kontrolle gehemmt.
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 6: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie Kelly. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweilige Konzentration.
Bei der Zelllinie SK-N-SH konnten ähnliche Werte wie bei den Zelllinien LS und Kelly erzielt
werden. Hier lag die Zellzahl bei den Konzentrationen von 0,02 µg / ml bis 20 µg / ml
Bortezomib bei ca. 9% (p<0,001). Bei einer Konzentration von 0,002 µg / ml kam es lediglich
zu einem leichten Abfall auf 96% (p< 0,05) des Kontrollwachstums.
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 7: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie SK-N-SH. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweiligen Konzentrationen.
Bei der Zelllinie SH-SY5Y zeigte sich bereits ab einer Konzentration des Bortezomibs von
0,002 µg / ml ein massiver Abfall auf 39% (p<0,001) des Kontrollwachstums. Hier lag die
Ergebnisse
25
stärkste Inhibition bei einer Konzentration von 0,02 µg / ml. Hierbei konnte die Proliferation
auf 2,5% der Kontrolle (p<0,001) gehemmt werden.
10-4 10-3 10-2 10-1 100 101 102
0
50
100
150
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 8: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie SH-SY5Y. Dargestellt sind Mittelwert ± SEM (n = 20)
Bei der Zelllinie IMR-32 zeigte sich eine statistisch signifikante Änderung der Zellzahl ab 0,02
µg / ml Bortezomib (p<0,001). Hierbei wurde die Proliferation auf 23% des Kontrollwertes
gehemmt. Wie bei der Zelllinie SH-SY5Y lag auch hier die stärkste Proliferationshemmung
(16,4% der Kontrolle, p<0,001) bei einer Bortezomibkonzentration von 0,2 µg / ml.
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 9: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie IMR-32. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweiligen Konzentrationen.
Ergebnisse
26
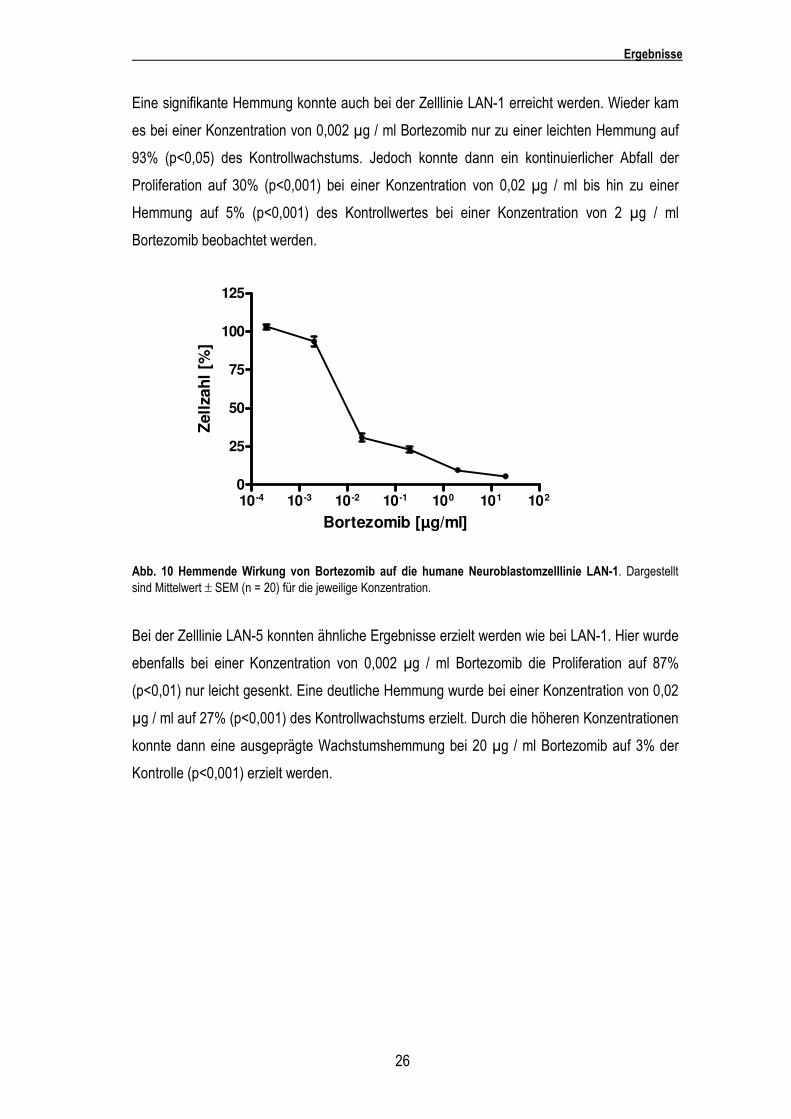
Eine signifikante Hemmung konnte auch bei der Zelllinie LAN-1 erreicht werden. Wieder kam
es bei einer Konzentration von 0,002 µg / ml Bortezomib nur zu einer leichten Hemmung auf
93% (p<0,05) des Kontrollwachstums. Jedoch konnte dann ein kontinuierlicher Abfall der
Proliferation auf 30% (p<0,001) bei einer Konzentration von 0,02 µg / ml bis hin zu einer
Hemmung auf 5% (p<0,001) des Kontrollwertes bei einer Konzentration von 2 µg / ml
Bortezomib beobachtet werden.
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 10 Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie LAN-1. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweilige Konzentration.
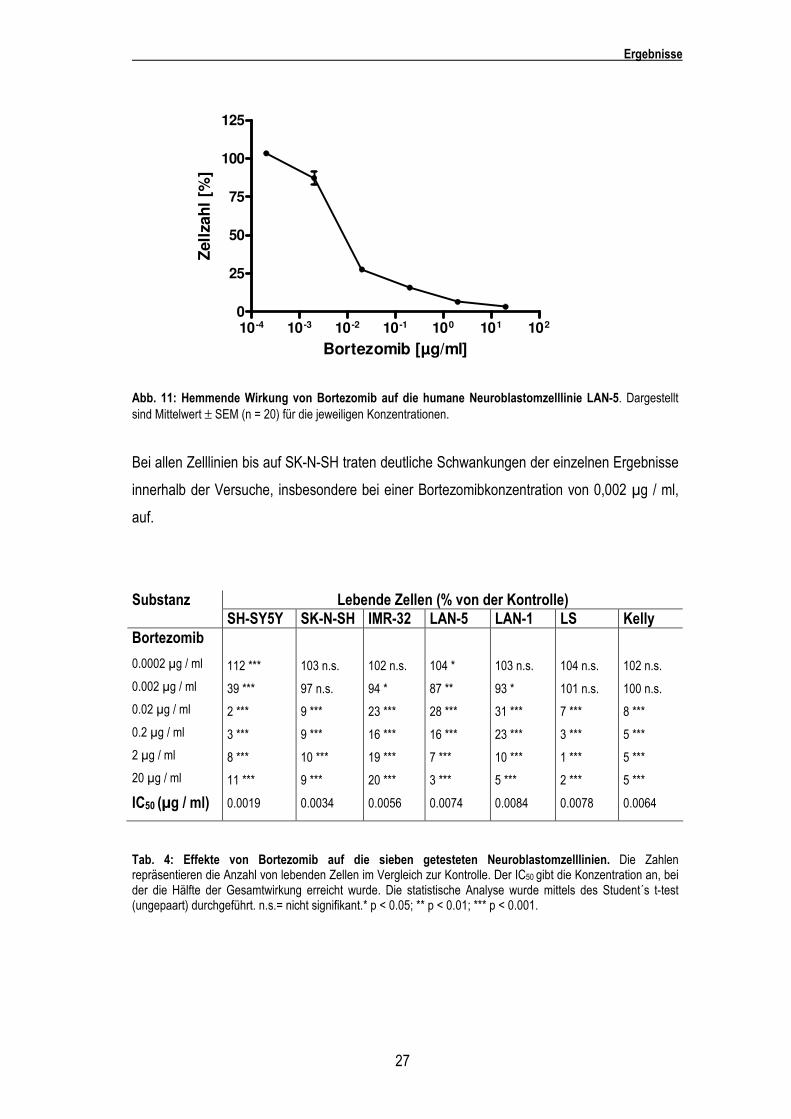
Bei der Zelllinie LAN-5 konnten ähnliche Ergebnisse erzielt werden wie bei LAN-1. Hier wurde
ebenfalls bei einer Konzentration von 0,002 µg / ml Bortezomib die Proliferation auf 87%
(p<0,01) nur leicht gesenkt. Eine deutliche Hemmung wurde bei einer Konzentration von 0,02
µg / ml auf 27% (p<0,001) des Kontrollwachstums erzielt. Durch die höheren Konzentrationen
konnte dann eine ausgeprägte Wachstumshemmung bei 20 µg / ml Bortezomib auf 3% der
Kontrolle (p<0,001) erzielt werden.
Ergebnisse
27
10-4 10-3 10-2 10-1 100 101 102
0
25
50
75
100
125
Bortezomib [µg/ml]
Zell
zah
l [%
]
Abb. 11: Hemmende Wirkung von Bortezomib auf die humane Neuroblastomzelllinie LAN-5. Dargestellt sind Mittelwert ± SEM (n = 20) für die jeweiligen Konzentrationen.
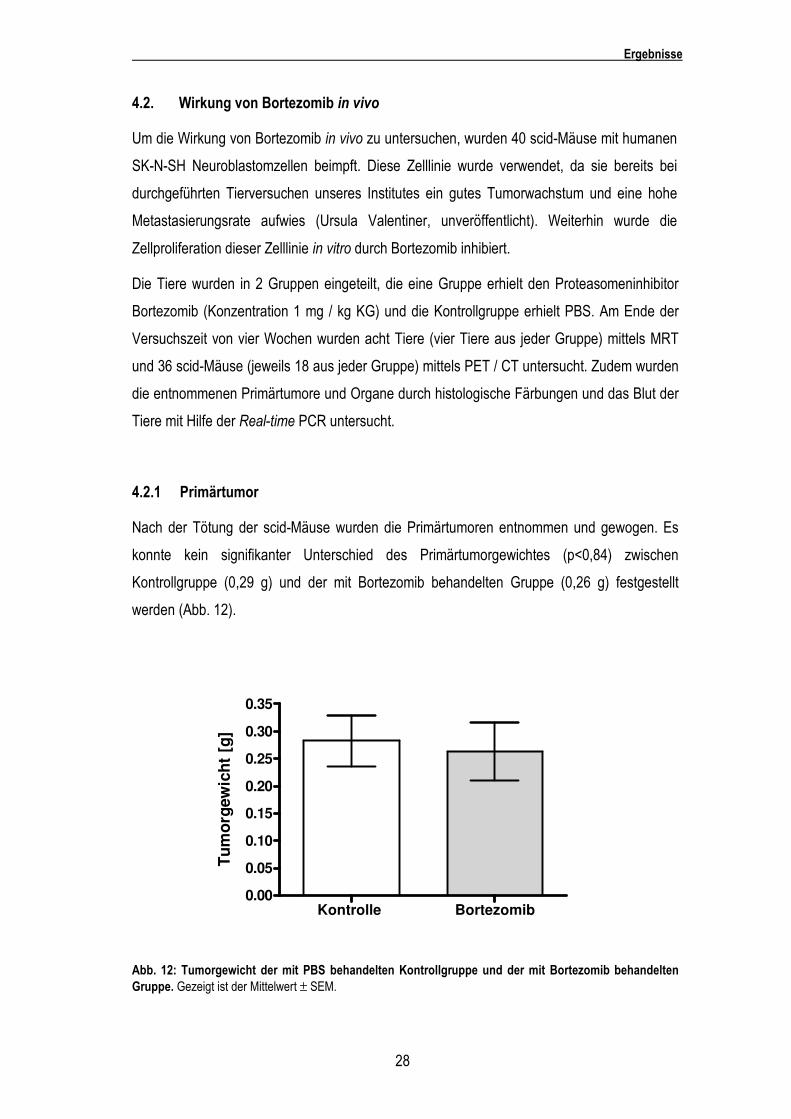
Bei allen Zelllinien bis auf SK-N-SH traten deutliche Schwankungen der einzelnen Ergebnisse
innerhalb der Versuche, insbesondere bei einer Bortezomibkonzentration von 0,002 µg / ml,
auf.
Substanz Lebende Zellen (% von der Kontrolle) SH-SY5Y SK-N-SH IMR-32 LAN-5 LAN-1 LS Kelly Bortezomib
0.0002 µg / ml
0.002 µg / ml
0.02 µg / ml
0.2 µg / ml
2 µg / ml
20 µg / ml
IC50 (µg / ml)
112 ***
39 ***
2 ***
3 ***
8 ***
11 ***
0.0019
103 n.s.
97 n.s.
9 ***
9 ***
10 ***
9 ***
0.0034
102 n.s.
94 *
23 ***
16 ***
19 ***
20 ***
0.0056
104 *
87 **
28 ***
16 ***
7 ***
3 ***
0.0074
103 n.s.
93 *
31 ***
23 ***
10 ***
5 ***
0.0084
104 n.s.
101 n.s.
7 ***
3 ***
1 ***
2 ***
0.0078
102 n.s.
100 n.s.
8 ***
5 ***
5 ***
5 ***
0.0064
Tab. 4: Effekte von Bortezomib auf die sieben getesteten Neuroblastomzelllinien. Die Zahlen repräsentieren die Anzahl von lebenden Zellen im Vergleich zur Kontrolle. Der IC50 gibt die Konzentration an, bei der die Hälfte der Gesamtwirkung erreicht wurde. Die statistische Analyse wurde mittels des Student´s t-test (ungepaart) durchgeführt. n.s.= nicht signifikant.* p < 0.05; ** p < 0.01; *** p < 0.001.
Ergebnisse
28
4.2. Wirkung von Bortezomib in vivo
Um die Wirkung von Bortezomib in vivo zu untersuchen, wurden 40 scid-Mäuse mit humanen
SK-N-SH Neuroblastomzellen beimpft. Diese Zelllinie wurde verwendet, da sie bereits bei
durchgeführten Tierversuchen unseres Institutes ein gutes Tumorwachstum und eine hohe
Metastasierungsrate aufwies (Ursula Valentiner, unveröffentlicht). Weiterhin wurde die
Zellproliferation dieser Zelllinie in vitro durch Bortezomib inhibiert.
Die Tiere wurden in 2 Gruppen eingeteilt, die eine Gruppe erhielt den Proteasomeninhibitor
Bortezomib (Konzentration 1 mg / kg KG) und die Kontrollgruppe erhielt PBS. Am Ende der
Versuchszeit von vier Wochen wurden acht Tiere (vier Tiere aus jeder Gruppe) mittels MRT
und 36 scid-Mäuse (jeweils 18 aus jeder Gruppe) mittels PET / CT untersucht. Zudem wurden
die entnommenen Primärtumore und Organe durch histologische Färbungen und das Blut der
Tiere mit Hilfe der Real-time PCR untersucht.
4.2.1 Primärtumor
Nach der Tötung der scid-Mäuse wurden die Primärtumoren entnommen und gewogen. Es
konnte kein signifikanter Unterschied des Primärtumorgewichtes (p<0,84) zwischen
Kontrollgruppe (0,29 g) und der mit Bortezomib behandelten Gruppe (0,26 g) festgestellt
werden (Abb. 12).
Abb. 12: Tumorgewicht der mit PBS behandelten Kontrollgruppe und der mit Bortezomib behandelten Gruppe. Gezeigt ist der Mittelwert ± SEM.
Kontrolle Bortezomib0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
Tu
mo
rgew
ich
t [g
]

Ergebnisse
29
4.2.1.1 Lichtmikroskopie
Die Neuroblastomzellen des Primärtumores präsentierten sich als eher runde, undifferenzierte
Zellen und waren dicht gepackt. Einzig in der Zellperipherie waren die Zellen lockerer gepackt.
Die Primärtumoren enthielten wenig Stroma; in ihrem Zentrum waren ausgeprägte eosinophile
nekrotische Areale zu erkennen. Weiterhin wiesen die malignen Zellen eine zum Kern hin
verschobene Kern-Plasma-Relation auf und zeigten atypische polymorphe und polyzyklische
Zellkerne, prominente Nukleoli und Mitosen. In der HE Färbung wurden die apoptotischen
Zellen durch dunkel gefärbte und kondensierte Chromosomen dargestellt. Mitosen zeigten
sich durch mitotische Figuren von kondensierten Chromosomen.
In der vorliegenden Arbeit wurde bei den entnommenen Primärtumoren die Prozentzahl der
Apoptosen und der Mitosen bestimmt. Sowohl die Anzahl der Apoptosen als auch die Anzahl
der Mitosen wiesen einen hochsignifikanten Unterschied zwischen der Kontrollgruppe und der
mit Bortezomib behandelten Gruppe auf. In der Kontrollgruppe befanden sich 5% der Zellen in
Apoptose, während sich in der Behandlungsgruppe 10% der gezählten Zellen in Apoptose
befanden (p<0,001). Die Mitoserate der Kontrollgruppe betrug im Mittel 6,5% und die der
Bortezomibgruppe 5% (p<0,001). In den folgenden Abbildungen (Abb.13 – 16) sind die
histologischen Präparate und die dazugehörigen Ergebnisse der Anzahl an Apoptosen und
Mitosen graphisch dargestellt. Der Anteil an Nekrose im Primärtumor und das histologische
Präparat eines Kontrollgruppentieres sind in Abb. 17 und 18 dargestellt.
Ergebnisse
30
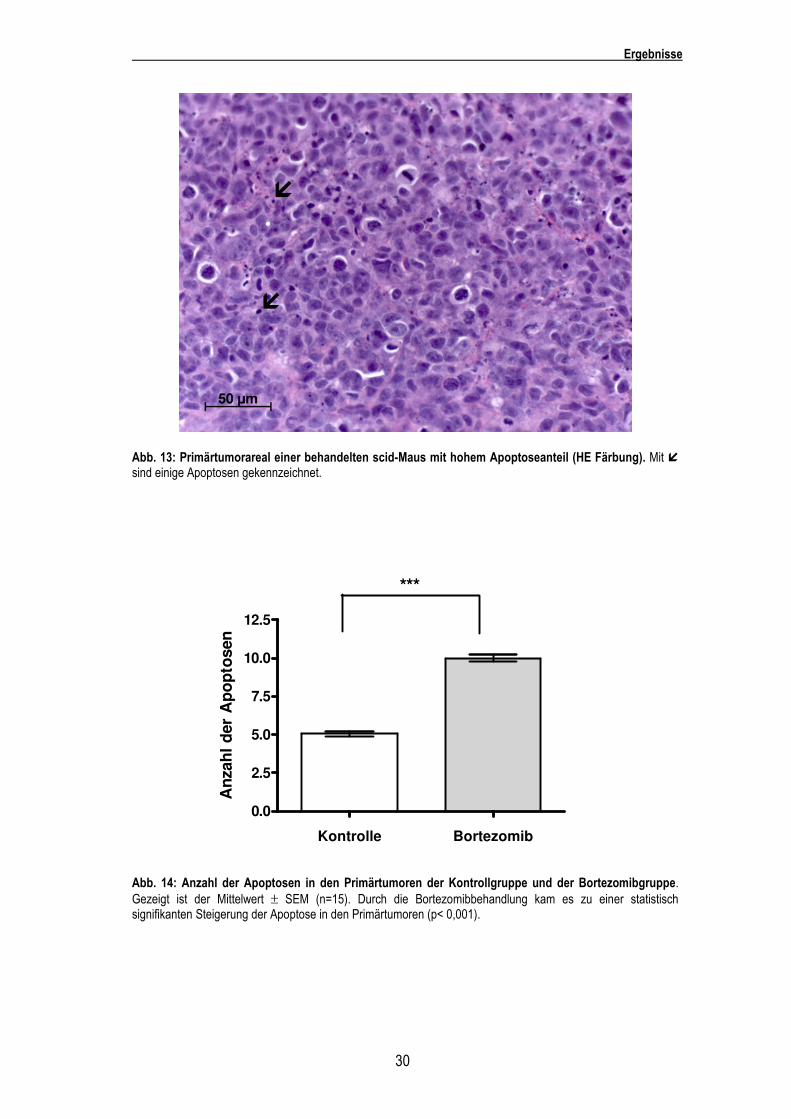
Abb. 13: Primärtumorareal einer behandelten scid-Maus mit hohem Apoptoseanteil (HE Färbung). Mit � sind einige Apoptosen gekennzeichnet.
Abb. 14: Anzahl der Apoptosen in den Primärtumoren der Kontrollgruppe und der Bortezomibgruppe. Gezeigt ist der Mittelwert ± SEM (n=15). Durch die Bortezomibbehandlung kam es zu einer statistisch signifikanten Steigerung der Apoptose in den Primärtumoren (p< 0,001).
Kontrolle Bortezomib
0.0
2.5
5.0
7.5
10.0
12.5
***
An
zah
l d
er
Ap
op
tosen
Ergebnisse
31
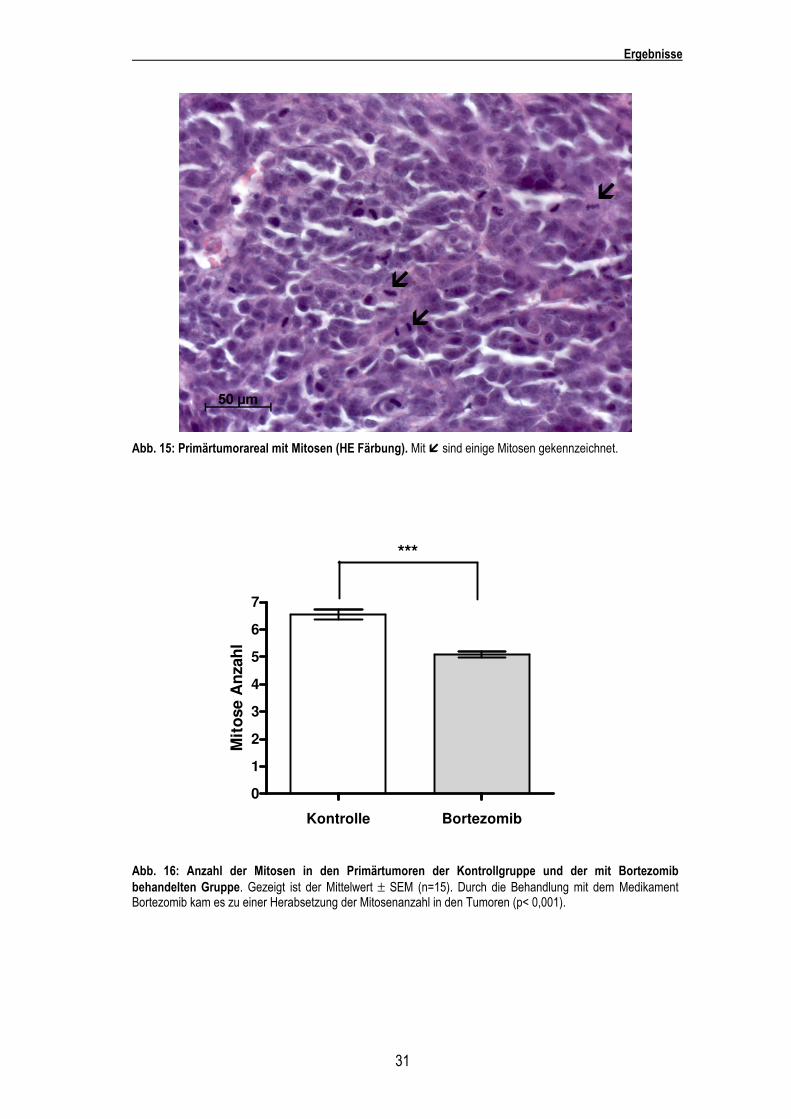
Abb. 15: Primärtumorareal mit Mitosen (HE Färbung). Mit � sind einige Mitosen gekennzeichnet.
Abb. 16: Anzahl der Mitosen in den Primärtumoren der Kontrollgruppe und der mit Bortezomib behandelten Gruppe. Gezeigt ist der Mittelwert ± SEM (n=15). Durch die Behandlung mit dem Medikament Bortezomib kam es zu einer Herabsetzung der Mitosenanzahl in den Tumoren (p< 0,001).
Kontrolle Bortezomib
0
1
2
3
4
5
6
7
***
Mit
ose A
nza
hl
Ergebnisse
32
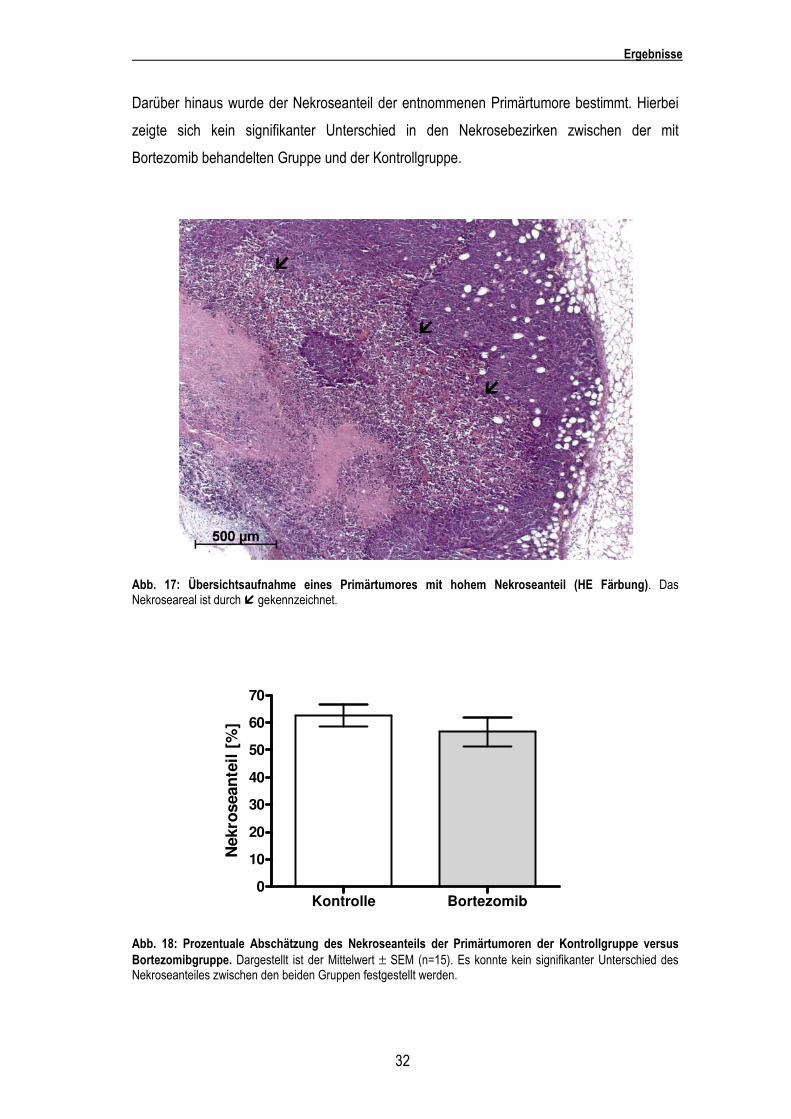
Darüber hinaus wurde der Nekroseanteil der entnommenen Primärtumore bestimmt. Hierbei
zeigte sich kein signifikanter Unterschied in den Nekrosebezirken zwischen der mit
Bortezomib behandelten Gruppe und der Kontrollgruppe.
Abb. 17: Übersichtsaufnahme eines Primärtumores mit hohem Nekroseanteil (HE Färbung). Das Nekroseareal ist durch � gekennzeichnet.
Abb. 18: Prozentuale Abschätzung des Nekroseanteils der Primärtumoren der Kontrollgruppe versus Bortezomibgruppe. Dargestellt ist der Mittelwert ± SEM (n=15). Es konnte kein signifikanter Unterschied des Nekroseanteiles zwischen den beiden Gruppen festgestellt werden.
Kontrolle Bortezomib0
10
20
30
40
50
60
70
Nekro
sean
teil
[%
]
Ergebnisse
33
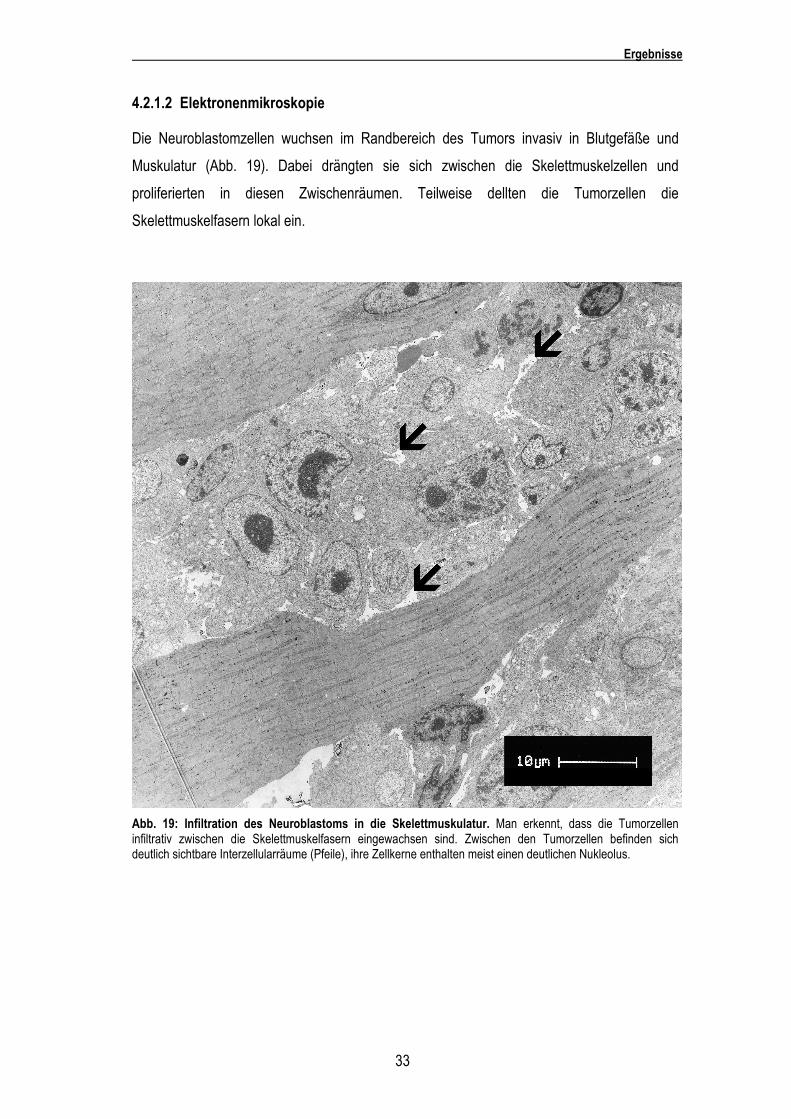
4.2.1.2 Elektronenmikroskopie
Die Neuroblastomzellen wuchsen im Randbereich des Tumors invasiv in Blutgefäße und
Muskulatur (Abb. 19). Dabei drängten sie sich zwischen die Skelettmuskelzellen und
proliferierten in diesen Zwischenräumen. Teilweise dellten die Tumorzellen die
Skelettmuskelfasern lokal ein.
Abb. 19: Infiltration des Neuroblastoms in die Skelettmuskulatur. Man erkennt, dass die Tumorzellen infiltrativ zwischen die Skelettmuskelfasern eingewachsen sind. Zwischen den Tumorzellen befinden sich deutlich sichtbare Interzellularräume (Pfeile), ihre Zellkerne enthalten meist einen deutlichen Nukleolus.
Ergebnisse
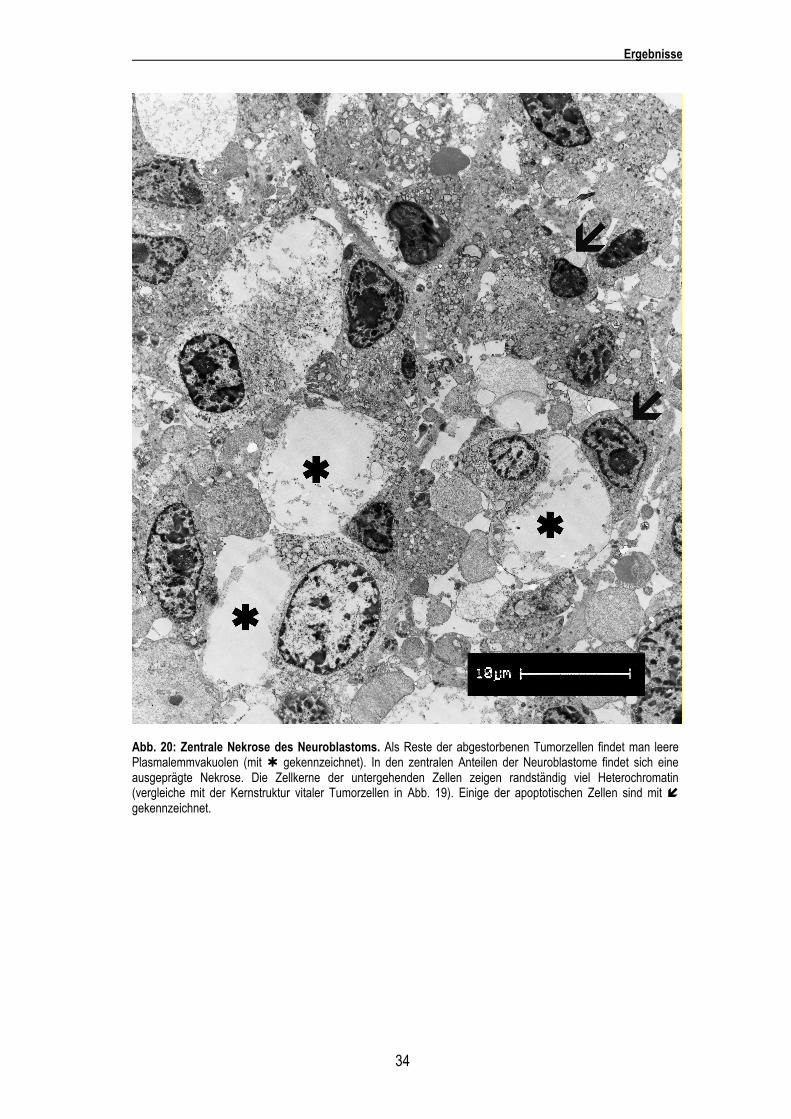
34
Abb. 20: Zentrale Nekrose des Neuroblastoms. Als Reste der abgestorbenen Tumorzellen findet man leere Plasmalemmvakuolen (mit � gekennzeichnet). In den zentralen Anteilen der Neuroblastome findet sich eine ausgeprägte Nekrose. Die Zellkerne der untergehenden Zellen zeigen randständig viel Heterochromatin (vergleiche mit der Kernstruktur vitaler Tumorzellen in Abb. 19). Einige der apoptotischen Zellen sind mit � gekennzeichnet.
Ergebnisse
35
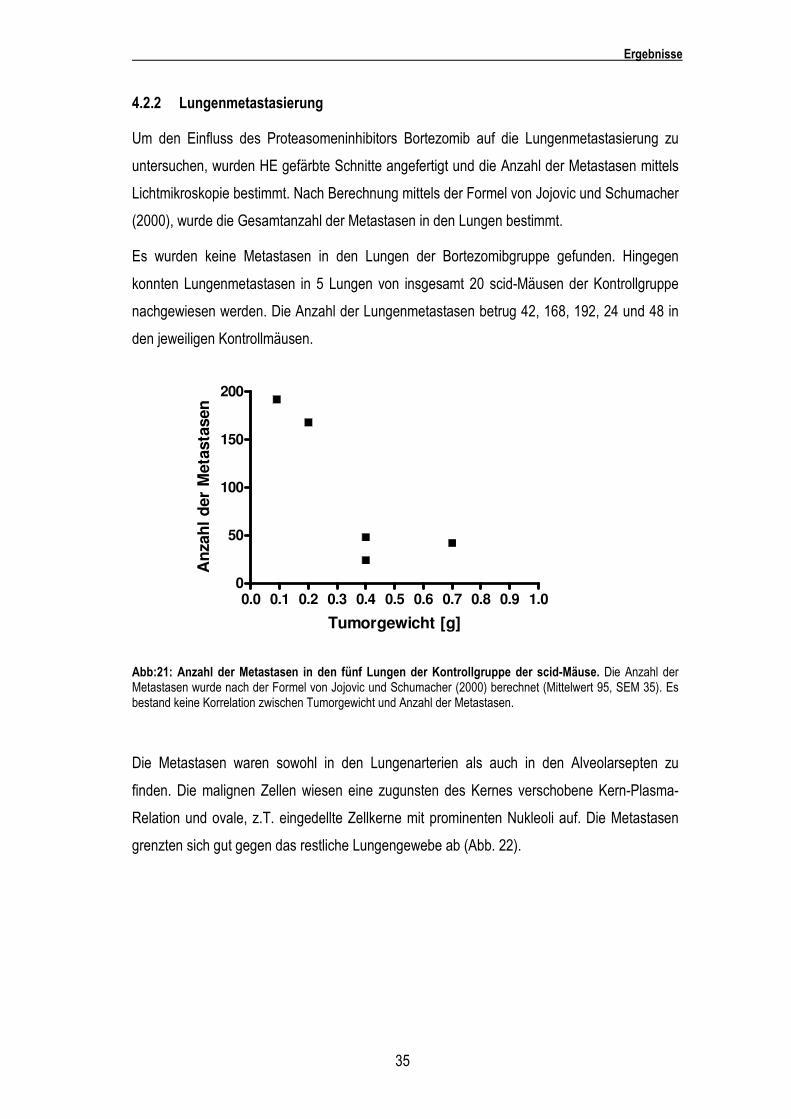
4.2.2 Lungenmetastasierung
Um den Einfluss des Proteasomeninhibitors Bortezomib auf die Lungenmetastasierung zu
untersuchen, wurden HE gefärbte Schnitte angefertigt und die Anzahl der Metastasen mittels
Lichtmikroskopie bestimmt. Nach Berechnung mittels der Formel von Jojovic und Schumacher
(2000), wurde die Gesamtanzahl der Metastasen in den Lungen bestimmt.
Es wurden keine Metastasen in den Lungen der Bortezomibgruppe gefunden. Hingegen
konnten Lungenmetastasen in 5 Lungen von insgesamt 20 scid-Mäusen der Kontrollgruppe
nachgewiesen werden. Die Anzahl der Lungenmetastasen betrug 42, 168, 192, 24 und 48 in
den jeweiligen Kontrollmäusen.
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
50
100
150
200
Tumorgewicht [g]
An
zah
l d
er
Meta
sta
sen
Abb:21: Anzahl der Metastasen in den fünf Lungen der Kontrollgruppe der scid-Mäuse. Die Anzahl der Metastasen wurde nach der Formel von Jojovic und Schumacher (2000) berechnet (Mittelwert 95, SEM 35). Es bestand keine Korrelation zwischen Tumorgewicht und Anzahl der Metastasen.
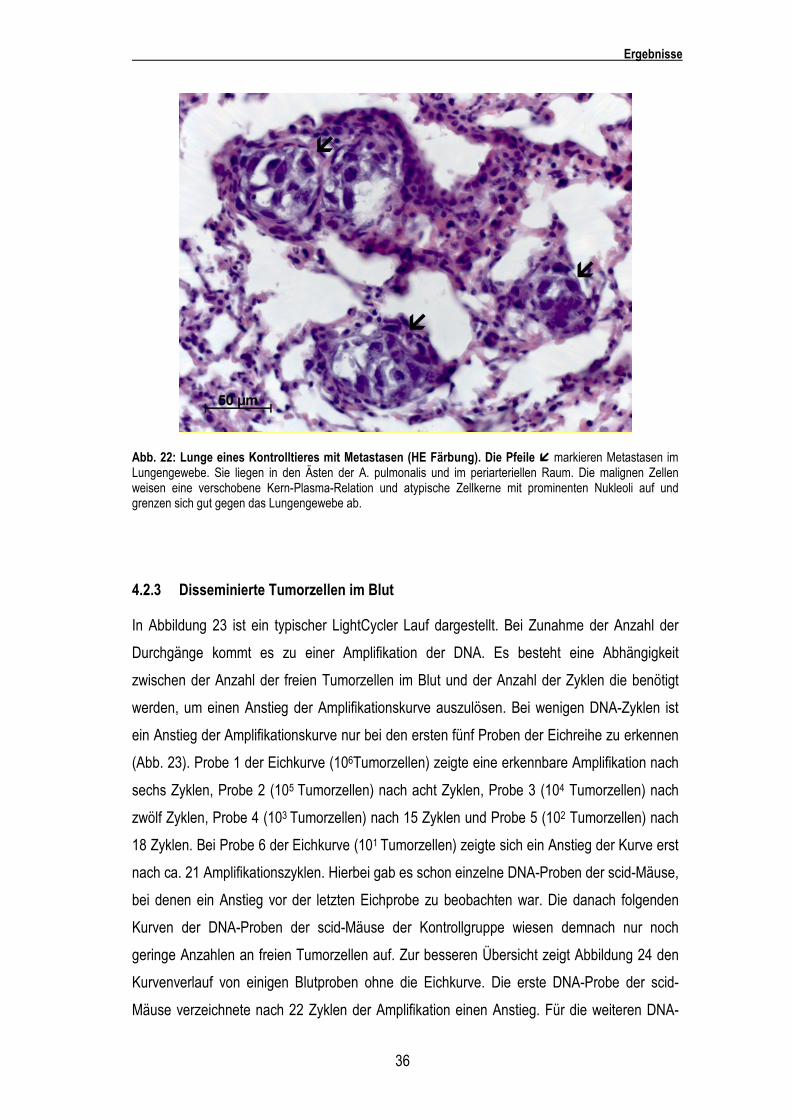
Die Metastasen waren sowohl in den Lungenarterien als auch in den Alveolarsepten zu
finden. Die malignen Zellen wiesen eine zugunsten des Kernes verschobene Kern-Plasma-
Relation und ovale, z.T. eingedellte Zellkerne mit prominenten Nukleoli auf. Die Metastasen
grenzten sich gut gegen das restliche Lungengewebe ab (Abb. 22).
Ergebnisse
36
Abb. 22: Lunge eines Kontrolltieres mit Metastasen (HE Färbung). Die Pfeile � markieren Metastasen im Lungengewebe. Sie liegen in den Ästen der A. pulmonalis und im periarteriellen Raum. Die malignen Zellen weisen eine verschobene Kern-Plasma-Relation und atypische Zellkerne mit prominenten Nukleoli auf und grenzen sich gut gegen das Lungengewebe ab.
4.2.3 Disseminierte Tumorzellen im Blut
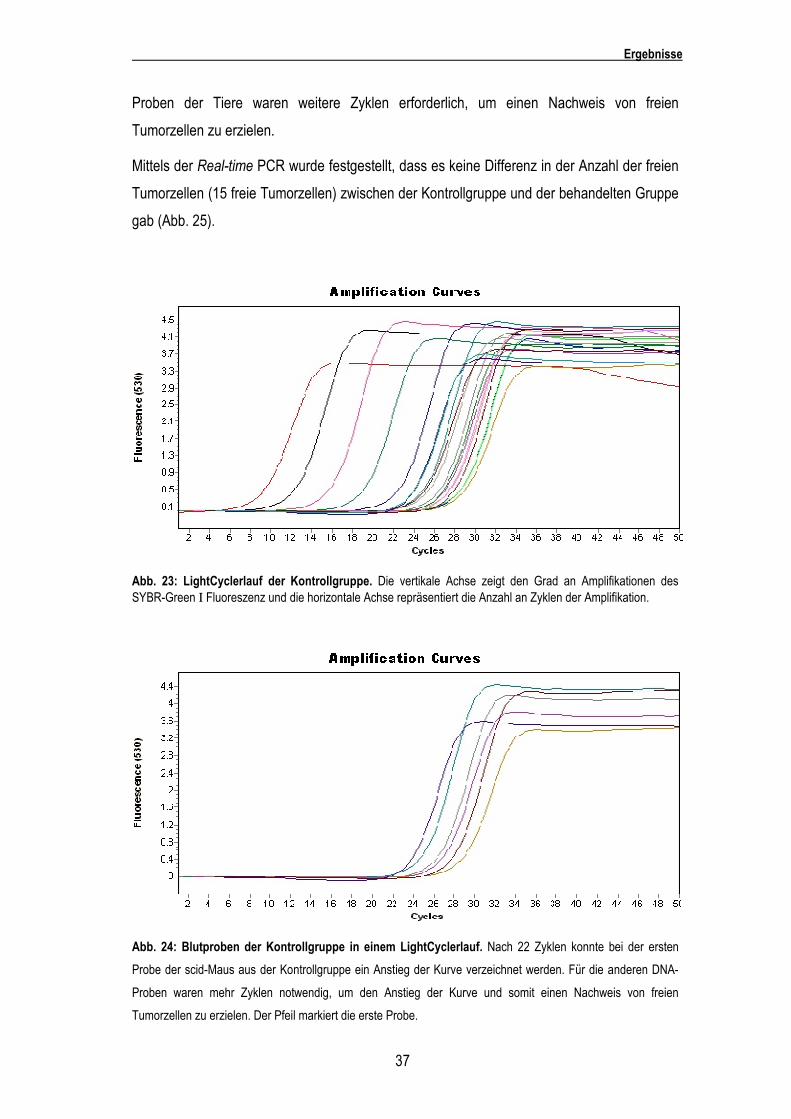
In Abbildung 23 ist ein typischer LightCycler Lauf dargestellt. Bei Zunahme der Anzahl der
Durchgänge kommt es zu einer Amplifikation der DNA. Es besteht eine Abhängigkeit
zwischen der Anzahl der freien Tumorzellen im Blut und der Anzahl der Zyklen die benötigt
werden, um einen Anstieg der Amplifikationskurve auszulösen. Bei wenigen DNA-Zyklen ist
ein Anstieg der Amplifikationskurve nur bei den ersten fünf Proben der Eichreihe zu erkennen
(Abb. 23). Probe 1 der Eichkurve (106Tumorzellen) zeigte eine erkennbare Amplifikation nach
sechs Zyklen, Probe 2 (105 Tumorzellen) nach acht Zyklen, Probe 3 (104 Tumorzellen) nach
zwölf Zyklen, Probe 4 (103 Tumorzellen) nach 15 Zyklen und Probe 5 (102 Tumorzellen) nach
18 Zyklen. Bei Probe 6 der Eichkurve (101 Tumorzellen) zeigte sich ein Anstieg der Kurve erst
nach ca. 21 Amplifikationszyklen. Hierbei gab es schon einzelne DNA-Proben der scid-Mäuse,
bei denen ein Anstieg vor der letzten Eichprobe zu beobachten war. Die danach folgenden
Kurven der DNA-Proben der scid-Mäuse der Kontrollgruppe wiesen demnach nur noch
geringe Anzahlen an freien Tumorzellen auf. Zur besseren Übersicht zeigt Abbildung 24 den
Kurvenverlauf von einigen Blutproben ohne die Eichkurve. Die erste DNA-Probe der scid-
Mäuse verzeichnete nach 22 Zyklen der Amplifikation einen Anstieg. Für die weiteren DNA-
Ergebnisse
37
Proben der Tiere waren weitere Zyklen erforderlich, um einen Nachweis von freien
Tumorzellen zu erzielen.
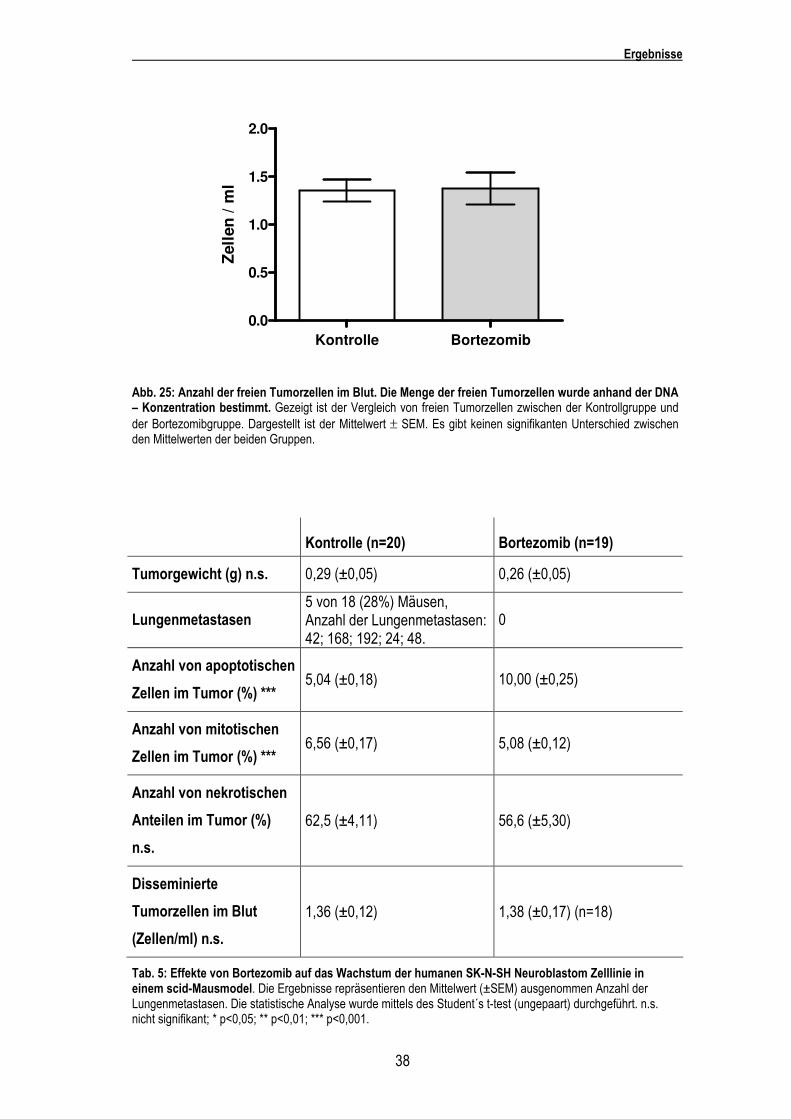
Mittels der Real-time PCR wurde festgestellt, dass es keine Differenz in der Anzahl der freien
Tumorzellen (15 freie Tumorzellen) zwischen der Kontrollgruppe und der behandelten Gruppe
gab (Abb. 25).
Abb. 23: LightCyclerlauf der Kontrollgruppe. Die vertikale Achse zeigt den Grad an Amplifikationen des SYBR-Green Ι Fluoreszenz und die horizontale Achse repräsentiert die Anzahl an Zyklen der Amplifikation.
Abb. 24: Blutproben der Kontrollgruppe in einem LightCyclerlauf. Nach 22 Zyklen konnte bei der ersten
Probe der scid-Maus aus der Kontrollgruppe ein Anstieg der Kurve verzeichnet werden. Für die anderen DNA-
Proben waren mehr Zyklen notwendig, um den Anstieg der Kurve und somit einen Nachweis von freien
Tumorzellen zu erzielen. Der Pfeil markiert die erste Probe.
Ergebnisse
38
Abb. 25: Anzahl der freien Tumorzellen im Blut. Die Menge der freien Tumorzellen wurde anhand der DNA – Konzentration bestimmt. Gezeigt ist der Vergleich von freien Tumorzellen zwischen der Kontrollgruppe und der Bortezomibgruppe. Dargestellt ist der Mittelwert ± SEM. Es gibt keinen signifikanten Unterschied zwischen den Mittelwerten der beiden Gruppen.
Kontrolle (n=20) Bortezomib (n=19)
Tumorgewicht (g) n.s. 0,29 (±0,05) 0,26 (±0,05)
Lungenmetastasen 5 von 18 (28%) Mäusen, Anzahl der Lungenmetastasen: 42; 168; 192; 24; 48.
0
Anzahl von apoptotischen
Zellen im Tumor (%) *** 5,04 (±0,18) 10,00 (±0,25)
Anzahl von mitotischen
Zellen im Tumor (%) *** 6,56 (±0,17) 5,08 (±0,12)
Anzahl von nekrotischen
Anteilen im Tumor (%)
n.s.
62,5 (±4,11) 56,6 (±5,30)
Disseminierte
Tumorzellen im Blut
(Zellen/ml) n.s.
1,36 (±0,12) 1,38 (±0,17) (n=18)
Tab. 5: Effekte von Bortezomib auf das Wachstum der humanen SK-N-SH Neuroblastom Zelllinie in einem scid-Mausmodel. Die Ergebnisse repräsentieren den Mittelwert (±SEM) ausgenommen Anzahl der Lungenmetastasen. Die statistische Analyse wurde mittels des Student´s t-test (ungepaart) durchgeführt. n.s. nicht signifikant; * p<0,05; ** p<0,01; *** p<0,001.
Kontrolle Bortezomib
0.0
0.5
1.0
1.5
2.0
Zell
en
/ m
l
Ergebnisse
39
4.2.4 Imaging
4.2.4.1 MRT
Zur Bestimmung der Tumorvolumina wurden MRT-Bilder angefertigt und diese mit dem Voxel
Programm (Voxel Man Gruppe, Hamburg) bearbeitet. Die Aufnahmen wurden im MRT sowohl
in T1 als auch in T2 Gewichtung angefertigt. Die anatomischen Strukturen waren mit der T1
Gewichtung sehr gut zu beurteilen, jedoch waren die Abgrenzung des Neuroblastoms, die
nekrotischen Anteile und die perifokalen Ödeme in den T2 gewichteten Aufnahmen deutlich
besser zu bestimmen. Für alle weiteren Beurteilungen und Berechnungen wurden deswegen
die sagittal oder coronal geschnittenen T2 Bilder benutzt.
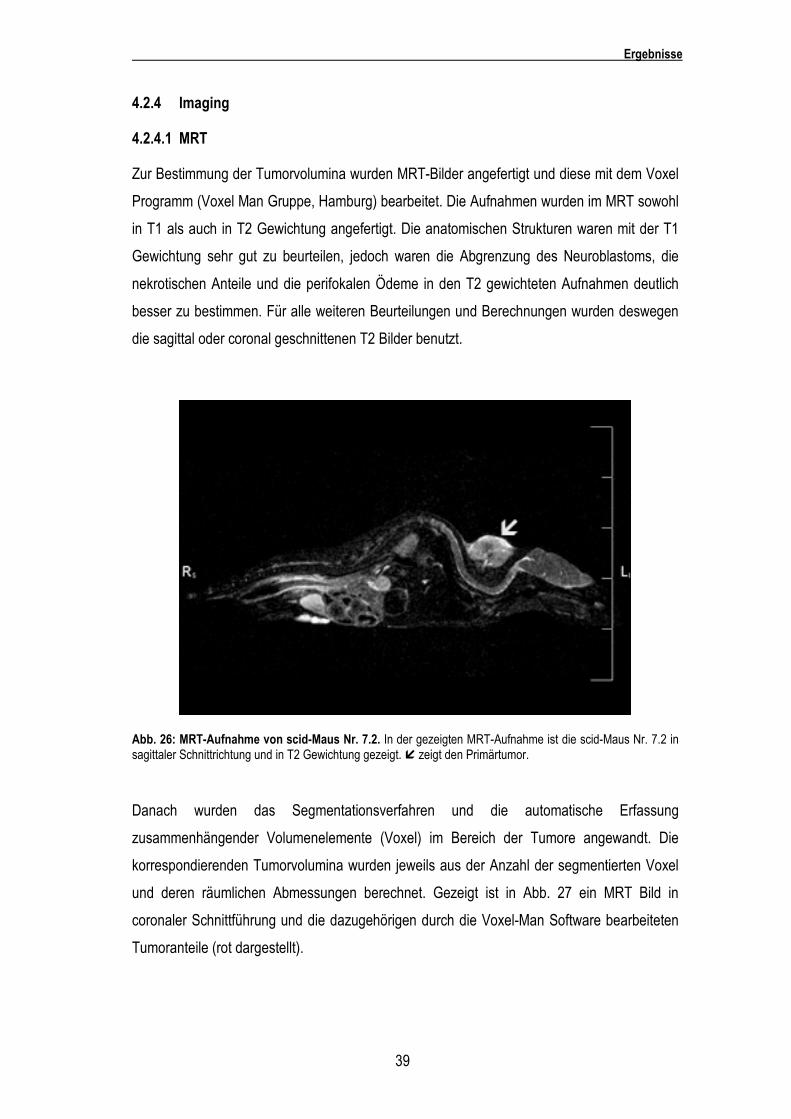
Abb. 26: MRT-Aufnahme von scid-Maus Nr. 7.2. In der gezeigten MRT-Aufnahme ist die scid-Maus Nr. 7.2 in sagittaler Schnittrichtung und in T2 Gewichtung gezeigt. � zeigt den Primärtumor.
Danach wurden das Segmentationsverfahren und die automatische Erfassung
zusammenhängender Volumenelemente (Voxel) im Bereich der Tumore angewandt. Die
korrespondierenden Tumorvolumina wurden jeweils aus der Anzahl der segmentierten Voxel
und deren räumlichen Abmessungen berechnet. Gezeigt ist in Abb. 27 ein MRT Bild in
coronaler Schnittführung und die dazugehörigen durch die Voxel-Man Software bearbeiteten
Tumoranteile (rot dargestellt).
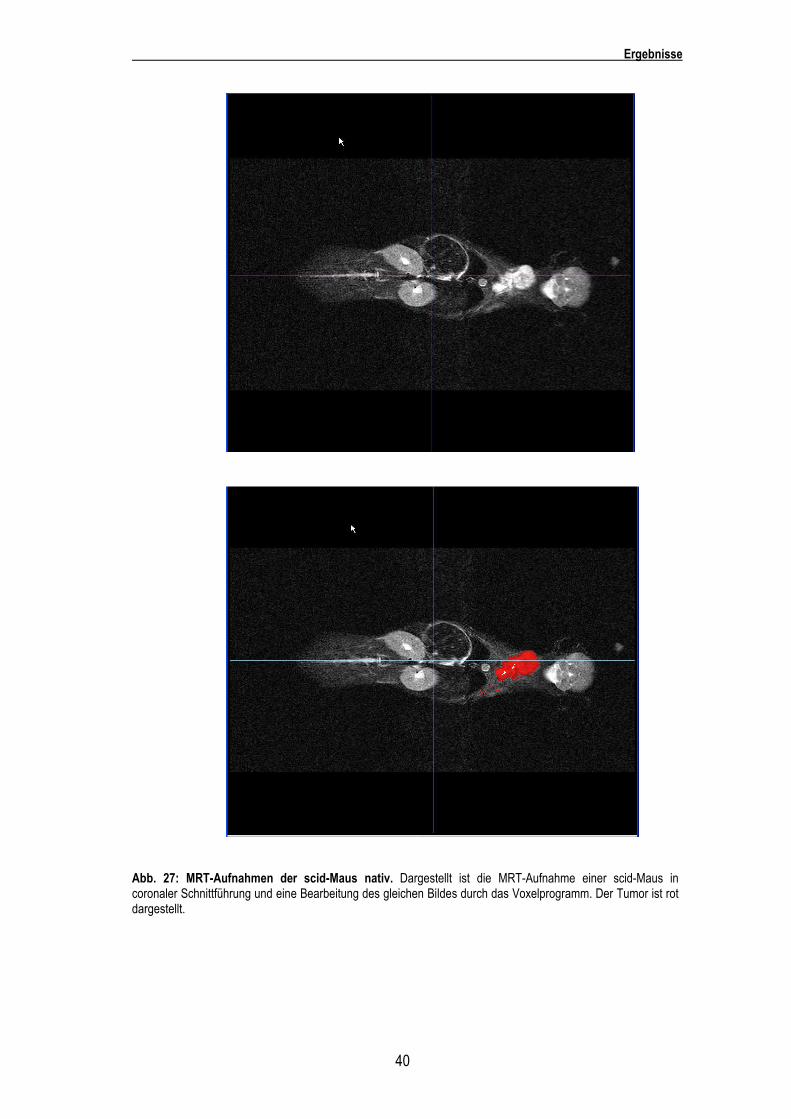
Ergebnisse
40
Abb. 27: MRT-Aufnahmen der scid-Maus nativ. Dargestellt ist die MRT-Aufnahme einer scid-Maus in coronaler Schnittführung und eine Bearbeitung des gleichen Bildes durch das Voxelprogramm. Der Tumor ist rot dargestellt.
Ergebnisse
41
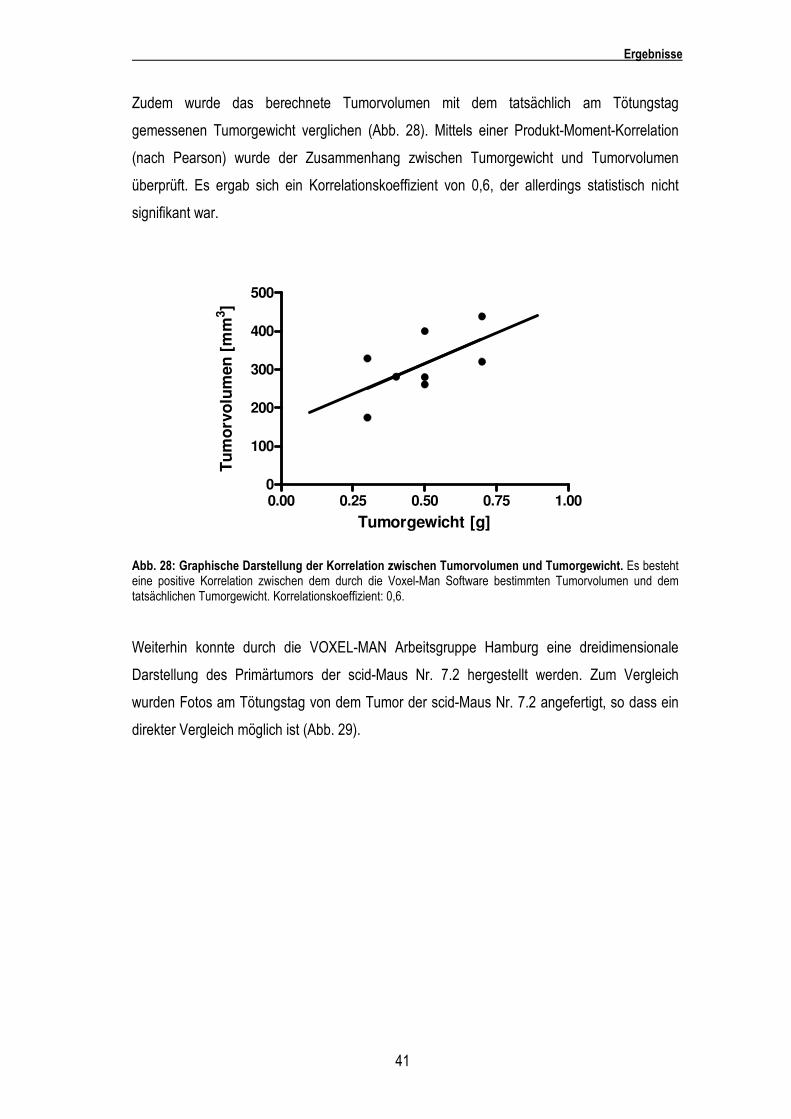
Zudem wurde das berechnete Tumorvolumen mit dem tatsächlich am Tötungstag
gemessenen Tumorgewicht verglichen (Abb. 28). Mittels einer Produkt-Moment-Korrelation
(nach Pearson) wurde der Zusammenhang zwischen Tumorgewicht und Tumorvolumen
überprüft. Es ergab sich ein Korrelationskoeffizient von 0,6, der allerdings statistisch nicht
signifikant war.
Abb. 28: Graphische Darstellung der Korrelation zwischen Tumorvolumen und Tumorgewicht. Es besteht eine positive Korrelation zwischen dem durch die Voxel-Man Software bestimmten Tumorvolumen und dem tatsächlichen Tumorgewicht. Korrelationskoeffizient: 0,6.
Weiterhin konnte durch die VOXEL-MAN Arbeitsgruppe Hamburg eine dreidimensionale
Darstellung des Primärtumors der scid-Maus Nr. 7.2 hergestellt werden. Zum Vergleich
wurden Fotos am Tötungstag von dem Tumor der scid-Maus Nr. 7.2 angefertigt, so dass ein
direkter Vergleich möglich ist (Abb. 29).
0.00 0.25 0.50 0.75 1.000
100
200
300
400
500
Tumorgewicht [g]
Tu
mo
rvo
lum
en
[m
m3]

Ergebnisse
42
Abb. 29: Mittels Computerbearbeitung erstelltes 3D Bild eines Tumors sowie Foto desselben Tumors direkt nach Entnahme am Tötungstag. Die Abbildungen ermöglichen den direkten Vergleich zwischen der durch die Voxel-Man Software bearbeiteten MRT Tumordarstellung und dem Foto des resizierten nativen Tumors. Hierbei ist zu sehen, dass die Bearbeitung mittels des Voxel Programms eine gute Methode darstellt, um das Aussehen und die Form eines Tumors in situ nachzubilden.
Ergebnisse
43
4.2.4.2 PET-CT
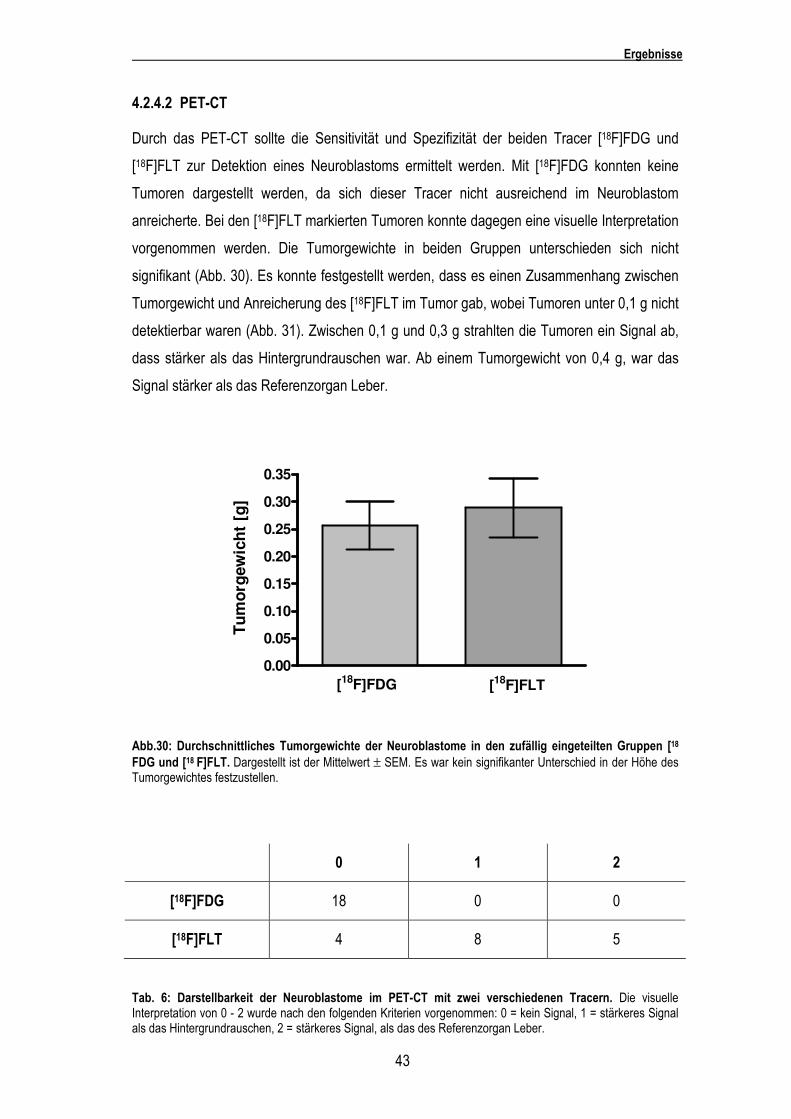
Durch das PET-CT sollte die Sensitivität und Spezifizität der beiden Tracer [18F]FDG und
[18F]FLT zur Detektion eines Neuroblastoms ermittelt werden. Mit [18F]FDG konnten keine
Tumoren dargestellt werden, da sich dieser Tracer nicht ausreichend im Neuroblastom
anreicherte. Bei den [18F]FLT markierten Tumoren konnte dagegen eine visuelle Interpretation
vorgenommen werden. Die Tumorgewichte in beiden Gruppen unterschieden sich nicht
signifikant (Abb. 30). Es konnte festgestellt werden, dass es einen Zusammenhang zwischen
Tumorgewicht und Anreicherung des [18F]FLT im Tumor gab, wobei Tumoren unter 0,1 g nicht
detektierbar waren (Abb. 31). Zwischen 0,1 g und 0,3 g strahlten die Tumoren ein Signal ab,
dass stärker als das Hintergrundrauschen war. Ab einem Tumorgewicht von 0,4 g, war das
Signal stärker als das Referenzorgan Leber.
Abb.30: Durchschnittliches Tumorgewichte der Neuroblastome in den zufällig eingeteilten Gruppen [18
FDG und [18 F]FLT. Dargestellt ist der Mittelwert ± SEM. Es war kein signifikanter Unterschied in der Höhe des Tumorgewichtes festzustellen.
0 1 2
[18F]FDG 18 0 0
[18F]FLT 4 8 5
Tab. 6: Darstellbarkeit der Neuroblastome im PET-CT mit zwei verschiedenen Tracern. Die visuelle Interpretation von 0 - 2 wurde nach den folgenden Kriterien vorgenommen: 0 = kein Signal, 1 = stärkeres Signal als das Hintergrundrauschen, 2 = stärkeres Signal, als das des Referenzorgan Leber.
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
[18F]FDG [18F]FLT
Tu
mo
rgew
ich
t [g
]

Ergebnisse
44
Abb. 31: Korrelation des Tumorgewichtes mit der Anreicherung des Tracers [18F]FLT. Bei Zunahme des Tumorgewichtes kam es zu einer stärkeren Anreicherung von [18F]FLT in den Neuroblastomen. Ab einem Schwellenwert von ca. 0,1 g Tumorgewicht war eine visuelle Interpretation der Neuroblastome möglich. Die Beurteilung wurde nach den oben genannten Kriterien durchgeführt (siehe Tab. 6).
Abb. 32: Axial geschnittene CT- Aufnahme der scid-Mäuse im Reck des PET-CT. Die CT- Aufnahmen wurden dann zur Zuordnung der anatomischen Strukturen in den PET Aufnahmen benutzt.
[18F]FLT
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
0
1
2
Tumorgewicht [g]
vis
uell
e I
nte
rpre
tati
on
Ergebnisse
45
A B
B
T
B
T
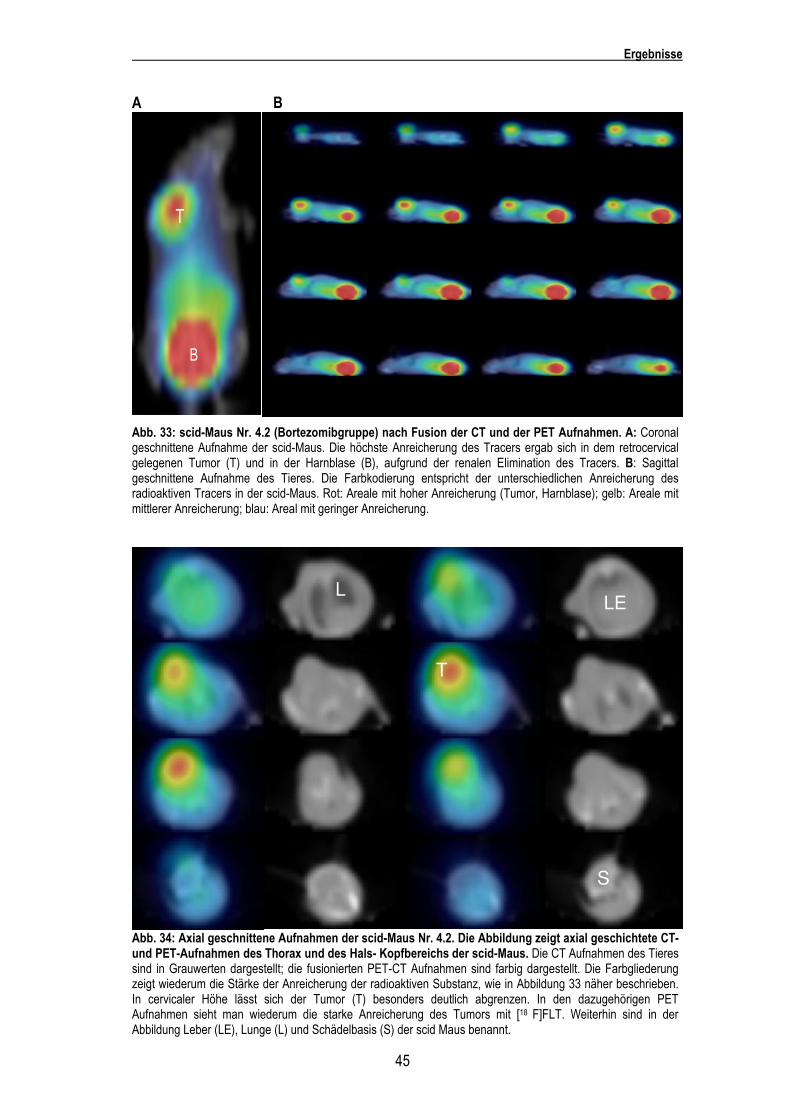
Abb. 33: scid-Maus Nr. 4.2 (Bortezomibgruppe) nach Fusion der CT und der PET Aufnahmen. A: Coronal geschnittene Aufnahme der scid-Maus. Die höchste Anreicherung des Tracers ergab sich in dem retrocervical gelegenen Tumor (T) und in der Harnblase (B), aufgrund der renalen Elimination des Tracers. B: Sagittal geschnittene Aufnahme des Tieres. Die Farbkodierung entspricht der unterschiedlichen Anreicherung des radioaktiven Tracers in der scid-Maus. Rot: Areale mit hoher Anreicherung (Tumor, Harnblase); gelb: Areale mit mittlerer Anreicherung; blau: Areal mit geringer Anreicherung.
Abb. 34: Axial geschnittene Aufnahmen der scid-Maus Nr. 4.2. Die Abbildung zeigt axial geschichtete CT- und PET-Aufnahmen des Thorax und des Hals- Kopfbereichs der scid-Maus. Die CT Aufnahmen des Tieres sind in Grauwerten dargestellt; die fusionierten PET-CT Aufnahmen sind farbig dargestellt. Die Farbgliederung zeigt wiederum die Stärke der Anreicherung der radioaktiven Substanz, wie in Abbildung 33 näher beschrieben. In cervicaler Höhe lässt sich der Tumor (T) besonders deutlich abgrenzen. In den dazugehörigen PET Aufnahmen sieht man wiederum die starke Anreicherung des Tumors mit [18 F]FLT. Weiterhin sind in der Abbildung Leber (LE), Lunge (L) und Schädelbasis (S) der scid Maus benannt.
L LE
T
S
Ergebnisse
46
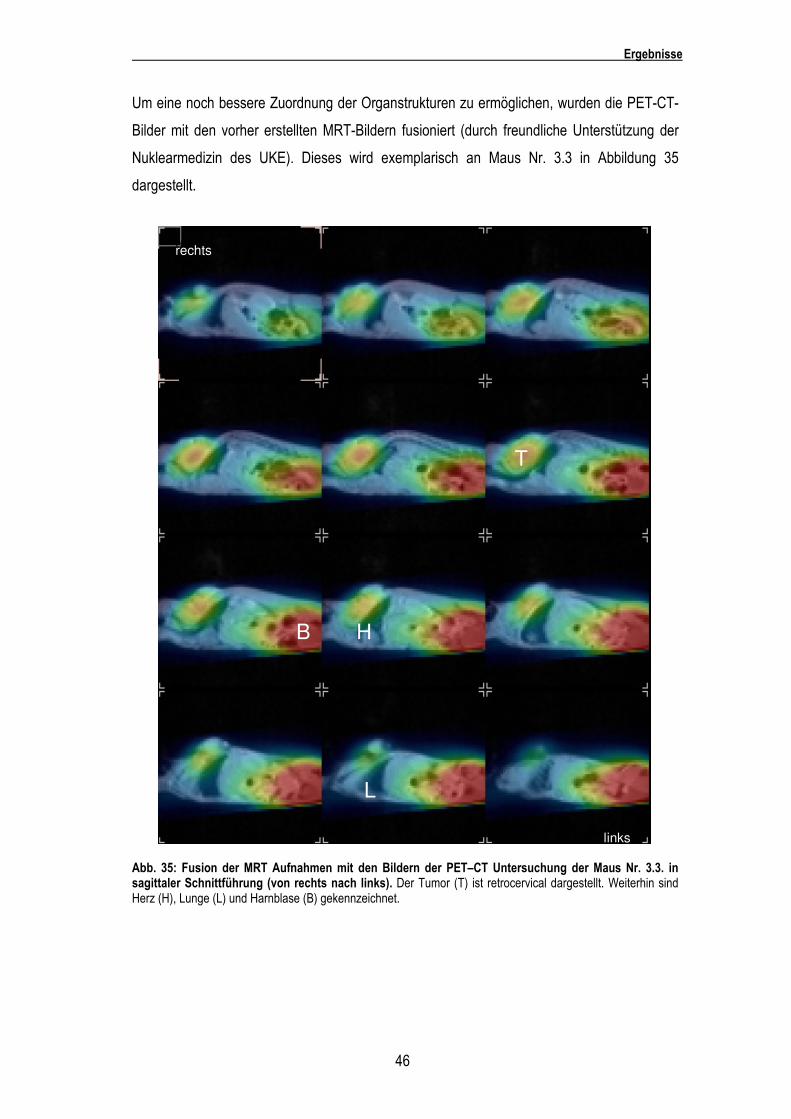
Um eine noch bessere Zuordnung der Organstrukturen zu ermöglichen, wurden die PET-CT-
Bilder mit den vorher erstellten MRT-Bildern fusioniert (durch freundliche Unterstützung der
Nuklearmedizin des UKE). Dieses wird exemplarisch an Maus Nr. 3.3 in Abbildung 35
dargestellt.
Abb. 35: Fusion der MRT Aufnahmen mit den Bildern der PET–CT Untersuchung der Maus Nr. 3.3. in sagittaler Schnittführung (von rechts nach links). Der Tumor (T) ist retrocervical dargestellt. Weiterhin sind Herz (H), Lunge (L) und Harnblase (B) gekennzeichnet.
L
T
HB
rechtsrechts
links
L
T
HB
rechtsrechts
links

Diskussion
47
5 Diskussion
Das Neuroblastom ist bei Kindern der häufigste solide extrakranielle Tumor, dessen Prognose
vom Alter (> ein Jahr = schlechtere Prognose) und vom Ausbreitungsstadium dieses
embryonalen Tumors abhängt. Wird bei einem Kind, das älter als ein Jahr alt ist, ein
Neuroblastom des Stadium IV diagnostiziert, beträgt die mediane
Fünfjahresüberlebenswahrscheinlichkeit 20 % (Koletzko, 2003). Die bisherigen
Therapieansätze, bestehend aus Kombinationen von Operationen, Chemotherapeutika und
homologer Stammzelltherapie, zeigten bisher nicht den erhofften Erfolg (Morgenstern et al.,
2004). Daher ist es dringend erforderlich nach neuen Therapieansätzen zu suchen.
In der vorliegenden Arbeit wurde zunächst in vitro überprüft, ob der Proteasomeninhibitor
Bortezomib auf die von uns getesteten humanen Neuroblastomzelllinien einen
antiproliferativen Effekt besitzt.
Es konnte gezeigt werden, dass das Medikament auf alle sieben getesteten Zelllinien eine
konzentrationsabhängige inhibitorische Wirkung bezüglich des Tumorzellwachstums hatte.
Alle Zelllinien zeigten einen massiven Abfall der Zellzahl bereits bei einer
Bortezomibkonzentration von 0,02 µg / ml. Bei der Zelllinie LS bewirkte dieses eine
Wachstumshemmung auf 6,9% der Kontrolle, bei der Zelllinie Kelly auf 7,8% der Kontrolle, bei
SK-N-SH auf 9% der Kontrolle. Bei den Zelllinien LAN-1 und LAN-5 zeigte sich eine
Hemmung auf 27% des Kontrollwertes. Bei der Zelllinie IMR-32 konnte das
Tumorzellwachstum auf 23% des Kontrollwertes gehemmt werden. Einzige Ausnahme war die
Zelllinie SH-SY5Y, die bereits bei einer Konzentration von 0,002 µg / ml einen starken Abfall
der Zellproliferation (auf 39% der Kontrolle) bewirkt hatte, welches sich in dem niedrigen IC50
Wert von 1,9 ng / ml widerspiegelt. Im Allgemeinen lagen die IC50 Werte zwischen den sieben
Zelllinien zwischen 1,9 ng / ml (SH-SY5Y) und 8,4 ng / ml (LAN-1). Auffallend ist die hohe
Standardabweichung von SH-SY5Y bei der Konzentration von 0,002 µg / ml von 39,6. Bei
allen anderen Zelllinien lag die höchste Standardabweichung ebenfalls bei dieser
Konzentration.
In der Arbeit von Brignole et al., (2006) konnte ebenfalls gezeigt werden, dass Bortezomib
eine statistisch signifikante Inhibierung des Zellwachstums bewirkt, welche zeit- und
dosisabhängig war. Der IC50 Wert lag nach 24-stündiger Behandlung (bei zehn getesteten
Neuroblastomzelllinien) im Mittel bei 9,6 nM, nach 48 Stunden bei 7,3 nM und nach 72
Stunden bei 6,1 nM. Bei uns lag der IC50 Wert im Mittel (bei sieben Zelllinien) nach 48
Stunden bei 15,1 nM. Die Forschungsgruppe von Brignole et al., (2006) benutzte in vitro

Diskussion
48
teilweise folgende andere Zelllinien (GIE-N, GI-LI-N, GI-CA-N, HTLA-230, SK-NBE2c und
CAN), wodurch die unterschiedlichen IC50 Werte erklärt werden könnten, wobei die Werte in
der gleichen Größenordnung liegen. Brignole et al., (2006) testeten zusätzlich SH-SY5Y
(Bortezomib sensible Zelllinie) und die Bortezomib resistente Zelllinie HTLA 230 mit kurzen
Behandlungszeiten von einer und vier bzw. acht Stunden mittels wash-out-Assay. Bei der
Zelllinie SH-SY5Y lag der IC50 Wert nach vierstündiger Behandlung bei 60 nM und nach einer
Stunde bei 150 nM. Bei den HTLA 230 Zellen zeigte sich nach einstündiger Behandlung ein
IC50 Wert von 200 nM. Weiterhin zeigte sich, dass es nach einer 72-stündigen Behandlung mit
20 nM Bortezomib zu einer fast 100% igen Inhibierung des Zellwachstums bei der Zelllinie
SH-SY5Y gekommen war. Bei den weiter getesteten Zelllinien (LAN-1, SK-NBE2c, IMR-32
und GI-LI-N) war es zu ähnlichen Resultaten gekommen (Brignole et al., 2007) In der
vorliegenden Studie lag die nahezu 100% ige Wachstumsinhibierung von SH-SY5Y nach 48
Stunden bei 52 nM.
Valentiner et al., (2005) konnten bei der Testung von PPAR-γ Agonisten ebenso bei SH-SY5Y
die stärkste Inhibierung des Neuroblastomzellwachstums in vitro nachweisen. Den
Ergebnissen von Valentiner et al., (2005) und Brignole et al., (2006) zufolge, weist die Zelllinie
SH-SY5Y anscheinend eine gute Chemosensibilität auf. Gründe für die hohe Sensitivität
dieser Neuroblastomzelllinie sind in der Literatur nicht beschrieben.
Hamner et al., (2007) untersuchten die Neuroblastomzelllinien NB-1691, CHLA-255 und SK-
N-AS mittels verschiedener Konzentrationen des Medikamentes Bortezomib und
unterschiedlichen Behandlungszeiten. Hierbei konnte ebenfalls eine zeit- und dosisabhängige
antiproliferierende Wirkung erzielt werden. Nach 24 stündiger Behandlungszeit (1 µmol / l)
konnte eine dosisabhängige Abnahme der Proliferation bei den 3 Zelllinien auf 13% (NB-
1691), 15% (CHLA-255) und 26% (SK-N-AS) bestimmt werden (p<0,1). Bei der geringsten
Dosis (10 nmol / l) wurde eine Zunahme der antiproliferativen Wirkung mit steigender
Behandlungsdauer (24, 48 und 72 Stunden) nachgewiesen (Hamner et al., 2007).
Weiterhin berichtete die Forschungsgruppe von Michaelis et al., (2006), dass Bortezomib bei
den von ihnen getesteten chemosensiblen Neuroblastomzelllinien (IMR-32 und Be(2)-C) und
medikamentenresistenten Zelllinien (UKF-NB-3rVCR10, UKF-NB-3 und UKF NB-3rDox20) bei
bereits niedrigsten Konzentrationen (Bsp. UKF-NB-3rVCR10 = 8,49 ± 2,07 nM; UKF NB-
3rDox20 = 4,0 ± 0,32 nM) eine Inhibierung des Zellzyklus bewirkte und zusätzlich Apoptose
induzierte. Ling et al., (2003) konnten eine Zunahme der Apoptose nach Behandlung mit
Bortezomib am nichtkleinzelligen Lungenkarzinom feststellen, wobei es zu einer Arretierung

Diskussion
49
des Zellzyklus in der G2 – M – Phase kam. Weitere Testungen mit Bortezomib wurden von
Sunwoo et al., (2001) an humanen Plattenepithelkarzinomen durchgeführt und wiederum kam
es zu einer Abnahme der Zellzahl und zudem zu einer Inaktivierung von NF-κB. Studien von
Ruiz et al., (2006) haben die Effekte von Bortezomib mit einem neuen irreversiblen
Proteasomeninhibitor (NPI-0052) an 20S Chymotryptic Proteasomen verglichen. Dazu wurden
isolierte chronisch lymphatische Leukämie (CLL) Zellen in vitro bezüglich ihrer Apoptose
getestet. Es konnte gezeigt werden, dass Bortezomib und NPI-0052 einen proapoptotischen
Effekt an isolierten CLL Zellen besaßen, wobei NPI-0052 sogar eine noch stärkere Wirkung
aufwies.
Nach den hervorragenden antiproliferativen Effekten in vitro wurde dann die Zelllinie SK-N-SH
für die in vivo Versuche ausgewählt. Diese Zelllinie wurde verwendet, da SK-N-SH in bereits
durchgeführten Tierversuchen ein schnelles Tumorwachstum und eine hohe
Metastasierungsrate aufwies (Ursula Valentiner, in Vorbereitung).
Die 40 scid-Mäuse wurden in zwei Gruppen eingeteilt und zwei Tage nach der s.c. Injektion
der Tumorzellen erhielt dann die eine Gruppe den Proteasomeninibitor Bortezomib in einer
Konzentration von 1 mg / kg KG (Michaelis et al., 2006; Brignole et al., 2006). Diese
Dosierung wurde ausgewählt, weil Sunwoo et al., (2001) berichteten, dass bei einer Gabe von
1 mg / kg KG eine signifikante Inhibierung des Tumorwachstums der humanen
Plattenzellkarzinomzelllinie UM-SCC-11B zu erzielen war. Eine höher dosierte
Medikamentengabe von 2 mg / kg KG hatte zum Tod (drei von sechs Mäusen) geführt. In der
von uns durchgeführten Studie erhielt die Kontrollgruppe PBS.
In dieser Studie erhielten die Tiere die Testsubstanz relativ direkt nach der Tumorzellinjektion.
Teicher (2006) berichtete zwar, dass Studien, bei denen ein Medikament erst nach erfolgtem
Tumorwachstum verabreicht wurde, eine viel höhere klinische Relevanz haben als Studien, in
denen das Medikament bereits vor Anwachsen des Tumors injiziert wurde. Dabei ist aber zu
bedenken, dass solide Tumoren fast nie primär mit einer Chemotherapie behandelt werden,
sondern der Primärtumor meist erst operativ entfernt oder verkleinert wird. Anschließend
erfolgt dann eine Chemotherapie, durch die okkulte Metastasen abgetötet werden sollen
(Koletzko, 2003).
Es hat sich gezeigt, dass eine Injektion von Bortezomib alle 72 Stunden für die Inhibierung der
Aktivität des 26S Proteasomen optimal ist (Adams et al., 1999). So wurde von uns in der
vorliegenden Arbeit das Medikament ebenfalls zweimal pro Woche verabreicht.

Diskussion
50
Es zeigte sich nach der Tötung der Tiere, dass das Medikament Bortezomib keinen
signifikanten Unterschied hinsichtlich des Primärtumorgewichtes zwischen den beiden
Gruppen bewirkt hatte. Im Mittel hatte die Kontrollgruppe ein Primärtumorgewicht von 0,29 g
und die Bortezomibgruppe von 0,26 g (Abb. 12). Dieses Tumorgewicht ist im Vergleich zu
bisherigen Tierversuchstudien eher gering. Thies et al., (2007) zeigten in den von ihnen
getesteten Melanomzelllinien (G361, MV3, FemX1, UISO-Mel 6,MeWo und Lox) auf, dass das
durchschnittliche Tumorgewicht eine Spannbreite zwischen 0,1 - 2,4 g bei unterschiedlichen
Anwachszeiten von 25 - 90 Tagen aufwies. Dieses zeigte, dass das Tumorgewicht unserer
Tiere (0,1 – 0,7 g) eher im unteren Drittel der Gewichtsskala anzusiedeln wäre. Im Gegensatz
zu vorhergehenden Versuchen, bei denen eine vierwöchige Anwachszeit ausreichend war,
wäre im vorliegenden Experiment eine längere Versuchsdauer vorteilhaft gewesen. Diese
Überlegungen werden durch die Arbeit von Teicher (2006) bestätigt, da das Tumorwachstum
logarithmisch mit der Versuchsdauer ansteigt. Durch die zeitliche Planung der bildgebenden
Untersuchungen war eine Terminänderung jedoch während des Tierversuches nicht mehr
möglich.
Michaelis et al., (2006) beschrieben, dass in Nacktmäusen die drei Neuroblastomzelllinien
(UKF-NB-3rVCR10, UKF-NB-3 and UKF NB-3rDox20) nach Behandlung mit Bortezomib eine
Verminderung des Tumorvolumens der Bortezomibgruppe im Vergleich zu der Kontrollgruppe
zeigten. Ein direkter Vergleich dieser Ergebnisse mit unseren Resultaten ist nicht möglich, da
diese Arbeitsgruppe das Tumorvolumen bestimmt hat und nicht das Tumorgewicht. Gründe
für eine unterschiedliche Wirksamkeit in vivo können sowohl in den unterschiedlich
verwendeten Zelllinien liegen, als auch in der Art des Versuchsaufbaus. So wurden von
Michaelis et al., (2006) nur die Nacktmäuse für die Testung mit Bortezomib benutzt, bei denen
der Tumor auch tastbar angewachsen war. Desgleichen zeigten Brignole et al., (2006) eine
statistisch signifikante Inhibierung des Tumorwachstums der Neuroblastome, welche sich
ebenfalls in einer Reduzierung des Tumorvolumens zeigte. Diese Arbeitsgruppe wendete
verschiedene Tiermodelle an, bei denen sowohl Nacktmäuse als auch scid-Mäuse benutzt
wurden. Die Nacktmäuse erhielten 2 x 106 HTLA-230 Tumorzellen, da diese Zelllinie eine der
resistentesten gegenüber Bortezomib ist, in die Schwanzvene. Die Behandlung mit
Bortezomib oder Kochsalzlösung setzte drei Tage nach Inokulation ein. Die scid-Mäuse
bekamen 1,5 x 106 Zellen der Neuroblastomzelllinie SH-SY5Y in die Nebennierenrinde
gespritzt. Ihre Behandlung mit Bortezomib begann 10 Tage nach Inkorporation der
Tumorzellen. In der vorliegenden Arbeit wurde ein Behandlungszeitraum von 30 Tagen
gewählt. Durch die Experimente von Brignole et al., (2006) zeigte sich eine zunehmende

Diskussion
51
Differenz des Tumorvolumens zwischen behandelter und nichtbehandelter Gruppe mit
steigender Anzahl (30, 40, 50 Tage) der Behandlungstage. Die Unterschiede bezüglich der
Versuchsdurchführung können die differierenden Ergebnisse in Hinblick auf die Wirkung von
Bortezomib im Neuroblastom in vivo erklären.
Bei den Versuchsreihen von Amiri et al., (2004) bezüglich der Therapie vom malignen
Melanom mit Bortezomib, konnte eine signifikante Inhibierung (p<0,001) des Tumorvolumens
nach Gabe von 1,25 mg / kg KG und einer Behandlungsdauer von 36 Tagen im Vergleich zur
Kontrollgruppe erzielt werden. Das geringere Tumorvolumen wurde durch eine Inhibierung der
Angiogenese des Tumors erklärt. Nach Testungen mit Antikörpern gegen Endothelzellmarker
CD31 konnte gezeigt werden, dass mit Bortezomib behandelte Tumoren signifikant weniger
Vaskularisierung aufwiesen als Tumoren der Kontrollgruppen. Le Blanc et al., (2002) konnten
ebenfalls beim malignen Myelom eine signifikante Inhibition des Tumorwachstums
nachweisen, die auf eine Verminderung der Angiogenese und Induzierung der Apoptose
zurückzuführen war. Kahn et al., (2006) beschäftigten sich in ihrer Neuroblastomstudie mit
Kombinationstherapien. Es wurde beschrieben, dass scid-Mäuse, die eine Therapie
bestehend aus Bortezomib und IL-2 erhalten haben, eine Limitierung der Vaskularisation der
Tumorperipherie zeigten, die mit einer Abnahme der Tumorgröße einherging. In Hinsicht auf
Kombinationstherapien gab Teicher (2006) zu bedenken, dass sich immer die Frage stelle, ob
die kombinierten Medikamente einen sich addierenden Effekt hätten oder eine faktisch
unterschiedliche Wirkung aufweisen würden.
Es sollte ferner überprüft werden, ob Bortezomib trotz der Unwirksamkeit bezüglich des
Tumorgewichtes einen Einfluss auf die Anzahl der Apoptosen und Mitosen der Primärtumoren
hat. In der lichtmikroskopischen Untersuchung konnte ein statistisch signifikanter Unterschied
zwischen der Apoptosen- und Mitosenanzahl der beiden getesteten Gruppen eruiert werden.
Dabei hatte die Kontrollgruppe im Mittel einen Apoptoseanteil von 5 % der Gesamtzellzahl
und einen Mitoseanteil von 6,5%. Im Gegensatz dazu konnte bei den Primärtumoren der
Bortezomibgruppe ein Apoptoseanteil von 10 % und ein Mitoseanteil von 5 % ermittelt werden
(Abb. 14 und 16). Michaelis et al., (2006) haben ebenfalls gezeigt, dass Bortezomib einen
Anstieg der Apoptose in den Neuroblastomzelllinien UKF-NB-3 rVCR10, UKF-NB-3 und UKF
NB-3rDox20 im Xenograft bewirkt hatte. Es bestand allerdings ein deutlicher Unterschied
zwischen den verschiedenen Zelllinien bezüglich der Wirkung auf die Apoptoserate. Hierbei
scheint ein Zusammenhang zur Expression des Tumorsuppressorgens p53 zu bestehen. So
reagierten die Wildtyp p53 exprimierenden Zelllinien UKF-NB-3 rVCR10 und UKF-NB-3
sensitiver als die p53 mutierte Zelllinie UKF NB-3rDox20 auf die Behandlung mit Bortezomib.

Diskussion
52
Brignole et al., (2006) haben DNA Fragmente der behandelten Zellen mittels TUNEL und PI-
Färbungen gemessen. Auch diese Arbeitsgruppe konnte zeigen, dass die mit Bortezomib
behandelte Gruppe 62 % TUNEL positive Zellen aufwies und es konnte ein statistisch
signifikant höherer Anteil an apoptotischen Zellen mittels PI Färbung erzielt werden
(Bortezomib 29 % versus Kontrolle 5 %, p<0,001). Die Induktion von Apoptose erfolgte durch
die Aktivierung von Aspartat spezifischen Cystein Proteasen oder Caspasen (Thornberry et
al., 1998). Brignole et al., (2006) konnten diesbezüglich demonstrieren, dass es zu einer
zeitabhängigen signifikant erhöhten Caspaseaktivität der Bortezomibgruppe gegenüber der
Kontrollgruppe gekommen war. Es wurde daraus geschlossen, dass das Tumorsupressorgen
Caspase einen direkten Einfluss auf die Apoptose bei Neuroblastomen aufweist (Brignole et
al., 2006).
Durch vorausgegangene Studien wird deutlich, dass die Induktion der Apoptose nicht der
einzig wichtige Mechanismus zur Behandlung des Neuroblastoms zu sein scheint. Durch
Untersuchungen von Michaelis et al., (2006), Brignole et al., (2006) und Hamner et al., (2007)
wird ersichtlich, dass antiangiogenetische Effekte höchstwahrscheinlich eine sehr wichtige
Rolle in der Wachstumsinhibierung von Neuroblastomen in vivo spielen. Hamner et al., (2007)
gehen weiterhin von einer Herunterregulation der Expression von VEGF (vascular edothelial
growth factor) aus.
Wie auch schon die vorherige Studie von Michaelis et al., (2006) aufzeigte, konnte in der
vorliegenden Arbeit kein signifikanter Unterschied bezüglich des nekrotischen Anteiles des
Tumors zwischen der mit Bortezomib behandelten Gruppe und der Kontrollgruppe festgestellt
werden.
Um den Unterschied der Metastasierung in die Lungen der scid-Mäuse zwischen der
Behandlungsgruppe und der Kontrollgruppe zu bestimmen, wurden die Lungen
lichtmikroskopisch untersucht. Es konnte dargestellt werden, dass es zu einem Unterschied
hinsichtlich der Metastasierung in die Lungen der scid-Mäuse gekommen war. In der
Kontrollgruppe konnten bei fünf von 20 scid-Mäusen Lungenmetastasen nachgewiesen
werden. Hingegen entwickelten die Tiere, die mit Bortezomib behandelt wurden, keine
Lungenmetastasen. Diese Ergebnisse zeigen erstmalig, dass es zu einer pharmakologischen
Beeinflussung der Metastasierungsrate der Neuroblastome in die Lunge der scid-Mäuse
kommen kann.
Insgesamt war die Anzahl der Metastasen in den Lungen der Kontrollgruppe gering. Dieses
könnte durch das geringe Primärtumorgewicht bei einer festgelegten Wachstumsdauer von

Diskussion
53
vier Wochen bedingt sein. In der Literatur wird dieser Zusammenhang zwischen
Primärtumorgewicht und Metastasierung kontrovers diskutiert. Man muss aber davon
ausgehen, dass der Zusammenhang zwischen Primärtumor und Metastasen
zelllinienspezifisch ist. So berichteten Thies et al., (2007), dass es bei den von ihnen
getesteten humanen Melanomzelllinien keine signifikante Beziehung zwischen Tumorgewicht
und Anzahl an Metastasen in der Lunge gab. Jojovic und Schumacher (2000) zeigten, dass
die von ihnen getesteten humanen HT29 Kolonkarzinomzellen einen Zusammenhang
zwischen Gewicht des Primärtumors und Anzahl der Lungenmetastasen aufwiesen. Falls ein
Zusammenhang zwischen Primärtumor und Anzahl an Metastasen existieren sollte, dann
könnte die Verringerung des Gewichts des Primärtumors schon per se eine Reduktion der
Metastasenanzahl bewirken. Nach unveröffentlichten Ergebnissen unseres Labors und den
Resultaten der vorliegenden Arbeit, besteht aber für die Zelllinie SK-N-SH kein
Zusammenhang zwischen Tumorgröße und Metastasen (Abb. 21); außerdem waren die
Tumorgewichte in unseren beiden Untersuchungsgruppen gleich (Abb.12). Daher kann in
dieser Studie von einer spezifisch antimetastatischen Wirkung ausgegangen werden. Eine
antimetastatische Wirkung von Bortezomib bei Neuroblastomen wird in den Untersuchungen
von Khan et al., (2006) bestätigt. Es wurden Unterschiede zwischen behandelten Gruppen
(Bortezomib, IL-2 oder Kombination von Bortezomib und IL-2) und der Kontrollgruppe
hinsichtlich der Metastasen von Neuroblastomen in die Leber beschrieben. Nach der i.v.
Injizierung von TBJ-RFP Zellen, waren die Lebermetastasen der Kontrollgruppe wesentlich
größer und besser vaskularisiert, als die der behandelten Gruppe. Die scid-Mäuse, welche
lediglich mit Bortezomib behandelt wurden, zeigten eine Abnahme der Lebermetastasengröße
sowie der Vaskularisierung. Eine Kombinationstherapie mit Bortezomib und IL-2 hat zu einer
Potenzierung dieser Effekte geführt. Hinzuzufügen ist aber, dass es sich hierbei nicht um eine
Spontanmetastasierung des Primärtumors handelt, sondern um eine i.v. Injektion der
Tumorzellen, sodass der erste Schritt der Metastasierungskaskade fehlt. Es handelt sich also
vielmehr um disseminierte Tumorzellen, die sich nach Durchtritt des Gefäßsinuswalls in der
Leber zu Metastasen formiert haben, welches auch als extravaskuläre Tumorzellproliferation
bezeichnet wird (Nischwitz, 2007). Damit zeigen unsere Versuche erstmalig, dass es zu einer
echten Hemmung von Metastasierung unter Bortezomib kommen kann.
Um weiterhin zu überprüfen, ob Bortezomib einen Einfluss auf die Disseminierung der freien
Tumorzellen im Blut besitzt, wurde eine Real-time PCR durchgeführt. In der vorliegenden
Arbeit konnte gezeigt werden, dass es keinen Unterschied (15 freie Tumorzellen) zwischen

Diskussion
54
der mit Bortezomib behandelten Gruppe und der Kontrollgruppe bezüglich der freien
Tumorzellen im Blut der scid-Mäuse gab.
Schneider et al., (2002) haben beschrieben, dass die Real-time PCR eine gute Möglichkeit
darstellt, um ALU-Sequenzen diverser humaner Tumorentitäten im Xenograftmodell zu
detektieren. Für die gegenwärtige Studie wurde ebenfalls ALU-PCR benutzt, da im haploiden
Genom ungefähr 500.000 Kopien (entspricht 5% der menschlichen DNA) an ALU-Sequenzen
vorhanden sind (Alberts et al., 1999). Schneider (2002) beschrieb, dass ALU-PCR den
Nachweis und eine Quantifizierung der Metastasen zu einem Zeitpunkt ermöglichen, zu dem
eine exakte Auswertung mit der klassischen Histologie noch nicht möglich ist. Demnach bietet
sich diese Methode an, um eine Kontrolle der Metastasierungsrate während des
Tierversuches durchzuführen.
Vor der Tötung der Tiere wurden zur näheren Charakterisierung des Tumors und seiner
möglichen Metastasen bildgebende Untersuchungen durchgeführt. Es wurde ein
halbautomatisches Segmentationsverfahren basierend auf der interaktiven Festlegung von
Schwellenwertbereichen (verschiedene Grauabstufungen des Tumors) und der
anschließenden, automatischen Erfassung zusammenhängender Volumenelemente (Voxel)
im Bereich der Tumore angewandt. Die korrespondierenden Tumorvolumina wurden jeweils
aus der Anzahl der segmentierten Voxel und deren räumlichen Abmessungen berechnet.
Hinsichtlich des Tumorwachstumsverhaltens konnte allerdings kein deutlicher Unterschied
zwischen der Bortezomib- und der Kontrollgruppe eruiert werden. Ein Befund, der der
späteren ex vivo Untersuchung der Tumorgewichte entsprach.
Mit der Voxel-Man Software hatte man durch die Bestimmung der Gesamtmenge der Voxel
des Tumors die Möglichkeit, das tatsächliche Tumorvolumen zu ermitteln. In Abbildung 30
wird gezeigt, wie sich eine Rekonstruktion des Tumors einer scid-Maus darstellt.
Anschließend wurde das Tumorvolumen berechnet, welches mit dem am Tötungstag
tatsächlichen gemessenen Gewicht mit einer leichten Korrelation (r = 0,6; statistisch n.s.)
übereinstimmte. Diese Ergebnisse demonstrieren an sich zwar schon die guten Ansätze der
intravitalen Tumorvolumetrie mittels MRT-Bildgebung und Bildnachbearbeitung mit der
Software Voxel-Man, zeigen aber dennoch, dass es zu weiteren Verfeinerungen kommen
muss, um eine noch höhere Korrelation zwischen Tumorgewicht und Tumorvolumen zu
erzielen, die dann auch für die klinische Onkologie von großem Interesse sein dürfte.
Verschiedene Faktoren könnten die Qualität der Segmentationen beeinflusst haben, wodurch
sich kein höherer Korrelationskoeffizient erzielen ließ. Bei unseren Untersuchungen waren die

Diskussion
55
Formen der Tumoren sehr unterschiedlich (kugelig bzw. länglich auslaufend, kompakt oder
gelappt). Bei gelappten Tumoren war die Korrelation zwischen Tumorgewicht und
Tumorvolumen schlechter als bei kugelförmigen Tumoren.
Die Forschungsgruppe von Cornelissen et al., 2005 verglich zwei MRT - basierte Methoden
(die manuelle Messmethode und die halbautomatische Messmethode) mit zwei klassischen
Methoden zur Tumorvolumenbestimmung (Messschieber und Wasserprobe). Bei der
manuellen Messmethode wurde das Tumorvolumen durch Multiplikation von Schnittdicke und
Tumorareal bestimmt. Das Prinzip der halbautomatischen Methode bestand darin, dass aus
dem intensivsten farblich markierten Teil des Tumors erst einmal ein Punkt bestimmt wurde.
Alle Voxel in der Umgebung dieses Punktes, die eine höhere Signalintensität als ein
festgelegter Schwellenwert aufwiesen, wurden in das Tumorvolumen einbezogen. Die
Messschieberauswertung funktionierte nach dem Prinzip der Messung der kurzen und langen
Achse des Tumors mittels eines Messschiebers, woraus das Volumen dann berechnet wurde.
Die weit verbreitete Tumorvolumenbestimmung durch die Wasserprobe erforderte eine
Resektion des Tumors. Das Tumorvolumen wurde danach in einem speziell dafür
hergestellten System gemessen. Dieses System bestand aus drei korrespondierenden
Pipetten, von denen alle drei mit Wasser gefüllt wurden. Durch die Zugabe des Tumors
änderte sich die Wasserstandshöhe, die dann abgelesen werden konnte. Nachteil dieser
Methode liegt darin, dass die Versuchstiere getötet werden müssen. Die Arbeitsgruppe konnte
bei dieser Versuchsreihe eine positive Korrelation von r2 = 0,95 bei p<0,0001 zwischen den
vier Methoden feststellen. Allerdings spielt der Nekroseanteil des Tumors bei dem
halbautomatischen Segmentationsverfahren eine Rolle (Cornelissen et al., 2005). Bei
Tumoren mit einen hohen Anteil an Nekrose (Tumoren > 1,5 cm3) unterschieden sich die
Ergebnisse der Tumorvolumenbestimmung mittels der halbautomatischen Methode deutlich
von der Messung mittels der Wasserprobe. Dies liegt darin begründet, dass die nekrotischen
Anteile bei der halbautomatischen Methode automatisch detektiert werden können und nicht
in die Messung mit einbezogen werden (Cornelissen et al., 2005). In der von uns
ausgeführten Tumorauswertung mittels der Voxel-Man Software könnten die Nekrosen
ebenfalls herausgerechnet werden. Mit halbautomatischen MRT - basierten Methoden zur
Tumorvolumenbestimmung ergäbe sich daher die Möglichkeit der genaueren Bestimmung der
Tumormasse. Dieses ist von Bedeutung, da einige Medikamente die apoptotischen Vorgänge
der Tumorzelle stimulieren. Daraus resultiert aber nicht zwangsläufig eine Abnahme des
Tumorvolumens.

Diskussion
56
Weiterhin haben halbautomatische Methoden den Vorteil, dass sie zur
Tumorwachstumskontrolle im Therapieverlauf eingesetzt werden können. Teicher (2006)
beschreibt in ihrer Arbeit die Wichtigkeit der Kontrolle des Tumorwachstums während des
Versuches, da das Tumorwachstum bis zum letztendlichen Tumorvolumen zwischen
Medikamenten- und Kontrollgruppe unterschiedlich ablaufen kann. Die MRT Kontrolle der
Tumormasse unter medikamentöser Therapie ist für die Beurteilung der Wirkung einer
Substanz von großem Interesse. Hierdurch wird nochmals deutlich, wie wichtig die
Weiterentwicklung der Methoden der nicht invasiven Tumorvolumenbestimmung ist.
Weiterhin könnte das Angrenzen an andere signalintensive Strukturen (z. B. Fett im
Nackenbereich) dafür verantwortlich sein, dass die Korrelation zwischen Tumorgewicht und
Tumorvolumen in dieser Untersuchung nicht noch höher ausgefallen ist. Hinzu kommen
Rundungsfehler bei der Angabe des tatsächlich gemessenen Tumorgewichtes, da nur eine
Kommastelle betrachtet wurde.
Mittels des PET-CTs sollte ein Vergleich der radioaktiven Tracer [18F]FDG und [18F]FLT
hinsichtlich ihrer Sensitivität beim Neuroblastom ermittelt werden. Meist wird zur
Diagnosestellung, Staging und Restaging von Tumoren der Tracer [18F]FDG benutzt
(Hazelton TR und Coppage L, 2005; Avril NE und Weber WA, 2005). Krieger-Hinck et al.,
(2006) zeigten, dass der normalerweise benutzte Tracer [18F]FDG sich im Neuroblastom
schlechter angereichert hatte als [18F]FLT. Dieses konnte durch die Untersuchungen in der
vorliegenden Arbeit bestätigt werden. Es zeigte sich, dass bei den PET-CT Aufnahmen, die
mit [18F]FDG durchgeführt wurden, trotz vergleichbarer Tumorgewichte in beiden Gruppen
(Abb. 30) keine Anreicherung der radioaktiven Substanz in den Tumoren nachweisbar war. Im
Gegensatz dazu konnte bei den Tumoren, die mit [18F]FLT markierten wurden, eine visuelle
Interpretation vorgenommen werden. Abbildung 31 zeigt nochmals, dass es auch bei [18F]FLT
zu keiner Anreicherung bei einem Tumorgewicht < 0,1 g gekommen war. Zwischen 0,1 g und
0,3 g zeigten die Tumoren ein Signal, dass stärker als das Hintergrundrauschen war. Ab
einem Tumorgewicht von 0,4 g war das Signal stärker als das Referenzorgan Leber.
Die Resultate der Untersuchung ergaben eine positive Korrelation zwischen Tumorgewicht
und Anreicherung des [18F]FLT im Tumor. Pacak et al., (2004) berichteten bereits, dass
Tumoren mit einer Tumorgröße unter 0,7 mm nicht im konventionellen humanen PET zu
beurteilen wären. Die Auflösung, die mit diesen Geräten erreicht werden kann, ist niedriger als
die eines Kleintier- PETs und somit ist die Darstellung und Beurteilung sehr kleiner Tumoren
schwierig. Im Gegensatz dazu und vor allem scheinbar kontrovers zu Krieger-Hinck et al.,
(2006), konnte in der vorliegenden Studie eine Anreicherung von [18F]FLT auch bei einem

Diskussion
57
Tumorgewicht < 1,0 g erzielt werden. Der Grund der unterschiedlichen Sensitivitäten der
Detektion der Tumoren liegt in der Benutzung unterschiedlicher PET-CT Geräte. Das in
unserer Studie verwendete Gerät besaß eine höhere Auflösung als das benutzte Gerät von
Krieger-Hinck et al., (2006) und somit konnten auch Tumoren mit sehr niedrigen
Tumorgewichten dargestellt werden.
Vorherige Studien belegten bereits, dass keine Korrelation zwischen der Expression von Ki 67
und der Anreicherung von [18F]FDG und [18F]FLT bestand. Beispiele hierfür sind das
Ösophaguskarzinom (van Westreenen et. al., 2005), das Pankreaskarzinom (Buck et al.,
2001) sowie die Tumoren der Mamma (Smyczek-Gargya et al., 2004) und des Thorax
(Dittmann et al., 2003). Krieger-Hinck et al., (2006) konnten bezüglich der Immunreaktion mit
Ki 67 ebenfalls keine positive Korrelation zwischen der Zellproliferationsrate und dem
[18F]FDG und [18F]FLT uptake bei humanen Neuroblastomen im scid-Mausmodel ermitteln.
Aus diesen Gründen wurden von uns keine weiteren Untersuchungen bezüglich der
Expression von Ki 67 in Neuroblastomen durchgeführt.
Weiterhin wurde in dieser Arbeit im Rahmen einer Pilotstudie untersucht, ob eine Fusion der
Aufnahmen von PET-CT und MRT zu einer weiteren Verbesserung der anatomischen
Zuordnung führen würde. Nach der Fusion der beiden Bilddatensätze konnte gezeigt werden,
dass die anatomischen Strukturen wesentlich besser sichtbar waren, als in den PET Bildern
allein. Eine Anreicherung der radioaktiven Substanz im Tumor war klarer vom umliegenden
Gewebe abzugrenzen. Eine genauere Beurteilbarkeit von Tumoren mittels Fusion der
bildgebenden Verfahren könnte eine bessere Verlaufskontrolle während der Tumortherapie in
der onkologischen Medizin ermöglichen.
Zu bedenken ist aber, dass das MRT und das PET-CT zu unterschiedlichen Zeitpunkten
durchgeführt wurden und somit die Mäuse nicht exakt in der gleichen Position lagen. Es
wurde für diese Arbeit eine retrospektive Fusion durchgeführt. Für weitere Studien könnte
man eine Verbesserung der Untersuchungsergebnisse erzielen, indem das CT Protokoll für
die Mausuntersuchung z.B. durch weniger Kilovolt (kV) optimiert würde, denn dieses würde zu
mehr Weichteilkontrast führen. Durch ein Kleintier-PET würde man noch eine weitere
Optimierung der Auflösung erhalten.
Zusammengefasst haben diese Studien ergeben, dass der Proteasomeninhibitor Bortezomib
hervorragende Wirkungen auf die Proliferation humaner Neuroblastomzelllinien in vitro gezeigt
hatte, die in vivo nur teilweise zu erzielen waren. Es konnte keine Wirkung auf das
Primärtumorgewicht erzielt werden. Jedoch zeigte sich eine statistisch signifikante Zunahme

Diskussion
58
der Apoptoserate und Abnahme der Mitoserate in der Behandlungsgruppe im Vergleich zur
Kontrollgruppe. Erstmalig konnte eine antimetastatische Wirkung in Bezug auf die spontane
Lungenmetastasierung festgestellt werden. Diese Ergebnisse sind aber nur begrenzt zu
verallgemeinern, da nur eine Zellreihe in vivo untersucht wurde und eine längere
Versuchsdauer von Vorteil gewesen wäre. Aufgrund der Ergebnisse der vorliegenden Arbeit
und der aktuellen Ergebnisse anderer Forschungsgruppen sollten weitere Untersuchungen
zur Bedeutung von Proteasomeninhibitoren für die Therapie von Neuroblastomen
durchgeführt werden.

Zusammenfassung
59
6 Zusammenfassung
Insgesamt ist die Prognose von Kindern mit Neuroblastomen im fortgeschrittenen Stadium
schlecht, so dass ein dringender Bedarf an neuen Therapieansätzen besteht. Der
Proteasomeninhibitor Bortezomib (Velcade) zeigte bereits antiproliferative Wirkungen bei
verschiedenen Tumorentitäten sowie nach neuen Studien auch wachstumshemmende
Einflüsse auf das humane Neuroblastom in vitro und in vivo. Ziel dieser Arbeit war es, die
Wirkung des Medikament Bortezomib auf humane Neuroblastomzellen in vitro und in vivo zu
untersuchen. Das Medikament wurde in vitro an sieben humanen Neuroblastomzelllinien
(Kelly, LS, SH-SY5Y, SK-N-SH, IMR-32, LAN-1 und LAN-5) getestet. In vitro konnte bei allen
Zelllinien eine dosisabhängige Inhibierung des Zellwachstums erzielt werden. Für die in vivo
Versuche wurde die Zelllinie SK-N-SH in 40 scid-Mäuse injiziert und die Wirkung von
Bortezomib getestet. Diese Zelllinie wurde ausgewählt, weil sie in vorhergehenden scid
Mausversuchen eine gute Anwachsrate und eine hohe Metastasierungsrate aufwies; in den in
vitro Versuchen zeigte sie eine durchschnittliche Empfindlichkeit auf Bortezomib. Zusätzlich
wurde ein Imaging der Tumoren mittels MRT und PET-CT durchgeführt. Nach Tötung der
scid-Mäuse wurden die Primärtumore und Organe entnommen, histologisch aufgearbeitet und
die Apoptose- und Mitoserate in den Primärtumoren sowie die Lungenmetastasierung
bestimmt. Das Blut der Tiere wurde auf disseminierte Tumorzellen mittels der Real-time PCR
untersucht. In vivo reduzierte Bortezomib das Gewicht der Primärtumoren nicht. Es fand sich
jedoch ein statistisch signifikant höherer Apoptoseanteil und niedrigerer Mitoseanteil bei den
mit Bortezomib behandelten scid-Mäusen. Weiterhin konnte erstmalig gezeigt werden, das
Bortezomib die Lungenmetastasierungsrate von Neuroblastomen pharmakologisch
beeinflusst, da die Medikamentengruppe im Gegensatz zur Kontrollgruppe (5 von 20 Tieren)
keine Lungenmetastasen aufwies. Bei der Untersuchung des Blutes (n = 15 / Maus) gab es
keinen Unterschied bezüglich der disseminierten Tumorzellen zwischen den beiden Gruppen.
Durch MRT und anschließende Tumorrekonstruierung mit Hilfe des Voxel-Man Programms
konnte eine leichte Korrelation (r = 0,6; n.s.) zwischen dem Tumorvolumen und Tumorgewicht
erzielt werden. Mittels PET-CT konnte bestätigt werden, dass sich [18F]FLT zur Untersuchung
des Neuroblastoms besser eignet als [18F]FDG. Eine Fusion der Aufnahmen des MRT und
des PET-CT konnte eine deutliche Verbesserung der Zuordnung anatomischer Strukturen
erzielt werden. Aufgrund der Ergebnisse dieser vorliegenden Arbeit könnte man erwägen,
Bortezomib als adjuvante Therapie beim Neuroblastom in klinischen Studien zu testen.

Literaturverzeichnis
60
7 Literaturverzeichnis
Adams J, Palombella VJ, Sausville EA, Johnson J, Destree A, Lazarus DD, et al. (1999).
Proteasome inhibitors: a novel class of potent and effective antitumor agents. Cancer Res
59:2615 - 2622.
Adams J (2003). The Proteasome: structure, function and role in the cell. Cancer Treat Rev
29: 3 – 9.
Ahn KS, Sethi G, Aggarwal BB (2006). Embelin, an Inhibitor of XIAP, Blocks Nuclear Factor-
{kappa}B (NF-{kappa}B) Signaling Pathway Leading to Suppression of NF-{kappa}B-regulated
Anti-apoptotic and Metastatic Gene Products. Mol Pharmacol 71:209 – 219.
Alberts B, Bray D, Johnson A, Lewis J, Raff K, Walter RP (1999). Lehrbuch der Molekularen
Zellbiologie. Seite: 315.
Amiri KI, Horton LW, La Fleur BJ, Sosman JA, Richmond A (2004). Augmenting
Chemosensitivity of Malignant Melanoma Tumors via Proteasome Inhibition: Implication for
Bortezomib (Velcade, PS-341) as a Therapeutic Agent for Malignant Melanoma. Cancer Res
64:4912 - 4918.
Avril NE und Weber WA (2005). Monitoring response to treatment in patients utilizing PET.
Radio Clin North Am 43:189 – 204.
Berthold F, Boos J, Burdach S, Erttmann R, Henze G, Hermann J, Klingebiel T, Kremens B,
Schilling FH, Schrappe M, Simon T, Hero B (2005). Myeloablative megatherapy with
autologous stem-cell rescue versus oral maintenance chemotherapy as consolidation
treatment in patients with high-risk neuroblastoma: a randomised controlled trial. Lancet Oncol
6:649 - 658.

Literaturverzeichnis
61
Blaney SM, Bernstein M, Neville K, Ginsberg G, Kitchen B, Horton T, Berg SL, Krailo M,
Adamison PC (2004). Phase I Study of the Proteasome Inhibitor Bortezomib in pediatric
patients with refractory solid tumours: A children´s oncology group study (ADVL0015). J clin
Oncol 22:4804 - 4809.
Brignole C, Marimpietri D, Pastorino F, Nico B, Di Paolo D, Cioni M, Piccardi F, Cilli M,
Pezzolo A, Corrias MV, Pistoia V, Ribatti D, Pagnan G, Ponzoni M (2006). Effekt of
Bortezomib on Human Neuroblatoma Cell Growth, Apoptosis and Angiogenesis. J Nat Cancer
Inst 98:1142 – 1157.
Bold RJ, Virudachalam S, Mc Coney DJ (2001). Chemosensitization of Pancreatic Cancer by
Inhibition of the 26 S Proteasome. J Surg Res 100:11 - 17.
Brodeur GM (2003). Neuroblastoma: biological insights into a clinical enigma. Nat Rev Cancer
3:203 – 216.
Brown JM, Attardi LD (2005). The role of apoptosis in cancer development and treatment
response. Nat Rev Cancer 5:231 – 237.
Buck AC, Schirrmeister HH, Guhlmann CA, Diedereichs CG, Shen C, Buchmann I, Kozerke J,
Birk D, Mattfeldt T, Reske SN (2001). Ki-67 immunostaining in pancreatic cancer and chronic
active pancreatitis: does in vivo FDG uptake correlate with proliferative activity? J Nucl Med
42: 721 - 725.
Cornelissen B, Kersemans V, Jans L, Staelens L, Oltenfreiter R, Thonissen T, Achten E,
Slegers G (2005). Comparison between 1 T MRI and non-MRI based volumetry in inoculated
tumours in mice. Br J Radiol 78:338 - 342.

Literaturverzeichnis
62
Deutsche Krebsgesellschaft e.V., Informationszentrum für Stadards in der Onkologie (2003).
Kurzgefasste Interdisziplinäre Leitlinien 2004, Diagnostik und Therapie maligner
Erkrankungen. Zuckschwerdt Verlag München – Bern – Wien – New York, 4. Auflage:423 -
429.
Dittmann H, Dohmen BM, Paulsen F, Eichhorn K, Eschmann SM, Horger M, Wehrmann M,
Machulla HJ, Bares R (2003). [18F]FLT PET for diagnosis and staging of thoracic tumours. Eur
J Nucl Med Imaging 30:1407 - 1412.
Fang J, Quinones QJ, Holman TL, Morowitz MJ, Wang Q, Zhao H, Sivo F, Maris JM, Wahl ML
(2006). H+ linked monocarboxylate transporter (MCT1/SLC16A1): a potential treapeutic target
for high-risk neuroblastoma. Mol Pharmacol 70:2108 – 2115.
Frankel A, Man S, Elliot P, Adams J, Kerbel RS (2000). Lack of Multicellular Drug Resistance
Observed in Human Ovarian and Prostate Carcinoma treated with the Protesome Inhibitor PS-
341. Clin Cancer Res 6:3719 - 3728.
Fulda S, Proemba C, Berwanger B, Hacker S, Eilers M, Christiansen H, Hero B, Debatin KM
(2006). Loss of caspase-8 expression does not correlate with MYCN amplification aggressive
disease or prognosis in neuroblastoma. Cancer Res 66:10016 - 10023.
Hamner JB, Dickson PV, Sims TL, Zhou J, Spence Y, Ng CY, Davidoff AM (2007). Bortezomib
inhibits angiogenesis and reduces tumor burden in a murine model of neuroblastoma. Surgery
142:185 – 191.
Hazelton TR und Coppage L (2005). Imaging for lung cancer restaging. Semin Röntgenol
40:182 – 192.
Hero B und Berthold F (2002). Neuroblastom. Monatszeitschrift Kinderheilkunde 150:775 –
788.

Literaturverzeichnis
63
Hidshma T, Chanhan D, Podar K, Schlussman RL, Richardson P, Anderson KC (2001). Novel
therapies targeting the myeloma cell and its bone marrow microenvironment. Semin Oncol 28:
607 - 612.
Jojovic M, Schumacher U (2000). Quantitative assessment of spontaneous lung metastases
of human HT29 colon cancer cells transplanted into SCID mice. Cancer Lett 152:151 – 156.
Karin M, Cao Y, Greten FR, Li ZW (2002). NF-kappa B in cancer: from innocent bystander to
major culprit. Nat Rev Cancer 2:301 – 310.
Katayoun AI, Horton LW, La Fleur BJ, Sosman J, Rochmond A (2004). Augmenting
Chemosensitivity of Malignant Melanoma Tumors via Proteasome Inhibition: Implication for
Bortezomib (Velcade, PS-341) as a therapeutic agent for Malginant Melanoma. Cancer Res
64:4912 - 4918.
Khan T, Stauffer JK, Williams SR, Hixon JA, Salcedo R, Lincoin E, Back TC, Powell D, Lockett
S, Arnold AC, Sayers TJ, Yigginton JM (2006). Proteasome Inhibiton to Maximize the
Apoptotic Potential of Cytokine Therapy for Murine Neuroblastoma Tumours. J Immunol
176:6302 - 6312.
Kolezko B: Kinderheilkunde und Jugendmedizin (2003). 12. Auflage, Springer Verlag Berlin:
331 - 350.
Krieger-Hinck N, Gustke H, Valentiner U, Mikecz P, Buchert R, Mester J, Schumacher U
(2006). Visualisation of neuroblastoma growth in a scid mouse model using [18F]FDG and
[18F]FLT PET. Anticancer Res 26:3467 - 3472.
Kruger A, Soeltl R, Sopov I, Kopitz C, Arlt M, Magdolen V, Harbeck N, Gänsbacher B, Schmitt
M (2001). Hydroxamate-type matrix metalloproteinase inhibitor batimastat promotes liver
metastasis. Cancer Res 61:1272 - 1275.

Literaturverzeichnis
64
Le Blanc R, Caley LP, Hideshima T, Lentzsch S, Mitsiades C, Mitsiades N, Neuberg D,
Goloubeva O, Pien CS, Adams J, Gupta D, Richardson PG, Munshi NC, Anderson KC (2002).
Proteasome Inhibtior PS-341 inhibits human myeloma cell growth in vivo and prolongs
survival in a murine model. Cancer Res 62:4996 - 5000.
Ling YH, Liebes L, Jiang JD, Holland JF, Eliot J, Adams J, Muggia FM, Soler RP (2003).
Mechanisms of Proteasome Inhibitor PS-341-induced G2-M-Phase Arrest and Apoptosis in
Human Non-small Cell lug Cancer cell lines. Clin Cancer Res 9:1145 - 1154.
Ludwig C (2004). Bortezomib (Velcade) - ein neues Wirkungsprinzip in der Behandlung des
multiplen Myeloms. Schweiz Med Forum 4:1226 - 1227.
Michaelis M, Fichtner I, Behrens D, Haider W, Rothweiler F, Mack A, Cinatl J, Doerr HW,
Cinatl J (2006). Anti-cancer effects of bortezomib against chemoresistant neuroblastoma cell
lines in vitro and in vivo. Int J Oncol 28:439 - 446.
Morgenstern BZ, Krivoshik AP, Rodriguez V, Anderson PM (2004). Wilms tumor and
neuroblastoma. Acta Pädiatr Suppl 445:78 - 85.
Nischwitz M (2007). Nachweis disseminierter Tumorzellen operabler gastrointestinaler
Tumoren mittels quantitativer RT-PCR von Melanom Antigen (MAGE) – A – Transkription.
Dissertation der medizinischen Fakultät der Ludwig-Maximilians-Universität zu München: 9 –
10.
Pacak K, Eisenhofer G, Goldstein DS (2004). Functional imaging of endocrine tumours: role of
positron emission tomography. Endocr Rev 25:568 - 580.
Pham LV, Tamayo AT, Yoshimura LC, Lo P, Ford RJ (2003). Inhibition of Constitutive NFkB
Activation in Mantle cell Lymphoma B Cells leads to Induction of Cell Cycle Arrest and
Apoptosis. J Immunol 171:88 – 95.

Literaturverzeichnis
65
Richardson PG und Anderson KC (2003). Bortezomib: A Novel Therapy Approved for Multiple
Myeloma. Clin Adv in Hem & Onc 1:596 – 600.
Ruiz S, Krupnik Y, Keatin M, Chandra J, Palladino M, Mc Conkey D (2006). The proteasome
inhibitor NPE-0052 is a more effective inducer of apoptosis than bortezomib in lymphocytes
from patients with chronic lymphocytic leukemia. Mol Cancer Ther 5:1836 - 1843.
Schwab M, Westermann F, Hero B, Berthold F (2003). Neuroblastoma: biology and molecular
and chromosomal pathology. Lancet Oncol 4:472 - 480.
Schneider T (2002). Humane Alu-Sequenzen als Surrogat-Marker für die Metastasierung in
humanen Xenograft - Tiermodellen, Dissertation, Fakultät für Biologie der Ludwig Maximilians-
Universität München: 93.
Schneider T et al. (2002). Quantification of human Alu sequences by real-time PCR - - an
improved method to measure therapeutic efficacy of anti-metastatic drugs in human
xenotransplants. Clin Exp Metastasis 19:571 - 582.
Shah SA, Potter MW, Mc Dade TP, Ricciardi R, Perugini RA, Elliot PJ, Adams J, Callery MP
(2001). 26S Proteasome inhibition induces apoptosis and limits growth of human pancreatic
cancer. Cell Biochem 82:110 – 122.
Simon T, Spitz R, Faldum A, Hero B, Berthold F (2004). New Defintion of Low-Risk
Neuroblastoma using stage, age and 1p and MYCN Status. Pediatr Hematol Oncol 26:791 -
796.
Simon T (2005). Neuroblastom: Der Urologe 44:543 - 555.

Literaturverzeichnis
66
Smyczek-Gargya B, Fersis N, Dittmann H, Vogel U, Reischl G, Machulla HJ, Wallwiener D,
Bares R, Dohmen BM (2004). PET with [18F]Fluorothymidine for imaging of primary breast
cancer: a pilot study. Eur J Nucl Med Mol Imaging 31:720 - 724.
Sunwoo JB, Chen Z, Dong G, Yeh N, Crowl Bancroft C, Sausville E, Adams J, Elliott P, van
Waes C (2001). Novel proteasome Inhibitor PS-341 inhibits activation of Nuclear factor-κB,
cell survival, tumor growth, and angiogenesis in squamos cell carcinoma. Clin Cancer Res
7:1419 - 1428.
Teicher BA (2006). Tumor models for efficacy determination. Mol Cancer Ther 5:2435 - 2443.
Thies A, Mauer S, Fodstad O, Schumacher U (2007). Clinically proven markers of metastasis
predict metastatic spread of human melanoma cells engrafted in scid mice. Br J Cancer
96:609 – 616.
Valentiner U, Carlsson M, Erttmann R, Hildebrandt H, Schumacher U (2005). Ligands for the
peroxisome proliferator-activated receptor-γ have inhibitory effects on growth of human
neuroblastoma cells in vitro. Toxicol 213:157 - 168.
Thornberry NA, Lazebnik Y (1998). Caspases: enemies within. Science 281:1312 – 1316.
Van Waes C, Chang AA, Lebowitz, PF, Druzgal CH, Chen Z, Elsayed YA, Sunwoo JB, Rudy
SF, Morris JC, Mitchell JB, Camphausen K, Gius D, Adams J, Sausville EA, Conley BA
(2005). Inhibition of nuclear factor-kappaB and target genes during combined therapy with
proteasome inhibitor bortezomib and reirradiation in patients with recurrent head-and-neck
squamous cell carcinoma. Int J Radiat Oncol Biol Phys 63:1400 - 1412.
Van Westreenen HL, Cobben DX, Jager PL, van Dullemen HM, Wesseling J, Elsinga PH,
Plukker JT (2005). Comparison of [18F]FLT-PET and [18F]FDG PET in esophageal cancer. J
Nucl Med 46:400 - 404.

Danksagung
67
Danksagung
Allen, die zum Gelingen an dieser Arbeit beigetragen haben, möchte ich an dieser Stelle
meinen Dank aussprechen:
Mein ganz besonderer Dank gilt meinem Lehrer und Doktorvater Herrn Prof. Dr. med. Udo
Schumacher für die Bereitstellung des interessanten Themas, die Möglichkeit dieses im
Anatomischen Institut ΙΙ durchführen zu können und für seine immer motivierenden
Anregungen und Ideen, die diese Arbeit erst wachsen ließen.
Bei meiner Betreuerin Frau Dr. med. Ursula Valentiner bedanke ich mich sehr für die
uneingeschränkte Betreuung während der gesamten Durchführung der Arbeit, den vielen
hervorragenden Tipps und Ratschlägen und ihrer immer unterstützenden Art.
Herrn Prof. Dr. Erttmann und Herrn Prof. Dr. Hildebrandt danke ich sehr herzlich für die zur
Verfügung gestellten Neuroblastomzelllinien.
Dem gesamten MTA-Laborteam möchte ich für die hervorragende Unterstützung und der
„umsorgenden“ Art während der ganzen Zeit recht herzlich danken. Dieses gilt vor allem Herrn
Klaus Siebert, Frau Renate Gehrke, Frau Maike Märker und ganz besonders Frau Kornelia
Babista.
Den wissenschaftlichen Assistenten, besonders Frau Dr. rer. nat. Heike Gustke und Frau Dr.
rer. nat. Nina Nehmann, möchte ich sowohl für die ausgezeichneten Hilfestellungen bei
praktischen Fragestellungen als auch beim Verfassen der Doktorarbeit danken.
Herrn Prof. Dr. med. Brenner, Herrn Florian Wilke und besonders Herrn Dr. med. Kersten
Peldschus danke ich sehr für ihre Unterstützung bezüglich des Imagings. Weiterhin möchte
ich mich bei Herrn Dr. Ulf Tiede und Herrn Dr. Michael Owsijewitsch für die Bearbeitung der
MRT – Bilder mittels der Voxel-Man Software recht herzlich bedanken.
Den Mitarbeitern des “Tierstalles“ danke ich sehr für die gute Versorgung der Versuchstiere.
Bei meinen Mitdoktoranden bedanke ich mich für eine tolle gemeinsame Zeit mit vielen
anregenden Gesprächen, gegenseitiger Unterstützung und Motivation.
Meiner Familie danke ich besonders für ihre stetige Begleitung und Unterstützung während
meines gesamten Studiums.
Zu guter Letzt möchte ich mich bei meinen Freunden bedanken, die durch Motivation und
Abwechslung ebenfalls sehr zum Gelingen beigetragen haben.

Lebenslauf
68
Lebenslauf
Persönliche Daten
Name Haane
Vorname Christina
Geburtsdatum-, ort 02/03/1979 in Dorsten
Schulbildung
08/1985 – 07/1989 Grundschule Don Bosco, Dorsten – Lembeck
08/1989 – 06/1995 St. Elisabeth Realschule, Reken
08/1995 – 06/1997 Fachoberschule für Sozial- und Gesundheitswesen, Dorsten, Erlangen der Fachhochschulreife
Berufsausbildung
10/1997 – 09/2000 Timmermeister Schule, Lehranstalt für Physiotherapie, Münster, Staatsexamen
Berufserfahrung
01/2001 – 03/2002 Berufsgenossenschaftliches Unfallkrankenhaus Hamburg-Boberg, Abteilung für Physiotherapie
Hochschulausbildung
12/2001 Studienberechtigung § 38 des Hamburgischen Hochschulgesetzes
04/2002 Aufnahme des Studiums der Humanmedizin am Universitätsklinikum Hamburg-Eppendorf der Universität Hamburg
03/2004 Physikum
04/2004 – 08/2006 Klinische Ausbildung Universitätsklinikum Hamburg-Eppendorf
09/2006 – 07/2007 Université Victor Segalen Bordeaux 2, ERASMUS Studienjahr in Frankreich
seit 08/2007 Praktisches Jahr

Selbstständigkeitserklärung
69
Selbstständigkeitserklärung
Ich versichere ausdrücklich, dass ich die Arbeit selbstständig und ohne fremde Hilfe verfasst,
andere als die von mir angegebenen Quellen und Hilfsmittel nicht benutzt und die aus den
benutzten Werken wörtlich oder inhaltlich entnommenen Stellen einzeln nach Ausgabe
(Auflage und Jahr des Erscheinens), Band und Seite des benutzten Werkes kenntlich
gemacht habe, und dass ich die Dissertation bisher nicht einem Fachvertreter an einer
anderen Hochschule zur Überprüfung vorgelegt oder mich anderweitig um Zulassung zur
Promotion beworben habe.
Hamburg, den 24.10.2007
Christina Haane


















